Effects of anthropogenic seawater acidification on acid–base balance in the sea urchin Psammechinus miliaris Hayley Miles a , Stephen Widdicombe b, * , John I. Spicer a , Jason Hall-Spencer a a Marine Biology and Ecology Research Centre, School of Biological Sciences, University of Plymouth, Drake Circus, Plymouth, Devon PL4 8AA, UK b Plymouth Marine Laboratory, Prospect Place, West Hoe, Plymouth, Devon PL1 3DH, UK Abstract The purple-tipped sea urchin, Psammechinus miliaris, was exposed to artificially acidified seawater treatments (pH w 6.16, 6.63 or 7.44) over a period of 8 days. Urchin mortality of 100% was observed at pH w 6.16 after 7 days and coincided with a pronounced hypercapnia in the coelomic fluid producing an irrecoverable acidosis. Coelomic fluid acid–base measures showed that an accumulation of CO 2 and a significant reduction in pH occurred in all treatments compared with controls. Bicarbonate buffering was employed in each case, reducing the resultant acidosis, but compensation was incomplete even under moderate environmental hypercapnia. Significant test dissolution was inferred from observable increases in the Mg 2+ concentration of the coelomic fluid under all pH treatments. We show that a chronic reduction of surface water pH to below 7.5 would be severely detrimental to the acid–base balance of this predominantly intertidal spe- cies; despite its ability to tolerate fluctuations in pCO 2 and pH in the rock pool environment. The absence of respiratory pigment (or any substantial protein in the coelomic fluid), a poor capacity for ionic regulation and dependency on a magnesium calcite test, make echi- noids particularly vulnerable to anthropogenic acidification. Geological sequestration leaks may result in dramatic localised pH reduc- tions, e.g. pH 5.8. P. miliaris is intolerant of pH 6.16 seawater and significant mortality is seen at pH 6.63. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Carbon dioxide; Hypercapnia; Ocean acidification; Carbon sequestration; Acid–base balance; Psammechinus miliaris 1. Introduction Over 80% of global energy use releases carbon dioxide (Sundquist, 1993) and this has resulted in an increase in the atmospheric CO 2 concentration of over 30% since 1800 (Houghton et al., 2001) with the prospect of concen- trations exceeding 1500 ppm sometime between 2100 and 2200 (Po ¨rtner et al., 2004). This rate of increase is thought to be 100 times greater than the natural fluctuations seen over recent millennia (Blackford and Gilbert, in press). The Intergovernmental Panel on Climate Change (IPCC) predicted that by 2100, atmospheric levels of CO 2 could reach 800 ppm (Feely et al., 2004). A situation in contrast to the stability of the past 24 million years during which time levels have remained below 500 ppm (Pearson and Palmer, 2000). Atmospheric CO 2 diffuses passively into global ocean surface waters and causes not only an increase in the partial pressure of the CO 2 but also a reduction in seawater pH. With the current rate of pH change standing at 0.015 units per decade (Haugan and Drange, 1996) it is probable that by 2100 a global surface water reduction in pH of up to 0.5 units will occur. Pressure to reduce CO 2 emissions in response to the threat of climate change has led to governments seeking new options for CO 2 mitiga- tion. One option is geological sequestration. This method of storage involves injecting CO 2 into underground porous reservoir rocks (Holloway, 2005) and has been in use at the Sleipner West gas field in the Norwegian sector of the North Sea since 2000. At this site, around 1 · 10 6 ty 1 CO 2 are currently being sequestered (Holloway, 2005). Whilst geological storage appears to be a practical tool in reducing emissions (Gibbins et al., 2005), little is known about the long term issues which will arise from underground storage 0025-326X/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2006.09.021 * Corresponding author. Tel.: +44 1752 633100; fax: +44 1752 633101. E-mail address: [email protected] (S. Widdicombe). www.elsevier.com/locate/marpolbul Marine Pollution Bulletin 54 (2007) 89–96

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/marpolbul
Marine Pollution Bulletin 54 (2007) 89–96
Effects of anthropogenic seawater acidification on acid–base balancein the sea urchin Psammechinus miliaris
Hayley Miles a, Stephen Widdicombe b,*, John I. Spicer a, Jason Hall-Spencer a
a Marine Biology and Ecology Research Centre, School of Biological Sciences, University of Plymouth, Drake Circus, Plymouth, Devon PL4 8AA, UKb Plymouth Marine Laboratory, Prospect Place, West Hoe, Plymouth, Devon PL1 3DH, UK
Abstract
The purple-tipped sea urchin, Psammechinus miliaris, was exposed to artificially acidified seawater treatments (pHw 6.16, 6.63 or 7.44)over a period of 8 days. Urchin mortality of 100% was observed at pHw 6.16 after 7 days and coincided with a pronounced hypercapniain the coelomic fluid producing an irrecoverable acidosis. Coelomic fluid acid–base measures showed that an accumulation of CO2 and asignificant reduction in pH occurred in all treatments compared with controls. Bicarbonate buffering was employed in each case, reducingthe resultant acidosis, but compensation was incomplete even under moderate environmental hypercapnia. Significant test dissolutionwas inferred from observable increases in the Mg2+ concentration of the coelomic fluid under all pH treatments. We show that a chronicreduction of surface water pH to below 7.5 would be severely detrimental to the acid–base balance of this predominantly intertidal spe-cies; despite its ability to tolerate fluctuations in pCO2 and pH in the rock pool environment. The absence of respiratory pigment (or anysubstantial protein in the coelomic fluid), a poor capacity for ionic regulation and dependency on a magnesium calcite test, make echi-noids particularly vulnerable to anthropogenic acidification. Geological sequestration leaks may result in dramatic localised pH reduc-tions, e.g. pH 5.8. P. miliaris is intolerant of pH 6.16 seawater and significant mortality is seen at pH 6.63.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Carbon dioxide; Hypercapnia; Ocean acidification; Carbon sequestration; Acid–base balance; Psammechinus miliaris
1. Introduction
Over 80% of global energy use releases carbon dioxide(Sundquist, 1993) and this has resulted in an increase inthe atmospheric CO2 concentration of over 30% since1800 (Houghton et al., 2001) with the prospect of concen-trations exceeding 1500 ppm sometime between 2100 and2200 (Portner et al., 2004). This rate of increase is thoughtto be 100 times greater than the natural fluctuations seenover recent millennia (Blackford and Gilbert, in press).The Intergovernmental Panel on Climate Change (IPCC)predicted that by 2100, atmospheric levels of CO2 couldreach 800 ppm (Feely et al., 2004). A situation in contrastto the stability of the past 24 million years during whichtime levels have remained below 500 ppm (Pearson and
0025-326X/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.marpolbul.2006.09.021
* Corresponding author. Tel.: +44 1752 633100; fax: +44 1752 633101.E-mail address: [email protected] (S. Widdicombe).
Palmer, 2000). Atmospheric CO2 diffuses passively intoglobal ocean surface waters and causes not only an increasein the partial pressure of the CO2 but also a reduction inseawater pH. With the current rate of pH change standingat 0.015 units per decade (Haugan and Drange, 1996) it isprobable that by 2100 a global surface water reduction inpH of up to 0.5 units will occur. Pressure to reduce CO2
emissions in response to the threat of climate change hasled to governments seeking new options for CO2 mitiga-tion. One option is geological sequestration. This methodof storage involves injecting CO2 into underground porousreservoir rocks (Holloway, 2005) and has been in use at theSleipner West gas field in the Norwegian sector of theNorth Sea since 2000. At this site, around 1 · 106 t y�1
CO2 are currently being sequestered (Holloway, 2005). Whilstgeological storage appears to be a practical tool in reducingemissions (Gibbins et al., 2005), little is known about thelong term issues which will arise from underground storage

90 H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96
of CO2. Subsurface storage leaks are possible over time(Hawkins, 2004) and ocean currents may carry acidifiedwater to vulnerable areas nearer to shore and in the deepocean.
Little is known about how marine organisms will beaffected by both long and short term seawater acidification,increased carbon dioxide partial pressure and other pHassociated changes to ocean chemistry (Turley et al.,2006). However, it is likely that marine organisms will beaffected on a number of levels including acid–base regula-tion, growth, reproduction, feeding and ultimately mortal-ity (Kurihara et al., 2004; Kurihara and Shirayama, 2004;Raven et al., 2005). In the few studies conducted manymarine animals were only able to tolerate brief periods ofexposure to pH values below 7. Furthermore, reductionof pH below 7.5 may sufficient to cause severe damageand should be avoided (Ormerod and Angel, 1996).
Carbon dioxide induced changes in seawater chemistrycould affect different groups of marine organisms in differ-ent ways. Particularly vulnerable are those organismswhich rely on the production of calcified tests or shellsfor survival, e.g. corals (Seibel and Fabry, 2003), molluscs(Lindinger et al., 1984; Michaelidis et al., 2005), crusta-ceans (DeFur and McMahon, 1984) and calcified algal spe-cies (Riebesell et al., 2000). Of these, echinoderms could beespecially at risk as they use aragonite (MgCO3) in test con-struction both as adults and juveniles (Raven et al., 2005).Aragonite is 30 times more soluble than non-magnesiumcontaining calcite (Politi et al., 2004) and therefore dissolu-tion could occur more readily. The primary problemcaused by seawater acidification for organisms that relyon biogenic calcification is the reduced availability of car-bonate ions (CO�3 ). If atmospheric CO2 concentrationsreach 800 ppm, the corresponding drop in CO�3 availabilityin surface waters worldwide would be around 60% (Feelyet al., 2004). This drop in CO�3 availability means organ-isms will be less able to deposit calcium carbonate struc-tures and, if the reduction in CO�3 availability is severeenough, calcite dissolution can occur (Raven et al., 2005).
The current study focuses on the acid–base response ofthe predominantly intertidal, purple-tipped sea urchin,Psammechinus miliaris, to anthropogenic environmentalhypercapnia. This species was chosen for a number of rea-sons. Firstly, P. miliaris is instrumental in key ecosystemprocesses, e.g. the control of macro-epiphytic biomass(Guillou et al., 2002) and the decay of kelp debris (Bedfordand Moore, 1985). This species is even used as a biologicalcontrol of fouling organisms in suspended scallop cultiva-tion (Ross et al., 2004). Secondly, the acid–base balanceof P. miliaris is known to be sensitive to hypercapnia asso-ciated with emersion (Spicer et al., 1988) and at night inintertidal rockpools (Spicer, 1995). This sensitivity hasbeen attributed to the fact that the composition of coelomicfluid in echinoderms varies with that of the surroundingseawater (Farmanfarmaian, 1966). In addition, P. miliaris
possess no respiratory pigment and relies only on favour-able diffusion gradients for gas exchange. The water vascu-
lar system in echinoids, particularly the tube feet, is theprimary medium for gas exchange (Johansen and Vadas,1967). For this reason, the CO2 content of the coelomicfluid is always slightly greater than that found in seawater.
The current study uses laboratory based experimentationto investigate the physiological consequences of acidifiedseawater exposure for the ecologically important speciesP. miliaris, an approach advocated by previous researchers,e.g. Langenbuch and Portner (2004) where the marinesipunculid Sipunculus nudus was exposed to elevated CO2.The study uses a wide range of pH exposure values to rep-resent scenarios encompassing both atmospheric CO2 emis-sions during the next 100 years and potentially dramaticsequestration leaks causing severe reductions in pH, i.e.down to 5.6. This leakage scenario is analogous to a naturalCO2 seep site located in the Bay of Naples, Italy (Hall-Spen-cer, J., unpublished observation). The study also uses longerexposure times than have previously been conducted inorder to assess the physiological response outside of condi-tions that the organisms would normally be exposed to dur-ing normal tidal cycles.
2. Materials and methods
2.1. Collection and maintenance of urchins
Urchins (test diameter = 3–5 cm) were collected by handfrom intertidal pools at Mount Batten, Plymouth, Devon,UK (50:21:34N, 04:07:45W) during July 2005. Urchinswere transported to Plymouth Marine Laboratory in con-tainers of seawater immediately after collection and main-tained in aquaria (vol. = 20 l) at stocking densities ofaround 20 individuals per aquarium. Each aquarium wassupplied with recirculating seawater (salinity = 35 PSU,temperature = 15 �C). Individuals were kept under theseconditions for up to 2 weeks before experimental use, thisensured that any individuals used were not weak or dis-tressed from the transfer. During the holding period urch-ins were fed ad libitum on Fucus serratus.
To assess the comparability of experimental controlspecimens with natural field populations, additional urch-ins were collected from the original collection site on26th July 2005. Samples of coelomic fluid were extractedfrom these urchins in the field and the coelomic fluid wasreturned to the laboratory and analysed for the same phys-iological parameters as described below for experimentalurchin samples.
2.2. Acidification of natural seawater
Seawater was collected from the Eddystone area, 20 kmoffshore (Plymouth, Devon) and used to fill four large(vol. = 450 l) header tanks. One of these tanks containedambient seawater at pH 7.95 and was used as a control.The three others were acidified by passing CO2 gas throughthe water as a fine flow of bubbles. Aeration and mixing ofthe gas was achieved using large aquarium pumps fitted in

H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96 91
each tank. When the required pH was achieved, the supplyof gaseous CO2 stopped. As the header tank water wasbeing constantly drained for use in the experimental tanks,the header tanks were automatically topped up with waterfrom a separate reservoir. This reservoir having been filledwith water collected at the same time and location as thewater used to fill the header tanks initially. The introduc-tion of ambient seawater (pH � 7.95) increased the pH ofthe acidified water in the header tanks, triggering therelease of further gaseous CO2 to counter this change. Inthis way the header tanks were kept at consistent, pro-grammed levels of acidification.
The four header tanks used in this experiment containedwater of differing pH values, allowing four treatment levelsto be run together. Values of pH were set and monitoredusing a controller (Walchem Webmaster-GI) in connectionwith four pH electrodes (S650CD), one for each headertank. The system measured pH to an accuracy of 0.01pH units and controlled CO2 release and maintenance ofpH values to set limits. Targets for pH maintenance wereset and programmed as follows: tank 1 – 5.6, tank 2 –6.5, tank 3 – 7.5, tank 4 – ambient seawater (pH 7.9). Theselevels were chosen as they represent scenarios which couldpotentially impact on marine life. In the Bay of Naples, aseawater pH of 5.6 can be found in association with a nat-ural CO2 seep site (99% CO2 in gas composition) whichreleases 1.5 million litres of gas per day (Hall-Spencer, J.,unpublished observation). This may reflect potential condi-tions after a geological CO2 sequestration site leak. A pHvalue of 7.5 is the Intergovernmental Panel on ClimateChange’s predicted global ocean surface water pH for theend of this century (Feely et al., 2004). The final treatmentlevel of pH 6.5 was chosen as an intermediate betweenthese two scenarios.
3. Exposure of urchins to acidified seawater
Exposure experiments were carried out at the PlymouthMarine Laboratory mesocosm facility. This is a large con-stant temperature room where marine environment simula-tion experiments are conducted. Five P. miliaris individualswere placed in each of four exposure tanks (vol. = 60 l)containing seawater as described in Table 1. Water flowedin from the header tanks at a constant rate of 100 ml min�1
and drained to waste, producing a turnover rate of twice a
Table 1Physicochemical parameters (±1 standard deviation) of the ambient andacidified seawater used during sea urchin exposure
Parameter Tank 1 Tank 2 Tank 3 Controltank 4
pH 6.16 (±0.11) 6.63 (±0.11) 7.46 (±0.03) 7.96 (±0.07)Total CO2
(mmol l�1)4.54 (±0.75) 3.26 (±0.55) 2.3 (±0.41) 1.66 (±0.45)
PartialpressureCO2 (Torr)
39.7 (±9.75) 17.3 (±0.11) 1.8 (±0.11) 0.39 (±0.11)
day. Circulation of water in the exposure tanks wasensured using small aquarium pumps and nylon bags con-taining activated charcoal (25 g) were fixed to the tank sideand submerged to minimise ammonia build up. Individualswere contained within labelled baskets of seasoned plasticmesh and opaque plastic sheeting was put over each tankto prevent water–air flux of CO2 and growth of contami-nant algae. Measures of water pH and total CO2 concen-tration were taken daily from these tanks to ensure thatany fluctuations over the experimental time were noted.
3.1. Measurement of pH and CCO2, calculation of pCO2 and
HCO3� concentration in coelomic fluid samples
Before the urchins were placed in the exposure tanks andexposed to acidified water, a 0.1 ml sample of coelomic fluidwas extracted from each individual P. miliaris. The urchinswere then placed into the exposure tanks and the experi-ment began. After 24 h a 0.1 ml sample of coelomic fluidwas again extracted from each urchin. This procedure con-tinued daily for 8 days or until an animal died. Sampleswere only taken from living urchins. To determine whetheran urchin was alive it was first inspected whilst in seawater.If the urchin’s spines and tube feet could be seen moving,the urchin was alive and a sample was taken. If there wasno movement, the urchin would be removed from the waterand examined closely for at least 10 min for any signs ofmovement, particularly in the area around the mouth parts.If there was no spine movement or all the spines had beendropped and the mouth parts had turned black the urchinwas assumed to be dead and no sample was taken. It wasnoted that in association with these visual indicators, deadurchins had a distinctive and unpleasant smell. If there wasany sign of movement in any remaining spines and therewas no smell the urchin was considered living and was sam-pled before being returned to the experimental system.
Coelomic fluid samples were collected using a 0.1 ml syr-inge, the needle (21 G) of which was inserted directly intothe pervisceral coelomic space. Subsamples were then usedto determine the total amount of CO2 dissolved in the coe-lomic fluid (termed CCO2) and coelomic fluid pH (methoddescribed below). The remaining fluid was transferred to amicrocentrifuge tube (Eppendorf, vol. = 1.6 ml) kept at atemperature of 2 �C. Coelomic fluid pH was measured usinga micro pH electrode (Mettler Toledo Inlab423, Switzer-land) inserted directly into the sample in vitro within sec-onds of collection. Total CO2 concentration (mmol l�1)was measured on duplicate 10 ll sub-samples using a CO2
analyser (Ciba-Corning 965, England). The partial pressureof CO2 (pCO2) in both the coelomic fluid of each individualand of the water in the experimental tanks which they wereexposed to during the experiment was then calculated usingvalues for CCO2 and pH using a modified form of theHenderson–Hasselbalch equation (Spicer et al., 1988):
pCO2 ¼ CCO2=að10pH�pK i þ 1Þ ð1Þ
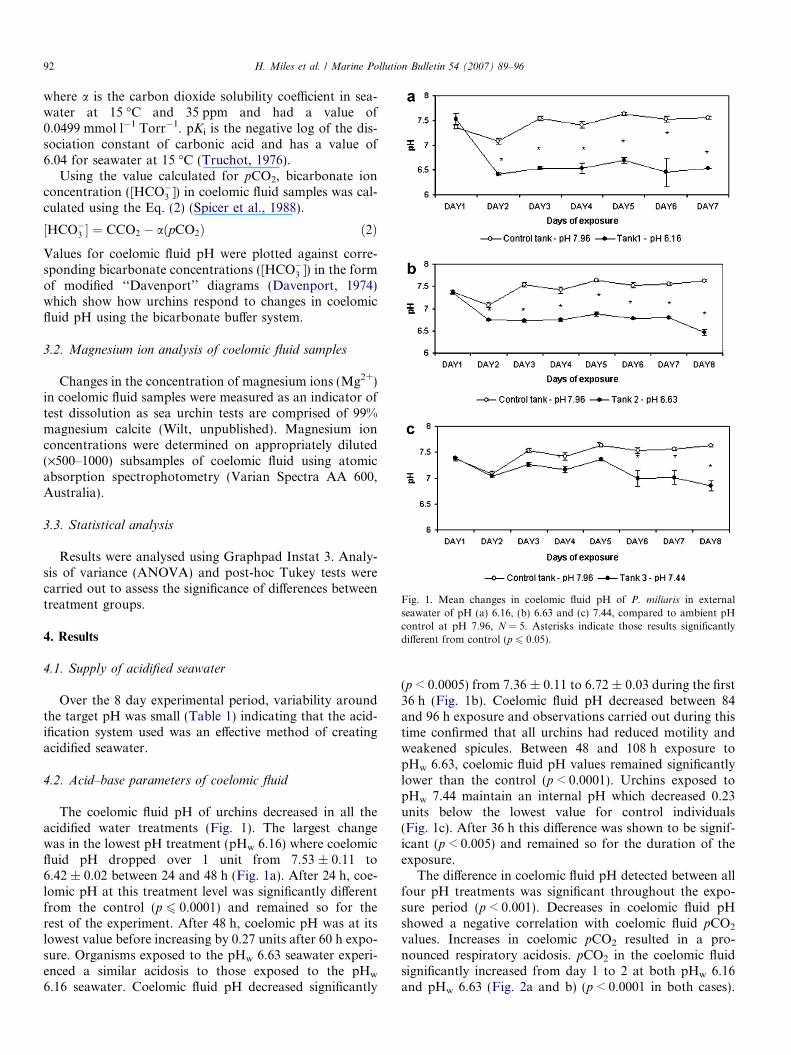
Fig. 1. Mean changes in coelomic fluid pH of P. miliaris in externalseawater of pH (a) 6.16, (b) 6.63 and (c) 7.44, compared to ambient pHcontrol at pH 7.96, N = 5. Asterisks indicate those results significantlydifferent from control (p 6 0.05).
92 H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96
where a is the carbon dioxide solubility coefficient in sea-water at 15 �C and 35 ppm and had a value of0.0499 mmol l�1 Torr�1. pKi is the negative log of the dis-sociation constant of carbonic acid and has a value of6.04 for seawater at 15 �C (Truchot, 1976).
Using the value calculated for pCO2, bicarbonate ionconcentration ([HCO�3 ]) in coelomic fluid samples was cal-culated using the Eq. (2) (Spicer et al., 1988).
½HCO�3 � ¼ CCO2 � aðpCO2Þ ð2Þ
Values for coelomic fluid pH were plotted against corre-sponding bicarbonate concentrations ([HCO�3 ]) in the formof modified ‘‘Davenport’’ diagrams (Davenport, 1974)which show how urchins respond to changes in coelomicfluid pH using the bicarbonate buffer system.
3.2. Magnesium ion analysis of coelomic fluid samples
Changes in the concentration of magnesium ions (Mg2+)in coelomic fluid samples were measured as an indicator oftest dissolution as sea urchin tests are comprised of 99%magnesium calcite (Wilt, unpublished). Magnesium ionconcentrations were determined on appropriately diluted(·500–1000) subsamples of coelomic fluid using atomicabsorption spectrophotometry (Varian Spectra AA 600,Australia).
3.3. Statistical analysis
Results were analysed using Graphpad Instat 3. Analy-sis of variance (ANOVA) and post-hoc Tukey tests werecarried out to assess the significance of differences betweentreatment groups.
4. Results
4.1. Supply of acidified seawater
Over the 8 day experimental period, variability aroundthe target pH was small (Table 1) indicating that the acid-ification system used was an effective method of creatingacidified seawater.
4.2. Acid–base parameters of coelomic fluid
The coelomic fluid pH of urchins decreased in all theacidified water treatments (Fig. 1). The largest changewas in the lowest pH treatment (pHw 6.16) where coelomicfluid pH dropped over 1 unit from 7.53 ± 0.11 to6.42 ± 0.02 between 24 and 48 h (Fig. 1a). After 24 h, coe-lomic pH at this treatment level was significantly differentfrom the control (p 6 0.0001) and remained so for therest of the experiment. After 48 h, coelomic pH was at itslowest value before increasing by 0.27 units after 60 h expo-sure. Organisms exposed to the pHw 6.63 seawater experi-enced a similar acidosis to those exposed to the pHw
6.16 seawater. Coelomic fluid pH decreased significantly
(p < 0.0005) from 7.36 ± 0.11 to 6.72 ± 0.03 during the first36 h (Fig. 1b). Coelomic fluid pH decreased between 84and 96 h exposure and observations carried out during thistime confirmed that all urchins had reduced motility andweakened spicules. Between 48 and 108 h exposure topHw 6.63, coelomic fluid pH values remained significantlylower than the control (p < 0.0001). Urchins exposed topHw 7.44 maintain an internal pH which decreased 0.23units below the lowest value for control individuals(Fig. 1c). After 36 h this difference was shown to be signif-icant (p < 0.005) and remained so for the duration of theexposure.
The difference in coelomic fluid pH detected between allfour pH treatments was significant throughout the expo-sure period (p < 0.001). Decreases in coelomic fluid pHshowed a negative correlation with coelomic fluid pCO2
values. Increases in coelomic pCO2 resulted in a pro-nounced respiratory acidosis. pCO2 in the coelomic fluidsignificantly increased from day 1 to 2 at both pHw 6.16and pHw 6.63 (Fig. 2a and b) (p < 0.0001 in both cases).
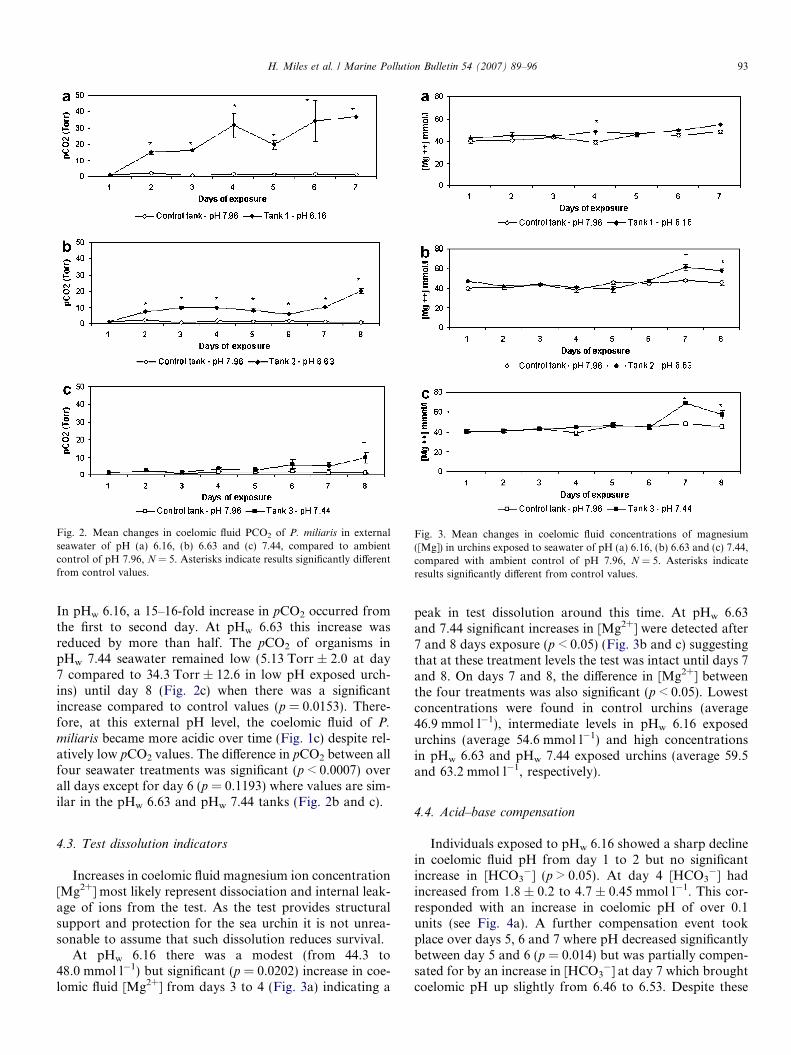
Fig. 2. Mean changes in coelomic fluid PCO2 of P. miliaris in externalseawater of pH (a) 6.16, (b) 6.63 and (c) 7.44, compared to ambientcontrol of pH 7.96, N = 5. Asterisks indicate results significantly differentfrom control values.
Fig. 3. Mean changes in coelomic fluid concentrations of magnesium([Mg]) in urchins exposed to seawater of pH (a) 6.16, (b) 6.63 and (c) 7.44,compared with ambient control of pH 7.96, N = 5. Asterisks indicateresults significantly different from control values.
H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96 93
In pHw 6.16, a 15–16-fold increase in pCO2 occurred fromthe first to second day. At pHw 6.63 this increase wasreduced by more than half. The pCO2 of organisms inpHw 7.44 seawater remained low (5.13 Torr ± 2.0 at day7 compared to 34.3 Torr ± 12.6 in low pH exposed urch-ins) until day 8 (Fig. 2c) when there was a significantincrease compared to control values (p = 0.0153). There-fore, at this external pH level, the coelomic fluid of P.
miliaris became more acidic over time (Fig. 1c) despite rel-atively low pCO2 values. The difference in pCO2 between allfour seawater treatments was significant (p < 0.0007) overall days except for day 6 (p = 0.1193) where values are sim-ilar in the pHw 6.63 and pHw 7.44 tanks (Fig. 2b and c).
4.3. Test dissolution indicators
Increases in coelomic fluid magnesium ion concentration[Mg2+] most likely represent dissociation and internal leak-age of ions from the test. As the test provides structuralsupport and protection for the sea urchin it is not unrea-sonable to assume that such dissolution reduces survival.
At pHw 6.16 there was a modest (from 44.3 to48.0 mmol l�1) but significant (p = 0.0202) increase in coe-lomic fluid [Mg2+] from days 3 to 4 (Fig. 3a) indicating a
peak in test dissolution around this time. At pHw 6.63and 7.44 significant increases in [Mg2+] were detected after7 and 8 days exposure (p < 0.05) (Fig. 3b and c) suggestingthat at these treatment levels the test was intact until days 7and 8. On days 7 and 8, the difference in [Mg2+] betweenthe four treatments was also significant (p < 0.05). Lowestconcentrations were found in control urchins (average46.9 mmol l�1), intermediate levels in pHw 6.16 exposedurchins (average 54.6 mmol l�1) and high concentrationsin pHw 6.63 and pHw 7.44 exposed urchins (average 59.5and 63.2 mmol l�1, respectively).
4.4. Acid–base compensation
Individuals exposed to pHw 6.16 showed a sharp declinein coelomic fluid pH from day 1 to 2 but no significantincrease in [HCO3
�] (p > 0.05). At day 4 [HCO3�] had
increased from 1.8 ± 0.2 to 4.7 ± 0.45 mmol l�1. This cor-responded with an increase in coelomic pH of over 0.1units (see Fig. 4a). A further compensation event tookplace over days 5, 6 and 7 where pH decreased significantlybetween day 5 and 6 (p = 0.014) but was partially compen-sated for by an increase in [HCO3
�] at day 7 which broughtcoelomic pH up slightly from 6.46 to 6.53. Despite these
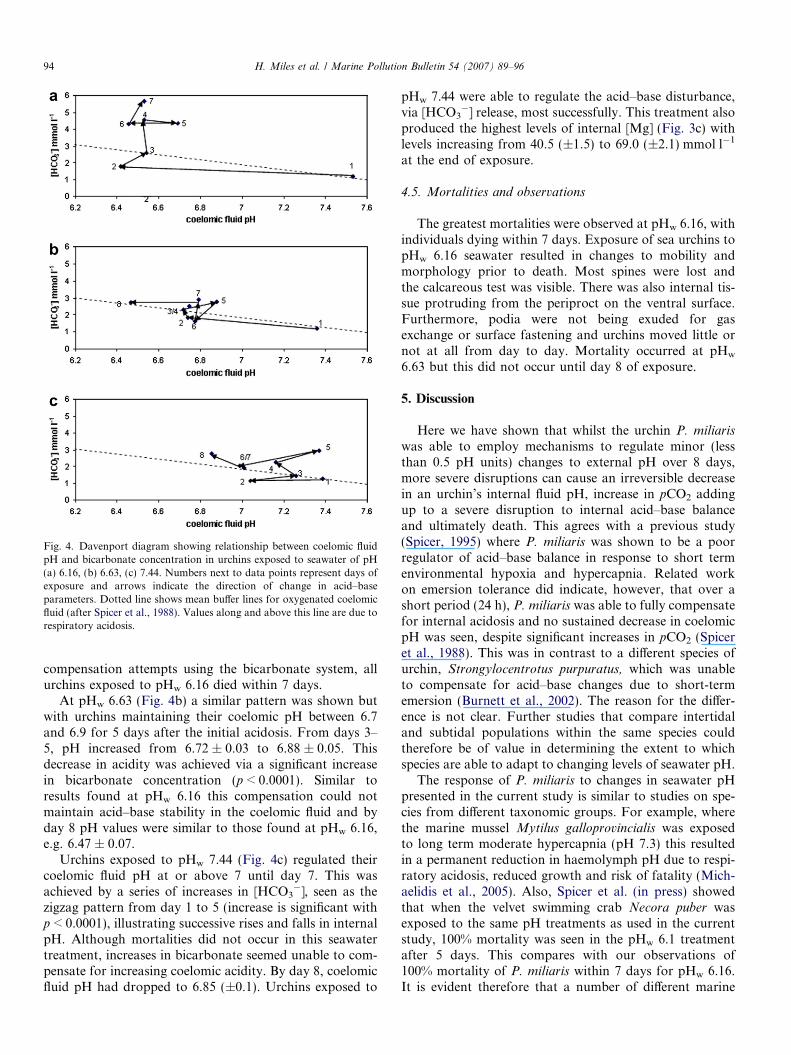
Fig. 4. Davenport diagram showing relationship between coelomic fluidpH and bicarbonate concentration in urchins exposed to seawater of pH(a) 6.16, (b) 6.63, (c) 7.44. Numbers next to data points represent days ofexposure and arrows indicate the direction of change in acid–baseparameters. Dotted line shows mean buffer lines for oxygenated coelomicfluid (after Spicer et al., 1988). Values along and above this line are due torespiratory acidosis.
94 H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96
compensation attempts using the bicarbonate system, allurchins exposed to pHw 6.16 died within 7 days.
At pHw 6.63 (Fig. 4b) a similar pattern was shown butwith urchins maintaining their coelomic pH between 6.7and 6.9 for 5 days after the initial acidosis. From days 3–5, pH increased from 6.72 ± 0.03 to 6.88 ± 0.05. Thisdecrease in acidity was achieved via a significant increasein bicarbonate concentration (p < 0.0001). Similar toresults found at pHw 6.16 this compensation could notmaintain acid–base stability in the coelomic fluid and byday 8 pH values were similar to those found at pHw 6.16,e.g. 6.47 ± 0.07.
Urchins exposed to pHw 7.44 (Fig. 4c) regulated theircoelomic fluid pH at or above 7 until day 7. This wasachieved by a series of increases in [HCO3
�], seen as thezigzag pattern from day 1 to 5 (increase is significant withp < 0.0001), illustrating successive rises and falls in internalpH. Although mortalities did not occur in this seawatertreatment, increases in bicarbonate seemed unable to com-pensate for increasing coelomic acidity. By day 8, coelomicfluid pH had dropped to 6.85 (±0.1). Urchins exposed to
pHw 7.44 were able to regulate the acid–base disturbance,via [HCO3
�] release, most successfully. This treatment alsoproduced the highest levels of internal [Mg] (Fig. 3c) withlevels increasing from 40.5 (±1.5) to 69.0 (±2.1) mmol l�1
at the end of exposure.
4.5. Mortalities and observations
The greatest mortalities were observed at pHw 6.16, withindividuals dying within 7 days. Exposure of sea urchins topHw 6.16 seawater resulted in changes to mobility andmorphology prior to death. Most spines were lost andthe calcareous test was visible. There was also internal tis-sue protruding from the periproct on the ventral surface.Furthermore, podia were not being exuded for gasexchange or surface fastening and urchins moved little ornot at all from day to day. Mortality occurred at pHw
6.63 but this did not occur until day 8 of exposure.
5. Discussion
Here we have shown that whilst the urchin P. miliaris
was able to employ mechanisms to regulate minor (lessthan 0.5 pH units) changes to external pH over 8 days,more severe disruptions can cause an irreversible decreasein an urchin’s internal fluid pH, increase in pCO2 addingup to a severe disruption to internal acid–base balanceand ultimately death. This agrees with a previous study(Spicer, 1995) where P. miliaris was shown to be a poorregulator of acid–base balance in response to short termenvironmental hypoxia and hypercapnia. Related workon emersion tolerance did indicate, however, that over ashort period (24 h), P. miliaris was able to fully compensatefor internal acidosis and no sustained decrease in coelomicpH was seen, despite significant increases in pCO2 (Spiceret al., 1988). This was in contrast to a different species ofurchin, Strongylocentrotus purpuratus, which was unableto compensate for acid–base changes due to short-termemersion (Burnett et al., 2002). The reason for the differ-ence is not clear. Further studies that compare intertidaland subtidal populations within the same species couldtherefore be of value in determining the extent to whichspecies are able to adapt to changing levels of seawater pH.
The response of P. miliaris to changes in seawater pHpresented in the current study is similar to studies on spe-cies from different taxonomic groups. For example, wherethe marine mussel Mytilus galloprovincialis was exposedto long term moderate hypercapnia (pH 7.3) this resultedin a permanent reduction in haemolymph pH due to respi-ratory acidosis, reduced growth and risk of fatality (Mich-aelidis et al., 2005). Also, Spicer et al. (in press) showedthat when the velvet swimming crab Necora puber wasexposed to the same pH treatments as used in the currentstudy, 100% mortality was seen in the pHw 6.1 treatmentafter 5 days. This compares with our observations of100% mortality of P. miliaris within 7 days for pHw 6.16.It is evident therefore that a number of different marine

H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96 95
organisms should be considered vulnerable to changes inseawater pH.
The extracellular acidosis in P. miliaris resulting fromexposure to pHw 6.16, pHw 6.63 and pHw 7.44 was accom-panied by significant increases in [HCO3
�] over the expo-sure period. We might expect this response as aquaticorganisms regularly use bicarbonate to compensate forrespiratory acidosis events (e.g. Henry et al., 1981; Taylorand Spicer, 1991). Spicer et al. (1988) found that respira-tory acidosis in P. miliaris induced by emersion over 24 hwas successfully buffered by increases in [HCO3
�], thusallowing acid–base balance to be maintained. This presentstudy was carried out over a longer time frame and here theurchins’ attempts to compensate using bicarbonate buffer-ing were not as complete, as shown by significant reductionin coelomic fluid pH over 8 days. This finding is in line withwork by Portner et al. (2004) who found that in response toacidosis, despite increases in [HCO3
�], acid–base parame-ters and ionic concentrations in marine invertebratesreached new steady state values rather than returning tothe pre-acidosis levels. Thus in the case of P. miliaris itseems that during long term hypercapnia the partially com-pensated pH values cannot be maintained. We suggest thismay be attributed to several echinoid characteristics relatedto the group’s ability to regulate acid–base balance. Firstly,echinoids have a low concentration of protein in the peri-visceral fluid. In many animal groups often the dominantprotein in the blood or haemolymph is a respiratory pig-ment. As far as we know no echinoid has a respiratory pig-ment. Of the echinoderms only the holothurians have beenshown to possess haemoglobin. Whilst the pigment echino-chrome may be present in echinoid eleocyte cells, thispigment contains no metal ions and has no respiratoryfunction (Farmanfarmaian, 1966). The absence of respira-tory protein in P. miliaris results in the species having a lowcapacity for CO2 storage (i.e. very similar to that of seawa-ter) within the coelomic fluid (Spicer et al., 1988). Further-more, echinoids reliance on a water vascular system andpodia unspecialised for respiratory gas exchange mayexplain the accumulation of respiratory CO2 and the con-comitant depressed pH in the coelomic fluid which remainsconstantly lower than that of seawater by around 0.5 units(Farmanfarmaian, 1966).
In a number of aquatic organisms, such as fish and crus-taceans, the primary method of acid–base regulation isionic exchange, i.e. reliance on H+/Na+ and HCO3
�/Cl�
transport (Portner et al., 2004). There should therefore bea positive relationship between a species’ capacity for ionicregulation and acid–base compensation. As an osmo-con-former, the ionic composition of sea urchin coelomic fluidvaries with that of the surrounding seawater (Farmanfar-maian, 1966; Johansen and Vadas, 1967) and as such it isthought these organisms are also poor acid–base compen-sators. This is supported by Burnett et al. (2002) in whichthe purple sea urchin Strongylocentrotus purpuratus under-went short term emersion and no mechanisms for compen-sation were employed in response to acidosis.
This study has shown that P. miliaris responds to aci-dosis by increasing levels of bicarbonate in the coelomicfluid, however, the source of this buffering ion is as yetunknown. Lindinger et al. (1984) report that 70–80% ofHCO3
� released by the mussel M. edulis in response tohypercapnia was released from the CaCO3 shell, via pas-sive dissolution. This mechanism is also suggested forechinoids during emersion (Spicer et al., 1988), terrestrialcrabs (Wheatly and Henry, 1992) and water breathingmolluscs during periods when the shell is closed (Henryet al., 1981), i.e. when internal fluids are cut off from theseawater ion pool.
Psammechinus miliaris shows some evidence of test dis-solution during hypercapnic exposure with significantincreases in [Mg2+] in all exposure treatments; most nota-bly at pHw 6.63 and pHw 7.44 treatments towards theend of the exposure period. The greatest elevation of[Mg2+] was seen at pHw 7.44 with coelomic fluid concentra-tions rising to 69.0 mmol l�1 ± 1.5 compared to typicalfield values of around 53 mmol l�1. This contrasted withthe response of P. miliaris to emersion where [Mg2+] roseto 66.7 mmol l�1 ± 10.7 over 24 h (Spicer et al., 1988).
The apparent difference in the use of the test as a sourceof HCO3
� between P. miliaris individuals subjected toemersion and those exposed to elevated aquatic pCO2
and reduced pH may be due to the availability of the sea-water ion pool. During emersion sea urchins may be forcedto utilise the magnesium calcite test as a buffer sourcealmost immediately whereas urchins immersed in acidifiedwater have access to the available ion pool containingHCO3
� which are exchanged and used in acid–base com-pensation. It may be that the increases in [Mg2+] and[Ca2+] are simply passive dissolutions of the magnesiumcalcite test caused by increased acidity in the coelomic flu-ids. As HCO3
� buffering begins at day 2 (Fig. 4) and testdissolution only releases ions at days 7 and 8 (Fig. 3), wesuggest that the test contributes little to the attempted com-pensation before this time. In conclusion, the intertidal seaurchin P. miliaris was unable to tolerate long term expo-sure to pHw 6.16 seawater. Significant mortality occurs atpHw 6.63 after 8 days and results suggest that at pHw
7.44 coelomic fluid pH and pCO2 values will continue todiverge from controls over longer term exposure. This isdespite the large natural daily fluctuations in both pHand pCO2 experienced by P. miliaris in rock pools (Morrisand Taylor, 1983; Spicer, 1995). This study therefore agreeswith Ormerod and Angel (1996) that a surface ocean pH ofbelow 7.5 should be avoided to prevent acid–base distur-bance, test dissolution and reduced survival of sea urchinadults.
We suggest here that if IPCC predictions for 2100 CO2
emissions are realised (800 ppm), there could be severeeffects on echinoids. Sessile organisms without respiratorypigments and poor ionic regulation mechanisms will bemost vulnerable to chronic exposure to hypercapnia. Mag-nesium calcite dependent species will be at particular riskfrom structural dissolution and reduced calcification.

96 H. Miles et al. / Marine Pollution Bulletin 54 (2007) 89–96
Acknowledgements
We thank Angela Raffo, Julie Soane, Roger Haslam,Fred Staff and Hazel Needham for help and technical sup-port. This study was part-funded by a joint DEFRA/DTIfunded project IMCO2 (http://www.dti.gov.uk/energy/coal/cfft/cct/pub) and a NERC standardgrant (NE/C510016/1) awarded to SW.
References
Bedford, A.P., Moore, P.G., 1985. Macrofaunal involvement in thesublittoral decay of kelp debris – the sea urchin Psammechinus miliaris
(Gmelin) (Echinodermata, Echinoidea). Estuarine Coastal and ShelfScience 20, 19–40.
Blackford, J.C., Gilbert, F., 2007. pH variability and CO2 inducedacidification in the North Sea. Journal of Marine Systems 64, in press,doi:10.1016/j.jmarsys.2006.03.016.
Burnett, L.E., Terwilliger, C.A., Jorgensen, D., Scholnick, D., 2002.Respiratory and acid–base physiology of the purple sea urchin,Strongylocentrotus purpuratus, during air exposure: presence andfunction of a facultative lung. Biological Bulletin of the MarineBiological Laboratory, Woods Hole (Massachusetts) 203, 42–50.
Davenport, H.W., 1974. The ABC of Acid–base Chemistry. University ofChicago Press, Chicago.
DeFur, P.L., McMahon, B.R., 1984. Physiological compensation to shortterm air exposure in red rock crabs Cancer productus from littoral andsublittoral habitats: Acid–base balance. Physiological Zoology 57,151–160.
Farmanfarmaian, A., 1966. The respiratory physiology of echinoderms.In: Boolootian, R.A. (Ed.), Physiology of Echinodermata, 10. Wiley,New York, USA (Chapter 10).
Feely, R.A., Sabine, C.L., Lee, K., Berelson, W., Klepas, J., Fabry, V.J.,Millero, F.J., 2004. Impact of anthropogenic CO2 on the CaCO3
system in the oceans. Science 305, 362–366.Gibbins, J., Haszeldine. S., Holloway, S., Pearce, J., Oakey, J,, Shackley,
S., Turley, C.. 2005. Scope for future CO2 emission reductions fromelectricity generation through the deployment of carbon capture andstorage technologies. In: Proceedings of the Avoiding DangerousClimate Change Conference, Exeter, 1–3 February 2005.
Guillou, M., Grall, J., Connan, S., 2002. Can low sea urchin densitiescontrol macro-epiphytic biomass in a north-east Atlantic maerl bedecosystem (Bay of Brest, Brittany, France)? Journal of the MarineBiological Association of the United Kingdom 82, 867–876.
Haugan, P.M., Drange, H., 1996. Effects of CO2 on the ocean environ-ment. Energy Conversion and Management 37, 1019–1022.
Hawkins, D.G., 2004. No exit: Thinking about leakage from geologiccarbon storage sites. Energy 29, 1571–1578.
Henry, R.P., Kormanik, G.A., Smatresk, N.J., Cameron, J.N., 1981. Therole of CaCO3 dissolution as a source of [HCO3
�] for the buffering ofhypercapnic acidosis in aquatic and terrestrial decapod crustaceans.Journal of Experimental Biology 94, 269–274.
Holloway, S., 2005. Underground sequestration of carbon dioxide – aviable greenhouse gas mitigation option. Energy 30, 2318–2333.
Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Linden, P.J.,Dai, X., Maskell, K., Johnson, C.A., 2001. Climate change 2001: Thescientific basis. In: IPCC Third assessment report: Climate Change2001. Cambridge University Press, Cambridge, p. 944.
Johansen, K., Vadas, R.L., 1967. Oxygen uptake and responses torespiratory stress in sea urchins. Biological Bulletin of the MarineBiological Laboratory, Woods Hole (Massachusetts) 132, 16–22.
Kurihara, H., Shirayama, Y., 2004. Effects of increased atmospheric CO2
on sea urchin early development. Marine Ecology Progress Series 274,161–169.
Kurihara, H., Shimode, S., Shirayama, Y., 2004. Effects of raised CO2
concentration on the egg production rate and early development of
two marine copepods (Acartia steueri and A. erythraea). MarinePollution Bulletin 49, 721–727.
Langenbuch, M., Portner, H.O., 2004. High sensitivity to chronicallyelevated CO2 levels in a eurybathic marine sipunculid. AquaticToxicology 70, 55–61.
Lindinger, M.I., Lawren, D.J., McDonald, D.G., 1984. Acid–base balancein the sea mussel Mytilus edulis. Effects of environmental hypercapniaon intra and extracellular acid–base balance. Marine Biology Letters 5,371–381.
Michaelidis, B., Ouzounis, C., Paleras, A., Portner, H.O., 2005. Effects oflong-term moderate hypercapnia on acid–base balance and growthrate in marine mussels Mytilus galloprovincialis. Marine EcologyProgress Series 293, 109–118.
Morris, S., Taylor, A.C., 1983. Diurnal and seasonal variation in physico-chemical conditions within intertidal rock pools. Estuarine, Coastaland Shelf Sciences 17, 339–355.
Ormerod, B., Angel, M.V., 1996. Ocean storage of carbon dioxide. In:Ormerod, B., Angel, M.V. (Eds.), Workshop 2: Environmental impact.International Energy Agency, Greenhouse Gas R&D Program,Cheltenham, pp. 9–40.
Pearson, P.N., Palmer, M.R., 2000. Atmospheric carbon dioxide concen-trations over the past 60 million years. Nature 406, 695–699.
Politi, Y., Arad, T., Klein, E., Weiner, S., Addadi, L., 2004. Sea urchinspine calcite forms via a transient amorphous calcium carbonate phase.Science 306, 1161–1164.
Portner, H.O., Langenbuch, M., Reipschlager, A., 2004. Biological impactof elevated ocean CO2 concentrations: lessons from animal physiologyand earth history. Journal of Oceanography 60, 705–718.
Raven, J., Caldeira, K., Elderfield, H., Hoegh-Guldberg, O., Liss, P.,Riebesell, U., Shepherd, J., Turley, C., Watson, A., 2005. Oceanacidification due to increasing atmospheric carbon dioxide. TheRoyal Society policy document 12/05. The Clyvedon Press Ltd.,Cardiff, UK.
Riebesell, U., Zondervan, I., Rost, B., Tortell, P.D., Zeebe, R.E., Morel,F.M.M., 2000. Reduced calcification of marine plankton in response toincreased atmospheric CO2. Nature 407, 364–367.
Ross, K.A., Thorpe, J.P., Brand, A.R., 2004. Biological control of foulingin suspended scallop cultivation. Aquaculture 229, 99–116.
Seibel, B.A., Fabry, V.J., 2003. Marine biotic response to elevated carbondioxide. Advances in Applied Biodiversity Science 4, 59–67.
Spicer, J.I., 1995. Oxygen and acid–base status of the sea urchinPsammechinus miliaris during environmental hypoxia. Marine Biology124, 71–76.
Spicer, J.I., Taylor, A.C., Hill, A.D., 1988. Acid–base status in the seaurchins Psammechinus miliaris and Echinus esculentus (Echinodermata:Echinoidea) during emersion. Marine Biology 99, 527–534.
Spicer, J.I., Raffo, A., Widdicombe, S., in press. Influence of CO2-relatedseawater acidification on extracellular acid-base balance in the velvetswimming crab Necora puber. Marine Biology.
Sundquist, E.T., 1993. The global carbon dioxide budget. Science 259,934–941.
Taylor, A.C., Spicer, J.I., 1991. Acid base disturbances in the haemolymphof the prawns, Palaemon elegans (Rathke) and P. serratus (Pennant)(Crustacea: Decapoda) during exposure to hypoxia. ComparativeBiochemistry and Physiology A 98, 445–452.
Truchot, J.P., 1976. Carbon dioxide combining properties of the blood ofthe shore crab Carcinus maenas (L.): Carbon dioxide solubilitycoefficient and carbonic acid dissociation constants. Journal ofExperimental Biology 64, 45–57.
Turley, C., Blackford, J.C., Widdicombe, S., Lowe, D., Nightingale, P.D.,Rees, A.P., 2006. Reviewing the impact of increased atmospheric CO2
on oceanic pH and the marine ecosystem. In: Schellnhuber, H.J.,Cramer, W., Nakicenovic, N., Wigley, T., Yohe, G. (Eds.), AvoidingDangerous Climate Change, 8. Cambridge University Press, Cam-bridge, pp. 65–70.
Wheatly, M.G., Henry, R.P., 1992. Extracellular and intracellular acid–base regulation in crustaceans. Journal of Experimental Zoology 263,127–142.
Related Documents











