Effect of post-teneral diets on the performance of sterile Anastrepha ludens and Anastrepha obliqua fruit flies P. Liedo 1 , D. Orozco 2 , L. Cruz-Lo ´ pez 1 , J. L. Quintero 2 , C. Becerra-Pe ´ rez 1 , M. del Refugio Herna ´ ndez 2 , A. Oropeza 1 & J. Toledo 1 1 El Colegio de la Frontera Sur (ECOSUR), Tapachula, Chiapas, Me ´ xico 2 Programa Moscamed Moscafrut, Tapachula, Chiapas, Me ´ xico Introduction Anastrepha fruit flies are important pests of fruits in the Americas (Aluja 1994). In Mexico, the technology has been developed to apply the sterile insect tech- nique (SIT) for the control of the Mexican fruit fly, Anastrepha ludens (Loew) and the West Indies fruit fly or mango fly, Anastrepha obliqua (Macquart). Over 100 million sterile flies of each species are produced weekly at the MOSCAFRUT facility, in Metapa, Chiapas, to be released in different places of Mexico and in the southern USA (Orozco et al. 2004). The advantages and benefits of using the SIT as part of an area-wide integrated pest control have been well acknowledged and the application of this technique for the control of fruit flies has expanded rapidly (Dyck et al. 2005). However, since the early days of its development, the possible harmful effects that mass-rearing, sterilization and handling, could Keywords Mexican fruit fly, pre-release feeding, sterile insect technique, West Indies fruit fly Correspondence Pablo Liedo, ECOSUR, Carretera Antiguo Aeropuerto Km 2.5, Tapachula, Chiapas, Me ´ xico 30700. E-mail: [email protected] Received: May 12, 2010; accepted: June 23, 2010. doi: 10.1111/j.1439-0418.2010.01568.x Abstract The effect of post-teneral diets on the mating performance, pheromone production and longevity of sterile Anastrepha ludens and Anastrepha obli- qua fruit flies (Diptera: Tephritidae) was investigated. Enriched pre- release diets improved male mating performance. Wild and sterile males fed on mango and orange fruits achieved the greatest copulatory suc- cess, followed by those fed on a 3 : 1 sugar : yeast (SY) diet. Males fed only on a sugar diet achieved the lowest number of matings. The mean numbers of copulas achieved by wild males were significantly higher than those by sterile males fed on a sugar-only diet, while there were no significant differences between wild males and sterile males fed on yeast diets. There was a trend of reduced mean number of copulas as the proportion of yeast was reduced in the diet, but differences were not significant. Pre-release diets had a significant effect on pheromone production. Males fed on a 3 : 1 SY diet produced the greatest amount of the three main pheromone components in A. ludens males and two major components in A. obliqua males, followed by males fed on fruits or pasteurized fruit juice. Males fed on a sugar only diet produced the lowest amounts. The longevity response to post-teneral diets was com- plex. The most yeast-rich diet and the poorest diet (sugar only) resulted in the lowest life expectancies. Flies fed on 24 : 1 SY diets showed the highest life expectancies for both males and females of the two species. Considering the tradeoffs between mating performance and longevity, the 24 : 1 SY diet would be recommended for programmes integrating the sterile insect technique, but the effects of these diets on field sur- vival and dispersal still need to be investigated. J. Appl. Entomol. J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 49

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Effect of post-teneral diets on the performance of sterileAnastrepha ludens and Anastrepha obliqua fruit fliesP. Liedo1, D. Orozco2, L. Cruz-Lopez1, J. L. Quintero2, C. Becerra-Perez1, M. del Refugio Hernandez2,A. Oropeza1 & J. Toledo1
1 El Colegio de la Frontera Sur (ECOSUR), Tapachula, Chiapas, Mexico
2 Programa Moscamed Moscafrut, Tapachula, Chiapas, Mexico
Introduction
Anastrepha fruit flies are important pests of fruits in
the Americas (Aluja 1994). In Mexico, the technology
has been developed to apply the sterile insect tech-
nique (SIT) for the control of the Mexican fruit fly,
Anastrepha ludens (Loew) and the West Indies fruit fly
or mango fly, Anastrepha obliqua (Macquart). Over
100 million sterile flies of each species are produced
weekly at the MOSCAFRUT facility, in Metapa,
Chiapas, to be released in different places of
Mexico and in the southern USA (Orozco et al.
2004).
The advantages and benefits of using the SIT as
part of an area-wide integrated pest control have
been well acknowledged and the application of this
technique for the control of fruit flies has expanded
rapidly (Dyck et al. 2005). However, since the early
days of its development, the possible harmful effects
that mass-rearing, sterilization and handling, could
Keywords
Mexican fruit fly, pre-release feeding, sterile
insect technique, West Indies fruit fly
Correspondence
Pablo Liedo, ECOSUR, Carretera Antiguo
Aeropuerto Km 2.5, Tapachula, Chiapas,
Mexico 30700. E-mail: [email protected]
Received: May 12, 2010; accepted: June 23,
2010.
doi: 10.1111/j.1439-0418.2010.01568.x
Abstract
The effect of post-teneral diets on the mating performance, pheromone
production and longevity of sterile Anastrepha ludens and Anastrepha obli-
qua fruit flies (Diptera: Tephritidae) was investigated. Enriched pre-
release diets improved male mating performance. Wild and sterile males
fed on mango and orange fruits achieved the greatest copulatory suc-
cess, followed by those fed on a 3 : 1 sugar : yeast (SY) diet. Males fed
only on a sugar diet achieved the lowest number of matings. The mean
numbers of copulas achieved by wild males were significantly higher
than those by sterile males fed on a sugar-only diet, while there were
no significant differences between wild males and sterile males fed on
yeast diets. There was a trend of reduced mean number of copulas as
the proportion of yeast was reduced in the diet, but differences were
not significant. Pre-release diets had a significant effect on pheromone
production. Males fed on a 3 : 1 SY diet produced the greatest amount
of the three main pheromone components in A. ludens males and two
major components in A. obliqua males, followed by males fed on fruits
or pasteurized fruit juice. Males fed on a sugar only diet produced the
lowest amounts. The longevity response to post-teneral diets was com-
plex. The most yeast-rich diet and the poorest diet (sugar only) resulted
in the lowest life expectancies. Flies fed on 24 : 1 SY diets showed the
highest life expectancies for both males and females of the two species.
Considering the tradeoffs between mating performance and longevity,
the 24 : 1 SY diet would be recommended for programmes integrating
the sterile insect technique, but the effects of these diets on field sur-
vival and dispersal still need to be investigated.
J. Appl. Entomol.
J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 49

have on the performance of sterile insects were
recognized (Knipling 1955, 1959). Methods have
been developed to overcome or minimize some of
these limitations. The total quality control approach,
colony management, larval diets, irradiation doses,
atmospheres and ages for sterilization are examples
of improvements at the mass-rearing factory (Ohi-
nata et al. 1977; Chang et al. 2001; FAO/IAEA/
USDA 2003; Toledo et al. 2004; Calkins and Parker
2005; Caceres et al. 2007; Liedo et al. 2007; Resilva
et al. 2007; Rull et al. 2007; Collins et al. 2008).
Development of pupal packing and shipping, adult
emergence and release methods, use of hormones,
nutrients and semiochemicals to improve mating
performance and implementation of field cage evalu-
ation of mating behaviour, are examples of post-fac-
tory improvements (Sivisnki et al. 1994; Shelly and
McInnis 2001; Hendrichs et al. 2002; Shelly et al.
2002b, 2007).
There is a large body of evidence that the addition
of protein to the diet of adult fruit flies enhances
their mating performance and other fitness attri-
butes. This could have positive effects in the applica-
tion of the SIT (Blay and Yuval 1997; Papadopoulos
et al. 1998; Yuval et al. 1998, 2002, 2007; Field and
Yuval 1999; Taylor and Yuval 1999; Kaspi and Yuval
2000; Kaspi et al. 2000; Shelly and Kennelly 2002;
Shelly et al. 2002a, 2005, 2006; Shelly and McInnis
2003; Cresoni-Pereira and Zucoloto 2006; Manrak-
han and Lux 2006; Perez-Staples et al. 2007, 2008a,
2009; Gavriel et al. 2009). In the case of Anastrepha
fruit flies, there is some evidence on the positive
effect of protein on their sexual behaviour (Aluja
et al. 2001, 2008; Cresoni-Pereira and Zucoloto
2006; Perez-Staples et al. 2008b). The purpose of this
research was to assess the effect of various pre-
release diets on the mating performance, pheromone
production and longevity of sterile A. ludens and
A. obliqua males.
Materials and Methods
Insects
Wild A. ludens flies were obtained as larvae from
infested sour oranges (Citrus aurantium L.) and wild
A. obliqua were obtained as larvae from infested yel-
low mombin fruits or jobos (Spondias mombin L.) col-
lected in the Soconusco region of Chiapas, Mexico.
The mass-reared non-irradiated and irradiated flies
were obtained as pupae from the MOSCAFRUT
mass-rearing facility at Metapa, Chiapas, Mexico.
Sterile flies were c-irradiated as pupae at 80 Gy
2 days before adult emergence. The mass-reared flies
were from strains with approximately 130 and 110
generations under mass rearing conditions, for
A. ludens and A. obliqua, respectively.
Mating performance
Initially, a series of five field cage mating tests were
carried out with A. ludens and one with A. obliqua.
The tests were done following the quality control
protocol commonly used at the MOSCAFRUT facility
(FAO/IAEA/USDA 2003).
After emergence, adult flies were sorted by sex
and placed in groups of 40 individuals in 30
cm · 30 cm · 30 cm Plexiglas cages. Before the tests,
these cages were maintained at the laboratory at
24 � 2�C, 70 � 10% RH, 550 � 50 lux light inten-
sity, with a 12 : 12 L : D photoperiod. Water and
food were provided ad libitum. The types of foods
were: (i) orange, Citrus sinensis (L.) Osbeck cv Valen-
cia or mango, Mangifera indica L. cv Ataulfo (�200 g
pieces); (ii) full diet, which was the standard 3 : 1
sugar : hydrolysed yeast laboratory diet; and (iii)
only sugar (standard cane sugar, dry sucrose). Wild
females always were fed a full diet.
Five field cages, 2.9 m diameter · 2 m height (Cal-
kins and Webb 1983), were set up at a mango orch-
ard near Tapachula, Chiapas, Mexico (14� 55¢ 08.9¢¢N, 92� 16¢34.2¢¢W and 137 msl). Six small orange
and mango potted trees (�1.5 m tall) were placed at
the periphery and central part of each field cage.
Each field cage was considered as a replicate. Thir-
teen males per treatment and 39 wild females were
released per field cage in the five A. ludens tests
(table 1). Twelve males per treatment and 48 wild
females in the A. obliqua test (table 2). Forty-eight
hours before the test, flies were individually marked
by gluing a small piece of paper printed with a num-
ber on their thorax for treatment identification. This
type of mark does not interfere with the sexual
activity of the flies (Meza et al. 2005). Mating pairs
were vial collected and the number and type of mat-
ings was recorded. The five tests for A. ludens were
made between 15:00 and 19:00 hours, and the test
for A. obliqua was between 06:00 and 12:00 hours.
In the tests for A. ludens we first compared wild
males fed on orange with sterile males fed on sugar
only and full diet. In the second, third and fourth
tests we compared the effect of orange, sugar only
and full diet on the performance of sterile, fertile
and wild males, respectively. In the fifth test the
effect of two fruits, mango and orange and full diet
was compared on sterile males. For A. obliqua only
Pre-release feeding on sterile Anastrepha male performance P. Liedo et al.
50 J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH
one test was done where wild males were fed on
orange and sterile males were fed on orange, full
diet and only sugar.
The age of the flies, at the time of the test was,
18–19 days for the wild flies and 12 days for the
mass-reared fertile and sterile flies in the case of A.
ludens, and 15 days for wild and 8 days for mass-
reared sterile A. obliqua flies.
A second set of tests were carried out to look at
the effect of different sugar : hydrolysed yeast (SY)
ratios. Laboratory and field cage conditions were as
described above. Groups of 200 males were provided
one of the following four diets: Only sugar (1 : 0),
and 24 : 1, 9 : 1 and 3 : 1 SY ratios. In the case of A.
ludens, eight sterile males from each diet treatment,
eight wild males and 40 wild females fed with the
standard 3 : 1 SY diet were released per field cage.
In the case of A. obliqua, 10 sterile males of each diet
treatment, 10 wild males and 50 wild females fed on
full diet were released in each field cage. Six repli-
cates were carried out per species.
Pheromone analysis
Pheromone volatiles emitted by males were collected
using an air-entrainment technique. Ten males were
confined in a 100 ml glass entrainment container
[4.8 cm inner diameter (ID) · 12.5 cm long]. Vola-
tiles were drawn from the container, using purifier
air that had previously passed through an activated
charcoal trap, onto a glass volatile collection trap
(4 mm ID · 40 mm long) containing 50 mg of Super
Q adsorbent (Alltech Associates, Deerfield, IL) (Heath
and Manukian 1992). Air was drawn through the
trap at a rate of 1 l/min by a vacuum pump. At the
conclusion of each air entrainment, which lasted 4 h,
the volatiles were eluted from the adsorbent with
200 ll of methylene chloride (Baker, HPLC grade)
and 100 gg of tridecane was added as an internal
standard for subsequent quantification. The samples
were kept at )20�C before analysis.
Volatiles were analysed by means of gas chromatog-
raphy-mass spectrometry using a Varian Star 3400 CX
gas chromatograph linked to a Varian Saturn 4D mass
spectrometer (GC-MS). The samples were analysed
using a fused silica column (30 m · 0.25 mm) coated
with poly (5%-diphenyl-95%-dimethylsiloxane) pro-
grammed from 50�C to 250�C at 15 �C/min. The
carrier gas was helium. The injector port temperature
was held at 200�C. Compounds were identified using
their retention times, Kovat index (KI), and mass
spectra and comparing these data with those of
synthetic standards. Synthetic standards of farnesene
(mixture of isomers that includes (E,E)-a-farnesene)
and (Z)-3-nonenol were supplied by Aldrich (Toluca,
Mexico).
Adult A. ludens sterile males were exposed to five
types of food: (i) standard laboratory full diet (3 : 1
SY dry mixture); (ii) pulp of orange fruits cv. Valen-
cia; (iii) pulp of mango fruits cv Ataulfo; (iv) com-
mercial pasteurized peach juice (Herdez �); and (v)
sugar only. These males were placed in 30 cm ·30 cm · 30 cm Plexiglas cages after emergence.
Every day a sample of 10 males per treatment was
taken and this was repeated during six consecutive
days, from 6- to 12-day-old flies. Four known main
pheromone components were quantified, (E,E)-a-
farnesene, suspensolide, anastrephin and epianastre-
phin.
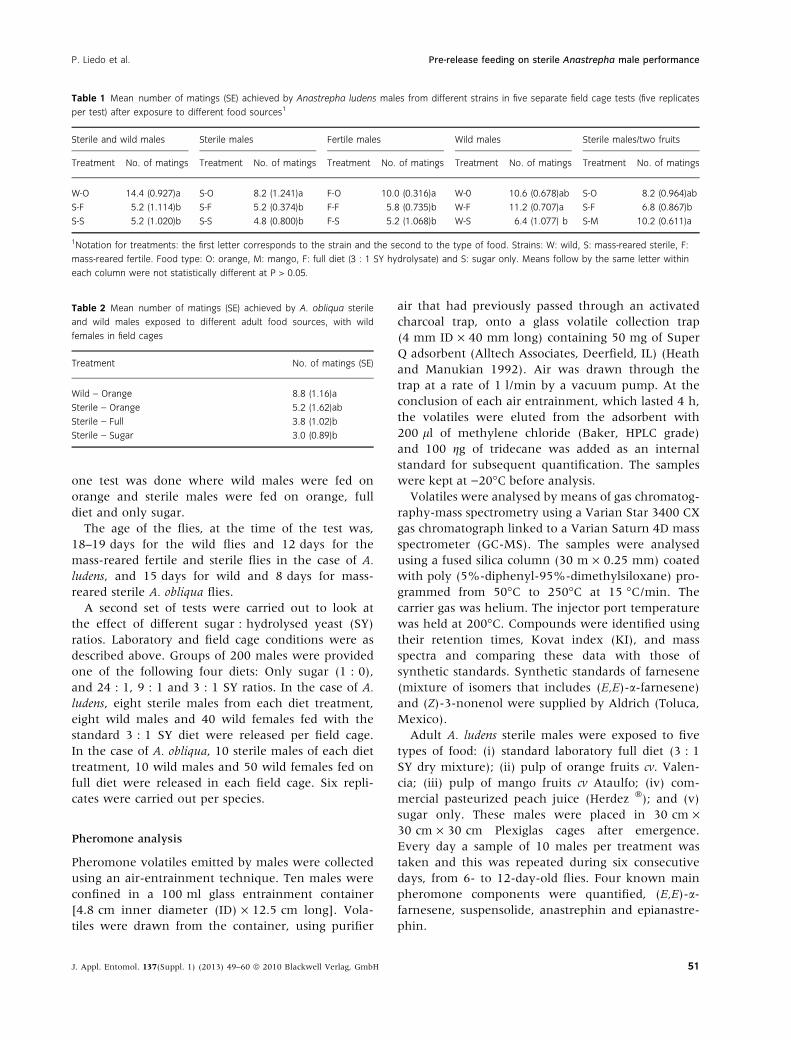
Table 1 Mean number of matings (SE) achieved by Anastrepha ludens males from different strains in five separate field cage tests (five replicates
per test) after exposure to different food sources1
Sterile and wild males Sterile males Fertile males Wild males Sterile males/two fruits
Treatment No. of matings Treatment No. of matings Treatment No. of matings Treatment No. of matings Treatment No. of matings
W-O 14.4 (0.927)a S-O 8.2 (1.241)a F-O 10.0 (0.316)a W-0 10.6 (0.678)ab S-O 8.2 (0.964)ab
S-F 5.2 (1.114)b S-F 5.2 (0.374)b F-F 5.8 (0.735)b W-F 11.2 (0.707)a S-F 6.8 (0.867)b
S-S 5.2 (1.020)b S-S 4.8 (0.800)b F-S 5.2 (1.068)b W-S 6.4 (1.077) b S-M 10.2 (0.611)a
1Notation for treatments: the first letter corresponds to the strain and the second to the type of food. Strains: W: wild, S: mass-reared sterile, F:
mass-reared fertile. Food type: O: orange, M: mango, F: full diet (3 : 1 SY hydrolysate) and S: sugar only. Means follow by the same letter within
each column were not statistically different at P > 0.05.
Table 2 Mean number of matings (SE) achieved by A. obliqua sterile
and wild males exposed to different adult food sources, with wild
females in field cages
Treatment No. of matings (SE)
Wild – Orange 8.8 (1.16)a
Sterile – Orange 5.2 (1.62)ab
Sterile – Full 3.8 (1.02)b
Sterile – Sugar 3.0 (0.89)b
P. Liedo et al. Pre-release feeding on sterile Anastrepha male performance
J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 51

Adult A. obliqua males were exposed to three food
sources: (i) standard laboratory 3 : 1 SY full diet: (ii)
pulp of mango fruits cv Ataulfo; and (iii) sugar only.
Pheromone components were collected from males
that were 8 to 10 days old. Three known com-
pounds, z-3-nonenol, (Z, E)-a-farnesene, and (E, E)-
a-farnesene, and one unknown compound, were
quantified.
Longevity
The effect of sugar : hydrolysed yeast ratios on the
longevity of mass-reared non-irradiated A. ludens
and A. obliqua fruit flies was evaluated under labora-
tory conditions by comparing four diets (only sugar
1 : 0, and 24 : 1, 9 : 1, and 3 : 1 S: : Y). Aluminium
frame, mesh covered, 80 cm · 50 cm · 15 cm cages,
were used, three cages per treatment. In each cage
about 2000 adult flies were released. Food, according
to each treatment, and water, were provided ad libi-
tum. Dead flies were removed daily from the cages.
The number and sex of the dead flies were recorded.
Data analysis
The data from the mating tests were analysed by
Fisher’s PLSD statistical test and P values were cal-
culated for pair-wise comparisons in each test. For
the pheromone analysis, the relative amount of
each compound was estimated from the GC graphs.
These figures were square root transformed for anal-
ysis of variance (anova). In the case of A. obliqua, a
Kruskal–Wallis test was used. Life tables were con-
structed for demographic analysis of longevity (Ca-
rey 1993). Survival data were analysed using the
Cox proportional hazard model (Everitt and Pickles
2004). Independent analyses were done for males
and females. Survival of flies exposed to 3 : 1, 9 : 1
and 24 : 1 SY diets was compared with survival of
flies fed with only sugar (control). Treatments were
compared with 95% CI. Statistical analyses were
performed using R software (R v.2.9.2. 2009, the R
Foundation for Statistical Computing, http://www.
r-project.org/).
Results
Mating
A total of 713 copulas were observed in the five ini-
tial tests with A. ludens, 56.1% were achieved by
males fed on fruit (even though only 40% of males
had been exposed to fruit), 28.7% by males fed on
full diet (33% of males on this sugar-yeast hydroly-
sed diet) and 15.1% by males on a sugar only diet
(27% of males present). The mean number (SE) of
matings recorded for A. ludens males for different
combinations of strain (wild, mass-reared fertile and
mass-reared sterile) and food type (orange and
mango fruit pieces, sugar only and full diet) in the
five initial tests is shown in table 1.
In the first of the five tests, with wild males fed
on orange and sterile males fed on sugar only or full
diet, the difference between wild males and sterile
males was significant (P < 0.0001), with the wild
ones achieving significantly more matings. The dif-
ference between sterile males fed on a full diet or
sugar only was not significant (P > 0.05). In order to
separate the effect of ‘wild’, from the effect of ‘fruit’,
the second test was designed using only mass-reared
sterile males (table 1). In this second test, the differ-
ences in matings between orange fruit and full diet
and orange fruit and sugar fed males were significant
in favour of fruit feeding (P = 0.0328 and P =
0.0181, respectively). The difference between full
diet and sugar only fed males was not significant
(P > 0.05). In the third test, mass-reared not steril-
ized males showed a similar pattern (table 1). The
difference in mating success between males fed on
fruit and males fed on full diet and sugar only was
significant (P = 0.0023 and P = 0.0009, respectively),
whereas the difference between males fed on full
diet and males fed on sugar only was not significant
(P > 0.05). Wild males showed a slightly different
mating pattern in the fourth test. There was no sig-
nificant difference between orange fruit and the full
diet (P > 0.05) and full diet was significantly differ-
ent from sugar only fed males (P = 0.0341) (table 1).
In the last test, while sterile males fed on mango
showed a significantly better mating performance
than those fed on the full diet (P = 0.0213), there
was no significant difference between males fed on
mango and those fed on orange, nor between those
fed on orange and those fed on full diet (P > 0.05)
(table 1).
In the field cage test with A. obliqua, a total of 104
matings were observed, 42.3% were by wild males
fed on orange, 25.0% by sterile males fed on orange,
18.3% by sterile males fed on full diet and 14.4% by
males fed on sugar only. The mean numbers of mat-
ings (SE) are shown in table 2. There was no signifi-
cant difference between sterile and wild males fed
with orange, while orange fed wild males performed
significantly better than sterile males fed on the full
diet or sugar. No significant differences were found
among sterile insect treatments.
Pre-release feeding on sterile Anastrepha male performance P. Liedo et al.
52 J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH

Results from the second series of field cage mating
tests with males exposed to different SY ratios are
shown in table 3. A total of 123 and 86 matings
were observed for A. ludens and A. obliqua, respec-
tively. In both species, the mean numbers of matings
by wild males were significantly different from those
by sterile males fed on a sugar-only diet. There were
no significant differences between wild males and
sterile males fed on yeast diets, although the mean
number of matings tended to decline as the propor-
tion of yeast was reduced in the diet.
Pheromone
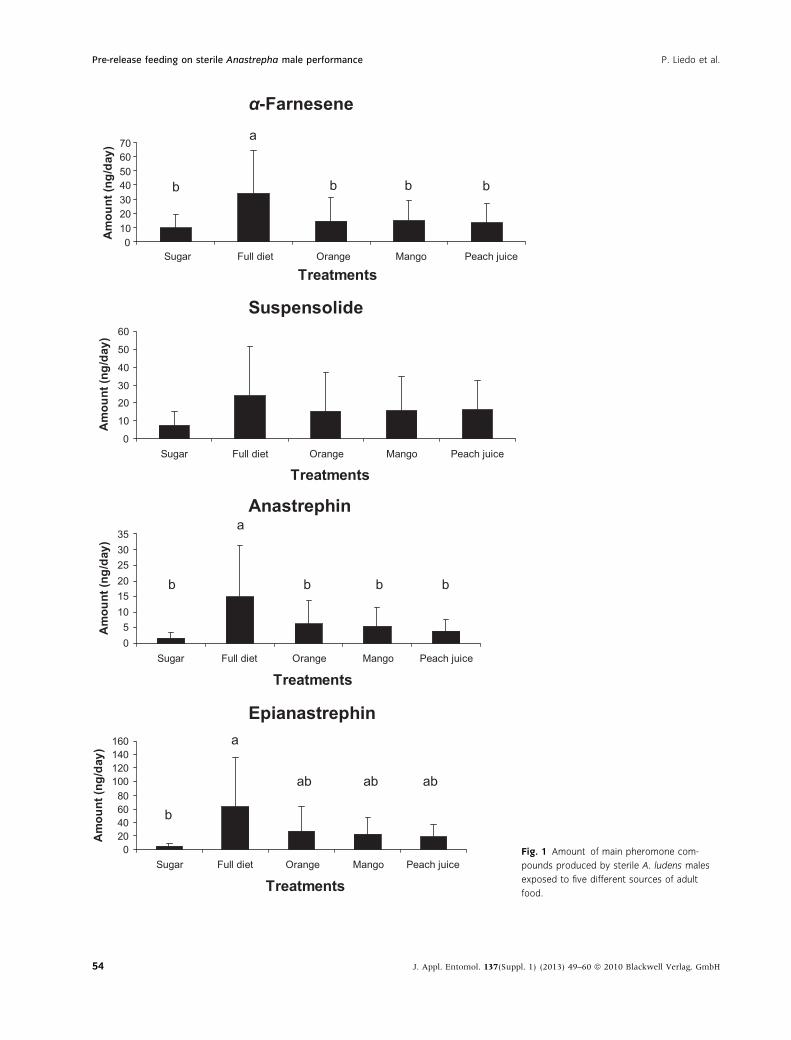
In A. ludens, diet induced differences in the amount
of the main pheromone compounds produced by
males were significant for (E,E)-a-farnesene
(F = 4.27, d.f. = 4, 12, P = 0.0223), anastrephin (F =
4.20, d.f. = 4, 12, P = 0.0234) and epianastrephin
(F = 5.57, d.f. = 4, 12, P = 0.0089). Differences in
the amount produced of suspensolide were not sig-
nificant (F = 2.42, d.f. = 4, 12, P = 0.1057). In A. ob-
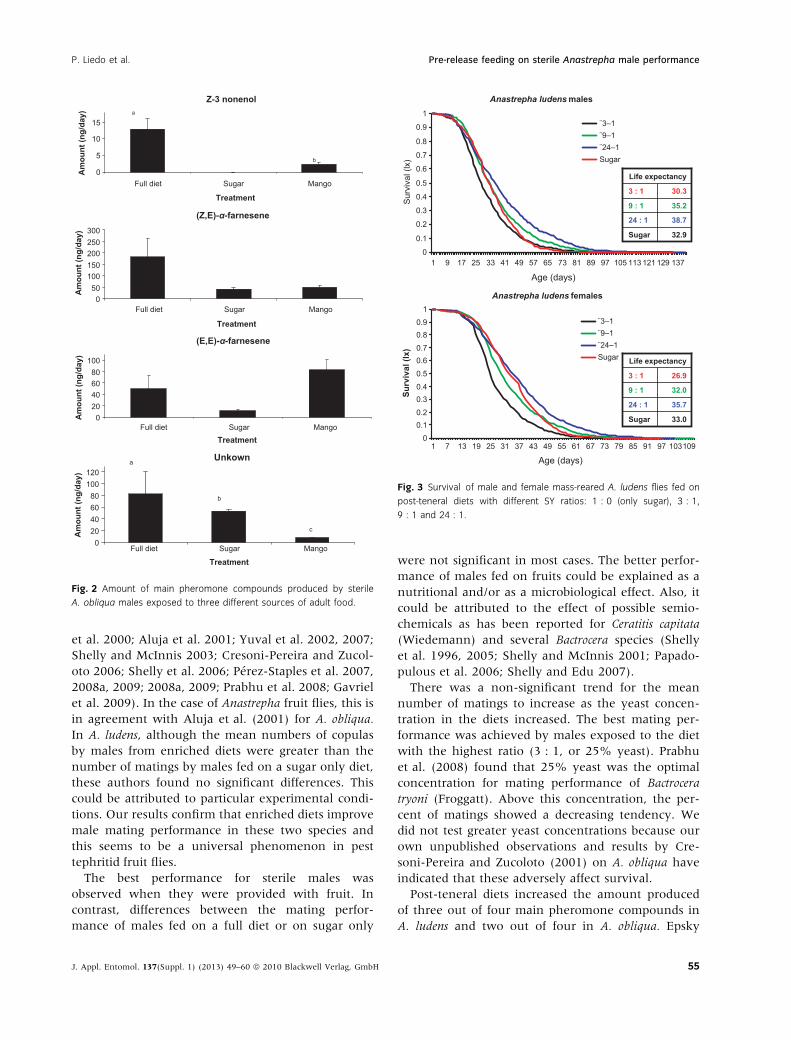
liqua, differences were significant for Z-3-nonenol
(H = 3.86, d.f = 1, P = 0.05) and the unknown com-
pound (H = 6.49, d.f. = 2, P = 0.039), but non-sig-
nificant in the other two compounds (H = 4.36,
d.f. = 2, P = 0.113 and H = 4.27, d.f. = 2, P = 0.118
for Z,E-a-farnesene, and E,E-a-farnesene, respec-
tively).
In most cases, males fed with the standard full diet
produced the greatest amount of each compound
and males fed with only sugar produced the lowest
amount (figs 1 and 2). A. ludens males fed with
mango, orange or peach juice produced an interme-
diate amount of each compound. The only exception
was with (E, E)-a-farnesene in A. obliqua, where
males fed on mango produced greater amounts than
those fed on the full diet.
Longevity
Anastrepha ludens longevity response to pre-release
diets was complex. In males, the lowest expectation
of life was observed on the most yeast rich diet
(3 : 1 SY), followed by the non-yeast diet (sugar-
only). In females, the lowest life expectancy was
observed on the 3 : 1 diet, followed by the 9 : 1 diet
and the sugar only diet. The effect of the four SY
ratios on the survival of A. ludens can be seen in
fig. 3. According to the Cox proportional hazard
model and the Likehood Ratio test, differences in
survival were highly significant (table 4). The SY
ratio that resulted in the highest life expectancy was
the 24 : 1 ratio for both males and females.
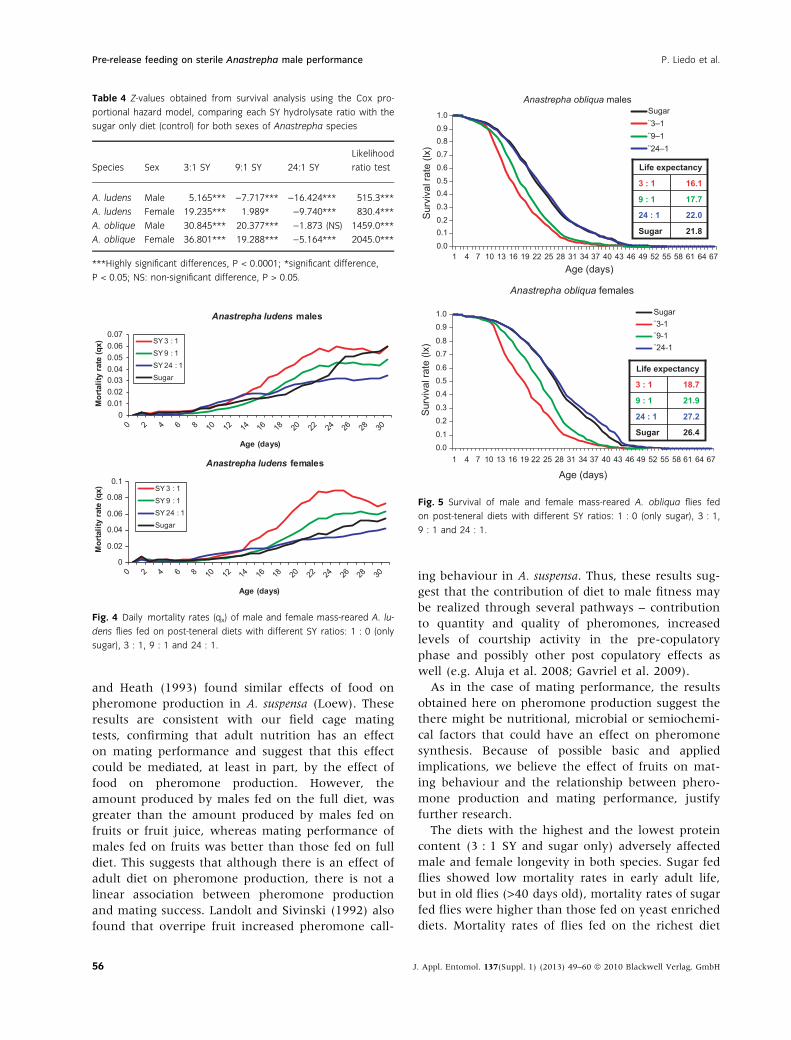
The daily mortality rates (qx) during the first
30 days of A. ludens adult life are shown in fig. 4. In
males, the sugar only diet showed the lowest mortal-
ity rates during the first 20–24 days, then mortality
increased and was the highest after 30 days. In
females, the mortality rates under full diet (3 : 1 SY)
were the highest during the initial 30 days. The 9 : 1
and 24 : 1 SY diets showed the lowest mortality rates
through most of the lifespan of males and females.
The survival response of A. obliqua to the four SY
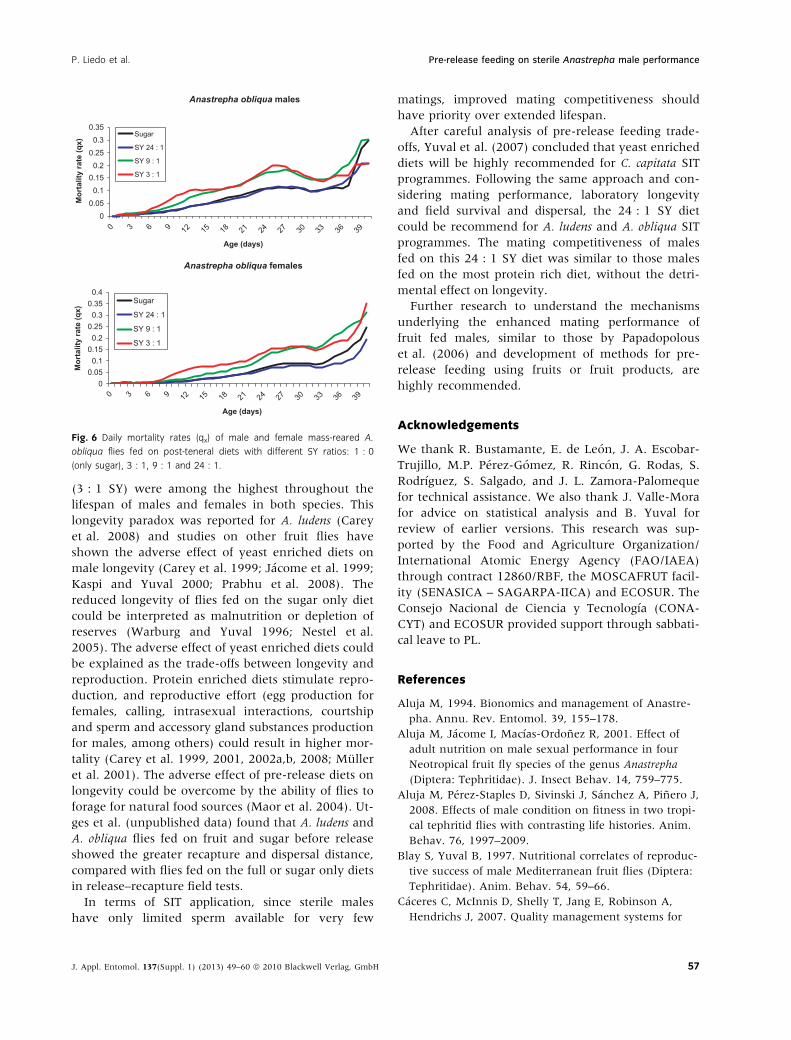
diets are shown in fig. 5 and the daily mortality rates
(qx) in fig. 6. Differences between flies fed on sugar
only and all other treatments were significant, except
with A. obliqua males fed on the 24 : 1 SY diet, where
the difference was non-significant (table 4). The most
protein rich diet (SY 3 : 1) showed the lowest life
expectancies for both, males and females. The highest
life expectancy was observed with the SY 24 : 1 diet
for both males and females.
Correspondingly, the highest mortality rates for
both A. obliqua males and females were found in the
3 : 1 SY diet and the lowest mortality rates were
observed in males and females fed on sugar only
and the 24 : 1 SY diet. The cross-over of the mortal-
ity schedules observed in A. ludens (fig. 4) was not
seen in A. obliqua females during the first 40 days of
adult life and was observed in males around 37 to
40 days old (fig. 6).
Discussion
Mating performance of A. ludens and A. obliqua males
was significantly affected by adult pre-release diet.
Fruit, and to a lesser extent, yeast enriched diets,
improved mating performance of wild, mass-reared
and sterile males. Generally, this is in agreement with
what has been reported for other fruit fly species
(Blay and Yuval 1997; Kaspi and Yuval 2000; Kaspi
Table 3 Mean number (SE) of matings achieved by wild and sterile
males of A. ludens and A. obliqua fed on diets with different SY
hydrolysate ratios in tests conducted in field cages
Strain – SY ratio A. ludens A. obliqua
Wild –3 : 1 5.00 (0.58)a 4.50 (0.81)a
Sterile –3 : 1 4.67 (0.61)ab 3.50 (1.34)ab
Sterile –9 : 1 4.33 (0.42)ab 2.50 (1.02)ab
Sterile –24 : 1 3.50 (0.76)ab 2.33 (0.56)ab
Sterile –1 : 0 3.00 (0.77)b 1.50 (0.43)b
Means followed by the same letter were not statistically different at
P > 0.05 according to Fisher’s PLSD.
P. Liedo et al. Pre-release feeding on sterile Anastrepha male performance
J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 53
a
b b b b
a
b b b b
a
ab ab ab
b
α-Farnesene
0
10
20
30
40
50
60
70
Sugar Full diet Orange Mango Peach juice
Treatments
Am
ount
(ng/
day)
Suspensolide
0
10
20
30
40
50
60
Sugar Full diet Orange Mango Peach juice
Treatments
Am
ount
(ng/
day)
Anastrephin
0
5
10
15
20
25
30
35
Sugar Full diet Orange Mango Peach juice
Treatments
Am
ount
(ng/
day)
Epianastrephin
0
20
40
60
80
100
120
140
160
Sugar Full diet Orange Mango Peach juice
Treatments
Am
ount
(ng/
day)
Fig. 1 Amount of main pheromone com-
pounds produced by sterile A. ludens males
exposed to five different sources of adult
food.
Pre-release feeding on sterile Anastrepha male performance P. Liedo et al.
54 J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH
et al. 2000; Aluja et al. 2001; Yuval et al. 2002, 2007;
Shelly and McInnis 2003; Cresoni-Pereira and Zucol-
oto 2006; Shelly et al. 2006; Perez-Staples et al. 2007,
2008a, 2009; 2008a, 2009; Prabhu et al. 2008; Gavriel
et al. 2009). In the case of Anastrepha fruit flies, this is
in agreement with Aluja et al. (2001) for A. obliqua.
In A. ludens, although the mean numbers of copulas
by males from enriched diets were greater than the
number of matings by males fed on a sugar only diet,
these authors found no significant differences. This
could be attributed to particular experimental condi-
tions. Our results confirm that enriched diets improve
male mating performance in these two species and
this seems to be a universal phenomenon in pest
tephritid fruit flies.
The best performance for sterile males was
observed when they were provided with fruit. In
contrast, differences between the mating perfor-
mance of males fed on a full diet or on sugar only
were not significant in most cases. The better perfor-
mance of males fed on fruits could be explained as a
nutritional and/or as a microbiological effect. Also, it
could be attributed to the effect of possible semio-
chemicals as has been reported for Ceratitis capitata
(Wiedemann) and several Bactrocera species (Shelly
et al. 1996, 2005; Shelly and McInnis 2001; Papado-
pulous et al. 2006; Shelly and Edu 2007).
There was a non-significant trend for the mean
number of matings to increase as the yeast concen-
tration in the diets increased. The best mating per-
formance was achieved by males exposed to the diet
with the highest ratio (3 : 1, or 25% yeast). Prabhu
et al. (2008) found that 25% yeast was the optimal
concentration for mating performance of Bactrocera
tryoni (Froggatt). Above this concentration, the per-
cent of matings showed a decreasing tendency. We
did not test greater yeast concentrations because our
own unpublished observations and results by Cre-
soni-Pereira and Zucoloto (2001) on A. obliqua have
indicated that these adversely affect survival.
Post-teneral diets increased the amount produced
of three out of four main pheromone compounds in
A. ludens and two out of four in A. obliqua. Epsky
Z-3 nonenol
0
5
10
15
MangoSugarFull diet
MangoSugarFull diet
MangoSugarFull diet
MangoSugarFull diet
Treatment
Am
ount
(ng/
day) a
b
(Z,E)-α-farnesene
050
100150200250300
Treatment
Am
ount
(ng/
day)
(E,E)-α-farnesene
020406080
100
Treatment
Am
ount
(ng/
day)
Unkown
020406080
100120
Treatment
Am
ount
(ng/
day)
a
b
c
Fig. 2 Amount of main pheromone compounds produced by sterile
A. obliqua males exposed to three different sources of adult food.
Anastrepha ludens females
0
0.10.2
0.30.4
0.5
0.60.7
0.80.9
1
1 7 13 19 25 31 37 43 49 55 61 67 73 79 85 91 97 103109
Age (days)
Surv
ival
(lx)
¨3–1¨9–1¨24–1Sugar
Anastrepha ludens males
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
1 9 17 25 33 41 49 57 65 73 81 89 97 105 113 121 129 137
Age (days)
Sur
viva
l (lx
)
¨3–1¨9–1¨24–1Sugar
32.9Sugar
38.724 : 1
35.29 : 1
30.33 : 1
Life expectancy
33.0Sugar
35.724 : 1
32.09 : 1
26.93 : 1
Life expectancy
Fig. 3 Survival of male and female mass-reared A. ludens flies fed on
post-teneral diets with different SY ratios: 1 : 0 (only sugar), 3 : 1,
9 : 1 and 24 : 1.
P. Liedo et al. Pre-release feeding on sterile Anastrepha male performance
J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 55
and Heath (1993) found similar effects of food on
pheromone production in A. suspensa (Loew). These
results are consistent with our field cage mating
tests, confirming that adult nutrition has an effect
on mating performance and suggest that this effect
could be mediated, at least in part, by the effect of
food on pheromone production. However, the
amount produced by males fed on the full diet, was
greater than the amount produced by males fed on
fruits or fruit juice, whereas mating performance of
males fed on fruits was better than those fed on full
diet. This suggests that although there is an effect of
adult diet on pheromone production, there is not a
linear association between pheromone production
and mating success. Landolt and Sivinski (1992) also
found that overripe fruit increased pheromone call-
ing behaviour in A. suspensa. Thus, these results sug-
gest that the contribution of diet to male fitness may
be realized through several pathways – contribution
to quantity and quality of pheromones, increased
levels of courtship activity in the pre-copulatory
phase and possibly other post copulatory effects as
well (e.g. Aluja et al. 2008; Gavriel et al. 2009).
As in the case of mating performance, the results
obtained here on pheromone production suggest the
there might be nutritional, microbial or semiochemi-
cal factors that could have an effect on pheromone
synthesis. Because of possible basic and applied
implications, we believe the effect of fruits on mat-
ing behaviour and the relationship between phero-
mone production and mating performance, justify
further research.
The diets with the highest and the lowest protein
content (3 : 1 SY and sugar only) adversely affected
male and female longevity in both species. Sugar fed
flies showed low mortality rates in early adult life,
but in old flies (>40 days old), mortality rates of sugar
fed flies were higher than those fed on yeast enriched
diets. Mortality rates of flies fed on the richest diet
Table 4 Z-values obtained from survival analysis using the Cox pro-
portional hazard model, comparing each SY hydrolysate ratio with the
sugar only diet (control) for both sexes of Anastrepha species
Species Sex 3:1 SY 9:1 SY 24:1 SY
Likelihood
ratio test
A. ludens Male 5.165*** )7.717*** )16.424*** 515.3***
A. ludens Female 19.235*** 1.989* )9.740*** 830.4***
A. oblique Male 30.845*** 20.377*** )1.873 (NS) 1459.0***
A. oblique Female 36.801*** 19.288*** )5.164*** 2045.0***
***Highly significant differences, P < 0.0001; *significant difference,
P < 0.05; NS: non-significant difference, P > 0.05.
Anastrepha ludens males
00.010.020.030.040.050.060.07
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Age (days)
Mor
talit
y ra
te (q
x) SY 3 : 1
SY 9 : 1SY 24 : 1
Sugar
Anastrepha ludens females
0
0.02
0.04
0.06
0.08
0.1
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Age (days)
Mor
talit
y ra
te (q
x) SY 3 : 1
SY 9 : 1SY 24 : 1
Sugar
Fig. 4 Daily mortality rates (qx) of male and female mass-reared A. lu-
dens flies fed on post-teneral diets with different SY ratios: 1 : 0 (only
sugar), 3 : 1, 9 : 1 and 24 : 1.
Anastrepha obliqua males
0.0
0.10.2
0.3
0.40.5
0.6
0.7
0.80.9
1.0
Age (days)
Sur
viva
l rat
e (lx
)
Sugar¨3–1¨9–1¨24–1
Anastrepha obliqua females
0.00.1
0.20.3
0.40.50.6
0.70.8
0.91.0
1 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 58 61 64 67
1 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 58 61 64 67
Age (days)
Sur
viva
l rat
e (lx
)
Sugar¨3-1¨9-1¨24-1
21.8Sugar
22.024 : 1
17.79 : 1
16.13 : 1
Life expectancy
26.4Sugar
27.224 : 1
21.99 : 1
18.73 : 1
Life expectancy
Fig. 5 Survival of male and female mass-reared A. obliqua flies fed
on post-teneral diets with different SY ratios: 1 : 0 (only sugar), 3 : 1,
9 : 1 and 24 : 1.
Pre-release feeding on sterile Anastrepha male performance P. Liedo et al.
56 J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH
(3 : 1 SY) were among the highest throughout the
lifespan of males and females in both species. This
longevity paradox was reported for A. ludens (Carey
et al. 2008) and studies on other fruit flies have
shown the adverse effect of yeast enriched diets on
male longevity (Carey et al. 1999; Jacome et al. 1999;
Kaspi and Yuval 2000; Prabhu et al. 2008). The
reduced longevity of flies fed on the sugar only diet
could be interpreted as malnutrition or depletion of
reserves (Warburg and Yuval 1996; Nestel et al.
2005). The adverse effect of yeast enriched diets could
be explained as the trade-offs between longevity and
reproduction. Protein enriched diets stimulate repro-
duction, and reproductive effort (egg production for
females, calling, intrasexual interactions, courtship
and sperm and accessory gland substances production
for males, among others) could result in higher mor-
tality (Carey et al. 1999, 2001, 2002a,b, 2008; Muller
et al. 2001). The adverse effect of pre-release diets on
longevity could be overcome by the ability of flies to
forage for natural food sources (Maor et al. 2004). Ut-
ges et al. (unpublished data) found that A. ludens and
A. obliqua flies fed on fruit and sugar before release
showed the greater recapture and dispersal distance,
compared with flies fed on the full or sugar only diets
in release–recapture field tests.
In terms of SIT application, since sterile males
have only limited sperm available for very few
matings, improved mating competitiveness should
have priority over extended lifespan.
After careful analysis of pre-release feeding trade-
offs, Yuval et al. (2007) concluded that yeast enriched
diets will be highly recommended for C. capitata SIT
programmes. Following the same approach and con-
sidering mating performance, laboratory longevity
and field survival and dispersal, the 24 : 1 SY diet
could be recommend for A. ludens and A. obliqua SIT
programmes. The mating competitiveness of males
fed on this 24 : 1 SY diet was similar to those males
fed on the most protein rich diet, without the detri-
mental effect on longevity.
Further research to understand the mechanisms
underlying the enhanced mating performance of
fruit fed males, similar to those by Papadopolous
et al. (2006) and development of methods for pre-
release feeding using fruits or fruit products, are
highly recommended.
Acknowledgements
We thank R. Bustamante, E. de Leon, J. A. Escobar-
Trujillo, M.P. Perez-Gomez, R. Rincon, G. Rodas, S.
Rodrıguez, S. Salgado, and J. L. Zamora-Palomeque
for technical assistance. We also thank J. Valle-Mora
for advice on statistical analysis and B. Yuval for
review of earlier versions. This research was sup-
ported by the Food and Agriculture Organization/
International Atomic Energy Agency (FAO/IAEA)
through contract 12860/RBF, the MOSCAFRUT facil-
ity (SENASICA – SAGARPA-IICA) and ECOSUR. The
Consejo Nacional de Ciencia y Tecnologıa (CONA-
CYT) and ECOSUR provided support through sabbati-
cal leave to PL.
References
Aluja M, 1994. Bionomics and management of Anastre-
pha. Annu. Rev. Entomol. 39, 155–178.
Aluja M, Jacome I, Macıas-Ordonez R, 2001. Effect of
adult nutrition on male sexual performance in four
Neotropical fruit fly species of the genus Anastrepha
(Diptera: Tephritidae). J. Insect Behav. 14, 759–775.
Aluja M, Perez-Staples D, Sivinski J, Sanchez A, Pinero J,
2008. Effects of male condition on fitness in two tropi-
cal tephritid flies with contrasting life histories. Anim.
Behav. 76, 1997–2009.
Blay S, Yuval B, 1997. Nutritional correlates of reproduc-
tive success of male Mediterranean fruit flies (Diptera:
Tephritidae). Anim. Behav. 54, 59–66.
Caceres C, McInnis D, Shelly T, Jang E, Robinson A,
Hendrichs J, 2007. Quality management systems for
Anastrepha obliqua males
00.050.1
0.150.2
0.250.3
0.35
0 3 6 9 12 15 18 21 24 27 30 33 36 39
Age (days)
Mor
talit
y ra
te (q
x)
Sugar
SY 24 : 1
SY 9 : 1
SY 3 : 1
Anastrepha obliqua females
00.050.1
0.150.2
0.250.3
0.350.4
0 3 6 9 12 15 18 21 24 27 30 33 36 39
Age (days)
Mor
talit
y ra
te (q
x)
Sugar
SY 24 : 1
SY 9 : 1
SY 3 : 1
Fig. 6 Daily mortality rates (qx) of male and female mass-reared A.
obliqua flies fed on post-teneral diets with different SY ratios: 1 : 0
(only sugar), 3 : 1, 9 : 1 and 24 : 1.
P. Liedo et al. Pre-release feeding on sterile Anastrepha male performance
J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 57

fruit fly (Diptera: Tephritidae) sterile insect technique.
Fla. Entomol. 90, 1–9.
Calkins CO, Parker A, 2005. Sterile insect quality. In:
Sterile insect technique. Principles and practice in area-
wide integrated pest management. Ed. by Dyck VA,
Hendrichs J, Robinson AS, Springer, Dordrecht, The
Netherlands, 269–296.
Calkins CO, Webb JC, 1983. A cage and support frame-
work for behavioral tests of fruit flies in the field. Fla.
Entomol. 66, 512–514.
Carey JR, 1993. Applied demography for biologists with
special emphasis on insects. Oxford University Press,
New York.
Carey JR, Liedo P, Muller HG, Wang JL, Chiou JM,
1999. Mortality oscillations induced by periodic starva-
tion alter sex-mortality differentials in Mediterranean
fruit flies. J. Geront. Biol. Series 54A, B424–B431.
Carey JR, Liedo P, Muller H-G, Wang J-L, Love B, Harsh-
man L, Partridge L, 2001. Female sensitivity to diet
and irradiation treatments underlies sex-mortality
differentials in the Mediterranean fruit fly. J. Geront.
Biol. Series 56A, B89–B93.
Carey JR, Liedo P, Harshman L, Liu X, Muller H-G,
Partridge L, Wang J-L, 2002a. Food pulses increase
longevity and induce cyclical egg production in Medi-
terranean fruit flies. Funct. Ecol. 16, 313–325.
Carey JR, Liedo P, Harshman L, Zhang Y, Muller H-G,
Partridge L, Wang J-L, 2002b. Life history response of
Mediterranean fruit flies to dietary restriction. Aging
Cell 1, 140–148.
Carey JR, Harshman LG, Liedo P, Muller H-G, Wang J-L,
Zhang Z, 2008. Longevity-fertility trade-offs in the te-
phritid fruit fly Anastrepha ludens, across dietary-restric-
tion gradients. Aging Cell 7, 470–477.
Chang C, Kurashima R, Albrecht CP, 2001. Larval devel-
opment of Ceratitis capitata (Diptera: Tephritidae) on a
meridic diet. Ann. Entomol. Soc. Am. 94, 433–437.
Collins SR, Weldon CW, Banos C, Taylor PW, 2008.
Effects of irradiation dose rate on quality and sterility
of Queensland fruit flies, Bactrocera tryoni (Froggatt).
J. Appl. Entomol. 132, 398–405.
Cresoni-Pereira C, Zucoloto FS., 2001. Influence of quan-
tities of brewer yeast on the performance of Anastrepha
obliqua wild females (Diptera, Tephritidae). Iheringia,
Ser. Zool., Porto Alegre, 91, 53–60.
Cresoni-Pereira C, Zucoloto FS, 2006. Influence of male
nutritional conditions on the performance and alimen-
tary selection of wild females of Anastrepha obliqua
(Macquart) (Diptera: Tephritidae). Rev. Brasileira Ento-
mol. 50, 287–292.
Dyck VA, Hendrichs J, Robinson AS (Eds), 2005. Sterile
insect technique. Principles and practice in area-wide
integrated pest management. Springer, Dordrecht, The
Netherlands.
Epsky ND, Heath RR, 1993. Food availability and phero-
mone production by males of Anastrepha suspensa
(Diptera: Tephritidae). Environ. Entomol. 22, 942–
947.
Everitt BS, Pickles A, 2004. Statistical aspects of the
design and analysis of clinical trials (revised edition).
Imperial College Press, London, UK.
FAO/IAEA/USDA, 2003. Manual for product quality
control and shipping procedures for sterile mass-reared
Tephritid fruit flies. Version 5.0. International Atomic
Energy Agency. Vienna, Austria. 85 pp. http://
www.iaea.org/programmes/nafa/d4/index.html
Field SA, Yuval B, 1999. Nutritional status affects copula
duration in the Mediterranean fruit fly, Ceratitis capitata
(Insecta: Tephritidae). Ethol. Ecol. Evol. 11, 61–70.
Gavriel S, Gazit Y, Yuval B, 2009. Remating by female
Mediterranean fruit flies Ceratitis capitata, (Diptera:
Tephritidae): temporal patterns and modulation by
male condition. J. Insect Physiol. 55, 637–642.
Heath RR, Manukian A, 1992. Development and evalua-
tion of systems to collect volatile semiochemicals from
insects and plants using charcoal-infused medium for
air purification. J. Chem. Ecol. 18, 1209–1226.
Hendrichs J, Robinson AS, Cayol JP, Enkerlin W., 2002.
Medfly areawide sterile insect technique programmes
for prevention, suppression or eradication: the impor-
tance of mating behavior studies. Fla. Entomol. 85,
1–13.
Jacome I, Aluja M, Liedo P, 1999. Impact of adult diet on
demographic and population parameters of the tropical
fruit fly Anastrepha serpentina (Diptera: Tephritidae).
Bull. Entomol. Res. 89, 165–175.
Kaspi R, Yuval B, 2000. Post-teneral protein feeding
improves sexual competitiveness but reduces longevity
of mass reared sterile male Mediterranean fruit flies.
Ann. Entomol. Soc. Am. 93, 949–955.
Kaspi R, Taylor P W, Yuval B, 2000. Diet and size influ-
ence sexual advertisement and copulatory success of
males in Mediterranean fruit fly leks. Ecol. Entomol.
25, 279–284.
Knipling EF, 1955. Possibilities of insect control or eradi-
cation through the use of sexually sterile males. J.
Econ. Entomol. 48, 459–462.
Knipling EF, 1959. Sterile-male method of population
control. Science 130, 902–904.
Landolt PJ, Sivinski J, 1992. Effects of time of day, adult
food and host fruit on incidence of calling by male
Caribbean fruit flies (Diptera: Tephritidae). Environ.
Entomol. 21, 382–387.
Liedo P, Salgado S, Oropeza A, Toledo J, 2007. Improving
mating performance of mass-reared sterile Mediterra-
nean fruit flies (Diptera: Tephritidae) through changes
in adult holding conditions: demography and mating
competitiveness. Fla. Entomol. 90, 33–40.
Pre-release feeding on sterile Anastrepha male performance P. Liedo et al.
58 J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH

Manrakhan A, Lux SA, 2006. Contribution of natural
food sources to reproductive behaviour, fecundity and
longevity of Ceratitis cosyra, C. fasciventris and C. capitata
(Diptera: Tephritidae). Bull. Entomol. Res. 96, 259–
268.
Maor M, Kamensky B, Shloush S, Yuval B, 2004. Effects
of post-teneral diet on foraging success of sterile mal
Mediterranean fruit flies. Entomol. Exp. Appl. 10,
225–230.
Meza JS, Dıaz-Fleischer F, Orozco D, 2005. Pupariation
time as a source of variability in mating performance
in mass reared Anastrepha ludens (Diptera: Tephritidae).
J. Econ. Entomol. 98, 1930–1936.
Muller H-G, Carey JR, Wu D, Liedo P, Vaupel JW, 2001.
Reproductive potential predicts longevity of female
Mediterranean fruit flies. Proc. R. Soc. B Biol. Sci. B
268, 445–450.
Nestel D, Papadopoulos NT, Liedo P, Gonzalez-Ceron L,
Carey JR, 2005. Trends in lipid and protein contents
during medfly aging: an harmonic path to death. Arch.
Insect Biochem. Physiol. 60, 130–139.
Ohinata K, Ashraf M, Harris EJ, 1977. Mediterranean
fruit flies: sterility and sexual competitiveness in the
laboratory after treatment with gamma irradiation in
air, carbon dioxide, helium, nitrogen or partial
vacuum. J. Econ. Entomol. 70, 165–168.
Orozco D, Domınguez J, Reyes J, Villasenor A, Gutierrez
JM, 2004. SIT and biological control of Anastrepha fruit
flies in Mexico. In: Proceedings of the 6th International
Symposium on Fruit Flies of Economic Importance,
Stellenbosch, South Africa, 6–10 May, 2002. Ed. by
Barnes B, Isteg Scientific Publications, Stellenbosch,
245–249.
Papadopoulos NT, Katsoyannos BI, Kouloussis NA,
Economopoulos AP, Carey JR, 1998. Effect of adult
age, food, and time of day on sexual calling incidence
of wild and mass-reared Ceratitis capitata males. Ento-
mol. Exp. Appl. 89, 175–182.
Papadopulous NT, Shelly TE, Niyazi N, Jang E, 2006.
Olfactory and behavioral mechanisms underlying
enhanced mating competitiveness following exposure
to ginger root oil or orange oil in males of the Mediter-
ranean fruit fly Ceratitis capitata (Diptera: Tephritidae).
J. Insect Behav. 19, 403–418.
Perez-Staples D, Prabhu V, Taylor PW, 2007. Post-teneral
protein feeding enhances sexual performance of
Queensland fruit flies. Physiol. Entomol. 32, 225–232.
Perez-Staples D, Harmer AMT, Collins SR, Taylor PW,
2008a. Potential for pre-release diet supplements to
increase sexual performance and longevity of male
Queensland fruit flies. Agic. Forest Entomol. 10, 255–
262.
Perez-Staples D, Aluja M, Macıas-Ordonez R, Sivinski J,
2008b. Reproductive trade-offs from mating with a
successful male: the case of the tephritid fly Anastrepha
obliqua. Behav. Ecol. Sociobiol. 62, 1333–1340.
Perez-Staples D, Weldon CW, Smallridge C, Taylor PW,
2009. Pre-release feeding on yeast hydrolysate
enhances sexual competitiveness of sterile male
Queensland fruit flies in field cages. Entomol. Exp.
Appl. 131, 159–166.
Prabhu V, Perez-Staples D, Taylor PW, 2008. Protein: car-
bohydrate ratios promoting sexual activity and longev-
ity of male Queensland fruit flies. J. Appl. Entomol.
132, 575–582.
Resilva S, Obra G, Zamora N, Gaitan E, 2007. Develop-
ment of quality control procedures for mass produced
and released Bactrocera philippinensis (Diptera: Tephriti-
dae) for sterile insect technique programs. Fla. Ento-
mol. 90, 58–63.
Rull J, Diaz-Fleischer F, Arredondo J, 2007. Irradiation of
Anastrepha ludens (Diptera: Tephritidae) revisited: opti-
mizing sterility induction. J. Econ. Entomol. 100,
1153–1159.
Shelly TE, Edu J, 2007. Exposure to the ripe fruit of
tropical almond enhances the mating success of male
Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol.
90, 757–758.
Shelly TE, Kennelly S, 2002. Influence of male diet on
male mating success and longevity and female remating
in the Mediterranean fruit fly (Diptera: Tephritidae)
under laboratory conditions. Fla. Entomol. 85, 572–579.
Shelly TE, McInnis DO, 2001. Exposure to ginger root oil
enhances mating success of irradiated mass-reared
males of the Mediterranean fruit fly (Diptera: Tephriti-
dae). J. Econ. Entomol. 94, 1413–1418.
Shelly TE, McInnis DO, 2003. Influence of adult diet on
the mating success and survival of male Mediterranean
fruit flies (Diptera: Tephritidae) from two mass-rearing
strains on field-caged host trees. Fla. Entomol. 86,
340–344.
Shelly TE, Resilva S, Reyes M, Bignayan H, 1996. Methyl
eugenol and mating competitiveness of irradiated male
Bactrocera phillipinensis (Diptera: Tephritidae). Fla. Ento-
mol. 79, 482–488.
Shelly TE, Kennelly SS, McInnis DO, 2002a. Effect of
adult diet on signaling activity, mate attraction, and
mating success in male Mediterranean fruit flies
(Diptera: Tephritidae). Fla. Entomol. 85, 150–155.
Shelly TE, Robinson AS, Caceres C, Wornoayporn V,
Islam A, 2002b. Exposure to ginger root oil enhances
mating success of male Mediterranean fruit flies
(Diptera: Tephritidae) from a genetic sexing strain. Fla.
Entomol. 85, 440–445.
Shelly TE, Edu J, Pahio E, 2005. Influence of diet and
methyl eugenol on the mating success of males of the
Oriental fruit fly, Bactrocera dorsalis (Diptera: Tephriti-
dae). Fla. Entomol. 88, 307–313.
P. Liedo et al. Pre-release feeding on sterile Anastrepha male performance
J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH 59

Shelly TE, Edu J, Pahio E, 2006. Dietary protein and
mating competitiveness of sterile males of the Mediter-
ranean fruit fly (Diptera: Tephritidae): measurements
of induced egg sterility in large field enclosures. Fla.
Entomol. 89, 277–278.
Shelly TE, McInnis DO, Rodd C, Edu J, Pahio E, 2007.
Sterile insect technique and Mediterranean fruit fly
(Diptera: Tephritidae). Assessing the utility of aroma-
therapy in a Hawaiian coffee field. J. Econ. Entomol.
100, 273–282.
Sivisnki J, Calkins C, Baranowski R, 1994. A container
for eclosion and holding adult insects prior to mass
release. Fla. Entomol. 77, 513–515.
Taylor PW, Yuval B, 1999. Postcopulatory sexual selec-
tion in Mediterranean fruit flies: advantages for large
and protein-fed males. Anim. Behav. 58, 247–254.
Toledo J, Rull J, Oropeza A, Hernandez E, Liedo P, 2004.
Irradiation of Anastrepha obliqua (Diptera: Tephritidae)
revisited: Optimizing sterility induction. J. Econ. Ento-
mol. 97, 383–389.
Warburg MS, Yuval B, 1996. Effects of diet and activity
on lipid levels of adult Mediterranean fruit flies. Phys-
iol. Entomol. 21, 151–158.
Yuval B, Kaspi R, Shloush S, Warburg MS, 1998.
Nutritional reserves regulate male participation in
Mediterranean fruit fly leks. Ecol. Entomol. 23, 211–
215.
Yuval B, Kaspi R, Field SA, Blay S, Taylor PW, 2002.
Effects of post-teneral nutrition on reproductive success
of male Mediterranean fruit flies (Diptera: Tephritidae).
Fla. Entomol. 85, 165–170.
Yuval B, Maor M, Levy K, Kaspi R, Taylor P, Shelly T,
2007. Breakfast of champions or kiss of death? Survival
and sexual performance of protein-fed, sterile Mediter-
ranean fruit flies (Diptera: Tephritidae) Fla. Entomol.
90, 115–122.
Pre-release feeding on sterile Anastrepha male performance P. Liedo et al.
60 J. Appl. Entomol. 137(Suppl. 1) (2013) 49–60 ª 2010 Blackwell Verlag, GmbH
Related Documents











