Effects of bud phenology and foliage chemistry of balsam fir and white spruce trees on the efficacy of Bacillus thuringiensis against the spruce budworm, Choristoneura fumiferana Nathalie Carisey, E ´ ric Bauce, Alain Dupont* and Sylvain Miron Faculte ´ de Foresterie et de Ge ´omatique, CRBF, Universite ´ Laval, Sainte-Foy, Que ´bec, G1K 7P4, Canada and *Socie ´te ´ de Protection des Fore ˆts contre les Insectes et Maladies (SOPFIM), 1780 rue Semple, Que´bec, G1N 4B8 Abstract 1 Efficacy of commercial formulations of Bacillus thuringiensis ssp. kurstaki (Btk) against spruce budworm Choristoneura fumiferana was investigated in mixed balsam fir-white spruce stands. Btk treatments were scheduled to coincide with early flaring of balsam fir shoots, and later with flaring of white spruce shoots. Btk efficacy on the two host trees was compared and examined according to the foliar content of nutrients and allelochemicals and the insect developmental stage at the time of spray. 2 Larvae fed white spruce foliage were less vulnerable to Btk ingestion than larvae fed balsam fir foliage. Higher larval survival on white spruce, observed 10 days after spray, was related to higher foliage content in tannins and a lower N/ tannins ratio, which might have induced inactivation of Btk toxins. 3 Larval mortality due to Btk did not depend on spruce budworm larval age. 4 Foliage protection of both host trees was similar in plots treated with Btk: larval mortality due to Btk treatment reduced insect grazing pressure on balsam fir trees; meanwhile, suitability of white spruce foliage seemed to decrease very rapidly, which induced high larval mortality among spruce budworm fed on white spruce trees. Nevertheless, following Btk sprays, 50% more foliage remained on white spruce than on balsam fir trees, because of the higher white spruce foliage production. 5 Both spray timings achieved similar protection of white spruce trees, but Btk treatments had to be applied as early as possible (i.e. during the flaring of balsam fir shoots to optimally protect balsam fir trees in mixed balsam fir-white spruce stands). Keywords Bacillus thuringiensis, balsam fir, Btk efficacy, bud phenology, Choristoneura fumiferana, foliage chemistry, foliage production, spray timing, tannins, white spruce. Introduction The spruce budworm Choristoneura fumiferana (Clem.) is an oligophagous tortricid moth, common to the eastern boreal forests of North America, that feeds primarily on the current-year foliage of balsam fir, Abies balsamea (L.) Miller, and white spruce, Picea glauca (Moench) Voss (Blais, 1983; Sanders, 1991). Boulet (2001) reported that 180 million m 3 of timber were lost during the last spruce budworm outbreak (1967–92) in the Province of Quebec. Current-year foliage has to be protected against spruce budworm to prevent extensive defoliation and reduction in tree volume growth that lead to tree mortality after 4–5 years of severe annual defoliation (Hardy, 1979; Gagnon & Chabot, 1991). Forest protection against spruce budworm in the Province of Quebec (Canada) currently relies on aerial sprays of the microbial insecticide Bacillus thuringiensis Berliner ssp. kurstaki (Btk). Btk is a gram-positive soil bacterium that produces a proteinaceous crystalline Correspondence: Nathalie Carisey. Tel: þ1 418 656 2131 ext. 13850; fax: þ1 418 656 3177; e-mail: [email protected] Agricultural and Forest Entomology (2004) 6, 55–69 # 2004 The Royal Entomological Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Effects of bud phenology and foliage chemistry of balsam fir andwhite spruce trees on the efficacy of Bacillus thuringiensis againstthe spruce budworm, Choristoneura fumiferana
Nathalie Carisey, Eric Bauce, Alain Dupont* and Sylvain Miron
Faculte de Foresterie et de Geomatique, CRBF, Universite Laval, Sainte-Foy, Quebec, G1K 7P4, Canada and *Societe de Protection des Forets
contre les Insectes et Maladies (SOPFIM), 1780 rue Semple, Quebec, G1N 4B8
Abstract 1 Efficacy of commercial formulations of Bacillus thuringiensis ssp. kurstaki (Btk)against spruce budworm Choristoneura fumiferana was investigated in mixedbalsam fir-white spruce stands. Btk treatments were scheduled to coincide withearly flaring of balsam fir shoots, and later with flaring of white spruce shoots.Btk efficacy on the two host trees was compared and examined according to thefoliar content of nutrients and allelochemicals and the insect developmentalstage at the time of spray.
2 Larvae fed white spruce foliage were less vulnerable to Btk ingestion than larvaefed balsam fir foliage. Higher larval survival on white spruce, observed 10 daysafter spray, was related to higher foliage content in tannins and a lower N/tannins ratio, which might have induced inactivation of Btk toxins.
3 Larval mortality due to Btk did not depend on spruce budworm larval age.4 Foliage protection of both host trees was similar in plots treated with Btk:
larval mortality due to Btk treatment reduced insect grazing pressure on balsamfir trees; meanwhile, suitability of white spruce foliage seemed to decrease veryrapidly, which induced high larval mortality among spruce budworm fed onwhite spruce trees. Nevertheless, following Btk sprays, 50% more foliageremained on white spruce than on balsam fir trees, because of the higherwhite spruce foliage production.
5 Both spray timings achieved similar protection of white spruce trees, but Btktreatments had to be applied as early as possible (i.e. during the flaring ofbalsam fir shoots to optimally protect balsam fir trees in mixed balsam fir-whitespruce stands).
Keywords Bacillus thuringiensis, balsam fir, Btk efficacy, bud phenology,Choristoneura fumiferana, foliage chemistry, foliage production, spray timing,tannins, white spruce.
Introduction
The spruce budworm Choristoneura fumiferana (Clem.) is
an oligophagous tortricid moth, common to the eastern
boreal forests of North America, that feeds primarily on
the current-year foliage of balsam fir, Abies balsamea (L.)
Miller, and white spruce, Picea glauca (Moench) Voss
(Blais, 1983; Sanders, 1991). Boulet (2001) reported that
180millionm3 of timber were lost during the last spruce
budworm outbreak (1967–92) in the Province of Quebec.
Current-year foliage has to be protected against spruce
budworm to prevent extensive defoliation and reduction in
tree volume growth that lead to tree mortality after
4–5 years of severe annual defoliation (Hardy, 1979;
Gagnon & Chabot, 1991).
Forest protection against spruce budworm in the
Province of Quebec (Canada) currently relies on aerial
sprays of the microbial insecticide Bacillus thuringiensis
Berliner ssp. kurstaki (Btk). Btk is a gram-positive
soil bacterium that produces a proteinaceous crystallineCorrespondence: Nathalie Carisey. Tel:þ1 418 656 2131 ext. 13850;
fax:þ1 418 656 3177; e-mail: [email protected]
Agricultural and Forest Entomology (2004) 6, 55–69
# 2004 The Royal Entomological Society

inclusion during sporulation. This inclusion, the d-endotoxin,exhibits highly specific insecticidal activity against Lepi-
doptera (Beegle & Yamamoto, 1992; Dent, 1993). Improve-
ments have been made to Btk formulations, application
technology as well as knowledge of effective droplet size
and lethal dose, but Btk aerial sprays still show variable
efficacy, in part due to several constraints inherent to the
product (van Frankenhuyzen, 1993; van Frankenhuyzen
et al., 1995). First, Btk formulation needs to be ingested by
the larva to be effective. Once activated by the alkaline pH
and proteolytic activity of the insect gut fluids, the toxin
perforates the midgut epithelium, which leads to feeding
cessation and, ultimately, to death through septicemia
(Dent, 1993; van Frankenhuyzen, 1993). Second, foliage on
which the insect feeds needs to be exposed to the Btk aerial
spray, which is why Btk applications do not begin before the
flushing and flaring of new shoots (opened buds with well-
exposed needles). Third, Btk efficacy is potentially affected by
the nutritional and allelochemical composition of food
ingested by the insect. For example, the vulnerability of
gypsy moth (Lymantria dispar L.) and forest tent caterpillar
(Malacosoma distriaHubner) larvae to Btk appear to depend
on host plant species on which the insects grow (Moldenke
et al., 1994; Farrar et al., 1996; Kouassi et al., 2001). Bauce
et al. (2002) reported that spruce budworm larvae that were
fed artificial diet of medium quality were more vulnerable to
Btk than larvae that were fed high quality food. Tannins are
common secondary plant compounds that could reduce the
effectiveness of Btk formulations to L. dispar (Appel &
Schultz, 1994). Moreover, tannins have been shown to
negatively affect the efficacy of Btk d-endotoxins to both
Pieris brassicae L. (Luthy et al., 1985) andHeliothis virescens
Fabricius (Navon et al., 1993). Other chemicals have been
found to enhance Bt sp. effects. Cholorogenic acid and
polyphenol oxidase increase the toxicity ofBtk toHelicoverpa
armigeraHubner (Ludlum et al., 1991). Simple phenols, such
as gallic acid and resorcinol, have also been shown to increase
the activity of Bt ssp. galleriae endotoxin againstH. armigera
(Sivamani et al., 1992).
Most of Canada’s boreal forests are mixed coniferous
stands in which host trees, such as balsam fir and white
spruce, display considerable variation in phenotype, bud
phenology, foliage production and vulnerability to spruce
budworm (Blais, 1976; Hardy, 1979;MacLean&MacKinnon,
1997). Because of the complexity of Btk mode of action, the
challenge of optimally protecting these stands relies on an
understanding of the tri-trophic relationships among Btk,
spruce budworm and host trees. Budbreak in white spruce
occurs 1–4 days later than in balsam fir (Blais, 1957;
Greenbank, 1963). Although balsam fir needles flare quite
rapidly after budbreak, the protective bud scales of white
spruce persist on the tips of the expanding shoots for a long
time following budbreak (Dimond, 1985; Volney & Cerezke,
1992). Thus, the appropriate timing ofBtk applicationsmay be
later for white spruce than for balsam fir (Dimond, 1985). The
first objective of this study was to determine which Btk spray
timing, based on the flaring of balsam fir and white spruce
needles, optimally protects mixed coniferous stands. The
second objective was to compare Btk efficacy between the
two host trees based on foliar nutritional and allelochemical
composition. Lysyk (1989) demonstrated that phenological
development of spruce budworm larvae was different depend-
ing on whether they fed on balsam fir or white spruce trees,
whereas van Frankenhuyzen et al. (1997) showed that lethal
dose requirements per larva increased with larval stage. To
dissociate the influence of foliage quality from that of larval
stage on Btk efficacy, a laboratory experiment using foliage
from the field was conducted. The aims of this study were (i) to
examine Btk efficacy at different larval instars (third, fourth,
fifth and sixth instars) relative to host trees; (ii) to determine
the ingested amounts of Btk-contaminated foliage; and (iii) to
define relationships between Btk mortality rate and amounts
of nutrients (N, P, K, Ca, Mg, total soluble sugars) and
allelochemical compounds (total tannins, total phenolics)
ingested per larva.
Materials and methods
Field experiment
Site description. The experimental site is located in the
Ottawa River Valley of Quebec, Canada (between 45�380-46�010N and 75�330 to 76�330W; mean altitude, 165m asl).
Spruce budworm populations have been at epidemic levels
in the mixed wood stands of this region since 1992. Twenty-
four plots were selected according to the following criteria:
high second-instar spruce budworm populations in autumn;
more than 30% of the basal area in balsam fir and white
spruce; 30–50-year-old spruce-fir stands; moderate previous
defoliation; and easy site access. Plot areas averaged 25 ha.
In each plot, 12 balsam fir and 12 white spruce codominant
trees were randomly selected along transects perpendicular
to the flight lines of the aircraft applying Btk formulations.
Experimental design. Three factors were studied: (i) the
scheduling of the first Btk formulation application that cor-
responded to the moment when buds of balsam fir and white
spruce trees reached the fourth phenological bud stage
(described in the next paragraph); (ii) Btk application rates
that consisted of 0, 1 or 2 applications (with 8–13 days apart)
of 30 billion international units per hectare (BIU/ha); and (iii)
host trees, balsam fir vs. white spruce. Each interaction ofBtk
schedule�Btk application rate, was replicated in four ran-
domly allocated plots. The experimental design is summar-
ized in Table 1.
Bud phenology. The condition of balsam fir buds and
shoots was described according to the scheme of Auger
(Dorais & Kettela, 1982; Juneau, 1989): Stage 1, no appar-
ent bud development (bud in winter condition); Stage 2,
buds swelling and 10–35% of the needles visible; Stage 3,
budbreak, all needles visible but not flaring; Stage 4, needles
flushing and flaring and shoot elongation initiated; Stage 5,
shoots are supple and undergoing elongation.
The phenological stages of white spruce buds and shoots
were adapted from our own observations and from schemes
used by Volney & Cerezke (1992) and Lawrence et al.
(1997): Stage 1, no apparent bud development (bud in
winter condition); Stage 2, buds swelling without separation
of bud scales; Stage 3a, green buds with expanding and
56 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

thinning scales, but still intact; Stage 3b, all needles are
visible with bud cap separated from the twig base (bud-
break); Stage 4, needles flaring with or without bud cap;
Stage 5, shoot elongation.
Btk formulation applications. Foray 76B, a Btk strain
HD-1 commercial formulation at nominal potency of
20.0BIU/L [Abbott Laboratories (Chicago, IL) on behalf
of Valent Bio-Sciences Corporation (Libertyville, IL)] was
applied to the test plots, described above. Two aircraft were
used, a Piper-Pawnee PA-25 and a Dromader M-18, which
were equipped with six and eight Micronair atomizers
(Micronair Sprayers Ltd, Bromyard, U.K.), respectively.
Micronair atomizers, spinning at 3195 g, were located
within 75% of the total wingspan. The Piper-Pawnee and
Table 1 Summary of the experimental design for the field experiment
Factor 1 Factor 2 Factor 3
Btk schedule No of Btk applications of 30BIU/ha Date of Btk sprays No of plots Host tree
Fourth phenological bud 0 – 1 12 balsam fir
stage of balsam fir tree 12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 28–29 May 2000 1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
2 28–29 May 2000 1 12 balsam fir
7-11 June 2000 12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
Fourth phenological bud
stage of white spruce tree
0 – 1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 3–4 June 2000 1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
2 3–4 June 2000 1 12 balsam fir
11�12 June 2000 12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
1 12 balsam fir
12 white spruce
Btk efficacy against spruce budworm in mixed coniferous stand 57
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

theDromaderwere flownat 161km/hand193km/h,with30-m
and50-m spraywidths, respectively.Aircraft spraying occurred
early in the morning or at dusk under good weather conditions
(wind speed¼ 6km/h; no rain). The flow rate through the
nozzles was calibrated to deliver 1.5L/ha.
Evaluation of treatment efficacy. A 45-cm branch tip was
collected in the midcrown of the 12 balsam fir and 12 white
spruce trees in each plot on three occasions: 24 h before the
first application (Day 0), 10 days after the first application
(just before the second spray in plots that received two
successive Btk applications; day 10) and at the time when
85% of the larvae had reached the pupal stage (pupal stage
day). Numbers of spruce budworm larvae and their larval
instars (McGugan, 1954) were recorded per branch; an
insect development index (IDI) was calculated according
to the methods of Dorais & Kettela (1982) and Juneau
(1989) where:
IDI ¼ [(no. second-instar larvae � 2) þ (no. third-instar
larvae � 3) þ (no. fourth-instar larvae � 4) þ (no. fifth-
instar larvae � 5) þ (no. sixth-instar larvae � 6) þ (no.
pupae larvae � 7) þ (no. moths � 8)]/(total larvae/branch)
Mortality rates observed on day 10 and pupal stage day
corresponded to larval density reductions between day 0
and day 10 and pupal stage day, respectively.
The phenological bud and shoot stage per branch was
recorded on day 0 and a bud development index (BDI) was
calculated according to the methods of Dorais & Kettela
(1982):
For balsam fir tree, BDI ¼ [(no. first � stage
buds � 1) þ (no. second � stage buds � 2) þ (no. third �stage buds � 3) þ (no. fourth � stage buds � 4) þ (no.
fifth � stage buds � 5)]/(total buds/branch)
For white spruce tree, BDI ¼ [(no. first�stage
buds � 1) þ (no. second�stage buds � 2) þ (no. third (a)
�stage buds � 3) þ (no. third (b)�stage buds � 3.5) þ (no.
fourth�stage buds � 4) þ (no. fifth�stage buds � 5)]/(total
buds/branch)
Bud and shoot defoliation was estimated by the method
of Fettes (1950) (Dorais & Hardy, 1976; Sanders, 1980) on
day 0, day 10 and pupal stage day.
The length and number of new shoots were recorded for
branches collected on pupal stage day. Three shoots per tree
(three trees per host tree and plot) were used to estimate
(i) the number of needles per centimeter of shoot (no.
needles/cm); (ii) the mean dry weight of one needle (mg/
needle); (iii) the dry weight of one shoot axis (mg/shoot
axis); and (iv) the length of one shoot axis (cm/shoot axis).
Foliage was dried in an oven at 70 �C for 3 days. The mean
dry weight of shoot per centimetre (mg/cm) was calculated as:
[(no. needles/cm) � (mg/needle)] þ [(mg/shoot axis)/(cm/
shoot axis)]
Subsequently, to calculate the total dry weight of current-
year foliage produced per branch, the mean dry weight of
shoot per centimetre was multiplied by the mean shoot
length and number of shoots per branch.
Statistical analysis. Bud development index (BDI), insect
development index (IDI) and larval density data on day 0
were subjected to analyses of variance (PROC GLM; SAS
Institute Inc., 1988) in a three-stage crossed nested design.
Four replicate plots were nested within the Btk schedule
(two timings) and Btk application rate (three treatment
levels) factors. The host tree factor was crossed with plots
nested within the Btk schedule and Btk application rate
factors. Larval mortality, defoliation and foliage produc-
tion data were submitted to analyses of covariance with the
initial larval density as a covariate in the previously
described design and are summarized in Table 2. The aver-
age of each parameter, measured on a tree, was firstly
calculated per each host tree and plot. The LSMEANS state-
ment (SAS Institute Inc., 1988), performed for each effect
and interaction, computed least-squares means and multiple
comparisons (least significant difference; LSD), with Bon-
ferroni adjustment for the P-value and confidence limits for
the differences of least squares-means (Bonferroni method
of pairwise comparisons where the adjusted P-value corres-
ponds to the traditional P-value of 0.05 divided by the
number of pairwise comparisons; SAS Institute Inc., 1988).
Foliar bioassays in laboratory
Foliage treatment. Two plots were selected according to
criteria described in the first experiment. The first plot had
Table 2 Sources of variation of the analysis of covariance in the three-stage crossed nested design used for the field experiment
Sources of variation Degrees of freedom
Initial larval density (covariate) 1
Btk schedule (a�1)¼ 1
Btk application rate (b� 1)¼ 2
Btk schedule�Btk application rate (a�1)(b� 1)¼2
Error (a)
Plots� (Btk schedule�Btk application rate) (r�1)ab¼ 3�2� 3¼18
Host tree (c�1)¼ 1
Host tree�Btk schedule (c�1)(a� 1)¼ 1
Host tree�Btk application rate (c�1)(b� 1)¼2
Host tree�Btk schedule�Btk application rate (c�1)(a� 1)(b� 1)¼ 2
Error (b)
Plots�Host tree� (Btk schedule�Btk application rate) (r�1)(c� 1)ab – 1¼ 17
Total rabc – 1¼ 47
58 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

no Btk treatment whereas the second was sprayed with
30BIU/ha on 3 June 2000, after white spruce needles had
flushed and flared. The Btk strain HD-1 commercial for-
mulation was Foray 48B (Abbott Laboratories on behalf of
Valent Bio-Sciences Corporation) at nominal potency of
12.7 BIU/L and delivered at a rate of 2.37L/ha. In each
plot, five balsam fir and five white spruce trees were ran-
domly selected. In the treated plot, trees were situated along
lines perpendicular to the flight direction of the aircraft, a
Piper-Pawnee PA-25.
Pre-spray foliage collection and laboratory insect rearing. On
20 May 2000, 25 mid-crown 45-cm branches per host
tree were harvested from different trees in the two selected
plots to sample and rear spruce budworm larvae in the
laboratory; foliage from each tree species was collected
every two days.
A total of 1000 spruce budworm larvae per each host tree
were sampled from branches and reared in 180mL clear
plastic containers (15 larvae per container) filled with either
balsam fir or white spruce buds. A dampened piece of filter
paper was placed on the bottom of the container. Foliage in
each container was replaced every 2 days with fresh material.
For the experiment with postspray foliage (described below),
similar numbers of larvae per each instar (third, fourth, fifth
and sixth) were required at the same moment. Thus, it was
necessary to slow down or increase larval development, and
clear plastic containers filled with larvae were randomly
distributed in four different thermo-electric coolers
(KoolatronsTM, Rochester, U.S.A.) set up at 10, 15, 20 and
25 �C, respectively, for 10–12 days prior to use in the experi-
ment with postspray foliage. These different temperature
treatments (cold or warm conditions) could not have affected
the further results of the experiment, because, first, spruce
budworm larvae in the field are well-adapted to undergo
periods of cold or warm temperatures and, second, larvae
fed on the two host trees were reared in the same thermo-
electric cooler that corresponded to one specific instar.
Postspray foliage collection and laboratory insect rear-
ing. Four hours after the Btk application, five branches
per tree were harvested in the five balsam fir and white
spruce trees previously selected. The current-year shoots
from the five branches per tree were clipped, stored in
plastic boxes and taken to the laboratory where resident
spruce budworm larvae were carefully removed from shoots
to avoid tearing needles and crushing Btk droplets, and
were used as described below.
First, the shoots were used to estimate the foliar water
content. Five shoots per tree were weighed, freeze-dried and
weighed again.
Second, the shoots were used to measure the number of
needles per shoot, the mean dry weight of one needle and
the needle and shoot areas. One hundred needles collected
on five shoots from each tree were freeze-dried and weighed.
Moreover, the projected area of each needle was estimated
using a Digital Image Analysis System (WinFoliaTM,
Regent Instruments Inc., Canada) after placing 50 needles
sampled from each tree on a white sheet. Balsam fir and
white spruce needles have two and four sides, respectively.
Thus, the mean projected area of a needle, for balsam fir
and white spruce was multiplied by 2 and 4, respectively, to
obtain the appropriate needle surface areas. The mean leaf
area of one shoot (mm2) was calculated by multiplying the
needle mean area by the mean number of needles per shoot.
Third, the shoots were used to analyse for N, P, K, Ca,
Mg, total soluble sugar, total tannin and total phenolic
contents, according to methods described by Bauce et al.
(1994) and Bauce (1996). Total tannin concentrations were
quantified by the radial diffusion method (Dement &
Mooney, 1974; Wisdom et al., 1987), which relies on the
formation of binding complexes between tannins and
bovine serum albumin (BSA) protein in agar medium
(Hagerman, 1987). A known quantity of foliage extract is
deposited in a well, cut into agar containing BSA, and the
area of radial diffusion is measured. Thus, total tannin
concentrations are expressed in cm2/mg dry weight foliage.
Fourth, the shoots were used to quantify the Btk toxins
deposited on foliage using the Abbott Deposit Assessment
Method (ADAMKITTM
). This kit, developed on behalf of
Abbott Laboratories by Agdia Inc. (Elkhart, U.S.A.), is
based on the enzyme linked immunosorbent assay (DAS
ELISA) method (van Frankenhuyzen et al., 1998). Five
shoots per tree were placed together in a 50-mL milk sample
bag in which 5mL of solubilization buffer (0.05M trisodium
phosphate, pH12.0, PBS-TA) were added and incubated at
room temperature for 1 h. After agitation, a 1-mL aliquot
was transferred to a 2-mL microcentrifuge tube and an
equal volume of neutralization buffer (0.05M monosodium
phosphate, pH2.2, PBS-TN) was added to obtain a final
pH of 7.5 (van Frankenhuyzen et al., 1998). Because the
foliar samples were too concentrated in Btk toxins to be
directly submitted to the ELISA research kit, they were
diluted with a solution comprising half trisodium phosphate
(pH12.0, PBS-TA) and half monosodium phosphate (pH2.2,
PBS-TN) buffers. For the standard curve, samples of Btk
commercial formulation were collected in the aircraft tank
after spraying, and Btk deposit was expressed in terms of
IU/shoot. Then, the mean quantity of IU deposited on
foliage surface (IU/mm2) per each tree was calculated by
dividing the mean IU/shoot by the mean leaf area of one
shoot (mm2).
Five, the shoots were used to feed the insects and study
the influence of the two host trees on the effect of Btk on
spruce budworm larvae. Shoots collected from each tree
were randomly distributed in four 180mL clear plastic
containers. Fresh shoots in each cup were weighed before
larvae were added. The four containers per host tree
received 15 third-, fourth-, fifth- and sixth-instar larvae,
respectively. The containers were incubated for 5 days at
25 �C, after which the number of dead and surviving larvae
were counted; the remaining foliage was dried (oven at
70 �C for 3 days) and weighed to determine foliage
ingestion. Five days before the Btk treatment, similar
manipulations were carried out with foliage collected in
the control plot. This interval of 5 days between the two
treatments was due to bad weather conditions that delayed
the Btk aerial spray.
Btk efficacy against spruce budworm in mixed coniferous stand 59
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

Quantity of foliage ingested per larva. Initial fresh foliage
weight, dry weight of the remaining foliage, foliar water
content and mean needle dry weight were used to determine
the number of needles ingested per container (IN), and the
mean number of needles ingested per larva (IN15) when IN
was divided by the 15 spruce budworm larvae. The amounts
of each nutritional (N, P, K, Ca, Mg and total soluble
sugars) and allelochemical (total phenols and tannins) com-
pounds ingested per larva were calculated by multiplying
the nutritional and allelochemical compound concentration
by the amount of ingested foliage dry weight per larva.
Total needles Ingested/container ¼ IN ¼ [([(fresh foliage
weight on day 0a) � (100 � foliar water content)]/100) �remaining foliage dry weight on day 5a)]/mean needle dry
weight
Mean number of needles ingested/larva tested ¼ IN15
¼ IN/15; where 15 ¼ number of larvae per container at the
beginning of the test
Nitrogen consumed (mg)/larva tested ¼ [([(fresh foliage
weight on day 0a) � (100 � foliar water content)]/100 �remaining foliage dry weight on day 5a)/15] � N%; idem
for P, K, Ca, Mg, soluble sugars and phenols consumed
Tannins consumed (cm2)/larva tested ¼ [([(fresh foliage
weight on day 0a) � (100 � foliar water content)]/100 �remaining foliage dry weight on day 5a)/15] � tannins
(cm2/mg)b
[aper container of 15 larvae; bsee earlier for the method to
quantify tannins in foliage]
Statistical analyses. Mean number of needles ingested/
larva tested (IN15) (Btk�treated and control larvae) data
were subjected to an analysis of variance (PROC GLM; SAS
Institute Inc., 1988) in a three-stage crossed nested design
with trees (5 trees) nested within the Btk treatment (control
and Foray 48B) and host tree (two species) factors. The
larval instar factor (four instars) was crossed with trees
nested within the Btk treatment and host tree factors.
IN15 data were ln-transformed to meet assumptions of
normality and homogeneity of variances. The LSMEANS
statement (SAS Institute Inc., 1988), performed for each
effect and interaction, computed least-squares means and
multiple comparisons (Bonferroni method of pairwise com-
parisons).
Mortality rate
No larva died in the control containers during the assay
period and therefore these control treatments were excluded
from the analysis of variance. The mortality data were
subjected to an analysis of variance with a two-stage crossed
nested design with tree samples (five trees) nested within the
host tree factor (two species) (error term for the host tree
factor) and the larval instar factor (four instars) crossed
with trees and nested within the host tree factor (error
term for the larval intar effect and the host tree–larval
intar interaction).
Mean number of needles ingested/larva tested (IN15)
(Btk-treated larvae) data were ln-transformed and subjected
to the previously described analysis of variance with a
two-stage crossed nested design. Transformed data were
normally distributed and homoskedastic.
Mean leaf area of one shoot and IU/mm2 data were
subjected to an analysis of variance in a complete rando-
mized design with host tree as factor and using Btk-treated
trees as experimental units. Data had normal distribution
and homogeneous variances.
Nutritional and allelochemical compound contents were
measured for the two host trees from the control and treated
plots. Due to a 5-day delay between the control and Btk-
treated foliage collection, the analysis of these data indicated
changes in foliar compound contents in time. Thus, data
were subjected to an analysis of variance in a 2� 2 factorial
design with host tree factor (balsam fir and white spruce) and
collection day factor (29 May vs. 3 June) using trees as
experimental units.
Relationships between mortality rate and quantities of
nutritional and allelochemical ingested compounds per
tested larva (independent variables) were analysed by linear
regression using stepwise method at P< 0.1 (PROC STEPWISE;
SAS Institute Inc., 1988). Separate regression analyses were
performed on fifth- and sixth-instar data sets, respectively.
Results
Field experiment
Bud development index (BDI) at Btk application
timing. Six days separated the two Btk timings and within
this period, differences between balsam fir and white spruce
phenological shoot stages were nearly halved. At the balsam
fir Btk spray timing, balsam fir shoots were 14% more
developed than white spruce shoots. At the white spruce
Btk spray timing, they were just 8% more developed than
white spruce shoots (Tables 3 and 4). The first Btk applica-
tion was delayed 4–5 days due to bad weather conditions,
and explains why the Btk sprays were carried out when
balsam fir and white spruce shoots had exceeded the stage
4 (Table 4). Nevertheless, this delay could not significantly
affect conclusion on Btk timing: by the time, most balsam
fir shoots were at stage 4, the white spruce bud development
index was equal to 3.68 (data not shown). This index was
very close to that of white spruce (3.98) when Btk sprays
timed on balsam fir schedule were conducted (Table 4). This
small difference in shoot development is unlikely to have
affected the number of Btk droplets per area unit.
Insect development index (IDI) at Btk application
timing. At the time of balsam fir Btk sprays, spruce bud-
worm larvae fed on balsam fir or white spruce trees showed
similar larval development. Six days later, at the time of the
white spruce Btk sprays, larvae fed on white spruce trees
appeared to have grown more rapidly than on balsam fir
trees (Tables 3 and 4).
Initial larval density. Larval densities (larvae per 45-cm
branch tip) evaluated on balsam fir trees were similar at the
time of balsam fir and white spruce Btk sprays, but larval
density on white spruce increased from the balsam fir Btk
spray timing to the white spruce Btk spray timing (Tables 3
and 4). During their larval development, spruce budworm
60 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

larvae tended to move vertically as well as horizontally
within trees, as reported previously by Regniere et al.
(1989). Because the initial larval density (i.e. density one
day before the first Btk sprays) depended on host tree and
Btk schedule, which could affect larval mortality and defo-
liation, initial larval density was used as a covariate for the
analyses of covariances of the different parameters mea-
sured.
Larval mortality. Larval mortality (%) observed 10 days
after the first Btk application was significantly influenced by
the Btk application rate, the effect of which depended on the
Btk schedule (Table 5). In the controls, larval mortality rate
observed on day 10 was higher at the white spruce Btk spray
timing than at the balsam fir Btk spray timing (Table 5;
Fig. 1), which is consistent with literature that reported
increase in spruce budworm natural mortality during the
season (Royama, 1984; Regniere & You, 1991). Btk sprays,
carried out at the balsam fir timing, induced higher larval
mortality than those carried out at the white spruce timing
(Fig. 1). Larval mortality, observed on day 10, depended on
both host tree and Btk application rate factors (Table 5). On
this day, differences in larval mortality between balsam fir
and white spruce trees were 42% and 64%, for the control
and Btk-treated plots (at this time, all treated plots had
received only one Btk application), respectively (Fig. 2a).
On average, larval mortality in Btk-treated plots was
much higher on balsam fir trees (approximately 66%) than
on white spruce trees (around 24%) (Fig. 2a). In fact, Btk
application rate did not influence significantly larval mor-
tality from day 0 to day 10 on white spruce trees (Fig. 2a).
Btk spray timing did not influence larval mortality from
day 0 to pupal stage day (Table 5), but mortality during this
period was higher on white spruce control trees than on
balsam fir control trees (Table 5; Fig. 2b). By contrast, one
or two Btk applications induced higher larval mortality on
balsam fir trees (90%) than on white spruce trees (84%)
(Fig. 2b).
Defoliation. Analysis of covariance indicated that initial
defoliation (day 0) was higher in control plots than in Btk-
treated plots (Tables 6 and 7). The choice of plots in the
Ottawa River Valley was subjected to logistic constraints;
some owners did not allow SOPFIM to conduct Btk sprays
on their forests, which then were used as control plots.
Although the result was not statistically significant, these
control plots had 62% higher larval population densities
than plots dedicated to the Btk treatment (Table 3), and
this might have influenced the results of the initial defolia-
tion. Pre-spray defoliation was nearly two-fold higher at
white spruce Btk spray timing than at balsam fir Btk spray
timing (Tables 6 and 7).
Btk schedule and Btk application rate factors signifi-
cantly influenced defoliation observed on day 10 (Table 6).
Table 3 Summaries of the analyses of variance, in a three-stage crossed nested design, on bud development index (BDI), insect development
index (IDI) and larval density measured at the beginning of the field experiment (day 0)
BDI (Day 0) IDI (day 0)
Larval density
(larvae/45-cm branch tip) (day 0)
Sources of variation d.f. P MSE P MSE P MSE
Btk schedule 1 0.0005 0.0001 0.64
Btk application rate 2 0.74 0.13 0.07
Btk schedule�Btk application rate 2 0.95 0.99 0.66
Mean square error (a) 18 0.0593 0.0335 801.76
Host tree 1 0.0001 0.0001 0.0001
Host tree�Btk schedule 1 0.008 0.008 0.005
Host tree�Btk application rate 2 0.53 0.89 0.22
Host tree�Btk schedule�Btk application rate 2 0.37 0.4 0.44
Mean square error (b) 18 0.0141 0.0056 68.37
d.f., P-values for F-tests and the mean square errors (MSE) used for the F-tests (in italic). Numbers in bold indicate statistically significant effects
at P< 0.05. When interactions are significant, the corresponding simple effects are not in bold type.
Table 4 Statistically significant interaction (P< 0.05) between Btk schedule (Balsam fir, BF) and white spruce (WS) timing, and host tree on
foliage and larval population characteristics measured on day 0 in the field experiment.
Btk schedule� host tree
Day 0 BF timing�BF BF timing�WS WS timing�BF WS timing�WS
Bud development index 4.55�0.07b 3.98� 0.07d 4.75� 0.07a 4.38�0.07c
Insect development index 4.21�0.04c 4.27� 0.04c 4.89� 0.04b 5.07�0.04a
Larval density 35�5b,c 42� 5b 32� 5c 54�5a
(larvae/45-cm branch tip)
Mean�2SE followed by the same letter do not differ significantly according to the Bonferroni pairwise comparison method.
Btk efficacy against spruce budworm in mixed coniferous stand 61
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

Defoliation on day 10 was 247% higher in the control plots
than that in the Btk-treated plots (Table 7), and this defoli-
ation was nearly two-fold higher at white spruce timing
than at balsam fir timing (Table 6). Defoliation from day 0
to day 10 increased only in control plots.
At the end of larval development, Btk application rate
resulted in different levels of foliage protection against
spruce budworm, depending on the host trees (Table 6).
Although defoliation of balsam fir and white spruce trees
was similar in plots sprayed with one or two Btk applica-
tions, defoliation in control plots was higher on balsam fir
trees than on white spruce trees (Fig. 3a). Defoliation
observed on pupal stage day was 133% higher in the control
plots than that in plots treated with one or two Btk sprays
(Fig. 3a). Moreover, at the pupal stage day, defoliation on
balsam fir trees was higher when Btk sprays were carried
out at white spruce timing instead of balsam fir timing.
Defoliation on white spruce trees was not influenced by
the Btk schedule (Table 6; Fig. 4).
Table 5 Summaries of the analyses of covariance, in a three-stage crossed nested design, on larval mortality measured on day 10 and pupal
stage day
Mortality (%) 10days
after the first Btk spray
Mortality (%) from the first Btk
spray (day 0) to pupal stage day
Sources of variation d.f. P MSE P MSE
Initial larval density (covariate) 1 0.0019 0.0044
Btk schedule 1 0.66 0.64
Btk application rate 2 0.016 0.08
Btk schedule�Btk application rate 2 0.011 0.32
Mean square error (a) 18 192.88 53.92
Host tree 1 0.0002 0.47
Host tree�Btk schedule 1 0.95 0.08
Host tree�Btk application rate 2 .0016 0.0001
Host tree�Btk schedule�Btk application rate 2 0.53 0.69
Mean square error (b) 17 125.35 14.01
d.f., P-values for F-tests and the mean square errors (MSE) used for the F-tests (in italic). Numbers in bold indicate statistically significant effects
at P< 0.05. When interactions are significant, the corresponding simple effects are not in bold type.
Figure 1 Larval mortality (%) on both host trees 10days after the first
Btk spray, according to the three Btk application rates (control,
1�30BIU, and 2�30BIU, respectively, 0, 1, and 2 applications of
30BIU/ha) and the Btk schedule based on bud flaring stage of either
balsam fir (BF) or white spruce (WS).
100
90
80
70
60
50
40
30
20
10
0Control 1×30BIU 2×30BIU
±2SE
Balsam firWhite spruce
(a)
Larv
al m
orta
lity
(%)
from
Day
0 to
Day
10
100
90
80
70
60
50
40
30
20
10
0Control 1×30BIU 2×30BIU
±2SE
(b)
Larv
al m
orta
lity
(%)
from
Day
0 to
Pup
al S
atge
Day
Figure 2 (a) Larval mortality (%) 10 days after the first Btk spray and
(b) larval mortality (%) at pupal stage day depending on three Btk
application rates and the host trees.
62 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

Table
6Summariesoftheanalysesofcovariance,in
athree-stagecrossednesteddesign,ondefoliation(%
)measuredonday0,day10andpupalstagedayandonremainingfoliage(g/45-cm
branchtip)
Defoliation(%
)onday0
Defoliation(%
)onday10
Defoliation(%
)atpupalstageday
New
foliageproduction
(g/45-cm
branchtip)
Remainingnew
foliage
(g/45-cm
branchtip)
Sourcesofvariation
d.f.
PMSE
PMSE
PMSE
PMSE
PMSE
Initiallarvaldensity(covariate)
1<0.0001
<0.0001
<0.0001
–<0.0001
Btk
schedule
10.0018
<0.0001
0.4
0.13
0.11
Btk
applicationrate
20.03
0.0002
0.001
0.006
0.03
Btk
schedule
�Btk
applicationrate
20.39
0.08
0.37
0.5
0.38
Meansquare
error(a)
86
18
0.0057
0.0138
224.6
213.86
130.56
Hosttree
10.64
0.3
0.68
<0.0001
0.02
Hosttree�Btk
schedule
10.8
0.09
0.049
0.21
0.52
Hosttree�Btk
applicationrate
20.23
0.11
0.0003
0.041
0.49
Hosttree�Btk
schedule�
Btk
applicationrate
20.23
0.09
0.96
0.5
0.62
Meansquare
error(b)
17
0.0024
0.0034
49.2
63.32
46.45
d.f.,P-valuesforF-testsandthemeansquare
errors
(MSE)use
dfortheF-tests(inita
lic).Ananalysisofvariancewasperform
edonnewfoliageproductiondata
because
theinitiallarvald
ensity
usedas
covariate
wasnotstatistically
significant.Numbers
inbold
indicate
statistic
ally
significanteffects
atP<0.05.Wheninteractio
nsare
significant,thecorresp
ondingsim
ple
effects
are
notin
bold
type.
Table
7Statistically
significanteffects
(atP<0.05)ofBtk
schedule,Btk
applicationrate
orhosttreeondefoliation(%
)measuredonday0andday10andonamounts
ofremainingfoliage
(g/45-cm
branchtip)atpupalstageday
Btk
schedule
Btk
applicationrate
Hosttree
BFtiming
WStiming
Control
1�30BIU
2�30BIU
Balsam
fir
Whitespruce
Defoliationonday0
12�4b
21�4a
26�6a
13�5b
12�5b
––
Defoliationonday10
22�5b
40�5a
59�11a
17�8b
17�7b
––
Remainingfoliageonpupalstageday
––
6.57�11.08b
24.35�7.76a,b
29.27�7.48a
15.41�4.09b
24.71�3.99a
Mean�2SEfollowedbythesameletterwithin
thesamerowperfactordonotdiffersignificantlyaccordingto
theBonferronip
airwisecomparisonmethod.Horizontalb
ars
indicate
thattheeffectof
this
factorwasnotstatistically
significant.BF,Balsam
fir;WS,whitespruce.
Btk efficacy against spruce budworm in mixed coniferous stand 63
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69
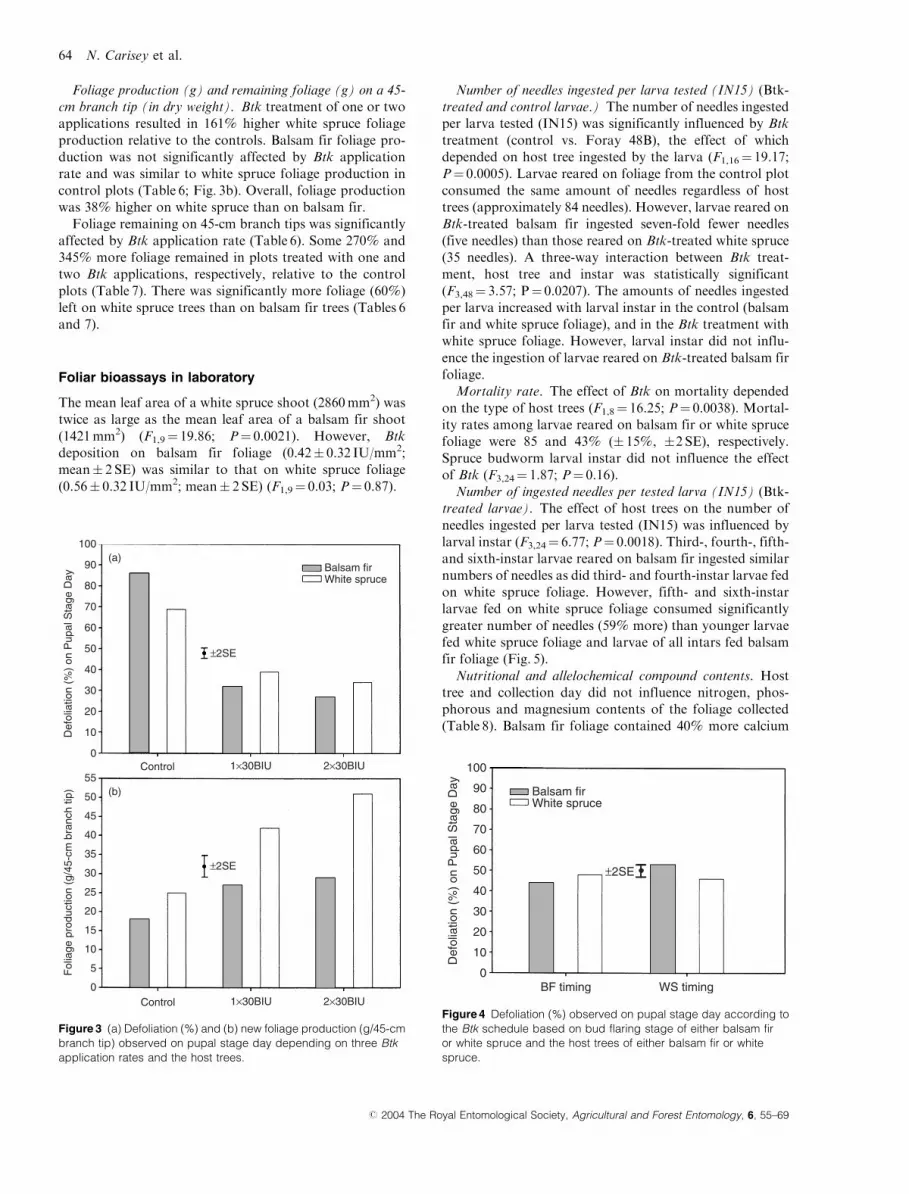
Foliage production (g) and remaining foliage (g) on a 45-
cm branch tip (in dry weight). Btk treatment of one or two
applications resulted in 161% higher white spruce foliage
production relative to the controls. Balsam fir foliage pro-
duction was not significantly affected by Btk application
rate and was similar to white spruce foliage production in
control plots (Table 6; Fig. 3b). Overall, foliage production
was 38% higher on white spruce than on balsam fir.
Foliage remaining on 45-cm branch tips was significantly
affected by Btk application rate (Table 6). Some 270% and
345% more foliage remained in plots treated with one and
two Btk applications, respectively, relative to the control
plots (Table 7). There was significantly more foliage (60%)
left on white spruce trees than on balsam fir trees (Tables 6
and 7).
Foliar bioassays in laboratory
The mean leaf area of a white spruce shoot (2860mm2) was
twice as large as the mean leaf area of a balsam fir shoot
(1421mm2) (F1,9¼ 19.86; P¼ 0.0021). However, Btk
deposition on balsam fir foliage (0.42� 0.32 IU/mm2;
mean� 2 SE) was similar to that on white spruce foliage
(0.56� 0.32 IU/mm2; mean� 2 SE) (F1,9¼ 0.03; P¼ 0.87).
Number of needles ingested per larva tested (IN15) (Btk-
treated and control larvae.) The number of needles ingested
per larva tested (IN15) was significantly influenced by Btk
treatment (control vs. Foray 48B), the effect of which
depended on host tree ingested by the larva (F1,16¼ 19.17;
P¼ 0.0005). Larvae reared on foliage from the control plot
consumed the same amount of needles regardless of host
trees (approximately 84 needles). However, larvae reared on
Btk-treated balsam fir ingested seven-fold fewer needles
(five needles) than those reared on Btk-treated white spruce
(35 needles). A three-way interaction between Btk treat-
ment, host tree and instar was statistically significant
(F3,48¼ 3.57; P¼ 0.0207). The amounts of needles ingested
per larva increased with larval instar in the control (balsam
fir and white spruce foliage), and in the Btk treatment with
white spruce foliage. However, larval instar did not influ-
ence the ingestion of larvae reared on Btk-treated balsam fir
foliage.
Mortality rate. The effect of Btk on mortality depended
on the type of host trees (F1,8¼ 16.25; P¼ 0.0038). Mortal-
ity rates among larvae reared on balsam fir or white spruce
foliage were 85 and 43% (� 15%, �2 SE), respectively.
Spruce budworm larval instar did not influence the effect
of Btk (F3,24¼ 1.87; P¼ 0.16).
Number of ingested needles per tested larva (IN15) (Btk-
treated larvae). The effect of host trees on the number of
needles ingested per larva tested (IN15) was influenced by
larval instar (F3,24¼ 6.77; P¼ 0.0018). Third-, fourth-, fifth-
and sixth-instar larvae reared on balsam fir ingested similar
numbers of needles as did third- and fourth-instar larvae fed
on white spruce foliage. However, fifth- and sixth-instar
larvae fed on white spruce foliage consumed significantly
greater number of needles (59% more) than younger larvae
fed white spruce foliage and larvae of all intars fed balsam
fir foliage (Fig. 5).
Nutritional and allelochemical compound contents. Host
tree and collection day did not influence nitrogen, phos-
phorous and magnesium contents of the foliage collected
(Table 8). Balsam fir foliage contained 40% more calcium
100
90
80
70
60
50
40
30
20
10
0Control 1×30BIU 2×30BIU
±2SE
Balsam firWhite spruce
(a)
Def
olia
tion
(%)
on P
upal
Sta
ge D
ay
55
50
45
40
35
30
25
20
15
10
0
5
Control 1×30BIU 2×30BIU
±2SE
(b)
Fol
iage
pro
duct
ion
(g/4
5-cm
bra
nch
tip)
Figure 3 (a) Defoliation (%) and (b) new foliage production (g/45-cm
branch tip) observed on pupal stage day depending on three Btk
application rates and the host trees.
100
90
80
70
60
50
40
30
20
10
0BF timing WS timing
Balsam firWhite spruce
±2SE
Def
olia
tion
(%)
on P
upal
Sta
ge D
ay
Figure 4 Defoliation (%) observed on pupal stage day according to
the Btk schedule based on bud flaring stage of either balsam fir
or white spruce and the host trees of either balsam fir or white
spruce.
64 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

than white spruce foliage, but calcium content in foliage did
not change over 5 days in either species (Table 8). Total
soluble sugars in either host trees were significantly lower
on 29 May than on 3 June, and there was a host tree effect;
white spruce foliage contained 29% more sugars than bal-
sam fir foliage (Table 8). Potassium levels increased 12%with time from 29May to 3 June (Table 8). There were more
phenols in white spruce foliage on 29 May than 3 June, but
the opposite was true for balsam fir foliage (Table 8). White
spruce foliage contained 19% more total tannins than
balsam fir foliage, and tannin contents were higher on 29
May than on 3 June in both host trees (Table 8). The ratio
of N to tannins, which is an index of the amounts of nitro-
gen not bound to tannins in foliage (Feeny, 1969), was 33%higher in balsam fir foliage than in white spruce foliage
(Table 8), and was higher on 29 May than on 3 June in
both host trees.
Relationships between mortality rate and ingested amounts
of nutrient and allelochemical compounds. Stepwise regres-
sions on mortality rate vs. ingested quantities of nutritional
and allelochemical compounds per larva tested produced
two significant equations for the fifth and sixth instars,
respectively:
Fifth instar: mortality rate (%) ¼ � 3.14 � (tannins
ingested/larva tested) þ 91.17; (r2 ¼ 70%); the standard
errors of the slope and intercept estimates were 9.6 and
0.72, respectively.
Sixth instar: mortality rate (%) ¼ � 11.39 � (tannins
ingested/larva tested) þ 550.18 � (K tested/ingested larva) þ85.47; (r2 ¼ 87%); tannins ingested/larva tested: partial
r2 ¼ 69%; K ingested/larva tested: partial r2 ¼ 18%9 the stand-ard errors of the slope (tannins ingested and K ingested/tested
larva) and intercept estimates were 3.3, 182 and 6.7, respectively.
These results indicated that mortality among fifth- and
sixth-instar larvae that had ingested Btk, decreased with the
amount of ingested tannins. In sixth-intar larvae, the
amounts of ingested K had positive effects on Btk mortality
rate.
Discussion
Experiments in the field and laboratory clearly showed that
Btk caused lower mortality when spruce budworms fed on
white spruce foliage compared to balsam fir foliage. Ten
days after field application of Foray 76B, larval mortality
was 2.8-fold higher on fir than on spruce trees. Moreover,
Btk applications on white spruce trees did not significantly
increase mortality among budworm population beyond
untreated controls. Under warm and constant temperature
(25 �C), larval mortality due to Foray 48B ingestion was
higher in the laboratory than in the field, but mortality
among larvae fed balsam fir foliage was also two-fold
higher than that among larvae fed white spruce foliage.
This response could not be linked to differences in the
quantity of Btk toxins deposited on foliage. When Btk
formulation was applied at the flaring of balsam fir buds,
white spruce buds were still closed, and Btk droplets prob-
ably did not reach the insect target. However, at the flaring
of white spruce, young shoots of both host trees were fully
opened to receive a uniform mist of Btk spray. Results of
the Btk toxin dosage experiment also indicated that Btk
deposits were similar on balsam fir and white spruce foliage.
Thus, other factors must have influenced the efficacy of Btk
on spruce budworm fed balsam fir compared to white
spruce foliage.
Larvae fed white spruce developed faster than larvae fed
balsam fir and were particularly advanced when Btk was
applied at white spruce Btk timing, which was consistent
with reports in the literature (Blais, 1957; Webb &McLeod,
1957; Lavallee & Hardy, 1988; Lysyk, 1989). However, the
laboratory experiment showed that differences in Btk
efficacy between the two host trees could not be attributed
to differences in larval development. Third, fourth, fifth and
sixth instars fed balsam fir foliage ingested similar amounts
of Btk-contaminated needles and there was no indication
that they differed in mortality (mean¼ 85%). Although
ingestion of Btk-contaminated needles increased with larval
instar (from 12 to 58 needles/larva for third–fourth and
fifth–sixth instars, respectively) on white spruce, larvae fed
white spruce foliage had similar mortality rate (43%)
regardless of larval instar. The increase in food consump-
tion as larvae grew resulted undoubtedly in ingestion of
higher Btk doses, which is consistent with the findings of
van Frankenhuyzen et al. (1997) for spruce budworm larvae
fed artificial diet, that ‘the 3.5-fold increase in lethal dose
requirement per larva from the fourth to the sixth instar is
more than compensated for by the 20-fold increase in
potential feeding rates’. In the present study, larvae fed
white spruce foliage ingested between two- and 12-fold
more Btk-contaminated needles than larvae fed balsam fir
foliage and less than 30% of the larvae died in the field. The
results indicate that larvae fed on white spruce had a high
tolerance to Btk and feeding inhibition may have set in just
before they acquired the lethal dose. By contrast, larvae fed
balsam fir foliage rapidly acquired the lethal dose, although
they ingested just a few needles.
At the time spruce budworm larvae were subjected to
Btk treatment, total tannin contents was higher in white
4.5
4.0
3.5
3.0
2.5
2.0
1.5
1.0
0.5
0.03rd Instar 4th Instar 5th Instar 6th Instar
Balsam firWhite spruce
±2SE
Ln (
Num
ber
of n
eedl
es in
gest
ed/la
rva
test
ed)
Figure 5 Effect of the host trees on the natural logarithm of the
number of needles ingested per larva according to the larval
instar.
Btk efficacy against spruce budworm in mixed coniferous stand 65
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

spruce foliage than in balsam fir foliage. Moreover, mor-
tality due to Btk ingestion was negatively correlated to the
amount of tannins ingested by a larva. This effect may be
related to the protein precipitating ability of tannins
(Zucker, 1983), which may have led to inactivation of the
Btk proteinaceous toxin. Appel & Schultz (1994) showed
that the efficacy of Thuricide, another commercial formu-
lation of Btk, towards gypsy moth also depended on host
trees; red oak foliage was more inhibitory to Btk toxins
than chestnut and quaking aspen, probably because of its
higher concentrations of total phenolics and tannins. Lord
& Undeen (1990) found that tannins released by decaying
vegetation in mosquito breeding sites tended to reduce
Bacillus thuringiensis ssp. israelensis efficacy. Luthy et al.
(1985) demonstrated that Btk protoxin could be inacti-
vated by increasing concentrations of hydrolysable gallo-
tannins and they also showed that Btk d-endotoxin was
strongly inactivated by extracts prepared from spruce nee-
dles. Formation of complexes between tannins and pro-
teins depends on the relative concentrations of tannins and
protein, and ‘a threshold level of tannin is required before
any protein precipitation occurs’ (Zucker, 1983). In the
present study, the ratio of N to tannins from white spruce
foliage was significantly lower than that from balsam fir
foliage, which indicates that more tannins were available
to form complexes with protein and Btk toxin in the mid-
gut of larvae fed white spruce than in that of larvae fed
balsam fir.
The midgut pH of lepidopteran larvae is alkaline and, in
many species, tends to reach pH10 (Berenbaum, 1980;
Gringorten et al., 1992), and this creates a favourable
environment for solubilization and digestion of Btk proto-
xin (Dent, 1993; van Frankenhuyzen, 1993). Larval midgut
pH tended to decrease when larvae fed on acidic food
(foliage or artificial diet) (Keating et al., 1990; Bauce et al.,
2002), and even slight reductions in pH to low alkaline pH
(above 8) could decrease the vulnerability of insects to
nuclear polyhedrosis virus (Stiles & Paschkles, 1980;
Keating et al., 1990) and suppress toxicity of the enzyme-
activated d-endotoxin of Btk (Gringorten et al., 1992).
Further investigations should verify if midgut pH of larvae
fed white spruce is lower than that of larvae fed balsam fir,
and whether this plays a major role in the low efficacy
of Btk formulations on white spruce trees. The protein-
precipitating ability of tannins is also favoured at pH
under 8 (Zucker, 1983) and lower alkaline pH in midgut
of larvae fed white spruce foliage could strengthen the
previous hypothesis of the potential role of white spruce
tannins in the inactivation of Btk toxins.
The positive correlation between the amount of potas-
sium ingested by the larva and mortality due to Btk inges-
tion indicated that potassium availability could enhance Btk
efficacy. Knowles (1994) highlighted the major role of
potassium in the destruction of midgut epithelial cells after
Bt-toxins had induced pores in the epithelium. Morris et al.
(1995) reported that a commercial Btk formulation
supplemented with 0.05% of potassium carbonates improved
Btk efficacy against Mamestra configurata Walker fed on
canola (Brassica napus L.or Brassica rapa L.).Table
8Chemicalc
ontents
(mean�2SE)in
balsam
firandwhitesprucefoliagecollectedon29Mayand3JuneFoliagewasusedto
testBtk
efficacyonsprucebudworm
larvaefedtwodifferent
hosttrees
Hosttree
Collectionday
Balsam
fir
Whitespruce
29May
3June
Hosttree�collectionday
Nutritionalcompounds
N(%
dry
weight)
F1,19¼3.52;p¼0.079
2.38�0.18a
2.15�0.18a
F1,19¼0.82;p¼0.38
2.21�0.18a
2.32�0.18a
F1,19¼0.25;p¼0.62
P(p.p.m
)F1,19¼1.36;P¼0.26
3725�202a
3558�202a
F1,19¼3.69;P¼0.073
3779�202a
3504�202a
F1,19¼2.25;P¼0.15
K(p.p.m
)F1,19¼1.18;P¼0.29
13626�596a
13168�596a
F1,19¼12.34;P¼0.0029
14138�596a
12656�596b
F1,19¼3.43;P¼0.083
Ca(p.p.m
)F1,19¼8.12;P¼0.012
1926�272a
1378�272b
F1,19¼1.20;P¼0.29
1757�272b
1547�272b
F1,19¼0.44;P¼0.52
Mg(p.p.m
)F1,19¼0.14;P¼0.71
1152�66a
1170�66a
F1,19¼2.07;P¼0.17
1194�66a
1127�66a
F1,19¼0.27;P¼0.61
Totalsoluble
sugars
(%dry
weight)
F1,19¼4.87;P¼0.042
5.91�1.1
b7.64�1.1
aF1,19¼5.3;P¼0.035
5.88�1.1
b7.68�1.1
aF1,19¼1.66;P¼0.22
Allelochemicalcompounds
Totalphenols
(%)
F1,19¼0.81;P¼0.38
11.74�1.46a
10.81�1.46a
F1,19¼0.44;P¼0.52
11.62�1.46a
10.94�1.46a
F1,19¼5.62;P¼0.0306
Totaltannins(cm
2/m
gfoliagedry
weight)
F1,19¼14.36;P¼0.0016
0.642�0.046b
0.763�0.046a
F1,19¼19.6;P¼0.0004
0.773�0.046a
0.632�0.046b
F1,19¼1.74;P¼0.21
N/totaltannins
F1,19¼12.21;P¼0.003
3.81�0.38a
2.87�0.38b
F1,19¼9.93;P¼0.0062
2.91�0.38b
3.77�0.38a
F1,19¼0.1;P¼0.75
Means�
2SEfollowedbythesameletterwithin
thesamerow
perfactordonotdiffersignificantlyaccordingto
theBonferronipairwisecomparisonmethod.
66 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

In control plots, defoliation was higher on balsam fir
than on white spruce trees at the end of larval development.
Insect grazing pressure was undoubtedly higher on balsam
fir than on white spruce because larval mortality during the
total larval development was lower when larvae fed on
untreated balsam fir foliage than on untreated white spruce
foliage. An increase in raw fibre foliar content (i.e. lignin,
silica, pectin, cellulose and hemicellulose) may be the cause
of this result because it negatively affects larval survival of
spruce budworm (Bauce & Hardy, 1988) as well as larch
budmoth (Zeiraphera diniana (Gn.) (Benz, 1974). The rapid
cessation of white spruce shoot elongation and the start of
shoot lignification have been reported to rapidly make
white spruce foliage less suitable for spruce budworm
nutrition (Thomas, 1987; Lawrence et al., 1997) and these
changes in foliage quality lead to decrease drastically larval
survival. By contrast, balsam fir foliage appears to remain
suitable for a longer period of time than white spruce foliage
because of its slower rate of shoot growth (Greenbank,
1963).
Larvae that ingested sublethal Btk doses exhibit longer
larval development (van Frankenhuyzen et al., 1997; Bauce
et al., 2002). In the Btk-treated plots, the more important
the increase in larval development, the more important are
the risks to encountering unsuitable foliage, especially for
larvae fed on white spruce trees, because of the shoot lig-
nification. Feeding on unsuitable white spruce foliage
tended to decrease larval survival, which led to a reduction
in defoliation of white spruce trees. At the end of the grow-
ing season, defoliation was similar in Btk-treated balsam fir
and white spruce trees, although Btk-related larval mortal-
ity on balsam fir trees was nearly three-fold higher than that
on white spruce trees. White spruce foliage protection was
probably achieved by the combination of a shorter window
of foliage suitability and Btk effects, lethal and sublethal
(feeding inhibition and increase in larval development time).
The present field experiment showed that balsam fir
foliage production was similar in control and Btk-treated
plots. Insect grazing probably compromised shoot elonga-
tion prior to Btk applications, or was able to overtake
balsam fir production (Koller & Leonard, 1981), even
when budworm populations were low and reduced by Btk
treatment (under 16 larvae per 45-cm branch tip). White
spruce shoots can grow longer than balsam fir shoots and
thus, provide more food per unit of larval population
(Greenbank, 1963; Koller & Leonard, 1981; Regniere
et al., 1989). However, this phenomenon was not observed
at high larval densities (42 larvae per 45-cm branch tip);
control balsam fir and white spruce trees showed similar
foliage production, probably because insect grazing by high
larval population densities had outpaced expansion of both
balsam fir and white spruce shoots. On Btk-treated white
spruce, most of the spruce budworm larvae ceased to feed
and defoliation did not increase during the 10 days follow-
ing Btk application (unpublished data). Thus, shoot elonga-
tion was not impeded and foliar production was 60% higher
in white spruce than in balsam fir in Btk-treated plots.
More foliage remained on white spruce than on balsam
fir trees. In control plots, white spruce trees underwent
lower defoliation, because of an increase in budworm mor-
tality at the end of the larval development. In Btk-treated
plots, white spruce and balsam fir trees exhibited similar
levels of defoliation, but 50% more foliage remained on
spruce compared to fir due to the higher foliage production
of the former.
During the last budworm epidemic (1967–92) in the Pro-
vince of Quebec, the main objective of forest protection
against spruce budworm was tree survival. When aerial
spray programmes were planned, one of the decision criteria
was the death risk of host trees. White spruce trees were less
vulnerable to spruce budworm than balsam fir trees, with
spruce dying after 5–7 years of severe successive defoliation
compared to 4–5 years for balsam fir trees (Hardy, 1979;
Gagnon & Chabot, 1991). However, in the present managed
forests, it should be more important to protect stand yield
and more efficient strategies have to be found. The impact
of standard Btk sprays (one or two applications of 30BIU/
ha) on spruce budworm fed on white spruce appeared to
rely more on sublethal effects than on mortality. If the host
tree reduces pest vulnerability to Btk, applications of higher
Btk concentrated formulations than those currently used
could increase the chance that larvae ingest a lethal dose
more rapidly (Appel & Schultz, 1994).
Differences in bud phenology among host trees compli-
cated the scheduling of Btk applications in mixed-coniferous
stands, and the inter-relationship between spruce budworm,
host tree, and Btk at bud flaring needed to be understood.
The present field experiment clearly demonstrated that pro-
tection of mixed balsam fir-white spruce stands, required that
Btk treatment had to be applied as early as possible (i.e. on
the flaring of balsam fir buds). When Btk treatments were
scheduled at the later white spruce flaring, defoliation had
already reached 21% for both tree species, and tree growth
begins to be negatively affected at this level of defoliation
(MacLean, 1985). Furthermore, defoliation of white spruce
at the end of the season was similar at the two Btk timings,
whereas balsam fir defoliation at the end of the season was
higher when Btk was applied at the white spruce flaring
instead of at the balsam fir flaring. The main advantage of
scheduling Btk application at balsam fir bud expansion was
to provide the best protection for this species, and finally the
mixed balsam fir-white spruce stands. These results were in
accordance with those of Cadogan & Scharbach (1993) who
showed that balsam fir defoliation was significantly higher
when stands were sprayed at the flaring of black spruce
(28%) instead of the flaring of balsam fir (18%). They also
found that Btk application timing did not influence the black
spruce final defoliation.
This experiment demonstrates the importance of acquir-
ing a fundamental knowledge of the relationships between
Btk, spruce budworm and host trees (chemical foliage
profile as well as bud phenology) for developing the best
forest protection strategy according to ecological and
economic concerns of the day. Further research of this
kind should be pursued to determine the appropriate and
efficient use of the Btk insecticide in forest protection for this
specific complex (Btk and spruce budworm) in other geograph-
ical locations and for other tree species and defoliators.
Btk efficacy against spruce budworm in mixed coniferous stand 67
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

Acknowledgements
The authors would like to thank the laboratory and field
team of the forestry department at SOPFIM and at the
Laval University for their helpful technical assistance.
Thanks are addressed to Dan Quiring (Population Ecology
Group, Faculty of Forestry and Environmental Manage-
ment, University of New Brunswick, Fredericton, Canada)
for his critical review of this manuscript. Research funding
was provided by the National Sciences and Engineering
Council of Canada and the Societe de Protection des Forets
contre les Insectes et Maladies.
References
Appel, H.M. & Schultz, J.C. (1994) Oak tannins reduce effective-
ness of Thuricide (Bacillus thuringiensis) in the gypsy moth
(Lepidoptera: Tortricidae). Journal of Economic Entomology, 87,
1736–1742.
Bauce, E. (1996) One and two years impact of commercial thinning
on spruce budworm feeding ecology and host tree foliage
production and chemistry. Forestry Chronicle, 72, 1–6.
Bauce, E. & Hardy, Y. (1988) Effects of drainage and severe
defoliation on the rawfiber content of balsam fir needles and
growth of the spruce budworm (Lepidoptera: Tortricidae).
Environmental Entomology, 17, 671–674.
Bauce, E., Crepin, M. & Carisey, N. (1994) Spruce budworm
growth, development and food utilization on young and old
balsam fir trees. Oecologia, 97, 499–507.
Bauce, E., Bidon, Y. & Berthiaume, R. (2002) Effects of food
nutritive quality and Bacillus thuringiensis on feeding behaviour,
food utilization and larval growth of spruce budworm
Choristoneura fumiferana (Clem.) when exposed as fourth and
sixth-instar larvae. Agricultural and Forest Entomology, 4, 1–14.
Beegle, C.C. & Yamamoto, T. (1992) Invitation paper (C.P.
Alexander Fund): history of Bacillus thuringiensis Berliner
research and development. Canadian Entomologist, 124, 587–616.
Benz, G. (1974) Negative feedback by competition for food and
space and by cyclic induced changes in the nutritional base
as regulatory principles in the population dynamics of the
larch bud moth, Zeiraphera diniana (Gn.) (Lep., Tortricidae).
Zeitschrift fuer Angewandte Entomologie, 76, 196–228.
Berenbaum, M. (1980) Adaptive significance of midgut pH in
larval Lepidoptera. American Naturalist, 115, 138–146.
Blais, J.R. (1957) Some relationships of the spruce budworm,
Choristoneura fumiferana (Clem.) to black spruce, Picea mariana
(Moench) Voss. Forestry Chronicle, 13, 364–372.
Blais, J.R. (1976) Vulnerabilite de l’epinette blanche a la defoliation
par la tordeuse des bourgeons de l’epinette. Revue Bimestrielle de
Recherches, 32, 21.
Blais, J.R. (1983) Trends in the frequency, extent, and severity of
spruce budworm outbreaks in eastern Canada. Canadian Journal
of Forest Research, 13, 539–617.
Boulet, B. (2001) Retrospective-Les enseignements de la derniere
epidemie de tordeuses des bourgeons de l’epinette. Tordeuse des
Bourgeons de l’epinette: l’Apprivoiser dans nos Strategies
d’Amenagement pp. 3–13, Fo18–48/2001F, 27–29 March. Service
Canadien Des Forets, Shawinigan, Quebec, Canada.
Cadogan, B.L. & Scharbach, R.D. (1993) Efficacy of Foray 48B
(Bacillus thuringiensis Berliner) applications against the spruce
budworm, Choristoneura fumiferana (Clemens) (Lepidoptera:
Tortricidae), timed for phenological development of balsam fir
and black spruce. Canadian Entomologist, 125, 479–488.
Dement, W.A. & Mooney, H.A. (1974) Seasonal variation in the
production of tannins and cyanogenic glucosides in the
chaparral shrubs, Heteromeles arbutifolia. Oecologia, 15, 65–76.
Dent, D.R. (1993) The use of Bacillus thuringiensis as an
insecticide. Exploitation of Microorganisms (ed. by D. G. Jones),
pp. 19–43. Chapman & Hall, London.
Dimond, J.B. (1985) Considerations in timing insecticidal sprays
on different host trees of the spruce budworm. Recent advances
in spruce budworms research. CANUSA Spruce Budworms
Research Symposium, pp. 409. Bangor, Maine, 16–20 September
1984. Canadian Forest Service, Ottawa.
Dorais, L.G. & Hardy, Y.J. (1976) Methode d’evaluation de la
protection accordee au sapin baumier par les pulverisations
aeriennes contre la tordeuse des bourgeons de l’epinette.
Canadian Journal of Forest Research, 6, 86–92.
Dorais, L.G. & Kettela, E. (1982) Revue, par region, des techniques
d’inventaire entomologique et d’evaluation des programmes de
pulverisation a grande echelle contre la tordeuse des bourgeons de
l’epinette Choristoneura fumiferana (Clem.). Conseil de l’Est de
la tordeuse des bourgeons de l’epinette. Rapport du comite pour
la standardisation des techniques entomologiques, p. 51.
Ministere de l’Energie et des Resources du Quebec, Quebec,
Canada.
Farrar, R.R.J., Phyllis, A.W.M. & Ridgway, R.L. (1996) Host
plant effects on activity of Bacillus thuringiensis against gypsy
moth (Lepidoptera: Lymantriidae) larvae. Environmental
Entomology, 25, 1215–1223.
Feeny, P.P. (1969) Inhibitory effect of oak tannins on the
hydrolysis of proteins by trypsin. Phytochemistry, 8, 2119–2126.
Fettes, J.J. (1950) Investigations of sampling techniques for
population studies of the spruce budworm on balsam fir in
Ontario. Forest Insect Laboratory, Sault Ste Marie, Annual
Technical Report, 4, 163–401.
Gagnon, R.R. & Chabot, M. (1991) Prevention des pertes de bois
attribuables a la tordeuse des bourgeons de l’epinette, p. 52.
Gouvernement du Quebec, Ministere des forets. Service d’ame-
nagement forestier et service de protection contre les insectes et
les maladies, Quebec.
Greenbank, D.O. (1963) Host trees and the spruce budworm.
Memoirs of the Entomological Society of Canada, 31, 219–223.
Gringorten, J.L., Milne, R.E., Fast, P.G., Sohi, S.S. & Van
Frankenhuyzen, K. (1992) Suppression of Bacillus thuringiensis
d-endotoxin activity by low alkaline pH. Journal of Invertebrate
Pathology, 60, 47–52.
Hagerman, A.E. (1987) Radial diffusion method for determining
tannin in plant extracts. Journal of Chemical Ecology, 13, 437–449.
Hardy, Y.J. (1979) Les recherches sur la tordeuse des bourgeons de
l’epinette. Revue Forestiere Francaise, 31, 217–222.
Juneau, A. (1989) A Review of Aerial Spraying Technology for
Spruce Budworm Control in Private Woodlots in Eastern Quebec,
p. 219. Canadian Forest Service, Ste Foy, Quebec.
Keating, S.T., Schultz, J.C. & Yendol, W.G. (1990) The effect of
diet on gypsy moth (Lymantria dispar) larval midgut pH, and its
relationship with larval susceptibility to a baculovirus. Journal of
Invertebrate Pathology, 56, 317–326.
Knowles, B.H. (1994) Mechanism of action of Bacillus thuringiensis
insecticidal d-endotoxins. Advances in Insect Physiology, 24,
275–308.
Koller, C.N. & Leonard, D.E. (1981) Comparison of energy
budgets for spruce budworm Choristoneura fumiferana
(Clemens) on balsam fir and white spruce. Oecologia, 49, 14–20.
Kouassi, K.C., Lorenzetti, F., Guertin, C., Cabana, J. &Mauffette, Y.
(2001) Variation in the susceptibility of the forest tent caterpillar
(Lepidoptera: Lasiocampidae) to Bacillus thuringiensis variety
68 N. Carisey et al.
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

kurstaki HD-1: effect of the host plant. Journal of Economic
Entomology, 94, 1135–1141.
Lavallee, R. & Hardy, Y. (1988) Etude en laboratoire du developpe-
ment du Choristoneura fumiferana sur l’Abies balsamea, le Picea
glauca et le Picea rubens. Phytoprotection, 69, 79–86.
Lawrence, R.K., Mattson, W.J. & Haack, R.A. (1997) White
spruce and the spruce budworm: defining the phenological
window of susceptibility. Canadian Entomologist, 129, 291–318.
Lord, J.C. & Undeen, A.H. (1990) Inhibition of the Bacillus
thuringiensis var. israelensis toxin by dissolved tannins. Environ-
mental Entomology, 19, 1547–1551.
Ludlum, C.T., Felton, G.W. & Duffey, S.S. (1991) Plant defenses:
Chlorogenic acid and polyphenol oxidase enhance toxicity of
Bacillus thuringiensis subsp. kurstaki to Heliothis zea. Journal of
Chemical Ecology, 17, 217–237.
Luthy, P., Hofmann, C. & Jaquet, F. (1985) Inactivation of delta-
endotoxin of Bacillus thuringiensis by tannin. FEMS Microbiol-
ogy Letters, 28, 31–33.
Lysyk, T.J. (1989) Stochastic model of eastern spruce budworm
(Lepidoptera: Tortricidae) phenology in white spruce and
balsam fir. Journal of Economic Entomology, 82, 1161–1168.
MacLean, D.A. (1985) Effects of spruce budworm outbreaks on
forest growth and yield. CANUSA Spruce Budworms Research
Symposium, pp. 148–175. Bangor, Maine, 16–20 September
1984. Canadian Forest Service, Ottawa.
MacLean, D.A. & MacKinnon, W.E. (1997) Effects of stand and
site characteristics on susceptibility and vulnerability of balsam
fir and spruce to spruce budworm in New Brunswick. Canadian
Journal of Forest Research, 27, 1859–1871.
McGugan, B.M. (1954) Needle-mining habits and larval instars of
the spruce budworm. Canadian Entomologist, 86, 439–454.
Moldenke, A.F., Berry, R.E., Miller, J.C., Wernz, J.G. & Li, X.H.
(1994) Toxicity of Bacillus thuringiensis subsp. kurstaki to gypsy
moth, Lymantria dispar, fed with alder or douglas fir. Journal of
Invertebrate Pathology, 64, 143–145.
Morris, O.N., Converse, V. & Kanagaratnam, P. (1995) Chemical
additive effects on the efficacy of Bacillus thuringiensis Berliner
subsp. kurstaki against Mamestra configurata (Lepidoptera:
Noctuidae). Journal of Economic Entomology, 88, 815–824.
Navon, A., Hare, J.D. & Federici, B.A. (1993) Interactions among
Heliothis virescens larvae, cotton condensed tannin and the
CryIA (c) d-endotoxin of Bacillus thuringiensis. Journal of
Chemical Ecology, 19, 2485–2499.
Regniere, J. & You, M. (1991) A simulation model of spruce
budworm (Lepidoptera: Tortricidae) feeding on balsam fir and
white spruce. Ecological Modelling, 54, 277–297.
Regniere, J., Lysyk, T.J. & Auger, M. (1989) Population density
estimation of spruce budworm, Choristoneura fumiferana
(Clem.) (Lepidoptera: Tortricidae) on balsam fir and white
spruce from 45-cm mid-crown branch tips. Canadian Entomol-
ogist, 121, 267–281.
Royama, T. (1984) Population dynamics of the spruce budworm
Choristoneura fumiferana. Ecological Monographs, 54, 429–462.
Sanders, C.J. (1980) A Summary of Current Techniques Used for
Sampling Spruce Budworm Populations and Estimating Defoli-
ation in Eastern Canada, p. 33. Environment Canada, Canadian
Forest Service Information Report: O-X-306, Great Lakes
Forest Research Center, Sault Ste Marie, Ontario.
Sanders, C.J. (1991) Biology of North American spruce budworms.
Tortricid Pests, Their Biology, Natural Enemies and Control (ed.
by L. P. S. van der Geest and H. H. Evenhuis), pp. 579–620.
Elsevier Science Publishers, Amsterdam, The Netherlands.
SAS Institute Inc (1988) SAS/STAT User’s Guide, version 6.03.
SAS Institute, Inc., Cary, NC.
Sivamani, E., Rajendran, N., Senrayan, R., Ananthakrishnan, T.N.,
Jayaraman, K. (1992) Influence of some plant phenolics on the
activity of d-endotoxin of Bacillus thuringiensis var. galleriae on
Heliothis armigera. Entomologia Experimentalis et Applicata, 63,
243–248.
Stiles, B. & Paschkles, J.D. (1980) Midgut pH in different instars of
three Aedes mosquito species and the relationship between pH
and susceptibility of larvae to a nuclear polyhedrosis virus.
Journal of Invertebrate Pathology, 35, 58–64.
Thomas, A.W. (1987) The effect of age of current-year shoots of
Picea glauca on survival, development time, and feeding
efficiency of 6th-intar larvae of Choristoneura fumiferana.
Entomologia Experimentalis et Applicata, 43, 251–260.
van Frankenhuyzen, K. (1993) The challenge of Bacillus
thuringiensis. Bacillus Thuringiensis, an Environmental Biopesti-
cide: Theory and Practice (ed. by P. F. Entwistle, J. S. Cory,
M. J. Bailey and S. Higgs), pp. 1–35. John Wiley & Sons, New
York.
van Frankenhuyzen, K., Nystrom, C.W. & Tabashnik, B.E. (1995)
Variation in tolerance to Bacillus thuringiensis among and within
populations of the spruce budworm (Lepidoptera: Tortricidae)
in Ontario. Journal of Economic Entomology, 88, 97–105.
van Frankenhuyzen, K., Gringorten, L., Dedes, J. & Gauthier, D.
(1997) Susceptibility of different instars of the spruce budworm
(Lepidoptera: Tortricidae) to Bacillus thuringiensis var kurstaki
estimated with a droplet-feeding method. Journal of Economic
Entomology, 90, 560–565.
van Frankenhuyzen, K., Nystrom, C. & Dedes, J. (1998) The Use
of the Abbott Deposit Assessment Method and a Foliar Bioassay
to Evaluate Spray Deposition and Treatment Effects Resulting
From Experimental Applications of Foray 48B in the 1997
Gatineau Program. Canadian Forest Service, Great Lakes
Forestry Centre, Sault Ste Marie, Ontario, Canada.
Volney, W.J.A. & Cerezke, H.F. (1992) The phenology of white
spruce and the spruce budworm in northern Alberta. Canadian
Journal of Forest Research, 22, 198–205.
Webb, F.E. & McLeod, J.M. (1957) Studies of Aerial Spraying
Against the Spruce Budworm in New Brunswick IX. Comparative
Studies of the Spruce Budworm on Balsam Fir and Spruce in New
Brunswick, p. 28. Forest Biology Laboratory, Fredericton, New
Brunswick. Department of Agriculture, Science Service, Canada.
Wisdom, C.S., Gonzales-Coloma, A. & Rundel, P.W. (1987)
Ecological tannin assays. Evaluation of proanthocyanidins,
protein binding assays and protein precipitating potential.
Oecologia, 72, 395–401.
Zucker, W.V. (1983) Tannins: Does structure determine function?
An ecological perspective. American Naturalist, 121, 335–365.
Accepted 4 March 2003
Btk efficacy against spruce budworm in mixed coniferous stand 69
# 2004 The Royal Entomological Society, Agricultural and Forest Entomology, 6, 55–69

Related Documents








![Choristoneura fumiferana (Clem.)cfs.nrcan.gc.ca/pubwarehouse/pdfs/9561.pdfCHORISTONEURA FUMIFERANA {CLEM.) INTRODUCTION The spruce budworm {Chonstoiieurafumifsrana [Clem.]) is a major](https://static.cupdf.com/doc/110x72/5f0b027b7e708231d42e6847/choristoneura-fumiferana-clemcfsnrcangccapubwarehousepdfs9561pdf-choristoneura.jpg)


