Journal of Neuro-Oncology 36: 113–121, 1998. 1998 Kluwer Academic Publishers. Printed in the Netherlands. Laboratory Investigation Effects of DFMO on glioma cell proliferation, migration and invasion in vitro A-Jorge A. Terzis, 1 Paal-Henning Pedersen, 2 Burt G. Feuerstein, 3 Hans Arnold, 1 Rolf Bjerkvig, 2 and Dennis F. Deen 3 1 Department of Neurosurgery, Luebeck, Germany; 2 Institute of Anatomy and Cell Biology, Bergen, Norway; 3 Brain Tumor Research Center, San Francisco, USA Key words: glioma, polyamines, invasiveness, DFMO, three dimensional culture Summary The polyamine inhibitor DL-α-difluoromethylornithine (DFMO) is a specific irreversible inhibitor of ornith- ine decarboxylase which is a rate-limiting enzyme in the polyamine bio-synthesis pathway. The present study describes the effects of DFMO on glioma cell proliferation, migration and invasion using multicellular sphe- roids from three glioma cell lines (GaMg, U-251 Mg and U-87 Mg). 10 mM DFMO reduced cell migration in the three cell lines by about 30–50%. 1 mM putrescine, added together with DFMO inhibited the DFMO effect. A stronger effect was observed in the growth assay where 10 mM DFMO reduced the spheroid growth, for all cell lines, by 90%. This effect was also reversed by adding 1 mM of putrescine. In vitro tumor cell invasion experiments indicated after 3 days of confrontation, an extensive invasion also after 10 mM DFMO treatment. The brain aggregate volumes were reduced to about the same extent as in the absence of drug, suggesting essentially no effects of DFMO on the invasive process. It is concluded that the tumor spheroids retained their ability to invade normal brain tissue even after DFMO exposure. However, DFMO inhibited spheroid growth and cell migration which supports the notion that cell growth, migration and invasion are biological properties that are not necessarily related to each other. Introduction Cerebral tumors are responsible for approximately 2% of all cancer deaths and high grade glioma is the most common cerebral tumor in adults. The glio- mas invade the brain extensively and there is no sat- isfactory treatment. The best available treatment, using surgery, radiation therapy and systematic che- motherapy, results in a median survival of z 1 year [1–3]. Most failures of treatment occur because of local recurrence of the tumor. During the past dec- ade, it has been suggested that α-Difluoromethylor- nithine (DFMO), which is an ornithine decarboxy- lase (ODC) inhibitor, may be used in the treatment of gliomas either alone or in combination with other therapy regimens [4, 5]. It is well known that the aliphatic polyamines pu- trescine, spermidine and spermine are widely dis- tributed in eukaryotic cells [6], and changes in their concentrations have been associated with a number of important cellular activities [7, 8]. For instance, the polyamine biosynthetic enzymes and the con- centrations of the polyamines increase precipitous- ly when growth and differentiation is induced. It is also well known that growth stimulation of quies- cent cells and tissues invariably results in an in- creased rate of polyamine synthesis [9, 10]. In addi- tion, a large body of circumstantial evidence sug-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
Journal of Neuro-Oncology 36: 113–121, 1998. 1998 Kluwer Academic Publishers. Printed in the Netherlands.
Laboratory Investigation
Effects of DFMO on glioma cell proliferation, migration and invasion invitro
A-Jorge A. Terzis,1 Paal-Henning Pedersen,2 Burt G. Feuerstein,3 Hans Arnold,1 Rolf Bjerkvig,2 and Dennis F.Deen3
1Department of Neurosurgery, Luebeck, Germany; 2Institute of Anatomy and Cell Biology, Bergen, Norway;3Brain Tumor Research Center, San Francisco, USA
Key words: glioma, polyamines, invasiveness, DFMO, three dimensional culture
Summary
The polyamine inhibitor DL-α-difluoromethylornithine (DFMO) is a specific irreversible inhibitor of ornith-ine decarboxylase which is a rate-limiting enzyme in the polyamine bio-synthesis pathway. The present studydescribes the effects of DFMO on glioma cell proliferation, migration and invasion using multicellular sphe-roids from three glioma cell lines (GaMg, U-251 Mg and U-87 Mg). 10 mM DFMO reduced cell migration inthe three cell lines by about 30–50%. 1 mM putrescine, added together with DFMO inhibited the DFMOeffect.
A stronger effect was observed in the growth assay where 10 mM DFMO reduced the spheroid growth, forall cell lines, by 90%. This effect was also reversed by adding 1 mM of putrescine.
In vitro tumor cell invasion experiments indicated after 3 days of confrontation, an extensive invasion alsoafter 10 mM DFMO treatment. The brain aggregate volumes were reduced to about the same extent as in theabsence of drug, suggesting essentially no effects of DFMO on the invasive process. It is concluded that thetumor spheroids retained their ability to invade normal brain tissue even after DFMO exposure. However,DFMO inhibited spheroid growth and cell migration which supports the notion that cell growth, migrationand invasion are biological properties that are not necessarily related to each other.
Introduction
Cerebral tumors are responsible for approximately2% of all cancer deaths and high grade glioma is themost common cerebral tumor in adults. The glio-mas invade the brain extensively and there is no sat-isfactory treatment. The best available treatment,using surgery, radiation therapy and systematic che-motherapy, results in a median survival of z 1 year[1–3]. Most failures of treatment occur because oflocal recurrence of the tumor. During the past dec-ade, it has been suggested that α-Difluoromethylor-nithine (DFMO), which is an ornithine decarboxy-lase (ODC) inhibitor, may be used in the treatment
of gliomas either alone or in combination with othertherapy regimens [4, 5].
It is well known that the aliphatic polyamines pu-trescine, spermidine and spermine are widely dis-tributed in eukaryotic cells [6], and changes in theirconcentrations have been associated with a numberof important cellular activities [7, 8]. For instance,the polyamine biosynthetic enzymes and the con-centrations of the polyamines increase precipitous-ly when growth and differentiation is induced. It isalso well known that growth stimulation of quies-cent cells and tissues invariably results in an in-creased rate of polyamine synthesis [9, 10]. In addi-tion, a large body of circumstantial evidence sug-

Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
114
gests that polyamine synthesis is associated withDNA replication, cell division, and in stabilizingDNA structure [11, 12]. The decarboxylation of L-ornitine, catalyzed by ODC, may be considered theinitial step in mammalian polyamine synthesis [13,14]. DFMO belongs to the class of enzyme-activa-ted irreversible inhibitors known as kcat inhibitors orsuicide enzyme inactivators [15]. DFMO has beenused in cell cultures and in intact animals to induce arapid decrease in putrescine and spermidine con-centrations [16, 17]. The rationale for utilizingDFMO in therapy is based on the fact that this com-pound is highly selective, has no acute pharmaco-logic activity, and is essentially nontoxic. As indicat-ed above, DFMO specifically inactivates ODC anddoes not influence other enzymes employing or-nithine as a substrate [16, 18].
The in vitro sensitivity of cancer cells to DFMOhas usually been determined by clonogenic survivalassays or growth inhibition of treated cells culturedas monolayer or in suspension. However, it seemsreasonable to assume that cells growing under dif-ferent micro-environmental conditions character-ized by, variations in nutrient supply, oxygen ten-sion and pH also differ in their sensitivities toDFMO. It has been shown that multicellular sphe-roids both from normal and malignant tissues canmaintain several biochemical and morphologicalfeatures similar to those of the corresponding tissuein vivo [19–22]. Therefore, by studying effects ofDFMO on spheroids, valid information may be pro-vided regarding the cellular effects of DFMO in vi-vo.
For this purpose, spheroids from three perma-nent human glioma cell lines (GaMg, U-251 Mg andU-87 Mg) were used. Furthermore, tumor cell in-vasion was studied in a co-culture assay involvingnormal rat brain tissue as a target for the tumorcells. Thus, the present work focus on some key bi-ological features which characterise gliomas (cellgrowth, migration and invasion) and how DFMOinduced inhibition of polyamines affect these prop-erties.
Material and methods
Drug
DFMO (α-Difluoromethylornithine) was obtainedfrom Marion-Merrill Dow Research Institute (Cin-cinnati, Ohio) and stored at 4 °C. DFMO (lyophil-ized powder) was prepared 1hour before use by dis-solving it in ethanol (96%). The solution was fur-ther diluted in Dulbecco’s modified Eagle’s medi-um (DMEM) to a final concentration of 10 mM.DFMO in this concentration has been used in cellcultures and intact animals to produce a rapid fall inthe concentrations of putrescine and spermidine.
In order to revert the DFMO effects, 1 mM pu-trescine was added to the cultures. Putrescine wasobtained from Sigma (St. Louis, MO). Putrescine(lyophilized powder) was prepared 1 hour beforeuse by dissolving it in DMEM to a final concentra-tion of 1 mM.
Cell lines and cell culture conditions
Multicellular tumor spheroids from 3 human glio-ma cell lines were used. The human U-87 Mg andU-251 Mg cell lines were kindly supplied by Dr. J.Carlsson, Uppsala University, Uppsala, Sweden.The GaMg was obtained from a 42-year-old femaleand histologically identified as a Glioblastoma [23].All three cell lines were grown in DMEM supple-mented with 10% heat-inactivated new-born bo-vine serum, 4 times the prescribed concentration ofnon-essential amino acids, 2% L-glutamine, peni-cillin (100 IU/ml) and streptomycin (100 µg/ml).The cells were cultured at 37 °C, in 100% relativehumidity, 95% air and 5% CO2.
Tumor spheroids
Tumor spheroids were initiated by the agar overlayculture method as described by Yuhas et al. [24].Briefly, spheroids were formed by seeding 5 × 106
cells in 20 ml growth medium into 80 cm2 agar-coat-ed tissue-culture flasks (Nunc, Roskilde; Den-mark). After 10 days in culture, spheroids with dia-

Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
115
meters between 200 and 250 µm were selected forfurther experiments (see below).
Spheroid growth
For each experiment, the spheroids were divided in-to 4 groups (control, 1 mM putrescine alone, 10 mMDFMO alone, and putrescine + DFMO). For eachgroup, 12 spheroids were transferred individuallyinto 16 mm multiwell dishes (Nunc). The disheswere base-coated with 0.5 ml 0.75% DMEM-agarand filled with 1 ml of DMEM. The diameters of thespheroids were measured regularly over a 15-dayperiod using a phase-contrast microscope and thespheroid volume was calculated. The experimentswere performed in triplicate.
Tumor cell migration
For each experiment, the spheroids were divided in-to 4 groups (control putrescine alone, DFMO alone,and putrescine + DFMO). For each group, 12 sphe-roids were transferred individually into uncoated16 mm multiwell dishes (Nunc) filled with 1 mlDMEM. The spheroids plated within 2 hr, and thecellular outgrowth from one spheroid was definedas a colony. Colony diameters were measured regu-larly in the phase-contrast microscope over a 4-dayperiod (96 hr), and the colony outgrowth areaswere then calculated. The experiments were per-formed in triplicate.
Brain aggregates
Fetal rat brain cell aggregates were obtained from18-day-old fetuses of inbred Wistar rats. The brainswere dissected out under aseptic conditions and themeningeal coverings were removed. The brainswere placed in sterile petri dishes containing calci-um and magnesium free phosphate buffered saline(PBS). The tissue was cut into small pieces, washedin PBS and dissociated by serial trypsination using0.025% trypsin (Whittaker Bioproducts, Walkers-ville, MD) into a single cell suspension. The brain
cell aggregates were produced by seeding 6 × 106
cells in 1 ml of growth medium into 16 mm multiwelldishes (Nunc), base coated with 0.5 ml of a non-ad-herent 0.75% medium-agar substrate. During a 4-day period, immature brain cell aggregates wereformed. After 20 days in tissue culture, the cellulardifferentiation within the aggregates was completeforming a defined three layered structure contain-ing mature astrocytes, oligodendrocytes and neu-rons with myelinated axons and synapses present ina well developed neuropil. Thus, the aggregatesshow a close resemblance to adult brain tissue [25–29].
Tumor-cell invasion
6 days before confrontation with brain aggregates,the tumor spheroids were exposed to 10 mMDFMO alone, or to 1 mM putrescine alone, or to acombination of 10 mM DFMO and 1 mM putres-cine. This was done in order to obtain an effectivegrowth inhibition, based on the results obtainedfrom the protocol described for spheroid growth.There were 5 replicate samples for each of the 4groups. The spheroids were then transferred to agarcoated 96-multiwell dishes with a sterile Pasteurpipette and confronted individually with the brainaggregates. The co-cultures were incubated for a 3-days period at 37 °C, and then fixed for light-micro-scopic examination and morphometric analyses.The reduction of brain-aggregate volume was de-termined as a percentage of the initial brain volumeand this was used to quantify the invasive process(see below). The experiments were done in dupli-cate.
Light microscopy
The co-cultures were fixed in 2% glutaraldehyde in0.1 M sucrose-adjusted sodium-cacodylate buffer(300 ± 25 mOsm). After 24 hr, the specimens werewashed in the same buffer, without glutaraldehyde,and post-fixed for 1 hr in 1% OsO4 before serial de-hydration in increasing gradients of ethanol up to100%. Embedding of the co-cultures in Epon 812
Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
116
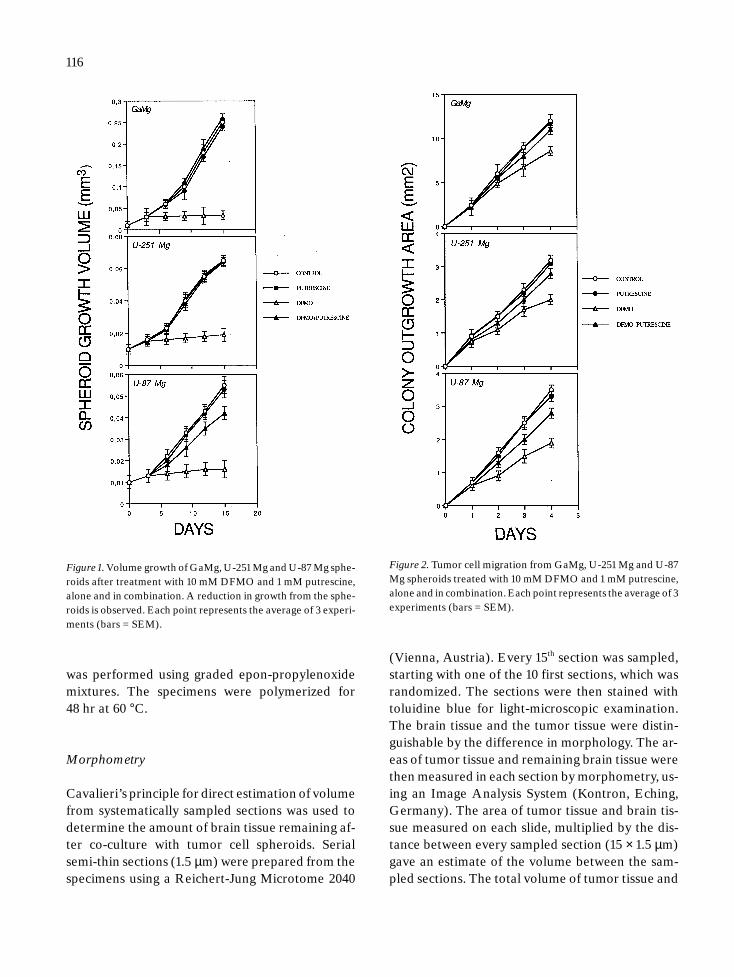
Figure 1. Volume growth of GaMg, U-251Mg and U-87 Mg sphe-roids after treatment with 10 mM DFMO and 1 mM putrescine,alone and in combination. A reduction in growth from the sphe-roids is observed. Each point represents the average of 3 experi-ments (bars = SEM).
Figure 2. Tumor cell migration from GaMg, U-251 Mg and U-87Mg spheroids treated with 10 mM DFMO and 1 mM putrescine,alone and in combination. Each point represents the average of 3experiments (bars = SEM).
was performed using graded epon-propylenoxidemixtures. The specimens were polymerized for48 hr at 60 °C.
Morphometry
Cavalieri’s principle for direct estimation of volumefrom systematically sampled sections was used todetermine the amount of brain tissue remaining af-ter co-culture with tumor cell spheroids. Serialsemi-thin sections (1.5 µm) were prepared from thespecimens using a Reichert-Jung Microtome 2040
(Vienna, Austria). Every 15th section was sampled,starting with one of the 10 first sections, which wasrandomized. The sections were then stained withtoluidine blue for light-microscopic examination.The brain tissue and the tumor tissue were distin-guishable by the difference in morphology. The ar-eas of tumor tissue and remaining brain tissue werethen measured in each section by morphometry, us-ing an Image Analysis System (Kontron, Eching,Germany). The area of tumor tissue and brain tis-sue measured on each slide, multiplied by the dis-tance between every sampled section (15 × 1.5 µm)gave an estimate of the volume between the sam-pled sections. The total volume of tumor tissue and

Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
117
Figure 3. Photomicrographs (× 225) of co-cultures between U-251 Mg spheroids (T) and rat brain aggregates (N). (A) Control co-cultureshowing extensive invasion and degradation of brain tissue. (B) U-251 Mg spheroids treated with 1 mM putrescine, (C) with 10 mMDFMO, and (D) with 1 mM putrescine + 10 mM DFMO showing also an extensive invasion into the brain aggregates.
of remaining brain tissue was calculated by summa-tion of all the individual volumes. The coefficient oferror in this method for estimating volume of sec-tions is below 5% [30].
Statistics
The Student’s t test was used to determine the levelof significance of the difference between individualgroups. The drug response in the growth, migrationand invasion experiments was assessed statisticallyby regression analysis.
Results
Spheroid volume growth
For all three cell lines (GaMg, U-251 Mg, and U-87Mg), tumor spheroids continuously exposed to10 mM DFMO showed an inhibition of growthwhich became apparent after 3 days of drug expo-sure. There were 6 to 10-fold volume increases inthe untreated groups, in contrast to treated sphe-roids where only a slight volume increasing duringthe observation period was observed.
1 mM putrescine prevented the DFMO inhibito-ry effects in all three cell lines. For GaMg and U-251Mg, the growth rates were identical to those foundin the control groups. However, the putrescine re-
Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
118
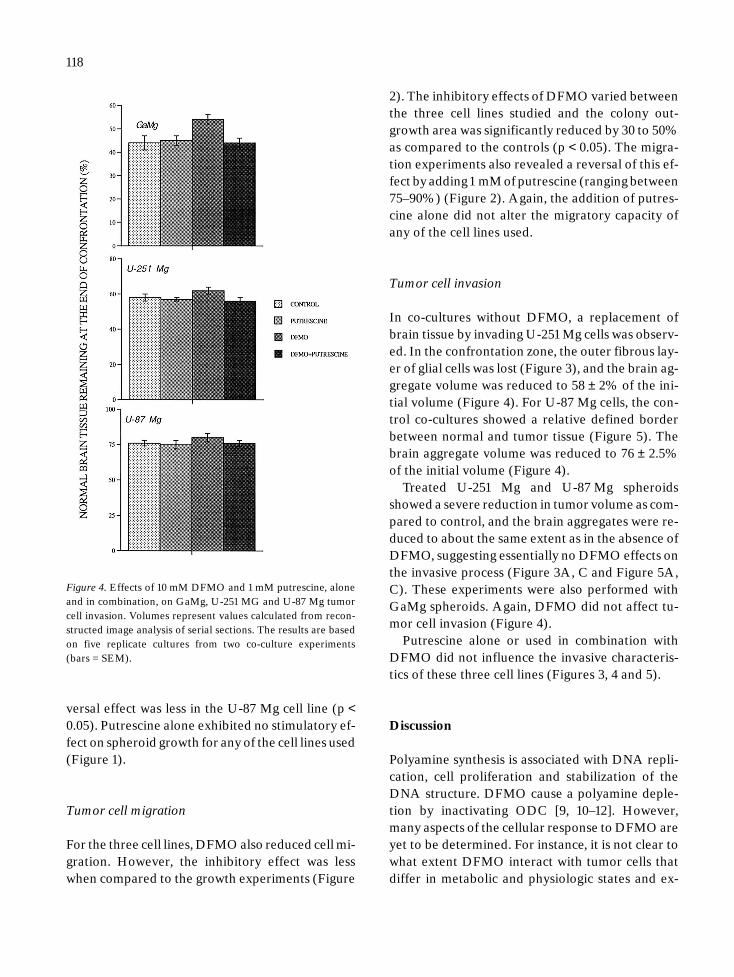
Figure 4. Effects of 10 mM DFMO and 1 mM putrescine, aloneand in combination, on GaMg, U-251 MG and U-87 Mg tumorcell invasion. Volumes represent values calculated from recon-structed image analysis of serial sections. The results are basedon five replicate cultures from two co-culture experiments(bars = SEM).
versal effect was less in the U-87 Mg cell line (p <0.05). Putrescine alone exhibited no stimulatory ef-fect on spheroid growth for any of the cell lines used(Figure 1).
Tumor cell migration
For the three cell lines, DFMO also reduced cell mi-gration. However, the inhibitory effect was lesswhen compared to the growth experiments (Figure
2). The inhibitory effects of DFMO varied betweenthe three cell lines studied and the colony out-growth area was significantly reduced by 30 to 50%as compared to the controls (p < 0.05). The migra-tion experiments also revealed a reversal of this ef-fect by adding 1 mM of putrescine (ranging between75–90%) (Figure 2). Again, the addition of putres-cine alone did not alter the migratory capacity ofany of the cell lines used.
Tumor cell invasion
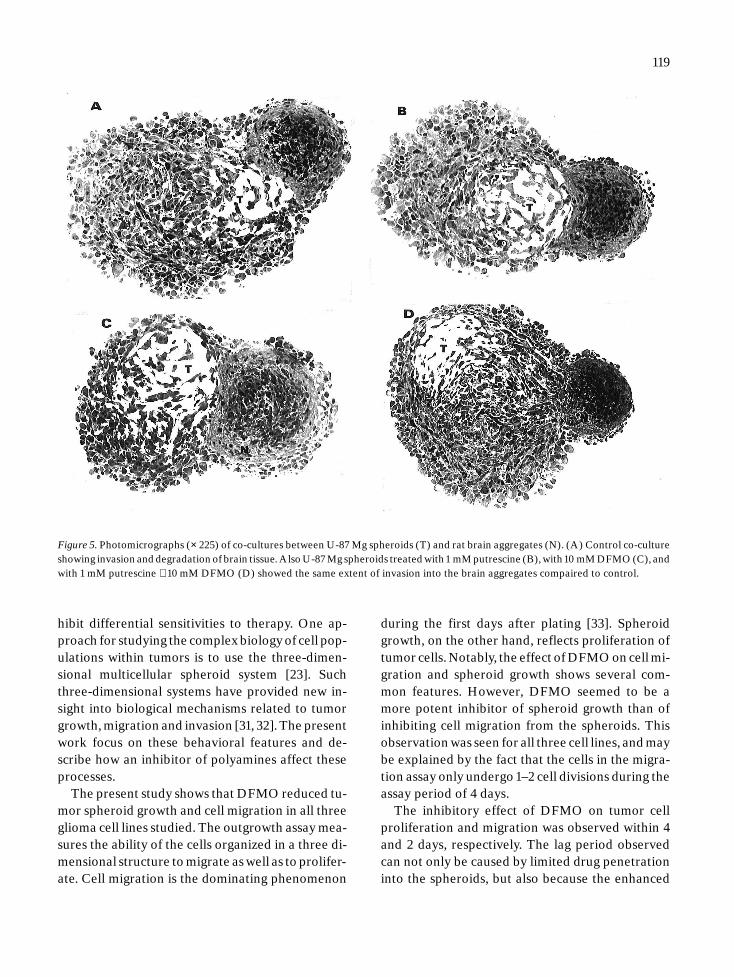
In co-cultures without DFMO, a replacement ofbrain tissue by invading U-251 Mg cells was observ-ed. In the confrontation zone, the outer fibrous lay-er of glial cells was lost (Figure 3), and the brain ag-gregate volume was reduced to 58 ± 2% of the ini-tial volume (Figure 4). For U-87 Mg cells, the con-trol co-cultures showed a relative defined borderbetween normal and tumor tissue (Figure 5). Thebrain aggregate volume was reduced to 76 ± 2.5%of the initial volume (Figure 4).
Treated U-251 Mg and U-87 Mg spheroidsshowed a severe reduction in tumor volume as com-pared to control, and the brain aggregates were re-duced to about the same extent as in the absence ofDFMO, suggesting essentially no DFMO effects onthe invasive process (Figure 3A, C and Figure 5A,C). These experiments were also performed withGaMg spheroids. Again, DFMO did not affect tu-mor cell invasion (Figure 4).
Putrescine alone or used in combination withDFMO did not influence the invasive characteris-tics of these three cell lines (Figures 3, 4 and 5).
Discussion
Polyamine synthesis is associated with DNA repli-cation, cell proliferation and stabilization of theDNA structure. DFMO cause a polyamine deple-tion by inactivating ODC [9, 10–12]. However,many aspects of the cellular response to DFMO areyet to be determined. For instance, it is not clear towhat extent DFMO interact with tumor cells thatdiffer in metabolic and physiologic states and ex-
Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
119
Figure 5. Photomicrographs (× 225) of co-cultures between U-87 Mg spheroids (T) and rat brain aggregates (N). (A) Control co-cultureshowing invasion and degradation of brain tissue. Also U-87 Mg spheroids treated with 1 mM putrescine (B), with 10 mM DFMO (C), andwith 1 mM putrescine + 10 mM DFMO (D) showed the same extent of invasion into the brain aggregates compaired to control.
hibit differential sensitivities to therapy. One ap-proach for studying the complex biology of cell pop-ulations within tumors is to use the three-dimen-sional multicellular spheroid system [23]. Suchthree-dimensional systems have provided new in-sight into biological mechanisms related to tumorgrowth, migration and invasion [31, 32]. The presentwork focus on these behavioral features and de-scribe how an inhibitor of polyamines affect theseprocesses.
The present study shows that DFMO reduced tu-mor spheroid growth and cell migration in all threeglioma cell lines studied. The outgrowth assay mea-sures the ability of the cells organized in a three di-mensional structure to migrate as well as to prolifer-ate. Cell migration is the dominating phenomenon
during the first days after plating [33]. Spheroidgrowth, on the other hand, reflects proliferation oftumor cells. Notably, the effect of DFMO on cell mi-gration and spheroid growth shows several com-mon features. However, DFMO seemed to be amore potent inhibitor of spheroid growth than ofinhibiting cell migration from the spheroids. Thisobservation was seen for all three cell lines, and maybe explained by the fact that the cells in the migra-tion assay only undergo 1–2 cell divisions during theassay period of 4 days.
The inhibitory effect of DFMO on tumor cellproliferation and migration was observed within 4and 2 days, respectively. The lag period observedcan not only be caused by limited drug penetrationinto the spheroids, but also because the enhanced

Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
120
synthesis of polyamines, precedes or coincides withthe initiation of DNA synthesis. Therefore, the cellcycle distribution within the spheroids may affectthe observed growth inhibitory effects [34].
The addition of putrescine completely reversedthe inhibition of migration as well as the antiprolif-erative effect of DFMO. These observations corre-spond well with earlier studies, where it has beenshown that putrescine, which is the immediate pre-cursor of spermidine, promptly replenishes the in-tracellular putrescine and spermidine pools revers-ing very effectively the DFMO effects [35].
Local invasion into normal brain tissue is an in-herent feature of malignant gliomas. Since DFMOextensively suppressed cell growth and migration(Figures 1 and 2), we studied whether DFMO af-fected invasion. The present work demonstratesthat DFMO did not inhibit invasion of any of thethree cell lines (Figure 3), despite the fact that thespheroids were pretreated with DFMO and com-pletely growth inhibited at the time of confronta-tion. Although the three cell lines exhibited differ-ent invasive properties, the DFMO effect seems tobe the same. The failure of DFMO to inhibit inva-sion of tumor cells is an observation of considerablebiological as well as clinical interest, and some ex-planations should be considered. The presence ofdifferentiating cells in the brain aggregate at thetime of confrontation [24] may be a source of fac-tors affecting mitosis [36] or migration [37]. It is alsoconceivable that metabolic cooperativity exists, im-plying that the normal non-proliferative brain tis-sue furnishes nutritional components, such as po-lyamines, to the malignant neighboring cancer cells.
In conclusion, the inhibitory effects observed bythe DFMO on tumor cell growth and migration didnot affect the invasiveness of the tumor cells duringthe time period studied. This finding indicates thatcell growth, migration and invasion are biologicalproperties that are not necessarily related to eachother.
Acknowledgements
This work was supported by grants from the De-partment of Neurosurgery, Lubeck, the Norwegian
Cancer Society, a donations from Inger Margretheand Per Jaeger, Frank Mohn A/S, and the Norwe-gian Research Council.
Technical assistance from Ms E. Pawlak is greatlyappreciated.
References
1. Chang CH, Horton J, Schoenfeld D, Salazer O, Perez-Tamayo R, Kramer S, Weinstein A, Nelson JS, Tsakuda Y:Comparison of postoperative radiotherapy and combinedpostoperative radiotherapy and chemotherapy in the multi-disciplinary management of malignant gliomas. Cancer 52:997–1007, 1983
2. EORTC Brain Tumor Group: Evaluation of CCNU, VM-26plus CCNU, and procarbazine in supratentorial brain glio-mas. J Neurosurg 55: 27–31, 1981
3. Walker MD: Brain tumor study group: a survey of currentactivities. Natl Cancer Inst Monogr 46: 209–212, 1977
4. Levin VA, Chamberlain MC, Prados MD, Choucair AK,Berger MS, Silver P, Seager M, Gutin PH, Davis RL, WilsonCB: Phase I/II study of the combination of DFMO andMGBG for the treatment of primary brain tumors. CancerTreat Rep 71: 459–464, 1987
5. Prados MD, Rodriguez L, Chamberlain MC, Silver P, LevinVA: Treatment of recurrent gliomas with 1,3-bis(2-chloro-ethyl)-1-nitrosourea and α-difluoromethylornithine. Neu-rosurgery 24(6): 806–809, 1989
6. Janne J, Poso H, Raina A: Polyamines in rapid growth andcancer. Biochim Biophys Acta 473: 241–293, 1978
7. Takami H, Romsdahl MM, Nishioka K: Polyamines in bloodcells as a cancer markers. Letter Lancet 2: 912–914, 1979
8. Caldarera CM, Barbiroli B, Moruzzi G: Polyamines and nu-cleic acids during development of the chick embryo. Bio-chem J 97: 84–88, 1965
9. Pegg AE, McCann PP: Polyamine metabolism and function.Am J Physiol 243: C212–C221, 1982
10. Luk GD, Casero RA: Polyamines in normal and cancer cells.Adv Enzyme Regul 26: 91–105, 1987
11. Behe M, Felsenfeld G: Effects of methylation on a syntheticpolynucleotide: the Z-transition in poly(dG-me5dC)-poly(dG-me5dC). Proc Natl Acad Sci USA 78: 1619–1623, 1981
12. Basu HS, Feuerstein BG, Deen DF, Lubich WP, BergeronRJ, Samejima K, Marton LJ: Correlation between the ef-fects of polyamine analogs on DNA conformation and cellgrowth. Cancer Res 49: 5591–5597, 1989
13. Tabor CW, Tabor H: Polyamines. Annu Rev Biochem 53:749–790, 1984
14. Pegg AE, Williams-Ashman HG: Biosynthesis of putrescin.In: Morris DR, Marton LJ (eds) Polyamines in Biology andMedicine. Marcel Dekker, Inc., New York, 1979, pp 3–42
15. Metcalf BW, Bey P, Danzin C, Jung MJ, Casara P, Vevert JP:Catalytic irreversible inhibition of mammalian ornithine de-

Please indicate author’s corrections in blue, setting errors in red
138167 NEON ART.NO 850-96 (684) ORD.NO 234684.Z
121
carboxylase (E.C.4.1.1.17) by substrate and products ana-logues. J Am Chem Soc 100: 2551–2553, 1978
16. Mamont PS, Duschene M-C, Grove J, Bey P: Antiprolifer-ative properties of D,L-α-difluoromethyl ornithine in cul-tured cells. A consequence of the irreversible inhibition ofornithine decarboxylase. Biochem Biophys Res Commun81: 58–66, 1978
17. Sunkara PS, Prakash NJ, Mayer GD, Sjoerdsma A: Tumorsuppression with a combination of α-difluoromethyl-orni-thine and interferon. Science 219: 851–853, 1983
18. Sunkara PS, Baylin SB, Luk GD: Inhibitors of polyaminebiosynthesis: cellular and in vivo effects on tumor prolifer-ation. In: McCann PP, Pegg AE, Sjoerdsma A (eds) Inhib-ition of Polyamine Metabolism, FL: Academic Press, Orlan-do, 1987, pp 121–140
19. Carlsson J, Nilsson K, Westermark B, Ponten J, SundstromC, Carlsson E, Bergh J, Pahlman S, Busch C, Collins VP:Formation and growth of multicellular spheroids of humanorigin. Int J Cancer 31: 523–533, 1983
20. Mueller-Klieser W: Multicellular spheroids. A review oncellular aggregates in cancer research. J Cancer Res ClinOncol 113: 101–122, 1987
21. Sutherland RM: Cell and environment interactions in tumormicroregions: The multicellular spheroid model. Science240: 177–184, 1988
22. Sutherland RM, Freyer J, Mueller-Klieser W: Cellulargrowth and metabolic adaptations to nutrient stress envi-ronments in tumor microregions. Int J Rad Oncol Biol Phys12: 611–615, 1986
23. Akslen LA, Andersen KJ, Bjerkvig R: Characteristics of hu-man and rat glioma cell grown in a defined medium. Anti-cancer Res 8: 797–804, 1988
24. Yuhas JM, Li AP, Martinez AO, Ladman AJ: A simplifiedmethod for production and growth of multicellular sphe-roids. Cancer Res 37: 3639–3643, 1977
25. Bjerkvig R: Reaggregation of fetal rat brain cells in a sta-tionary culture system. II: Ultrastructure characterisation.In vitro Cell Dev Biol 22: 193–200, 1986
26. Bjerkvig R, Steinvaag SK, Laerum OD: Reaggregation offetal rat brain cells in a stationary culture system. I: Method-ology and cell identification. In vitro Cell Dev Biol 22: 180–192, 1986
27. Honegger P, Lenoir P, Farvrod P: Growth and differentia-tion of aggregating fetal brain cells in serum free definedmedium. Nature 282: 305–307, 1979
28. Trapp BD, Honegger P, Richelson E: Morphological differ-entiation of mechanically dissociated fetal rat brain in ag-gregating cell cultures. Brain Res 160: 235–252, 1982
29. Trapp BD, Webster HD, Johnson D, Quarles RH, CohenSR, Murray MR: Myelin formation in rotation-mediatedaggregating cell cultures: Immunocytochemical, electronmicroscopic and biochemical observations. J Neurosci 2:986–993, 1982
30. Gundersen HJG, Jensen EB: The efficiency of systematicsampling in stereology and its prediction. J Microscopy 147:229–263, 1987
31. Pedersen PH, Rucklidge G, Mork S, Terzis AJA, Engebraa-ten O, Lund-Johansen M, Backlund EO, Laerum OD,Bjerkvig R: Leptomeningeal tissue: a barrier against gliomacell invasion. J Natl Cancer Inst 86: 1593–1599, 1994
32. Pedersen PH, Marienhagen K, Mork S, Bjerkvig R: The mi-gratory pattern of fetal rat brain cells and human gliomacells in the adult rat brain. Cancer Res 53: 5158–5165, 1993
33. Storme G, Mareel MM, De Bruyne G: Influence of cellnumber on directional migration of MO4 cells in vitro. ArchGesschwulstforsch 51: 45–50, 1981
34. Carlsson J, Nederman T: Tumor spheroids as a model instudies of drug effects. In: Bjerkvig R (ed) Spheroid Culturein Cancer Research. CRP Press, Boca Raton (FL), AnnArbor, London, 1992, pp 199–216
35. Mamont PS, Duschesne MC, Joder-Ohlenbusch AM, GroveJ: Effects of ornithine decarboxylase inhibitors on culturedcells. In: Seiler N, Jung MJ, Koch-Weser J (eds) Enzyme-activated Irreversible Inhibitors. Elsevier/North HollandBiomedical Press, Amsterdam, 1978, pp 43–54
36. Prehn RT: Many growth factors may not be growth factors.Cancer Res 52: 501–507, 1992
37. Lund-Johansen M, Bjerkvig R, Rucklidge GJ: Tumor sphe-roids from monolayer cultures. In: Bjerkvig R (ed) SpheroidCulture in Cancer Research. CRP Press, Boca Raton (FL),Ann Arbor, London, 1992, pp 199–216
Address for offprints: A-Jorge A. Terzis, Department of Neuro-surgery, University of Luebeck, Ratzeburger Allee 160, 23538Luebeck, Germany
Related Documents










![Research Paper A Noninvasive Assessment of Tumor Proliferation in Lung cancer … · 2020. 11. 3. · widely used to predict the prognoses of breast cancer [3], glioma [4], and lung](https://static.cupdf.com/doc/110x72/609e220c5a8cb135d265239a/research-paper-a-noninvasive-assessment-of-tumor-proliferation-in-lung-cancer-2020.jpg)
