Eur. J. Biochem. 67, 177-186 (1976) Effect of Inhibitors on the Substrate-Dependent Quenching of 9-Aminoacridine Fluorescence in Inside-Out Membrane Vesicles of Escherichia coli Akhand P. SINGH and Philip D. BRAGG Department of Biochemistry, University of British Columbia, Vancouver (Received March 1 /May 14. 1976) The effect of various inhibitors on the substrate-dependent quenching of the fluorescence of 9- aminoacridine was measured in inside-out membrane vesicles of Eschrriclzia coli. The rate of fluores- cence quenching in the presence of inhibitors was dependent on the rate of electron transfer through the respiratory chain with NADH, succinate, D-lactate or DL-glycerol 3-phosphate as substrates. Several patterns of response were given by the inhibitors. Inhibitors competitive with substrate, or those acting only on the dehydrogenases, gave a direct relationship between the extent of inhibition of oxidase activity and the rate of quenching. A biphasic relationship was given by 2-heptyl-4-hydroxy- quinoline N-oxide and piericidin A which was due to these compounds acting both as inhibitors of the respiratory chain and, at higher concentrations, as uncoupling agents. Uncouplers inhibited fluorescence quenching with minimal inhibition of oxidase activity. The transmembrane pH difference was calculated from the extent of fluorescence quenching and the intravesicular volume. The maximum pH difference of 3.3-3.7 units was generated by each of the substrates tested. The fluorescence of acridine dyes has been shown to alter with changes in the energized state of phos- phorylating membranes from various sources [I - 51 including Escherichia coli [6- 121. In inside-out mem- brane vesicles of this organism the fluorescence of acridine dyes can be quenched either by substrate oxidation through the respiratory chain or by hydro- lysis of ATP [6-121. When the electron transport chain is incomplete [8,9], or blocked by inhibitors or anaerobiosis, fluorescence quenching by substrate oxidation is abolished but will occur if ATP is added. The involvement of the (Ca2 +, Mg2 +)-activated ATP- ase in this process has been shown by the use of mutants or by treatment with ATPase inhibitors [6,10- 121. Mitchell [13]has suggested that the electrochemical gradient of protons which can be formed across certain membranes of cells is of primary importance in the mechanism of biological energy conversion. The electrochemical potential difference for protons (“pro- tonmotive force”, dp) [13] in electrical units is given by the relationship dp = A$ - Z.ApH where A$ is the membrane potential, dpH is the trans- membrane pH difference, and 2 is a factor to convert pH into electrical units. The energy-dependent quench- Enzyntes. ATPase (EC 3.6.1.3); succinale dehydrogenase (EC 1.3.99.1); D-lactate dehydrogenase (cytochrome) (EC 1.1.2.4). ing of the fluorescence of 9-aminoacridine was found to occur when the dye entered the internal space of chloroplasts, subchloroplast particles, chromatopho- res, and submitochondrial particles, in response to the transmembrane pH difference. Thus, it represents a sensitive measure of alterations in ApH with the further advantage that continuous measurements can be made [ 1,2,4,5,14]. Other energy-linked functions in E. coli, including the active transport of various solutes [15], respond to the presence and absence of respiratory chain sub- strates and inhibitors, ATP, anaerobiosis, and muta- tions involving the (Ca2+, Mg’+)-activated ATPase, in a similar manner to fluorescence quenching. This is consistent with the involvement of the transmem- brane pH difference in these processes. It is probable, as Niven and Hamilton [16] have shown for amino acid transport in Staphylococcus nureus, that the uptake of only uncharged and anionic solutes will take place in response to dpH. (The membrane potential will also be involved in the uptake of uncharged solu- tes.) In agreement with this view, it has been shown in E. coli that active transport of P-galactosides, arabi- nose, galactose, glucose 6-phosphate, succinate, fuma- rate, malate and aspartate occur with solute : proton symport [17 - 211. In experiments using right-side-out membrane vesicles of E. coli and other organisms [22] there appeared to be no relationship between the rate at

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. 67, 177-186 (1976)
Effect of Inhibitors on the Substrate-Dependent Quenching of 9-Aminoacridine Fluorescence in Inside-Out Membrane Vesicles of Escherichia coli Akhand P. SINGH and Philip D. BRAGG
Department of Biochemistry, University of British Columbia, Vancouver
(Received March 1 /May 14. 1976)
The effect of various inhibitors on the substrate-dependent quenching of the fluorescence of 9- aminoacridine was measured in inside-out membrane vesicles of Eschrriclzia coli. The rate of fluores- cence quenching in the presence of inhibitors was dependent on the rate of electron transfer through the respiratory chain with NADH, succinate, D-lactate or DL-glycerol 3-phosphate as substrates. Several patterns of response were given by the inhibitors. Inhibitors competitive with substrate, or those acting only on the dehydrogenases, gave a direct relationship between the extent of inhibition of oxidase activity and the rate of quenching. A biphasic relationship was given by 2-heptyl-4-hydroxy- quinoline N-oxide and piericidin A which was due to these compounds acting both as inhibitors of the respiratory chain and, at higher concentrations, as uncoupling agents. Uncouplers inhibited fluorescence quenching with minimal inhibition of oxidase activity. The transmembrane pH difference was calculated from the extent of fluorescence quenching and the intravesicular volume. The maximum pH difference of 3.3-3.7 units was generated by each of the substrates tested.
The fluorescence of acridine dyes has been shown to alter with changes in the energized state of phos- phorylating membranes from various sources [I - 51 including Escherichia coli [6- 121. In inside-out mem- brane vesicles of this organism the fluorescence of acridine dyes can be quenched either by substrate oxidation through the respiratory chain or by hydro- lysis of ATP [6-121. When the electron transport chain is incomplete [8,9], or blocked by inhibitors or anaerobiosis, fluorescence quenching by substrate oxidation is abolished but will occur if ATP is added. The involvement of the (Ca2 +, Mg2 +)-activated ATP- ase in this process has been shown by the use of mutants or by treatment with ATPase inhibitors [6,10- 121.
Mitchell [13] has suggested that the electrochemical gradient of protons which can be formed across certain membranes of cells is of primary importance in the mechanism of biological energy conversion. The electrochemical potential difference for protons (“pro- tonmotive force”, dp) [13] in electrical units is given by the relationship
dp = A $ - Z . A p H
where A $ is the membrane potential, dpH is the trans- membrane pH difference, and 2 is a factor to convert pH into electrical units. The energy-dependent quench-
Enzyntes. ATPase (EC 3.6.1.3); succinale dehydrogenase (EC 1.3.99.1); D-lactate dehydrogenase (cytochrome) (EC 1.1.2.4).
ing of the fluorescence of 9-aminoacridine was found to occur when the dye entered the internal space of chloroplasts, subchloroplast particles, chromatopho- res, and submitochondrial particles, in response to the transmembrane pH difference. Thus, it represents a sensitive measure of alterations in ApH with the further advantage that continuous measurements can be made [ 1,2,4,5,14].
Other energy-linked functions in E. coli, including the active transport of various solutes [15], respond to the presence and absence of respiratory chain sub- strates and inhibitors, ATP, anaerobiosis, and muta- tions involving the (Ca2+, Mg’+)-activated ATPase, in a similar manner to fluorescence quenching. This is consistent with the involvement of the transmem- brane pH difference in these processes. It is probable, as Niven and Hamilton [16] have shown for amino acid transport in Staphylococcus nureus, that the uptake of only uncharged and anionic solutes will take place in response to dpH. (The membrane potential will also be involved in the uptake of uncharged solu- tes.) In agreement with this view, it has been shown in E. coli that active transport of P-galactosides, arabi- nose, galactose, glucose 6-phosphate, succinate, fuma- rate, malate and aspartate occur with solute : proton symport [17 - 211.
In experiments using right-side-out membrane vesicles of E. coli and other organisms [22] there appeared to be no relationship between the rate at

178 Inhibition of Fluorescence Quenching
which a substrate was oxidized and its ability to drive active transport. This suggested that transport was coupled to respiratory chains concerned with the oxidation of only certain substrates, for example, D-lactate. From this viewpoint, the fact that NADH was a poor substrate to support active transport in right-side-out membrane vesicles of E. coli indicated that the NADH oxidase respiratory pathway was not linked to the transport system [22]. Recently, Futai has shown that the most likely reason for this result is that the vesicles were not permeable to NADH [23]. If NADH was generated within the vesicle then it was one of the most effective substrates for supporting transport. Presumably, the differential effects of the other substrates on transport could be explained on the same basis although this has not been demon- strated.
In this paper we have examined the relationship between membrane energization and the rate of oxida- tion of various substrates. In order to avoid the pro- blem of substrate permeability we have used inside-out membrane vesicles and have measured the quenching of the fluorescence of 9-aminoacridine as an index of the transmembrane pH difference. We have found that the rate of energization was dependent on the rate of electron transfer through the respiratory chain with NADH, succinate, D-lactate, and DL-glycerol 3-phos- phate as substrates. The maximum transmembrane pH difference which could be generated by each substrate was similar. Furthermore, this technique appeared to be useful in investigating the site of action of inhibitors as a number of different types of fluor-. escence response in the presence of inhibitors could be detected.
MATERIALS AND METHODS
Chem ica Is All chemicals were of reagent grade purity. Lithium
D-lactate, NADH, valinomycin, and carbonylcyanide m-chlorophenylhydrazone were purchased from Cal- biochem. DL-Glycerol 3-phosphate and 2-heptyl-4- hydroxyquinoline N-oxide were obtained from Sigma Chemicals Co., and 9-aminoacridine hydrochloride from Aldrich Chemical Co. Nigericin and piericidin A were generous gifts from Drs. R. J. Hosley (Lilly Research Laboratories) and S. Tamura (University of Tokyo), respectively. Inulin [lSC]carboxylic acid (13 Ci/mol) and 3Hz0 (1.14 pCi/ml) were obtained from Amersham-Searle Corporation and Packard Instrument Company, respectively.
Organism and Growth
E. coli NRC 482 was grown anaerobically at 22 "C in 4-1 flasks (filled to the top) in a minimal salts medium
[9] containing 20 mM potassium fumarate, 68 mM glycerol and 0.2 (w/v) vitamin-free casamino acids. Dissolved oxygen was removed from the flasks by flushing with nitrogen for 20 min before sealing with a fermentation lock.
Preparation of Inside-Out Membrane Vesicles
Inside-out membrane vesicles were prepared from cells harvested at the end of the exponential phase of growth by the method of Bragg et al. [24] except that the vesicles were not washed.
9-Aminoacridine Fluorescence
The fluorescence of 9-aminoacridine was measured at 22 "C with a Turner model 420 spectrofluorometer connected to a Varian strip chart recorder. The reac- tion mixture in a cuvette of 1-cm light path and in a final volume of 2 ml contained 300 mM KC1, 5 mM MgC12, 10 mM N-2-hydroxyethylpiperazine-N'- ethanesulfonate/KOH buffer, pH 7.5, 2- 10 pM 9- aminoacridine, and 1-4 mg vesicle protein. The energy sources were used at the concentrations indicat- ed in the legends to the figures. Fluorescence was excited by light at 420 nm and emission was measured at 500 nm [4]. At these wavelengths there was no inter- ference from the fluorescence of NADH.
The transmembrane pH difference (dpH) was cal- culated from the fluorescence data by the method of Schuldiner et al. [25] using the value of 0.5 pl/mg protein determined below for the intravesicular volume.
Measurement of Oxidase Activity
The oxidase activity was calculated from the time required to deplete the oxygen in the cuvette during the measurement of 9-aminoacridine quenching.
Protein was determined by the Folin method [26].
Determination of /ntvavesicular Volume
Inside-out membrane vesicles suspended in 7.5 ml 10 mM N-2-hydroxyethylpiperazine-N'-2-ethanesul- fonate/KOH buffer, pH 7.5, containing 300 mM KC1 and 5 mM MgClz, were supplemented with 1 pCi of both H2O and in~lin-['~C]carboxylic acid. After 30 min at 22 "C the vesicles were collected by centrif- ugation at 140000 x g for 1 h. Both the pellet and the supernatant were treated with 8 ml10 "/, trichloroacetic acid. Aliquots (250 pl) of the deproteinized samples were added to PCS (Amersham-Searle) and the 3H and 14C activities determined in a Packard Tri-Carb liquid scintillation counter (model 2425). The intra- vesicular volume of the inside-out membrane vesicles calculated by the method of Hunter and Brierley [27]

A. P. Singli and P. D. Bragg 179
was 0.5 pl/mg protein over the range of 45 to 140 mg used in our experiments.
RESULTS
In this study inside-out membrane vesicles prepared from cells grown anaerobically on glycerol with fumarate were used. Vesicles prepared in this way had fully-induced levels of the membrane-bound glycerol 3-phosphate and succinate dehydrogenases.
Effect of Substrates on Quenching of 9-Aminoacridine Fluorescence
The effect of DL-glycerol 3-phosphate, D-lactate, succinate, and NADH on the fluorescence of 9-amino- acridine in the presence of inside-out membrane vesicles is shown in Fig. 1 A. When limiting amounts of these electron donors were used the fluorescence of the dye could be quenched sequentially until the maxi- mum level of quenching was attained. In the presence of malonate or of oxalate, competitive inhibitors for succinate and D-lactate dehydrogenase [28,29], re- spectively, sequential quenching of fluorescence by limiting amounts of the other substrate could still be demonstrated. Analogous results were obtained when saturating concentrations of the various electron donors were used (Fig. 1 B) although in this instance each substrate could cause the maximum extent of quenching possible. Quenching of fluorescence was reversed when the system became anaerobic but could be restored by the addition of ATP.
J
r
I
Fluorescence Quenching and the Transmembrane p H Difference
As discussed in the Introduction the quenching of the fluorescence of 9-aminoacridine has been used to measure the transmembrane pH difference (ApH) in chloroplast and subchloroplast particles, chromato- phores, liposomes and submitochondrial particles [ I , 2,4,5,14]. The following experiments were per- formed to see if the E. coli inside-out membrane vesicles behaved in a similar fashion to these sys- tems.
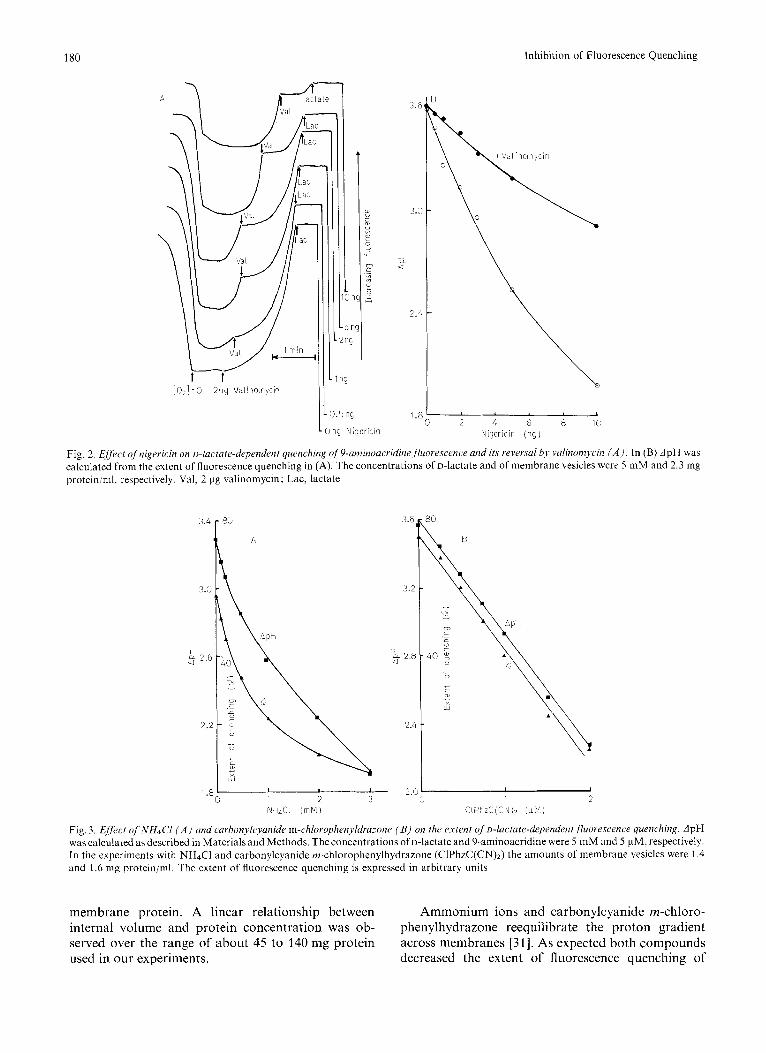
The effect of nigericin and valinomycin on the quenching of 9-aminoacridine fluorescence by u-lac- tate oxidation is shown in Fig. 2A. Preincubation of the inside-out membrane vesicles in a KCl but not a NaC1-containing buffer with various concentrations of nigericin resulted in varying degrees of inhibition of quenching presumably by electroneutral H '/K +
exchange across the vesicle membrane with consequent reduction in the size of ApH [30]. Addition of valino- mycin partially reversed the effect of nigericin by permitting exit of K + from the vesicles and allowing ApH to reform. At high nigericin concentrations valinomycin could not reverse the effect of this inhibi- tor. Similar results were obtained when succinate was used as substrate. These findings are similar to those obtained with submitochondrial particles by Rottenberg and Lee [4]. The etfect of nigericin on the ApH before and following addition of valinomycin is plotted in Fig. 2 B. The transmembrane pH difference was calculated from the extent of fluorescence quench- ing as described by Schuldiner et al. [25]. The internal volume of the vesicles was determined to be 0.5 pl/mg
La. G r o - P
[O?! -0 / /
I
Fig. 1. E&ct of malonate and oxalate on succirzate, o-lactate, u~.-glycerol3-plio.sp/iutr arid NA DH-dep~ndr.nt qucwching of 9-arninoacridine fluorescence. The concentrations of malonate, oxalate, succinate (Suc), D-lactate (Lac), Di.-glycerol 3-phosphate (Gro-P) and NADH were 1.25,0.25, 1.0, 0.25, 5.0 and 0.25 mM, respectively, in (A), In (B) o-lactate, succinate, and DL glycerol 3-phosphate were used at a final concen- tration of 10 m M ; oxalate (0xl)and malonate (Mal) were at 0.5 and 5 mM, respectively. The concentration of 9-aminoacridine was 10 pM. Membrane vesicles, 2.8 mg protein
180 Inhibition of Fluorescence Quenching
2 rnp
1 ng
i O 2 1 zo 2pg Valinomycin I1
3.6
3.0
L, a
2.4
I 1.8 C. 2 4 6 8 1C
Nigericin ( n g )
Fig. 2. Eflect qf'nigericin on o-lactate-dependent quenching qf9-amitioacridinefluorrscencr and its reversul bJ, valinonTycin ( A ) , In (B) ApH was calculated from the extent of fluorescence quenching in (A). The concentrations of D-lactate and of membrane vesicles were 5 mM and 2.3 mg protein/ml, respectively. Val, 2 pg valinomycin; Lac, lactate
3.4
3.0
I 3 2.6
2.2
8G
A
" NHICl (mM) C I ? ~ Z C ( C N ) ~ (pr,i)
Fig. 3. E.frct qfNHdc'l ( A ) and carhonylcyunide m-chlorophen~ldra--one ( B ) on the extent ofo-lactate-dependent.fluo~es~ence quenching. ApH was calculated as described in Materials and Methods. The concentrations of D-lactate and 9-aminoacridine were 5 niM and 5 pM, respectively. In the experiments with NH4C1 and carbonylcyanide in-chlorophenylhydrazone (CIPhzC(CN)2) the amounts of membrane vesicles were 1.4 and 1.6 nig protein/ml. The extent of fluorescence quenching is expressed in arbitrary units
membrane protein. A linear relationship between internal volume and protein concentration was ob- served over the range of about 45 to 140 mg protein used in our experiments.
Ammonium ions and carbonylcyanide m-chloro- phenylhydrazone reequilibrate the proton gradient across membranes [31]. As expected both compounds decreased the extent of fluorescence quenching of
181 A. P. Singh and P. D. Bragg
A o - L a c t a t e
3 x a l a t e (WI
250 200
150
100 80
40 20
10 0
l a lona ie i m M )
5
4
2.5
0
S u c c i n a t e
/ 1 min
D - L a c t a t e + oxalate / 6 12 18 24 30
I n i t i a l r a t e of f luorescence quenching
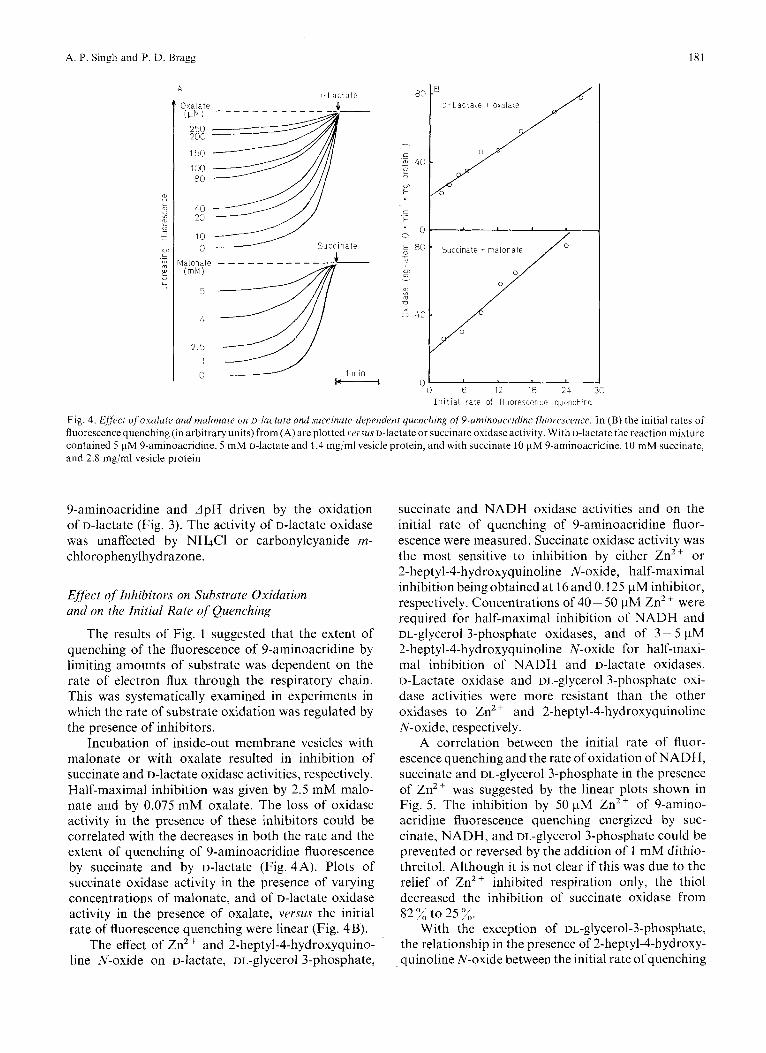
Fig. 4. f i f fecf qfoxulafe and nafonace on ~-1ai~tate and surrirzate-drJpertdeni quenching of~-aniinouc.i.idine Jluorescmre. In ( B ) the initial rates or fluorescence quenching (in arbitrary units) from (A) are plottcd ve~sus D-lactate or succinate oxidase activity. With D-lactate the reaction mixture contained 5 pM 9-aminoacridine, 5 mM D-hctale and 1.4 mg/ml vesicle protein, and with succinate 10 pM 9-aminoacridine. 10 mM succinate, and 2.8 mg/ml vesicle protein
9-aminoacridine and ApH driven by the oxidation of D-lactate (Fig. 3) . The activity of D-lactate oxidase was unaffected by NH4Cl or carbonylcyanide rn- chlorophenylhydrazone.
Effect of Inhibitors on Substrate Oxidation and on the Initial Rate of Quenching
The results of Fig. 1 suggested that the extent of quenching of the fluorescence of 9-aminoacridine by limiting amounts of substrate was dependent on the rate of electron flux through the respiratory chain. This was systematically examined in experiments in which the rate of substrate oxidation was regulated by the presence of inhibitors.
Incubation of inside-out membrane vesicles with malonate or with oxalate resulted in inhibition of succinate and D-lactate oxidase activities, respectively. Half-maximal inhibition was given by 2.5 mM malo- nate and by 0.075 mM oxalate. The loss of oxidase activity in the presence of these inhibitors could be correlated with the decreases in both the rate and the extent of quenching of 9-aminoacridine fluorescence by succinate and by D-lactate (Fig. 4A). Plots of succinate oxidase activity in the presence of varying concentrations of malonate, and of D-lactate oxidase activity in the presence of oxalate, versus the initial rate of fluorescence quenching were linear (Fig. 4 B).
The effect of Zn2' and 2-heptyl-4-hydroxyquino- line N-oxide on D-lactate. DL-dvcerol 3 - ~ h o s ~ h a t e .
succinate and NADH oxidase activities and on the initial rate of quenching of 9-aminoacridine fluor- escence were measured. Succinate oxidase activity was the most sensitive to inhibition by either Zn2+ or 2-heptyl-4-hydroxyquinoline N-oxide, half-maximal inhibition being obtained at 16 and 0.125 pM inhibitor, respectively. Concentrations of 40- 50 pM Zn2+ were required for half-maximal inhibition of NADH and DL-glycerol 3-phosphate oxidases, and of 3 - 5 pM 2-heptyl-4-hydroxyquinoline N-oxide for half-maxi- ma1 inhibition of NADH and D-lactate oxidases. D-Lactate oxidase and DL-glycerol 3-phosphate oxi- dase activities were more resistant than the other oxidases to Zn2+ and 2-heptyl-4-hydroxyquinoline N-oxide, respectively.
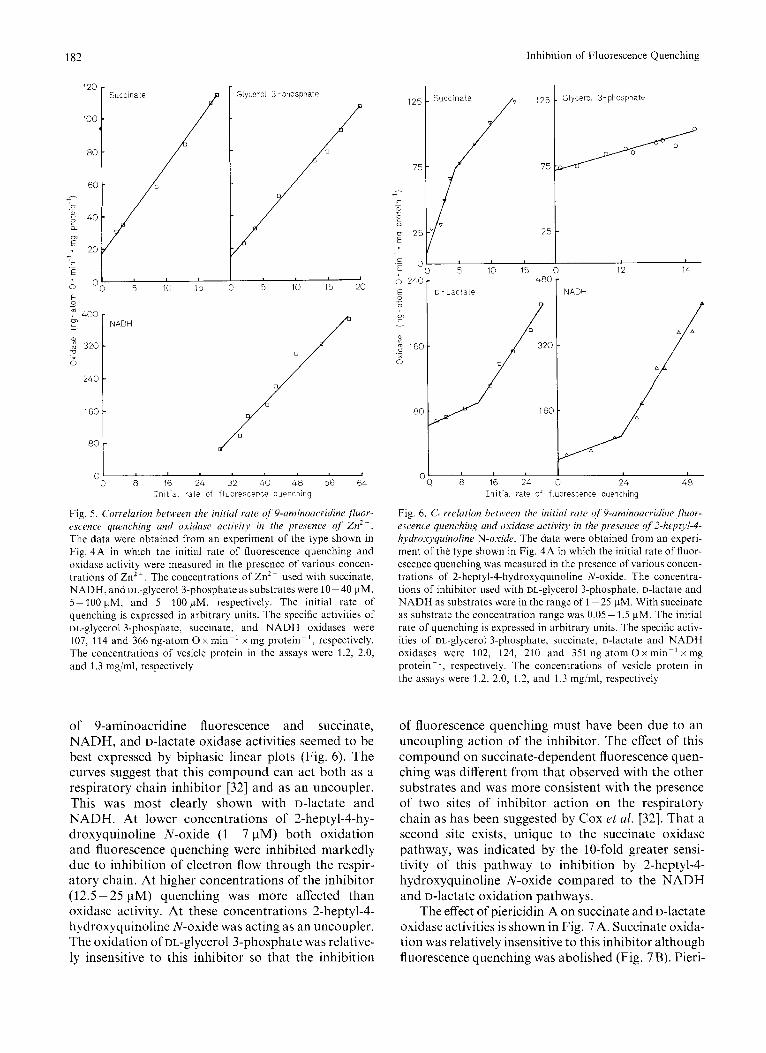
A correlation between the initial rate of fluor- escence quenching and the rate of oxidation of NADH, succinate and DL-glycerol 3-phosphate in the presence of ZnZ+ was suggested by the linear plots shown in Fig. 5. The inhibition by 50 pM ZnZ+ of 9-amino- acridine fluorescence quenching energized by suc- cinate, NADH, and DL-glycerol 3-phosphate could be prevented or reversed by the addition of I mM dithio- threitol. Although it is not clear if this was due to the relief of Zn2 + inhibited respiration only, the thiol decreased the inhibition of succinate oxidase from 82% to 25%.
With the exception of ~~-glycerol-3-phosphate, the relationship in the presence of 2-heptyl-4-hydroxy- auinoline N-oxide between the initial rate of quenching
182 Inhibition of Fluorescence Quenching
0 "
g ? 400 m i 1
2 3 2 0 n x ._ 0
2 4 0
160
8C
C
5 10 15
NADH
Glycerol 3 -phosphate
5 10 15 20
I I
8 16 2 4 3 2 40 48 56 6 4 I n i t i a l rate of fluorescence quenching
Fig. 5. Correlation between the initial rate of 9-aminoacridine ,fluor- escence quenching and oxidase activity in the pre.sence of Znz+. The data were obtained from an experiment of the type shown in Fig. 4A in which the initial rate of fluorescence quenching and oxidase activity were measured in the presence of various concen- trations of Zn2+. The concentrations of Zn2+ used with succinate, NADH, and oL-glycerol 3-phosphate as substrates were 10-40 pM, 5- 100 pM, and 5-100 pM, respectively. The initial rate of quenching is expressed in arbitrary units. The specific activities of DL-glycerol 3-phosphate, succinate, and NADH oxidases were 107, 114 and 366 ng-atom 0 x min-' x mg protein-', respectively. The concentrations of vesicle protein in the assays were 1.2, 2.0, and 1.3 mgiml, respectively
of 9-aminoacridine fluorescence and succinate, NADH, and D-lactate oxidase activities seemed to be best expressed by biphasic linear plots (Fig. 6). The curves suggest that this compound can act both as a respiratory chain inhibitor [32] and as an uncoupler. This was most clearly shown with D-lactate and NADH. At lower concentrations of 2-heptyl-4-hy- droxyquinoline N-oxide (1 - 7 pM) both oxidation and fluorescence quenching were inhibited markedly due to inhibition of electron flow through the respir- atory chain. At higher concentrations of the inhibitor (12.5 - 25 pM) quenching was more affected than oxidase activity. At these concentrations 2-heptyl-4- hydroxyquinoline N-oxide was acting as an uncoupler. The oxidation of DL-glycerol 3-phosphate was relative- ly insensitive to this inhibitor so that the inhibition
125 Glycerol 3-phosphate t
.- '0 5 10 15 0 12 14
6 24C
5 m
m i Y
a, 0)
," 16C .- 0
8 C
c
1 480
D - Lactate NADH
8 16 2 4 0 2 4 48 I n i t i a l rate of fluorescence quenching
Fig. 6. C, rrelation hetiveen the initial rate of 9-aminoucridine , jhor- escence quenching and oxidase activity in the presence of2-heptyl-4- hydro.xyquinoline N-oxide. The data were obtained from an experi- ment of the type shown in Fig. 4A in which the initial rate of fluor- escence quenching was measured in the presence of various concen- trations of 2-heptyl-4-hydroxyquinoline N-oxide. The concentra- tions of inhibitor used with DL-glycerol 3-phosphate, o-lactate and NADH as substrates were in the range of 1-25 pM. With succinate as substrate the concentration range was 0.05- 1.5 pM. The initial rate of quenching is expressed in arbitrary units. The specific activ- ities of DL-glycerol 3-phosphate, succinate, D-laCtate and NADH oxidases were 102, 124, 210 and 351 ng-atom 0 x min-' x mg protein- ', respectively. The concentrations of vesicle protein in the assays were 1.2, 2.0, 1.2, and 1.3 mgiml, respectively
of fluorescence quenching must have been due to an uncoupling action of the inhibitor. The effect of this compound on succinate-dependent fluorescence quen- ching was different from that observed with the other substrates and was more consistent with the presence of two sites of inhibitor action on the respiratory chain as has been suggested by Cox rt al. [32]. That a second site exists, unique to the succinate oxidase pathway, was indicated by the 10-fold greater sensi- tivity of this pathway to inhibition by 2-heptyl-4- hydroxyquinoline N-oxide compared to the NADH and D-lactate oxidation pathways.
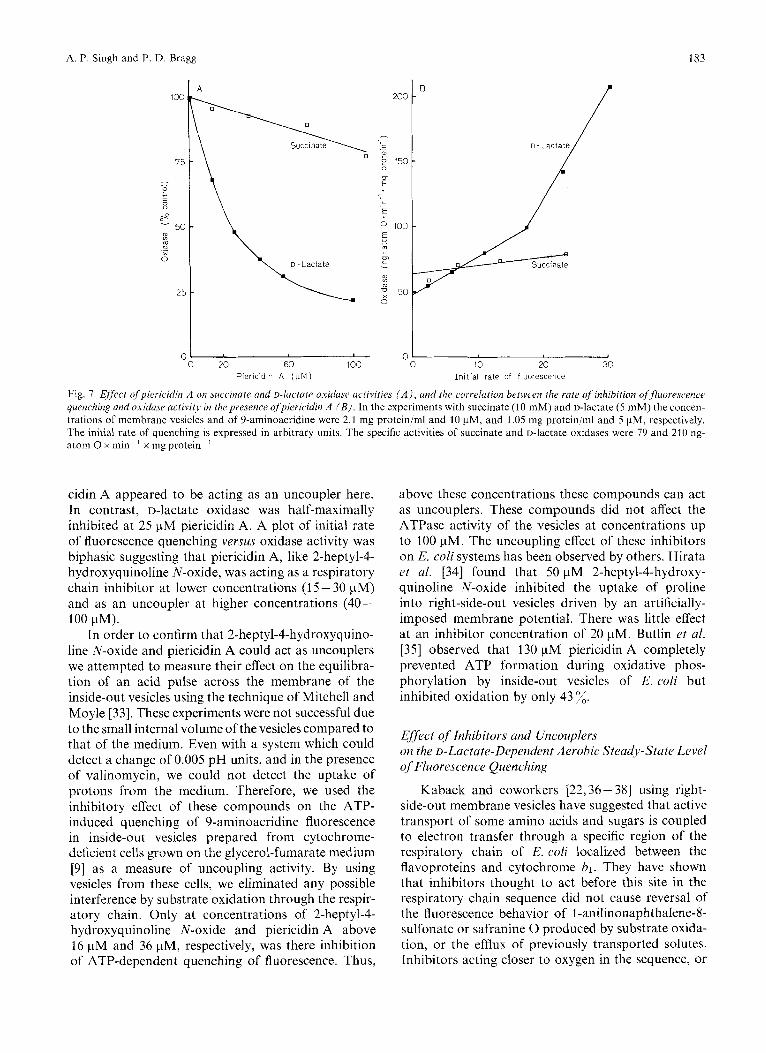
The effect of piericidin A on succinate and D-lactate oxidase activities is shown in Fig. 7A. Succinate oxida- tion was relatively insensitive to this inhibitor although fluorescence quenching was abolished (Fig. 7 B). Pieri-
A. P. Singh and P. D. Bragg 183
100
75
- - e c 0 " ."
50 % n_ 0
25
C C 20 60 100
Piericidin A (pM) 10 20 30
I n i t i a l rate of fluorescence
Fig. 7. Effeect ofpiericidin A on succinate and D-lactate oxiduse uctzvities ( A ) , and the correlation beween the rate of inhibition offluorescence quenching and oxiduse activity in thepresence ofpiericidin A ( B ) . In the experiments with succinate (10 mM) and D-lactate (5 mM) the concen- trations of membrane vesicles and of 9-aminoacridine were 2.1 mg protein/ml and 10 pM, and 1.05 mg protein/ml and 5 pM, respectively. The initial rate of quenching is expressed in arbitrary units. The specific activities of succinate and D-lactate oxidases were 79 and 210 ng- atom 0 x min-' x mg protein-'
cidin A appeared to be acting as an uncoupler here. In contrast, D-lactate oxidase was half-maximally inhibited at 25 pM piericidin A. A plot of initial rate of fluorescence quenching versus oxidase activity was biphasic suggesting that piericidin A, like 2-heptyl-4- hydroxyquinoline N-oxide, was acting as a respiratory chain inhibitor at lower concentrations (1 5 - 30 pM) and as an uncoupler at higher concentrations (40- 100 pM).
In order to confirm that 2-heptyl-4-hydroxyquino- line N-oxide and piericidin A could act as uncouplers we attempted to measure their effect on the equilibra- tion of an acid pulse across the membrane of the inside-out vesicles using the technique of Mitchell and Moyle [33]. These experiments were not successful due to the small internal volume of the vesicles compared to that of the medium. Even with a system which could detect a change of 0.005 pH units, and in the presence of valinomycin, we could not detect the uptake of protons from the medium. Therefore, we used the inhibitory effect of these compounds on the ATP- induced quenching of 9-aminoacridine fluorescence in inside-out vesicles prepared from cytochrome- deficient cells grown on the glycerol-fumarate medium [9] as a measure of uncoupling activity. By using vesicles from these cells, we eliminated any possible interference by substrate oxidation through the respir- atory chain. Only at concentrations of 2-heptyl-4- hydroxyquinoline N-oxide and piericidin A above 16 pM and 36 pM, respectively, was there inhibition of ATP-dependent quenching of fluorescence. Thus,
above these concentrations these compounds can act as uncouplers. These compounds did not affect the ATPase activity of the vesicles at concentrations up to 100 pM. The uncoupling effect of these inhibitors on E. coli systems has been observed by others. Hirata et al. [34] found that 50 pM 2-heptyl-4-hydroxy- quinoline N-oxide inhibited the uptake of proline into right-side-out vesicles driven by an artificially- imposed membrane potential. There was little effect at an inhibitor concentration of 20 pM. Butlin et al. [35] observed that 130 pM piericidin A completely prevented ATP formation during oxidative phos- phorylation by inside-out vesicles of E. coli but inhibited oxidation by only 43 x. Eflect o j Inhibitors and Uncouplers on the D-Lactate-Dependent Aerobic Steady-State Level of'Fluorescence Quenching
Kaback and coworkers [22,36- 381 using right- side-out membrane vesicles have suggested that active transport of some amino acids and sugars is coupled to electron transfer through a specific region of the respiratory chain of E. coli localized between the flavoproteins and cytochroine bl. They have shown that inhibitors thought to act before this site in the respiratory chain sequence did not cause reversal of the fluorescence behavior of l-anilinonaphthalene-8- sulfonate or safranine 0 produced by substrate oxida- tion, or the efflux of previously transported solutes. Inhibitors acting closer to oxygen in the sequence, or
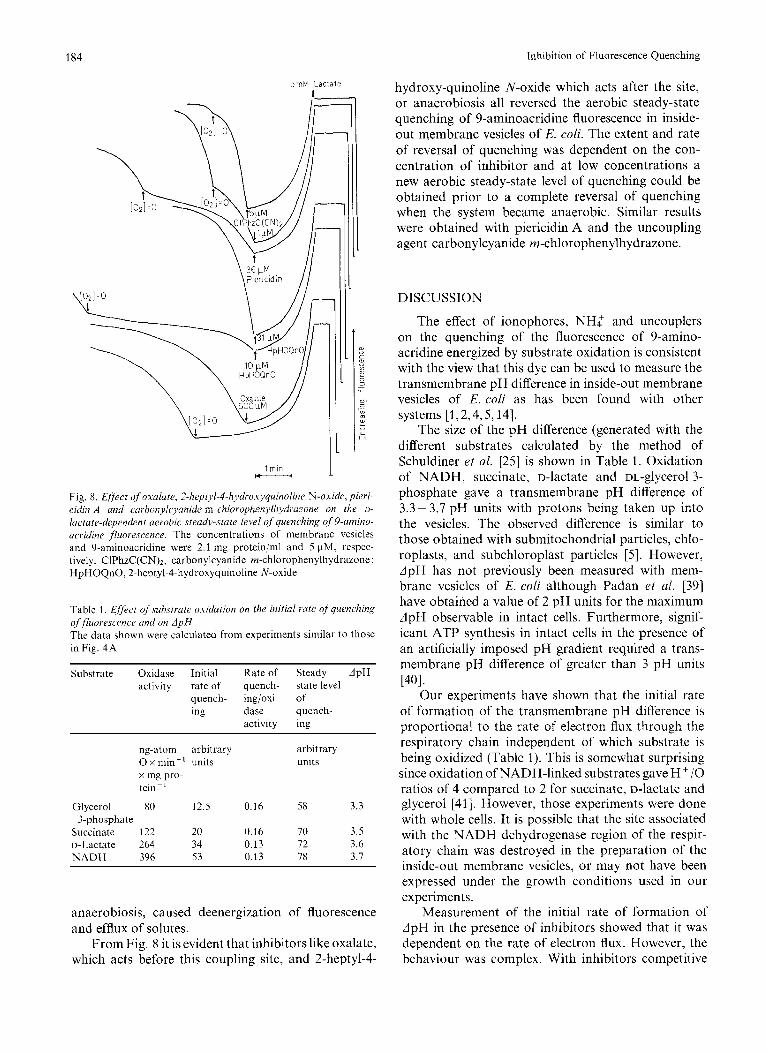
184 Inhibition of Fluorescence Quenching
\1
1 rnin H----r L
Fig. 8. E/ject of' oxalate, 2-heptyl-4-hydroxyquinoline N-oxide, pieri- cidin A and carbonykyanide m-chlorophenyli~~dl.al.one on the u- lactate-dependent aerobic .sceady-state level ?f quenching of 9-umino- ucridine fluorescence. The concentrations of membrane vesicles and 9-aminoacridine were 2.1 mg protein/ml and 5 pM, respec- tively. CIPhzC(CN)2, carbonylcyanide m-chlorophenylhydrazone; HpHOQnO, 2-heptyl-4-hydroxyquinoline N-oxide
Table 1. ,Eflect of suhstrale oxidation on the initial rate ofquenching offluorescetice and on A p H The data shown were calculated from experiments similar to those in Fig. 4 A
Substrate Oxidase Initial Rate of Steady dpH activity rate of quench- state level
quench- ing/oxi- of ing dase quench-
activity ing
ng-atom arbitrary arbitrary 0 x min-' units units x mg pro- tein ~ '
Glycerol 80 12.5 0.16 58 3.3 3-phosphate
Succinate 122 20 0.16 70 3.5 D-Lactate 264 34 0.13 72 3.6 N A D H 396 53 0.13 78 3.7
anaerobiosis, caused deenergization of fluorescence and efflux of solutes.
From Fig. 8 it is evident that inhibitors like oxalate, which acts before this coupling site, and 2-heptyl-4-
hydroxy-quinoline N-oxide which acts after the site, or anaerobiosis all reversed the aerobic steady-state quenching of 9-aminoacridine fluorescence in inside- out membrane vesicles of E. coli. The extent and rate of reversal of quenching was dependent on the con- centration of inhibitor and at low concentrations a new aerobic steady-state level of quenching could be obtained prior to a complete reversal of quenching when the system became anaerobic. Similar results were obtained with piericidin A and the uncoupling agent carbonylcyanide m-chlorophenylhydrazone.
DISCUSSION
The effect of ionophores, NH2 and uncouplers on the quenching of the fluorescence of 9-amino- acridine energized by substrate oxidation is consistent with the view that this dye can be used to measure the transmembrane pH difference in inside-out membrane vesicles of E. coli as has been found with other systems [1,2,4,5,14].
The size of the pH difference (generated with the different substrates calculated by the method of Schuldiner et al. [25] is shown in Table 1. Oxidation of NADH, succinate, D-lactate and DL-glycerol 3- phosphate gave a transmembrane pH difference of 3.3-3.7 pH units with protons being taken up into the vesicles. The observed difference is similar to those obtained with submitochondrial particles, chlo- roplasts, and subchloroplast particles [5]. However, ApH has not previously been measured with mem- brane vesicles of E. coli although Padan et al. 1391 have obtained a value of 2 pH units for the maximum ApH observable in intact cells. Furthermore, signif- icant ATP synthesis in intact cells in the presence of an artificially imposed pH gradient required a trans- membrane pH difference of greater than 3 pH units
Our experiments have shown that the initial rate of formation of the transmembrane pH difference is proportional to the rate of electron flux through the respiratory chain independent of which substrate is being oxidized (Table 1). This is somewhat surprising since oxidation of NADH-linked substrates gave H + / O ratios of 4 compared to 2 for succinate, D-lactate and glycerol 1411. However, those experiments were done with whole cells. It is possible that the site associated with the NADH dehydrogenase region of the respir- atory chain was destroyed in the preparation of the inside-out membrane vesicles, or may not have been expressed under the growth conditions used in our experiments.
Measurement of the initial rate of formation of ApH in the presence of inhibitors showed that it was dependent on the rate of electron flux. However, the behaviour was complex. With inhibitors competitive
~401.

A. P. Singh and P. D. Bragg 185
Right ~ side out vesic les Insine - o u t v e s i c i e i
.1 Oxamatt D ~ L a c t a t e
9-Gminoacridine , F'
dine
D - ILactate Oxalate
Fig. 9. Location of D-luctate ckelzydrogenase, polurity of proton lrunslocalion, and movement qffluoresctnt prohes in right-side-out and inside-out vesicles. See text for explanation. Ansyl, 1 -anilinonaphthalene-8-sulfonic acid. The filled areas indicate the location of D-lactate dehydro- genase
with the substrates there was a direct relationship between oxidase activity and the rate of fluorescence quenching, although the lines did not pass through the origin. This could be due to the presence of respiratory activity not coupled to the formation of a proton gradient. Zinc ions had a similar effect on succinate and DL-glycerol 3-phosphate oxidation. Here it is probable that the main site of inhibition is at the level of the dehydrogenases and probably involves thiol groups [42]. The second type of response which was observed is exemplified by the biphasic inhibition curves obtained by plotting the succinate, NADH or D-lactate oxidase activities measured in the presence of 2-heptyl-4-hydroxyquinoline N-oxide, or D-lactate oxidase with piericidin A, versus the initial rate of quenching. This behaviour is consistent with piericidin A and 2-heptyl-4-hydroxyquinoline N-oxide acting both as inhibitors of the respiratory chain and, at higher concentrations, as uncoupling agents. Finally, inhibition of fluorescence quenching with only slight effects on substrate oxidation as shown by piericidin A for succinate oxidase, and NH: and carbonylcyanide m-chlorophenylhydrazone for D-lactate oxidase, is typical of the action of an uncoupling agent. Thus, fluorescence quenching is a useful technique for the investigation of the action of inhibitors on the respir- atory chain.
Reeves et ul. [37] have examined the substrate- dependent quenching of the fluorescence of 1 -aniline- naphthalene 8-sulfonate in right-side-out membrane vesicles of E. coli. They found that the behaviour of l-anilinonaphthalene-8-sulfonate with substrates and inhibitors correlated nearly perfectly with the proper- ties of the D-lactate dehydrogenase-coupled transport systems of membrane vesicles. Thus, inhibitors such as 2-heptyl-4-hydroxyquinoline N-oxide which seem to act between the coupling site for transport in the respiratory chain and oxygen caused efflux of accu-
mulated solutes and reversed the energy-dependent quenching of 1-anilinonaphthalene-8-sulfonate fluor- escence. In contrast, inhibitors such as oxamate acting between D-lactate and the coupling site had little effect on accumulated solute and did not decrease energy- dependent quenching. However, both types of in- hibitors prevented the accumulation of solute and the quenching of fluorescence if added before the energiz- ing substrate [37]. In our experiments with inside-out membrane vesicles both oxalate, which like oxamate is a competitive inhibitor of D-lactate dehydrogenase, and 2-heptyl-4-hydroxyquinoline N-oxide decreased the D-lactate-dependent aerobic steady state level of quenching of 9-aminoacridine fluorescence.
The difference in the effect of oxalate on the quench- ing of 9-aminoacridine fluorescence in inside-out membrane vesicles, and of oxamate on fluorescence of l-anilinonaphthalene-8-sulfonate in right-side-out membrane vesicles, can be explained as follows (Fig. 9). On energization by oxidation of D-lactate, the interior of the inside-out membrane vesicle be- comes more acidic and a membrane potential, interior positive, is generated. The cationic weak base 9-amino- acridine will penetrate the membrane in its neutral form to enter the vesicle where it is protonated and its fluorescence is quenched. Thus, its distribution de- pends on the pH difference across the membrane [ 5 ] . In right-side-out vesicles l-anilinonaphthalene-8-sul- fonate is bound to the membrane and in this state is fluorescent. On energization of right-side-out vesicles by oxidation of D-hCtate protons are extruded from the vesicle and a membrane poteqtial, exterior posi- tive, is formed. The anionic l-anilinonaphthalene-8- sulfonate is released from the membrane into the medium in response to both the pH difference and the membrane potential [43] and its fluorescence is quench- ed. Oxamate must enter the right-side-out membrane vesicle to inhibit D-lactate dehydrogenase, the active

186
site of which is accessible on the inner face of the vesicular membrane [44]. When it is added prior to D-lactate sufficient oxamate will have penetrated into the vesicle to compete effectively with the substrate. Thus, the D-lactate-dependent quenching of 1 -aniline- naphthalene-8-sulfonate fluorescence is inhibited. Ad- dition of oxamate after D-lactate does not result in inhibition since oxamate is not able to build up in concentration within the vesicle sufficient rapidly to compete with the accumulated D-lactate for the active site of the dehydrogenase. In contrast, in inside-out membrane vesicles oxalate and D-lactate have im- mediate access to the active site of the D-lactate dehydrogenase and their relative concentrations are the same as in the medium. Thus, oxalate even when added after D-lactate is at a sufficiently high concentra- tion to successfully compete at the active site of the enzyme. It is probable that the different effects of oxamate on transport depending on whether it was added before or some time after D-lactate can be explained in the same way [45]. Our mechanism also explains the finding that in reconstituted membrane vesicles in which D-lactate dehydrogenase was on the outside of the vesicle the addition of oxamate caused efflux of preaccumulated solute [46,47]. Thus, the effects of oxamate on transport do not support mechanisms for active transport in which oxidation and reduction of the solute carrier is an essential part [36,38].
This work was supported by a grant from the Medical Research Council of Canada
REFERENCES
1. Rottenberg, H. & Grunwaid, T. (1972) Eur. J . Biochern. 25, 71 - 74.
2. Casadio, R., Baccarini-Melandri, A. & Melandri, B. A. (1974) Eur. J . Biochem. 47, 121 - 128.
3. Barsky, E. L., Bonch-Osmolovskaya, S. A,, Ostroumov, V. D., Samuilov, V. D. 6i Skulachcv, V . P. (1975) Biochim. Biophps. Acfa, 387, 388-395.
4. Rottenberg, H. & Lee, C.-P. (1975) Biochemistry, 14, 2675- 2680.
5. Rottenberg, H. (1975) J . Bioenergetics, 7, 61 - 74. 6. Nieuwenhuis, F. J. R. M., Kanner, B. I., Gutnick, D. L.,
Postma, P. W. & Van Dam. K. (1973) Bioehini. Biophys. Acta, 325, 62 - 71.
7. Van Thienen, G . & Postma, P. W. (1973) Biochim. Biophys. Acfa, 323, 429-440.
8. Haddock, B. A. & Downie, J. A. (1974) Biochem. J . 142,
9. Singh, A. P. & Bragg, P. D. (1975) Biuchim. Biophys. Acta,
10. Rosen, B. P. & Adler, L. W. (1975) Biochim. Biophys. Acta,
11. Tsuchiya, T. & Rosen, B. P. (1975) J . Biol. Chem. 250, 8404-
703 - 706.
396, 229-241.
387,23 - 36.
8415.
A. P
12.
13. 14.
15.
16.
17.
18. 19.
20.
21
22.
23 24
25
26
27
28
29
30 31 32
33
34
35
36
37
38
39
40
41
42 43
44 45
46
47
Singh and P. D. Bragg: Inhibition of Fluorescence Quenching
Kanner, B. I., Nelson, N. & Gutnick, D. L. (1975) Biochim.
Mitchell, P. (1966) Bid. Rev. 41, 445-502. Deamer, D. W., Prince, R. C. & Crofts, A. R. (1972) Biochim.
Sinioni, R. D. & Postma, P. W. (1975) Annu. Rev. Biochrm.
Niven, D. F. & Hamilton, W. A. (1974) Eur. J . Biochem. 44,
West, I . C. (1970) Biochem. Biophys. Res. Commun. 41, 655-
West, I. C. & Mitchell, P. (1973) Biochet??. J . 132, 587-592. Henderson, P. J. F. & Skinner, A. (1974) Trans. Biochem. Soc.
Gutowski, S. J . & Rosenberg, H. (1975) Biochem. J . 152,
Essenberg, R. C. & Kornberg, H. L. (1975) J . Biol. Chem. 250,
Kaback, H. R. & Hong, J. (1973) CRC Critical Rev. Microbiol.
Futdi, M. (1974) J . Bacteriol. 120, 861 - 865. Bragg, P. D., Davies, P. L. & Hou, C. (1974) Biochem. Biophys.
Schuldiner, S., Rottenberg, H. & Avron, M. (1972) Eur. J .
Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J .
Hunter, G. R. & Brierley, G. P. (1969) Biochim. Biophys. Acta,
Kim, I. C. & Bragg, P. D. (1971) Can. J. Biochem. 49, 1098-
Barnes, E. M. & Kaback, H. R. (1971) J . Biol. Chem. 246,
Harold, F. M. (1972) Bacteriol. Rev. 36. 172-230. Greville, G. D. (1969) Curr. Top. Bioenergefics, 3, 1 - 78. COX, G. B., Newton, N. A., Gibson, P., Snoswell, A. M. &
Hamilton, J. A. (1970) Biochem. J . 117, 551-562. Mitchell, P. & Moyle, J. (1967) in Biochemistry ofMitochondriu
@later, E. C., Kaniugd, Z. & Wojtczak, L., eds) pp. 53-74. Academic Press, New York.
Hirata, H., Altendorf, K. & Harold, F. M. (1973) Proc. Natl Acad. Sci. U.S.A. 70, 1804-1808.
Butlin, J. D., Cox, G. B. & Gibson, F. (1971) Biochem. 1. 124,
Kaback, H. R. & Barnes, E. M. (1971) J . B id . Chenz. 246,
Reeves, J. P., Lombardi, F. J. & Kaback, H. R. (1972) J . B id .
Schuldiner, S . & Kaback, H. R. (1975) Biochemisiry, 14,5451 -
Padan, E., Zilberstein, D. & Rollenberg, H. (1976) Eur. J .
Wilson, D. M., Alderete, J. F., Maloney. P. C. & Wilson, T. H.
Lawford, H. G . & Haddock, B. A. (1973) Biochem. J . 136,
Kasahara, M. &Anraku, Y. (1974) J . Biochem. 76,967 -976. Barsky, E. L., Bonch-Osmolovskaya, E. A., Ostroumov, S. A.,
Samuilov, V. D. & Skulachev, V. P. (1975) Biochim. Biophys. Acta, 387, 388-395.
Biophys. Acta, 396, 347- 359.
Biophy,r.. Acta, 274, 323 - 335.
44,523 - 554.
517-522.
661.
2, 543 - 545.
647 - 654.
939 - 945.
2, 333- 376.
Res. Conzmun. 47, 1248- 1255.
Biochem. 25,64 - 70.
(1951) J . Biol. Chem. 193, 265-275.
180, 68 - 80.
1104.
5518- 5522.
75-81.
5523-5531.
Chem. 247,6204-6211.
5461.
Biochem. 63, 533-541.
(1976) J . Bacteriol. 126, 327-337.
21 7 - 220.
Futai, M. & Tanaka, Y. (1975) J . Bacteriol. 124, 470-475. Altendorf, K., Hirata, H. &Harold, F. M. (1975)J. Bioi. Chen7.
Reeves, J. P., Hong, J.-S. & Kaback, H. R. (1973) Proc. Nut/
Futai, M. (1974) Biochemistr.y, 13, 2327-2333.
250, 1405- 1412.
Acad. Sci. U.S.A. 70, 1917-1921.
Akhand P. Singh and P. D. Bragg, Department of Biochemistry, University of British Columbia, Vancouver. British Columbia, Canada, V6T lW5
Related Documents











