Prostaglandins Leukotriencs and Medicine 17: 43-54, 1985 EFFECT OF PGF-2a ON PROGESTERONE PRODUCTION IN SWINE LUTEAL CELLS AT DIFFERENT STAGES OF THE LUTEAL PHASE. M.Mattioli: G.Galeati, A.Prandi, E.Seren. Istituto di Fisiologia Veteri- naria, Universita di Bologna, Via Belmeloro, 8/2: 40126 Bologna, Italy. (reprint request to M.M.). ABSTRACT Suspensions of luteal cells were prepared by enzymatic dispersion of pig corpora lutea obtained at specific times during the estrous cycle. Luteal cells from early corpora lutea produced more progesterone (4.73 + 0.84 nmol/106 cells: day 3) than those from late diestrus (0.73 2 0.04 nmol/106 cells, day 15); (PC 0.05). Bovine LH enhanced progestero- ne production in a dose dependent manner particularly in cells from 9 to 1.5 day corpora lutea. Also PGF-2a enhanced progesterone output in cells from mid-late corpora lutea. PGF-2a did not exert any antigonado- tropic effect since it further increased the progesterone production induced by LH. Luteal cells produced PGF-2a with levels ranging between 1.6 and 2.7 pmol/106 cells throughout the whole luteal phase. The cellular content of CAMP was markedly increased by LH (556 2 60%) while it was not affected by PGF-2a. Plasma membrane receptors for PGF-2a were not detected in the analyzed tissue. INTRODUCTION There is considerable evidence to support the hypothesis that uterine PGF-2a is the luteolytic factor in the pig (1,2). However, at present: it is not known how this factor induces luteal regression. Two main hypotheses.have been advanced to explain the luteolytic action of PGF- 2a : 1) reduction of ovarian blood flow as observed in the rabbit (3) and in the sheep (4:5) after PGF-2a administration; 2) antagonism of the steroidogenic effect of LH as described in vitro in luteal tissues of sheep (6), guinea pigs (7) and rats (8,9). The finding of specific plasma membrane receptors for PGF-2a in the corpus luteum of a number 43

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Prostaglandins Leukotriencs and Medicine 17: 43-54, 1985
EFFECT OF PGF-2a ON PROGESTERONE PRODUCTION IN SWINE LUTEAL CELLS AT DIFFERENT STAGES OF THE LUTEAL PHASE.
M.Mattioli: G.Galeati, A.Prandi, E.Seren. Istituto di Fisiologia Veteri- naria, Universita di Bologna, Via Belmeloro, 8/2: 40126 Bologna, Italy. (reprint request to M.M.).
ABSTRACT
Suspensions of luteal cells were prepared by enzymatic dispersion of pig corpora lutea obtained at specific times during the estrous cycle. Luteal cells from early corpora lutea produced more progesterone (4.73 + 0.84 nmol/106 cells: day 3) than those from late diestrus (0.73 2 0.04 nmol/106 cells, day 15); (PC 0.05). Bovine LH enhanced progestero- ne production in a dose dependent manner particularly in cells from 9 to 1.5 day corpora lutea. Also PGF-2a enhanced progesterone output in cells from mid-late corpora lutea. PGF-2a did not exert any antigonado- tropic effect since it further increased the progesterone production induced by LH. Luteal cells produced PGF-2a with levels ranging between 1.6 and 2.7 pmol/106 cells throughout the whole luteal phase. The cellular content of CAMP was markedly increased by LH (556 2 60%) while it was not affected by PGF-2a. Plasma membrane receptors for PGF-2a were not detected in the analyzed tissue.
INTRODUCTION
There is considerable evidence to support the hypothesis that uterine PGF-2a is the luteolytic factor in the pig (1,2). However, at present: it is not known how this factor induces luteal regression. Two main hypotheses.have been advanced to explain the luteolytic action of PGF- 2a : 1) reduction of ovarian blood flow as observed in the rabbit (3) and in the sheep (4:5) after PGF-2a administration; 2) antagonism of the steroidogenic effect of LH as described in vitro in luteal tissues of sheep (6), guinea pigs (7) and rats (8,9). The finding of specific plasma membrane receptors for PGF-2a in the corpus luteum of a number
43

of species such as the cow (IO.1 I .I21 and the hol,\kt ( 1,:. IJ. 12) >3tlppot’1 \ the hypothesis that PI:&& acts ;rt thz zol‘pus Iutt‘llm Iev(>l . lo L\~hl‘
knowledge PGF-2a receptors h,l vc Ilot tlec?ll descr‘hcd Ill sl\‘Ln<~ \‘irl’/k~l’,l
lutea. Another unclear ;lspcct of the f’GF-2a-itidu~~~~d I uteolyxl:, i \r 1 hL> resistance in vivo of early corpor,l lute;c to thi, 1101~r11orit. 1x5 i yt,int.k’ which is particularly protracted in the pir whercl I’GI”-2a admin i \.t 1.~1 t iL>n can induce luteolysis only after day 12 of the cst[‘<>us cycle (l,.Io). The present experiment was designed to evaluate the effect of IVY-2a on pi6 luteal cells obtained from corpora lutea at different staq:c‘x of the estrous cycle and to investigate if the sensitivity of pir corpux Iu- teum observed in vivo on-ly after day 12 of the cyc I e is the rexu I t of the appearance of receptors for this hormone around this period.
MATERIALS AND METHODS
Chemicals. Medium 199 with Hank’s salt. 2 5 mM Hepes and Hank ’ s ha I nnced salt solution (HBSS) were purchased from Gibco. Kanamicina and Funqlzo- ne obtained from Serva were added to the medium before use at a final concentration of 70 mg/l and 2 mg/l respectively. ColI.agen;kse type 1. hyaluronidase type 11: DNAse and bean trypsin inhibitor were purchased from Sigma. Trypsin was obtained from Gibco. Indomethacin employed (Liometacen) was a product of Chiesi Farmaceutici. A purified prenarit- tion of bovine luteinizing hormone(bLH;tiIH-LH-B9) wrls used to stlmul,ite progesterone production. PGF-2a was a generous eift of Dr.J.I’lkr.iJpjohn CO-j progesterone was purchased from Sigma. Tritiated hormones. proFe- sterone (1 ,2,6,7,21 jH Progesterone 120 Ci,/mmol) and PGF-2a( 5.ti:x.O. I I. 12: 14: 15 38 PGF-2a 150 Ci/mmol) and cyclic AMP RIA kits were purchased from NEN(New England Nuclear) .Antisera used for the r.ndioilnmurloassay of progesterone and prostaglandin F-2a were raised in the rabbit as dcscri- bed by Motta et al.. 198.1 (17) and Tamanini et al. .l’)j() (18) respectlvcly.
Dissociation of luteal. tissue. Mature gilts (pure bred Large White). aged 12 to 15 months were used. The animals were housed in individual crates and estrus detection was carried out by using a mature boar. Three animals were slaughtered ,3.5Z7Z9~ll.l,3 ,lnd I5 days after estrus (day 0). Preliminary experiments were carried out on ovaries collected from 4 sows slaughtered on day l&l:! of the estrous cycle. The ovaries were obtained immediately after slaughter and transported to the lnbo- ratory where they were dissected out in a cooled bench at J°C. The in- terval between slaughter and ovaries removal never exceeded IO min. About 2 g of tissue were used to prepare dispersed cells and the re- maining tissue was treated to obtain crude plasma membranes. The disso- ciation of corpora lutea was carried out essentially as described by Gospodarowicz and Gospodarowicz. 1972 (19) . Corpora lutca were excized and cut into small pieces using two razor blades. The tissue was first
44

incubated in 15 ml of HBSS containing 0.1% collagenase and hyaluronida- se and 4% BSA for 30 min. at 37OC under continuous stirring. The tissue was then washed 3 times with HBSS and incubated in 15 ml of medium 199, 25 mM Hew, containing 0.125% trypsin. The medium was renewed every 1.5 min. After 4x1.5 min. incubations, when most of the tissue was reduced to small clumps, the dissociation was hastened by repeatedly drawing these clumps into a Pasteur pipette. The cells were collected by centrifugation and resuspended in medium containing 0.1% bean tryp- sin inhibitor. If a mucous like substance prevented resuspension of the cells it was dissolved by adding two drops of 0.04% DNAse solution/ml of cell suspension. Finally the cells were washed twice, resuspended in culture medium 199, 25 mM Hepes and filtered through four Sayers of cheesecloth to eliminate large particles. The viability of the disper- sed cells was about 80% as judged by trypan blue exclusion (20). Cel.Ls were routinely counted in a standard hemocytometer. The effect of the time of incubation was evaluated by incubating about 1.5 x 106 viable cells,obtained by pooling 4 different cell samp1es:i.n 3 ml medium 199, 25 mM Hepes. After a 30 min. preincubation period: an aliquot of medium (30 ~1) was removed (0 h) and LH or PGF-2a were added (30 ~1) to give a final concentration of 0.1 FM. Aliquots of medium (10 ~1) were then removed 15,.?0,60,120,180,240:300 min. after the addi- tion of the hormone. Control incubations were also run. There were 3 replicates of each treatment. The progesterone production rate was eva- luated from the slope of the plot representing progesterone accumula- tion after increasing time of incubation. Data from these preliminary experiments, reported in the result section, suggested to us to carry out all the subsequent incubations, consisting of 5x105 cells in 1 ml medium 199: 25 mM Hepes, in humidified atmosphere at 37OC for a period of 2 hours. The incubations were stopped by freezing the cell samples (-2OOC).
PGF-2a receptors. Crude plasma membranes were obtained immediately after slaughter as previously described (12). An aliquot of the plasma membrane preparation was used for protein determination as described by Lowry et al.:1951 (21) using BSA as stan- dard. Incubations of varying amounts of plasma membrane preparations (0.5-2.5 mg protein/tube) and tritiated PGF-2a (0.3-60 fmol/tube) were carried out as previously described (12) and run at room temperature or at 37OC for 2:$r8:16 hours. Excess unlabelled ligand was used to de- termine non specific binding.
Measurement of progesterone, prostaglandin F-2o and cyclic AMP. The con- centrations of progesterone were determined by KIA. The antiserum was raised in rabbits to lla-OH progesterone-hemisuccinate-BSA and used at a working dilution of 1:15,000. Cross reactions % were: llo hydroxypro-
45

gesteronc Y.l..j; 11~ hydroxyprogesteront 15.7; 21 tlydroxyproSe~terorlr 4.Oj IylX hydroxyprogesteronr I.;: 20~1 hydroxyproee<terone < 0.1. The antigen was used at 80.000 d.p.m .;‘ml. The samples of medium were clilu- ted I:100 (v:v) with 0.05 N PRS-EDTA and 0.1 ml ,lliyuot was incubated overnight at 4OC with 0.1 ml antiserum and 0.1 ml antigen. Separation of free from bound hormone was obtained by adding 1 ml O..$ dextran coated charcoal and by centrifugation at 3200 g for 15 min. at j°C. The sensitivity of the assay was 6 pg/tuhe (the sensitivity is expressed as the amount of steroid which gives a 10% inhibition of the binding occurring in absence of unlabelled ligand). The intra and inter assay precision, expressed as the coefficient of variation for replicate (10) determinations of a pool of medium samples was 4.1 and 7..$ re- spectively. PGF-2a assay, conducted on 0.5 ml of medium: was carried out by using a specific antiserum as described by Chiesa et al..1979 (22). The sen- sitivity of the assay was of 2 pg/tube. The intra assay and inter as- say precision was 1.3.6 and 10.8% respectively (values based on 10 de- terminations). The measurement of cyclic AMP was carried out on 0.2 ml of cell suspen- sion. The samples were cooled in ice and sonicated for JO sec. Cyclic AMP was extracted by boiling the sample for 3 min. and removing the precipitate by centrifugation. Cyclic AMP was determined on 100 ~1 of supernatant diluted 1:lO with 0.05 M acetate buffer (pH 6.2) by using CAMP RIA kits. The acetylnted system was used. The assay was sensitive to 0.005 pmol/tube with minimal cross reactivity to other nucleotides (AMP: ATP, GMP <O.OOOOj%).The intra and inter assay precision was 3.2 and 7.7% respectively (IO determinations). The production of progesterone: PGF-2~r and CAMP was calculated follo- wing correction for the concentrations of these substances obtained for non incubated samples of cell suspensions.
KESULTS
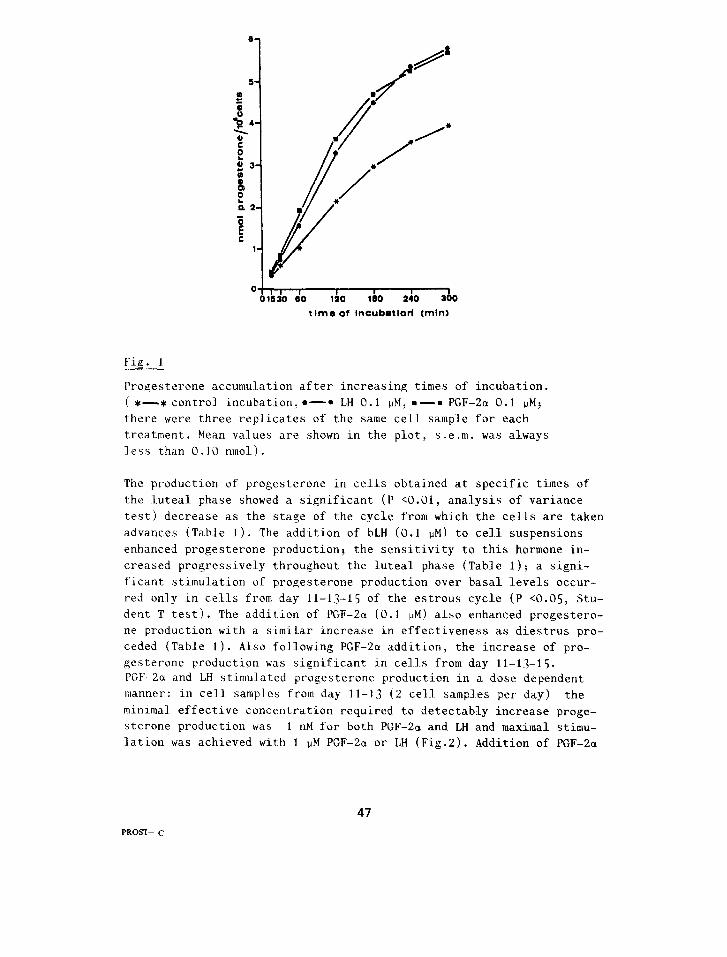
Progesterone production. Experiments with cells obtained from day lo- 12 of tne estrous cycle indicated that Juteal cells in our culture sy- stem produced progesterone at a constant rate(16.8 pmoI/min/IG'ceI Is) from the beginning of the experiment up to 120 min.. then this produ- tion progressively decreased(5.4 pmol/min/lO6 cells after 240 min. of culture). When LH or PGF-2cr (0.1 PM) were added to the culture medium an increased rate of progesterone production occurred(23.2 and 26.8pmol/ min./lo6 cells respectively for LH and PGF-2a ). This increased rate of progesterone production was constant during the first 120 min. of incubation then it progressively decreased to levels similar to those of control after 240 min. of incuhation(7.0 and 5.4pmol/min/i06 cells for LH and PGF-2a respectively)(Fig. I ).
46
o!,, , I 1 I
01630 60 120 180 240 2 0 time of Inoubatlod (mlnl
Fig. I
Progesterone accumulation after increasing times of incubation. (*-_* control incubation,*-* LH 0.1 PM: .-= PGF-2a 0.1 ~Mj there were three replicates of the same cell sample for each treatment. Mean values are shown in the plot, s.e.m. was always less than 0.10 nmol).
The production of progesterone in cells obtained at specific times of the Suteal phase showed a significant (P (0.01, analysis of variance test) decrease as the stage of the cycle from which the cells are taken advances (Table I). The addition of bLH (0.1 PM) to cell suspensions enhanced progesterone production: the sensitivity to this hormone in- creased progressively throughout the luteal. phase (Table 1); a signi- ficant stimulation of progesterone production over basal levels occur- red only in cells from day 11-13-15 of the estrous cycle (P ~0.05: Stu- dent T test). The addition of PGF-2a (0.1 PM) also enhanced progestero- ne production with a similar increase in effectiveness as diestrus pro- ceded (Table 1). Also following PGF-2a addition, the increase of pro- gesterone production was significant in cell-s from day 11-13-15. PGF-2a and LH stimulated progesterone production in a dose dependent manner: in cell samples from day 11-13 (2 cell samples per day) the minlmal effective concentration required to detectably increase proge- sterone production was 1 nM for both PGF-2a and LH and maximal stimu- lation was achieved with 1 PM PGF-2a or LH (Fig.2). Addition of PGF-2a
47 PROSTm- c
IO.1 PM) to the medium further incrc;L\ed the production of proersteron~~ ~nduccd by LH (0.1 PM). however when nraxitttal rffcctivc dose\ of thex~ hormones wcrc used together no additive effect was obxcrvcd.
Table 1 Progesterone production, PGF-2a production and effect of LH or PGI’-2c( (0.1 1M) on progesterone production in swine luteal ceJls at different stages of the luteal phase (mean !. sem of 3 cell samples per day).
day of the ___ Progesterone nmol/lOCI cells estrous cycle control LH PGF-2a
PCF-2; pmol/lO cells
.1 4.7.3 + 0.84 5.01 i 1.01 4.9.3 2 0.96 1.59 + 0.18
5 2.63 7 0.25 .j.29 ? 0.26 ,j.,JO-+O.,@ l.r;otO.26
7 2.63 i 0.43 2.86 L 0.53 2.88 5 0.59 1.74 i 0.10
9 2.04 + 0.22 3.0.? I 0.49 2.91 i 0.50 2.13 : O.,J8
11 1.82 IO.18 2.76 + 0.28 3.22 * 0.33 2.27 + 0.07
1 3 0.94 + 0.14 1 .7.3 i 0.18 2.10 + 0.14 2.64 : 0.36
15 0.7.3 _f 0.04 I .40 i 0.04 I .59 f 0.09 I .8,? + 0.26
PGF-2a production. Luteal cells produced PGF-2cr with levels ranging between 1.6 and 2.6 pmol/106 cells depending on the day of the estrous cycle (Table J 1. No significant changes occurred throughout the luteal phase (P>O.O5, analysis of variance test). Regardless of the stage of the cycle bLH did not affect PGF-2c( production. When indomethacin (JpM) was used an over 80% inhibition of PGF-2a synthesis occurred. Moreover indotnethacin induced a constant, although non statistically significant, inhibition of progesterone production of about 20% which could be re- versed by the addition of 2 pmol PGF-2a /ml of medium. The inhibition of the synthesis of PGF-2~ did not affect LH stimulated progesterone production. A representative example of the effect of indomethacin and LH is given in Table 2.
Table 2 Effect of indomethacin and LH on the production of progesterone and PGF-213 in ,I cell samples obtained from day I .j of the estrous cycle.
control IH indcnlethacin itYdanetkKin indanetacin (0.1l.N) (3i.N -t H +2 pnol. E&
progesterone 0.91 b 0.14 1.67 * 0.17 0.69 + 0.04 1.61 * 0.08 0.89 i 0.10 rsnol/lo6 ce1.l.s PGF-2a 2.64 + 0.36 2.75 + 0.24 0.50 + 0.05 0.47 i 0.07 2.53 2 0.21 pno7/106 cells
48
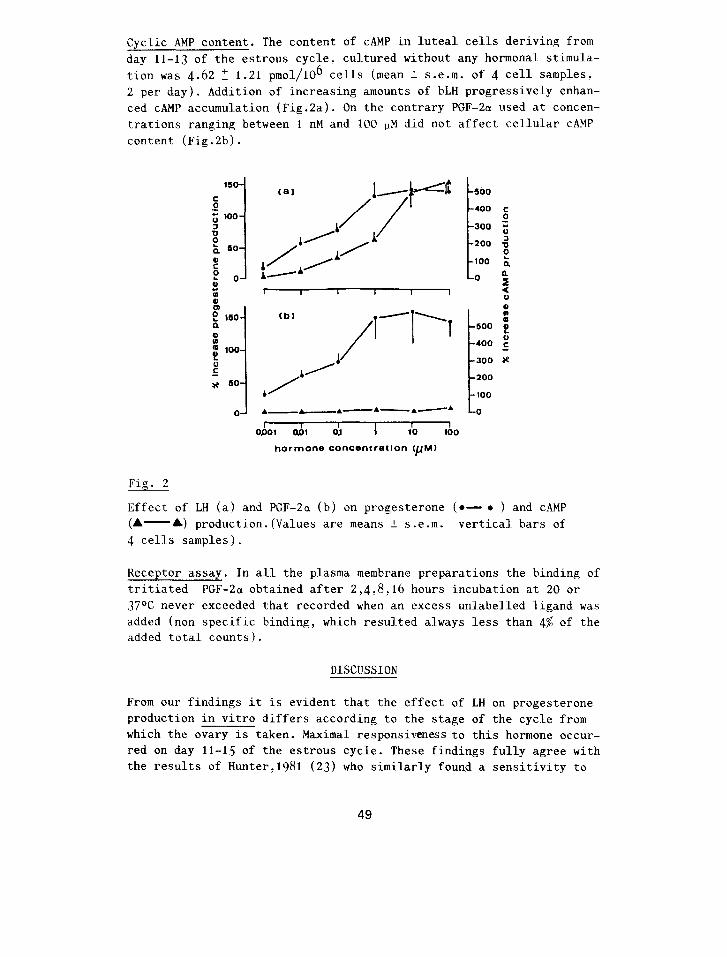
Cyclic AMP content. The content of CAMP in luteal cells deriving from day 11-13 of the estrous cycle, cultured without any hormonal stimula- tion was 4.62 T 1.21 pmol/1& cells (mean + s.e.m. of 4 cell samples: 2 per day). Addition of increasing amounts of bLH progressively enhan- ced CAMP accumulation (Fig.2a). On the contrary PGF-2a used at concen- trations ranging between 1 nM and 100 11M did not affect cellular CAMP content (Fig.2b).
(bl
1 500
400 0’
300 i
200 : f 100 0
O “I * 0
iz 500 ;
400 :
I
300 ae
200
100
0
OPOl oD1 0.l 1 10 too
hormone COI’ICentretlOn CpM)
Fig. 2
Effect of LH (a) and PGF-2a (b) on progesterone (e- ?? ) and CAMP (A- A) production.(Values are means 2 s.e.m. vertical bars of 4 cells samples).
Heceptor assay. In all the plasma membrane preparations the binding of tritiated PGF-2a obtained after 2,4:8:16 hours incubation at 20 or 37OC never exceeded that recorded when an excess unlabelled ligand was added (non specific binding, which resulted always less than 4% of the added total counts).
DISCUSSION
From our findings it is evident that the effect of LH on progesterone production in vitro differs according to the stage of the cycle from which the ovary is taken. Maximal responsiveness to this hormone occur- red on day 11-15 of the estrous cycle. These findings fully agree with the results of Hunter,1981 (23) who similarly found a sensitivity to
49

LH higher on day 12. 5 than on day 0. j in cul tut-cd swine I Lltc,t I \‘L> I I\ and those of du Mesnil du Buisson and Lcelise. I UO,i (21 1 a11d :\tldzrxon et al.: 1907 (25) reporting that in vivo pir cot-pus I utc‘um ilo~‘b not ii’txm to need any gonadotropic support before day I 0 0 t‘ the 62 h t r ou h c,v I‘ I t‘ . Therefore the progressive reduction of progesterone secretion ohser-vcd in cells taken from early to late corpora 1 utea. suggests that the l‘Cltt of steroidogenesis proceeds in an automatic way in newly formed corpol’a lutea while it becomes more and more LH dependent as dirstrus proceeds. Our experiments showed that PGF-2a stimulated the product ion of proetx- sterone by dispersed swine luteal cells in a doxe dependent Imanner. This stimulating effect is: in some ways: similar to that of LH. f3oth PGF-2a and LH: in fact. could significantJy enhanc: the production of progesterone only in cells deriving from day II-lj. they both showed the same dose dependent effect and their effect had the same kinetics. Despite this: the mechanism through which PGF-2~t stimulates the produ- ction of progesterone seemed totaLly different from that of LH. In fact we did not find any detectable amounts of membrane receptors for PGF-2a and : in agreement with this finding. the increased production of pro- gesterone induced by this hormone was not mediated via a CAMP accumula- tion which is recognized as the signal of the hormone-membrane receptor interaction for a variety of hormones among which is LH. A progesterone secretion not regulated via a CAMP dependent mechanism has however a- ready heen described in sheep large luteal eel 1 s (26). To explain the similar stimulating effect of LH and PGF-2a it could he hypothesized that PGF-2a is a messenger of LH acting at a site distal to CAMP for- mation, hut this was not confirmed hy our experiments where LH did not influence PGF-2a production and indomethacin. i.e. PGF-2a withdra- wal, did not affect LH induced progesterone production. Further studies are requested to elucidate the mechanism of action of PGF-2a In vitro. In agreement with our findings the stimulating effect of PGF-213 has also been described in human (27 1 and in bovine (28) dispersed luteal cells. On the contrary Watson et al.. 1977 (29) reported that PGF-2a re- duces progesterone secretion by porcine luteal slices.although an ini- tial stimulation may he seen particularly at higher doses of PGF-2a (1-j pg/ml). These authors however could obtain rj complete inhibition of progesterone production only by using endometrlum super-fusate thus indicating that endometrial factors. other than PGF-2a .may he invo I ved in lutes1 regression. Recent experiments carried out in vitro on acute- I.y cultured luteal tissue of guinea pigs (7): rats (3.9) and sheep (6). suggest the hypothesis that the luteolytic effect of PGF-2a is not due to an inhibition of hasaL levels of progesterone production but to an anti-gonadotropic effect of this hormone which could in fact prevent the LH-induced increase of progesterone production. This was not confir med in swine l.uteal cells where PGF-2a further increased the levels of progesterone production induced by LH. In the experiments in vivo so far
50

reported administration of PGF-2a after day 12 of the estrous cycle in the pig exertsa luteolytic effect. In particular Kotwica, 1980 130) found that the infusion of PGF-2a into the anterior uterine vein of sows, after day 12 of the cycle: significantly depressed the progeste- rone plasma levels within 90 min. of the beginning of the infusion. Si- milar results were reported by Krzymowski et al.,1978 (?I). Two main hypotheses may be advanced in order to explain the different effects of PGF-2a observed in vitro and in vivo: A) PGF-2a does not exert its luteolytic effect at the luteal tissue level but on the ova- rian vasculature. Thanks to its vasoconstrictive properties it might reduce ovarian blood flow resulting in a limited luteal. steroidogenetic activity. A marked reduction of ovarian blood flow around the time of I.uteolysis has recently been described in the sow (.32:33) and this re- duced blood flow was strictly correlated with the drop of plasma pro- gesterone levels occurring at luteolysis (3.1). Such an effect of PGF-2a on the ovarian blood flow has been described in the sheep by Nett & Niswender, 1981,(5); B) PGF-2a acts directly at the luteal tissue level.: in this case it must he supposed that for some reason dispersed -Luteal cells behaviour does not reflect the situation in vivo. The presence of the whole tissue with consequent cell to cell interactionorthe presen- ce of factors missed in our in vitro system might be necessary for the luteolytic effect of PGF-2n to occur. Patek & Watson, 1976 (34): have proposed that luteal PGF-2a synthesis might be involved in the process of luteolysis. They reported that as well as endometrial also luteal PGF-2a release in vitro was highest in tissues collected during the late luteal phase. On the contrary we did not find any significant change in PGF-2a release throughout the whole Luteal phase in agreement with the results of Guthrie & Rexroad: (35). Therefore, apart from uterine PGF-213 ,secreted mainly around day 14-18 of the estrous cycle and generally accepted as a factor involved in the luteolysis in the pig, we should consider that the corpus luteum itself is capable,at least in vitro, of synthesizing PGF-2a throughout the whole luteal phase. If the results in vitro reflect the situation in viva; it could he suggested that PGF-2a: produced and acting inside the corpus luteurn: might represent an important local factor involved in the control of the luteal function.
Acknowledgments. We wish to thank the staff of the slaughterhouse of Bologna and Prof. A.Morselli for their assistence during the collection of the luteal tissue. This work was in part supported by Regione Emilia Romagna.
REFERENCES
1. Horton EW & Poiser NL. Uterine luteolytic hormone - A physiological
51

role for prostaglandin P‘-2a. Physiol .liev. 50: 505. 1070. 2. Moleonio MPE. Razer FW R Tntchcr WW. A study of prostaglanditl F-“a
as the Luteolysin in swLne. I .Effects of prostaelandin I’-2n in hystc _ rectomized gil tx. I’rostagl andiris I 1 : 7.3;. 1970.
.i. Novg NJ & Cook MJ . Redistribution of‘ blood flow hy prostaglandin I;-2a in the rabbit ovary. Am..J.Ohstet.GynecoL. I I7:,iYl .107.3.
4. Nett TM: McClellan MC 8. Niswender GD. Effects of prostaglandins on the ovine corpus luteum: hlood flow secretion of progesterone and morphol.ogy. Biol .Keprod. I j:ih. 19ih.
5. Nett TM &: Niswender GD. Luteal blood flow and receptor for LH during PGF-2a-induced luteolysis : production of PGE-2a and PGF-2a during early pregnancy. Acta Vet.Scand. Suppl. 177: 117.1()81.
6. Fletcher PW & Niswender GD. Effect of PGF-2a on progesterone sccre- tion and adenylate cyclase activity in ovine luteal tissue. Prosta- glandins 2.3:80.?. 1982.
7. Richardson MC. Peddie MJ. Inhibitory effect of PGF-2a on hCG-stimu- lated progesterone production by luteal cel_ls from guinea pigs at different stages of the oestrus cycJe. J.Keprod.Fert.66:117.1982.
8. Lahav M: Freud A R Lindner HR. Ahrogation by prostaglandin F-2n of LH-stimulated cyclic AMP accumulation in isolated rat corpora lutea of pregnancy. Biochem.Hiophys.lies.Com. 68:J294,J976.
9. .Jordan AW. Effects of Prostaglandin F-2a treatment on LH and Dibuty- ryl cyclic AMP-stimulated progesterone secretion by isolated rat Lu- teal cells. Biol .&prod. 25:.327.IOhL.
10. Kitnball FA & Lauerdale .JW. Prostaglandin E-l and F-213 specific hin- ding in bovine corpora Lutea: comparison with IuteoLytic effect. Prostaglandins 10:.11.3.l975.
11. QowelL WS. Hammarstrom S & Samuelsson B. Occurrence and properties of PGF-2a receptor in bovine corpora lutea. Eur. J . Hiochem. 56: i.3.1975.
12. Mattioli M. tialeati G & Seren K. Characteristics of receptors for prostagl andin F-2a in bovine and equine corpora lutea. Prostagl andins Leukotr ienes and Medicine I I : 2 5(9: IO8.1.
1.3. ,\rtnhall FA. t,rostagland~n F-2a specific binding in equine corpora lutea. Prostaglandins 13: 55.1.1977.
14. Vernon MU. Strauss .J: Simonelli Z. Zavy MT & Sharp DC. Specific PGF-2a binding hy the corpus luteum of the pregnant and non pregnant mare. J.Lieprod.Fert. 27:42l:l979.
15. Diehl JH & Day BN. Effect of prostaglandin F-2a on l~uteal function in swine. J.Anim.Sci. .19:.392:1974.
16. Guthrie HD & Polge C. Luteal function and oestrus in gilts treated with a synthetic anaLogue of PGF-2a (ICI 79,939) at various time during the estrous cycle. J.lieprod.Fert. 48:42,3:19$6.
17. Motta M: Bono G> Bolelii G & Seren E. Mathematical analysis antise- rum titre and affinity in rabbits injected with IlaOH-progesterone-
52

hemisuccinate-BSA. J.Steroid Biochem. 19:1769:1983. Ih. Tamanini C: Bono G & Mongiorgi S. Production of anti PGF-2cx and
anti PGE-2 antisera. Atti S.T.S.VET. 333160.1979. 19. Gospodarowicz D & Gospodarowicz F. A technique for the isolation
of bovine luteal cells and its application to metabolic studies of luteal cells in vitro. Endocrinology 90:1427.1972.
20. Tennant JR. Evaluation of the trypan blu techniques for determina- tion of cell viability. Transplantation 2:685,1964.
21. Lowry OH, Kosenbroueh NJ. Far AL & Kandall KJ. Protein measurement with the Folin phenol reagent.J.Biol.Chem. 19.3:265:1951.
22. Chiesa F‘: Seren E. Bono G, i;aiani K & Tamanini C. Plasma concentra- tions of PGF-2o in dairy cows at parturition. Atti Soc.It.Buiatria, I1:37.3.1979.
2.3. Hunter MG. Kesponsivness in vitro of porcine Luteal tissue recove- red at two stages of the luteal phase. J.Keprod.Fert. 63:471~1981.
24. du Mesnil du Buisson F & Leglise PC. Effect de T'hypophysectomie sur les corpus jaunes de la truie. C.K. Hebd.Seanch.Acad.Sci.D 257: 25j:lQh.j.
25. Anderson LL. Dyck GM, Mori H. Henricks DM & Melapy KM. Ovarian function in pigs following hypophyseal stalk transection or hypo- physectomy. Am.J.Physiol. 21:1188.1967.
26. Hoyer f'B> Fitz T.A. & Niswender GD. Progesterone secretion by large ovine luteal cells is not regulated :',5'-cyclic adenosine mono- phosphate. Biol.fieprod. Suppl.1~28:34~1983.
27. Kichardson MC: Masson GM. Progesterone production by dispersed cells from human corpus 1uteum:stimulation by gonadotrophins and prostaglandins F-2a; lack of response to adrenaline and isopre- naline. J.Endocr. 87:247:19&I.
28. Weston PG & Hixon JE. Effects of in vivo prostaglandin F-2a admini- stration on in vitro progesterone synthesis by bovine corpora lu- tea. Biol.Reprod. 22:259:1980.
20. Watson J & Felicity H Maule Walker. Effect of prostaglandin F-2a and uterine extracts on progesterone secretion in vitro by super- fused pig corpora lutea. J.Keprod.Fert. 51:393:1977.
.i0. Kotwlca J. Mechanism of prostaglandin F-2a penetration from the horn of the uterus to the ovaries in pigs. J.Keprod.Fert.59:227: 1980.
.Il. Krzymowski T, Kotwica J, Okrasa S: Doboszynska T & Ziecik A. Luteal function in sows after unilateral infusion of PGF-2o into the ante- rior uterine vein on different days of the oestrous cycle. J.Keprod Fert. 54:21,1978.
32. Ford SK: Keynold LP & Magness HK. Blood flow to the uterine and ovarian vascular beds of gilts during the estrous cycle or early pregnancy. Biol.Keprod. 27:878:1982.
53

.l.i . ~l,reness lilt. Chrixtetlson IiK R Ford SP. Ovarian blood f low throughout the estrus cycle dnd early pregnancy in sowb. HioL.lieprod. 28: li)W. 1 o’i,i .
,7j.. Pate& CE Py. Watson J. Prostaglandin F and progesterone secretion by porcine endometrium and corpus luteum in vitro. ProstagSandinh ____ 12:98.1y76.
.3 5. Guthrie HD & tiexroad CE Jr. Progesterone secretion and prostaglan- din E‘ release in vitro by endometrial and lutea tissue of cyclic pigs. J.Heprod.Fert. 60:157.1980.
54
Related Documents











