ELSEVIER Life Sciences, Vol. 61, No. 2, pp. lOSllS,l!I97 CoWright 0 1997 Eck3evier sciellce Inc. Printed in the USA. All rights ltservcd oo24al5/97 $17.00 t .oa PI1 soo24-3205(97)00365- EFFECTS OF BIOCHANIN A ON THE GROWTH AND DIFFERENTIATION OF MYELOID LEUKEMIA WEHI3B (JCS) CELLS M.C. Fung’, Y .Y. Szeto’, K.N. Leung2 , Y.L. Wong-Leung3, and N.K. Mk4 ‘Department of Biology, 2Department of Biochemistry, The Chinese University of Hong Kong; 3Department of Chemistry,4Department of Biology, Hong Kong Baptist University, Hong Kong. (Received in final form April 2,1997) Summary The effects of biochanin A on the growth and differentiation of a recently characterized myeloid leukemia cell line WEHI3B (JCS) were investigated. Biochanin A not only inhibited the growth of JCS cells in a dose-dependent manner (0 - 200 pM) but also induced the morphological differentiation of JCS cells. The phagocytic activity of biochanin A-treated JCS cells was also increased. Flow cytometric analysis showed that the expression of macrophage differentiation markers Mac-l and F4/80 was up-regulated in biochanin A-treated JCS cells. The expression level of Mac-l was higher than that of F4/80. The expression of cytokine genes was studied by reverse transcription-polymerase chain reaction (RT-PCR) and cycle titration. mRNA levels of IL-la, IL-lb and IL-4 were found to be up-regulated at 46 hours after incubation of JCS cells with biochanin A. Although the expression of LIF was also up-regulated, the LIF receptor gene was not expressed in the uninduced or induced JCS cells. Our results suggest that IL-la, IL-1 p and IL-4 may act on the later stage of biochanin A-mediated differentiation of JCS cells. I&y word: anti-leukemia, biochanin A, myeloid leukemia Isoflavonoids are a large distinctive subclass of flavonoids (1). The naturally occurring isoflavonoids are mostly restricted to plant species. Isoflavones constitute the largest group of natural isoflavonoids. Certain isoflavones, such as biochanin A and genistein, have been shown to mediate a wide range of biological activities, such as inhibition of the in vitro growth of stomach, breast and prostate tumour cells (2-4), reduction of in vitro mutagenic activities of chemical mutagens (5-6), and suppression of in vivo tumour development and tumour promotion in mice (7-8). Corresponding author: Dr. K.N. Leung, Department of Biochemistry, The Chinese University of Hong Kong, Shatin, N.T., Hong Kong. Tel: (852)-26096802, Fax: (852)-26035123

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER
Life Sciences, Vol. 61, No. 2, pp. lOSllS,l!I97 CoWright 0 1997 Eck3evier sciellce Inc.
Printed in the USA. All rights ltservcd oo24al5/97 $17.00 t .oa
PI1 soo24-3205(97)00365-
EFFECTS OF BIOCHANIN A ON THE GROWTH AND DIFFERENTIATION OF MYELOID LEUKEMIA WEHI3B (JCS) CELLS
M.C. Fung’, Y .Y. Szeto’, K.N. Leung2 , Y.L. Wong-Leung3, and N.K. Mk4
‘Department of Biology, 2Department of Biochemistry, The Chinese University of Hong Kong;
3Department of Chemistry,4Department of Biology, Hong Kong Baptist University, Hong Kong.
(Received in final form April 2,1997)
Summary
The effects of biochanin A on the growth and differentiation of a recently characterized myeloid leukemia cell line WEHI3B (JCS) were investigated. Biochanin A not only inhibited the growth of JCS cells in a dose-dependent manner (0 - 200 pM) but also induced the morphological differentiation of JCS cells. The phagocytic activity of biochanin A-treated JCS cells was also increased. Flow cytometric analysis showed that the expression of macrophage differentiation markers Mac-l and F4/80 was up-regulated in biochanin A-treated JCS cells. The expression level of Mac-l was higher than that of F4/80. The expression of cytokine genes was studied by reverse transcription-polymerase chain reaction (RT-PCR) and cycle titration. mRNA levels of IL-la, IL-lb and IL-4 were found to be up-regulated at 46 hours after incubation of JCS cells with biochanin A. Although the expression of LIF was also up-regulated, the LIF receptor gene was not expressed in the uninduced or induced JCS cells. Our results suggest that IL-la, IL-1 p and IL-4 may act on the later stage of biochanin A-mediated differentiation of JCS cells.
I&y word: anti-leukemia, biochanin A, myeloid leukemia
Isoflavonoids are a large distinctive subclass of flavonoids (1). The naturally occurring isoflavonoids are mostly restricted to plant species. Isoflavones constitute the largest group of natural isoflavonoids. Certain isoflavones, such as biochanin A and genistein, have been shown to mediate a wide range of biological activities, such as inhibition of the in vitro growth of stomach, breast and prostate tumour cells (2-4), reduction of in vitro mutagenic activities of chemical mutagens (5-6), and suppression of in vivo tumour development and tumour promotion in mice (7-8).
Corresponding author: Dr. K.N. Leung, Department of Biochemistry, The Chinese University of Hong Kong, Shatin, N.T., Hong Kong. Tel: (852)-26096802, Fax: (852)-26035123

106 Biochauiu A and Leukemic Cell Differentiation Vol. 61, No. 2, 1997
We have been investigating the growth and differentiation of a murine myeloid WEHI-3B (JCS) leukemia cell line, and have shown that monocytic differentiation of this cell line can be induced by tumour necrosis factor-a (TNF-a) (9). The differentiation-inducing effect of TNF-a could be enhanced by cytokines such as interleukin-4 (IL-4) , interferon-y, or microbial products such as pertussis toxin (9-l 1). Although biochanin A has been known to have anti-mutagenic activity against carcinogens, the differentiation-inducing effect of biochanin A on leukemia cells has not yet been reported. In this investigation, the effects of biochanin A on the growth and differentiation of the JCS leukemia cells were studied. The expression of cytokine IL- 1 a, IL-l 0, IL-4, TNF-a, and leukemia inhibitory factor (LIF) genes in biochanin A-treated JCS cells was
also examined.
Materials and Methods
Cell line: The. murine myeloid leukemia cell line WEHI-3B (JCS) was maintained in RPM1 1640 medium supplemented with 10% fetal calf serum (FCS, Gibco), 2 mM glutamine, and 50 U/ml penicillin, 50 pg/ml streptomycin, and 10 pg/ml neomycin. The cultures were incubated in a humidified CO2 incubator (37°C 5% CO*).
Proliferation assay: Thymidine incorporation assay was used to measure the proliferation of
leukemia cells. 4~10~ cells were cultured with 200 ~1 of serially diluted biochanin A (5,7-
dihydroxy4’-methoxyisoflavone, Sigma) in 96-well flat-bottom microplates. The cultures were incubated at 37°C in a humidified CO, incubator for 48 hours. Biochanin A was freshly prepared by dissolving in absolute ethanol. Appropriately diluted ethanol was used as a solvent control for biochanin A. The cultures were then pulsed with 0.5 pCi of 3H-methyl-thymidine (3H-TdR, specific activity = 5 Ci/mmol, Amersham) in the last 6 hours of incubation. The cells were harvested and the incorporation of 3H-TdR was measured using liquid scintillation counter (Beckman liquid scintillation counter LS1801).
Flow cytometry: The expression of Mac-l, F4/80 and Gr- 1 cell surface antigens on uninduced or differentiated JCS cells was analyzed by flow cytometry (10). JCS cells were initially blocked with 10 pg/ml isotype-specific antibody at 4Y! for 30 minutes. The cells were then washed and incubated at 4°C for 30 minutes with anti-Mac-l (ATCC TIB 128), anti-F4/80 (ATCC HB 198) or anti-Gr-1 monoclonal antibodies (Pharmingen, USA). The cells were further stained with appropriately diluted fluorescein isothiocyanate-conjugated sheep anti-rat Ig antibodies (Southern Biotechnology Associates Inc.) at 4°C for 30 minutes followed by fixation with 1% (w/v) paraformaldehyde in normal saline (12). Between each step, the cells were washed with phosphate-buffered saline (PBS) containing 2% heat-inactivated FCS and 0.02 M sodium azide. At least 1x10’ cells for each sample were analyzed by a fluorescence activated cell sorter (Becton Dickinson FACSort flow cytometer).
Cell morphology: Cytospin preparations from control and biochanin A-treated cultures were stained with Hemacolor (Merck). The numbers of cells with different degrees of differentiation were determined by their morphological characteristics (9). Six hundred cells were counted for each slide and the results were presented as mean * S.D.

Vol. 61, No. 2, 1997 Biochanio A and Leukemic Cell Differentiation 107
Phagocytosis assay: Phagocytic activity was determined by the modified yeast phagocytosis
assay as described previously (9). Briefly, opsonized yeasts were prepared by incubating glutaraldehyde-inactivated yeast cells (Saccharomyces cerevisiae) with fresh mouse serum at 37°C for 30 minutes. 2~10~ opsonized yeasts were added to untreated or biochanin A-treated JCS cell cultures in a 96-well microplate. The mixtures were incubated at 37°C for 16 hours. The cells were harvested and the percentages of phagocytic cells were counted. A total of 500 cells were counted in each sample and the results were expressed as mean f S.D.
Reverse transcription-polymerase chain reaction (RT-PCR): Guanidinium thiocyanate I cesium chloride centrifitgation method was used in the preparation of total RNA (13). RT-PCR was used to determine the expression of cytokine genes (14). Briefly, 1 ug of total RNA isolated from each sample was heated to 65°C for 10 minutes and reverse-transcribed at 37°C for 1 hour by adding 10 ul of master mix containing 40 U RNase inhibitor, 0.1 ug oligo dT,,,,, 0.5 mM dNTP mixture, 50 mM Tris-HCl pH 8.3, 75 mM KCl, 3 mM MgC12, 10 mM DTT and 200 U
Moloney murine leukemia virus reverse transcriptase. The RT samples were diluted in DEPC- treated water (equivalent to 0.1 ug/ul of total RNA), boiled for 10 minutes and quick-chilled on ice. 10 ul of the diluted RT sample was amplified in a 50 ul PCR reaction mixture containing
1.5 mM MgC12, 20 mM (NH,),SO,, 75 mM Tris-HCl (pH 9.0) 0.01% Tween-20, 0.2 mM of
each dNTP, 1 pmol/ul of each primer and 0.25 U Thermoprime plus DNA polymerase (Thermoprime plus 207G ). PCR involved denaturation at 94°C for 1 minute, annealing at 10°C below the melting temperature of the primers for 1 minute, and extension at 72°C for 1 minute. The sequences of primers are shown in Table I. GAPDH mRNA was used as the standard for normalization of RNA samples.
Southern blot and dot blot hybridization: The specificity of RT-PCR products was confirmed by Southern blot analysis. PCR products were loaded in a 2% TAE agarose gel and transferred in 0.4 M NaOH onto positively-charged nylon membranes (Boehringer Mannheim). The membranes were neutralized in 2X SSC and baked at 120°C for 30 minutes. Dot blot hybridization was used to determine the relative amount of PCR products obtained from cycle titration. 10 ul of PCR products from each sample were denatured in 0.2 M NaOH in a total volume of 200 yl for 15 minutes at room temperature. The denatured products were applied to a
pre-wetted membrane assembled in the dot-blot apparatus (BioRad) and 400 ul of 20X SSC was added to each well for neutralization. The membrane was removed, air-dried and baked at 120°C for 30 minutes. The membrane was then pre-hybridized for 2 hours followed by hybridization with the appropriate internal probe (Table I) labelled with digoxigenin-dUTP using the DIG oligonucleotide 3’end labelling kit ( Boehringer Mannheim # 362372). Hybridization was carried out at 10°C below the melting temperature of the probe for 12 hours. The membrane was washed twice with 0.5X SSC, 0.1% SDS for 15 minutes. CSPD (Disodium 3 - (4- methoxyspiro (1,2 - dioxetane - 3,2’ - (5’-chloro) tricycle (3.3.1.1) decan) - 4~1) phenyl phosphate was used as a substrate in the detection of hybridization products (DIG Luminescent
Detection Kit, Boehringer Mannheim #13635 14). The membrane was exposed to X-ray film for 5 to 10 minutes. The signals developed on the X-ray film were analyzed using the Molecular AnalystW System (BioRad).
108 Biochanin A and Leukemic CeU Differentiation Vol. 61, No. 2, 1997
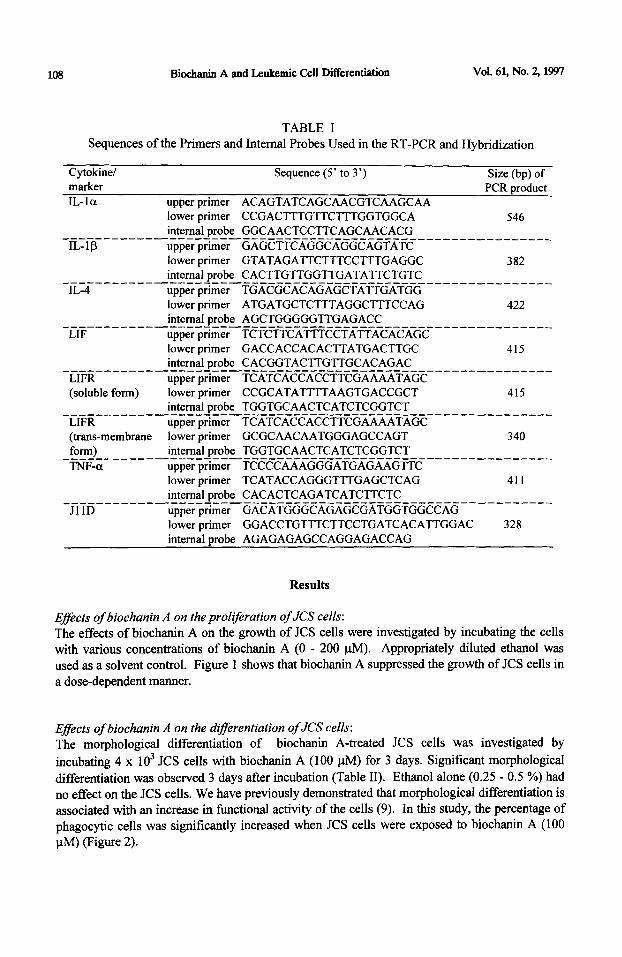
TABLE I Sequences of the Primers and Internal Probes Used in the RT-PCR and Hybridization
Cytokinel Sequence (5’ to 3’) Size (bp) of marker
IL-la upper primer ACAGTATCAGCAACGTCAAGCAA lower primer CCGACTTTGTTCTTTGGTGGCA
PCR product
546 internal probe GGCAACTCCTTCAGCAACACG _------_-_-------~-----_-----------------_-_-----_____-___
IL-IS upper primer GAGCTTCAGGCAGGCAGTATC lower primer GTATAGATTCTTTCCTTTGAGGC 382 internal probe CACTTGTTGGTTGATATTCTGTC _-_-______-_-_---~-----------------_-------_--_-__-_-_-___
IL-4 upper primer TGACGCACAGAGCTATTGATGG lower primer ATGATGCTCT’M’AGGCTTTCCAG 422 internal probe AGCTGGGGGTTGAGACC ___-______-_-_--7-----------------------~-~_-_-~--_----__
LIF upper prrmer TCTCTTCATTTCCTATTACACAGC lower primer GACCACCACACTTATGACTTGC 415 internal probe CACGGTACTTGTTGCACAGAC _---------------~-------------------------_--_-~_-_-----_
LIFR upper prrmer TCATCACCACCTTCGAAAATAGC (soluble form) lower primer CCGCATATTTTAAGTGACCGCT 415
internal probe TGGTGCAACTCATCTCGGTCT _~I~~--__----_---‘-_-_---_--___---_--___------_-__-_-----_ upper primer TCATCACCACCTTCGAAAATAGC
(trans-membrane lower primer GCGCAACAATGGGAGCCAGT 340 form) internal probe TGGTGCAACTCATCTCGGTCT __________-_--___----_-_------_--------_--_-----___-_____ TNF-a upper primer TCCCCAAAGGGATGAGAAGTTC
lower primer TCATACCAGGG’TTTGAGCTCAG 411 internal probe CACACTCAGATCATCTTCTC ________-_-----_~---__----------------~-_--_~~~~-_-_--_--
JllD upper prrmer GACATGGGCAGAGCGATGGTGGCCAG lower primer GGACCTGTTTCTTCCTGATCACATTGGAC 328 internal probe AGAGAGAGCCAGGAGACCAG
Results
Eflects of biochanin A on the proliferation of JCS cells:
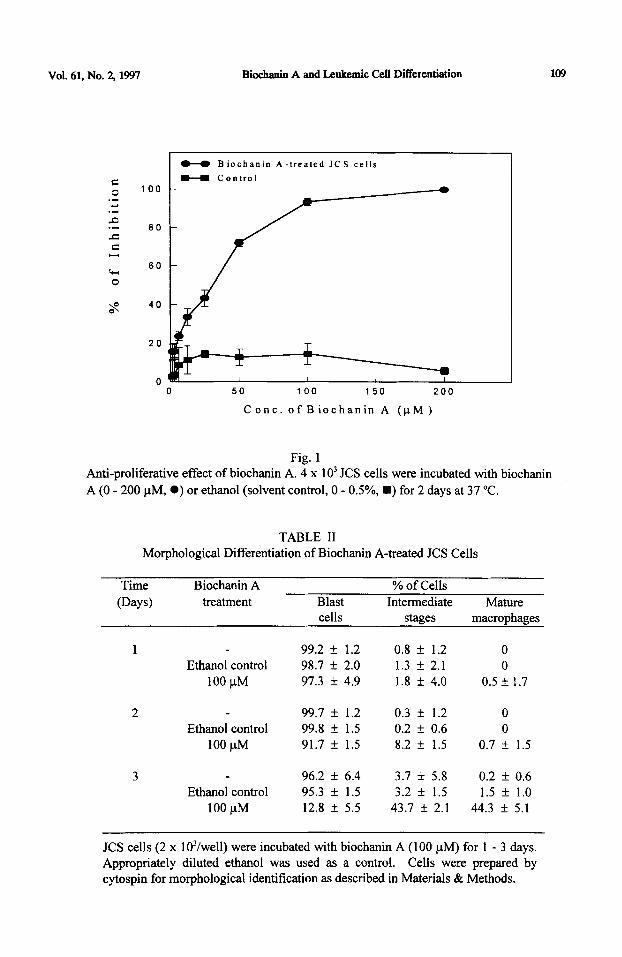
The effects of biochanin A on the growth of JCS cells were investigated by incubating the cells with various concentrations of biochanin A (0 - 200 l.tM). Appropriately diluted ethanol was used as a solvent control. Figure 1 shows that biochanin A suppressed the growth of JCS cells in a dose-dependent manner.
Effects of biochanin A on the difSerentiation of JCS cells:
The morphological differentiation of biochanin A-treated JCS cells was investigated by
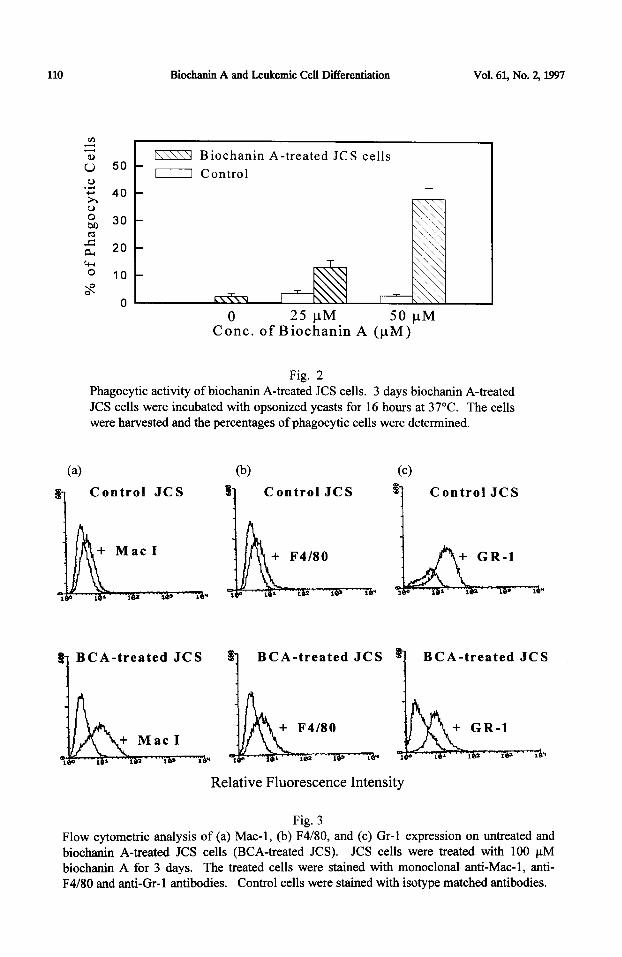
incubating 4 x 1 O3 JCS cells with biochanin A (100 PM) for 3 days. Significant morphological differentiation was observed 3 days after incubation (Table II). Ethanol alone (0.25 - 0.5 %) had no effect on the JCS cells. We have previously demonstrated that morphological differentiation is associated with an increase in functional activity of the cells (9). In this study, the percentage of phagocytic cells was significantly increased when JCS cells were exposed to biochanin A (100 PM) (Figure 2).
Vol. 61, No. 2, 1997 Biochania A and Leukemic Cell Differentiation 109
100
80
80
40
20
0
0-O Biochanin A-treated JCS cells
H Control
0 50 100 150 200
C enc. of B iochanin A (pM )
Fig. 1 Anti-proliferative effect of biochanin A. 4 x 10’ JCS cells were incubated with biochanin
A (0 - 200 pM, 0) or ethanol (solvent control, 0 - OS%, n ) for 2 days at 37 “C.
TABLE II Morphological Differentiation of Biochanin A-treated JCS Cells
Time Biochanin A
(Days) treatment Blast % of Cells
Intermediate Mature cells stages macrophages
1 99.2 + 1.2 0.8 k 1.2 0 Ethanol control 98.7 + 2.0 1.3 f 2.1 0
100 uM 97.3 k 4.9 1.8 f 4.0 0.5 f I.7
2 99.7 k 1.2 0.3 + 1.2 0 Ethanol control 99.8 + 1.5 0.2 f 0.6 0
100 PM 91.7 k 1.5 8.2 f 1.5 0.7 + 1.5
3 96.2 + 6.4 3.7 f 5.8 0.2 f 0.6 Ethanol control 95.3 f 1.5 3.2 k 1.5 1.5 f 1.0
100 PM 12.8 f 5.5 43.7 f 2.1 44.3 + 5.1
JCS cells (2 x lO’/well) were incubated with biochanin A (100 PM) for 1 - 3 days. Appropriately diluted ethanol was used as a control. Cells were prepared by cytospin for morphological identification as described in Materials & Methods.
110 Biochanin A and Leukemic Cell Differentiation Vol. 61, No. 2,1997
I Control
0 25 pM 50 pM Cont. of Biochanin A (PM)
Fig. 2 Phagocytic activity of biochanin A-treated JCS cells. 3 days biochanin A-treated JCS cells were incubated with opsonized yeasts for 16 hours at 37°C. The cells were harvested and the percentages of phagocytic cells were determined.
(4
8-l Control JCS
4 B CA-treated JCS
@I % 1
Control JCS
f
1 BCA-treated JCS
Cc) 3 7 Control JCS
Relative Fluorescence Intensity
1 BCA-treated JCS
Fig. 3 Flow cytometric analysis of (a) Mac- 1, (b) F4/80, and (c) Gr- 1 expression on untreated and biochanin A-treated JCS cells (BCA-treated JCS). JCS cells were treated with 100 PM biochanin A for 3 days. The treated cells were stained with monoclonal anti-Mac-l, anti- F4/80 and anti-Gr-1 antibodies. Control cells were stained with isotype matched antibodies.

Vol. 61, No. 2, 1997 Biochanin A and Leukemic Cell Differentiation 111
Phenotypic characterization biochanin A-treated JCL’S cells: The expression of macrophage differentiation antigens (Mac-l and F4/80) and granulocyte marker Gr-1 were analyzed by flow cytometry (Figure 3). After 3 days’ treatment with biochanin A, JCS cells showed an increase in expression of both Mac-l and F4l80. The
expression of Mac-l on the biochanin A-treated cells was slightly higher than F4/80. In
contrast, the expression of Gr-1 was down-regulated in the treated cells as compared to the untreated control cells.
mRNA phenovping: Cytokines, such as IL-l, LIF, TNF-a, and IL-4, have been shown to regulate the growth and differentiation of hematopoietic cells (15-18). In this study, we examined the expression of cytokine genes after treatment of the cells with biochanin A. Cycle titration, dot blot and Southern blot analysis were used to evaluate the change in gene expression in a semi-quantitative approach. Total RNA was collected from JCS cells treated with 50 pM biochanin A for 18 and 46 hours. Primers specific for IL-la, IL-I p, IL-4, LIF, TNF-a and LIF receptors were used in this study (Table I). No IL- 1 a mRNA transcript was detected at 0 and 18 hours of induction, but IL-la was slightly up-regulated after treatment with biochanin A for 46 hours (Figure 4).
IL- la
(546 bp)
IL-1 fi
(382 bp)
IL-4
(422 bp)
LIFR
0 Cl8 C46 B18 B46
0 Cl8 C46 B18 846 0 Cl8 C46 B18 846 +
(transmembrane form) (340 bp)
LIFR (soluble form)
(415 bp)
35 cycles 50 cycles
Fig. 4 Southern blot analysis of the RT-PCR products of IL-la, IL-lp, IL-4, and LIF receptor (both transmembrane and soluble forms). RT-PCR products amplified from the RNA samples of the JCS cells which were untreated (0), treated with 0.25% ethanol for 18 (Cl 8) or 46 (C46) hours, or treated with 50 yM biochanin A for 18 (B18) or 46 (B46) hours were separated on 2% TAE agarose gel, blotted to the nylon membrane and hybridized with the appropriate internal probe. Signals were developed as described in Materials and Methods. + : PCR fragment of the corresponding gene used as a positive control.
112 Biochanin A and Leukemic Cell Ditrerentiation Vol. 61, No. 2, 1997
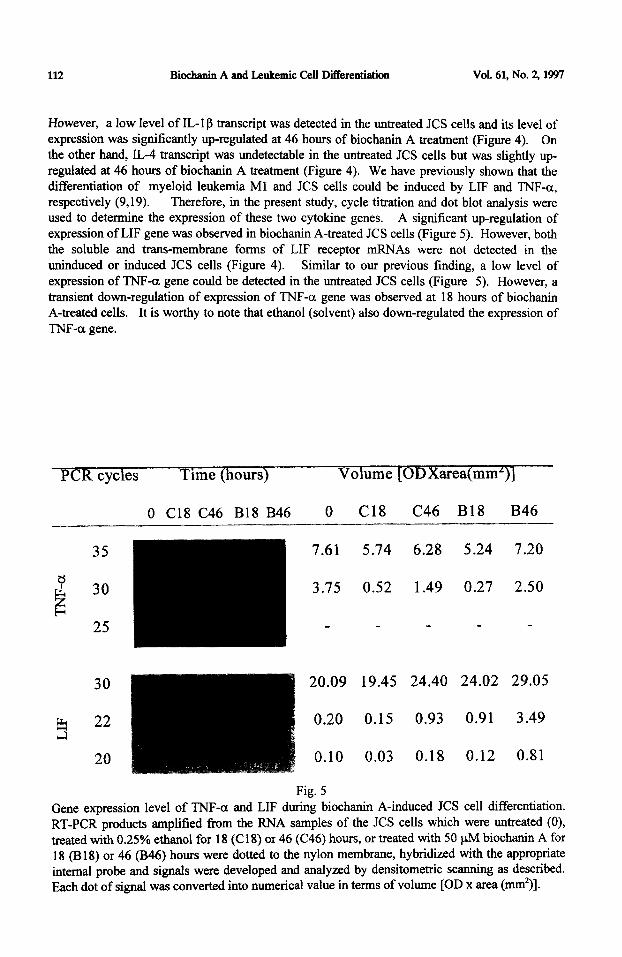
However, a low level of IL- 1 g transcript was detected in the untreated JCS cells and its level of expression was significantly up-regulated at 46 hours of biochanin A treatment (Figure 4). On the other hand, IL-4 transcript was undetectable in the untreated JCS cells but was slightly up- regulated at 46 hours of biochanin A treatment (Figure 4). We have previously shown that the differentiation of myeloid leukemia Ml and JCS cells could be induced by LIF and TNF-a, respectively (9,19). Therefore, in the present study, cycle titration and dot blot analysis were used to determine the expression of these two cytokine genes. A significant up-regulation of expression of LIF gene was observed in biochanin A-treated JCS cells (Figure 5). However, both the soluble and trans.-membrane forms of LIF receptor mRNAs were not detected in the uninduced or induced JCS cells (Figure 4). Similar to our previous finding, a low level of expression of TNF-cr gene could be detected in the untreated JCS cells (Figure 5). However, a transient down-regulation of expression of TNF-a gene was observed at 18 hours of biochanin A-treated cells. It is worthy to note that ethanol (solvent) also down-regulated the expression of
TNF-a gene.
1 cyc es ime ours
0 Cl8 C46 B18 B46 0 Cl8 C46 B18 B46
35
e & 30
E 25
30
b 22 cl
20
m 7.61 5.74 6.28 5.24 7.20
3.75 0.52 1.49 0.27 2.50
20.09 19.45 24.40 24.02 29.05
0.20 0.15 0.93 0.91 3.49
0.10 0.03 0.18 0.12 0.81
Fig. 5
Gene expression level of TNF-a and LIF during biochanin A-induced JCS cell differentiation. RT-PCR products amplified from the RNA samples of the JCS cells which were untreated (0), treated with 0.25% ethanol for 18 (C18) or 46 (C46) hours, or treated with 50 uM biochanin A for 18 (B 18) or 46 (B46) hours were dotted to the nylon membrane, hybridized with the appropriate internal probe and signals were developed and analyzed by densitometric scanning as described. Each dot of signal was converted into numerical value in terms of volume [OD x area (mm*)].

Vol. 61, No. 51597 Biochanin A and Leukemic Cell Differentiation 113
Discussion
Many naturally occurring anti-tumour compounds were identified and isolated from plant species. Epidemiological studies have also indicated the relationship between the incidence of cancer and dietary factors (20-21). These natural anti-tumour compounds include proteins (22) polysaccharides (23) and small molecules such as flavonoids (24). Biochanin A, a naturally occurring isoflavone, has been identified and detected in various diets and plant species (25-26). Biochanin A has been shown to mediate multiple biological effects including anti-mutagenic activity in mammalian cells (27-28), anti-tumour activity (2,4) and modulation of intracellular signal transduction (29). The anti-tumour effect of biochanin A has been correlated with its cytotoxic and cytostatic properties (2,4). To gain more insight in the possible application of biochanin A as an anti-oncogenic agent, we have investigated the biological effects of this compound in controlling the growth and differentiation of a murine myeloid leukemia JCS cell line. We have previously shown that TNF-a and some other chemical compounds such as phorbol 12-myristate 13-acetate (PMA) could induce monocytic differentiation of the JCS cells. The present study demonstrated that biochanin A not only inhibited the growth of JCS cells, but also induced monocytic differentiation of JCS cells to macrophages. Morphological differentiation of leukemia JCS cells was associated with the increased expression of macrophage differentiation markers (Mac-l and F4/80), but not the granulocyte specific marker Gr-1. We have previously demonstrated that cytokine-induced maturation of JCS cells is associated with an increase in the phagocytic activity. Our results also showed that the phagocytic activity of biochanin A-treated JCS cells was also significantly increased (32).
It is well documented that cytokines are pleiotropic and have multiple biological activities on normal or neoplastic cells (30). The activation of cytokine genes is temporal and is usually restricted to the target cells. The anti-tumour effects of flavonoids such as flavone acetic acid was partly due to the up-regulation of expression of cytokine genes in the immune cells (31). In the present study, the expression of some regulatory cytokines during the differentiation process induced by biochanin A was examined. We found that both IL-la and IL-l l3 mRNAs were not significantly expressed at 18 hours but were up-regulated at 46 hours of induction. These results indicate that IL-l may be involved at the later stage rather than at the early stage of biochanin A-mediated differentiation of the JCS cells. More recently, we have shown that
endogenously produced IL-la and IL-l p are involved, at least in part, in the TNF-a-induced monocytic differentiation of JCS cells (32). In addition, exogenous IL-la and IL-l p were found to be capable of inhibiting the growth as well as inducing the differentiation of JCS cells, indicating that IL-l alone may directly trigger myeloid leukemic cell differentiation (32).
We had previously demonstrated that PMA-induced differentiation of JCS cells was mediated by TNF-a (9). The differentiation-inducing effect of TNF-a could be synergized by IL-4 (10). In this study, IL-4 mENA was detected at 46 hours after biochanin A treatment. On the other hand, endogenous TNF-a gene expression could be detected in uninduced JCS cells and its level of expression was down-regulated at 18 and 46 hours after biochanin A treatment. This decrease in TNF-a mRNA is most likely due to the solvent (ethanol) rather than biochanin A’s effect. The mechanism by which ethanol can induce a down-regulation of TNF-a gene expression in JCS cells remains obscure and is currently under investigation. Nevertheless, it is conceivable that IL4-induced by biochanin A may act synergistically with endogenous TNF-a at the later stage of biochanin A-triggered differentiation of JCS cells. Moreover, it has been demonstrated that LIF is a differentiation inducer for Ml myeloid leukemia cells (33). In this study, we found that the expression of LIF transcript was significantly up-regulated as early as 18 hours after biochanin A treatment. However, both the soluble and tram+membrane forms of LIF

114 Bioehanin A and Leukemic Cell Differentiation Vol. 61, No. 2, 1997
receptor mRNAs were not detected in the uninduced or induced JCS cells. Likewise, exogenous LIF was found to be incapable of inducing the differentiation of JCS cells (Fung et al., unpublished observation). These results, when taken together, suggest that the endogenous produced cytokines such as IL-la, IL-lo, IL-4 and TNF-a but not LIF, appear to be involved in the later stage of biochanin A-induced monocytic differentiation of myeloid leukemia JCS cells.
Acknowledgment
This work was supported by the CUHK direct grant, the Faculty Development Grant of HKBU, and the Hong Kong Research Grant Council (RGC /95-96/21).
1.
2.
3. 4. 5.
6.
7.
8. 9.
10.
11.
12. 13.
14.
15.
16. 17.
18. 19.
P.M. DEWICK, The Flavonoids: Advances in research since 1986, J.B. Harborne (Ed), Chapman & Hall (1993). K. YANAGIHARA, A. ITO, T. TOGE, and M. NUMOTO, Cancer Res. 53 5815-5821 (1993). G. PETERSON and S. BARNES, Biochem. Biophy. Res. Commun. 179 661- 667 (1991). G. PETERSON and S. BARNES, Prostate 22. 335-345 (1993). A.R. FRANCIS, T.K. SHETTY and R.K. BHATTACHARYA, Carcinogenesis u 1953-1955 (1989). R. EDENHARDER, I. VON-PETERSDORFF, and R. RAUSCHER, Mutation Res. 287 261-274 (1993). Y.S. LEE, J.S. SEO, H.T. CHUNG, and J.J. JANG, J. Korean Med. Sci. 6 325-328 (1991). Y.S. LEE, T.H. KIM, K.J. CHO, and J.J. JANG, In-Vivo 6 283-286 (1992). N.K. MAK, M.C. FLING, K.N.LEUNG, and A.J. I-LAPEL, Cell. Immunol. 150 1-14
(1993). K.N. LEUNG, N.K. MAK, M.C. FUNG, and A.J. HAPEL, Immunology &I 65-7
(1994). N.K. MAK, K.N. LEUNG, M.C. FLING, and A.J. HAPEL, Immunobiology 190 1-12
(1994). L.L. LANIER, andN.L. WARNER, J. Immunol. Methods 47 25-30 (1981). J.M. CHIRGWIN, A.E. PRZYBYLA, R.J. MACDONALD, and W.J. RUTTER, Biochemistry 18 5294-5299 (1979). C. F. MORRIS, C. J. SIMEONOVIC, M. C. FUNG, J. D. WILSON and A. J. HAPEL, J. Immunol. 154 2470-2482 (1995). K. ONOZAKI, H. URAWA, T. TAMATANI, Y. IWAMUFL4, T. HASHIMOTO, T. BABA, H. SUZUKI, M. YAMADA, S. YAMAMOTO, J.J. OPPENHEIM, and K. MATSUSHIMA, J. Immunol. 140 112-119 (1988). C. SCHEIBENBOGEN and R. ANDREESEN, J. Leuk. Biol. 50 35-42 (1991). J. LOTEM and L. SACHS, Leukemia Res. II. 13 - 21(1989).
J. LOTEM, Y. SHABO, and L. SACHS, Cell Growth & Differentiation 2 421-427 (1991). M.C. FlJNG, N.K. MAK, K.N. LEUNG, and A.J HAPEL, Cell. Immunol. 141 121-130 (1992).
20. R. DOLL, and R. PETO, J. Natl. Cancer Inst. &i 1191-1308 (1981). 21. K.A. STEINMETZ and J.D. POTTER, Cancer-Causes-Control 2 427-442 (1991). 22. P.C. SHAW, W.L. CHAN, H.W. YEUNG, and T.B. NG, Life Sci. 51 253-264 (1994).
References

Vol. 61, No. 2, 1997 Biochauh A and Leukemic Cell Differentiation 115
23. M. HASHI, Bulletin of the Forestry and Forest Products Research Institute 360 121-148 (1991).
24. T. HIRANO, M. GOTOH, and K. OKA, Life Sci. 55 1061-1069 (1994). 25. J.T. DWYER, B.R. GOLDIN, N. SAUL, L. GUALTIERI, S. BARAKAT, and
H. ADLERCREUTZ, J. Am. Diet. Assoc. 94 739-743 (1994). 26. W.M. KEUNG, Alcohol Clin. Exp. Res. 17 1254-1260 (1993). 27. J.M. CASSADY, T.M. ZENNIE, Y.H. CHAE, M.A. FERIN, N.E. PORTUONDO, and
W.M. BAIRD, Cancer Res. 48 6257-6261 (1988). 28. Y.H. CHAE, D.K. HO, J.M. CASSADY, V.M. COOK, C.B. MARCUS, and W.M. BAIRD,
Chem-Biol. Interact. 82 181-193 (1992). 29. C.H. VERSANTVOORT, G.J. SCHUURHUIS, H.M. PINEDO, CA. EEKMAN,
C.M. KUIPER, J. LANKELMA, and H.J. BROXTERMAN, Br. J. Cancer @ 939-946 (1993).
30. K.I. ARAI, F. LEE, A. MIYAJIMA, S. MIYATAKE, N. ARAI, and T. YOKOTA, Amm. Rev. Biochem. 59 783-836 (1990).
3 1. K.F. MACE, R.L. HORNUNG, R.H. WILTROUT , and H.A. YOUNG, Cancer Res. s 1742-1747 (1990).
32. SC. CHAN, M.C. FUNG, N.K. MAK, and K.N. LEUNG, Int. J. Oncol. (in press). 33. N.M. GOUGH, R.L. WILLIAMS, D.J. HILTON, S. PEASE, T.A. WILLSON, J. STAHL,
D.P. GEARING, N.A. NICOLA, and D. METCALF, Reprod. Fertil. Dev. 1281-288 (1989).
Related Documents







![Phenolic Profiles and Polyphenol Oxidase (PPO) Gene ... · p-ethylphenol, a metabolite of biochanin A, have been linked to decreased bovine reproduction . However, [15] biochanin](https://static.cupdf.com/doc/110x72/607469627bc5c160f7329b6c/phenolic-profiles-and-polyphenol-oxidase-ppo-gene-p-ethylphenol-a-metabolite.jpg)



