HORTICULTURAL ENTOMOLOGY Effect of Root Feeding by Diaprepes abbreviatus (Coleoptera: Curculionidae) Larvae on Leaf Gas Exchange and Growth of Three Ornamental Tree Species ALEX P. DIAZ, CATHARINE MANNION, AND BRUCE SCHAFFER Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Homestead, FL 33031 J. Econ. Entomol. 99(3): 811Ð821 (2006) ABSTRACT Diaprepes abbreviatus L. (Coleoptera: Curculionidae), feeds on a variety of ornamental plants grown in southern Florida. Studies were conducted to evaluate the effects of root feeding by D. abbreviatus larvae on leaf gas exchange and growth of three ornamental tree species commonly grown in southern Florida that are known hosts of this weevil: green buttonwood, Conocarpus erectus L.; live oak, Quercus virginiana Mill.; and pygmy date palm, Phoenix roebelenii OÕBrien. These hosts were grown in containers and infested with weevil larvae. Net CO 2 assimilation, transpiration, and stomatal conductance of CO 2 were measured monthly. Leaf, stem, and root fresh and dry weights of each species also were determined. In one of two tests, larval root feeding signiÞcantly reduced net CO 2 assimilation, transpiration, and stomatal conductance of CO 2 of infested green buttonwood trees. Leaf gas exchange of live oak was not affected by larval infestation. In addition to testing cumulative effects of multiple infestations of larvae, the effects of incremental infestations on leaf gas exchange and fresh and dry weights also were tested for each plant species. Net CO 2 assimilation, transpiration, stomatal conductance of CO 2 , and dry weights of green buttonwood were reduced as a result of larval root feeding, whereas there was no effect of incremental larval infestations on leaf gas exchange of live oak or pygmy date palm within the experimental time frame. There was no effect of incremental larval infestations on dry weights of live oak, but leaf, stem, and dry root weight of pygmy date palm were lower for infested plants than for noninfested plants. Overall, green buttonwood was more susceptible to larval root feeding damage than either live oak or pygmy date palm. KEY WORDS Diaprepes abbreviatus, net CO 2 assimilation, transpiration, stomatal conductance Diaprepes abbreviatus L. (Coleoptera: Curculionidae) is a polyphagous weevil species native to the Carib- bean. It was Þrst reported in the United States in 1964 in a nursery in Apopka, FL. The weevil is thought to have been introduced into Florida in an ornamental plant shipment from a nursery in Puerto Rico (Wood- ruff 1964). D. abbreviatus has become a serious pest and is currently plaguing citrus orchards and many ornamental plant species throughout most of central and southern Florida. In Florida, it is estimated that this weevil causes $70 million in damage annually (Weissling et al. 2002, LaPointe 2000). Larvae of D. abbreviatus cause extensive damage to the root systems of host plants. Larvae feed on all underground portions of the plant, including roots and tubers. Neonate larvae enter the soil, feed on Þbrous roots, and then begin feeding on larger lateral roots as they mature. Larvae cause damage by feeding their way through the outer bark and cambium layer (Futch and McCoy 1993). Large root weevil larvae have been known to girdle roots and cause tree death by either continuous root feeding or by allowing entry for root diseases associated with certain root fungi that destroy crops (McCoy et al. 2003). D. abbreviatus has a host range of nearly 293 plant species that are known to support either adult or larval stages and in many cases both (Simpson et al. 2000). Several native and ornamental plant species in Florida are known to be hosts of D. abbreviatus, and both native and nonnative ornamental plant species sup- port larval development (Schroeder et al. 1979). This weevil is currently injuring many ornamental nursery plants in southern Florida, but the amount of damage to nursery crops has not been quantiÞed. It has been reported that many ornamental trees can sustain a large amount of larval feeding before leaf yellowing and other symptoms occur (Knapp et al. 2000). There are many economically important ornamen- tal plant species grown in southern Florida that are affected by this weevil. Three ornamental plant spe- ciesÑpygmy date palm, Phoenix roebelenii OÕBrien; green buttonwood, Conocarpus erectus L.; and live oak, Quercus virginiana Mill.Ñare commonly grown to- gether in southern Florida Þeld nurseries. These plant species are known to support larval and adult stages of D. abbreviatus and green buttonwood has been shown to be a preferred host (Mannion et al. 2003). Serious root injury has been observed on Þeld-grown live oaks, 0022-0493/06/0811Ð0821$04.00/0 2006 Entomological Society of America

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

HORTICULTURAL ENTOMOLOGY
Effect of Root Feeding by Diaprepes abbreviatus(Coleoptera: Curculionidae) Larvae on Leaf Gas Exchange and
Growth of Three Ornamental Tree Species
ALEX P. DIAZ, CATHARINE MANNION, AND BRUCE SCHAFFER
Tropical Research and Education Center, University of Florida, 18905 SW 280 Street, Homestead, FL 33031
J. Econ. Entomol. 99(3): 811Ð821 (2006)
ABSTRACT Diaprepes abbreviatusL. (Coleoptera: Curculionidae), feeds on a variety of ornamentalplants grown in southern Florida. Studies were conducted to evaluate the effects of root feeding byD. abbreviatus larvae on leaf gas exchange and growth of three ornamental tree species commonlygrown in southern Florida that are known hosts of this weevil: green buttonwood, Conocarpus erectusL.; live oak, Quercus virginiana Mill.; and pygmy date palm, Phoenix roebelenii OÕBrien. These hostswere grown in containers and infested with weevil larvae. Net CO2 assimilation, transpiration, andstomatal conductance of CO2 were measured monthly. Leaf, stem, and root fresh and dry weights ofeach species also were determined. In one of two tests, larval root feeding signiÞcantly reduced netCO2 assimilation, transpiration, and stomatal conductance of CO2 of infested green buttonwood trees.Leaf gas exchange of live oak was not affected by larval infestation. In addition to testing cumulativeeffects of multiple infestations of larvae, the effects of incremental infestations on leaf gas exchangeand fresh and dry weights also were tested for each plant species. Net CO2 assimilation, transpiration,stomatal conductance of CO2, and dry weights of green buttonwood were reduced as a result of larvalroot feeding, whereas there was no effect of incremental larval infestations on leaf gas exchange oflive oak or pygmy date palm within the experimental time frame. There was no effect of incrementallarval infestations on dry weights of live oak, but leaf, stem, and dry root weight of pygmy date palmwere lower for infested plants than for noninfested plants. Overall, green buttonwood was moresusceptible to larval root feeding damage than either live oak or pygmy date palm.
KEY WORDS Diaprepes abbreviatus, net CO2 assimilation, transpiration, stomatal conductance
Diaprepes abbreviatusL. (Coleoptera: Curculionidae)is a polyphagous weevil species native to the Carib-bean. It was Þrst reported in the United States in 1964in a nursery in Apopka, FL. The weevil is thought tohave been introduced into Florida in an ornamentalplant shipment from a nursery in Puerto Rico (Wood-ruff 1964). D. abbreviatus has become a serious pestand is currently plaguing citrus orchards and manyornamental plant species throughout most of centraland southern Florida. In Florida, it is estimated thatthis weevil causes �$70 million in damage annually(Weissling et al. 2002, LaPointe 2000).
Larvae ofD. abbreviatus cause extensive damage tothe root systems of host plants. Larvae feed on allunderground portions of the plant, including roots andtubers. Neonate larvae enter the soil, feed on Þbrousroots, and then begin feeding on larger lateral roots asthey mature. Larvae cause damage by feeding theirway through the outer bark and cambium layer (Futchand McCoy 1993). Large root weevil larvae have beenknown to girdle roots and cause tree death by eithercontinuous root feeding or by allowing entry for rootdiseases associated with certain root fungi that destroycrops (McCoy et al. 2003).
D. abbreviatus has a host range of nearly 293 plantspecies that are known to support either adult or larvalstages and in many cases both (Simpson et al. 2000).Several native and ornamental plant species in Floridaare known to be hosts of D. abbreviatus, and bothnative and nonnative ornamental plant species sup-port larval development (Schroeder et al. 1979). Thisweevil is currently injuring many ornamental nurseryplants in southern Florida, but the amount of damageto nursery crops has not been quantiÞed. It has beenreported that many ornamental trees can sustain alarge amount of larval feeding before leaf yellowingand other symptoms occur (Knapp et al. 2000).
There are many economically important ornamen-tal plant species grown in southern Florida that areaffected by this weevil. Three ornamental plant spe-ciesÑpygmy date palm, Phoenix roebelenii OÕBrien;green buttonwood,Conocarpus erectusL.; and live oak,Quercus virginiana Mill.Ñare commonly grown to-gether in southern Florida Þeld nurseries. These plantspecies are known to support larval and adult stages ofD. abbreviatus and green buttonwood has been shownto be a preferred host (Mannion et al. 2003). Seriousroot injury has been observed on Þeld-grown live oaks,
0022-0493/06/0811Ð0821$04.00/0 � 2006 Entomological Society of America

and adults and larvae have been found on all threespecies in several Þeld nurseries in Homestead, FL(C.M., personal observation).
A basic understanding of the physiological re-sponses of plants to herbivory may provide criticalinformation for predicting and preventing crop dam-age (Welter 1989). Measurements of leaf gas exchangeprovide information about the effects of insect feedingon plant physiology and stress often before the ap-pearance of any visual symptoms of stress occur(Welter 1989, Peterson et al. 1992, Peterson et al.1998). The majority of studies of plant responses toinsect herbivory focused on adult insects feeding onleaves (Johnson et al. 1983, Li and Proctor 1984, Peter-son et al. 1998, Klingeman et al. 2000), includingD. abbreviatus (Syvertsen and McCoy 1985). Therehave been a few studies reported on the effect of rootdamage from insect larvae on leaf gas exchange (God-frey et al. 1993, Riedell and Reese 1999), and there areno known reports on the effects ofD. abbreviatus rootherbivory on leaf gas exchange.
The objective of this study was to evaluate theeffects of root feeding by larvae of D. abbreviatus onnet gas exchange and growth of pygmy date palm,green buttonwood, and live oak.
Materials and Methods
Studies were conducted from October 2003 throughJuly 2004 at the University of Florida, Tropical Re-search and Education Center, Homestead, FL (25.5� N,85.5� W). Two separate experiments were conductedto determine the effects of 1) root feeding by larvaeof two different ages on leaf gas exchange and plantgrowth, and 2) root feeding damage by multiple larvalinfestations on leaf gas exchange and plant growth.The Þrst experiment was conducted on green button-wood from October 2003 to March 2004 and on liveoak from December 2003 to May 2004. The secondexperiment was conducted on green buttonwood,live oak, and pygmy date palm from January toJuly 2004.
Plant and Insect Material
Experiment 1.Green buttonwood in 3.79-liter con-tainers were purchased from a commercial nursery(Princeton Nurseries II, Inc., Homestead, FL) andtransplanted into 11.35-liter containers with welldrained media consisting of 40% Florida peat, 30% pinebark, 10% sand, 20% cypress sawdust, and 6.80 kgdolomite/91.44 cm purchased from a local soil distrib-utor (Lantana Peat & Soil, Boynton Beach, FL). Liveoaks in 3.79-liter containers were purchased from acommercial nursery (Albert Livingston Tree Farms,Homestead, FL) and transplanted into 11.35-liter con-tainers in the same media as described for green but-tonwood trees.Experiment 2.Young live oak and pigmy date palms
in 3.79-liter containers were purchased from a com-mercial nursery (Bill Ingram & Grandsons NSY,Homestead, FL). Green buttonwood trees in 3.79-liter
containers were purchased from another nursery(Princeton Nurseries II, Inc.). Plants were selectedbased on relative uniformity of shoot and canopy sizewithin each species. All plants were transplanted into11.35-liter containers in the same media as describedfor experiment 1.
All plants in both experiments were fertilized oncebefore treatment initiation with PlantacotePlus14-9-15 (NÐPÐK) (Helena Chemical Company, Col-lierville, TN) controlled release fertilizer (32.5 g percontainer). Plants were irrigated twice daily, at 5 a.m.and at 5 p.m. for 30 min each time. Larvae ofD. abbreviatus were obtained from the Florida Divi-sion of Plant Industry (Division of Agriculture andConsumer Services, Gainesville, FL).
Treatments
Experiment 1.Larvae were sorted according to sizeso that each treatment would have similar aged in-stars. The larvae were Þrst separated into medium(1Ð1.5-cm) or large (2-cm) size classes, measuredfrom the head to the tip of the abdomen. The headcapsules from 20 randomly selected larvae from eachsize class were measured, and average instars per sizeclass were determined as described by Quintela et al.(1998). The medium size class contained Þfth to sixthinstars, and the large size class contained eighth in-stars.
Ten holes were made in the soil surface around eachplant with small spatulas, 4Ð8 cm from the stem, with2.54-cm spacing between holes. The depth of the holesalternated at 5 and 10 cm from the soil surface. Twolarvae were placed into each hole by placing one larvacovering it with soil, then placing the second larva andcovering it with soil. Each container was infested with20 larvae. All containers were irrigated after infesta-tion to cool the soil and keep larvae from being af-fected by high soil temperatures.
There were three treatments with 16 replicationsper treatment: 1) noninfested controls, 2) infestedwith medium larvae, and 3) infested with large larvae.Three months after infestation, eight replications ofeach treatment were harvested to determine whetherlarvae were surviving and feeding on the roots. Theremaining eight plants in each treatment were rein-fested two times at 2-mo intervals with 20 larvae percontainer each time. Larvae were selected and placedinto containers as described previously for the Þrstinfestation. All remaining plants were harvested 6 moafter the initial infestation.Leaf Gas Exchange. Net CO2 assimilation, stomatal
conductanceofCO2, and transpirationweremeasuredmonthly with CIRAS-2 portable gas analyzer (PP Sys-tems, Hitchin, Hetfordshire, United Kingdom) be-tween 10 a.m. and noon. Leaf gas exchange was mea-sured at a photosynthetic photon ßux �900 �mol m�2
s�1 with a halogen lamp Þtted on the leaf cuvette asthe light source. Leaf gas exchange was determinedfor two fully expanded leaves of each plant, and theaverage of the two leaves was used to represent gasexchange values for each single-plant replication.
812 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 99, no. 3
Root Biomass and Larval Recovery. The roots werecarefully separated from the rooting media and rinsedwith tap water to remove excess soil. Soil from theroots was placed into bins and carefully inspected forlarvae. Recovered larvae were separated and placedinto holding containers containing 75% ethanol. Ex-cess water was allowed to drain from roots for 12 h, androot fresh weights were determined. Roots were oven-dried at 70�C to a constant weight, and root dryweights were determined.Experiment 2. In this experiment, Þfth to sixth in-
stars (determined by head capsule size as describedpreviously) were used to infest plants. Thirty-twoplants each of green buttonwood, live oak and pygmydate palm were infested with 20 larvae per containeras described for experiment 1 and 32 plants were leftnoninfested as controls. Two months after infestation,a set of eight infested plants and eight noninfestedplants of each species were harvested. Thereafter,eight plants of each treatment were harvestedmonthly. After each harvest, all remaining plants inthe infested treatment were reinfested with 20 larvaeper container as described for experiment 1. Thus, forthe infested treatment, the Þrst set of plants harvestedwas infested with a total of 20 larvae, plants harvestedafter 3 mo were infested with a total of 40 larvae (20per month), plants harvested after 4 mo were infested
with a total of 60 larvae, and plants harvested after 5mo were infested with a total of 80 larvae.Leaf Gas Exchange. Leaf gas exchange was mea-
sured monthly before each harvest for each set ofplants as described for experiment 1. For the set ofplants in the Þnal harvest, leaf gas exchange was mea-sured on the week of infestation and monthly there-after until the week of harvest.PlantGrowth andLarvalRecovery. Plant height and
trunk diameter were measured just before treatmentinitiation and again before the Þnal harvest. Plantheight was measured from the base of the stem to thetip of the apical bud by using a measuring tape. For oakand green buttonwood trunk diameter was measuredat 10 cm above the soil surface with an electronicdigital caliper (Fisher, Pittsburgh, PA). Leaves, stems,and roots were harvested monthly beginning at theend of the second month, and the number of recov-ered larvae and fresh and dry weights of plant tissueswere determined as described for experiment 1. Theratio of root dry weight to root fresh weight wasdetermined by dividing the difference between thefresh and dry root weights by the fresh root weight.Root dry weight/fresh weight ratios were comparedbetween treatments at each harvest to determinewhether there was a relationship between root suc-culence (ratio) and treatment.
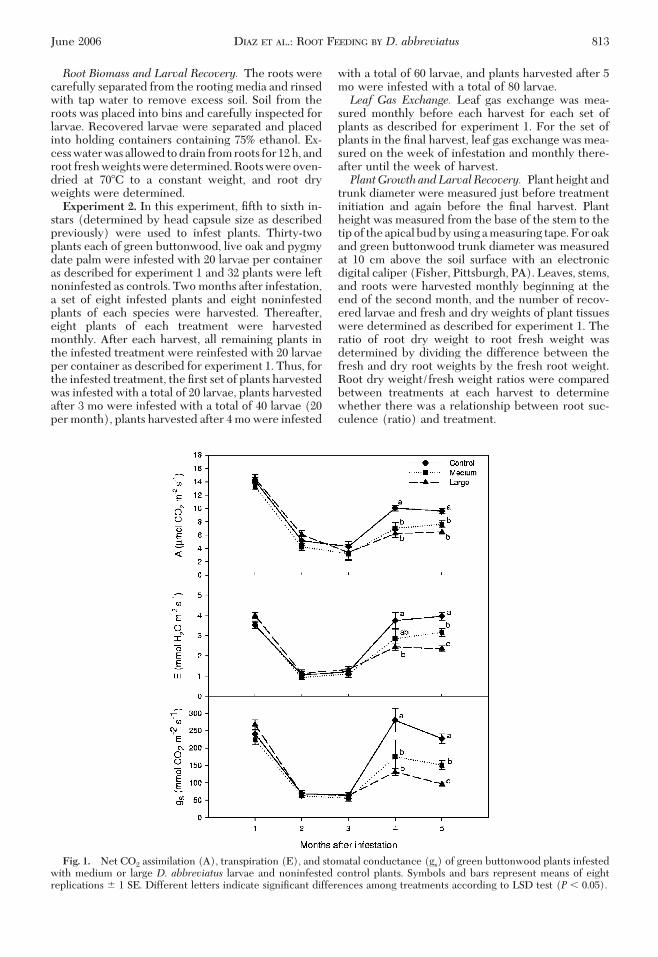
Fig. 1. Net CO2 assimilation (A), transpiration (E), and stomatal conductance (gs) of green buttonwood plants infestedwith medium or large D. abbreviatus larvae and noninfested control plants. Symbols and bars represent means of eightreplications � 1 SE. Different letters indicate signiÞcant differences among treatments according to LSD test (P � 0.05).
June 2006 DIAZ ET AL.: ROOT FEEDING BY D. abbreviatus 813
Temperature Measurements
Air and soil temperatures were measured withHOBO StowAway data loggers (Onset ComputerCorp., Pocasset, MA). Data loggers used for soil mea-surements were placed at a depth from the soil surfaceof �10 cm in eight 11.35-liter containers with no plantsin them and the same media as the containers withplants. Air temperature was measured with a datalogger placed under a bench next to the experimentalplants. Mean daily temperatures were recorded (datanot shown).
Statistical Analysis
Data were analyzed by a standard t-test or analysisof variance, and means were compared with a test forleast signiÞcant difference (LSD) using SAS software(SAS Institute, Cary, NC).
Results
Experiment 1. Leaf Gas Exchange. Leaf gas ex-change of green buttonwood and live oak ßuctuatedwith temperature, decreasing with dropping temper-atures and increasing when temperatures increasedfor all treatments. At the beginning of the experimentin October, average monthly temperature was �25�C.
Average air and soil temperatures decreased by theend of November to 18�C. Average air and soil tem-peratures began to increase by February to above 21�C(data not shown).
Larval feeding did not affect leaf gas exchange ofgreen buttonwood until average temperatures in-creased above 21�C. At that temperature, net CO2
assimilation, transpiration, and stomatal conductanceof CO2 for all treatments increased from previouslevels. However, control trees had signiÞcantly highernet CO2 assimilation, transpiration, and stomatal con-ductance of CO2 than plants infested with medium orlarge larvae 4 mo (net CO2 assimilation: F� 8.70; df �2, 23; P� 0.0018; transpiration: F� 3.47; df � 2, 23; P�0.05; and stomatal conductance of CO2: F� 5.03; df �2, 23; P� 0.016) and 5 mo (net CO2 assimilation: F�15.74; df � 2, 23; P � 0.0001; transpiration: F � 20.06;df � 2, 23; P � 0.0001; stomatal conductance of CO2:F� 32.97; df � 2, 23;P� 0.0001) after treatments wereinitiated (Fig. 1).
Leaf gas exchange of live oak ßuctuated slightlywith temperature. Treatments were initiated duringDecember when temperatures were relatively lowand net CO2 assimilation, transpiration, and stomatalconductance of CO2 were also relatively low and thenincreased as average temperature increased. There
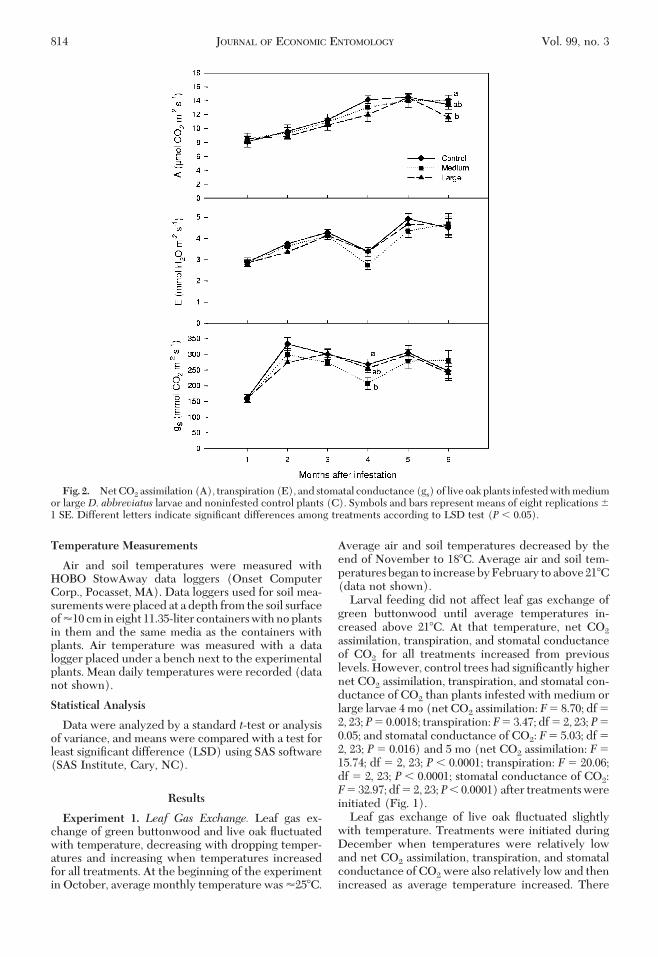
Fig. 2. Net CO2 assimilation (A), transpiration (E), and stomatal conductance (gs) of live oak plants infested with mediumor largeD. abbreviatus larvae and noninfested control plants (C). Symbols and bars represent means of eight replications �1 SE. Different letters indicate signiÞcant differences among treatments according to LSD test (P � 0.05).
814 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 99, no. 3
were no signiÞcant differences in net CO2 assimila-tion, transpiration, and stomatal conductance of CO2
among treatments with a couple of exceptions (Fig. 2).One exception was 4 mo after the initial infestationwhen stomatal conductance of CO2 of the treatmentinfested with medium larvae was signiÞcantly lowerthan that of the control treatment (Fig. 2). There wasno difference in stomatal conductance of CO2 amongtreatments on the last two measurement dates. Thesecond exception was the signiÞcant reduction in netCO2 assimilation of the large larvae treatment com-pared with the control treatment on the Þnal mea-surement date.Root Weights and Larval Recovery. Fresh root
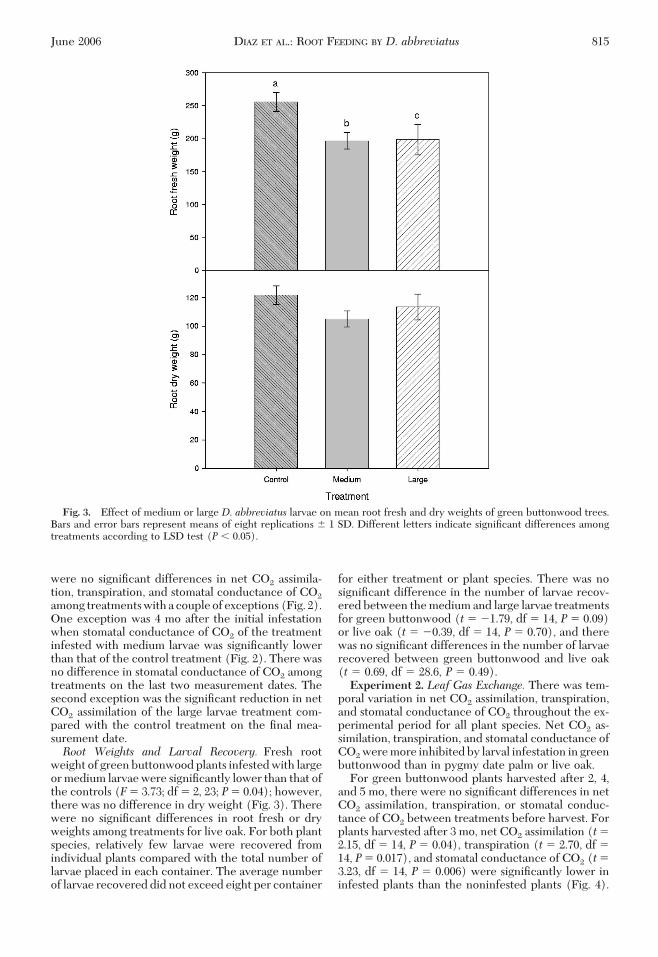
weight of green buttonwood plants infested with largeor medium larvae were signiÞcantly lower than that ofthe controls (F� 3.73; df � 2, 23; P� 0.04); however,there was no difference in dry weight (Fig. 3). Therewere no signiÞcant differences in root fresh or dryweights among treatments for live oak. For both plantspecies, relatively few larvae were recovered fromindividual plants compared with the total number oflarvae placed in each container. The average numberof larvae recovered did not exceed eight per container
for either treatment or plant species. There was nosigniÞcant difference in the number of larvae recov-ered between the medium and large larvae treatmentsfor green buttonwood (t � �1.79, df � 14, P � 0.09)or live oak (t � �0.39, df � 14, P � 0.70), and therewas no signiÞcant differences in the number of larvaerecovered between green buttonwood and live oak(t � 0.69, df � 28.6, P � 0.49).Experiment 2. Leaf Gas Exchange. There was tem-
poral variation in net CO2 assimilation, transpiration,and stomatal conductance of CO2 throughout the ex-perimental period for all plant species. Net CO2 as-similation, transpiration, and stomatal conductance ofCO2 were more inhibited by larval infestation in greenbuttonwood than in pygmy date palm or live oak.
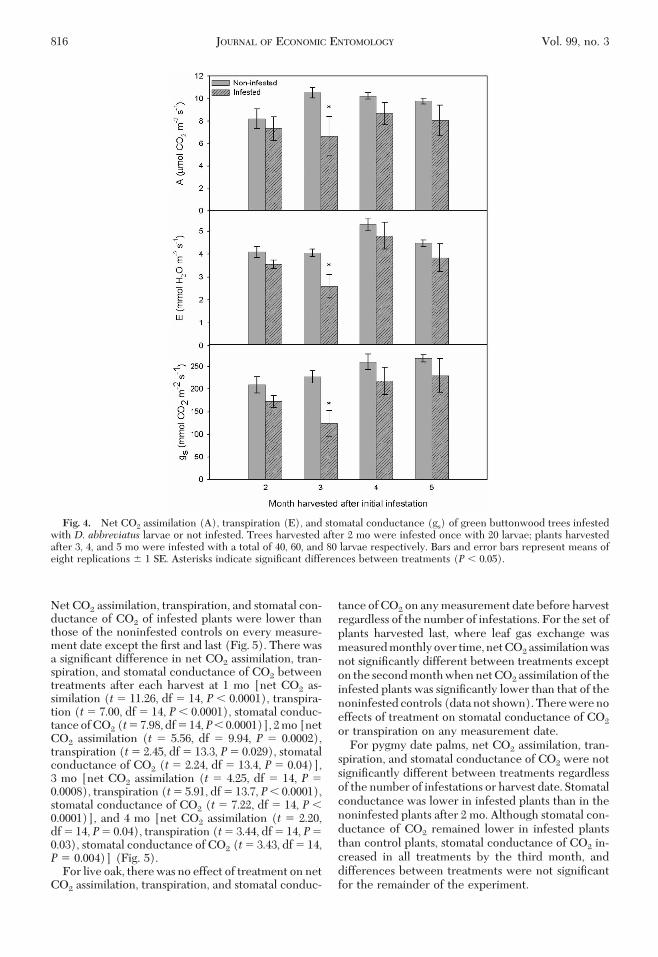
For green buttonwood plants harvested after 2, 4,and 5 mo, there were no signiÞcant differences in netCO2 assimilation, transpiration, or stomatal conduc-tance of CO2 between treatments before harvest. Forplants harvested after 3 mo, net CO2 assimilation (t�2.15, df � 14, P � 0.04), transpiration (t � 2.70, df �14, P� 0.017), and stomatal conductance of CO2 (t�3.23, df � 14, P � 0.006) were signiÞcantly lower ininfested plants than the noninfested plants (Fig. 4).
Fig. 3. Effect of medium or large D. abbreviatus larvae on mean root fresh and dry weights of green buttonwood trees.Bars and error bars represent means of eight replications � 1 SD. Different letters indicate signiÞcant differences amongtreatments according to LSD test (P � 0.05).
June 2006 DIAZ ET AL.: ROOT FEEDING BY D. abbreviatus 815
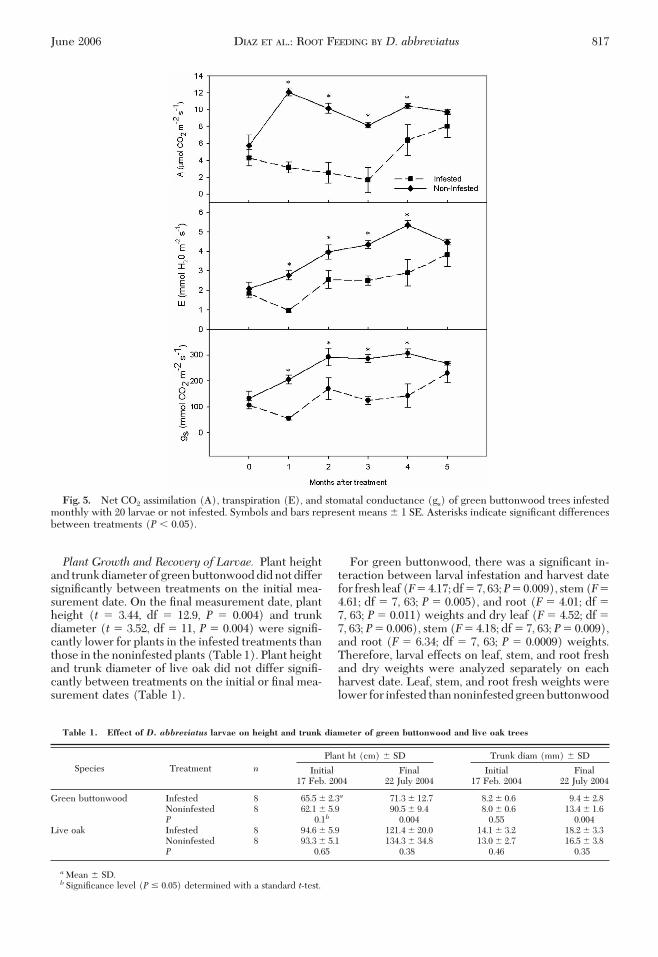
Net CO2 assimilation, transpiration, and stomatal con-ductance of CO2 of infested plants were lower thanthose of the noninfested controls on every measure-ment date except the Þrst and last (Fig. 5). There wasa signiÞcant difference in net CO2 assimilation, tran-spiration, and stomatal conductance of CO2 betweentreatments after each harvest at 1 mo [net CO2 as-similation (t � 11.26, df � 14, P � 0.0001), transpira-tion (t� 7.00, df � 14, P� 0.0001), stomatal conduc-tance of CO2 (t� 7.98, df � 14,P� 0.0001)], 2 mo [netCO2 assimilation (t � 5.56, df � 9.94, P � 0.0002),transpiration (t� 2.45, df � 13.3, P� 0.029), stomatalconductance of CO2 (t � 2.24, df � 13.4, P � 0.04)],3 mo [net CO2 assimilation (t � 4.25, df � 14, P �0.0008), transpiration (t� 5.91, df � 13.7, P� 0.0001),stomatal conductance of CO2 (t � 7.22, df � 14, P �0.0001)], and 4 mo [net CO2 assimilation (t � 2.20,df � 14, P� 0.04), transpiration (t� 3.44, df � 14, P�0.03), stomatal conductance of CO2 (t� 3.43, df � 14,P � 0.004)] (Fig. 5).
For live oak, there was no effect of treatment on netCO2 assimilation, transpiration, and stomatal conduc-
tance of CO2 on any measurement date before harvestregardless of the number of infestations. For the set ofplants harvested last, where leaf gas exchange wasmeasuredmonthlyover time,netCO2 assimilationwasnot signiÞcantly different between treatments excepton the second month when net CO2 assimilation of theinfested plants was signiÞcantly lower than that of thenoninfested controls (data not shown). There were noeffects of treatment on stomatal conductance of CO2
or transpiration on any measurement date.For pygmy date palms, net CO2 assimilation, tran-
spiration, and stomatal conductance of CO2 were notsigniÞcantly different between treatments regardlessof the number of infestations or harvest date. Stomatalconductance was lower in infested plants than in thenoninfested plants after 2 mo. Although stomatal con-ductance of CO2 remained lower in infested plantsthan control plants, stomatal conductance of CO2 in-creased in all treatments by the third month, anddifferences between treatments were not signiÞcantfor the remainder of the experiment.
Fig. 4. Net CO2 assimilation (A), transpiration (E), and stomatal conductance (gs) of green buttonwood trees infestedwith D. abbreviatus larvae or not infested. Trees harvested after 2 mo were infested once with 20 larvae; plants harvestedafter 3, 4, and 5 mo were infested with a total of 40, 60, and 80 larvae respectively. Bars and error bars represent means ofeight replications � 1 SE. Asterisks indicate signiÞcant differences between treatments (P � 0.05).
816 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 99, no. 3
Plant Growth and Recovery of Larvae. Plant heightand trunk diameter of green buttonwood did not differsigniÞcantly between treatments on the initial mea-surement date. On the Þnal measurement date, plantheight (t � 3.44, df � 12.9, P � 0.004) and trunkdiameter (t � 3.52, df � 11, P � 0.004) were signiÞ-cantly lower for plants in the infested treatments thanthose in the noninfested plants (Table 1). Plant heightand trunk diameter of live oak did not differ signiÞ-cantly between treatments on the initial or Þnal mea-surement dates (Table 1).
For green buttonwood, there was a signiÞcant in-teraction between larval infestation and harvest datefor fresh leaf (F� 4.17; df � 7, 63;P� 0.009), stem (F�4.61; df � 7, 63; P � 0.005), and root (F � 4.01; df �7, 63; P � 0.011) weights and dry leaf (F � 4.52; df �7, 63; P� 0.006), stem (F� 4.18; df � 7, 63; P� 0.009),and root (F � 6.34; df � 7, 63; P � 0.0009) weights.Therefore, larval effects on leaf, stem, and root freshand dry weights were analyzed separately on eachharvest date. Leaf, stem, and root fresh weights werelower for infested than noninfested green buttonwood
Fig. 5. Net CO2 assimilation (A), transpiration (E), and stomatal conductance (gs) of green buttonwood trees infestedmonthly with 20 larvae or not infested. Symbols and bars represent means � 1 SE. Asterisks indicate signiÞcant differencesbetween treatments (P � 0.05).
Table 1. Effect of D. abbreviatus larvae on height and trunk diameter of green buttonwood and live oak trees
Species Treatment n
Plant ht (cm) � SD Trunk diam (mm) � SD
Initial17 Feb. 2004
Final22 July 2004
Initial17 Feb. 2004
Final22 July 2004
Green buttonwood Infested 8 65.5 � 2.3a 71.3 � 12.7 8.2 � 0.6 9.4 � 2.8Noninfested 8 62.1 � 5.9 90.5 � 9.4 8.0 � 0.6 13.4 � 1.6P 0.1b 0.004 0.55 0.004
Live oak Infested 8 94.6 � 5.9 121.4 � 20.0 14.1 � 3.2 18.2 � 3.3Noninfested 8 93.3 � 5.1 134.3 � 34.8 13.0 � 2.7 16.5 � 3.8P 0.65 0.38 0.46 0.35
aMean � SD.b SigniÞcance level (P � 0.05) determined with a standard t-test.
June 2006 DIAZ ET AL.: ROOT FEEDING BY D. abbreviatus 817

plants at each harvest date except the Þrst when therewere no differences in leaf and stem weights betweentreatments (Table 2). Leaf dry weight of green but-tonwood was lower for infested than noninfestedplants harvested after 3 or 5 mo. Stem dry weightswere not signiÞcantly different between treatmentsfor plants that were harvested after 2 mo. However,stem dry weight of infested plants was signiÞcantlylower than that of the control plants harvested after 3,4, or 5 mo (Table 2). Root dry weights on all fourharvestdateswere signiÞcantly lower for infested thancontrol plants (Table 2). The root fresh weight/dryweight ratio for green buttonwood did not differ be-tween treatments for the Þnal two harvests (P� 0.05);however, there was a signiÞcantly greater ratio for thecontrol treatment than for the larval infested treat-ment at the Þrst two harvests (P � 0.5; data notshown).
There were no signiÞcant interactions betweentreatment and harvest date for leaf, stem, and rootfresh and dry weights of live oak or pygmy date palm(P � 0.05). Therefore, harvest dates were pooled foreach species to test the effect of larval root feeding onleaf, stem, and root fresh and dry weights. For live oak,there were no signiÞcant effects of treatment on leaf,stem, and root fresh or dry weights (Table 3). Infestedpygmy date palms had signiÞcantly lower leaf, stem,and root fresh and dry weights than noninfested plants(Table 3). The root dry weight/fresh weight ratio forlive oak and pygmy palm did not differ between in-fested and control plants (P � 0.05; data not shown).For both pygmy palm and live oak, the number oflarvae recovered at each harvest was �10.
There was a signiÞcant effect of harvest date on thenumber of larvae recovered from green buttonwood(F� 27.64; df � 3, 31; P� 0.0001), live oak (F� 10.15;df � 3, 31;P� 0.0001), and pygmy date palm (F� 7.79;df � 3, 31; P� 0.0006) (Table 4). There tended to bemore larvae recovered from green buttonwood thanfrom live oak or pygmy date palm; however, the dif-ferences were not signiÞcant until the last harvest datewhen more larvae were recovered from green but-tonwood (F� 7.00; df � 2, 23; P� 0.004) than live oakor pygmy date palm (Table 4).
Discussion
In experiment 1, average monthly temperatures haddropped from �27�C in October to �18�C in Decem-ber, including several days with minimum tempera-tures below 10�C. In green buttonwood, the reductionin net CO2 assimilation, transpiration, and stomatalconductance of CO2 of trees in all treatments 2 moafter infestation was presumably the result of lowtemperature because leaf gas exchange of all treat-ments, including controls was reduced. As the averagemonthly air and soil temperatures increased to �21�Cby the end of February, net CO2 assimilation, tran-spiration, and stomatal conductance of CO2 of alltreatments increased.These results are similar to thoseof Taylor and Rowley 1971 who observed an imme-diate reduction in leaf photosynthesis in sorghum(Sorghum hybrid), maize (Zea mays L.) and pennis-etum (Pennisetum typhoides Burm.) as temperaturesdeclined from 25 to 10�C and then increased as tem-perature returned to 25�C. Reductions in net CO2
Table 3. Effect of D. abbreviatus larvae on leaf stem and root fresh and dry weights of live oak and pygmy date palm
Species Treatment nFresh wt (g) � SD Dry weight (g) � SD
Leaf Stem Root Leaf Stem Root
Live oak Noninfested 32 89.0 � 28.3a 233.4 � 87.1 394.8 � 115.8 45.9 � 16.0 136.8 � 53.4 221.4 � 65.5Infested 32 84.6 � 31.7 245.4 � 84.9 372.8 � 164.6 41.8 � 16.2 143.8 � 49.6 211.2 � 97.9P 0.55b 0.57 0.53 0.31 0.58 0.62
Pygmy palm Noninfested 32 171.0 � 49.1 182.9 � 57.2 322.5 � 151.4 67.9 � 19.0 94.5 � 36.0 113.3 � 43.7Infested 32 148.0 � 49.7 146.2 � 58.3 239.7 � 177.1 58.5 � 19.4 72.9 � 38.2 84.6 � 49.1P 0.06 0.01 0.04 0.05 0.02 0.01
aMean � SD.b SigniÞcance level (P � 0.05) determined with a standard t-test.
Table 2. Effect of D. abbreviatus larvae on green buttonwood leaf, stem, and root fresh and dry weights at each harvest date
Planttissue
Treatment nFresh wt (g) � SD Dry wt (g) � SD
April 2004 May 2004 June 2004 July 2004 April 2004 May 2004 June 2004 July 2004
Leaf Noninfested 8 197.2 � 30.8a 219.2 � 45.4 247.6 � 26.7 254.5 � 41.4 73.5 � 12.0 80.3 � 14.9 99.7 � 9.6 97.8 � 15.7Infested 8 186.3 � 28.9 112.4 � 70.2 188.0 � 91.7 100.3 � 97.8 74.2 � 16.1 43.4 � 25.8 75.9 � 39.1 37.5 � 38.1P 0.47b 0.003 0.09 0.001 0.91 0.004 0.11 0.001
Stem Noninfested 8 86.2 � 8.3 102.5 � 23.3 130.3 � 16.9 142.3 � 27.6 36.0 � 3.5 45.4 � 8.0 66.7 � 12.4 65.3 � 15.7Infested 8 76.7 � 14.1 58.5 � 31.0 103.0 � 28.9 68.8 � 38.9 32.8 � 7.3 28.5 � 13.3 49.1 � 15.0 31.8 � 15.7P 0.13 0.006 0.04 0.001 0.28 0.009 0.02 0.001
Root Noninfested 8 143.6 � 24.8 128.0 � 21.3 184.9 � 34.5 244.0 � 49.5 54.5 � 7.8 65.7 � 8.2 84.2 � 8.7 100.4 � 12.3Infested 8 85.0 � 19.9 40.9 � 23.3 109.2 � 56.9 79.4 � 93.0 37.1 � 5.8 25.5 � 13.7 57.5 � 26.9 33.4 � 32.3P 0.0002 �0.0001 0.007 0.001 0.0002 �0.0001 0.01 �0.0001
a Mean � SD.b SigniÞcance level (P � 0.05) determined with a standard t-test.
818 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 99, no. 3

assimilation also have been observed in tropicalwoody plant species such as avocado (Persea ameri-cana Mill.) and mango (Mangifera indica L.) in re-sponse to low temperatures (Whiley et al. 1999).Green buttonwood is a tropical plant species native tosouthern Florida and its distribution into central andnorthern Florida is limited by low winter tempera-tures (Tomlinson 1980). The signiÞcant difference innet CO2 assimilation, transpiration, and stomatal con-ductance of CO2 between treatments when temper-ature increased was presumably a result of increasedlarval activity and root feeding in the infested treat-ment when root temperature had warmed. LaPointe(2000) found that the growth rate of D. abbreviatuslarvae increased exponentially with temperature from15 to 30�C and the optimal temperature for larvaldevelopment was between 22 and 26�C. Although netCO2 assimilation, transpiration, and stomatal conduc-tance of CO2 were very low during periods of low soiltemperature, the average midday soil temperaturepresumably warmed enough for some larval root feed-ing to occur. There are no other known reports of theeffects of root feeding byD. abbreviatus larvae on netCO2 assimilation, transpiration, and stomatal conduc-tance of CO2. However, western corn root worm,Diabrotica virgifera virgifera LeConte, caused reduc-tions in net CO2 assimilation when larvae were ac-tively feeding on maize root systems but did not affectnet CO2 assimilation when the larvae reached thequiescent pupal stage (Godfrey et al. 1993, Riedell andReese 1999). In experiment 1, green buttonwoodplants infested with large larvae (eighth instar) ex-hibited lower net CO2 assimilation, transpiration, andstomatal conductance of CO2 than those infested withmedium-sized larvae (Þfth to sixth instar). It has beenreported that weight gain is greatest in larvae betweensixth and ninth instars because at these stages larvaerequire large amounts of nutrients and therefore causesigniÞcant injury to plants (Quintela et al. 1998, Rog-ers et al. 2000). After the Þrst infestation in experiment1, soil temperatures below 15�C may have caused somelarval mortality (LaPointe 2000), and some larvae mayhave become quiescent and pupated, thereby reduc-ing the amount of damage caused by root feeding.However, reinfestation of larvae may have allowed thelarger larvae to continue to cause signiÞcantly moredamage.
Net CO2 assimilation, transpiration, and stomatalconductance of CO2 were not as sensitive to larvalinfestation in live oak as in green buttonwood. The oak
treatments were initiated later than the green button-wood when temperature, on average, had increased toabove 21�C. Therefore, the temperature was presum-ably not low enough during the oak study to affect leafgas exchange. Larval infestation did not signiÞcantlyand consistently affect leaf gas exchange of oak duringthe 6-mo infestation period.
For green buttonwood, infested plants had lowerroot fresh and dry weights than the controls. However,the differences were signiÞcant only on fresh rootweights. Others also have reported reductions in rootweight caused by D. abbreviatus larval feeding (Rog-ers et al. 2000, Nigg et al. 2001, Mannion et al. 2003).For live oak, root fresh and dry weights were notsigniÞcantly different from the control trees. Greenbuttonwood root systems seemed to have more dam-age than those of the live oak, which may be due, inpart, to the differences in root system anatomy of bothspecies. The root system of green buttonwood wasmore succulent and Þbrous than that of live oak. Thefast-growing succulent roots of green buttonwoodmay be the reason that the weevil larvae prefer greenbuttonwood as a host over many other ornamentalplant species (Mannion et al. 2003). Live oak had someroot damage in the form of tunneling through the barkand cambium layer on the larger lateral roots and anoticeable reduction in the number of smaller Þbrousfeeder roots. However, the majority of root mass wasin the large primary tap and lateral roots that wererelatively thick and root feeding did not seem to re-duce root weight.
In experiment 2, green buttonwood trees were af-fected by larval root feeding more than pygmy datepalm or live oak. Infestation by D. abbreviatus larvaecaused a reduction in net CO2 assimilation, transpi-ration, and stomatal conductance of CO2 of greenbuttonwood. This is consistent with results from ex-periment 1, in which there were reductions in net CO2
assimilation, transpiration, and stomatal conductanceof CO2 of infested plants. Several green buttonwoodplants were showing signs of leaf yellowing, defolia-tion, and wilting before harvest. These symptoms aretypical of intense weevil damage (Knapp et al. 2000).Live oak responded as they did in experiment 1, in thatthere were no visible signs of damage to the canopyand no signiÞcant reductions in net CO2 assimilation,transpiration, and stomatal conductance of CO2
caused by larval root feeding. For pygmy date palms,there were no visible signs of stress to the canopy andnet CO2 assimilation, transpiration, and stomatal con-
Table 4. Number of D. abbreviatus larvae recovered from plants harvested 2, 3, 4, or 5 mo after infestation
Species nMean no. of larvae (� SD)
2 mo 3 mo 4 mo 5 mo
Green buttonwood 8 8.6 � 1.3aAa 10.0 � 2.0aA 4.3 � 2.2bA 2.8 � 1.8bALive oak 8 7.9 � 3.9aA 9.9 � 4.9aA 3.9 � 3.0bA 1.1 � 0.8bBPygmy palm 8 7.6 � 2.4aA 5.8 � 4.1abA 3.8 � 3.9bA 0.5 � 0.8cB
Total number of larvae infested for Þrst harvest, 20; second harvest, 40; third harvest, 60; and fourth harvest, 80.aMeans within a row followed by different lower case letters and means within a column with different capital letters are signiÞcantly
different according to LSD test (P � 0.05).
June 2006 DIAZ ET AL.: ROOT FEEDING BY D. abbreviatus 819

ductance of CO2 were not signiÞcantly affected bytreatment. Again, green buttonwood may have beenmore sensitive than live oak and pygmy date palmbecause of its ßeshier root system, which may haveencouraged more larval root feeding.
Leaf, stem, and root fresh and dry weights weresigniÞcantly lower for infested green buttonwoodthan for control plants. Most infested plants, especiallythose with canopies showing signs of stress, had ob-vious root feeding damage and very limited root sys-tems. Only one green buttonwood plant showing se-vere stress died before harvest; all the other greenbuttonwood plants survived. The roots, stems, andleaves of infested green buttonwood trees were no-ticeably smaller than those of the control trees. Greenbuttonwood trees infested monthly and harvested atthe end of the experiment had signiÞcantly less trunkdiameter and height than the controls. A reduction inroot weight and trunk diameter because of D. abbre-viatus larval root feeding also has been observed byothers (Nigg et al. 2001, Mannion et al. 2003). Inexperiment 2, green buttonwood plants that showedsigns of leaf yellowing, and wilt had the greatest re-duction of growth. As in experiment 1, infested liveoak did not have lower root weight than noninfestedtrees. Trunk diameter and plant height of live oakharvested at the end of the experiment 2 were notsigniÞcantly affected by treatment. The reasons forthis Þnding are not clear because larvae were recov-ered from live oak, and there were signs of feedingdamage. It seems that oaks are more tolerant to rootdamage than green buttonwood, and it may take alonger period of feeding to cause signiÞcant damage tolive oak.
For pygmy date palm, root fresh and dry weightswere lower for infested than those of noninfestedtrees. Pygmy date palm is a monocot with an adven-titious root system (Broschat and Meerow 2000). Al-though these adventitious roots are relatively tenderand succulent and had obvious signs of root feeding,there were fewer larvae recovered at each harvestcompared with green buttonwood and live oak. Thismay explain why green buttonwood had more damagethan pygmy date palm despite the palmÕs succulentadventitious root system, i.e., there presumably wasgreater feeding pressure on green buttonwood, whichis a preferred host (Mannion et al. 2003). In contrast,live oak has a much tougher root system, which pre-sumably was more resistant to larval feeding than rootsof the other species.
Although there was obvious feeding damage foundon the root systems of green buttonwood, live oak, andpygmy date palm, only green buttonwood showedsigns of severe stress in the form of leaf yellowing, wilt,and a signiÞcant reduction in leaf gas exchange andplant growth. These tests were performed in contain-ers with young trees, and although larval feeding pres-sure was high due to reinfestation with larvae, thetreatment period in these experiments lasted only afew months. The negative effects of larval root feedingmost likely are cumulative with constant feeding pres-sure as it occurs in the Þeld with multiple generations
feeding year after year. Thus, under the conditionsand parameters of these studies, D. abbreviatus couldbe considered an economic pest of green buttonwood.Further studies need to be conducted on plants in theÞeld whereD. abbreviatus are present year-round andwhere there are larvae continually feeding and dam-aging roots over a long period.
Acknowledgments
We thank S. Webb for advice throughout this study andDrs. F. S. Davies and N. Epsky for helpful comments andsuggestions during preparation of this manuscript. This isFlorida Agriculture Experiment Station Journal Series No.R-11012.
References Cited
Broschat, T. K., and A. Meerow. 2000. Ornamental palmhorticulture. University Press of Florida, Gainesville, FL.
Futch, S.H., andC.W.McCoy, Jr. 1993. Citrus root weevils.Circ. 1065. Fla. Coop. Ext. Serv., Inst. Food Agric. Sci.,Univ. of Florida, Gainesville, FL.
Godfrey, L. D., L. J. Mienke, and R. J. Wright. 1993. Effectsof larval injury by western corn rootworm (Coleoptera:Chrysomelidae) on gas exchange parameters of Þeldcorn. J. Econ. Entomol. 86: 1546Ð1556.
Johnson, M. W., S. C. Welter, N. C. Toscano, I. P. Ting, andJ. T. Trumble. 1983. Reduction of tomato leaßet photo-synthetic rates by mining activity of Liriomyza sativae(Diptera: Agromyzidae). J. Econ. Entomol. 76: 1061Ð1063.
Knapp, J. L., S. E. Simpson, J. E. Pena, and H. N. Nigg. 2000.Diaprepes root weevil: what Floridians need to know.ENY-640 Fla. Coop. Ext. Serv., Inst. Food Agric. Sci., Univ.of Florida, Gainesville, FL.
Klingeman, W. E., M. W. Van Iersel, G. D. Buntin, andS. K. Braman. 2000. Whole-plant CO2 exchange mea-surements on azaleas injured by azalea lace bug (Het-eroptera: Tingidae) feeding. J. Econ. Entomol. 93: 352Ð356.
LaPointe, S. L. 2000. History and importance of Diaprepesto agriculture in the Caribbean region, pp. 8Ð12. In Dia-prepes short course. Citrus Research and Education Cen-ter, Lake Alfred, FL.
Li, J., and J.T.A. Proctor. 1984. Simulated pest injury effectson photosynthesis and transpiration of apple leaves. Hort-Science 19: 815Ð817.
McCoy, C.W., S. H. Futch, J. H.Graham, L.W.Duncan, andH. N. Nigg. 2003. Florida citrus pest management guide:citrus root weevils. ENY-611. Fla. Coop. Ext. Serv., Inst.Food Agric. Sci., Univ. of Florida, Gainesville, FL.
Mannion,C.M.,A.Hunsberger, J.E.Pena, andL.S.Osborne.2003. Oviposition and larval survival ofDiaprepes abbre-viatus (Coleoptera: Curculionidae) on select host plants.Fla. Entomol. 86: 165Ð173.
Nigg, H. N., S. E. Simpson, N. E. El-Gholl, and F. G. Gmitter,Jr. 2001. Response of citrus rootstock seedlings to Dia-prepes abbreviatus L. (Coleoptera: Curculionidae) larvalfeeding. Proc. Fla. State Hortic. Soc. 114: 57Ð64.
Peterson, R. K., S. D. Danielson, and L. G. Higley. 1992.Photosynthetic responses of alfalfa to actual and simu-lated alfalfa weevil (Coleoptera: Curculionidae) injury.Environ. Entomol. 21: 501Ð507.
Peterson, R. K., L.G.Higley, F. J.Haile, and J.A.F. Barrigossi.1998. Mexican bean beetle (Coleoptera: Coccinellidae)
820 JOURNAL OF ECONOMIC ENTOMOLOGY Vol. 99, no. 3

injury affects photosynthesis of Glycine max and Phaseo-lus vulgaris. Environ. Entomol. 27: 373Ð381.
Quintela, E. D., J. Fan, and C. W. McCoy. 1998. Develop-ment of Diaprepes abbreviatus (Coleoptera: Curculion-idae) on artiÞcial and citrus root substrates. J. Econ.Entomol. 91: 1173Ð1179.
Riedell, W. E., and R. N. Reese. 1999. Maize morphologyand shoot CO2 assimilation after root damage by westerncorn rootworm larvae. Crop Sci. 39: 1332Ð1340.
Rogers, S., C. W. McCoy, and J. H. Graham. 2000. Larvalgrowth ofDiaprepes abbreviatus (Coleoptera: Curculion-idae) and resulting root injury to three citrus varieties intwo soil types. J. Econ. Entomol. 93: 380Ð387.
Schroeder, W. J., R. A. Hamlen, and J. B. Beavers. 1979.Survival of Diaprepes abbreviatus larvae on selected na-tive and ornamental Florida plants. Fla. Entomol. 62:309Ð312.
Simpson, S.E.,H.N.Nigg, and J.L.Knapp. 2000. Host plantsof diaprepes root weevil and their implications to theregulatory process, pp. 19Ð37. Diaprepes short course.Citrus Research and Education Center, Lake Alfred, FL.
Syvertsen, J. P., andC.W.McCoy. 1985. Leaf feeding injuryto citrus by root weevil adults: leaf area, photosynthesisand water use efÞciency. Fla. Entomol. 68: 386Ð393.
Taylor, A. O., and J. A. Rowley. 1971. Plants under climaticstress. Plant Physiol. 47: 713Ð718.
Tomlinson, P. B. 1980. The biology of trees native to trop-ical Florida, pp. 143Ð148. Harvard University Printing.Alliston, MA.
Weissling,T. J., J.E.Pena,R.M.Giblin-Davis, and J.L.KnappJr. 2002. Sugarcane rootstock borer weevil, Diaprepesabbreviatus (L.). Featured creatures, Univ. of Florida,Gainesville, FL. (http://creatures.ifas.uß.edu/citrus/sugarcane_rootstock_borer_weevil.htm).
Welter, S.C. 1989. Arthropod impact on plant gas exchange,pp. 135Ð147. In E. A. Bernays [ed.], InsectÐplant inter-actions, vol. 1. CRC, Boca Raton, FL.
Whiley, A. W., C. Searle, B. Schaffer, and B. N. Wolsten-holme. 1999. Cool orchard temperatures or growingtrees in containers can inhibit leaf gas exchange of avo-cado and mango. J. Am. Soc. Hortic. Sci. 124: 46Ð51.
Woodruff, R. E. 1964. A Puerto Rican weevil new to theUnited States (Coleoptera: Curculionidae). Fla. Dep.Agric. Div. Plant Ind. Entomol. Circ. No. 30: 1Ð2.
Received 5 August 2005; accepted 5 December 2005.
June 2006 DIAZ ET AL.: ROOT FEEDING BY D. abbreviatus 821
Related Documents











