Journal of Experimental Botany, Vol. 44, No. 262, pp. 893-899, May 1993 Effect of Nitrogen Supply on the Photosynthetic Performance of Leaves from Coffee Plants Exposed to Bright Light M. ANTONIETA NUNES 1 ,3, J. D. COCHICHO RAMALH0 1 and M. ADELAIDE DIAS 2 1 Centro de Estudos de Produc;8,o e Tecnologia Agricolas, Instituto de Investigac;8,o Cientifica Tropical, Tapada da Ajuda, Apartado 3014, Lisboa, Portugal 2 Departamento de Fisi%gia Vegetal, Estac;8,o Agron6mica Nacional, 2780 Oeiras, Portugal Received 27 October 1992; Accepted 16 February 1993 ABSTRACT Although Coffea arabica L. grows naturally in shaded habitats, it can be cultivated under high light intensity, but not without severe photoinhibition mainly during the period of transfer from the nursery into the field. The present work examines some of the changes in the photosynthetic performance induced by exposure to high light and the possibility of using enhanced nitrogen levels to overcome photoinhibition. For that purpose, young plants of Coffea arabica L. (cv. Catuai) grown in a shaded greenhouse were treated with 0, 1 and 2 mrnol of nitrogen and 4 weeks later exposed to full solar irradiation, outside. Visible damage due to exposure to full sunlight appeared within 2 d in all plants, resulting in a reduced photosynthetic leaf area and drastic shedding of leaves in the unfertilized plants. These effects were considerably less in plants with the highest N dose. After 130 d of exposure, there was 100% mortality in plants receiving no extra nitrogen, compared with 30% in the plants treated with 2 mmol nitrogen. Photosynthesis rates, leaf conductance and transpiration presented minimum values after 4 d of light stress. Large changes in the photosynthetic capacity (measured at high CO 2 concentration and high light intensity), quantum efficiency and fluorescence yield (FvlF m ) indicate that net photosynthesis rate in the air had been reduced by both stomatal closure and by changes at the photochemical level. All indicators show that N-fertilized plants were less affected by photoinhibition. r Key words: Coffee plant, nitrogen, photoinhibition, photosynthesis. INTRODUCTION The coffee tree (Coffea arabica L.) grows naturally in shaded habitats and its leaves suffer severe photoinhibi- tion after exposure to high light intensities as would be expected from its structural and physiological character- istics, which are typical of shade plants (Rhizopoulou and Nunes, 1981). However, cultivation in full sun may lead to higher yields in the short term and this has encouraged research into technological and genetic improvements to overcome the problems resulting from exposure to high light. It is well known that exposure of leaves to light levels higher than can be used in photosynthesis may cause photoinhibition (Powles, 1984). This phenomenon is typically manifested as a sustained decline in the quantum yield of photosynthesis and a decrease in the ratio Fv/F m (variable fluorescence of the chlorophyll/ 3 To whom correspondence should be addressed. © Oxford University Press 1993 maximum fluorescence), and in most cases by decreases in PSII photochemistry and maximal capacity of photosynthesis (Ogren, 1988, 1991; Rosenquist et al., 1991). Nitrogen fertilization, applied in a broad range of doses, has positive effects on growth and may improve plant performance in suboptimal environmental condi- tions such as shortage of water (Shimshi, 1970; Bennett et aI., 1986), marginal temperature (Lawlor et aI., 1987) and high light (Osmond, 1983, 1987; Ferrar and Osmond, 1986). Studies by Fahl (1989) and Rijo et al. (1989) indicated that increased nitrogen supply might alleviate problems of photoinhibition in coffee leaves. Thus we have explored this possibility further and considered some of the physio- logical and biochemical changes involved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Experimental Botany, Vol. 44, No. 262, pp. 893-899, May 1993
Effect of Nitrogen Supply on the PhotosyntheticPerformance of Leaves from Coffee PlantsExposed to Bright Light
M. ANTONIETA NUNES 1,3, J. D. COCHICHO RAMALH0 1 and M. ADELAIDE DIAS 2
1 Centro de Estudos de Produc;8,o e Tecnologia Agricolas, Instituto de Investigac;8,o Cientifica Tropical, Tapada da Ajuda,Apartado 3014, Lisboa, Portugal
2 Departamento de Fisi%gia Vegetal, Estac;8,o Agron6mica Nacional, 2780 Oeiras, Portugal
Received 27 October 1992; Accepted 16 February 1993
ABSTRACTAlthough Coffea arabica L. grows naturally in shaded habitats, it can be cultivated under high light intensity, but not withoutsevere photoinhibition mainly during the period of transfer from the nursery into the field. The present work examines some ofthe changes in the photosynthetic performance induced by exposure to high light and the possibility of using enhanced nitrogenlevels to overcome photoinhibition. For that purpose, young plants of Coffea arabica L. (cv. Catuai) grown in a shadedgreenhouse were treated with 0, 1 and 2 mrnol of nitrogen and 4 weeks later exposed to full solar irradiation, outside.
Visible damage due to exposure to full sunlight appeared within 2 d in all plants, resulting in a reduced photosynthetic leaf areaand drastic shedding of leaves in the unfertilized plants. These effects were considerably less in plants with the highest N dose.After 130 d of exposure, there was 100% mortality in plants receiving no extra nitrogen, compared with 30% in the plants treatedwith 2 mmol nitrogen. Photosynthesis rates, leaf conductance and transpiration presented minimum values after 4 d of light stress.Large changes in the photosynthetic capacity (measured at high CO2 concentration and high light intensity), quantum efficiencyand fluorescence yield (FvlFm) indicate that net photosynthesis rate in the air had been reduced by both stomatal closure and bychanges at the photochemical level. All indicators show that N-fertilized plants were less affected by photoinhibition. r
Key words: Coffee plant, nitrogen, photoinhibition, photosynthesis.
INTRODUCTIONThe coffee tree (Coffea arabica L.) grows naturally inshaded habitats and its leaves suffer severe photoinhibition after exposure to high light intensities as would beexpected from its structural and physiological characteristics, which are typical of shade plants (Rhizopoulou andNunes, 1981). However, cultivation in full sun may leadto higher yields in the short term and this has encouragedresearch into technological and genetic improvements toovercome the problems resulting from exposure to highlight.
It is well known that exposure of leaves to light levelshigher than can be used in photosynthesis may causephotoinhibition (Powles, 1984). This phenomenon istypically manifested as a sustained decline in thequantum yield of photosynthesis and a decrease in theratio Fv/Fm (variable fluorescence of the chlorophyll/
3 To whom correspondence should be addressed.
© Oxford University Press 1993
maximum fluorescence), and in most cases by decreasesin PSII photochemistry and maximal capacity ofphotosynthesis (Ogren, 1988, 1991; Rosenquist et al.,1991).
Nitrogen fertilization, applied in a broad range ofdoses, has positive effects on growth and may improveplant performance in suboptimal environmental conditions such as shortage of water (Shimshi, 1970; Bennettet aI., 1986), marginal temperature (Lawlor et aI., 1987)and high light (Osmond, 1983, 1987; Ferrar andOsmond, 1986).
Studies by Fahl (1989) and Rijo et al. (1989) indicatedthat increased nitrogen supply might alleviate problemsof photoinhibition in coffee leaves. Thus we have exploredthis possibility further and considered some of the physiological and biochemical changes involved.
1201008060
Days
4020
400
cr 300E0
til 200Q)
ro10Q)
...J 100
RESULTS
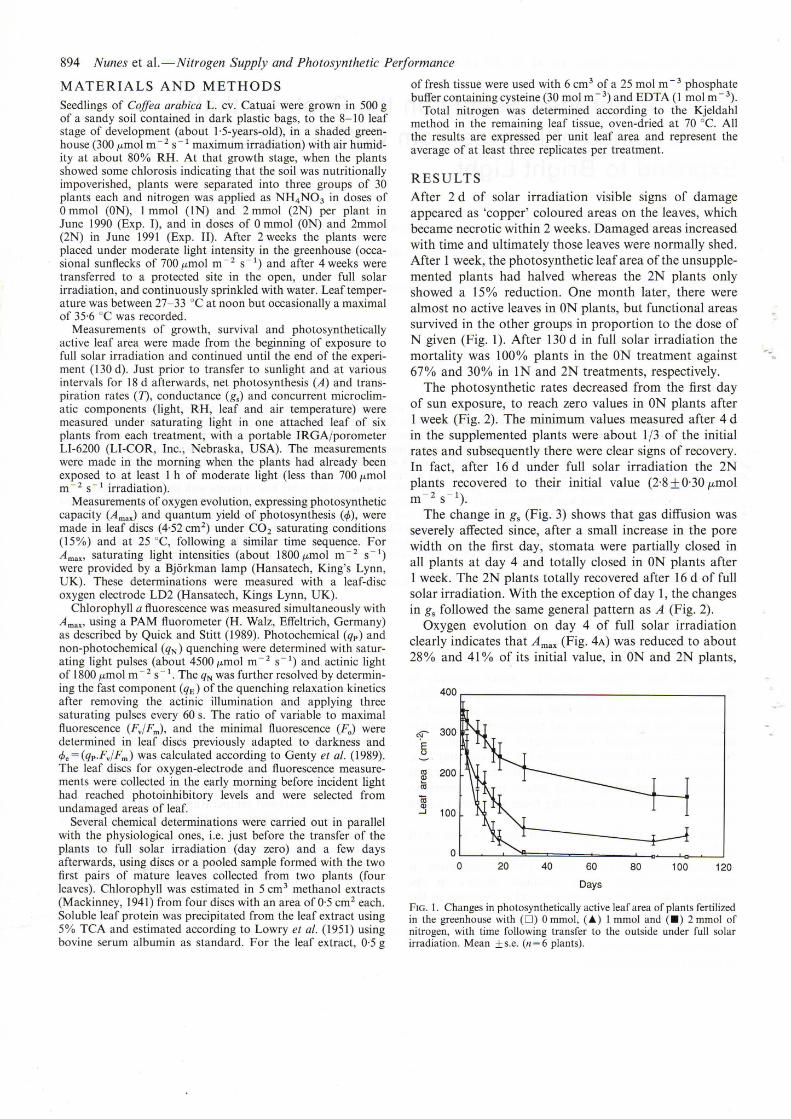
After 2 d of solar irradiation visible signs of damageappeared as 'copper' coloured areas on the leaves, whichbecame necrotic within 2 weeks. Damaged areas increasedwith time and ultimately those leaves were normally shed.After 1 week, the photosynthetic leaf area of the unsupplemented plants had halved whereas the 2N plants onlyshowed a 15% reduction. One month later, there werealmost no active leaves in ON plants, but functional areassurvived in the other groups in proportion to the dose ofN given (Fig. 1). After 130 d in full solar irradiation themortality was 100% plants in the ON treatment against67% and 30% in IN and 2N treatments, respectively.
The photosynthetic rates decreased from the first dayof sun exposure, to reach zero values in ON plants after1 week (Fig. 2). The minimum values measured after 4 din the supplemented plants were about 1/3 of the initialrates and subsequently there were clear signs of recovery.In fact, after 16 d under full solar irradiation the 2Nplants recovered to their initial value (2'8 ±O'30 /Lmolm- 2 S-l).
The change in gs (Fig. 3) shows that gas diffusion wasseverely affected since, after a small increase in the porewidth on the first day, stomata were partially closed inall plants at day 4 and totally closed in ON plants after1 week. The 2N plants totally recovered after 16 d of fullsolar irradiation. With the exception of day 1, the changesin gs followed the same general pattern as A (Fig. 2).
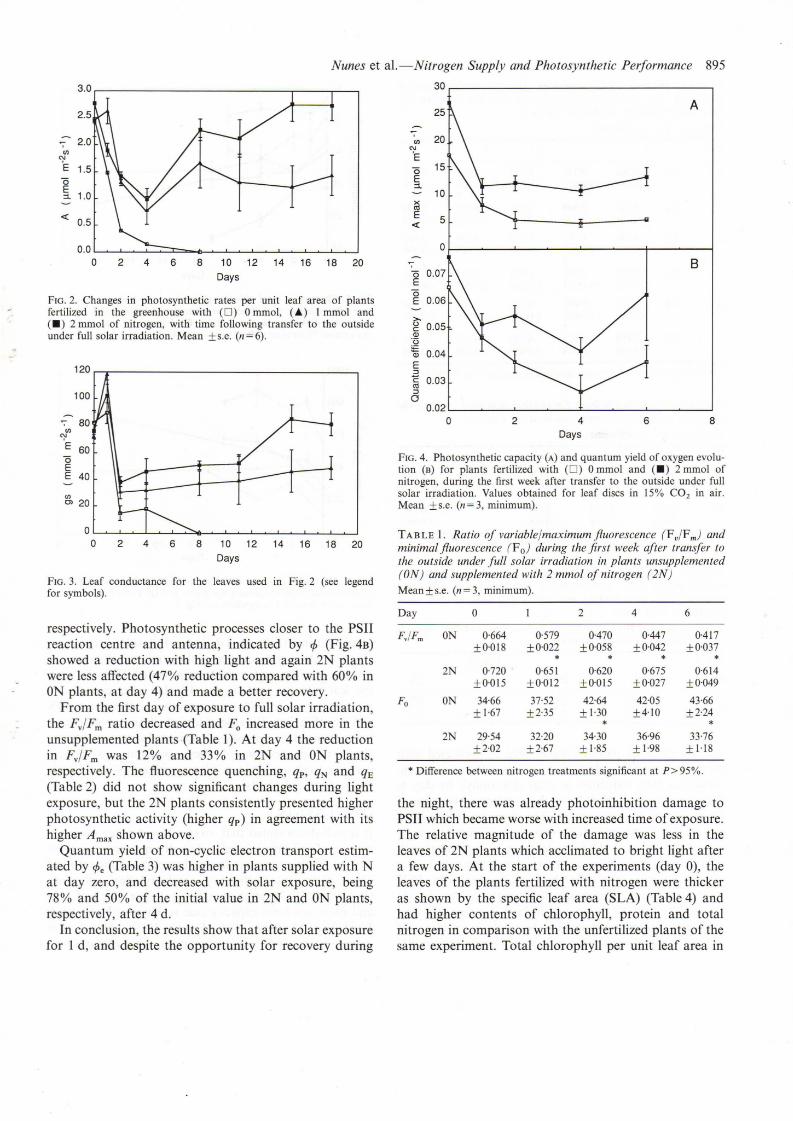
Oxygen evolution on day 4 of full solar irradiationclearly indicates that A max (Fig. 4A) was reduced to about28% and 41 % of its initial value, in ON and 2N plants,
FIG. I. Changes in photosynthetically active leaf area of plants fertilizedin the greenhouse with (D) 0 mmol, (.&) I mmol and (.) 2 mmol ofnitrogen, with time following transfer to the outside under full solarirradiation. Mean ± s.e. (n = 6 plants).
894 Nunes et al.-Nitrogen Supply and Photosynthetic Performance
MATERIALS AND METHODS of fresh tissue were used with 6 cm3 of a 25 mol m- 3 phosphatebuffer containing cysteine (30 mol m - 3) and EDTA (1 mol m - 3).
Total nitrogen was determined according to the Kjeldahlmethod in the remaining leaf tissue, oven-dried at 70°C. Allthe results are expressed per unit leaf area and represent theaverage of at least three replicates per treatment.
Seedlings of Coffea arabica L. cv. Catuai were grown in 500 gof a sandy soil contained in dark plastic bags, to the 8-10 leafstage of development (about 1'5-years-old), in a shaded greenhouse (300 /-Lmol m - 2 S -1 maximum irradiation) with air humidity at about 80% RH. At that growth stage, when the plantsshowed some chlorosis indicating that the soil was nutritionallyimpoverished, plants were separated into three groups of 30plants each and nitrogen was applied as NH4 N03 in doses ofommol (ON), 1 mmol (1 N) and 2 mmol (2N) per plant inJune 1990 (Exp. I), and in doses of 0 mmol (ON) and 2mmol(2N) in June 1991 (Exp. II). After 2 weeks the plants wereplaced under moderate light intensity in the greenhouse (occasional sunflecks of 700 /-Lmol m - 2 S - 1) and after 4 weeks weretransferred to a protected site in the open, under full solarirradiation, and continuously sprinkled with water. Leaf temperature was between 27-33 °C at noon but occasionally a maximalof 35·6 °C was recorded.
Measurements of growth, survival and photosyntheticallyactive leaf area were made from the beginning of exposure tofull solar irradiation and continued until the end of the experiment (130 d). Just prior to transfer to sunlight and at variousintervals for 18 d afterwards, net photosynthesis (A) and transpiration rates (T), conductance (gs) and concurrent microclimatic components (light, RH, leaf and air temperature) weremeasured under saturating light in one attached leaf of sixplants from each treatment, with a portable IRGA/porometerLI-6200 (LI-COR, Inc., Nebraska, USA). The measurementswere made in the morning when the plants had already beenexposed to at least 1 h of moderate light (less than 700 /-Lmolm - 2 S -1 irradiation).
Measurements of oxygen evolution, expressing photosyntheticcapacity (A max) and quantum yield of photosynthesis (1)), weremade in leaf discs (4'52 cm2
) under CO2 saturating conditions(15%) and at 25°C, following a similar time sequence. ForA max , saturating light intensities (about 1800/-Lmol m - 2 S-1)
were provided by a Bjorkman lamp (Hansatech, King's Lynn,UK). These determinations were measured with a leaf-discoxygen electrode LD2 (Hansatech, Kings Lynn, UK).
Chlorophyll a fluorescence was measured simultaneously withAmm using a PAM fluorometer (H. Walz, Effeltrich, Germany)as described by Quick and Stitt (1989). Photochemical (qp) andnon-photochemical (qN) quenching were determined with saturating light pulses (about 4500 !Lmol m - 2 S -1) and actinic lightof 1800 /-Lmol m - 2 S - 1. The qN was further resolved by determining the fast component (qE) of the quenching relaxation kineticsafter removing the actinic illumination and applying threesaturating pulses every 60 s. The ratio of variable to maximalfluorescence (Fv/Fm), and the minimal fluorescence (Fo) weredetermined in leaf discs previously adapted to darkness and1>. = (qp.Fv/Fm) was calculated according to Genty et al. (1989).The leaf discs for oxygen-electrode and fluorescence measurements were collected in the early morning before incident lighthad reached photoinhibitory levels and were selected fromundamaged areas of leaf.
Several chemical determinations were carried out in parallelwith the physiological ones, i.e. just before the transfer of theplants to full solar irradiation (day zero) and a few daysafterwards, using discs or a pooled sample formed with the twofirst pairs of mature leaves collected from two plants (fourleaves). Chlorophyll was estimated in 5 cm3 methanol extracts(Mackinney, 1941) from four discs with an area of 0·5 cm 2 each.Soluble leaf protein was precipitated from the leaf extract using5% TCA and estimated according to Lowry et al. (1951) usingbovine serum albumin as standard. For the leaf extract, 0·5 g
8
B
A
64Days
2
25~
";'20Vl
N
'E
(5 15E:1.
10)(
ellE 5«
0
";'(5 0.07E0
0.06E
>.(,) 0.05c:Q)
'0
~ 0.04E:::JC 0.03ell:::Ja
0.020
TABLE 1. Ratio of variable/maximum fluorescence (Fv/Fm) andminimal fluorescence (F0) during the first week after transfer tothe outside under full solar irradiation in plants unsupplemented(ON) and supplemented with 2 mmol of nitrogen (2N)Mean ± s.e. (n = 3, minimum).
FIG. 4. Photosynthetic capacity (A) and quantum yield of oxygen evolution (B) for plants fertilized with (D) 0 mmol and (.) 2 mmol ofnitrogen, during the first week after transfer to the outside under fullsolar irradiation. Values obtained for leaf discs in 15% CO2 in air.Mean ± s.e. (n = 3, minimum).
8 1012 14 16 18 20
Days
8 10 12 14 16 18 20
Days
6
6
2 4
2 4
120
100
";' 80Vl
C)lE 60(5EE 40
VlC> 20
3.0
2.5
2.0Vl
N
'E 1.5(5E
1.0:1.
« 0.5
0.00
Nunes et al.-Nitrogen Supply and Photosynthetic Performance 89530.---- --,
FIG. 2. Changes in photosynthetic rates per unit leaf area of plantsfertilized in the greenhouse with (D) 0 mmol, (.A) I mmol and(.) 2 mmol of nitrogen, with time following transfer to the outsideunder full solar irradiation. Mean ± s.e. (n = 6).
FIG. 3. Leaf conductance for the leaves used In Fig. 2 (see legendfor symbols).
respectively. Photosynthetic processes closer to the PSIIreaction centre and antenna, indicated by eP (Fig.4B)showed a reduction with high light and again 2N plantswere less affected (47% reduction compared with 60% inON plants, at day 4) and made a better recovery.
From the first day of exposure to full solar irradiation,the Fv/Fm ratio decreased and Fo increased more in theunsupplemented plants (Table 1). At day 4 the reductionin Fv/Fm was 12% and 33% in 2N and ON plants,respectively. The fluorescence quenching, qp, qN and qE(Table 2) did not show significant changes during lightexposure, but the 2N plants consistently presented higherphotosynthetic activity (higher qp) in agreement with itshigher A max shown above.
Quantum yield of non-cyclic electron transport estimated by ePe (Table 3) was higher in plants supplied with Nat day zero, and decreased with solar exposure, being78% and 50% of the initial value in 2N and ON plants,respectively, after 4 d.
In conclusion, the results show that after solar exposurefor 1 d, and despite the opportunity for recovery during
Day 0 2 4 6
Fv/Fm ON 0·664 0·579 0-470 0·447 0·417±0·018 ±0'022 ±0'058 ±0'042 ±0·037
* * * *2N 0·720 0·651 0·620 0·675 0·614
±0·015 ±0'012 ±0'015 ±0'027 ±0'049
Fo ON 34·66 37·52 42·64 42·05 43'66± 1·67 ±2·35 ± 1·30 ±4·10 ±2·24
* *2N 29·54 32·20 34·30 36·96 33·76
±2'02 ±2·67 ±1·85 ±1·98 ±1·18
* Difference between nitrogen treatments significant at P> 95%.
the night, there was already photoinhibition damage toPSII which became worse with increased time of exposure.The relative magnitude of the damage was less in theleaves of 2N plants which acclimated to bright light aftera few days. At the start of the experiments (day 0), theleaves of the plants fertilized with nitrogen were thickeras shown by the specific leaf area (SLA) (Table 4) andhad higher contents of chlorophyll, protein and totalnitrogen in comparison with the unfertilized plants of thesame experiment. Total chlorophyll per unit leaf area in

1614128 10Days
642
60
~ 50E()
OJ 40~
>.~
c.. 3000:c()
20
FIG. 5. Changes in total chlorophyll content per unit leaf area withtime after transfer to full solar irradiation, for plants of Exp. I: 0(without N fertilizer); ... (I mmol N); • (2 mmol N); and of Exp. II:o (without N fertilizer); • (2 mmol N). Means ± s.e. (n = 6 leaves).
0 2 4 6
0·264 0·318 0·253 0·263 0·242±0'031 ±0'022 ±0'023 ±0'023 ±0'014
*0·380 0·309 0·301 00410 0·300
±0'056 ±0·030 ±0·0l1 ±0'023 ±0'032
0·800 0·802 0·810 0·808 0·847±0·024 ±0'007 ±0'016 ±0'019 ±0'008
*0·838 0·759 0·808 0·824 0·781
±0·006 ±0·011 ±0·01O ±0·012 ±0'031
0·399 00407 0·365 0·365 0·390±0'049 ±0·020 ±0'029 ±0'016 ±0'061
*0·468 0·325 0·337 0·387 0·380
±0·030 ±0'019 ±0'069 ±0'059 ±0'044
ON
2N
2N
2N
Day
896 Nunes et al.-Nitrogen Supply and Photosynthetic Performance
TAB L E 2. Photochemical quenching (qp) , non-photochemicalquenching (qN) and quenching for energy (qE) for the same leafdiscs and treatments as used for Table iMean ± s.e. (n = 3, minimum).
* Difference between nitrogen treatments significant at P> 95%.
TAB L E 3. Quantum efficiency for electron transport (1)e) for thesame leaf discs and treatments as used for Table iMean±s.e. (n=3, minimum).
Day 0 2 4 6
ON 0·200 0·199 0·120 0·105 0·111±0'012 ±0'009 ±0'020 ±0'013 ±0'013
*2N 0·245 0·205 0·193 0·190
±0·038 ±0'014 ±0'018 ±0'042
* Difference between nitrogen treatments significant at P> 95%.
1000
800
~ 600E()
OJ~
400c:'0;
ea.. 200
00 2 4 6 8 10 12 14 16
Days
TABLE 4. Specific leaf area (cm 2g- 1) for leaves of unsupple
mented plants (ON) and for plants that received i (iN) and2 mmol (2N) of nitrogen, just before exposure to full solarirradiation (day zero)Experiments I and II were performed in June 1990 and 1991, respectively. Mean±s.e. (n=3, minimum).
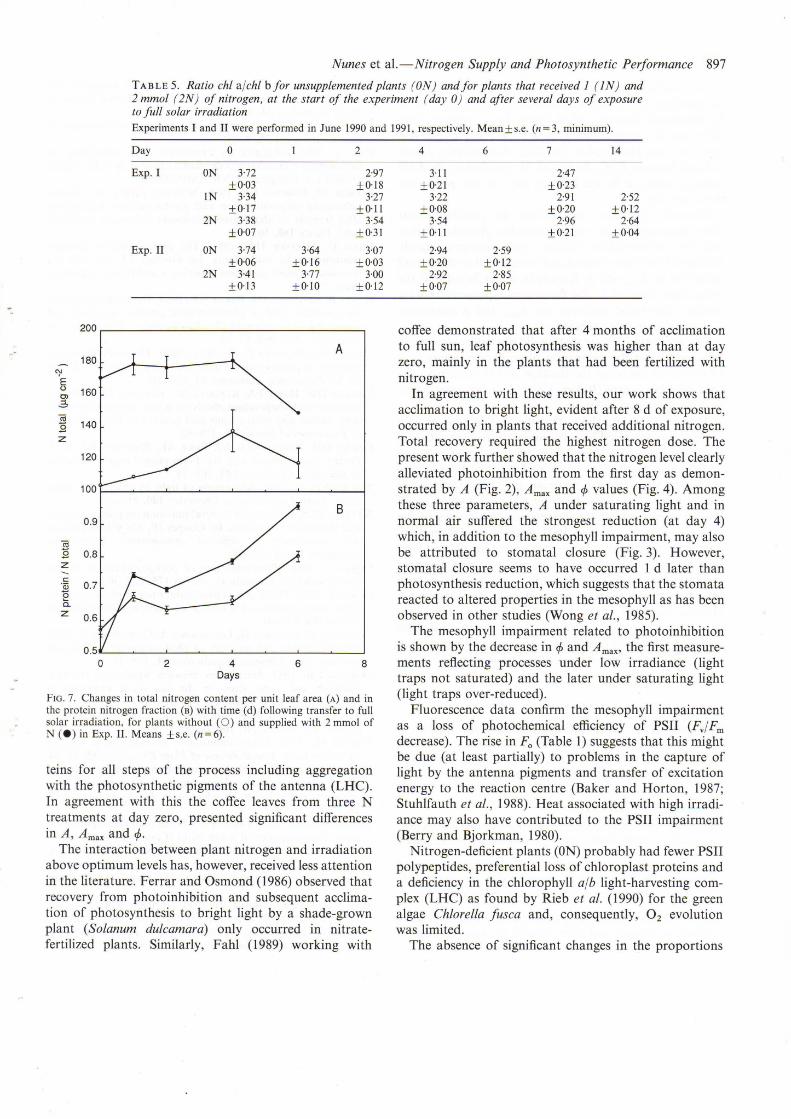
the two terminal pairs of leaves (Fig. 5) decreased withsolar exposure particularly in the plants with less nitrogen.Despite a 25% reduction in total chlorophyll by day 4,ON plants in Exp. II did not show chlorosis because oftheir initially higher content. The decrease in the ratiochI a/chI b (Table 5) indicates that a preferential loss ofchI a contributed to the total decrease, in agreement withresults from Williams et al. (1986) showing that PSI chI asuffers selective photobleaching associated with PSIIinhibition. Soluble protein content in the leaves (Fig. 6)increased in all the treatments in both experiments. Thetotal nitrogen content tended to be stable in the leaves offertilized plants while a slight increase showed in ON
Exp. I
ON
325·7± 18·6
IN
302·0±8'0
2N
285·0±21·3
Exp. II
ON
294·1±lB
2N
238·1±5'7
FIG. 6. Changes in protein content per unit leaf area with time followingtransfer to full solar irradiation, for plants of Exp. I and Exp. II. Means± s.e. (n = 6 leaves). For symbols see Fig. 5.
plants during the first 4 d under full solar irradiation(Fig. 7A). As a consequence of these relative changes, theprotein fraction under shade represented 50 to 57% ofthe leaf nitrogen and on the 4th day after exposure itrepresented 65 to 77%, respectively, in the groups ONand 2N (Fig. 7B). Because chlorophyll and protein contents changed in opposite directions during solar exposure, the proportion of these pigments to proteindecreased (about 50%) in plants without N and clearlyless in fertilized plants.
DISCUSSIONIt is well documented that, over a broad range of doses,nitrogen fertilization is related to dry matter productionas a result of increases in dry matter partitioning into leafarea, water use efficiency, carboxylation efficiency(Syvertsen, 1987), photosynthesis rate per unit leaf areaand photosynthetic capacity (see reviews by Natr, 1975;Field and Mooney, 1986; Evans, 1989). This is notsurprising since photosynthetic performance requires pro-
Nunes et al.-Nitrogen Supply and Photosynthetic Performance 897
TABLE 5. Ratio chI a/chI bfor unsupplemented plants (ON) andfor plants that received J (IN) and2 mmol (2N) of nitrogen, at the start of the experiment (day 0) and after several days of exposureto full solar irradiationExperiments I and II were performed in June 1990 and 1991, respectively. Mean±s.e. (n=3, minimum).
Day
Exp. I
Exp. II
0 2 4 6 7 14
ON 3·72 2·97 3·11 2-47±0'03 ±0'18 ±0'21 ±0'23
IN 3-34 3·27 3·22 2·91 2·52±0'17 ±O'II ±0'08 ±0·20 ±0'12
2N 3-38 3·54 3·54 2·96 2-64±0'07 ±0'31 ±O·II ±0·21 ±0'04
ON 3·74 3·64 3·07 2·94 2·59±0'06 ±0·16 ±0'03 ±0'20 ±0'12
2N HI 3·77 3-00 2·92 2·85±0·13 ±O'IO ±0'12 ±0·07 ±0·07
fIG. 7. Changes in total nitrogen content per unit leaf area (A) and inthe protein nitrogen fraction (B) with time (d) following transfer to fullsolar irradiation, for plants without (0) and supplied with 2 mmol ofN (e) in Exp. II. Means ±s.e. (n=6).
teins for all steps of the process including aggregationwith the photosynthetic pigments of the antenna (LHC).In agreement with this the coffee leaves from three Ntreatments at day zero, presented significant differencesin A, Amax and 4>.
The interaction between plant nitrogen and irradiationabove optimum levels has, however, received less attentionin the literature. Ferrar and Osmond (1986) observed thatrecovery from photoinhibition and subsequent acclimation of photosynthesis to bright light by a shade-grownplant (Solanum dulcamara) only occurred in nitratefertilized plants. Similarly, Fahl (1989) working with
--200
180~
E0 160Ol
.6] 140.8z
120
100
0.9
co:§ 0.8z--c:
0.7'0:;
ea.Z 0.6
2 4Days
6
A
B
8
coffee demonstrated that after 4 months of acclimationto full sun, leaf photosynthesis was higher than at dayzero, mainly in the plants that had been fertilized withnitrogen.
In agreement with these results, our work shows thatacclimation to bright light, evident after 8 d of exposure,occurred only in plants that received additional nitrogen.Total recovery required the highest nitrogen dose. Thepresent work further showed that the nitrogen level clearlyalleviated photoinhibition from the first day as demonstrated by A (Fig. 2), Amax and 4> values (Fig. 4). Amongthese three parameters, A under saturating light and innormal air suffered the strongest reduction (at day 4)which, in addition to the mesophyll impairment, may alsobe attributed to stomatal closure (Fig. 3). However,stomatal closure seems to have occurred 1 d later thanphotosynthesis reduction, which suggests that the stomatareacted to altered properties in the mesophyll as has beenobserved in other studies (Wong et al., 1985).
The mesophyll impairment related to photoinhibitionis shown by the decrease in 4> and Amax , the first measurements reflecting processes under low irradiance (lighttraps not saturated) and the later under saturating light(light traps over-reduced).
Fluorescence data confirm the mesophyll impairmentas a loss of photochemical efficiency of PSII (FyiFm
decrease). The rise in Fo (Table 1) suggests that this mightbe due (at least partially) to problems in. the capture oflight by the antenna pigments and transfer of excitationenergy to the reaction centre (Baker and Horton, 1987;Stuhlfauth et al., 1988). Heat associated with high irradiance may also have contributed to the PSII impairment(Berry and Bjorkman, 1980).
Nitrogen-deficient plants (ON) probably had fewer PSIIpolypeptides, preferential loss of chloroplast proteins anda deficiency in the chlorophyll alb light-harvesting complex (LHC) as found by Rieb et al. (1990) for the greenalgae Chlorella fusca and, consequently, O2 evolutionwas limited.
The absence of significant changes in the proportions

898 Nunes et al.-Nitrogen Supply and Photosynthetic Performance
of the available energy (Table 2) for photosynthesis (qp),for building up the transthylakoidal gradient (qE)' or tobe dissipated through other forms (qN), suggests thatthere was no excess of energy flow from PSII, instead fewelectrons were produced at that level. Accordingly, rPe wasreduced because Fv/Fmdecreased. In conclusion, the PSIIdamage seems to be the limiting step for the photosynthetic activity.
However, this does not exclude the possibility thatother processes such as electron transfer, phosphorylationand the carboxylation (Calvin cycle enzymes, mainlyRubisco) have been affected and contributed to the overallreduction in Amax and rP. It must be remembered that thereductions detected in the fluorescence parameters weresmaller than those observed for Amax and rP suggestingthat other steps were affected.
It is interesting to note that we did not detect alternativemechanisms for the dissipation of excitation energy (qN)which could minimize the damage to D 1 protein in thePSII reaction centre (Baker, 1991), thus we may expectthat this protein was hit.
The chemical data shows that the total soluble proteincontent was favoured in coffee plants with high nitrogensupply and clearly increased from the first day of exposureto full solar irradiation, in parallel with the tolerance tophotoinhibition. In other materials, e.g. Anacystis nidulansand barley leaves (Samuelsson et al., 1987; Oquist et al.,1987; Greer et aI., 1991) the relative repair rates of thephotosensitive proteins in PSII, mainly the D 1 protein,have been suggested as being responsible for photoinhibition tolerance. This suggests that the repair of damagedproteins may be a key process in the tolerance inducedby nitrogen fertilization in coffee leaves. Moreover,because the PSII complex of the fertilized plants wasprotected since the first day of exposure to full solarradiation the repair of PSII proteins seems to be involvedand start immediately, even with the leaf under brightlight conditions.
ACKNOWLEDGEMENTSThe authors are grateful to Dr M. Manuela Chaves ofInstituto Superior de Agronomia for the use of thefluorometer and valuable discussions and to Dr BryanLewis for the revision of the English manuscript.
LITERATURE CITED
BAKER NR. 1991. A possible role for photosystem II in environmental perturbations of photosynthesis. PhysiologiaPlantarum 81, 563-70.
BAKER NR, HORTON P. 1987. Fluorescence quenching duringphotoinhibition. In: Kyle DJ, Osmond CB, Arntzen CJ, eds.Photoinhibition. Amsterdam: Elsevier Science Publishers,145-67.
BENNET JM, JONES JW, ZUR B, HAMMOND LC. 1986. Interactiveeffects of nitrogen and water stresses on water relations offield-grown corn leaves. Agronomy Journal 78, 273-80.
BERRY J, BJORKMAN O. 1980. Photosynthetic response andadaptation to temperature in higher plants. Annual Review ofPlant Physiology 31, 491-638.
EVANS JR. 1989. Photosynthesis and nitrogen relationships inleaves. Oecologia 78, 9-19.
FAHL J. 1989. Influencia da irradiancia e do nitrogenio nafotossintese e crescimento de plantas jovens de cafe (Coffeaarabica L.). Campinas, UNICAMP (Thesis).
FERRAR PJ, OSMOND CB. 1986. Nitrogen supply as a factorinfluencing photoinhibition and photosynthetic acclimationafter transfer of shade-grown Solanum dulcamara to brightlight. Planta 168, 563-70.
FIELD C, MOONEY HA. 1986. The photosynthesis-nitrogenrelationships in wild plants. In: Givnish TJ, eds. On theeconomy of plant form and function. Cambridge: UniversityPress, 25-55.
GENTY B, BRIANTAIS JM, BAKER NR. 1989. The relation betweenthe quantum yield of photosynthetic electron transport andquenching of chlorophyll fluorescence. Biochimica etBiophysica Acta 990, 87-92.
GREER DH, OTTANDER C, OQUIST G. 1991. Photoinhibition andrecovery of photosynthesis in intact barley leaves at 5 °C and20 0c. Physiologia Plantarum 81, 203-10.
LAWLOR DW, BOYLE FA, KENDALL AC, KEyS AJ. 1987. Nitratenutrition and temperature effects on wheat: enzyme composition, nitrate and total amino acid content of leaves. Journalof Experimental Botany 38, 378-92.
LOWRY OH, ROSEBROUGH NJ, FARR AL, RANDALL RJ. 1951.Protein measurement with the Folin phenol reagent. Journalof Biological Chemistry 193, 265-75.
MACKINNEY G. 1941. Absorption of light by chlorophyll solutions. Journal of Biological Chemistry 140, 315-22.
NATR L. 1975. Influence of mineral nutrition on photosynthesisand the use of assimilates. In: Cooper JP, eds. Photosynthesisand productivity in different environments. Cambridge:University Press, 537-55.
OGREN E. 1988. Photoinhibition of photosynthesis in willowleaves under field conditions. Planta 175, 229-36.
OGREN E. 1991. Prediction of photoinhibition of photosynthesisfrom measurements of fluorescence quenching components.Planta 184, 538-44.
OQUIST G, SAMUELSSON G, LONNEBORG A, GUSTAFSSON P. 1987.Photoinhibition and recovery of photosynthesis in Anacystisnidulans. Acta Chemica Scandinavica 41, 108-11.
OSMOND CB. 1983. Interaction between irradiance, nitrogennutrition and water stress in the sun-shade responses ofSolanum dulcamara. Oecologia 57, 316-21.
OSMOND CB. 1987. Photosynthesis and carbon economy ofplants. New Phytologist 106, (supp!.), 161-75.
POWLES SB. 1984. Photoinhibition of photosynthesis inducedby visible light. Annual Review of Plant p'hysiology 35, 15-44.
QUICK WP, STITT M. 1989. An examination offactors contributing to non-photochemical quenching of chlorophyll fluorescence in barley leaves. Biochimica et Biophysica Acta 977,287-96.
RHIZOPOULOU S, NUNES MA. 1981. Some adaptative photosynthetic characteristics of a sun plant (Ceratonia siliqua) and ashade plant (Coffea arabica). In: Margaris NS, Mooney HA,eds. Components of productivity of Mediterranean-climateregions. Basic and applied aspects. The Hague: Dr W Junk,85-9.
RIEB MH, DAMM I, GRIMME H. 1990. Nitrogen deprivation andthe photosynthetic apparatus of the green algae Chlorellafusca. In: Baltscheffsky M, ed. Current research in photosynthesis. Dordrecht: Kluwer Academic, IV, 405-8.

Nunes et at-Nitrogen Supply and Photosynthetic Performance 899
Ruo P, NUNES MA, OIAS MA. 1989. Fotoinibi9:10 no cafeeiro. of plant water relations of beans (Phaseolus vulgaris L.). NewEfeito da ferti1iza9:10 azotada. VIII. Reunion Nacional de la Phytologist 69, 413-24.S.E.F.V.I Congreso Hispano-Luso de Fisiologia Vegetal. STUHLFAUTH T, SULTEMEYER OF, WEINZ S, FocK H. 1988.Barcelona. Fluorescence quenching and gas exchange in a water-stressed
ROSENQUIST E, WINGSLE G, OGREN E. 1991. Photoinhibition of C3 plant, Digitalis lanata. Plant Physiology 86, 246-50.photosynthesis in intact willow leaves in response to moderate SYVERTSEN JP. 1987. Nitrogen content and CO2 assimilationchanges in light and temperature. Physiologia Plantarum characteristics of Citrus leaves. HortScience 22, 289-91.83, 390-6. WILLIAMS WP, SEN A, FORK OC. 1986. Selective photobleaching
SAMUELSSON G, LONNEBORG A, GUSTAFSSON P, OQUIST G. 1987. of PSI-related chlorophylls in heat-stressed pea chloroplasts.The susceptibility of photosynthesis to photoinhibition and Photosynthesis Research 10, 75-92.the capacity for recovery in high- and low-light grown cyano- WONG SC, COWAN IR, FARQUHAR GO. 1985. Leaf conductancebacteria. Anacystis nidulans. Plant Physiology 83, 438-41. in relation to rate of CO2 assimilation. II. Influence of water
SHIMSHI D. 1970. The effect of nitrogen supply on some indices stress and photoinhibition. Plant Physiology 78, 830-4.
Related Documents






![Original Research Photosynthetic Responses of Tomato Leaves to … lycopersicum cv. Rio Grande), a variety that plays an important role in the Tunisian agricultural economy [44]. The](https://static.cupdf.com/doc/110x72/5f0bb5807e708231d431d46b/original-research-photosynthetic-responses-of-tomato-leaves-to-lycopersicum-cv.jpg)




