The Pennsylvania State University The Graduate School College of Medicine EFFECT OF EXERCISE MUSCLE TEMPERATURE ON RENAL AND SYMPATHETIC RESPONSES TO ISOMETRIC EXERCISE IN HUMANS A Dissertation in Integrative Biosciences by Nathan T. Kuipers 2008 Nathan T. Kuipers Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy August 2008

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Pennsylvania State University
The Graduate School
College of Medicine
EFFECT OF EXERCISE MUSCLE TEMPERATURE ON RENAL AND
SYMPATHETIC RESPONSES TO ISOMETRIC EXERCISE IN HUMANS
A Dissertation in
Integrative Biosciences
by
Nathan T. Kuipers
2008 Nathan T. Kuipers
Submitted in Partial Fulfillment of the Requirements
for the Degree of
Doctor of Philosophy
August 2008

ii
The dissertation of Nathan T. Kuipers was reviewed and approved* by the following: Chester A. Ray Professor of Medicine, and Cellular and Molecular Physiology Thesis Advisor Chair of Committee Lawrence I. Sinoway Professor of Medicine James A. Pawelczyk Associate Professor of Kinesiology Thomas C. Pritchard Associate Professor of Neural and Behavioral Sciences Peter J. Hudson
Professor of Biology Director of the Graduate Program in Integrative Biosciences
*Signatures are on file in the Graduate School

iii
Abstract
Abstract Chapter 2 – The purpose of the present study was to examine
the effect of heating and cooling the forearm muscles on renal vascular
responses to ischemic isometric handgrip (IHG). It was hypothesized that
heating and cooling the forearm would augment and attenuate renal vascular
responses to IHG, respectively. Renal vascular responses to IHG were studied
during forearm heating at 39 ºC (n=15, 26±1 yr) and cooling at 26 ºC (n=12, 26±1
yr). For a control trial subjects performed the experimental protocol while the
forearm was normothermic (~34 ºC). Muscle temperature (measured by
intramuscular probe) was controlled by changing the temperature of water
cycling through a water-perfused sleeve. The experimental protocol was: 3-min
baseline, 1 min of ischemia, ischemic IHG to fatigue, and 2 min of postexercise
muscle ischemia. At rest, renal artery blood velocity (RBV; Doppler ultrasound)
and renal vascular conductance (RVC; RBV / mean arterial blood pressure) were
not different between normothermia and the two thermal conditions. During
ischemic IHG there were greater decreases in RBV and RVC in the heating trial.
However, RBV and RVC were similar during postexercise muscle ischemia
during heating and normothermia. During cooling, RVC decreased less
compared to normothermia during ischemic IHG. During postexercise muscle
ischemia, RVC was greater during cooling compared to the normothermic trial.
These results indicate that heating augments mechanoreceptor-mediated renal
vasoconstriction whereas cooling blunts metaboreceptor-mediated renal
vasoconstriction.

iv
Abstract Chapter 3 – The purpose of the study was to determine the
interactive effect of aging and forearm muscle heating on renal blood flow and
muscle sympathetic nerve activity during ischemic isometric handgrip. A tube
lined water-perfused sleeve was used to heat the forearm in twelve young (27±1
yr) and nine older (63±1 yr) subjects. Ischemic isometric handgrip was performed
before and after heating. Muscle temperature (intramuscular thermistor) was
34.3±0.2 ºC and 38.7±0.1 ºC during normothermia and heating, respectively. At
rest, heating had no effect on renal blood velocity (Doppler ultrasound) or renal
vascular conductance in either group (young, n = 12; older, n = 8). During
ischemic isometric handgrip, heating caused a significantly greater decrease in
renal blood velocity and increase in renal vasoconstriction in both groups
compared to normothermia. However, the increase in renal vasoconstriction
during heating was greater in the older subjects (18±3% at fatigue) compared to
the young (8±3% at fatigue). Unlike the younger group, heating increased renal
vasoconstriction during postexercise muscle ischemia in the older group. During
handgrip, heating elicited comparable increases in muscle sympathetic nerve
activity responses in both groups (young, n = 12; older, n = 6). The interaction of
aging and muscle heating did not alter muscle sympathetic nerve activity
responses to exercise. In summary, aging augments renal vascular responses to
ischemic isometric handgrip during heating of the muscle that is not associated
with greater muscle sympathetic nerve activity.

v
Abstract Chapter 4 – This study examined if ACE-inhibition alters central
hemodynamic, vascular, and sympathetic responses to isometric handgrip with a
normothermic or hyperthermic forearm. Eight male (25±2yr) subjects were given
an ACE-inhibitor (20 mg of quinapril) or placebo on alternate visits. Subjects
performed ischemic isometric handgrip and postexercise muscle ischemia with
the forearm muscle normothermic (~35±0.3 ºC, intramuscular probe) and
hyperthermic (~38±0.3 ºC). Blood pressure, heart rate, renal blood flow velocity
(Doppler ultrasound), calf blood flow, and muscle sympathetic nerve activity
(MSNA) were recorded throughout all studies. Quinapril lowered mean arterial
blood pressure (~8±3 mmHg) at baseline, but did not alter cardiovascular and
MSNA responses to normothermic isometric handgrip or postexercise muscle
ischemia. Exercise with a hyperthermic forearm augmented increases in blood
pressure, renal vasoconstriction, and MSNA during both treatments, but there
were no significant differences between drug treatments. Likewise, central
hemodynamic, vascular, and MSNA responses to postexercise muscle ischemia
during heating were not significantly different between drug treatments. These
findings suggest ACE-inhibitor-induced changes in the renin-angiotensin and
kallikrein-kinin systems do not alter central hemodynamic, vascular, and MSNA
responses to isometric exercise in healthy humans.

vi
TABLE OF CONTENTS LIST OF FIGURES …………………………………………………...... ix LIST OF TABLES ……………………………………………………..... xii ACKNOWLEDGMENTS ……………………………………………….. xiii Chapter 1 Introduction ……………………………………………….. 1 1.1 Introduction ……………………………………………… 1
Do Muscle Afferents Contribute to Cardiovascular 1.2 Responses to Exercise? ………………………………..
3
Exercise Pressor Reflex and Local Muscle 1.3 Temperature ……………………………………………..
6
Influence of Temperature on Exercise-Induced 1.4 Changes in Peripheral and Renal Blood Flow ………
8
Aging, the Exercise Pressor Reflex, Heat, and 1.5 Renal Blood Flow ……………………………………….
11
Influence of ACE-inhibition on Cardiovascular and 1.6 MSNA Responses to Normothermic and Hyperthermic
13
Isometric Exercise ………………………………………. 1.7 References ……………………………………………… 16
Changes in Forearm Muscle Temperature Alter Renal Chapter 2 Vascular Responses to Isometric Handgrip ………….
20
2.1 Introduction ………………………………………………. 20 2.2 Methods ………………………………………………….. 21 2.2.1 Subjects …………………………………..
. 21
2.2.2 Experimental Design …………………….. 22 2.2.3 Experimental Protocol …………………… 23 2.2.3 Measurements …………………………… 23 2.2.4 Data Analysis …………………………….. 25 2.3 Results ……………………………………………………... 25 2.3.1 Heating Study ……………………………. 25 2.3.2 Cooling Study …………………………….. 26 2.4 Discussion ………………………………………………... 28 2.5 References ……………………………………………...... 35 Chapter 3 Renal Vasoconstriction During Isometric Handgrip:
Interactive Effect of Aging and Local Muscle Heating… 46
3.1 Introduction ………………………………………………. 46 3.2 Methods ………………………………………………....… 48

vii
3.2.1 Subjects …………………………………….. 48 3.2.2 Experimental Design ………………………. 48 3.2.3 Experimental Protocol ……………………… 49 3.2.4 Measurements ……………………………… 3.2.4 Data Analysis ………………………………. 51 3.3 Results …………………………………………………… 53 3.3.1 Baseline …………………………………….. 3.3.2 Exercise …………………………………….. 3.4 Discussion ……………………….………………………. 55 3.5 References ………………………………………………. 61
ACE-Inhibition Does Not Alter Sympathetic and Vascular Responses to Isometric Exercise During Forearm Heating …………………………………………
Chapter 4
74
4.1 Introduction ………………………………………………... 76 4.2 Methods …………….……………………………………… 77 4.2.1 Subjects …………………………………………. 77 4.2.2 Experimental Design …………………………… 78 4.2.3 Measurements ………………………………….. 79 4.2.4 Statistical Analysis ……………………………… 81 4.3 Results ……………………………………………………… 82 4.4 Discussion ……………………….………………….……… 85 4.5 References …………………………………………..…….. 90 Chapter 5 Conclusions ……………………………………………… 101 5.1 Introduction ………………………………………………. 101 5.2 Significance of Current Finding ………………………… 102 5.3 Perspectives and Future Directions …………………… 104 5.4 Summary ………………………………………………… 108 5.5 References ………………………………………………. 109 Appendix A: Subject Consent Form: Studies 1 and 2 ……………… 112 Appendix B: Subject Consent Form: Study 3 ………………………. 120 Appendix C: Raw Data For Young Normothermia &
Heating Studies ………………………………………….. 131
Appendix D: Raw Data For Young Normothermia & Cooling Studies …………………………………………..
157
Appendix E: Raw Data For Older Normothermia & Heating Studies …………………………………………..
178

viii
Appendix F: Raw Data For ACE-inhibitor Study: Drug and Placebo Trials During Normothermia and Heating ……………..
224
Appendix F: Blood Data For ACE-inhibitor Study: Drug and Placebo Trials During Normothermia and Heating …………….. 261

ix
LIST OF FIGURES
Figure 2.1: Changes from baseline in mean arterial blood pressure and heart rate during exercise and postexercise muscle ischemia during forearm
heating and normothermia. ………………………..... 40 Figure 2.2: Changes from baseline in renal artery blood flow
velocity and renal vascular conductance during exercise and postexercise muscle ischemia during
forearm heating and normothermia. ……………….. 41 Figure 2.3: Changes from baseline in calf vascular conductance
during exercise and postexercise muscle ischemia
during forearm heating and normothermia. ………. 42 Figure 2.4: Changes from baseline in mean arterial blood
pressure and heart rate during exercise and postexercise muscle ischemia during forearm
cooling and normothermia. …………………………. 44 Figure 2.5: Changes from baseline in renal artery blood velocity
and renal vascular conductance during exercise and postexercise muscle ischemia during forearm
cooling and normothermia. …………………………. 45 Figure 2.6: Changes from baseline in calf vascular conductance
during exercise and postexercise muscle ischemia
during forearm cooling and normothermia. ………. 46 Figure 3.1: Ratings of perceived exertion during exercise in the
normothermic and heated trials in the young and
older subjects. ………………………………………… 67 Figure 3.2: Changes in mean arterial blood pressure from
baseline during forearm heating and normothermia during exercise and postexercise muscle ischemia
in the young and older groups. ……………….......... 68 Figure 3.3: Changes in renal blood velocity from baseline during
forearm heating and normothermia during exercise and postexercise muscle ischemia in the
young and older groups. …………………………….. 69 Figure 3.4: Percent changes in renal vascular conductance from
baseline during forearm heating and normothermia

x
during exercise and postexercise muscle ischemia in the young and older groups. ……………………. 70 Figure 3.5: Renal blood velocity and renal vascular conductance
differences between heating and
normothermia in both age groups at fatigue. …….. 71 Figure 3.6: Percent changes in calf vascular conductance from
baseline during forearm heating and normothermia during exercise and postexercise muscle ischemia
in the young and older groups. ……………………… 72 Figure 3.7: Changes in muscle sympathetic nerve activity burst
frequency from baseline during forearm heating and normothermia during exercise and postexercise
muscle ischemia in the young and older groups. .... 73 Figure 3.8: Changes in total muscle sympathetic nerve activity
from baseline during forearm heating and normothermia during exercise and postexercise
muscle ischemia in the young and older groups. … 74 Figure 3.9: Percent changes in total muscle sympathetic nerve
activity from baseline during forearm heating and normothermia during exercise and postexercise
muscle ischemia in the young and older groups. … 75 Figure 4.1: Changes in mean arterial blood pressure during
exercise and postexercise muscle ischemia for the quinapril and placebo treatments with the forearm
normothermic or heated. ………………………….... 96 Figure 4.2: Changes in renal vascular conductance during
exercise and postexercise muscle ischemia for quinapril and placebo treatments with the forearm
normothermic or heated. ……………………………. 97 Figure 4.3: Changes in calf vascular conductance during
exercise and postexercise muscle ischemia for quinapril and placebo treatments with the forearm
normothermic or heated. …………………………… 98 Figure 4.4: Changes in muscle sympathetic nerve activity burst
frequency during exercise and postexercise muscle ischemia for the quinapril and placebo treatments
with the forearm normothermic or heated. ………... 99

xi
Figure 4.5 Changes in total muscle sympathetic nerve activity
during exercise and postexercise muscle ischemia during quinapril and placebo treatments with the
forearm normothermic or hyperthermic. …………. 100

xii
LIST OF TABLES
Baseline measurement during normothermia Table 2.1: heating. ………………………………………………………………….
39
Baseline measurement during normothermia and Table 2.2: cooling. ………………………………………………………………….
43
Baseline measurements in the young group during normothermia Table 3.1: and heating. ……………………………………………………………..
65
Baseline measurements in the older group during normothermia Table 3.2: and heating. ……………………………………………………………..
66
Baseline measurements during the drug trials during normothermia Table 4.1: and forearm heating. …………………………………………………...
94
Baseline measurements during the placebo trials during Table 4.2: normothermia and forearm heating. …………………………………
95

xiii
ACKNOWLEDGEMENTS The author would like to thank the following individuals for their guidance,
support, encouragement, and participation throughout this endeavor.
Doctoral Committee
Dr. Chester Ray Dr. Lawrence Sinoway Dr. James Pawelczyk Dr. Thomas Pritchard Experimental Assistance Charity Sauder Thad Wilson Damian Dyckman Kevin Monahan Matthew Kearney GCRC Staff Amy Fogelman Cardiology Research Group Erin Muldoon Family and Friends Heidi Kuipers The Epps Family William and Marie Kuipers The Belmont Family Ric and Shelly Hoyt The Melleby Family Zes and Vicki Kuipers Nancy Hoffmann Research Volunteers The author would like to pay special thanks to all the subjects that participated in this study. Without their participation this research endeavor could not have been completed.

1
Chapter 1
Introduction
1.1 Introduction
The optimal temperature range of the human body must be maintained or
severe pathological problems develop. Heat stress and exercise severely affect
the ability to maintain core temperature, and when exercise and heat stress are
combined, this challenge is multiplied significantly. The cardiovascular system
plays an integral part in heat dissipation and exercise performance by directing
blood to heat dissipating tissues and working muscles. However, the
cardiovascular system has a limited amount of blood to circulate to these tissues.
The combined challenge of heat stress and exercise creates a situation where
blood-pumping capacity of the heart (i.e., cardiac output) is insufficient.
Therefore, the body must limit heat loss to sustain performance or limit
performance to preserve heat loss.
The physiological responses to exercise include increases in blood pressure,
heart rate, muscle sympathetic nerve activity (MSNA), and vasoconstriction in
inactive tissues. This patterned response is referred to as the exercise pressor
reflex. Reflexes that control the exercise pressor reflex include central
command, the baroreflex, and muscle afferents. How these reflexes are
influenced by thermal stress, with their ability to alter cardiovascular control,
remains equivocal. There is evidence that changes in local muscle temperature
alter blood pressure and MSNA responses to exercise in humans (72, 73);
however, the various changes in systemic blood flow that produce the altered

2
pressor changes during thermal stress are unknown. Therefore, the first goal of
this project is to examine the influence of local muscle temperature on muscle
afferent control of renal blood flow during exercise. The renal vasculature will be
examined because of the large percentage of cardiac output it receives at rest
and its important role in regulation of blood pressure and body water (80).
Aging is associated with changes in cardiovascular responses to exercise in
the heat. The exact cause of age-related changes remains equivocal, but may
include changes in how local muscle temperature influences the exercise pressor
reflex. Therefore, the second goal of the present work is to examine if local
muscle temperature influences cardiovascular and sympathetic responses to
exercise. Because of the importance of the renal vascular in controlling blood
pressure and blood volume during exercise and heat stress, and because aging
influences these responses, this project will also examine the influence of aging
and local muscle temperature on renal vascular responses to exercise.
The last goal is to examine if inhibiting angiotensin converting enzyme (ACE)
alters how limb temperature influences cardiovascular and MSNA responses to
isometric handgrip. ACE-inhibitor treatment alters the renin-angiotensin and the
kallikrein-kinin systems, which might influence central hemodynamic, vascular,
and MSNA responses to isometric exercise.
To help understand the importance of these studies, I will first review muscle
afferent physiology and how exercising muscle temperature might effect
cardiovascular, renal blood flow, and MSNA responses to exercise.

3
1.2 Do Muscle Afferents Contribute to Cardiovascular Responses to
Exercise?
The possibility of a reflex originating from the exercising muscle to increases
arterial blood pressure during exercise was first proposed by Alam and Smirk (1).
Alam and Smirk observed that when blood flow was occluded to an exercising
limb during and after exercise, a greater increase in arterial blood pressure was
observed than when flow remained constant. They concluded that this response
was mediated by accumulation of metabolic byproducts that could not leave the
muscle during circulatory occlusion. The authors believed that during exercise,
when blood flow into the working muscle was inadequate, this reflex increased
perfusion pressure to augment blood flow into the working muscle.
Coote et al. (11) first demonstrated that the exercise pressor reflex was a
neuronally-mediated reflex originating in the exercising muscle. Coote et al.
studied blood pressure responses during electrically stimulated skeletal muscle
contraction in both decerebrate and anesthetized cats. When the investigators
pharmacologically inhibited muscle contraction or when dorsal roots L6-S1 were
sectioned, the pressor response was completely abolished. In contrast, articular
proprioreceptor and vagi nerves ablation did not alter the exercise pressor
response. Collectively, these ablation studies indicated that the response was
initiated by muscle afferents.
McCloskey and Mitchell (53) demonstrated that group III and IV sensory
afferents mediated exercise pressor reflex and not group I and II afferents.
McCloskey and Mitchell found that transecting the dorsal roots of the spinal cord

4
abolished the normal pressor responses to electrically-induced muscle
contraction. Moreover, anodal block of group I and II muscle afferents did not
block the pressor response to exercise, while local anesthetic block of group III
and IV muscle did. In a subsequent study by McCloskey et al. (52), vibrating the
muscle to stimulate the muscle spindle, a type Ia fiber, did not elicit an exercise
like pressor response, further supporting the concept that group III and IV
afferents were responsible for mediating the exercise pressor reflex.
Anatomical positioning of group III and IV fibers provides further evidence
that the afferents can contribute to the exercise pressor reflex. Using the Achilles
tendon of the cat as a model, Andres et al. (2) characterized the anatomical
positioning of the two afferent groups. Five types of group III afferents were
observed. Of the five types, one was located on the wall of venous vessels, one
on lymphatic vessels, two other types ended in the connective tissue of the
peritoneum externum and internum, and the fifth projected to the endoneurium.
Group IV afferents terminated adjacent to blood and lymphatic vessels. In the
muscle tissue, group III and IV muscle afferents mimic this distribution as well
(29, 99).
Location of these group III and IV afferents gives them the ability to sense
changes in connective tissue shape and chemical make up of the interstitial fluid.
Group III muscle afferents are largely thought to be responsive to mechanical
deformation of the muscle because of their proximity to muscle connective tissue.
Group IV afferents may be responsive to byproducts of muscle metabolism
because of their nearness to muscle vasculature. To demonstrate this, Kaufman

5
et al. (32) recorded cat group III and IV muscle afferent activity during muscle
contraction. During contraction, group III muscle afferent firing occurred early
and before changes in the muscle milieu, suggesting that the afferents were
responding to changes in muscle shape. In contrast, group IV muscle afferent
firing onset occurred later during contraction as metabolite concentration
increased. Moreover, group IV afferents were stimulated by chemical injections
into the muscle. The latter two findings indicated that metabolic by-products of
contraction were stimulating Group IV afferents. Based on these results the
authors concluded that group III muscle afferents were responsive to mechanical
deformation in the muscle while group IV muscle afferents were activated by
chemical changes in the interstitial fluid of the muscle. Because of their
propensity for being sensitive to mechanical changes in the muscle, group III
afferents are often termed mechanoreceptors. Group IV afferents are often
termed metaboreceptors because of their chemical sensing properties. Finally,
there was evidence that some group III muscle afferents were responsive to
metabolic byproducts and some group IV muscle afferents were responsive to
mechanical stimulation.
Studies have found several factors can alter both group III and IV muscle
afferent function. For example, bradykinin (32, 54), potassium (33, 54), lactic
acid (77, 95), arachidonic acid (77), and ATP (21, 46) all influence muscle
afferent physiology. Temperature is another factor that influences muscle
afferents and cardiovascular and MSNA responses to exercise (72, 73). To

6
better understand the influence of muscle temperature on the muscle afferents, I
will first review what is known from animal and humans studies.
1.3 Exercise Pressor Reflex and Local Muscle Temperature
Animal studies indicate that besides being sensitive to mechanical and
chemical stimuli, group III and IV muscle afferents are sensitive to thermal
stimulation. Hertel et al. (27) recorded single afferent activity, first at
normothermic temperatures in the gastrocnemius muscle of the cat, and then at
increased and decreased temperatures (± 6-8 ºC). Out of 49 group IV afferents
tested, 14 were sensitive to heat and 11 were sensitive to cold stimulation. For
the 10 group III afferents tested, 3 were sensitive to heat and 3 were sensitive to
cold. Kumazawa and Mizumura (40) studied thermal receptive properties of
group III and IV afferents in the dog. Applying radiant heat to the gastrocnemius
muscle increased the discharges of 19 out of 30 group IV muscle afferents and 6
out of 6 group III muscle afferents. Kumazawa and Mizumura (40) found that a
mean temperature of 43.1 ºC evoked discharges from group IV afferents while a
mean temperature of 41 ºC evoked discharges from group III muscle afferents.
Hertel et al. (27) concluded that the thermosensitivity of these afferents might
contribute to exercise-induced changes in cardiovascular and respiratory function
observed during exercise.
Unfortunately, in both animal studies the investigators did not report the
influence of muscle temperature changes on the cardiovascular and sympathetic
responses to exercise. Evidence that muscle temperature can influence group III
and IV muscle afferent control of cardiovascular function during exercise comes

7
from human studies. Ray and Gracey (72) studied cardiovascular and MSNA
responses to isometric handgrip during forearm heating in healthy young
subjects. The investigators found that local heating augmented the increase in
mean arterial blood pressure in response to isometric handgrip. The increase in
arterial blood pressure corresponded to a greater increase in MSNA during
handgrip that occurred within 30 s of exercise onset. At fatigue and during
postexercise muscle ischemia, no differences in mean arterial blood pressure or
MSNA were observed. Because differences in activity were measured within the
first 30 s of exercise when mechanoreceptor-mediated cardiovascular responses
would be greatest, and not during postexercise muscle ischemia when
metaboreceptors activation would be most prominent, Ray and Gracey
concluded that heating augmented sensitivity of mechanically sensitive muscle
afferents in the forearm.
Ray et al. (73) subsequently cooled the forearm, and elicited responses to
isometric handgrip that were different from the response to heating. Ray et al.
found cooling the forearm did not alter arterial blood pressure responses to
exercise but rather blunted the increase in heart rate. Furthermore, compared to
normothermia, forearm cooling delayed the exercise-induced increase in MSNA
after 1 minute of exercise. At fatigue and during postexercise muscle ischemia,
cardiovascular and MSNA responses were not different between temperatures.
Because the delay was observed after 1 min of exercise, Ray et al. concluded
that muscle cooling delayed metaboreceptor-mediated changes in cardiovascular
and MSNA responses to exercise.

8
In summary, limb temperature can alter muscle afferent control of blood
pressure and sympathetic responses to exercise (27, 40). Increasing muscle
temperature augments the increase in arterial blood pressure and MSNA to
exercise, and the response is mediated by an increase in sensitivity of the
muscle mechanoreceptors (72). Muscle cooling delays increases in MSNA
during exercise in humans, which is mediated by a delay in activation of the
muscle metaboreceptors (73).
1.4 Influence of Muscle Temperature on Exercise-Induced Changes in
Peripheral and Renal Blood Flow
Vasoconstriction of non-exercising muscles and the visceral vascular beds
is an important mechanism used to meet the pressure and flow demands
necessary to perfuse the muscle and skin during exercise and heat stress (79,
80). This next section will summarize some of this research and introduce the
first studies in the present body of work.
Isometric exercise in cats and dogs is characterized by reductions in blood
flow to the kidneys (12, 14); these responses might be mediated by group III and
IV muscle afferents (97). Victor et al. (97) found muscle contractions increased
renal sympathetic nerve activity in less than one second from contraction onset
and that renal sympathetic nerve activity synchronized with intermittent
contractions. The early onset of changes in renal sympathetic nerve activity
indicated that renal sympathetic nerve activity was controlled by muscle
mechanoreceptors and not metaboreceptors.

9
Middlekauf et al. (55) examined the influence of the muscle afferents on
renal blood flow during exercise in humans. Dynamic positron emission
tomography was used to measure renal cortical blood flow during isometric
handgrip onset and during postexercise muscle ischemia. Middlekauf et al.
found that renal cortical blood flow decreased in response to isometric handgrip
when central command and mechanoreceptors would be most active, and during
postexercise muscle ischemia when metaboreceptor activation would be
greatest. The investigators concluded that exercise-induced renal
vasoconstriction was controlled by central command and/or the muscle
mechano- and metaboreceptors, but the exact contribution of each remained
unclear.
Momen et al. (58) provided a better understanding of the role of central
command and the muscle mechano- and metaboreceptors in controlling renal
blood flow in humans during exercise. Momen et al. studied renal vascular
responses to different exercise paradigms using Doppler ultrasound because of
its time resolution capacities. During the first paradigm, subjects performed
handgrip to fatigue and postexercise muscle ischemia. Renal vascular
resistance increased throughout exercise and postexercise muscle ischemia;
during postexercise muscle ischemia however, renal vascular resistance
decreased from fatigue, suggesting that the muscle metaboreceptors had a
greater role in controlling renal vasoconstriction during exercise than the muscle
mechanoreceptors. During the next paradigm, the investigators examined renal
vascular responses during the first 15 s of handgrip and found that renal vascular

10
resistance increased within 6-10 s and 11- 15 s of exercise onset. These time
points occur within the expected latency of mechanoreceptor-mediated
responses in humans (26). In another paradigm, the investigators found that
renal vascular resistance did increase during involuntary muscle contraction
induced by electrical stimulation, which suggested that central command was not
playing a role in exercise-induced renal vasoconstriction. Based upon these
results, Momen et al. (58) concluded that renal vasoconstriction during exercise
was largely mediated by muscle mechanoreceptors.
From the studies by Victor et al. (97), Middlekauf et al. (55) and Momen et
al, (58) it is apparent that the muscle afferents are important in controlling renal
blood flow during exercise. Because muscle temperature can influence muscle
afferent control of blood pressure and sympathetic responses to exercise (72,
73), temperature might influence renal vascular responses to exercise.
Therefore, the first goal of this project is to characterize the influence local
muscle temperature has on renal vascular responses to isometric handgrip.
Because Ray and Gracey (72) observed that local heating augmented
cardiovascular and MSNA responses to exercise, which were mechanoreceptor-
mediated, and because Momen et al. (58) observed that renal vascular
responses are mechanoreceptor mediated in humans the following hypothesis
was made:
Hypothesis 1: Forearm heating would augment renal vasoconstriction
during isometric handgrip.

11
Because Ray et al. (73) observed that local cooling delayed exercise-
induced increases in MSNA, and because the metaboreceptors contribute to
exercise-induced renal vasoconstriction (55, 58), the following hypothesis was
made:
Hypothesis 2: Forearm cooling would delay renal vasoconstriction during
isometric handgrip.
1.5 Aging, the Exercise Pressor Reflex, Heat, and Renal Blood Flow
Aging is associated with an increased prevalence of heat-related illnesses
(60). Likewise, aging alters cardiovascular responses to exercise in the heat.
Older individuals increase skin blood flow and decrease renal and splanchnic
blood flow less during upright cycling in the heat (28, 35). The exact
mechanisms behind these age-related changes in control of regional blood flow
distribution during exercise in the heat remains equivocal. Because it is unknown
how the interaction of aging and muscle temperature will effect cardiovascular
and MSNA responses to exercise, the next study will address this question.
Furthermore, because the goal of the first two studies of this project was to
examine the influence of local muscle temperature and isometric exercise on
renal blood flow responses to exercise, the next aim of this work will examine if
aging will alter renal responses to isometric handgrip during forearm heating.
Before introducing the next hypotheses that will be tested in this study, I will
review what is known about the influence of aging on muscle afferent control of
the exercise pressor reflex.

12
Ng et al. (63) compared cardiovascular and MSNA responses to isometric
handgrip in a group of older individuals with a group of young individuals. The
investigators found that arterial blood pressure and MSNA responses were the
same to exercise and postexercise muscle ischemia. The authors concluded
that aging did not alter exercise-induced changes in cardiovascular function.
Moreover, the lack of a difference during exercise suggested that muscle
mechano- and metaboreceptor-mediated changes in cardiovascular function
during isometric exercise were unaltered with age.
In contrast to Ng et al. (63), Markel et al. (49) found that aging did alter
the exercise pressor reflex. Markel et al. compared cardiovascular and MSNA
during rhythmic handgrip in young and older individuals. During the study the
arm was enclosed in a tank and handgrip was performed in six 1-min stages at
increasing ambient pressures to limit blood flow into the arm. At the highest level
of pressure, the older individuals had significantly lower arterial blood pressure,
MSNA, and forearm H+ concentration. Because the main driving force of
activation of the exercise pressor reflex is a mismatch between blood flow and
muscular work and because the investigators observed differences in
cardiovascular and autonomic responses when this mismatch was the greatest,
Markel et al. concluded that aging attenuated muscle afferent control of the
exercise pressor reflex.
To date no study has examined the influence of aging and local muscle
temperature on cardiovascular and MSNA responses during activation of
exercise pressor reflex. Because the exercise paradigm that will be used in the

13
current study is similar Ng et al. (63), who found that aging did not alter MSNA
responses to isometric handgrip the following hypothesis was made:
Hypothesis 3: The interaction of local muscle heating and aging will not
alter blood pressure and MSNA responses to isometric exercise.
The first study to examine the influence of aging and renal vasoconsriction
during exercise in humans was conducted by Momen et al (57). The
investigators compared older individuals’ renal vascular responses during
handgrip to that of younger individuals. Momen et al. found that older individuals
had greater renal vasoconstriction during handgrip and the response was
mediated by an increase in muscle mechanoreceptor sensitivity because
compared to the young group, the older individuals constricted more at the onset
of exercise. The combined influence of local muscle heating and aging on renal
vascular responses to exercise remains equivocal, and because of the
importance of the renal vasculature in cardiovascular control during exercise and
heat stress the next goal is to examine the combined influence of aging and local
muscle heating on renal vascular blood flow.
Because aging increases mechanoreceptor-mediated renal
vasoconstriction during exercise, and because muscle heating increases
sensitivity of the muscle mechanoreceptors, (72) the following hypothesis was
made:
Hypothesis 4: The interaction of aging and local muscle heating will
augment renal vasoconstriction during exercise.

14
1.6 Influence of ACE-inhibition on Cardiovascular and MSNA Responses to
Normothermic and Hyperthermic Isometric Exercise
ACE-inhibitors are used in the treatment of hypertension and chronic heart
failure. ACE-inhibitors lower blood pressure by decreasing blood angiotensin II
levels and by increasing blood bradykinin levels. Both the renin-angiotensin and
kallikrein-kinin system modify cardiovascular control during exercise. For
example, 7 days of treatment with the ACE-inhibitor captopril blunted the
increase in mean arterial blood during isometric handgrip (42). Likewise, a
reduction in plasma angiotensin II levels is associated with the reduction in renal
sympathetic nerve activity in rabbits with chronic heart failure (61). Infusion of
bradykinin and blocking bradykinin receptors alters muscle afferent activity and
cardiovascular function in animals (31, 54, 67). Because both the renin-
angiotensin and kallikrein-kinin can be influenced by ACE-inhibition,
cardiovascular and MSNA responses to exercise might be altered by ACE-
inhibition. Little is known about the acute influence of ACE-inhibition on
cardiovascular, renal vascular, and MSNA responses to isometric exercise.
Therefore, the goal of this study is to examine the effect of ACE-inhibition on
cardiovascular, renal vascular, and MSNA responses to isometric exercise in
humans.
Because ACE-inhibition increases blood kinin levels, which have been
found to act on muscle afferents in animals (31, 54, 67), and because isometric
handgrip does not alter angiotensin II concentrations in the blood in humans
(100), the following hypothesis was made:

15
Hypothesis 5: Acute ACE-inhibition will augment blood pressure, renal
vasoconstriction, and MSNA responses to isometric handgrip.
Besides acting on factors that influence muscle afferent sensitivity at
normothermic temperatures, ACE-inhibition might influence factors that modify
muscle afferent sensitivity in the heat. Little is known about what causes
changes in muscle afferent sensitivity in the heat. However, in the skin
bradykinin sensitizes heat-sensitive mechanoreceptors. Whether or not the
increase in blood kinin levels during ACE-inhibition might influence muscle
afferent sensitivity inn the heat remains unclear. Examining the effect of local
heating and ACE-inhibition on central hemodynamic, vascular, and MSNA
responses to isometric exercise, might provide insight into heat-related changes
muscle afferent. The following hypothesis was made:
Hypothesis 6: ACE-inhibition will further augment the increases in blood
pressure, renal vasoconstriction, and MSNA during isometric handgrip with a
hyperthermic forearm because of increases in blood kinin levels, which increase
sensitivity of mechano-heat sensitive afferents in the skin (51, 69).

16
1.7 References
1. Alam M and Smirk FH. Observations in man upon a blood pressure raising reflex arising from the voluntary muscles. J Physiol 89: 372-383, 1937.
2. Andres KH, von During M, and Schmidt RF. Sensory innervation of the Achilles tendon by group III and IV afferent fibers. Anat Embryol (Berl) 172: 145-156, 1985.
3. Coote JH, Hilton SM, and Perez-Gonzalez JF. The reflex nature of the pressor response to muscular exercise. J Physiol 215: 789-804, 1971.
4. Crayton SC, Aung-Din R, Fixler DE, and Mitchell JH. Distribution of cardiac output during induced isometric exercise in dogs. Am J Physiol 236: H218-224, 1979.
5. Diepstra G, Gonyea W, and Mitchell JH. Distribution of cardiac output during static exercise in the conscious cat. J Appl Physiol 52: 642-646, 1982.
6. Hanna RL and Kaufman MP. Role played by purinergic receptors on muscle afferents in evoking the exercise pressor reflex. J Appl Physiol 94: 1437-1445, 2003.
7. Herr MD, Imadojemu V, Kunselman AR, and Sinoway LI. Characteristics of the muscle mechanoreflex during quadriceps contractions in humans. J Appl Physiol 86: 767-772, 1999.
8. Hertel HC, Howaldt B, and Mense S. Responses of group IV and group III muscle afferents to thermal stimuli. Brain Res 113: 201-205, 1976.
9. Ho CW, Beard JL, Farrell PA, Minson CT, and Kenney WL. Age, fitness, and regional blood flow during exercise in the heat. J Appl Physiol 82: 1126-1135, 1997.
10. Kaufman MP and Forster HV. Reflexes controlling circulatory, ventilatory and airway responses to exercise. In: Handbook of Physiology: "Exercise Regulation and Integration of Multiple Systems". Bethesda, MD: Am. Physiol. Soc., 1996, p. 381-447.

17
11. Kaufman MP, Iwamoto GA, Longhurst JC, and Mitchell JH. Effects of capsaicin and bradykinin on afferent fibers with ending in skeletal muscle. Circ Res 50: 133-139, 1982.
12. Kaufman MP, Longhurst JC, Rybicki KJ, Wallach JH, and Mitchell JH. Effects of static muscular contraction on impulse activity of groups III and IV afferents in cats. J Appl Physiol 55: 105-112, 1983.
13. Kaufman MP and Rybicki KJ. Discharge properties of group III and IV muscle afferents: their responses to mechanical and metabolic stimuli. Circ Res 61: I60-65, 1987.
14. Kenney WL and Ho CW. Age alters regional distribution of blood flow during moderate-intensity exercise. J Appl Physiol 79: 1112-1119, 1995.
15. Kumazawa T and Mizumura K. Thin-fibre receptors responding to mechanical, chemical, and thermal stimulation in the skeletal muscle of the dog. J Physiol 273: 179-194, 1977.
16. Lang CC, Stein CM, He HB, and Wood AJ. Angiotensin converting enzyme inhibition and sympathetic activity in healthy subjects. Clin Pharmacol Ther 59: 668-674, 1996.
17. Li J and Sinoway LI. ATP stimulates chemically sensitive and sensitizes mechanically sensitive afferents. Am J Physiol Heart Circ Physiol 283: H2636-2643, 2002.
18. Markel TA, Daley JC, 3rd, Hogeman CS, Herr MD, Khan MH, Gray KS, Kunselman AR, and Sinoway LI. Aging and the exercise pressor reflex in humans. Circulation 107: 675-678, 2003.
19. Mayer S, Izydorczyk I, Reeh PW, and Grubb BD. Bradykinin-induced nociceptor sensitisation to heat depends on cox-1 and cox-2 in isolated rat skin. Pain, 2006.
20. McCloskey DI, Matthews PB, and Mitchell JH. Absence of appreciable cardiovascular and respiratory responses to muscle vibration. J Appl Physiol 33: 623-626, 1972.

18
21. McCloskey DI and Mitchell JH. Reflex cardiovascular and respiratory responses originating in exercising muscle. J Physiol 224: 173-186, 1972.
22. Mense S. Nervous outflow from skeletal muscle following chemical noxious stimulation. J Physiol 267: 75-88, 1977.
23. Middlekauff HR, Nitzsche EU, Nguyen AH, Hoh CK, and Gibbs GG. Modulation of renal cortical blood flow during static exercise in humans. Circ Res 80: 62-68, 1997.
24. Momen A, Leuenberger UA, Handly B, and Sinoway LI. Effect of aging on renal blood flow velocity during static exercise. Am J Physiol Heart Circ Physiol 287: H735-740, 2004.
25. Momen A, Leuenberger UA, Ray CA, Cha S, Handly B, and Sinoway LI. Renal vascular responses to static handgrip: role of muscle mechanoreflex. Am J Physiol Heart Circ Physiol 285: H1247-1253, 2003.
26. Moore R, Mallonee S, Sabogal RI, Zanardi L, Redd J, and Malone J. From the Centers for Disease Control and Prevention. Heat-related deaths--four states, July-August 2001, and United States, 1979-1999. JAMA 288: 950-951, 2002.
27. Mousa TM, Liu D, Cornish KG, and Zucker IH. Exercise training enhances baroreflex sensitivity by an angiotensin II-dependent mechanism in chronic heart failure. J Appl Physiol 104: 616-624, 2008.
28. Ng AV, Callister R, Johnson DG, and Seals DR. Age and gender influence muscle sympathetic nerve activity at rest in healthy humans. Hypertension 21: 498-503, 1993.
29. Pan HL, Stebbins CL, and Longhurst JC. Bradykinin contributes to the exercise pressor reflex: mechanism of action. J Appl Physiol 75: 2061-2068, 1993.
30. Petho G, Derow A, and Reeh PW. Bradykinin-induced nociceptor sensitization to heat is mediated by cyclooxygenase products in isolated rat skin. Eur J Neurosci 14: 210-218, 2001.

19
31. Ray CA and Gracey KH. Augmentation of exercise-induced muscle sympathetic nerve activity during muscle heating. J Appl Physiol 82: 1719-1725, 1997.
32. Ray CA, Hume KM, Gracey KH, and Mahoney ET. Muscle cooling delays activation of the muscle metaboreflex in humans. Am J Physiol Heart Circ Physiol 273: H2436-2441, 1997.
33. Rotto DM and Kaufman MP. Effect of metabolic products of muscular contraction on discharge of group III and IV afferents. J Appl Physiol 64: 2306-2313, 1988.
34. Rowell LB. Human cardiovascular adjustments to exercise and thermal stress. Physiol Rev 54: 75-159, 1974.
35. Rowell LB. Human circulation: regulation during physical stress.: Oxford: Oxford University Press, 1986.
36. Thimm F and Baum K. Response of chemosensitive nerve fibers of group III and IV to metabolic changes in rat muscles. Pflugers Arch 410: 143-152, 1987.
37. Victor RG, Rotto DM, Pryor SL, and Kaufman MP. Stimulation of renal sympathetic activity by static contraction: evidence for mechanoreceptor-induced reflexes from skeletal muscle. Circ Res 64: 592-599, 1989.
38. von During M and Andres KH. Topography and ultrastructure of group III and IV nerve terminals of cat's gastrocnemius-soleus muscle. In: The Primary Afferent Neuron: A survey of Recent Morph-functional Aspects, edited by Zenker W and Neuhuber WL. New York: Pleneum Press, 1990, p. 35-41.
39. Warren JH, Lewis W, Wraa CE, and Stebbins CL. Central and peripheral effects of angiotensin II on the cardiovascular response to exercise. J Cardiovasc Pharmacol 38: 693-705, 2001.

20
Chapter 2
Changes in Forearm Muscle Temperature Alter Renal Vascular Responses to Isometric Handgrip
2.1 Introduction
Vasoconstriction of the renal vascular bed is an important mechanism for
meeting the blood pressure and flow demands necessary to perfuse the muscle
and skin during exercise and heat stress (3, 79, 80). During exercise, renal
vasoconstriction appears to be mediated by activation of the exercise pressor
reflex (25, 58, 97). This reflex is controlled by mechanically and metabolically
sensitive afferents in the working muscle. These afferents, aside from being
sensitive to mechanical and metabolic changes, are responsive to temperature
changes in the muscle (27, 40, 72, 73). Currently, little is known about how
changing temperature of the exercising muscle may alter exercise pressor reflex-
mediated renal vasoconstriction.
Previously, we have found that altering muscle temperature can alter
cardiovascular and autonomic responses to exercise in humans. For example,
heating the forearm increased mean arterial blood pressure and muscle
sympathetic nerve activity (MSNA) at the beginning of ischemic isometric
handgrip (72), whereas cooling the forearm muscles delayed increases in MSNA
during ischemic isometric handgrip (73). Ray et al. (72) concluded that the
greater increase in mean arterial blood pressure and MSNA during handgrip with
a heated forearm were mediated by an increase in sensitivity of mechanically
sensitive muscle afferents. This conclusion was based upon the finding that the
blood pressure and MSNA responses were observed near the onset of exercise,

21
when mechanoreceptor-mediated responses would be greatest, but not during
postexercise muscle ischemia when there would be no mechanoreceptor
mediated responses. In contrast, the attenuation MSNA responses during
muscle cooling appeared to be mediated by a delay in activation of metabolically
sensitive muscle afferents (73). This conclusion was based upon the fact that
MSNA increased less during exercise in the cold but did not differ at fatigue or
during postexercise muscle ischemia
Because in humans activation of the exercise pressor reflex can alter
renal vascular conductance (55, 58), and because muscle heating and cooling
augment and attenuate MSNA, respectively (73, 75), the following two
hypotheses were tested in the present study: 1) Forearm heating would augment
renal and calf vasoconstriction during ischemic isometric handgrip via
sensitization of mechanically sensitive afferents; and 2) Forearm cooling would
delay renal and calf vasoconstriction during ischemic isometric handgrip via
delayed activation of metabolically active muscle afferents.
2.2 Methods
2.2.1 Subjects
The study consisted of two experimental groups (heating and cooling
groups). Fifteen subjects participated in the heating study (age, 26 ± 1 yr; height,
174.4 ± 3.0 cm; weight, 70.7 ± 4.2 kg; 8 men and 7 women). Twelve subjects
participated in the cooling study (age, 25 ± 1 yr; height, 175.7 ± 3.3 cm; weight,
73.6 ± 4.3 kg; 7 men and 5 women). All subjects were normotensive, non-obese,
non-smokers, not taking any medications, and had no autonomic dysfunction or

22
cardiovascular disease. Subjects who were endurance or resistance trained
were excluded from study. Subjects arrived at the laboratory fasted and had
abstained from caffeine, alcohol, and exercise for 12 h. All testing procedures
were the same for the two groups except for the temperature of the forearm
during each respective thermal stress trial. To serve as their own controls,
subjects performed a normothermic trial during the same visit as the thermal
stress trial. The experimental protocol was approved by the Institutional Review
Board at the Pennsylvania State University College of Medicine and all subjects
gave written informed consent prior to participating.
2.2.2 Experimental Design
To regulate forearm muscle temperature, subjects wore a water-perfused
sleeve (Med-Eng Systems, Ottawa, ON, Canada) over their dominant arm. For
the heating group, water at 55 ºC was circulated through the sleeve for 30 min.
At the end of 30 min, the water was cooled to 50 ºC for the exercise protocol. In
the cooling group, a bag of ice was placed over the sleeve and water at 1 ºC was
circulated through the sleeve for approximately 1 h. At the end of one hour the
temperature of the circulating water was increased to 10 ºC. The order of the
normothermic trial and thermal stress trial was randomized. A minimum of 40
min separated the normothermia and thermal stress trial to allow all measures to
return to baseline, and the subsequent trial did not begin until baseline measures
were reached. When the normothermic trial was second, the forearm muscle
temperature was adjusted by varying the temperature of the circulating water

23
until the forearm temperature equaled those measured before heating or cooling
the forearm. Ambient temperature in the laboratory during testing was 21-23 ºC.
2.2.4 Experimental Protocol
The experimental protocol for all temperature conditions was as follows: 3-
min baseline, 1 min of ischemia, ischemic handgrip to fatigue, 1 min of
postexercise muscle ischemia, and 3-min recovery. During exercise, subjects
squeezed a hand dynamometer at 30% of their maximal voluntary contraction.
Maximal voluntary contraction was determined before the experimental protocol
and before muscle temperature probe insertion. Measurements during each trial
included muscle temperature, skin temperature, arterial blood pressure, heart
rate, renal artery blood velocity, rate of perceived exertion, and calf blood flow.
2.2.5 Measurements
Muscle temperature was measured using a 22-gauge hypodermic
intramuscular thermistor (YSI 552, Yellow Springs, OH). The thermistor was
placed 2-3 cm below the skin into the flexor muscles of the forearm. To limit the
possibility that heating or cooling the probe at the surface of the skin altered
temperature readings in the muscle, the top of the probe was insulated from
direct contact with the water-perfused sleeve. Measurements were taken every
minute during baseline and at 30 s intervals for the remainder of the experimental
protocol. Continuous skin temperature of the exercising limb was measured via
two thermocouples attached to the dorsal forearm skin and routed through a
thermocouple meter (model TC-1000, Sabel Systems, Henderson, NV).
Tympanic temperatures were recorded using a First Temp Genius Tympanic

24
Thermometer (Sherwood Medical, St. Loius, MO) after changing forearm muscle
temperature to monitor for possible changes in body temperature.
Doppler ultrasound (HDI 5000, ATL Ultrasound, Bothell, WA, USA) was
used to measure renal artery blood velocity. The renal artery was scanned using
the anterior abdominal approach. To scan the renal artery a curved-array
transducer (2–5 MHz) with a 2.5-MHz pulsed Doppler frequency was used. The
probe insonation angle to the artery was less than 60°. The focal zone was set at
the depth of the artery. The transducer was held in the same place to record
velocity tracings during each trial and the data were obtained in the same phase
of the respiratory cycle. Doppler tracings were analyzed using the software of
the ATL to obtain renal artery blood velocity measurements during each cardiac
cycle. The ratio of renal artery blood velocity and mean arterial blood pressure
was used as an index of renal artery conductance.
Calf blood flow was measured using venous occlusion plethysmography.
A mercury-in-silastic strain gauge (Hokanson, Bellevue, WA, USA) was placed
around the maximal circumference of the calf. The calf was positioned above the
heart. An ankle cuff was inflated to 220 mmHg to occlude blood to the foot. A
Hokanson CC 17 thigh cuff was placed around the thigh and inflated to 50 mmHg
to occlude venous outflow every 15 s for 7.5 s. Venous congestion caused by
the thigh cuff increased calf volume, which caused the mercury-in-silastic strain
gauge to stretch. The rate of change in electrical resistance in the mercury-in-
silastic strain gauge as it stretched is directly proportional to calf blood. The ratio

25
of calf blood flow and mean arterial blood pressure was used to calculate calf
vascular conductance.
Heart rate and arterial blood pressure were continuously recorded during
all trials using a Finometer (Finapres Medical Systems, Amsterdam,
Netherlands). Before all trials, resting brachial artery blood pressure (Dinamap,
General Electric, Waukesha, WI, USA) was recorded. Subjects were asked to
give ratings of perceived exertion every 30 s during exercise and at fatigue (7).
2.2.6 Data analysis
Data, except renal blood flow velocities, were analyzed offline using
Chart 5.4.2 software (ADI Instruments, Newcastle, Australia). Resting variables
for each temperature were compared using a paired t-test. Baseline and
postexercise muscle ischemia data were averaged over their respective time
periods. Because exercise time differed between temperature conditions data
were expressed as a percentage of time to fatigue. The five exercise time
periods averaged were 0-20%, 20-40%, 40-60%, 60-80%, and 80-100%. All
data were analyzed using a two-factor within-repeated-measures analysis of
variance (temperature x time). Significance was considered at a P value of <
0.05. Results were expressed as mean ± S.E.
2.3 Results
2.3.1 Heating Study.
Baseline. Baseline measurements during normothermia and heating are
presented in Table 2.1. Heating significantly increased forearm muscle and skin
temperatures but did not alter tympanic temperature, indicating that local heating

26
did not alter core body temperature. All other hemodynamic measurements were
the same during baseline for the two trials.
Exercise Responses. Mean arterial blood pressure increased significantly
more during heating compared to normothermia (Fig. 2.1). The decreases in
renal and calf vascular conductance were greater during heating than
normothermia (Fig. 2.2 & 2.3, respectively). Ratings of perceived exertion for the
heating trial were slightly higher throughout the first two minutes of exercise
compared to normothermia (~ ∆ 1 RPE unit) but did not differ at fatigue. Exercise
time was significantly longer during normothermia compared to heating (174 ± 15
and 128 ± 15 s, respectively). Skin temperatures did not change during
normothermia and heating and averaged 30.8 ± 0.5 ºC and 40.5 ± 0.3 ºC,
respectively, throughout exercise.
Postexercise Muscle Ischemia. Mean arterial blood pressure and heart
rate decreased from fatigue, but were above baseline for both temperatures.
Mean arterial blood pressure during heating was significantly higher than during
normothermia. Renal and calf vascular conductance were not different between
the two temperature conditions (Fig. 2.2 & 2.3, respectively).
2.3.2 Cooling Study.
Baseline. Baseline measurements during forearm cooling and
normothermia are presented in Table 2.2. Cooling significantly decreased

27
forearm muscle and skin temperatures, but did not alter tympanic temperature.
All other hemodynamic measurements were the same during baseline for the two
trials.
Exercise Responses. Mean arterial blood pressure and heart rate
increased during exercise at both forearm temperatures (Fig. 2.4). The increase
in mean arterial blood pressure was greater during normothermia compared to
cooling. Renal artery blood velocity approached significantly higher values
during cooling (P = 0.06) (Fig. 2.5). Renal vascular conductance was
significantly higher during cooling compared to normothermia (Fig. 2.5).
Changes in calf blood flow and vascular conductance (Fig. 2.6) were not different
between normothermia and cooling. Ratings of perceived exertion were not
different between normothermia and cooling throughout exercise and at fatigue.
Exercise time was not significantly different between normothermia and cooling
(184 ± 11 and 156 ± 16 s, respectively). Skin temperatures did not change during
normothermia and cooling and averaged 31.8 ± 0.5 ºC and 18.8 ± 0.6 ºC,
respectively, throughout exercise.
Postexercise Muscle Ischemia. During postexercise muscle ischemia, the
increase in mean arterial blood pressure was greater for normothermia compared
to cooling (Fig. 2.4). The increase in heart rate did not differ between
temperatures. Renal artery blood velocity and renal vascular conductance were

28
higher during cooling compared to the normothermic trial (Fig 2.5). Calf vascular
conductance did not differ between normothermia and cooling (Fig 2.6).
2.4 Discussion
The major new findings of these studies are as follows: 1) varying forearm
muscle temperature did not alter resting renal or calf vascular conductance; 2)
heating the forearm muscles augmented renal and calf vasoconstriction during
ischemic isometric handgrip via an increase in sensitivity of muscle
mechanoreceptors; and 3) cooling the forearm muscles attenuated renal
vasoconstriction during exercise due to blunted activation of the muscle
metaboreflex.
Our lab has found that forearm heating augments increases in MSNA
during the first few minutes of ischemic handgrip, but does not alter activity at
fatigue or during postexercise muscle ischemia (72). Ray and Gracey (72)
concluded that this increase in MSNA was mediated by increased sensitivity of
the mechanoreceptors. This conclusion was drawn because the augmented
response was observed early in exercise and because no differences were
observed during postexercise muscle ischemia. The results of the current study
indicate that in humans heating the forearm muscles elicits a similar influence on
mechanoreceptor-mediated renal vasoconstriction because the observed
differences in renal vasoconstriction occurred only during exercise but not when
the muscle metaboreceptors are engaged in isolation during postexercise muscle
ischemia. These findings are in agreement with other studies that have found

29
that mechanoreceptors contribute to decreases in renal conductance in humans
(55, 58) and decreases in both renal conductance and sympathetic nerve activity
in animals (37, 38, 97). The greater decrease in calf vascular conductance with
heating corresponds to greater increases in MSNA, which we have observed
previously using the same protocol (72).
Ray et al. (73) observed that forearm cooling delayed muscle
metaboreceptor-mediated increases in muscle sympathetic nerve activity during
ischemic isometric handgrip. This conclusion was based upon the fact that the
differences in MSNA between muscle cooling and normothermia occurred later
during exercise when metaboreceptor mediated increases in MSNA would be
greatest. In accordance with Ray et al. (73), the results of the present study may
be due to a decrease in sympathetic nerve activity to the kidney vasculature. In
the current study during postexercise muscle ischemia, which selectively
engages the metaboreflex only, renal vascular conductance was higher in the
cooling trial. If cooling did not attenuate metaboreflex-mediated renal
vasoconstriction, renal vascular conductance should have been similar during
postexercise muscle ischemia during cooling and normothermia. Although
cooling did not significantly blunt calf vasoconstriction during exercise there was
a definite trend for calf vascular conductance to be lower during cooling
compared to normothermia.
In the present study, the arterial baroreflexes and central command could
have contributed to measured changes in renal vasoconstriction during thermal
stress and exercise. In the current study we observed that mean arterial blood

30
pressure was greater during heating compared to normothermia and during
normothermia compared to cooling. Increased arterial blood pressure and
loading of the arterial baroreflexes decreases sympathetic outflow and vascular
tone thereby increasing vascular conductance (59). In the current study, if the
baroreflexes were contributing to the measured differences in arterial blood
pressure and blood flow between temperature conditions, we would have
expected greater renal and peripheral vasodilation when comparing heating to
normothermia and normothermia to cooling. We observed the opposite response
between thermal conditions; therefore, we believe that the arterial baroreflexes
may not contribute to the observed differences between thermal stimuli and
normothermia. Consistent with this, we found that MSNA and mean arterial blood
pressure were higher during ischemic isometric handgrip when comparing
responses between heated muscle and normothermic muscle, and between
normothermic muscle and cooled muscle (72, 73). The contribution of central
command to increases in MSNA and arterial blood pressure is thought to occur
mainly during intense exercise and at fatigue when volitional effort is greatest
(75, 98). In the current study, ratings of perceived exertion were slightly higher
during exercise and heating, but were not different at fatigue. During the cooling
trial, ratings of perceived exertion were not different during exercise or at fatigue.
Because perceived exertion was the same at fatigue in both trials, when volitional
effort was greatest, this suggests that central command did not contribute to
measured differences in renal vasoconstriction. Further support that central
command did not contribute to exercise-induced renal vasoconstriction is that in

31
humans, electrical stimulation of the bicep muscles and postexercise muscle
ischemia, two ways to selectively activate muscle afferents without central
command input, both increase renal vasoconstriction (55, 58).
Changing visceral blood flow is an important mechanism the body uses to
control core temperature during thermal stress and to increase perfusion of blood
into metabolically active tissues during exercise (79). Combined physical and
heat stress poses a severe challenge to maintaining blood pressure because the
demands of these two vascular beds can outstrip the available cardiac output.
Therefore, the cardiovascular system must reduce blood flow to these tissue
beds or direct blood flow from other tissues to prevent decreases in performance
and heat loss. Muscle blood flow has been found to be stable during exercise in
the heat (65, 82), and skin blood flow remains relatively unchanged as internal
temperature increases over 38 ºC during exercise (34). Therefore, the body
must limit blood flow to other vascular beds such as the renal or splanchnic
vascular beds. The results of the current study indicate that the augmentation of
the exercise pressor reflex during heating may be a mechanism that promotes
vasoconstriction of the visceral tissues and inactive skeletal muscle to help
maintain blood pressure. Changes in the exercise pressor reflex during the cold
may occur for a different reason than in the heat. During cold stress, the body
decreases peripheral blood flow to increase insulation and maintain internal
temperature (36, 92). By delaying increases in renal vasoconstriction during
exercise and limiting perfusion of the exercising muscle, higher peripheral
insulation may be maintained and decreases in core temperature delayed.

32
The mechanisms behind thermal induced changes in sensitivity of the
exercise pressor reflex remains equivocal. Mechanoreceptor sensitivity can be
altered by a variety of factors including prostaglandins, bradykinin, and lactic acid
(30, 86). Arachidonic acid derivatives selectively excite mechanoreceptors but
not metaboreceptors (76, 78). Therefore, it is possible that in the heating trials
the increased muscle temperature may have altered the chemical milieu of the
muscle and increased the concentration of a neurologically active substance that
could sensitize the mechanoreceptors. Several mechanisms may explain the
observed responses during the cooling trials. First, cooling the muscle
decreases firing of muscle afferents themselves (27, 40). Second, cooling the
muscle may have decreased the metabolic rate of the muscle, which could lower
the production of various exercise metabolites that activate the metaboreceptors
(e.g., lactic acid and hydrogen ions). Future investigations are needed to
elucidate thermal induced changes in sensitivity of the muscle pressor reflex.
It is possible that skin afferents could have contributed to the differences
between thermal conditions; however, there are several reasons to suggest this
is unlikely. First, at rest when only temperature of the exercising limb was
altered, baseline hemodynamic measurements and MSNA (72, 73) did not differ
between normothermia and heating and between normothermia and cooling.
Second, during exercise there was no change in skin temperature for any of the
thermal conditions. Third, subjects did not complain of pain related to the skin
and the temperatures of the skin recorded during heating and cooling were either
below or above those reported for causing pain in the arm (62, 101). Finally, two

33
subjects performed normothermic handgrip while the skin of the contralateral arm
was altered to equal skin temperatures measured in the normothermic, heating
and cooling trials using the water-perfused sleeve. Changes in mean arterial
blood pressure and renal and calf vascular conductance did not differ between
any of the temperatures. For these reasons, we do not believe that afferents in
the skin contributed to the observed differences between temperature conditions.
The goal of the present study was to examine if changing local muscle
temperature altered muscle afferent control of renal blood flow. To limit the
influence of the effect of changes in muscle temperature and metabolism
because of exercise-induced hyperemia, subjects performed handgrip while
ischemic. It is possible that changes in blood flow during contraction may alter
the influence of thermal stress on the exercise pressor reflex. However,
isometric contractions greater than 15% of maximal voluntary contraction do not
raise muscle blood flow and are therefore largely ischemic (22).
In the current study, the temperature of only a small muscle mass was
altered. The current study contributes to the understanding of the cardiovascular
responses to whole body exercise and thermal stress by isolating possible
contributions to cardiovascular changes from a peripheral limb. However, during
whole body exercise and thermal stress the challenges placed on the
cardiovascular system and sensory input will differ from that of the current study.
Therefore, the results of the current study may not extend to whole body exercise
during thermal stress.

34
Because of resolution limitations, it is not possible to accurately measure
renal artery diameters using Doppler Ultrasound. Consequently, we used blood
velocity as a surrogate for flow. Although it is possible that the diameters of the
renal arteries changed during the study and thus could have altered our
estimation of renal blood flow, pharmacological-mediated renal vasoconstriction
(50) and vasodilation (47) do not alter diameter of the renal artery. This, and the
fact that the vessel we examined was a conduit vessel and not a resistance
vessel, makes it unlikely that changes in renal artery diameter influenced the
results of the study.
In summary, heating the arm augmented renal and calf vasoconstriction
during forearm exercise, whereas cooling the arm attenuated renal
vasoconstriction at fatigue and during postexercise muscle ischemia. The
augmentation of renal vasoconstriction during exercise with a heated muscle is
associated with increases in sensitivity of the muscle mechanoreceptors,
whereas the attenuation of renal vasoconstriction during exercise with a cooled
muscle was related to a blunting of the muscle metaboreflex.

35
2.5 References
1. Armstrong RB, Delp MD, Goljan EF, and Laughlin MH. Distribution of blood flow in muscles of miniature swine during exercise. J Appl Physiol 62: 1285-1298, 1987.
2. Borg G. Subjective aspects of physical and mental load. Ergonomics 21: 215-220, 1978.
3. Hansen J, Jacobsen TN, and Amtorp O. The exercise pressor response to sustained handgrip does not augment blood flow in the contracting forearm skeletal muscle. Acta Physiol Scand 149: 419-425, 1993.
4. Hayes SG and Kaufman MP. MLR stimulation and exercise pressor reflex activate different renal sympathetic fibers in decerebrate cats. J Appl Physiol 92: 1628-1634, 2002.
5. Hertel HC, Howaldt B, and Mense S. Responses of group IV and group III muscle afferents to thermal stimuli. Brain Res 113: 201-205, 1976.
6. Kaufman MP and Hayes SG. The exercise pressor reflex. Clin Auton Res 12: 429-439, 2002.
7. Kellogg DL, Jr., Johnson JM, Kenney WL, Pergola PE, and Kosiba WA. Mechanisms of control of skin blood flow during prolonged exercise in humans. Am J Physiol Heart Circ Physiol 265: H562-568, 1993.
8. Kenney WL and Munce TA. Invited review: aging and human temperature regulation. J Appl Physiol 95: 2598-2603, 2003.
9. Kim JK, Hayes SG, Kindig AE, and Kaufman MP. Thin-fiber mechanoreceptors reflexly increase renal sympathetic nerve activity during static contraction. Am J Physiol Heart Circ Physiol 292: H866-873, 2007.
10. Koba S, Yoshida T, and Hayashi N. Renal sympathetic and circulatory responses to activation of the exercise pressor reflex in rats. Exp Physiol 91: 111-119, 2006.

36
11. Kumazawa T and Mizumura K. Thin-fibre receptors responding to mechanical, chemical, and thermal stimulation in the skeletal muscle of the dog. J Physiol 273: 179-194, 1977.
12. Manoharan G, Pijls NH, Lameire N, Verhamme K, Heyndrickx GR, Barbato E, Wijns W, Madaric J, Tielbeele X, Bartunek J, and De Bruyne B. Assessment of renal flow and flow reserve in humans. J Am Coll Cardiol 47: 620-625, 2006.
13. Marraccini P, Fedele S, Marzilli M, Orsini E, Dukic G, Serasini L, and L'Abbate A. Adenosine-induced renal vasoconstriction in man. Cardiovasc Res 32: 949-953, 1996.
14. Middlekauff HR, Nitzsche EU, Nguyen AH, Hoh CK, and Gibbs GG. Modulation of renal cortical blood flow during static exercise in humans. Circ Res 80: 62-68, 1997.
15. Momen A, Leuenberger UA, Ray CA, Cha S, Handly B, and Sinoway LI. Renal vascular responses to static handgrip: role of muscle mechanoreflex. Am J Physiol Heart Circ Physiol 285: H1247-1253, 2003.
16. Monahan KD. Effect of aging on baroreflex function in humans. Am J Physiol Regul Integr Comp Physiol 293: R3-12, 2007.
17. Neisser U. Temperature thresholds for cutaneous pain. J Appl Physiol 14: 368-372, 1959.
18. Nielsen B, Savard G, Richter EA, Hargreaves M, and Saltin B. Muscle blood flow and muscle metabolism during exercise and heat stress. J Appl Physiol 69: 1040-1046, 1990.
19. Ray CA and Gracey KH. Augmentation of exercise-induced muscle sympathetic nerve activity during muscle heating. J Appl Physiol 82: 1719-1725, 1997.
20. Ray CA, Hume KM, Gracey KH, and Mahoney ET. Muscle cooling delays activation of the muscle metaboreflex in humans. Am J Physiol Heart Circ Physiol 273: H2436-2441, 1997.

37
21. Ray CA, Secher NH, and Mark AL. Modulation of sympathetic nerve activity during posthandgrip muscle ischemia in humans. Am J Physiol Heart Circ Physiol 266: H79-83, 1994.
22. Rotto DM, Hill JM, Schultz HD, and Kaufman MP. Cyclooxygenase blockade attenuates responses of group IV muscle afferents to static contraction. Am J Physiol Heart Circ Physiol 259: H745-750, 1990.
23. Rotto DM, Schultz HD, Longhurst JC, and Kaufman MP. Sensitization of group III muscle afferents to static contraction by arachidonic acid. J Appl Physiol 68: 861-867, 1990.
24. Rowell LB. Human cardiovascular adjustments to exercise and thermal stress. Physiol Rev 54: 75-159, 1974.
25. Rowell LB. Human circulation: regulation during physical stress.: Oxford: Oxford University Press, 1986.
26. Savard GK, Nielsen B, Laszczynska J, Larsen BE, and Saltin B. Muscle blood flow is not reduced in humans during moderate exercise and heat stress. J Appl Physiol 64: 649-657, 1988.
27. Sinoway LI and Li J. A perspective on the muscle reflex: implications for congestive heart failure. J Appl Physiol 99: 5-22, 2005.
28. Stocks JM, Taylor NA, Tipton MJ, and Greenleaf JE. Human physiological responses to cold exposure. Aviat Space Environ Med 75: 444-457, 2004.
29. Victor RG, Rotto DM, Pryor SL, and Kaufman MP. Stimulation of renal sympathetic activity by static contraction: evidence for mechanoreceptor-induced reflexes from skeletal muscle. Circ Res 64: 592-599, 1989.
30. Victor RG, Secher NH, Lyson T, and Mitchell JH. Central command increases muscle sympathetic nerve activity during intense intermittent isometric exercise in humans. Circ Res 76: 127-131, 1995.

38
31. Wolf S and Hardy JD. Studies on Pain. Observations on Pain Due to Local Cooling and on Factors Involved in the "Cold Pressor" Effect. J Clin Invest 20: 521-533, 1941.

39
Table 2.1. Baseline measurement during normothermia heating. Heating Study (n=15)
Variable Normothermia Heating
Muscle Temperature (ºC) 33.9 ± 0.3 38.5 ± 0.2*
Tympanic Temperature (ºC) 36.8 ± 0.1 36.8 ± 0.1
Skin Temperature (ºC) 31.0 ± 0.4 40.2 ± 0.2*
MAP (mmHg) 87 ± 2 89 ± 2
Heart Rate (beats/min) 59 ± 2 62 ± 3
Renal Blood Velocity (cm/sec) 52.9 ± 2.2 53.6 ± 2.1
RVC (cm/sec/mmHg) 0.57 ± 0.04 0.60 ± 0.04
Calf Blood Flow (ml/min/dl) 1.8 ± 0.2 1.9 ± 0.2
CVC (ml/min/dl/mmHg) 0.018 ± 0.002 0.021 ± 0.002
MAP, mean arterial blood pressure; RVC, renal vascular conductance; CVC, calf vascular conductance. *Significantly different from respective normothermic trial (P < 0.05). Values are mean ± S.E.M.

40
0
10
20
30
40
50
60 34 °C
39 °C
temp x time p < 0.05
! M
AP
(m
mH
g)
34 °C
39 °C
*
0 20 40 60 80 1000
5
10
15
20
25
30
35
Time to Fatigue (%)
! H
R (
beats
/min
)
PEMI
Figure 2.1. Changes from baseline in mean arterial blood pressure (MAP) and
heart rate (HR) during exercise and postexercise muscle ischemia (PEMI) during forearm heating (39 ºC) and normothermia (34 ºC). Points on exercise graph are average of 20 percent of exercise time before each point. Heating the forearm augmented MAP during exercise and postexercise muscle ischemia. *Significantly different from normothermia (P < 0.05). Values are mean ± SE.

41
-15
-10
-5
0
34 °C
39 °C
temp x time p < 0.05
! R
BV
(c
m/s
ec
)
34 °C
39 °C
-0.4
-0.3
-0.2
-0.1
-0.0
temp x time p < 0.05! R
VC
(u
nit
s)
20 40 60 80 100
-60
-40
-20
0
temp x time p < 0.05
0Time to Fatigue (%)
! R
VC
(%
)
PEMI
Figure 2.2 Changes from baseline in renal artery blood flow velocity (RBV)
and renal vascular conductance (RVC) during exercise and postexercise muscle ischemia (PEMI) during forearm heating (39 ºC) and normothermia (34 ºC). Points on exercise graph are average of 20 percent of exercise time before each point. RBV and RVC decreased significantly more during the heating trial during exercise. Values are mean ± SE.

42
-0.8
-0.6
-0.4
-0.2
-0.0
0.2
0.4
0.6 34 °C
39 °C
temp x time p < 0.05
! C
VC
(u
nit
s)
34 °C
39 °C
20 40 60 80 100
-50
-30
-10
10
30
temp x time p < 0.05
0Time to Fatigue (%)
! C
VC
(%
)
PEMI
Figure 2.3. Changes from baseline in calf vascular conductance (CVC) during
exercise and postexercise muscle ischemia (PEMI) during forearm heating (39 ºC) and normothermia (34 ºC). Points on exercise graph are average of 20 percent of exercise time before each point. CVC decreased significantly more during the heating trial during exercise. Values are mean ± SE.

43
Table 2.2 Baseline measurement during normothermia and cooling. Cooling Study (n=12)
Variable Normothermia Cooling
Muscle Temperature (ºC) 34.4 ± 0.3 25.5 ± 0.4*
Tympanic Temperature (ºC) 36.9 ± 0.1 36.9 ± 0.1
Skin Temperature (ºC) 31.8 ± 0.5 18.4 ± 0.9*
MAP (mmHg) 89 ± 2 88 ± 1
Heart Rate (beats/min) 55 ± 5 57 ± 2
Renal Blood Velocity (cm/sec) 56.6 ± 3.3 57.1 ± 3.2
RVC (cm/sec/mmHg) 0.61 ± 0.05 0.59 ± 0.04
Calf Blood Flow (ml/min/dl) 2.2 ± 0.2 2.3 ± 0.2
CVC (ml/min/dl/mmHg) 0.023 ± 0.002 0.024 ± 0.003
MAP, mean arterial blood pressure; RVC, renal vascular conductance; CVC, calf vascular conductance. *Significantly different from respective normothermic trial (P < 0.05). Values are mean ± S.E.M.

44
0
10
20
30
40
50
60 34 °C26 ºC
temp x time p < 0.05
! M
AP
(m
mH
g)
*
34 °C
26 °C
0 20 40 60 80 1000
10
20
30
40
Time to Fatigue (%)
! H
R (
beats
/min
)
PEMI
Figure 2.4. Changes from baseline in mean arterial blood pressure (MAP) and
heart rate (HR) during exercise and postexercise muscle ischemia (PEMI) during forearm cooling (26 ºC) and normothermia (34 ºC). Points on exercise graph are average of 20 percent of exercise time before each point. Cooling the forearm attenuated the increase in MAP during exercise and postexercise muscle ischemia. *Significantly different from normothermia (P < 0.05). Values are mean ± SE.

45
-10
-5
0
5
34 °C26 ºC
! R
BV
(cm
/sec)
*
34 °C26 ºC
-0.3
-0.2
-0.1
-0.0
-0.3
temp x time p < 0.05
! R
VC
(u
nit
s)
*
20 40 60 80 100
-60
-40
-20
0
temp x time p < 0.05
0Time to Fatigue (%)
! R
VC
(%
)
*
PEMI
Figure 2.5. Changes from baseline in renal artery blood velocity (RBV) and
renal vascular conductance (RVC) during exercise and postexercise muscle ischemia (PEMI) during forearm cooling (26 ºC) and normothermia (34 ºC). Points on exercise graph are average of 20 percent of exercise time before each point. Cooling the forearm attenuated the decrease in RVC during exercise. During postexercise muscle ischemia RBV and RVC were higher in the cooling trial. *Significantly different from normothermia (P < 0.05). Values are mean ± SE.

46
-1.0
-0.5
0.0
0.5
1.0 34 °C26 ºC
! C
VC
(u
nit
s)
34 °C26 ºC
20 40 60 80 100
-40
-30
-20
-10
0
10
20
30
40
0Time to Fatigue (%)
! C
VC
(%
)
PEMI
Figure 2.6. Changes from baseline in calf vascular conductance (CVC) during
exercise and postexercise muscle ischemia (PEMI) during forearm cooling (26 ºC) and normothermia (34 ºC). Points on exercise graph are average of 20 percent of exercise time before each point. Cooling did not alter CVC compared to normothermia at any time. Values are mean ± SE.

47
Chapter 3
Renal Vasoconstriction During Isometric Handgrip: Interactive Effect of Aging and Local Muscle Heating
3.1 Introduction
Aging is associated with an increased prevalence of heat-related illnesses
(60). Moreover, cardiovascular responses to exercise in the heat are altered with
age. Older individuals increase skin blood flow and decrease renal and
splanchnic blood flow less during upright dynamic leg exercise (i.e., cycling) in
the heat compared to younger individuals (28, 35). The exact mechanisms
behind these age-related changes in control of regional blood flow distribution
during exercise in the heat remains equivocal.
Thinly myelinated skeletal muscle afferents respond to mechanical
deformation, metabolitic by-products, and changes in muscle temperature.
Activation of the muscle afferents contributes to renal vasoconstriction in humans
(39, 55, 58) and increases renal sympathetic nerve activity in cats (25). Older
individuals constrict the renal vasculature more during isometric handgrip than
younger individuals (57). Momen et al. (57) concluded that increased renal
vasoconstriction in older individuals was mediated by increased sensitivity of
mechanically sensitive muscle afferents because responses were observed at
the onset of exercise and not during postexercise muscle ischemia. Currently, it
remains unknown how aging and local muscle heating interact jointly affect renal
vascular responses to isometric exercise. Because aging augments
mechanoreceptor-mediated renal vasoconstriction during normothermic isometric

48
handgrip (57) and local heating augments muscle mechanoreceptor sensitivity
and renal vasoconstriction in younger individuals (39, 72), the primary goal of this
study was to test the hypothesis that aging augments renal vasoconstriction
during isometric handgrip with local muscle heating.
During exercise, muscle afferents modulate increases in muscle
sympathetic nerve activity (MSNA) (48). Heating the forearm augments MSNA
during isometric handgrip but not during postexercise muscle ischemia (72).
Because these responses were observed at the onset of exercise and not during
postexercise muscle ischemia, it was concluded that local heating increased
sensitivity of mechanically sensitive muscle afferents. The effect of aging and
local muscle temperature on MSNA during activation of the exercise pressor
reflex remains unclear. Therefore, the second goal of this study was to examine
the interaction between aging and heating on MSNA during exercise. Because
aging does not alter MSNA responses to handgrip exercise (64) and total
norepinephrine spillover is the same in young and older subjects during isometric
handgrip (18), we hypothesized that the interactive effect of heating and aging
would not alter MSNA responses to isometric handgrip during forearm heating.
3.2 Methods
3.2.1 Subjects
Twelve young (7 men and 5 women; 27 ± 1 yr of age, 175.0 ± 3.1 cm
height, 73.0 ± 4.4 kg body wt) and nine older volunteers (3 men and 6 women; 63
± 1 yr of age, 170.3 ± 1.8 cm height, 70.5 ± 2.8 kg body wt) participated in the
study. We have previously described the renal data of the young subjects (39);

49
however, their MSNA responses were not reported. All subjects were
normotensive, nonobese, nonsmokers, not taking any medications, and had no
autonomic dysfunction or cardiovascular diseases. Subjects who were
endurance or resistance trained were excluded from the study. Subjects arrived
at the laboratory fasted and had abstained from caffeine, alcohol, and exercise
for 12 h. The experimental protocol was approved by the Institutional Review
Board at the Pennsylvania State University College of Medicine and all subjects
gave written informed consent prior to participating.
3.2.2 Experimental Design
To serve as their own controls, subjects performed a normothermic and heating
trial during the same visit, but in random order. To regulate forearm temperature
subjects wore a water-perfused sleeve (Med-Eng Systems, Ottawa, ON, Canada)
over the exercising forearm (dominant arm was used). During the heating trial,
water at 55 ºC was circulated through the sleeve for 30 min. At the end of 30 min
the water was cooled to 50 ºC for the exercise protocol. For the normothermic
trial, water in the sleeve was regulated so that muscle temperature in the forearm
equaled initial measurements recorded during insertion of the muscle
temperature probe (~34 ºC). A minimum of 40 min separated the normothermic
and heating trials to allow all measures to return to baseline. Ambient
temperature in the laboratory during testing was 21-23 ºC.
3.2.3 Experimental Protocol.
The experimental protocol for both temperature conditions was as follows: 3-min
baseline, 1 min of ischemia, ischemic handgrip to fatigue, 1 min of postexercise

50
muscle ischemia, and 3-min recovery. During exercise, subjects squeezed a
hand dynamometer at 30% of their maximal voluntary contraction. Maximal
voluntary contraction was determined before the experimental protocol and
before muscle temperature probe insertion. The same workload was used for
each exercise trial. Measurements during each trial included muscle
temperature, arterial blood pressure, heart rate, MSNA, renal artery blood flow
velocity, calf blood flow, and ratings of perceived exertion.
3.2.4 Measurements
Muscle temperature was measured using a 22-gauge hypodermic
intramuscular thermistor (YSI 552, Yellow Springs, OH, USA). The thermistor
was placed 2-3 cm below the skin into the flexor muscles of the forearm. To limit
the possibility that heating or cooling the probe at the surface of the skin altered
temperature readings in the muscle, the top of the probe was insulated from
direct contact with the water-perfused sleeve. Measurements were taken every
minute during baseline and at 30-s intervals for the remainder of the
experimental protocol. Continuous skin temperature of the exercising limb was
measured via two thermocouples attached to the forearm skin and routed
through a thermocouple meter (model TC-1000, Sabel Systems, Henderson, NV,
USA). Tympanic temperatures were recorded using a First Temp Genius
Tympanic Thermometer (Sherwood Medical, St. Louis, MO, USA) to monitor
possible changes in core body temperature during the protocol.
Multifiber recordings of MSNA were made by inserting a tungsten
microelectrode into the peroneal nerve at the head of the fibula. A reference

51
electrode was inserted subcutaneously 2-3 cm from the recording electrode.
Both electrodes were connected to a differential preamplifier and then to an
amplifier (total gain between 40,000-80,000), where the nerve signal was band-
pass filtered (700-2000 Hz) and integrated (time constant, 0.1 s) to obtain a
mean voltage display of nerve activity. A satisfactory recording of MSNA was
defined as spontaneous, pulse-synchronous bursts that increased during end-
expiratory apnea and did not change during stroking of the skin or auditory
stimulation (yell).
Doppler ultrasound (HDI 5000, ATL Ultrasound, Bothell, WA, USA) was
used to measure renal artery blood velocity. The renal artery was scanned using
the anterior abdominal approach. To scan the renal artery, a curved-array
transducer (2–5 MHz) with a 2.5-MHz pulsed Doppler frequency was used. The
probe insonation angle to the artery was ≤60°. The focal zone was set at the
depth of the artery. The transducer was held in the same place to record velocity
tracings during each trial and the data were obtained in the same phase of the
respiratory cycle. Doppler tracings were analyzed using the software of the ATL
to obtain renal artery blood velocity for each cardiac cycle. Because of
technological limitations, it is not possible to accurately measure renal artery
diameter using the ATL Doppler ultrasound machine, therefore an index of renal
vascular conductance was calculated by dividing renal artery blood velocity by
mean arterial blood pressure. Because one older subject’s renal artery could not
be imaged, renal artery blood velocities were no recorded in that subject.

52
Calf blood flow was measured using venous occlusion plethysmography.
A mercury-in-Silastic strain gauge (Hokanson, Bellevue, WA, USA) was placed
around the maximal circumference of the calf. The calf was positioned above the
heart. Blood flow to the foot was occluded by inflating an ankle cuff to 220
mmHg. To occlude venous outflow a thigh cuff (model CC 17, Hokanson) was
placed around the thigh and inflated to 50 mmHg for 7.5 s every 15 s. Venous
congestion caused by the thigh cuff increased calf volume, which caused the
mercury-in-Silastic strain gauge to stretch. The rate of change in electrical
resistance in the mercury-in-Silastic strain gauge as it stretched is directly
proportional to calf blood flow. Calf vascular conductance was calculated by
dividing calf blood flow by mean arterial blood pressure.
Heart rate and blood pressure were continuously recorded during all trials
using a Finometer (Finapres Medical Systems, Amsterdam, Netherlands).
Before all trials, resting brachial blood pressure (Dinamap, General Electric,
Waukesha, WI, USA) was recorded. Subjects were asked to give ratings of
perceived exertion every 30 s during exercise and at fatigue (7).
3.2.4 Data analysis.
Data, except renal blood velocities, were analyzed offline using Chart
5.4.2 software (ADI Instruments, Newcastle, Australia). Sympathetic bursts were
identified from inspection of mean voltage neurograms. MSNA at rest was
quantified as bursts per 30 s and as the sum of the area under individual bursts
expressed as arbitrary units of activity per minute (a.u./30 s). The largest burst at
rest was assigned an amplitude of 1,000 arbitrary units, and a portion of the

53
nerve recording in which there was neural silence (i.e., no efferent discharges) for
at least 5 s was used to set the baseline to zero. For comparison during exercise
and postexercise ischemia muscle, MSNA was expressed as bursts per 30 s and
as arbitrary units of activity per 30 s (a.u./30 s). Sympathetic nerve recordings
were obtained in 12 young and 6 older individuals.
Resting variables for each trial were compared using a paired t-test within
age groups and an unpaired t-test across age groups. Because exercise time
was different between temperature conditions, data were expressed as a percent
of time to fatigue. The following exercise time periods were averaged: 0-20%,
20-40%, 40-60%, 60-80%, and 80-100%. Individual group responses were
compared within themselves using a two-within repeated measures analysis of
variance (temperature x time). To compare responses between the young and
older subjects, effect differences were calculated by subtracting measures during
normothermia from measures during the heating trial. Effect differences during
exercise were compared using a one-within, one-between repeated-measures
analysis of variance (time x age), while effect differences during fatigue and
during postexercise muscle ischemia were compared using a one-factor analysis
of variance (age). Significance was considered at P < 0.05. Results were
expressed as mean ± S.E.M.
3.3 Results
3.3.1 Baseline.

54
Measurements at baseline are presented in Tables 3.1 and 3.2. At rest, renal
artery blood velocity, renal vascular conductance, and MSNA were higher in the
older group compared to the young at both temperatures. Heating significantly
increased forearm skin and muscle temperature in both groups. Forearm heating
did not alter tympanic temperature in either group. All other measured variables
were the same between age groups and temperature conditions.
3.3.2 Exercise
Exercise workload, time, and ratings of perceived exertion Exercise workload
was not significantly different between the young and the older group (10.5 ± 1.0
kg and 9.5 ± 0.4 kg, respectively). Exercise time was not different between the
young and older groups during normothermia (179 ± 10 s vs. 186 ± 7 s,
respectively). Both groups gripped for a shorter duration during heating
compared to normothermia (young, 128 ± 9 s; older 154 ± 6 s). The older group
gripped significantly longer than the young during heating. Ratings of perceived
exertion in the young were significantly higher during exercise and heating (Fig.
3.1). No differences in ratings of perceived exertion were observed during
exercise between trials in the older group. However, comparison of these
changes in ratings of perceived exertion between trials across groups revealed
that they were not significantly different.
Mean arterial blood pressure. Forearm heating augmented mean arterial blood
pressure increases in both groups during exercise, but there was no difference in
responses between age groups (Fig. 3.2) (P = 0.72). During postexercise

55
muscle ischemia, mean arterial blood pressure was significantly higher in the
young subjects during heating compared to normothermia. However, changes in
mean arterial blood pressure from normothermia to heating were not different
between age groups (P = 0.47).
Heart rate. Heart rate increased with exercise, but responses to exercise were
not different between heating and normothermia in either the young or older
group. Heating did not alter heart rate responses during postexercise muscle
ischemia in either the young or older group.
Renal vascular responses. Forearm heating significantly augmented the
decrease in renal blood flow velocity and renal vascular conductance during
exercise in both the young and older subjects (Fig. 3.3, 3.4). However, the
decreases were significantly greater in the older group (∆ renal blood flow
velocity, P = 0.02; ∆ renal vascular conductance, P = 0.01) compared to the
young (Fig. 3.5). Renal vascular conductance was significantly lower during
postexercise muscle ischemia in the older group during heating compared to
normothermia. However, heat-induced changes in renal vascular conductance
during postexercise muscle ischemia were not different between age groups (P =
0.26).
Calf vascular responses. Heating significantly augmented the decrease in calf
vascular conductance in both age groups during exercise (Fig. 3.6). The mean

56
values between heating and normothermia in the young and older subjects were
not different (P = 0.11). During postexercise muscle ischemia, changes in calf
vascular conductance were not different between temperatures or age groups
(young vs. old, P = 0.16).
Muscle sympathetic nerve activity. Heating the forearm elicited similar increases
in burst frequency, total activity, and percent change in total activity during
exercise in the young and older subjects (Fig. 3.7, 3.8, 3.9). During postexercise
muscle ischemia in the older group, there was a tendency for changes in MSNA
to be higher during forearm heating, but there were no statistical differences. The
differences in burst frequency, total activity, and percent change in total activity
between heating and normothermia during postexercise muscle ischemia were
not different between age groups (P = 0.20, 0.51, and 0.63, respectively).
3.4 Discussion
The goal of this study was to test the following two hypotheses: 1) aging
augments renal vasoconstriction during isometric handgrip with local muscle
heating and 2) aging would not alter the augmented increase in MSNA in
response to isometric handgrip during local muscle heating. The results of the
study indicate that the interaction between forearm heating and aging elicits
greater renal vasoconstriction during isometric handgrip. Aging does not alter
the increase in MSNA observed during isometric handgrip with forearm heating.

57
In agreement with Momen et al. (57), the results of the current study
indicate that aging augments mechanoreceptor mediated increases in renal
vasoconstriction during isometric handgrip. The current study is the first study to
demonstrate that local forearm heating further augments renal vasoconstriction
during isometric handgrip. This augmented response appears to be partially
mediated by increases in muscle mechanoreceptor sensitivity because this
augmentation was observed at exercise onset when mechanoreceptor-mediated
responses are greatest in humans (26). Because renal vasoconstriction was
greater during heating and postexercise muscle ischemia compared to
normothermia in the older group, the muscle metaboreceptors could also be
contributing to the greater renal vasoconstriction.
The renal responses to isometric handgrip with a heated forearm in the
current study differ from vascular responses to dynamic upright exercise in the
heat in older individuals. During upright cycling in the heat (~36 ºC), older
individuals decrease renal and splanchnic blood flow and increase skin blood
flow less than younger individuals (28, 35). This response during dynamic
cycling is different than that observed during forearm isometric exercise. In the
current study, isometric exercise in the heat elicited greater renal
vasoconstriction in the old. The mechanism(s) responsible for eliciting these
contrasting results is unknown. Augmented sympathetic outflow would be
observed during both exercise paradigms in the heat (72, 81). However, the
extent of the thermal stress on the body is markedly different between local
heating of the forearm versus whole-body heating. The cardiovascular

58
consequences of this difference in thermal load might precipitate the difference in
renal vascular responses.
Without measuring renal sympathetic nerve activity, it is difficult to
ascertain the mechanism for age-related differences in renal blood flow
regulation. At rest, renal norepinephrine spillover does not differ between young
and older humans (19). Age-related changes in renal spillover, turnover, and
clearance during dynamic upright exercise and isometric exercise are unknown,
but some evidence suggests that renal clearance drops below normal during
supine bicycle ergometry in older individuals (23). The results of the current
study and Momen et al. (57), suggest that renal sympathetic outflow during
isometric handgrip might be higher in older individuals than in the young or that
renal vascular responsiveness to vasoconstrictors is augmented with age.
Ray and Gracey (72) concluded that local forearm heating increased
blood pressure and MSNA responses to exercise because heating increased
sensitivity of muscle mechanoreceptors. The current findings agree with Ray and
Gracey (72) because mean arterial blood pressure and MSNA were greater
during exercise and forearm heating compared to normothermic exercise. The
current study expands upon these findings because it is the first study to
demonstrate that blood pressure and MSNA responses to isometric exercise
mediated by muscle afferents are affected by muscle temperature in older
humans as well. The fact that we did not find age-related differences in MSNA
responses to exercise at either temperature is in agreement with other studies

59
that have reported that whole body norepinephrine spillover and MSNA
responses to isometric handgrip do not change with age (18, 64).
In agreement with previous findings, MSNA was elevated in the older
subjects at rest (16, 63, 74, 93). Because of this elevation we compared both
absolute and relative changes in MSNA between age groups. Previous studies
have found that during exercise in normothermic conditions, absolute changes in
MSNA in older individuals are either similar (64) or less (49) than in younger
individuals. We observed that aging did not alter absolute changes in MSNA
during isometric handgrip and forearm heating. Studies that have found
attenuation of the absolute increases in MSNA during exercise in older
individuals used dynamic handgrip (49), whereas, those reporting no differences
in absolute increases used isometric handgrip (64). Therefore, age-related
changes in MSNA responses during isometric and dynamic handgrip may be a
function of the mode of exercise utilized.
In both age groups heating augmented MSNA during exercise, this
increase was reflected by an increase in calf vasoconstriction during exercise.
The MSNA responses during postexercise muscle ischemia were also mirrored
by similar changes in calf vascular responses at both temperatures in each age
group. Increases in arterial blood pressure during heating in the young is related
to augmented peripheral and renal vasoconstriction (39). In the current study,
aging did not alter changes in mean arterial blood pressure during isometric
exercise with local muscle heating. However, renal vasoconstriction was greater
during exercise in the older subjects, indicating that the contribution of renal

60
vasoconstriction to the overall blood pressure response to exercise is different
between young and older subjects. One possible mechanism that would produce
comparable blood pressure responses to exercise despite lower renal
vasoconstriction is higher cardiac output in the young during the heating trial
compared to the older group. In agreement with this concept is that exercise-
induced increases in myocardial contractility decline with age (20). Another
possible mechanism is that younger subjects may have greater vasoconstriction
in another vascular bed not measured in the current study (e.g., splanchnic).
Finally, ratings of perceived exertion during exercise were not altered in the older
group during the heating compared to the normothermia trial; this finding
suggests that central command did not contribute to age-related differences in
renal vasoconstriction in the current study. Furthermore, the increase in MSNA
responses during exercise with heating was comparable between the young and
older groups. This result further supports the notion that central command was
not responsible for greater renal vasoconstriction in the older group.
This study has several limitations. First, to limit changes in muscle
temperature during exercise because of exercise-induced hyperemia, subjects
performed handgrip while ischemic. It is possible that changes in blood flow
during contraction may alter the effect of thermal stress on the exercise pressor
reflex. However, isometric contractions greater than 15% of maximal voluntary
contraction do not raise muscle blood flow and are therefore largely ischemic
(22). Second, in the current study Doppler ultrasound cannot accurately
measure renal artery diameter and be used to examine if changes in renal artery

61
diameter could account for age-related changes in renal vascular responses to
normothermic and forearm heated handgrip. There is some evidence to suggest
that pharmacological-mediated renal vasoconstriction (50) and vasodilation (47)
do not alter diameter of the renal artery. Furthermore, the vessel we examined
was a conduit artery and not a resistance vessel. Therefore, it is unlikely that
changes in renal artery diameter influenced the results of the study.
In summary, we compared renal and sympathetic responses to isometric
handgrip before and after local forearm heating in young and older subjects.
Heating elicited comparable increases in mean arterial blood pressure and
MSNA regardless of age. In the older subjects, the augmented pressure
response was mediated by a larger increase in renal vasoconstriction compared
to the young. Our results also reinforce the marked differences in hemodynamic
responses that are observed between isometric and dynamic exercise
modalities.

62
3.5 References 1. Borg G. Subjective aspects of physical and mental load. Ergonomics 21: 215-220, 1978.
2. Dinenno FA and Joyner MJ. Alpha-adrenergic control of skeletal muscle circulation at rest and during exercise in aging humans. Microcirculation 13: 329-341, 2006.
3. Esler MD, Thompson JM, Kaye DM, Turner AG, Jennings GL, Cox HS, Lambert GW, and Seals DR. Effects of aging on the responsiveness of the human cardiac sympathetic nerves to stressors. Circulation 91: 351-358, 1995.
4. Esler MD, Turner AG, Kaye DM, Thompson JM, Kingwell BA, Morris M, Lambert GW, Jennings GL, Cox HS, and Seals DR. Aging effects on human sympathetic neuronal function. Am J Physiol Regul Integr Comp Physiol 268: R278-285, 1995.
5. Ferrari AU, Radaelli A, and Centola M. Invited review: aging and the cardiovascular system. J Appl Physiol 95: 2591-2597, 2003.
6. Hansen J, Jacobsen TN, and Amtorp O. The exercise pressor response to sustained handgrip does not augment blood flow in the contracting forearm skeletal muscle. Acta Physiol Scand 149: 419-425, 1993.
7. Hasking GJ, Esler MD, Jennings GL, Dewar E, and Lambert G. Norepinephrine spillover to plasma during steady-state supine bicycle exercise. Comparison of patients with congestive heart failure and normal subjects. Circulation 78: 516-521, 1988.
8. Hayes SG and Kaufman MP. MLR stimulation and exercise pressor reflex activate different renal sympathetic fibers in decerebrate cats. J Appl Physiol 92: 1628-1634, 2002.
9. Herr MD, Imadojemu V, Kunselman AR, and Sinoway LI. Characteristics of the muscle mechanoreflex during quadriceps contractions in humans. J Appl Physiol 86: 767-772, 1999.

63
10. Ho CW, Beard JL, Farrell PA, Minson CT, and Kenney WL. Age, fitness, and regional blood flow during exercise in the heat. J Appl Physiol 82: 1126-1135, 1997.
11. Kenney WL and Ho CW. Age alters regional distribution of blood flow during moderate-intensity exercise. J Appl Physiol 79: 1112-1119, 1995.
12. Kuipers NT, Sauder CL, Kearney ML, and Ray CA. Changes in forearm muscle temperature alter renal vascular responses to isometric handgrip. Am J Physiol Heart Circ Physiol 293: H3432-3439, 2007.
13. Manoharan G, Pijls NH, Lameire N, Verhamme K, Heyndrickx GR, Barbato E, Wijns W, Madaric J, Tielbeele X, Bartunek J, and De Bruyne B. Assessment of renal flow and flow reserve in humans. J Am Coll Cardiol 47: 620-625, 2006.
14. Mark AL, Victor RG, Nerhed C, and Wallin BG. Microneurographic studies of the mechanisms of sympathetic nerve responses to static exercise in humans. Circ Res 57: 461-469, 1985.
15. Markel TA, Daley JC, 3rd, Hogeman CS, Herr MD, Khan MH, Gray KS, Kunselman AR, and Sinoway LI. Aging and the exercise pressor reflex in humans. Circulation 107: 675-678, 2003.
16. Marraccini P, Fedele S, Marzilli M, Orsini E, Dukic G, Serasini L, and L'Abbate A. Adenosine-induced renal vasoconstriction in man. Cardiovasc Res 32: 949-953, 1996.
17. Middlekauff HR, Nitzsche EU, Nguyen AH, Hoh CK, and Gibbs GG. Modulation of renal cortical blood flow during static exercise in humans. Circ Res 80: 62-68, 1997.
18. Momen A, Leuenberger UA, Handly B, and Sinoway LI. Effect of aging on renal blood flow velocity during static exercise. Am J Physiol Heart Circ Physiol 287: H735-740, 2004.
19. Momen A, Leuenberger UA, Ray CA, Cha S, Handly B, and Sinoway LI. Renal vascular responses to static handgrip: role of muscle mechanoreflex. Am J Physiol Heart Circ Physiol 285: H1247-1253, 2003.

64
20. Moore R, Mallonee S, Sabogal RI, Zanardi L, Redd J, and Malone J. From the Centers for Disease Control and Prevention. Heat-related deaths--four states, July-August 2001, and United States, 1979-1999. JAMA 288: 950-951, 2002.
21. Ng AV, Callister R, Johnson DG, and Seals DR. Age and gender influence muscle sympathetic nerve activity at rest in healthy humans. Hypertension 21: 498-503, 1993.
22. Ng AV, Callister R, Johnson DG, and Seals DR. Sympathetic neural reactivity to stress does not increase with age in healthy humans. Am J Physiol Heart Circ Physiol 267: H344-353, 1994.
23. Ray CA and Gracey KH. Augmentation of exercise-induced muscle sympathetic nerve activity during muscle heating. J Appl Physiol 82: 1719-1725, 1997.
24. Ray CA and Monahan KD. Aging attenuates the vestibulosympathetic reflex in humans. Circulation 105: 956-961, 2002.
25. Rowell LB, Brengelmann GL, and Freund PR. Unaltered norepinephrine-heart rate relationship in exercise with exogenous heat. J Appl Physiol 62: 646-650, 1987.
26. Sundlof G and Wallin BG. Human muscle nerve sympathetic activity at rest. Relationship to blood pressure and age. J Physiol 274: 621-637, 1978.

65
Table 3.1. Baseline measurements during normothermia and heating in the young group
Young (n=12)
Variable Normothermia Heating
Muscle Temperature (ºC) 34.0 ± 0.3 38.8 ± 0.2*
Tympanic Temperature (ºC) 36.7 ± 0.2 36.7 ± 0.1
Skin Temperature (ºC) 30.9 ± 0.4 40.3 ± 0.2*
MAP (mmHg) 87 ± 2 89 ± 2
Heart Rate (beats/min) 60 ± 3 63 ± 4
MSNA (bursts/30 s) 4 ± 1 5 ± 1
MSNA (a.u./30 s) 680 ± 110 960 ± 230
Renal Blood Velocity (cm/s) 52.7 ± 2.5 53.2 ± 2.5
RVC (cm/s/mmHg) 0.59 ± 0.05 0.62 ± 0.06
Calf Blood Flow (ml/min/dl) 1.8 ± 0.2 2.0 ± 0.2
CVC (ml/min/dl/mmHg) 0.019 ± 0.002 0.022 ± 0.002
MAP, mean arterial blood pressure; MSNA, muscle sympathetic nerve activity; RVC, renal vascular conductance; CVC, calf vascular conductance. *Significantly different from normothermia (P < 0.05). Values are mean ± S.E.M.

66
Table 3.2. Baseline measurements during normothermia and heating in the older group
Older (n=9)
Variable Normothermia Heating
Muscle Temperature (ºC) 34.5 ± 0.4 38.7 ± 0.2*
Tympanic Temperature (ºC) 36.3 ± 0.1 36.3 ± 0.1
Skin Temperature (ºC) 31.2 ± 0.5 39.6 ± 0.2*
MAP (mmHg) 90 ± 2 92 ± 1
Heart Rate (beats/min) 66 ± 2 66 ± 2
MSNA (bursts/30 s) 15 ± 3* 13 ± 2*
MSNA (a.u./ 30 s) 1928 ± 382† 1938 ± 499†
Renal Blood Velocity (cm/s) 71.6 ± 11.0† 77.8 ± 10.8†
RVC (cm/s/mmHg) 0.80 ± 0.25† 0.76 ± 0.21†
Calf Blood Flow (ml/min/dl) 1.6 ± 0.2 1.5 ± 0.1
CVC (ml/min/dl/mmHg) 0.018 ± 0.002 0.017 ± 0.001
MAP, mean arterial blood pressure; MSNA, muscle sympathetic nerve activity; RVC, renal vascular conductance; CVC, calf vascular conductance. †Significantly different from young. *Significantly different from normothermia (P < 0.05). Values are mean ± S.E.

67
0 20 40 60 80 100
6
8
10
12
14
16
18
20
Young
34 ºC
39 ºC
time x temperature p = 0.04
Time to Fatigue (%)
Ra
tin
gs
of
Pe
rce
ive
d E
xe
rtio
n
0 20 40 60 80 100
Older
time x temperature p = 0.66
Time to Fatigue (%)
Figure 3.1. Ratings of perceived exertion during exercise in the normothermic and heating trials in the young and older subjects. Heating increased ratings of perceived exertion in the young but not in the older subjects during exercise. However, heat-induced changes in ratings of perceived exertion during exercise were not significantly different between age groups. P < 0.05. Values are mean ± SE.

68
0
10
20
30
40
50
60
34 ºC
39 ºC
time x temperature p < 0.05
Young
! M
ea
n A
rte
ria
l B
loo
d P
res
su
re (
mm
Hg
)
0 20 40 60 80 1000
10
20
30
40
50
60
time x temperature p < 0.05
Older
Time to Fatigue (%)
! M
ea
n A
rte
ria
l B
loo
d
Pre
ss
ure
(m
mH
g)
34 ºC
39 ºC
*p = 0.01
PEMI
p = 0.20
Figure 3.2. Changes in mean arterial blood pressure (MAP) from baseline
during forearm heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. The increase in mean arterial blood pressure during exercise was comparable in the young and older groups. *Significantly different from normothermia (P < 0.05). Values are mean ± SE.

69
-40
-30
-20
-10
0
34 ºC
39 ºC
time x temperature p < 0.05
Young
! R
en
al
Blo
od
Ve
loc
ity
(c
m/s
)
20 40 60 80 100-40
-30
-20
-10
0
time x temperature p < 0.05
Older
Time to Fatigue (%)
! R
en
al
Blo
od
Ve
loc
ity
(c
m/s
)
34 ºC
39 ºC
p = 0.68
PEMI
p = 0.22
Figure 3.3. Changes in renal blood velocity from baseline during forearm
heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. Heat-induced decreases in renal blood velocity were significantly greater than normothermia in both groups during exercise. Heat-induced decreases in renal vascular conductance were significantly greater in the older than young subjects during exercise. Heat-altered responses were not significantly different between age groups during postexercise muscle ischemia. P< 0.05. Values are mean ± SE.

70
-75
-60
-45
-30
-15
0
34 ºC
39 ºC
time x temperature p < 0.05
Young
! R
en
al
Va
sc
ula
r C
on
du
cta
nc
e (
%)
20 40 60 80 100-75
-60
-45
-30
-15
0
time x temperature p < 0.05
Older
Time to Fatigue (%)
! R
en
al
Va
sc
ula
r C
on
du
cta
nc
e (
%)
34 ºC
39 ºC
p = 0.12
PEMI
*p = 0.04
Figure 3.4. Percent changes in renal vascular conductance from baseline
during forearm heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. Heat-induced decreases in renal vascular conductance were significantly greater than normothermia in both groups during exercise. Heat-induced decreases in renal vascular conductance were significantly greater in the older subjects than in the young during exercise. Heat-altered responses were not significantly different between age groups during postexercise muscle ischemia. *Significantly different from normothermia (P < 0.05). Values are mean ± SE.

71
-25
-20
-15
-10
-5
0
Young
Older
-25
-20
-15
-10
-5
0
* *
! R
BV
(cm
/s) !
RV
C (%
)
Figure 3.5. Renal blood velocity (RBV) and renal vascular conductance (RVC)
differences between heating and normothermia in both age groups at fatigue. The decrease in renal blood velocity and vascular conductance during exercise was greater in the older compared to the young. *Significantly different from young (P < 0.05). P < 0.05. Values are mean ± SE.

72
-40
-20
0
20
40
60
34 ºC 39 ºC
time x temperature p < 0.05
Young
! C
alf
Va
sc
ula
r C
on
du
cta
nc
e
(%)
20 40 60 80 100-40
-20
0
20
40
60
time x temperature p < 0.05
Older
Time to Fatigue (%)
! C
alf
Va
sc
ula
r C
on
du
cta
nc
e
(%)
34 ºC
39 ºC
p = 0.47
PEMI
p = 0.25
Figure 3.6. Percent change in calf vascular conductance from baseline during
forearm heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. Heating augmented calf vasoconstriction during exercise in both age groups. Heating did not alter calf vascular conductance during postexercise muscle ischemia in either group. P < 0.05. Values are mean ± SE.

73
0
5
10
15
20
25
34 ºC
39 ºC
time x temperature p < 0.05
Young
! M
SN
A (
bu
rsts
/ 3
0 s
)
0 20 40 60 80 1000
5
10
15
20
25
Older
time x temperature p < 0.05
Time to Fatigue (%)
! M
SN
A (
bu
rsts
/ 3
0 s
)
34 ºC
39 ºC
p = 0.07
PEMI
p = 0.13
Figure 3.7. Changes in muscle sympathetic nerve activity (MSNA) burst
frequency from baseline during forearm heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. Heating augmented burst frequency during exercise in both groups but the response was not significantly different between age groups. Heating did not alter burst frequency during postexercise muscle ischemia. P < 0.05. Values are mean ± SE.

74
0
2000
4000
6000
8000
10000
12000 34 ºC 39 ºC
time x temperature p < 0.05
Young
! M
SN
A (
a.u
. /
30
s)
0 20 40 60 80 100
0
2000
4000
6000
8000
10000
12000
Older
time x temperature p < 0.05
Time to Fatigue (%)
! M
SN
A (
a.u
. /
30
s)
34 ºC
39 ºC
p = 0.07
PEMI
p = 0.24
Figure 3.8. Change in total muscle sympathetic nerve activity (MSNA) from
baseline during forearm heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. Heating augmented total activity during exercise in both groups but the response was not significantly different between age groups. Heating did not alter total activity during postexercise muscle ischemia. P < 0.05. Values are mean ± SE.

75
0
500
1000
1500
2000
2500
34 ºC
39 ºC
time x temperature p < 0.05
Young
! T
ota
l M
SN
A (
%)
0 20 40 60 80 100
0
500
1000
1500
2000
2500
Older
time x temperature p < 0.05
Time to Fatigue (%)
! T
ota
l M
SN
A (
%)
34 ºC
39 ºC
p = 0.76
PEMI
p = 0.06
Figure 3.9. Percent change in total muscle sympathetic nerve activity (MSNA)
from baseline during forearm heating and normothermia during exercise and postexercise muscle ischemia (PEMI) in the young and older groups. Heating augmented relative total activity during exercise in both groups but the response was not significantly different between age groups. Heating did increase relative total activity in the older group during postexercise muscle ischemia; however, responses were not significantly different between age groups. P < 0.05. Values are mean ± SE.

76
Chapter 4
ACE-Inhibition Does Not Alter Sympathetic and Vascular Reponses to Isometric Exercise During Forearm Heating
4.1 Introduction
The normal cardiovascular response to isometric exercise includes
increases in blood pressure, heart rate, and sympathetic neural activity. These
responses are mediated by central command, the baroreflexes, and skeletal
muscle afferents (29). Drug treatments may alter physiological systems in the
body that influence these reflexes. For example, ACE-inhibition blocks the
breakdown of kinins in the blood, which can sensitize muscle afferents in animals
during exercise (67, 91). Likewise, blood prostaglandin levels , which increase
during ACE-inhibition (66), influence muscle afferent sensitivity (56, 76, 78). In
humans, little is known about the influences of ACE-inhibition on central
hemodynamic, vascular, and muscle sympathetic nerve activity (MSNA)
responses during exercise.
Changes in muscle temperature alter muscle afferent sensitivity, which
can alter cardiovascular and MSNA responses to exercise (72, 73). For
example, heating the forearm increases blood pressure, renal vasoconstriction,
and MSNA responses to isometric exercise in humans (39, 72). These heat-
induced changes are observed at the onset of exercise when mechanoreceptor-
mediated cardiovascular responses to exercise are greatest (72). Therefore,
heat-induced changes during exercise might be mediated by increased sensitivity
of muscle mechanoreceptors. The mechanisms that increase sensitivity of
muscle mechanoreceptors during exercise in the heat are unknown. Factors

77
changed by ACE-inhibition might have a role in altering mechanoreceptor
sensitivity in the heat. For example, bradykinin alters sensitivity of mechano-
heat-sensitive receptors in the skin (51, 69). Therefore, ACE-inhibition might
provide insight into heat-related changes in muscle afferent sensitivity.
The first goal of this study was to examine the effect of ACE-inhibition on
the exercise pressor reflex during normothermic isometric exercise. Because
ACE-inhibition increases blood kinin levels, which act on muscle afferents in
animals, it was hypothesized that acute ACE-inhibition would augment central
hemodynamic, vascular, and MSNA responses to isometric handgrip. The
second goal of this study was to examine if ACE-inhibition would alter muscle
afferent sensitivity in the heat. It was hypothesized that ACE-inhibition would
further augment blood pressure, renal vasoconstriction, and MSNA responses to
isometric handgrip when the forearm was hyperthermic because of higher blood
kinin levels, which are known to increase sensitivity of mechano-heat sensitive
afferents (51, 69).
4.2 Methods
4.2.1 Subjects
Eight male subjects participated in the study (age, 25 ± 2 yr; height, 177.6
± 2.3 cm; weight, 76.2 ± 5.9 kg). All subjects were normotensive, non-obese,
non-smokers, not taking any medications, and had no autonomic dysfunction or
cardiovascular diseases. Subjects who were endurance or resistance trained
were excluded from study. The experimental protocol was approved by the

78
Institutional Review Board at the Pennsylvania State University College of
Medicine and all subjects gave written informed consent prior to participating.
The study was conducted in a double-blinded, placebo controlled, and
randomized design. Subjects came to the lab on two separate days. Each day
subjects arrived at the laboratory in the morning fasted, and had abstained from
caffeine, alcohol, and physical activity for 12 h. During one visit subjects were
given 20 mg of the ACE-inhibitor quinapril (Pfizer, New York, NY, USA) and
during the other a placebo (sugar pill). In approximately 8 h quinapril doubles
blood kinin levels while blood angiotensin II levels return to normal. (68).
Subjects were given the same breakfast and snack each morning along with
plenty of water. Subjects were not allowed to eat 4 h before testing. The
experimental portion of the study was carried out in the afternoon beginning
approximately 7 to 8 h after the morning visit.
4.2.2 Experimental Design
The experimental portion of the study was the same each afternoon. To regulate
forearm muscle temperature, subjects wore a water-perfused sleeve (Med-Eng
Systems, Ottawa, ON, Canada) on their dominant forearm. For the heating trial,
water at 55 ºC was circulated through the sleeve for 30 min to increase muscle
temperature ~ 3 ºC above normal. At the end of 30 min, the water temperature
was reduced to 50 ºC for the exercise protocol. The order of the normothermic
and heating trials were randomized. When the normothermic trial was second,
the temperature of the circulating water was adjusted until forearm muscle
temperature equaled preheating measurements. A minimum of 40 min separated

79
the normothermia and heating trial. Ambient temperature in the laboratory during
testing was 21-23 ºC.
4.2.3 Experimental Protocol
The experimental protocol at each temperature was as follows: 5-min
baseline, 1 min of ischemia, ischemic handgrip to fatigue, 1 min of postexercise
muscle ischemia, and 3-min recovery. During exercise, subjects squeezed a
hand dynamometer at 30% of their maximal voluntary contraction. Maximal
voluntary contraction was determined before the experimental protocol and
before muscle temperature probe insertion. Measurements during each trial
included muscle temperature, skin temperature, arterial blood pressure, heart
rate, renal artery blood velocity, MSNA, and rate of perceived exertion.
4.2.4 Measurements
Muscle temperature was measured using a 22-gauge hypodermic
intramuscular thermistor (YSI 552, Yellow Springs, OH). The thermistor was
placed 2-3 cm below the skin into the flexor muscles of the forearm. To limit the
possibility that heat from the sleeve directly influenced probe measurements, the
thermistor was fully inserted into the subjects and the top of the probe was
insulated from direct contact with the water-perfused sleeve. Measurements
were taken every minute during baseline and at 30 s intervals for the remainder
of the experimental protocol. Continuous skin temperature of the exercising limb
was measured via two thermocouples attached to the dorsal forearm skin and
routed through a thermocouple meter (model TC-1000, Sabel Systems,
Henderson, NV). Tympanic temperatures were recorded using a First Temp

80
Genius tympanic thermometer (Sherwood Medical, St. Louis, MO) after changing
forearm muscle temperature to monitor for possible changes in body
temperature.
Multifiber recordings of MSNA were made with a tungsten microelectrode
inserted into the peroneal nerve at the head of the fibula. A reference electrode
was inserted subcutaneously 2-3 cm from the recording electrode. Both
electrodes were connected to a differential preamplifier and then to an amplifier
(total gain between 40,000-80,000), where the nerve signal was band-pass
filtered (700-2000 Hz) and integrated (time constant, 0.1 s) to obtain a mean
voltage display of nerve activity. A satisfactory recording of MSNA was defined
as spontaneous, pulse-synchronous bursts that increased during end-expiratory
apnea and did not change during stroking of the skin or auditory stimulation
(yell).
Doppler ultrasound (HDI 5000, ATL Ultrasound, Bothell, WA, USA) was
used to measure renal artery blood velocity. The renal artery was scanned using
the anterior abdominal approach. To scan the renal artery, a curved-array
transducer (2–5 MHz) with a 2.5-MHz pulsed Doppler frequency was used. The
probe insonation angle to the artery was ≤ 60°. The focal zone was set at the
depth of the artery. The transducer was held in the same place to record velocity
tracings during each trial and the data were obtained in the same phase of the
respiratory cycle. To obtain renal blood velocity measurements, the entire length
of each cardiac cycle waveform in the Doppler signal was traced by hand and
analyzed using the ATL computer’s software package. For the experimental

81
protocol 3 to 5 cardiac cycles were averaged every 15 s except at exercise onset
and fatigue where 15 s periods were averaged. An index of renal vascular
conductance was calculated by dividing renal artery blood velocity by mean
arterial blood pressure.
To measure blood pH, lactate, K+, and Na+ levels 1 ml of blood was drawn
at baseline and fatigue at each temperature through a retrograde catheter
inserted into a vein at the antecubital fossa of the exercising arm (Bayer Blood
Gas Analyzer, Bayer Healthcare Diagnostics Division, New York, NY, USA). For
both visits, blood was drawn in the morning, baseline, and fatigue at both
temperatures to measure blood bradykinin levels. Bloods samples were sent to
two different laboratories, but because of technical issues blood bradykinin levels
could not be measured.
Heart rate and arterial blood pressure were continuously recorded during
all trials using a Finometer (Finapres Medical Systems, Amsterdam,
Netherlands). Before all trials resting brachial artery blood pressure (Dinamap,
General Electric, Waukesha, WI, USA) was recorded. Subjects were asked to
give ratings of perceived exertion every 30 s during exercise and once at fatigue
(7).
4.2.5 Data analysis
Data, except renal blood flow velocities, were analyzed offline using Chart
5.4.2 software (ADI Instruments, Newcastle, Australia). Sympathetic bursts were
identified from inspection of mean voltage neurograms. MSNA at rest and during
exercise, was quantified as bursts per 30 s and as the sum of the area under

82
individual muscle sympathetic nerve bursts expressed as arbitrary units of activity
per 30 s (a.u./30 s). The largest burst at rest was assigned an amplitude of 1,000
arbitrary units, and a portion of the nerve recording in which there was neural
silence (i.e., no efferent discharges) for at least 5 s was used to set the baseline
to zero.
Resting variables for each temperature condition and treatment were
compared using a paired t-test. Baseline and postexercise muscle ischemia data
were averaged over their respective time periods. Because exercise time
differed between temperature conditions, data were expressed as a percent of
time to fatigue. The five exercise time periods averaged were 0-20%, 20-40%,
40-60%, 60-80%, and 80-100%. To compare drug and placebo effects on
responses at normothermia, a two-within repeated-measures analysis of variance
(condition x time) was used. To compare heating trials during both treatments,
effect differences were calculated by subtracting normothermia measurements
from heating measurements and compared using a two-factor within-repeated
measures analysis of variance (condition x time). Significance was considered at
a P value of < 0.05. Results were expressed as mean ± S.E.
4.3 Results
Baseline. Baseline measurements are presented in Table 4.1 and 4.2.
Quinapril decreased resting mean arterial blood pressure by 8 ± 3 mmHg from
morning to afternoon while placebo treatment did not alter resting blood

83
pressure. Heart rate did not change from morning to afternoon, with or without
quinapril.
Heating the forearm did not alter resting heart rate on either day. Blood
Na+ levels were significantly higher and blood lactate levels significantly lower
during the normothermic quinapril trial. Calf and renal conductance and MSNA
were comparable throughout the different drug and temperature trials. Forearm
muscle and skin temperatures were significantly elevated during the heating
trials.
Exercise Responses. Exercise handgrip time was not changed with quinapril.
Regardless of drug treatment, subjects gripped significantly longer during
normothermia compared to heating. Muscle and skin temperatures were not
altered by quinparil treatment and temperatures were not different between the
two respective heating and normothermic trials. Ratings of perceived exertion
were not different between drug treatments or forearm muscle temperatures.
Mean arterial blood pressure and heart rate responses to exercise. Quinapril did
not alter mean arterial blood pressure responses to handgrip or postexercise
muscle ischemia during normothermia (Fig. 4.1). Heart rate responses during
exercise and postexercise muscle ischemia were not different between drug
treatments and forearm muscle temperatures. Heating significantly augmented
mean arterial blood pressure responses to exercise during both drug trials.
Heating augmented the increase in mean arterial pressure during postexercise

84
muscle ischemia in the drug trial. Heat-induced changes in mean arterial blood
pressure during exercise and postexercise muscle ischemia were not different
between drug treatments.
Renal blood flow responses. Quinapril did not alter renal vascular responses to
handgrip or postexercise muscle ischemia during normothermia (Fig. 4.2).
Increasing forearm temperature augmented renal vasoconstriction during
exercise. The augmented renal vasoconstriction during heating was not different
between drug treatments. Heat-induced changes in renal vascular conductance
during postexercise muscle ischemia were not different between drug treatments.
Because blood pressure changes were comparable across trials, changes in
renal vascular conductance followed the same pattern as changes in renal blood
flow velocities for all trials.
Calf blood flow responses. Calf blood flow responses during exercise and
postexercise muscle ischemia were not different between treatments and
forearm temperatures (Fig. 4.3).
Muscle sympathetic nerve activity. Quinapril treatment did not alter MSNA
responses to handgrip or postexercise muscle ischemia during normothermia.
Increasing forearm temperature significantly augmented burst frequency (Fig.
4.4,4.5) and total activity during exercise for both treatments. Heat-induced

85
changes in burst frequency and total activity were not different between drug
treatments during exercise and postexercise muscle ischemia.
Blood metabolite measures. The changes in blood pH, lactate, sodium and
potassium from rest to fatigue were not different between temperatures or drug
treatments.
4.4 Discussion
The goal of this study was to examine if acute ACE-inhibition alters the
exercise pressor reflex in humans during isometric handgrip. The major finding
of the study is that acute ACE-inhibition, which alters resting blood pressure,
does not alter exercise-induced increases in arterial blood pressure, heart rate,
renal and calf vasoconstriction, and MSNA. These findings also were observed
when the temperature of the exercising muscle was increased.
Quinapril treatment lowered resting blood pressure indicating that ACE
activity was altered in the subjects. Despite the lower resting blood pressure,
ACE-inhibition does not alter central hemodynamic responses to isometric
exercise with normothermic muscle. More importantly, because of the increase
in muscle temperature during exercise, the current study is the first to find that
ACE-inhibition does not alter hemodynamic responses to isometric exercise with
hyperthermic muscle. These findings indicate that short-term ACE-inhibition, and
the subsequent alterations in the renin-angiotenin and kallikrein-kinin systems,
are insufficient to alter central hemodynamic responses to exercise.
Changes in renal and peripheral vasculature function are important

86
mechanisms used to control hemodynamic responses to exercise and thermal
stress. During treadmill running, ACE-inhibition will decrease renal blood flow
more compared to vehicle treatment in pigs (94). In contrast, during electrical
stimulation of rabbit renal sympathetic nerves, ACE-inhibition will blunt renal
vasoconstriction (44). The current study found that acute ACE-inhibition, at a
normally prescribed dosage, does not alter renal vasoconstriction during
isometric exercise in humans. Furthermore, even during periods of greater renal
vasoconstriction (i.e., exercise with a heated forearm), there is still no effect of
ACE-inhibition. In human peripheral vasculature, angiotensin II is a
vasoconstrictor and bradykinin is a vasodilator. The results of the current study
indicate that, like the renal vasculature, acute alterations in the renin-angiotensin
and kallikrein-kinin systems do not alter blood flow responses in non-exercising
limbs.
Angiotensin II and bradykinin can increase norepinephrine release from
sympathetic neurons in animals (8, 41). Therefore, it was unapparent if ACE-
inhibitor-induced changes in the renin-angiotensin and kallikrein-kinin systems,
and subsequent changes in norepinephrine release, might have been offset by
changes in MSNA during exercise. The current study found that ACE-inhibition
does not alter MSNA responses to isometric exercise in humans, indicating that
ACE-inhibitor-induced changes in norepinephrine release from the sympathetic
neuron are not offset by changes in MSNA in humans.
The reason why ACE-inhibition does not alter central hemodynamic,
vascular, and MSNA responses to isometric exercise in humans remains unclear.

87
In animal studies, kinins influence cardiovascular and sympathetic responses to
exercise; however, the dosages of bradykinin infused into the muscle vascular
bed were much higher than changes in blood bradykinin levels reported in
humans during ACE-inhibition (31, 54, 68). Therefore, the increases in blood
kinins caused by ACE-inhibition might be insufficient to alter muscle afferent
activity in humans. However, the concentrations observed by Pellacani et al.
(68) during quinapril treatment, as used in this study, were closer to those of
patients on ACE-inhibitor therapy indicating clinical relevance and importance to
the results of the current study.
A second effect of ACE-inhibition that might have influenced our results is
the decrease in bioavailability of angiotensin II because of a decrease in the
breakdown of angiotensin I to angiotensin II. However, in humans angiotensin II
levels do not increase during isometric handgrip (100), and 8 h after quinapril,
treatment blood angiotensin II levels are reported to be normal (68). Therefore,
the short-term changes in angiotensin II physiology are unlikely to have
influenced the results of the current study.
The results of the current study focused only on the acute influences of
exercise and ACE-inhibition on central hemodynamics, vascular responses, and
MSNA. Dynamic leg exercise and chronic ACE-inhibition might alter
hemodynamic and sympathetic responses to exercise differently. For example,
exercise training lowers plasma angiotensin II levels in chronic heart failure rats
(61), which might contribute to lowering sympathetic nerve activity. Future
studies are needed to address these questions.

88
Previously, we have reported that increasing local forearm temperature
augments blood pressure, renal vascular, and MSNA responses to isometric
handgrip (39, 72). Because heat-induced increases in cardiovascular and
MSNA responses occur at the onset of exercise when mechanoreceptors are
engaged and not during postexercise muscle ischemia when only the
metaboreceptors are engaged, it is believed that heat acts on the muscle
mechanoreceptors (72). The factors that increase mechanoreceptors sensitivity
during heating remain uncertain. The current study suggests that kinins do not
increase sensitivity of muscle mechanoreceptors in humans at elevated muscle
temperatures because neurovascular responses during hyperthermic isometric
exercise were not altered by ACE-inhibition. To determine if any metabolic
factors influence muscle afferent sensitivity during local heating and exercise we
measured blood pH, lactate, and potassium at baseline and fatigue. Hydrogen
ions, lactate, and potassium have all been found to alter the pattern of
cardiovascular responses to exercise mediated by muscle mechano- and
metaboreceptors (30, 86). During the placebo trial, local heating did not alter
these values at baseline or at fatigue. These findings suggest that changes in
these local metabolites do not alter muscle afferent sensitivity during local
forearm heating and isometric handgrip.
Based upon the results of the current study, the exact mechanisms that
increase in central hemodynamic, vascular, and MSNA responses to isometric
handgrip and local forearm heating remains equivocal. Prostaglandins might or
might not sensitize muscle mechanoreceptors to exercise (17, 76, 78). It is

89
known that prostaglandin production increases during heating of human cells (9),
indicating that they might be playing a role in heat-induced responses. Because
bradykinin can influence prostaglandin levels in tissue (96), our results suggest
that if prostaglandins do play a role in heat-induced mechanoreceptor sensitivity
changes, they would be produced by a kinin-independent mechanism. In our
previous studies we ruled out the possibility that central command played a role
in the augmented responses in the heat because subjects did not complain of
heat-induced pain and because central command’s contribution to increases in
MSNA occur during maximal exertion and fatigue (98).
In summary, ACE-inhibition does not alter neurovascular responses to
isometric handgrip with a normothermic or hyperthermic forearm. Furthermore,
ACE-inhibition does not alter muscle afferent sensitivity during isometric
exercise.

90
4.5 References
1. Borg G. Subjective aspects of physical and mental load. Ergonomics 21: 215-220, 1978.
2. Brasch H, Sieroslawski L, and Dominiak P. Angiotensin II increases norepinephrine release from atria by acting on angiotensin subtype 1 receptors. Hypertension 22: 699-704, 1993.
3. Calderwood SK, Bornstein B, Farnum EK, and Stevenson MA. Heat shock stimulates the release of arachidonic acid and the synthesis of prostaglandins and leukotriene B4 in mammalian cells. J Cell Physiol 141: 325-333, 1989.
4. Doerzbacher KJ and Ray CA. Muscle sympathetic nerve responses to physiological changes in prostaglandin production in humans. J Appl Physiol 90: 624-629, 2001.
5. Kaufman MP and Forster HV. Reflexes controlling circulatory, ventilatory and airway responses to exercise. In: Handbook of Physiology: "Exercise Regulation and Integration of Multiple Systems". Bethesda, MD: Am. Physiol. Soc., 1996, p. 381-447.
6. Kaufman MP and Hayes SG. The exercise pressor reflex. Clin Auton Res 12: 429-439, 2002.
7. Kaufman MP, Iwamoto GA, Longhurst JC, and Mitchell JH. Effects of capsaicin and bradykinin on afferent fibers with ending in skeletal muscle. Circ Res 50: 133-139, 1982.
8. Kuipers NT, Sauder CL, Kearney ML, and Ray CA. Changes in forearm muscle temperature alter renal vascular responses to isometric handgrip. Am J Physiol Heart Circ Physiol 293: H3432-3439, 2007.
9. Kurz T, Tolg R, and Richardt G. Bradykinin B2-receptor-mediated stimulation of exocytotic noradrenaline release from cardiac sympathetic neurons. J Mol Cell Cardiol 29: 2561-2569, 1997.

91
10. Lang CC, Stein CM, He HB, and Wood AJ. Angiotensin converting enzyme inhibition and sympathetic activity in healthy subjects. Clin Pharmacol Ther 59: 668-674, 1996.
11. Le Fevre ME, Guild SJ, Ramchandra R, Barrett CJ, and Malpas SC. Role of angiotensin II in the neural control of renal function. Hypertension 41: 583-591, 2003.
12. Mayer S, Izydorczyk I, Reeh PW, and Grubb BD. Bradykinin-induced nociceptor sensitisation to heat depends on cox-1 and cox-2 in isolated rat skin. Pain, 2006.
13. Mense S. Nervous outflow from skeletal muscle following chemical noxious stimulation. J Physiol 267: 75-88, 1977.
14. Momen A, Cui J, McQuillan P, and Sinoway LI. Local Prostaglandin Blockade Attenuates Muscle Mechanoreflex Mediated Renal Vasoconstriction during Muscle Stretch in Humans. Am J Physiol Heart Circ Physiol, 2008.
15. Mousa TM, Liu D, Cornish KG, and Zucker IH. Exercise training enhances baroreflex sensitivity by an angiotensin II-dependent mechanism in chronic heart failure. J Appl Physiol 104: 616-624, 2008.
16. Niarchos AP, Pickering TG, Morganti A, and Laragh JH. Plasma catecholamines and cardiovascular responses during converting enzyme inhibition in normotensive and hypertensive man. Clin Exp Hypertens A 4: 761-789, 1982.
17. Nishimura H, Kubo S, Ueyama M, Kubota J, and Kawamura K. Peripheral hemodynamic effects of captopril in patients with congestive heart failure. Am Heart J 117: 100-105, 1989.
18. Pan HL, Stebbins CL, and Longhurst JC. Bradykinin contributes to the exercise pressor reflex: mechanism of action. J Appl Physiol 75: 2061-2068, 1993.
19. Pellacani A, Brunner HR, and Nussberger J. Plasma kinins increase after angiotensin-converting enzyme inhibition in human subjects. Clin Sci (Lond) 87: 567-574, 1994.

92
20. Petho G, Derow A, and Reeh PW. Bradykinin-induced nociceptor sensitization to heat is mediated by cyclooxygenase products in isolated rat skin. Eur J Neurosci 14: 210-218, 2001.
21. Ray CA and Gracey KH. Augmentation of exercise-induced muscle sympathetic nerve activity during muscle heating. J Appl Physiol 82: 1719-1725, 1997.
22. Ray CA, Hume KM, Gracey KH, and Mahoney ET. Muscle cooling delays activation of the muscle metaboreflex in humans. Am J Physiol Heart Circ Physiol 273: H2436-2441, 1997.
23. Rotto DM, Hill JM, Schultz HD, and Kaufman MP. Cyclooxygenase blockade attenuates responses of group IV muscle afferents to static contraction. Am J Physiol Heart Circ Physiol 259: H745-750, 1990.
24. Rotto DM, Schultz HD, Longhurst JC, and Kaufman MP. Sensitization of group III muscle afferents to static contraction by arachidonic acid. J Appl Physiol 68: 861-867, 1990.
25. Sinoway LI and Li J. A perspective on the muscle reflex: implications for congestive heart failure. J Appl Physiol 99: 5-22, 2005.
26. Stebbins CL and Longhurst JC. Bradykinin in reflex cardiovascular responses to static muscular contraction. J Appl Physiol 61: 271-279, 1986.
27. Symons JD and Stebbins CL. Effects of angiotensin II receptor blockade during exercise: comparison of losartan and saralasin. J Cardiovasc Pharmacol 28: 223-231, 1996.
28. Veeravalli KK and Akula A. Involvement of nitric oxide and prostaglandin pathways in the cardioprotective actions of bradykinin in rats with experimental myocardial infarction. Pharmacol Res 49: 23-29, 2004.
29. Victor RG, Secher NH, Lyson T, and Mitchell JH. Central command increases muscle sympathetic nerve activity during intense intermittent isometric exercise in humans. Circ Res 76: 127-131, 1995.

93
30. Warren JH, Lewis W, Wraa CE, and Stebbins CL. Central and peripheral effects of angiotensin II on the cardiovascular response to exercise. J Cardiovasc Pharmacol 38: 693-705, 2001.

94
Table 4.1. Baseline measurements during the drug trials during normothermia and forearm heating (n=8)
Ace-Inhibitor Trial
Variable Normothermia Heating
Muscle Temperature (ºC) 34.7 ± 0.3 37.9 ± 0.3†
Tympanic Temperature (ºC) 36.9 ± 0.2 36.9 ± 0.2
Skin Temperature (ºC) 32.6 ± 0.4 39.9 ± 0.2†
MAP (mmHg) 73 ± 3* 72 ± 2*
Heart Rate (beats/min) 66 ± 3 68 ± 4
MSNA (bursts/30 s) 3 ± 1 3 ± 1
MSNA (a.u./30 s) 375 ± 126 285 ± 74
Renal Blood Velocity (cm/s) 58.9 ± 9.1 55.0 ± 6.4
RVC (cm/s/mmHg) 0.79 ± 0.11 0.76 ± 0.07
Calf Blood Flow (ml/min/dl) 2.6 ± 0.4 2.5 ± 0.4
CVC (ml/min/dl/mmHg) 0.36 ± 0.02 0.36 ± 0.02
Blood pH 7.39 ± 0.01 7.39 ± 0.01
Blood Na+ (mmol/L) 139.2 ± 0.7* 138.5 ± 1.2
Blood K+ (mmol/L) 3.7 ± 0.1 3.8 ± 0.1
Blood Lactate (mmol/L) 0.85 ± 0.1 0.79 ± 0.1
MAP, mean arterial pressure; MSNA, muscle sympathetic nerve activity; RVC, renal vascular conductance; CVC, calf vascular conductance. *Significantly different from placebo trials (P < 0.05). †Significantly different from normothermia. Values are mean ± S.E.

95
Table 4.2. Baseline measurements during the placebo trials during normothermia and forearm heating (n=8)
Placebo Trial
Variable Normothermia Heating
Muscle Temperature (ºC) 35.1 ± 0.3 37.9 ± 0.4†
Tympanic Temperature (ºC) 36.9 ± 0.2 37.0 ± 0.2
Skin Temperature (ºC) 32.9 ± 0.5 40.2 ± 0.6†
MAP (mmHg) 81 ± 3* 83 ± 6*
Heart Rate (beats/min) 66 ± 3 68 ± 3
MSNA (bursts/30 s) 3 ± 1 3 ± 1
MSNA (a.u./30 s) 254 ± 125 327 ± 107
Renal Blood Velocity (cm/s) 56.7 ± 7.8 57.4 ± 5.7
RVC (cm/s/mmHg) 0.72 ± 0.12 0.72 ± 0.10
Calf Blood Flow (ml/min/dl) 2.5 ± 0.3 2.9 ± 0.4
CVC (ml/min/dl/mmHg) 0.30 ± 0.01 0.35 ± 0.01
Blood pH 7.41 ± 0.03 7.39 ± 0.01
Blood Na+ (mmol/L) 136.8 ± 1.2 137.7 ± 0.9
Blood K+ (mmol/L) 4.1 ± 0.3 3.6 ± 0.2
Blood Lactate (mmol/L) 0.9 ± 0.1 0.8 ± 0.2
MAP, mean arterial pressure; MSNA, muscle sympathetic nerve activity; RVC, renal vascular conductance; CVC, calf vascular conductance. *Significantly different from drug trials (P < 0.05). †Significantly different from normothermia. Values are mean ± S.E.

96
Figure 4.1. Changes in mean arterial blood pressure (MAP) during exercise and postexercise muscle ischemia (PEMI) during quinapril and placebo treatments with the forearm normothermic or hyperthermic. ACE-inhibition did not alter MAP responses during normothermic handgrip or PEMI. Heating significantly increased MAP during exercise during both treatments. Heat-induced increases in MAP were not different between drug treatments. *Significantly different from normothermia (P < 0.05). Values are mean ± S.E.

97
Quinapril
BL 20 40 60 80 100
-60
-50
-40
-30
-20
-10
0
Normothermia
Heattemp p < 0.05
Time to Fatigue (%)
!
RV
C (
%)
Placebo
BL 20 40 60 80 100
temp x time p < 0.05
Time to Fatigue (%)
-60
-50
-40
-30
-20
-10
0
Normothermia
Heat
PEMI
!
RV
C (
%)
PEMI
Figure 4.2. Changes in renal vascular conductance (RVC) during exercise and postexercise muscle ischemia (PEMI) during quinapril and placebo treatments with the forearm normothermic or hyperthermic. ACE-inhibition did not alter RVC responses during normothermic handgrip or PEMI. Heating significantly decreased RVC from normothermia during exercise in both drug treatments. Heat-induced decreases in RVC were not different between drug treatments. *Significantly different from normothermia (P < 0.05). Values are mean ± S.E.

98
Quinapril
BL 20 40 60 80 100
-50
-25
0
25
50
Normothermia
Heat
Time to Fatigue (%)
!
CV
C (
%)
Placebo
BL 20 40 60 80 100Time to Fatigue (%)
-60
-50
-40
-30
-20
-10
0
Normothermia
Heat
PEMI
!
CV
C (
%)
PEMI
Figure 4.3. Changes in calf vascular conductance (CVC) during exercise and postexercise muscle ischemia (PEMI) for drug and placebo treatments with the forearm normothermic or heated. ACE-inhibitor treatment did not alter CVC responses handgrip or PEMI at either temperature. Values are mean ± S.E.

99
Quinapril
BL 20 40 60 80 100
0
5
10
15
20
25Normothermia
Heat
temp x time p < 0.05
Time to Fatigue (%)
Bu
rsts
(p
er
30s)
Placebo
BL 20 40 60 80 100
temp x time p < 0.05
Time to Fatigue (%)
0
5
10
15
20
25Normothermia
Heat *
PEMI
Bu
rsts
(p
er
30s)
Bursts Placebo
PEMI
Figure 4.4. Changes in muscle sympathetic nerve activity burst frequency during exercise and postexercise muscle ischemia (PEMI) during quinapril and placebo treatments with the forearm normothermic or hyperthermic. ACE-inhibition did not alter muscle sympathetic nerve activity burst frequency responses during normothermic handgrip or PEMI. Heating significantly increased burst frequency during exercise for both treatments and during PEMI in the drug trial. Heat-induced increases in burst frequency were not different between drug treatments. *Significantly different from normothermia (P < 0.05). Values are mean ± S.E.

100
Quinapril
BL 20 40 60 80 100
0
1000
2000
3000
4000
5000Normothermia
Heat
temp x time p < 0.05
Time to Fatigue (%)
To
tal
Ac
tiv
ity
(a
.u.)
Total Activity Placebo
BL 20 40 60 80 100
temp x time p < 0.05
Time to Fatigue (%)
0
1000
2000
3000
4000
5000Normothermia
Heat*
PEMI
To
tal
Ac
tiv
ty (
a.u
.) *
PEMI
Figure 4.5. Changes in total muscle sympathetic nerve activity during exercise and postexercise muscle ischemia (PEMI) during quinapril and placebo treatments with the forearm normothermic or hyperthermic. ACE-inhibition did not alter total activity during normothermic handgrip or PEMI. Heating significantly increased total activity during exercise for both treatments and during PEMI. Heat-induced increases in total activity were not different between drug treatments. *Significantly different from normothermia (P < 0.05). Values are mean ± S.E.

101
Chapter 5
Conclusion
5.1 Introduction
The three main goals of this project were: 1) to examine the effect of changing
forearm muscle temperature on renal blood flow during isometric handgrip, 2) to
examine the interactive effect of aging and forearm muscle heating on
cardiovascular, renal vascular and muscle sympathetic nerve activity (MSNA)
responses to isometric handgrip, and 3) to determine if ACE-inhibition alters
cardiovascular and MSNA responses to isometric handgrip with normothermic
and hyperthermic forearm muscle. The following hypotheses were tested and
these observations were made.
Hypothesis 1: Forearm heating would augment renal vasoconstriction during
isometric handgrip.
Observation 1: Forearm heating increased renal vasoconstriction during
isometric handgrip. This response might have been mediated by an increase in
muscle mechanoreceptor sensitivity.
Hypothesis 2: Forearm cooling would delay renal vasoconstriction during
isometric handgrip.
Observation 2: Forearm cooling decreased renal vasoconstriction during
isometric handgrip. This response might have been mediated by a delay in
activation of the muscle metaboreflex.
Hypothesis 3: The interaction of forearm muscle heating and aging would not
alter MSNA responses to isometric handgrip.

102
Observation 3: The combined interaction of muscle heating and aging did
not alter MSNA responses to isometric handgrip.
Hypothesis 4: The interaction of aging and local muscle heating would augment
renal vasoconstriction during isometric handgrip.
Observation 4: The combined interaction of aging and local muscle
heating increased renal vasoconstriction during isometric handgrip.
Hypothesis 5: ACE-inhibition would augment cardiovascular and MSNA
responses to isometric handgrip with a normothermic forearm.
Observation 5: ACE-inhibition did not alter cardiovascular and MSNA
function during isometric handgrip with a normothermic forearm.
Hypothesis 6: ACE-inhibition would augment cardiovascular and MSNA
responses to isometric handgrip with a hyperthermic forearm.
Observation 6: ACE-inhibition did not alter cardiovascular and MSNA
responses to isometric handgrip with a hyperthermic forearm.
5.2 Significance of Current Findings
The three studies in this project provide important insight on the
relationship between exercise and muscle temperature, renal blood flow, aging
and ACE-inhibition. This next section will review what insights can be gained
from the three studies.
Study 1. During isometric handgrip, forearm heating and cooling increase and
decrease renal blood flow compared to normothermia, respectively. These
thermal-induced changes in muscle afferent sensitivity might be a mechanism
that the body uses to control blood flow during exercise and thermal stress.

103
During periods of exercise in the heat, the greater renal vasoconstriction might
be a mechanism that increases blood flow to thermoregulatory and working
tissues or to help maintain blood pressure in the face of decreasesd total
peripheral resistance. During periods of exercise in the cold, a delay in activation
of the exercise pressor reflex caused by muscle cooling might blunt increases in
peripheral blood flow to attenuate heat loss to the environment.
Study 2. Compared to young individuals, older individuals who perform upright
dynamic lower body exercise in the heat produce less renal vasoconstriction and
skin vasodilation (28). In contrast, greater renal vasoconstriction occurred in
older individuals during isometric exercise with heated muscles compared to the
young. The importance of this finding is that it suggests that the pattern of renal
blood flow control during thermal stress in older individuals is different during
dynamic upright exercise with a large muscle mass compared to isometric
exercise with a small muscle mass, or that the thermal load during whole-body
heating in older individuals alters neural control of blood flow during exercise
differently than in the young. The age-related change in neural control of
circulation that causes this difference remains equivocal but could provide
important insight into why aging is associated with an increased prevalence of
thermal-related illnesses.
Study 2 provides insight into the possible mechanism that causes heat-
induced changes in muscle afferent sensitivity. As skeletal muscle ages it
becomes more oxidative (4, 24, 45), which causes a decrease in production of
metabolites known to sensitize muscle mechanoreceptors, including hydrogen

104
ions and lactate (49). If local metabolites influence muscle afferent sensitivity in
the heat, then the decreased production of metabolites in the older individuals
would have attenuated MSNA responses to handgrip with hyperthermic muscles.
This was not the case. Therefore metabolic by-products of exercise do not
contribute to heat-induced changes in muscle afferent sensitivity.
Study 3. ACE-inhibitors are used to treatcardiovascular disorders including
hypertension, congestive heart failure, and left ventricular dysfunction. The
exercise pressor reflex is might be exaggerated in congestive heart failure
patients because of changes in muscle afferent sensitivity (10, 70). The results
of this study suggests that acute use of an ACE-inhibitor, and subsequent
alterations in the renin-angiotensin and kallikrein-kinin systems, might not alter
cardiovascular and MSNA responses to isometric exercise in healthy subjects.
The results of the third study also help elucidate possible mechanisms
contributing to the increased sensitivity of the muscle afferents in the heat. The
results of the study suggest that, unlike the skin (51, 69), kinins might not
contribute to increased muscle afferent sensitivity in the heat. Furthermore, in
agreement with the second study, changes in local metabolites do not appear to
have a role in increasing sensitivity of the muscle afferents in the heat.
5.3 Perspectives and Future Studies
The goal of the current work was to examine the effect of local muscle
temperature on muscle afferent control of the exercise pressor reflex and
possible influences of aging and ACE-inhibition. The current studies answer
several questions, but created more. To better understand the collective

105
significance of the current studies in relation to each other, reviewing these
questions will be helpful.
For all studies in the current project, only a small muscle mass was
activated by isometric contraction. Cardiovascular and MSNA responses to
activation of the exercise pressor reflex at normal muscle temperatures are
influenced by the size of muscle mass activated (84). It is not apparent how
increasing the amount of muscle mass activated, or which particular muscle beds
are activated, influence cardiovascular and MSNA responses to isometric
exercise with hypo- and hyperthermic muscle. By performing isometric handgrip
with both forearms heated, we might better understand how muscle mass might
influence cardiovascular and MSNA responses to exercise. Furthermore, using a
model with dynamic contractions might be beneficial to understanding how mode
of contraction type might alter cardiovascular and MSNA responses to exercise
and changes in muscle temperature.
Endurance training alters muscle afferent control of the exercise pressor
reflex. For example, training decreases MSNA responses in the forearm and leg
during exercise and postexercise muscle ischemia (71, 85, 87, 89). The current
studies used untrained individuals. It is not apparent how exercise training would
influence muscle afferent sensitivity to thermal stress. Furthermore, endurance
training can have a marked influence on cardiovascular and sympathetic control
during exercise in older individuals (28, 36) and the possible influences of aging
and exercise training on muscle afferent sensitivity to thermal stress remains
equivocal.

106
Exposure to a hot environment at rest and exercise in a normothermic
environment both induce heat acclimatization. Following heat acclimatization,
heart rate is lower during exercise (83). It is not known if muscle afferent control
of the exercise pressor reflex might be altered by heat acclimatization.
Furthermore, the effects of cold acclimatization on muscle afferent contributions
to the exercise pressor reflex is unknown.
The current studies only examined the question of aging on cardiovascular
and MSNA responses to isomeric exercise with a hyperthermic forearm. Aging is
associated with changes in cardiovascular responses to the cold, including
attenuation of vasoconstrictor response in the skin in the cold (13). The influence
of muscle hypothermia and aging on cardiovascular and MSNA responses to
isometric exercise is unclear.
The results of the second study, where aging and heating increased renal
vasoconstriction during isometric exercise, is in agreement with Momen et al.
(57) who found that older individuals vasoconstrict the renal vasculature more
than younger individuals during normothermic exercise. The reasons for the
different renal vascular responses to isometric exercise between young and older
individuals remains equivocal. In the forearm and leg vasculature, aging is
associated with a reduction in α1-adrenergic responsiveness (15, 88) along with
increases in basal MSNA (16, 63, 74, 93). The results of the current aging study,
and Momen et al. (57), suggests that aging might be associated with changes in
either sympathetic outflow to the kidney or renal vascular responsiveness that
contrast with peripheral vascular and MSNA responses.

107
A major shortcoming of this study is that we examined the effects of ACE-
inhibition treatment in normal healthy individuals; however, ACE-inhibitors are
commonly used in older patients. Likewise, the study only examined the effect
of acute ACE-inhibition, whereas ACE-inhibitor treatment is usually long-term.
Future studies should examine if both acute ACE-inhibition and long-term
treatment with an ACE-inhibitor alter cardiovascular and MSNA responses to
exercise in older individuals.
ACE-inhibition alters blood angiotensin II and kinin levels (68). During
exercise, bradykinin is released from working muscles in both animals and
humans (5, 6, 43, 90). Whether or not an oral dose of ACE-inhibitor would
influence local muscle angiontensin II and kinins remains equivocal. Future,
studies should examine the influences of directly altering the renin-angiotensin
and kallikrein-kinin systems in muscle tissue on cardiovascular and MSNA
responses to exercise with hyperthermic muscles.
In the last study we tried to manipulate and measure factors that increase
muscle afferent sensitivity. The results of the study suggest that the kallikrein-
kinin and renin-angiotensin systems and local metabolism do not influence
muscle afferent sensitivity in the heat. What contributes to the increase in
muscle afferent sensitivity during both hyperthermia and hypothermia is not
known. However, possible-contributing factors could include prostaglandins,
heat and cold induced changes in chemical reaction kinetics and ion movement
in the tissues, and/or central command. Future studies are needed to elucidate

108
which of these factors, or combination of factors, contribute to altered muscle
afferent sensitivity during thermal stress.
5.4 Summary
The combination of thermal stress and exercise severely challenges the
cardiovascular system. The cardiovascular system must work simultaneously to
control body temperature and meet the metabolic demands of working muscle.
The results of the current work indicate that changes in muscle temperature alter
neural control of circulation to help meet the dual demand placed on the
cardiovascular system. As a result of this study we better understand the
following: 1) how exercising muscle temperature can influence renal circulation,
2) how aging and local muscle temperature interactively influence the exercise
pressor reflex, and 3) possible influences of short-term modulation of the renin-
angiotensin and the kallikrein-kinin system on cardiovascular and MSNA
responses to isometric exercise at normothermic and hyperthermic muscle
temperatures.

109
5.5 References 1. Bilodeau M, Henderson TK, Nolta BE, Pursley PJ, and Sandfort GL. Effect of aging on fatigue characteristics of elbow flexor muscles during sustained submaximal contraction. J Appl Physiol 91: 2654-2664, 2001.
2. Blais C, Jr., Adam A, Massicotte D, and Peronnet F. Increase in blood bradykinin concentration after eccentric weight-training exercise in men. J Appl Physiol 87: 1197-1201, 1999.
3. Boix F, Roe C, Rosenborg L, and Knardahl S. Kinin peptides in human trapezius muscle during sustained isometric contraction and their relation to pain. J Appl Physiol 98: 534-540, 2005.
4. Coats AJ, Clark AL, Piepoli M, Volterrani M, and Poole-Wilson PA. Symptoms and quality of life in heart failure: the muscle hypothesis. Br Heart J 72: S36-39, 1994.
5. Degroot DW and Kenney WL. Impaired defense of core temperature in aged humans during mild cold stress. Am J Physiol Regul Integr Comp Physiol 292: R103-108, 2007.
6. Dinenno FA, Dietz NM, and Joyner MJ. Aging and forearm postjunctional alpha-adrenergic vasoconstriction in healthy men. Circulation 106: 1349-1354, 2002.
7. Dinenno FA and Joyner MJ. Alpha-adrenergic control of skeletal muscle circulation at rest and during exercise in aging humans. Microcirculation 13: 329-341, 2006.
8. Hatakenaka M, Ueda M, Ishigami K, Otsuka M, and Masuda K. Effects of aging on muscle T2 relaxation time: difference between fast- and slow-twitch muscles. Invest Radiol 36: 692-698, 2001.
9. Ho CW, Beard JL, Farrell PA, Minson CT, and Kenney WL. Age, fitness, and regional blood flow during exercise in the heat. J Appl Physiol 82: 1126-1135, 1997.

110
10. Kenney WL and Munce TA. Invited review: aging and human temperature regulation. J Appl Physiol 95: 2598-2603, 2003.
11. Langberg H, Bjorn C, Boushel R, Hellsten Y, and Kjaer M. Exercise-induced increase in interstitial bradykinin and adenosine concentrations in skeletal muscle and peritendinous tissue in humans. J Physiol 542: 977-983, 2002.
12. Lee WS, Cheung WH, Qin L, Tang N, and Leung KS. Age-associated decrease of type IIA/B human skeletal muscle fibers. Clin Orthop Relat Res 450: 231-237, 2006.
13. Markel TA, Daley JC, 3rd, Hogeman CS, Herr MD, Khan MH, Gray KS, Kunselman AR, and Sinoway LI. Aging and the exercise pressor reflex in humans. Circulation 107: 675-678, 2003.
14. Mayer S, Izydorczyk I, Reeh PW, and Grubb BD. Bradykinin-induced nociceptor sensitisation to heat depends on cox-1 and cox-2 in isolated rat skin. Pain, 2006.
15. Momen A, Leuenberger UA, Handly B, and Sinoway LI. Effect of aging on renal blood flow velocity during static exercise. Am J Physiol Heart Circ Physiol 287: H735-740, 2004.
16. Ng AV, Callister R, Johnson DG, and Seals DR. Age and gender influence muscle sympathetic nerve activity at rest in healthy humans. Hypertension 21: 498-503, 1993.
17. Pellacani A, Brunner HR, and Nussberger J. Plasma kinins increase after angiotensin-converting enzyme inhibition in human subjects. Clin Sci (Lond) 87: 567-574, 1994.
18. Petho G, Derow A, and Reeh PW. Bradykinin-induced nociceptor sensitization to heat is mediated by cyclooxygenase products in isolated rat skin. Eur J Neurosci 14: 210-218, 2001.
19. Piepoli M, Ponikowski P, Clark AL, Banasiak W, Capucci A, and Coats AJ. A neural link to explain the "muscle hypothesis" of exercise intolerance in chronic heart failure. Am Heart J 137: 1050-1056, 1999.

111
20. Ray CA. Sympathetic adaptations to one-legged training. J Appl Physiol 86: 1583-1587, 1999.
21. Ray CA and Monahan KD. Aging attenuates the vestibulosympathetic reflex in humans. Circulation 105: 956-961, 2002.
22. Sawka NM and Young AJ. Physiological Systems and Their Responses to Conditions of Heat and Cold. In: ACSM's Advanced Exercise Physiology, edited by Tipton MJ. Baltimore, MD: Lippincott Williams & Wilkins, 2006, p. 535-563.
23. Seals DR. Influence of active muscle size on sympathetic nerve discharge during isometric contractions in humans. J Appl Physiol 75: 1426-1431, 1993.
24. Sinoway L, Shenberger J, Leaman G, Zelis R, Gray K, Baily R, and Leuenberger U. Forearm training attenuates sympathetic responses to prolonged rhythmic forearm exercise. J Appl Physiol 81: 1778-1784, 1996.
25. Sinoway LI, Rea RF, Mosher TJ, Smith MB, and Mark AL. Hydrogen ion concentration is not the sole determinant of muscle metaboreceptor responses in humans. J Clin Invest 89: 1875-1884, 1992.
26. Smith EG, Voyles WF, Kirby BS, Markwald RR, and Dinenno FA. Ageing and leg postjunctional alpha-adrenergic vasoconstrictor responsiveness in healthy men. J Physiol 582: 63-71, 2007.
27. Somers VK, Leo KC, Shields R, Clary M, and Mark AL. Forearm endurance training attenuates sympathetic nerve response to isometric handgrip in normal humans. J Appl Physiol 72: 1039-1043, 1992.
28. Stebbins CL, Carretero OA, Mindroiu T, and Longhurst JC. Bradykinin release from contracting skeletal muscle of the cat. J Appl Physiol 69: 1225-1230, 1990.
29. Sundlof G and Wallin BG. Human muscle nerve sympathetic activity at rest. Relationship to blood pressure and age. J Physiol 274: 621-637, 1978.

112
Appendix A
Subject Consent Form
CONSENT FOR RESEARCH Penn State College of Medicine
The Milton S. Hershey Medical Center Title of Project: The influence of aging on muscle sympathetic nerve activity during heating and cooling of exercising muscle. Principal Investigator: Chester A. Ray, Ph.D. Other Investigators: Kevin Monahan, Ph.D., Nathan Kuipers, M.S., Charity Sauder, M.S., Amy Fogelman, M.S., Damian Dyckman, B.S., Matthew Kearney, B.S., Erin Muldoon, B.S. Participant’s Printed Name: _____________________________ This is a research study. Research studies include only people who voluntarily choose to take part. This consent form gives you information about this research, which will be discussed with you. This consent form may contain words or procedures that you do not understand. You are urged to ask questions about anything that is unclear to you. Discuss it with your family and friends and take your time to make your decision. You will receive a copy of the signed and dated consent form to keep. 1. Purpose of the Research:
You are being offered the opportunity to take part in this research because you are a healthy volunteer.
The purpose of this study is to investigate how aging influences changes in blood pressure and heart rate during exercise at different muscle temperatures.
Approximately 60 people will take part in this research at the Hershey Medical Center.
2. Procedures to be Followed:
You will be asked to come into the laboratory for one 3-4 hour visit during which:
a. You will be given a medical history and physical examination by a physician.

113
b. If you are a woman of childbearing potential, a urine pregnancy test will be performed to insure that you are not pregnant. If the pregnancy test is positive, you will be excluded from participation in the study.
c. Patch electrodes (adhesive disks the size of a half dollar) will be placed on
your chest and will be used to monitor your heart rate while a small cuff on your finger will measure your blood pressure.
d. A small, sterilized needle will be inserted into the muscle in the forearm that
you will be exercising with. This needle is connected to a probe that measures the temperature of your muscle.
e. You will undergo a procedure called microneurography. The investigators
will look for your nerve by touching a pen-like device that gives off a small electrical stimulation to your skin. This will make your foot twitch. The electrical stimulation will last for about 5 minutes. Then, two very fine, sterilized nerve wires (microelectrodes), thinner than a sewing needle, will be introduced through the skin, one of which is inserted further into a superficial nerve behind your knee. This procedure is used to measure nerve activity from the brain to the leg. Locating the nerve with the microelectrodes will take a maximum of 1 hour.
f. A Velcro belt will be placed around your chest to measure your breathing.
This device is called a pneumotrace.
g. You will have blood flow measured in your arms and the arteries in your abdomen using Doppler ultrasound.
h. A small flexible plastic thermometer will be placed under your tongue to
measure your body temperature. Six small flexible thermometers will be taped to the skin on your arm, back, shoulder, side, stomach, thigh, and lower leg.
i. You will perform bouts of exercise while your arms are either heated, cooled
or remains at normal temperature. To heat and cool your arms you will wear special sleeves that are designed to allow water at different temperatures to circulate through them.
j. You will perform handgrip exercise at a percentage of your maximal gripping
capacity. You will perform this exercise until your arm fatigues and you cannot grip any more or the investigator tells you to stop. You will perform handgrip exercise with a blood pressure cuff inflated over your arm blocking blood flow to your arm. The blood pressure cuff will be inflated either during or after handgrip exercise.
k. You will be asked to place your hand in ice water.

114
l. A venous catheter (plastic tube) will be placed into your forearm to draw
blood and measure chemicals released by the muscles before, during, and after you exercise. The amount of blood drawn will be equal to six to seven tablespoons or 3 ounces. The chemicals that may be looked at include: hydrogen, lactate, electrolytes, prostaglandins, ATP, phosphate, blood gases, glucose, bradykinin, histamine, serotonin and substance P.
3. Discomforts and Risks:
Allergic reactions to adhesive patches, such as redness and rash, are minimal and are an unusual occurrence. There are no known risks of placing the Velcro belt around your chest, or the Finapres cuff on your finger. You may feel a slight pinch or pin prick when the sterile needle that measures your muscle temperature enters your forearm. Less commonly, a small blood clot will occur at the puncture site. This should feel similar to the feelings felt when getting a shot at the doctor's office. The placing of a microelectrode into the leg nerve is accompanied by a less than 10% chance of aching and/or pins and needles sensation in the area of the electrode insertion. These sensations may last for a few days. There is a remote chance of nerve soreness that may result in persistent or permanent sensations associated with microneurography tests. However, the investigators have done thousands of studies without permanent complications. Other risks include local infection or bleeding. There is a very small risk of skin burns from the ultrasound Doppler used to measure blood flow of the brachial arteries, which can be avoided by keeping the sound wave intensity at low levels. If you report warmth or discomfort, the ultrasound Doppler intensity will be readjusted downwards and/or moved to a different location. Handgrip exercise may be tiring, but this sensation will diminish rapidly once the exercise is finished. Blocking blood flow to the forearm with the blood pressure cuff may cause a pins and needles sensation in the arm and discomfort where the cuff is inflated. Deflating the cuff causes a warm rush (of blood) for approximately one minute. Heating or cooling of your muscles might cause slight discomfort. The temperatures your muscles will be heated to are those you would experience

115
during exercise. The temperature your muscles will be cooled to are those you would experience when using ice on an injury. We will use special thermometers to monitor your muscle and skin temperatures so that they do not get too hot or cold. It is important that a fetus (developing unborn baby) not be exposed to any unnecessary risks. If you are a female capable of becoming pregnant, you must not be pregnant at the beginning of this investigation. A pregnancy test will be required before you begin the research. Placing your hand in ice water will cause discomfort and pain but there are no major risks. The discomfort associated with removing blood by venipuncture (by needle from a vein) is a slight pinch or pin prick when the sterile needle enters the skin. The risks include mild discomfort and/or a black and blue mark at the site of puncture. Less common risks include a small blood clot, infection or bleeding at the puncture site, and on rare occasions fainting during the procedure.
4. Possible Benefits:
a. Possible benefits to the participant: You will not benefit from taking part in this research study.
b. Possible benefits to others: The potential benefit to others is better understanding of how temperature and aging influence muscle reflexes that control heart rate and blood pressure responses during exercise.
5. Other Options that Could be Used Instead of this Research:
You do not have to take part in this research study. 6. Time Duration of the Procedures and Study:
The study will last approximately 3-4 hours.
7. Statement of Confidentiality:
a. Privacy and Confidentiality Measures Your research records that are reviewed, stored, and analyzed at The Milton S. Hershey Medical Center (HMC) and Penn State College of Medicine (PSU) will be labeled with your initials and the date that your study was performed. The list that matches your name with the initials will be kept in a locked file in Dr. Ray’s office. The research records will be kept on a password protected computer in the PI’s laboratory (rooms C4520 or C4523.

116
Your samples collected for research purposes will be labeled with a letter code and will be stored and secured in a freezer in the General Clinical Research Center
In the event of any publication or presentation resulting from the research, no personally identifiable information will be shared.
b. The Use of Private Health Information: Health information about you will be collected if you choose to be part of this research study. Health information is protected by law as explained in the HMC Privacy Notice. If you have not received this notice, please request a copy from the researcher. At The Milton S. Hershey Medical Center (HMC) and Penn State College of Medicine (PSU) your information will only be used or shared as explained and authorized in this consent form or when required by law. It is possible that some of the other people/groups who receive your health information may not be required by Federal privacy laws to protect your information and may share it without your permission.
To participate in this research you must allow the research team to use your health information. If you do not want us to use your protected health information, you may not participate in this research. Your permission for the use, retention, and sharing of your identifiable health information will continue indefinitely. Should there be a time that the information is no longer needed the research information not already in your medical record will be destroyed. Any research information in your medical record will be kept indefinitely. If you choose to participate, you are free to withdraw your permission for the use and sharing of your health information at any time. You must do this in writing as indicated in the HMC Privacy Notice. Write to Dr. Ray and let him know that you are withdrawing from the research study. His mailing address is Penn State College of Medicine, Division of Cardiology, H047, 500 University Dr., Hershey, PA 17033.
If you withdraw your permission:
• We will no longer use or share medical information about you for the reasons covered by your written authorization, except when the law allows us to do so.
• We are unable to take back anything we have already done or any information we have already shared with your permission.
• We may continue using and sharing the information obtained prior to your withdrawal if it is necessary for the soundness of the overall research.
• We will need to keep our records of the care that we provided to you as long as the law requires.

117
The research team may use the following sources of health information.
• All data collected during the testing period including: ECG tracings, medical history, blood pressure measurements, muscle temperatures, exercise workloads, microneurography data, ultrasound data.
Representatives of the following people/groups within HMC/PSU are allowed to use your health information and to share it with other specific groups in connection with this research study.
• The principal investigator, Chester A. Ray, Ph.D. • The HMC/PSU Institutional Review Board • The HMC/PSU Human Subjects Protection Office • The research team • The study coordinators
The people or groups listed in the above paragraph may share your health information with the following people/groups outside HMC/PSU for their use in connection with this research study. These groups, while monitoring the research study, may also review and/or copy your original PSU/HMC records.
• The Office of Human Research Protections in the U. S. Department of Health and Human Services
• The National Institutes of Health
8a. Costs for Participation: a. Costs: There will be no added cost to you for participating in this research study b. Treatment and Compensation for Injury: Every effort to prevent injury as a result of your participation will be taken. It is possible, however, that you could develop complications or injuries as a result of participating in this research study. In the event of injury resulting from this research, medical treatment is available but will be provided at the usual charge. It is the policy of this institution to provide neither financial compensation nor free medical treatment for research-related injury.
Costs for the treatment of research-related injuries will be charged to your insurance carrier or to you. Some insurance companies may not cover costs associated with research studies. If for any reason these costs are not covered by your insurance, they will be your responsibility.
You are not waiving any legal rights you may have by signing this form.
9. Compensation for Participation:
You will receive $100 for completion of the entire study. If you do not complete the entire study, you will receive $25 per hour of your time.

118
10. Research Funding:
The institution and investigators are receiving a grant from The National Institutes of Health to support the activities that are required to conduct this research.
11. Voluntary Participation: Taking part in this research study is voluntary. If you choose to take part in this research, your major responsibilities will include: following the instructions of the investigators during the collection of data. You do not have to participate in this research. If you choose to take part, you have the right to stop at any time. If you decide not to participate or if you decide to stop taking part in the research at a later date, there will be no penalty or loss of benefits to which you are entitled. In other words, your decision to decline to participate in this research or to stop taking part in the research will not affect your medical care and academic standing and job status. Your research doctor or the sponsor may take you out of the research study without your permission. Possible reasons for this are: You do not adhere to the investigator’s instructions or follow the protocol as stated in this consent form. Also, the sponsor of the research may end the research study early. If your participation in the research ends early, you may be asked to visit the research doctor for a final visit.
If you will be participating in another clinical trial at Hershey Medical Center or elsewhere while in this research, you should discuss the procedures and/or treatments with your physician or the investigators. This precaution is intended to protect you from possible side effects from interactions of research drugs, treatments or testing.
12. Contact Information for Questions or Concerns: You have the right to ask any questions you may have about this research. If you have questions or concerns or believe you may have developed an injury that is related to this research, contact Dr. Ray at 717-531-3906 or you can page the Cardiology doctor on 24-hour call at 717-531-8521. If you have questions, complaints, or concerns regarding your rights as a research participant or you have general questions about the research or about your privacy and the use of your personal health information, you may contact the research protection advocate in the HMC Human Subjects Protection Office at 717-531-5687. You may also call this number if you cannot reach the research team and wish to talk to someone else.
For more information about participation in a research study and about the Institutional Review Board (IRB), a group of people who review the research to protect your rights, please visit the HMC IRB’s Web site at http://www.hmc.psu.edu/irb. Included on this web site, under the heading

119
“Participant Info”, you can access the federal regulations and information about the protection of human research participants. If you do not have access to the internet, copies of these federal regulations are available by calling the HSPO at (717) 531-5687.
Signature and Consent/Permission to be in the Research Before making the decision regarding enrollment in this research you should have:
• Discussed this study with an investigator, • Reviewed the information in this form, and • Had the opportunity to ask any questions you may have.
Your signature below means that you have received this information, have asked the questions you currently have about the research and those questions have been answered. You will receive a copy of the signed and dated form to keep for future reference. Participant: By signing this consent form, you indicate that you are voluntarily choosing to take part in this research. ___________________________ __________ ______ ________ Signature of Participant Date Time Printed Name
Person Explaining the Research: Your signature below means that you have explained the research to the participant/participant representative and have answered any questions he/she has about the research. ______________________________ ________ _____ Signature of person who explained this research* Date Time ______________________________ Printed Name (*Only approved investigators for this research may explain the research and obtain informed consent.)

120
Appendix B
Subject Consent Form
CONSENT FOR RESEARCH Penn State College of Medicine
The Milton S. Hershey Medical Center
Title of Project: Role of Bradykinin in the Augmentation of Renal and Peripheral Vasoconstriction During Activation of the Exercise Pressor Reflex in Heated Muscle. Principal Investigator: Chester A. Ray, Ph.D. Other Investigators: Urs. A. Leuenberger, Nathan T. Kuipers, Charity L. Sauder, Matthew L. Kearney, Kevin D. Monahan, Damian J. Dyckman Participant’s Printed Name: _____________________________ This is a research study. Research studies include only people who want to take part. This form gives you information about this research, which will be discussed with you. It may contain words or procedures that you don’t understand. Please ask questions about anything that is unclear to you. Discuss it with your family and friends and take your time to make your decision. 1. Purpose of the Research:
You are being offered the opportunity to take part in this research because you are a healthy volunteer. The purpose of this research is to examine if a substance released by your body, called bradykinin, plays a role in increasing blood pressure and decreasing blood flow to your limbs and kidneys when you exercise in the heat. During this study you will be given a dosage of quinapril or a placebo. Quinapril is a drug approved by the United States Food and Drug Administration for managing high blood pressure and treating congestive heart failure.
Approximately 20 people will take part in this research at the Hershey Medical Center.
2. Procedures to be Followed:

121
You will be asked to come into the laboratory for four visits. The visits will be structured as follows: one morning visit and afternoon visit one day and one morning and afternoon visit at least a week later. Before each morning visit, you will be asked not to eat or exercise for 8 hours or to eat and drink anything with caffeine for at least 24 hours before arriving. Because quinapril can cause low blood pressure you should drink plenty of water the night before and prior to your first morning visit. Twice you will come to the lab early in the morning for 60 minutes during which the following procedures will be followed: a. Upon arrival, female participants of childbearing age will submit a urine
sample for a pregnancy test. If the test is positive, you will not be allowed to participate in the study.
b. Medical staff will perform a brief medical history and physical examination
(this will be done on your first morning visit if you do not have a current physical examination from a licensed health care provider associated with the General Clinical Research Center).
c. A small needle will be inserted into a vein in your arm and approximately a
tablespoon of blood will be collected. This blood sample will be used to measure blood bradykinin levels.
d. You will be given a 20 mg quinapril pill or a sugar pill. Which pill you
receive at this visit will be determined based upon what number subject you are in the study and the treatment assigned to that number. The treatment condition associated with your subject number was randomly assigned by a computer. You will have an equal chance of receiving either pill at the first visit. If you receive quinapril at this visit, then at the next morning visit you will receive a sugar pill and vice versa. Neither you nor the research team will know which pill you are receiving, but we will be able to get this information quickly if we need it to ensure your safety.
e. The same breakfast will be provided for you after taking quinapril or
placebo on the other morning. Likewise, you will be given a snack to eat later that morning that will be identical on both testing days. You will be asked to not eat or drink anything but the food provided for you prior to the afternoon visit. You may drink water. You will be asked to not exercise prior to the afternoon visit.
f. During this time your blood pressure will be monitored in case the drug
causes you to suffer low blood pressure. You will remain at the lab for one hour after taking the pill and your blood pressure will be monitored throughout this time period and immediately before you leave. If you suffer from low blood pressure a nurse or doctor may give you an infusion of saline through an IV catheter that will be placed in your arm. You will be

122
encouraged to drink water up to an hour and a half before your afternoon visit to help you from developing low blood pressure.
You will return in the afternoon for a 3-4 hour visit during which the following procedures will be performed: a. You will lay flat on a bed and small adhesive patches will be placed on
your chest to record your heart rate. Cuffs will be placed on your upper arm and finger to measure your blood pressure.
b. A small plastic tube (IV catheter) will be inserted into a deep vein in one of
your arms. The IV will be used to withdraw blood samples before and after exercise. We will draw a total of 8 tablespoons per visit. The chemicals that may be looked at include: hydrogen, lactate, electrolytes, prostaglandins, ATP, phosphate, blood gases, glucose, bradykinin, histamine, serotonin and substance P.
c. A small, sterilized needle will be inserted into the muscle of the exercising
forearm. This needle is a probe that measures the temperature of your muscle.
d. A Velcro belt will be placed around your chest to measure your breathing.
This device is called a pneumotrace.
e. You will undergo a procedure called microneurography. The investigators will look for your nerve by touching a pen-like device that gives off a small electrical stimulation to your skin. This will make your foot twitch. The electrical stimulation will last for about 5 minutes. Then, two very fine, sterilized nerve wires (microelectrodes), thinner than a sewing needle, will be introduced through the skin, one of which is inserted further into a superficial nerve behind or next to your knee. This procedure is used to measure nerve activity from the brain to the leg. Locating the nerve with the microelectrodes will take a maximum of 1 hour.
f. You will have blood flow measured in arteries in your abdomen using
Doppler ultrasound. The Doppler ultrasound technique is similar to the one used to monitor the development of an unborn fetus. A probe six inches long and two inches wide will be placed over your abdomen and held there by an investigator.
g. Blood pressure cuffs will be placed around one thigh and one ankle. The
cuffs will be inflated from time to time to stop blood flow to your ankle and

123
out of your thigh, so that blood flow can be measured by a rubber-band like tube wrapped around your leg.
h. A thermometer will be placed under your tongue or in your ear to measure
your body temperature. Six small flexible thermometers may be taped to the skin on your arm.
i. You will perform bouts of exercise with your arm heated and at normal
temperature. To heat your arm you will wear a special sleeve designed to allow warm water to circulate through it. Your arm will be heated to a temperature that the muscles would experience during exercise on a hot day.
j. You will perform handgrip exercise at a percentage of your maximal gripping
capacity. You will perform this exercise until your arm fatigues and you cannot grip anymore or the investigator tells you to stop. You will perform handgrip exercise with a blood pressure cuff inflated over your arm blocking blood flow to your arm. The blood pressure cuff will be inflated before, during or after handgrip exercise.
k. Blood samples taken during your visits will be analyzed immediately or
stored and analyzed at a later date. Upon completion of the study all samples will destroyed.
l. You will return to the lab for the last two visits at least a week later and the
opposite drug will be given to you from the first trial. All measurements and testing procedures will be the same for the last two visits. Both you and the investigator will not know the drug being administered during each trial. If you experience any side effects during the duration of the study, you should inform one of the investigators or nurses involved with this research and you will receive treatment for the side effects you are experiencing. You will be given a questionnaire after each visit asking if you have had any side-effects from the study, please complete and mail this questionnaire back to the investigators approximately one week after your each afternoon visit.
m. If all measurements could not be taken during your visits you may be
asked to return to repeat those portions of the study.
3. Discomforts and Risks:
Common side effects to quinapril include: low blood pressure, chest pain, dizziness, headache, fatigue, rash, vomiting/nausea, diarrhea, muscle pain, back pain, cough and breathing difficulty. Immediately report to the investigators if you experience any swelling of your face, mouth, lips, tongue, or throat; chest pain or respiratory difficulty; persistent cough; persistent pain

124
in muscles, joints or back; skin rash or other adverse reactions. To help avoid low blood pressure you are encouraged to drink plenty of fluids during the morning sessions and continue up to an hour and a half before your afternoon visit. The placing of a microelectrode into the leg nerve is accompanied by a less than 10% chance of aching and/or pins and needles sensation in the area of the electrode insertion. These sensations may last for a few days. There is a remote chance of nerve soreness that may result in persistent or permanent sensations associated with microneurography tests. However, the investigators have done thousands of studies without permanent complications. Other risks include local infection, bruising, or bleeding.
Allergic reactions to adhesive patches, such as redness and rash, are minimal and are an unusual occurrence. There are no known risks of placing the Velcro belt around your chest, or the blood pressure cuffs on your arms or finger.
The discomfort associated with removing blood by venipuncture (by needle from a vein) is a slight pinch or pin prick when the sterile needle enters the skin. The risks include mild discomfort and/or a black and blue mark at the site of puncture. Less common risks include a small blood clot, infection or bleeding at the puncture site, and on rare occasions fainting during the procedure.
You may feel a slight pinch or pin prick when the sterile needle that measures your muscle temperature enters your forearm. The risks include mild discomfort and/or a black and blue mark at the site of puncture. Less commonly, a small blood clot will occur at the puncture site. This will feel similar to getting a shot at the doctor's office.
There is a very small risk of skin burns from the ultrasound Doppler used to measure blood flow to the renal arteries, which can be avoided by keeping the sound wave intensity at low levels. If you report warmth or discomfort, the ultrasound Doppler intensity will be readjusted downwards and/or moved to a different location. Handgrip exercise may be tiring, but this sensation will diminish rapidly once the exercise is finished. Blocking blood flow to the forearm with the blood pressure cuff may cause a pins and needles sensation in the arm and discomfort where the cuff is inflated. Deflating the cuff causes a warm rush (of blood) for approximately one minute. After exercising, when the blood flow is still stopped to the arm, you will experience a feeling in the muscle similar to what you experience during very hard exercise.

125
Heating your forearm might cause slight discomfort. After the study your arm may be sensitive and red for several hours because of the heating (it may feel like a sunburn), however, this feeling will go away within several hours. The temperatures your muscles will be heated to are those you would experience during exercise. We will use special thermometers to monitor your muscle and skin temperatures so that they do not get too hot. When measuring blood flow in your leg you may feel a “pins and needles” sensation in your foot that will go away when the blood pressure cuffs are turned off. There is no risk of measuring blood flow in the leg using the rubber band like tube. To limit the “pins and needles” sensation, blood flow to the foot will only be stopped for up to 15 min.
It is important that a fetus (developing unborn baby) not be exposed to any unnecessary risks. If you are a female capable of becoming pregnant, you must not be pregnant at the beginning of this investigation. A pregnancy test will be required before you begin the research. There are no risks from measuring blood pressure with the cuff around your finger or arm.
4. Possible Benefits: a. Possible benefits to the participant: You will not benefit from taking part in this research study.
b. Possible benefits to others: The potential benefit to others is better understanding of how temperature influences muscle reflexes that control heart rate and blood pressure responses during exercise.
5. Other Options that Could be Used Instead of this Research:
You do not have to take part in this research study. 6. Time Duration of the Procedures and Study:
If you agree to take part in this study, your involvement will require you to come to the laboratory four times. Two morning visits of 30-60 minute duration and two afternoon visits of 3 -4 hours.
7. Statement of Confidentiality:
a. Privacy and confidentiality measures Your electronic research records that are reviewed, stored, and analyzed at The Milton S. Hershey Medical Center (HMC) and Penn State College of Medicine (PSU) will be labeled with your initials and the date that your study was

126
performed. The research records will be kept on a password protected computer in the PI’s laboratory (rooms C4520 or C4523).
Your blood samples collected for research purposes will be labeled with a non-personal identifying code and will be stored and secured in a freezer in the General Clinical Research Center
All other paperwork completed with this study (e.g., health histories, questionnaires) will be kept in secure laboratories and offices.
In the event of any publication or presentation resulting from the research, no personally identifiable information will be shared.
7b. The use of private health information: Health information about you will be collected if you choose to be part of this research study. Health information is protected by law as explained in the HMC Privacy Notice. If you have not received this notice, please request a copy from the researcher. At The Milton S. Hershey Medical Center (HMC) and Penn State College of Medicine (PSU) your information will only be used or shared as explained and authorized in this consent form or when required by law. It is possible that some of the other people/groups who receive your health information may not be required by Federal privacy laws to protect your information and may share it without your permission.
To participate in this research you must allow the research team to use your health information. If you do not want us to use your protected health information, you may not participate in this research. People usually have a right to access their medical records. However, while the research is in progress, you may not be allowed to see or copy certain information that is related to this research study. This is only for the period of the research. You will be allowed to see that information when the entire research project is complete. Your permission for the use, retention, and sharing of your identifiable health information will continue indefinitely. Should there be a time that the information is no longer needed the research information not already in your medical record will be destroyed. Any research information in your medical record will be kept indefinitely.
If you choose to participate, you are free to withdraw your permission for the use and sharing of your health information and samples at any time. You must do this in writing. Write to Dr. Ray and let him know that you are withdrawing from the research study. His mailing address is Penn State College of Medicine, Division of Cardiology, H047, 500 University Dr., Hershey, PA 17033.

127
If you withdraw your permission: • We will no longer use or share medical information about you or your
samples for this research study, except when the law allows us to do so. • We are unable to take back anything we have already done or any
information we have already shared with your permission. • We may continue using and sharing the information obtained prior to
your withdrawal if it is necessary for the soundness of the overall research.
• We will keep our records of the care that we provided to you as long as the law requires.
The research team may use the following sources of health information.
• All data collected during the testing period including: ECG tracings, medical history, blood pressure measurements, muscle temperatures, exercise workloads, microneurography data, ultrasound data, blood samples, and blood flow measurements.
• Competed questionnaires
Representatives of the following people/groups within HMC/PSU may use your health information and share it with other specific groups in connection with this research study.
• The principal investigator, Chester A. Ray, Ph.D. • The HMC/PSU Institutional Review Board • The HMC/PSU Human Subjects Protection Office • The research team • The study coordinators
The above people/groups may share your health information with the following people/groups outside HMC/PSU for their use in connection with this research study. These groups, while monitoring the research study, may also review and/or copy your original PSU/HMC records.
• The Office of Human Research Protections in the U. S. Department of Health and Human Services
• The National Institutes of Health • Food and Drug Administration
8. Costs for Participation:
a. Costs: There will be no added cost to you for participating in this research study.
b. Treatment and compensation for injury: Every effort to prevent injury as a result of your participation will be taken. It is possible, however, that you could develop complications or injuries as a result of participating in this research study. In the event of injury resulting from this research, medical treatment is available but will be provided at the usual charge. It is the policy of this institution to provide neither financial compensation nor free medical treatment for research-related injury.

128
Costs for the treatment of research-related injuries will be charged to your insurance carrier or to you. Some insurance companies may not cover costs associated with research studies. If for any reason these costs are not covered by your insurance, they will be your responsibility. You will also be responsible for any deductibles, w-insurance, and or co-pay.
You are not waiving any legal rights you may have by signing this form.
10. Compensation for Participation:
You will be compensated $15/hour for each hour spent in the laboratory. If the study is terminated early, due to you with withdrawing or the investigator stopping the study, compensation will be $15/hour.
10. Research Funding: The institution and investigators are receiving a grant from The National Institutes of Health to support the activities that are required to conduct this research.
11. Voluntary Participation:
Taking part in this research study is voluntary. If you choose to take part in this research, your major responsibilities will include: following the instructions of the investigators during the collection of data. You do not have to participate in this research. If you choose to take part, you have the right to stop at any time. If you decide not to participate or if you decide to stop taking part in the research at a later date, there will be no penalty or loss of benefits to which you are entitled. In other words, your decision to decline to participate in this research or to stop taking part in the research will not affect your medical care and academic standing and job status. Your research doctor or the sponsor may take you out of the research study without your permission. Possible reasons for this are: You do not adhere to the investigator’s instructions or follow the protocol as stated in this consent form. Also, the sponsor of the research may end the research study early. If your participation in the research ends early, you may be asked to visit the research doctor for a final visit.
If you will be participating in another clinical trial at Hershey Medical Center or elsewhere while in this research, you should discuss the procedures and/or treatments with your physician or the investigators. This precaution is intended to protect you from possible side effects from interactions of research drugs, treatments or testing. During the course of the visit you will be provided with any significant new findings that may affect your willingness to continue participating in this research.

129
12. Contact Information for Questions or Concerns:
You have the right to ask any questions you may have about this research. If you have questions or concerns or believe you may have developed an injury that is related to this research, contact Dr. Ray at 717-531-3906 or you can page the Cardiology doctor on 24-hour call at 717-531-8521. If you have questions regarding your rights as a research participant or you have concerns or general questions about the research or about your privacy and the use of your personal health information, you may contact the research protection advocate in the HMC Human Subjects Protection Office at 717-531-5687. You may also call this number if you cannot reach the research team and wish to talk to someone else.
For more information about participation in a research study and about the Institutional Review Board (IRB), a group of people who review the research to protect your rights, please visit the HMC IRB’s Web site at http://www.hmc.psu.edu/irb. Included on this web site, under the heading “Participant Info”, you can access the federal regulations and information about the protection of human research participants. If you do not have access to the internet, copies of these federal regulations are available by calling the HSPO at (717) 531-5687.
Signature and Consent/Permission to be in the Research Before making the decision regarding enrollment in this research you should have:
• Discussed this study with an investigator, • Reviewed the information in this form, and • Had the opportunity to ask any questions you may have.
Your signature below means that you have received this information, have asked the questions you currently have about the research and those questions have been answered. You will receive a copy of the signed and dated form to keep for future reference. Participant: By signing this consent form, you indicate that you are voluntarily choosing to take part in this research. ___________________________ __________ ______ Signature of Participant Date Time ___________________________ Printed Name Person Explaining the Research: Your signature below means that you have explained the research to the participant/participant representative and have answered any questions he/she has about the research.

130
______________________________ _________ ______ Signature of person who explained this research Date Time ______________________________ Printed Name (Only approved investigators for this research may explain the research and obtain informed consent.)

131
Appendix C
Raw Data For Young Normothermia & Heating Studies

132
YOUNG NORMOTHERMIA
Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
1 SBP 122.67 122.10 129.00 131.30 141.80 1 DBP 64.54 66.03 70.38 76.16 82.03 1 MAP 83.67 84.00 90.00 95.00 102.00 1 HR 60.00 64.00 75.00 84.00 90.00 1 Skin Temp 1 29.86 30.28 30.34 30.37 30.40 1 Skin Temp 2 28.98 29.34 29.39 29.41 29.44 1 Skin Temp 3 30.49 30.43 30.44 30.42 30.43 1 Bursts 3.00 5.00 3.86 1.29 1.29 1 Total Activity 464.14 413.52 368.42 165.38 112.56 1 Muscle Temp 31.12 31.28 31.20 31.49 31.59 1 RPE 6 0 11 15 19 1 REN TAP 53.10 55.88 54.00 57.20 51.85 1 REN CON 0.63 0.67 0.60 0.60 0.51 1 CBF 1.25 2.14 2.95 2.67 2.61 1 CVC 0.015 0.025 0.033 0.028 0.026 2 SBP 157.47 163.40 171.00 186.50 208.90 2 DBP 84.81 87.62 91.63 99.78 111.30 2 MAP 105.33 110.00 116.00 125.00 141.00 2 HR 61.67 71.00 78.00 81.00 85.00 2 Skin Temp 1 31.53 31.31 31.22 31.15 31.12 2 Skin Temp 2 31.92 31.70 31.60 31.51 31.46 2 Skin Temp 3 29.72 29.55 29.45 29.42 29.41 2 Muscle Temp 33.99 33.92 34.00 34.00 33.91 2 RPE 6 7 11 13 17 2 REN TAP 64.97 65.28 56.30 61.00 58.28 2 REN CON 0.62 0.59 0.49 0.49 0.41 2 CBF 1.47 1.75 2.18 3.03 3.19 2 CVC 0.014 0.016 0.019 0.024 0.023 3 SBP 121.50 120.80 123.10 132.10 148.10 3 DBP 92.33 95.66 103.70 112.10 122.60 3 MAP 105.00 108.00 113.00 122.00 140.00 3 HR 70.67 71.00 83.00 88.00 95.00 3 Skin Temp 1 31.13 31.64 0.00 0.00 0.00 3 Skin Temp 2 30.49 30.58 30.64 30.68 30.71 3 Skin Temp 3 31.23 31.96 31.97 31.92 31.86 3 Muscle Temp 33.89 34.21 34.40 34.44 34.45 3 RPE 6 0 13 13 16 3 REN TAP 47.85 48.80 45.50 48.80 45.50 3 REN CON 0.46 0.45 0.40 0.40 0.33 3 CBF 0.80 0.84 0.81 0.73 0.86 3 CVC 0.008 0.008 0.007 0.006 0.006

133
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
1 SBP 148.40 154.90 154.80 156.40 144.70 1 DBP 86.99 93.44 94.37 87.10 82.11 1 MAP 107.00 115.00 115.00 109.00 99.00 1 HR 99.00 104.00 105.00 71.00 68.00 1 Skin Temp 1 30.41 30.41 30.42 30.47 40.16 1 Skin Temp 2 29.47 29.49 29.50 29.56 40.67 1 Skin Temp 3 30.44 30.46 30.46 30.45 32.40 1 Bursts 5.15 15.45 20.60 16.00 11.00 1 Total Activity 604.26 1642.50 2116.32 3690.23 3380.41 1 Muscle Temp 31.64 31.64 31.64 31.58 38.90 1 RPE 19 19 19 0 0 1 REN TAP 52.90 48.70 43.40 52.13 51.87 1 REN CON 0.49 0.42 0.38 0.48 0.52 1 CBF 3.03 2.95 2.95 1.50 1.37 1 CVC 0.028 0.026 0.026 0.01 0.01 2 SBP 222.90 228.40 228.40 216.10 187.00 2 DBP 122.00 126.70 127.60 114.80 91.84 2 MAP 154.00 161.00 161.00 145.00 118.00 2 HR 89.00 96.00 95.00 79.00 76.00 2 Skin Temp 1 31.10 31.07 31.07 31.06 30.99 2 Skin Temp 2 31.40 31.35 31.34 31.33 31.34 2 Skin Temp 3 29.36 29.35 29.34 29.30 29.17 2 Muscle Temp 33.96 33.96 33.95 33.69 33.87 2 RPE 19 20 20 0 0 2 REN TAP 54.15 56.30 54.10 64.20 65.70 2 REN CON 0.35 0.35 0.34 0.44 0.56 2 CBF 3.86 5.36 4.80 2.35 3.34 2 CVC 0.025 0.033 0.030 0.02 0.03 3 SBP 153.30 152.80 153.30 141.00 134.90 3 DBP 127.10 122.30 120.80 103.60 100.70 3 MAP 141.00 139.00 138.00 128.00 110.00 3 HR 97.00 99.00 103.00 79.00 61.00 3 Skin Temp 1 0.00 0.00 31.73 31.75 31.84 3 Skin Temp 2 30.72 30.72 30.72 30.76 30.73 3 Skin Temp 3 31.81 31.76 31.74 31.72 31.73 3 Muscle Temp 34.50 34.50 34.50 34.44 34.63 3 RPE 18 18 18 0 0 3 REN TAP 43.63 43.83 42.40 44.03 44.68 3 REN CON 0.31 0.32 0.31 0.34 0.41 3 CBF 0.84 0.74 0.77 0.93 0.77 3 CVC 0.006 0.005 0.006 0.01 0.01

134
YOUNG HEAT
Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20% 139.60 141.80 1 SBP 138.97 140.80 144.10
82.58 82.54 1 DBP 73.28 80.83 84.88 100.00 100.00 1 MAP 94.67 99.00 103.00
62.00 62.00 1 HR 72.33 84.00 91.00 39.28 38.95 1 Skin Temp 1 39.72 39.65 40.02 40.06 39.73 1 Skin Temp 2 39.96 39.92 40.32 32.35 32.36 1 Skin Temp 3 31.49 31.66 31.72 12.50 12.50 1 Bursts 3.00 4.00 4.30
3592.43 2879.64 1 Total Activity 514.40 317.58 432.39
38.75 38.47 1 Muscle Temp 38.50 38.37 38.33
0 0 1 RPE 6 0 15 56.80 50.83 1 REN TAP 48.00 44.48 48.70
0.57 0.51 1 REN CON 0.51 0.45 0.47 0.82 1.03 1 CBF 1.84 2.85 3.84 0.01 0.01 1 CVC 0.019 0.025 0.037
188.00 187.70 2 SBP 156.50 165.40 178.10 95.90 97.66 2 DBP 84.81 87.00 97.17
122.00 124.00 2 MAP 104.67 109.00 122.00 70.00 66.00 2 HR 64.33 71.00 84.00 30.88 30.95 2 Skin Temp 1 40.09 40.01 40.00 31.49 31.64 2 Skin Temp 2 40.90 40.72 40.69 29.12 29.06 2 Skin Temp 3 28.75 28.70 28.67
34.18 34.26 2 Muscle Temp 38.24 38.44 38.64
0 0 2 RPE 6 9 64.83 64.95 2 REN TAP 63.63 62.40 65.30
0.53 0.52 2 REN CON 0.61 0.57 0.54 2.20 1.81 2 CBF 2.05 1.28 2.34 0.02 0.01 2 CVC 0.020 0.012 0.019
117.60 113.10 3 SBP 133.47 141.50 147.30 90.57 86.44 3 DBP 92.35 93.38 106.50
103.00 99.00 3 MAP 109.33 111.00 127.00 65.00 64.00 3 HR 72.33 75.00 86.00 32.03 32.09 3 Skin Temp 1 38.61 38.60 0.00 30.66 30.65 3 Skin Temp 2 30.37 30.37 30.37 31.93 32.03 3 Skin Temp 3 39.20 39.09 39.23
34.59 34.57 3 Muscle Temp 39.24 39.18 39.16
0 0 3 RPE 6 0 14 48.53 46.60 3 REN TAP 53.02 50.23 48.25
0.47 0.47 3 REN CON 0.48 0.45 0.38 0.71 0.78 3 CBF 0.75 0.80 0.95 0.01 0.01 3 CVC 0.007 0.007 0.007

135
YOUNG HEAT Subject Variable 20-40% 40-60% 60-80% 80-100%
1 SBP 149.30 151.80 155.40 159.30 1 DBP 89.62 92.61 100.40 107.90 1 MAP 109.00 113.00 119.00 125.00 1 HR 101.00 96.00 104.00 112.00 1 Skin Temp 1 40.12 40.20 40.28 40.37 1 Skin Temp 2 40.43 40.54 40.64 40.74 1 Skin Temp 3 31.75 31.79 31.85 31.95 1 Bursts 12.90 19.35 15.05 21.50 1 Total Activity 1053.52 1748.36 2155.41 2977.77 1 Muscle Temp 38.41 38.41 38.41 38.41 1 RPE 19 19 19 19 1 REN TAP 50.80 47.60 45.50 45.50 1 REN CON 0.47 0.42 0.38 0.36 1 CBF 4.04 4.87 2.45 1.25 1 CVC 0.037 0.043 0.021 0.010 2 SBP 189.00 206.20 224.30 229.60 2 DBP 100.10 114.40 124.80 136.60 2 MAP 127.00 143.00 157.00 170.00 2 HR 89.00 88.00 96.00 104.00 2 Skin Temp 1 40.01 40.01 39.98 39.93 2 Skin Temp 2 40.70 40.72 40.71 40.65 2 Skin Temp 3 28.63 28.62 28.62 28.63 2 Muscle Temp 38.64 38.75 38.75 38.89 2 RPE 9 16 16 18 2 REN TAP 65.00 52.20 52.20 52.20 2 REN CON 0.51 0.37 0.33 0.31 2 CBF 2.28 2.69 1.33 0.98 2 CVC 0.018 0.019 0.009 0.006 3 SBP 152.50 156.70 155.60 154.70 3 DBP 112.20 121.50 126.50 123.40 3 MAP 129.00 139.00 143.00 144.00 3 HR 90.00 96.00 92.00 105.00 3 Skin Temp 1 0.00 0.00 0.00 0.00 3 Skin Temp 2 30.39 30.40 30.40 30.40 3 Skin Temp 3 39.32 39.40 39.47 39.52 3 Muscle Temp 39.16 39.22 39.24 39.24 3 RPE 14 16 19 19 3 REN TAP 47.70 44.40 41.65 41.10 3 REN CON 0.37 0.32 0.29 0.29 3 CBF 0.86 1.17 0.89 0.96 3 CVC 0.007 0.008 0.006 0.007

136
YOUNG HEAT Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
4 SBP 148.20 137.60 119.30 117.40 116.70 4 DBP 91.02 86.80 67.05 65.01 62.95 4 MAP 108.00 103.00 81.00 79.00 77.00 4 HR 72.00 64.00 60.00 55.00 54.00 4 Skin Temp 1 39.34 41.43 40.96 39.95 39.40 4 Skin Temp 2 40.02 42.53 41.82 40.79 40.15 4 Skin Temp 3 28.30 28.81 28.92 28.97 28.97 4 Bursts 20.00 19.50 11.00 11.50 9.00 4 Total Activity 8666.50 6760.36 2792.83 3342.60 2647.15 4 Muscle Temp 38.64 38.71 38.94 38.77 38.69 4 RPE 20 0 0 0 0 4 REN TAP 71.80 88.00 76.30 75.40 76.48 4 REN CON 0.66 0.85 0.94 0.95 0.99 4 CBF 1.29 1.56 1.36 1.06 1.50 4 CVC 0.012 0.015 0.017 0.013 0.019 5 SBP 213.50 194.50 166.40 151.80 151.50 5 DBP 135.00 121.20 90.70 78.23 74.88 5 MAP 164.00 151.00 112.00 101.00 99.00 5 HR 97.00 92.00 67.00 57.00 50.00 5 Skin Temp 1 38.54 38.63 38.83 38.57 38.37 5 Skin Temp 2 39.11 39.10 39.05 38.74 38.39 5 Skin Temp 3 29.92 29.95 30.05 30.05 30.01 5 Bursts 4.62 9.50 8.50 5.00 2.00 5 Total Activity 1260.49 2202.96 2074.57 1391.40 510.90 5 Muscle Temp 38.73 38.81 38.61 38.48 38.42 5 RPE 17 0 0 0 0 5 REN TAP 44.90 54.95 48.67 53.70 52.10 5 REN CON 0.27 0.36 0.43 0.53 0.53 5 CBF 1.48 1.28 1.41 1.76 1.48 5 CVC 0.009 0.008 0.013 0.017 0.015 6 SBP 156.80 143.30 127.60 124.90 119.40 6 DBP 91.76 80.01 66.25 65.39 62.59 6 MAP 119.00 101.00 84.00 83.00 80.00 6 HR 90.00 69.00 62.00 61.00 58.00 6 Skin Temp 1 39.75 39.83 39.44 38.66 38.42 6 Skin Temp 2 41.75 41.82 41.68 41.07 40.80 6 Skin Temp 3 30.57 30.65 30.58 30.55 30.44 6 Bursts 14.40 5.00 1.00 2.50 1.00 6 Total Activity 2852.06 1276.42 165.32 548.10 69.95 6 Muscle Temp 39.06 39.21 39.40 38.65 38.51 6 RPE 19 0 0 0 0 6 REN TAP 34.20 46.15 49.55 53.53 52.10 6 REN CON 0.29 0.46 0.59 0.64 0.65 6 CBF 0.98 1.69 1.89 1.45 1.85 6 CVC 0.008 0.017 0.023 0.018 0.023

137
YOUNG HEAT Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
1 SBP 158.10 166.30 143.00 137.60 129.40 1 DBP 110.30 105.80 76.70 73.66 74.43 1 MAP 127.00 126.00 95.00 91.00 90.00 1 HR 114.00 90.00 62.00 59.00 59.00 1 Skin Temp 1 40.39 40.43 30.53 30.45 30.39 1 Skin Temp 2 40.76 40.93 29.74 29.91 30.12 1 Skin Temp 3 31.97 32.20 30.40 30.47 30.48 1 Bursts 17.16 26.50 12.50 11.00 6.50 1 Total Activity 2688.54 5451.22 2152.68 1898.97 899.32
1 Muscle Temp 38.41 38.62 31.97 32.15 32.34
1 RPE 19 0 0 0 0 1 REN TAP 45.50 45.78 49.50 44.47 48.00 1 REN CON 0.36 0.36 0.52 0.49 0.53 1 CBF 1.25 1.07 1.83 1.15 1.46 1 CVC 0.010 0.008 0.019 0.013 0.016 2 SBP 230.40 222.90 222.90 218.10 179.00 2 DBP 141.30 120.20 120.20 118.10 93.76 2 MAP 174.00 154.00 154.00 151.00 119.00 2 HR 106.00 93.00 93.00 81.00 71.00 2 Skin Temp 1 39.91 39.84 39.84 39.72 39.43 2 Skin Temp 2 40.64 40.54 40.54 40.34 40.08 2 Skin Temp 3 28.63 28.55 28.55 28.53 28.48
2 Muscle Temp 38.89 39.31 38.64 37.98 37.83
2 RPE 18 0 0 0 0 2 REN TAP 52.20 71.10 65.80 66.80 62.40 2 REN CON 0.30 0.46 0.43 0.44 0.52 2 CBF 2.95 3.72 1.78 2.41 1.83 2 CVC 0.017 0.024 0.012 0.016 0.015 3 SBP 154.40 147.10 134.50 123.90 121.90 3 DBP 126.70 103.40 89.01 85.72 82.82 3 MAP 145.00 120.00 106.00 101.00 98.00 3 HR 107.00 76.00 69.00 70.00 69.00 3 Skin Temp 1 39.03 39.10 39.09 38.81 38.66 3 Skin Temp 2 30.40 30.34 30.31 30.31 30.30 3 Skin Temp 3 39.53 39.56 39.46 39.04 38.82
3 Muscle Temp 39.24 39.53 39.60 39.34 39.14
3 RPE 19 0 0 0 0 3 REN TAP 41.10 46.90 51.03 49.90 49.17 3 REN CON 0.28 0.39 0.48 0.49 0.50 3 CBF 1.05 0.76 0.71 0.74 0.73 3 CVC 0.007 0.006 0.007 0.007 0.007

138
Variable BL AVE ISCH 0-20% 20-40% 40-60%
SBP 119.93 127.00 133.50 136.40 139.10 DBP 69.40 71.11 75.61 78.76 80.91 MAP 81.00 84.00 90.00 93.00 98.00 HR 51.00 62.00 66.00 66.00 67.00 Skin Temp 1 31.40 31.34 31.29 31.30 31.31 Skin Temp 2 31.87 31.80 0.00 0.00 0.00 Skin Temp 3 30.89 30.84 30.81 30.85 30.82 Bursts 1.67 3.00 0.65 1.96 1.31 Total Activity 296.05 448.50 109.43 427.52 130.05 Muscle Temp 34.87 34.87 34.86 34.85 34.84 RPE 6 0 12 12 14 REN TAP 72.00 70.00 62.80 62.80 60.15 REN CON 0.89 0.83 0.70 0.68 0.61 CBF 1.58 2.51 1.91 1.80 1.54 CVC 0.019 0.030 0.021 0.019 0.016 SBP 139.07 147.10 144.80 170.50 167.60 DBP 80.95 90.07 92.05 110.40 109.60 MAP 101.33 110.00 116.00 134.00 132.00 HR 66.33 67.00 93.00 88.00 92.00 Skin Temp 1 30.07 30.37 30.49 30.64 30.77 Skin Temp 2 29.70 30.06 30.19 30.19 30.21 Skin Temp 3 29.56 29.53 29.48 29.51 29.56 Bursts 0.83 1.00 0.70 0.00 1.41 Total Activity 164.29 79.12 105.80 0.00 319.45 Muscle Temp 34.18 34.21 34.22 34.23 34.24 RPE 6 0 11 12 14 REN TAP 54.93 54.30 57.40 52.40 49.90 REN CON 0.54 0.49 0.49 0.39 0.38 CBF 2.03 1.57 2.34 2.13 1.70 CVC 0.020 0.014 0.020 0.016 0.013 SBP 137.20 141.30 147.50 154.00 158.00 DBP 77.17 77.61 84.32 89.40 90.93 MAP 95.67 97.00 105.00 112.00 114.00 HR 56.67 56.00 63.00 71.00 76.00 Skin Temp 1 31.55 31.62 31.63 31.63 31.63 Skin Temp 2 30.69 30.71 30.72 30.72 30.72 Skin Temp 3 30.18 30.02 29.98 29.96 29.97 Bursts 2.67 3.00 2.88 1.44 4.32 Total Activity 436.75 269.51 265.42 91.56 650.94 Muscle Temp 34.55 34.56 34.44 34.44 34.47 RPE 6 0 12 12 15 REN TAP 58.87 58.38 57.50 54.95 52.40 REN CON 0.62 0.60 0.55 0.49 0.46 CBF 1.24 1.55 1.57 1.48 1.59 CVC 0.013 0.016 0.015 0.013 0.014

139
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
4 SBP 146.20 150.90 153.30 147.50 134.40 4 DBP 85.92 86.37 88.84 76.15 62.16 4 MAP 106.00 110.00 110.00 97.00 80.00 4 HR 70.00 75.00 75.00 59.00 56.00 4 Skin Temp 1 31.31 31.32 31.32 31.36 31.44 4 Skin Temp 2 0.00 0.00 31.76 31.77 31.72 4 Skin Temp 3 30.83 30.86 30.85 30.84 30.80 4 Bursts 2.62 5.23 7.85 5.00 1.50 4 Total Activity 428.85 863.58 1775.74 794.59 148.40 4 Muscle Temp 34.84 34.84 34.84 34.76 34.76 4 RPE 17 18 19 0 0 4 REN TAP 71.80 79.00 75.40 72.70 74.80 4 REN CON 0.68 0.72 0.69 0.75 0.94 4 CBF 1.14 1.22 1.52 1.63 1.81 4 CVC 0.011 0.011 0.014 0.02 0.02 5 SBP 179.60 184.60 190.20 175.00 163.30 5 DBP 118.00 122.40 127.00 111.20 92.39 5 MAP 143.00 149.00 154.00 138.00 116.00 5 HR 96.00 98.00 98.00 89.00 64.00 5 Skin Temp 1 30.85 30.90 30.91 30.95 31.08 5 Skin Temp 2 30.25 30.30 30.32 30.38 30.53 5 Skin Temp 3 29.55 29.53 29.53 29.48 29.56 5 Bursts 1.41 2.11 2.82 2.00 4.00 5 Total Activity 561.01 834.93 1353.20 264.68 1168.00 5 Muscle Temp 34.23 34.23 34.23 34.20 34.34 5 RPE 16 17 17 0 0 5 REN TAP 47.70 46.20 46.20 53.67 47.83 5 REN CON 0.33 0.31 0.30 0.39 0.41 5 CBF 1.89 1.54 1.24 1.69 1.03 5 CVC 0.013 0.010 0.008 0.01 0.01 6 SBP 166.30 176.70 178.90 162.30 149.90 6 DBP 97.56 103.50 105.80 90.72 78.51 6 MAP 122.00 131.00 133.00 114.00 100.00 6 HR 85.00 87.00 89.00 67.00 65.00 6 Skin Temp 1 31.63 31.62 31.61 31.64 31.72 6 Skin Temp 2 30.70 30.66 30.66 30.67 30.73 6 Skin Temp 3 29.95 29.96 29.96 29.97 29.89 6 Bursts 8.64 12.96 14.40 8.00 3.00 6 Total Activity 1317.11 1545.39 1604.28 1507.67 642.66 6 Muscle Temp 34.47 34.53 34.53 34.55 34.52 6 RPE 15 18 18 0 0 6 REN TAP 51.30 46.70 44.40 46.15 55.80 6 REN CON 0.42 0.36 0.33 0.40 0.56 6 CBF 1.48 0.84 0.84 1.05 1.28 6 CVC 0.012 0.006 0.006 0.01 0.01

140
Rec 2 Rec 3 Subject Variable BL AVE ISCH 137.20 141.90 4 SBP 108.60 115.70
67.18 73.43 4 DBP 63.40 69.12 85.00 90.00 4 MAP 74.33 77.00 53.00 56.00 4 HR 54.00 56.00 31.58 31.75 4 Skin Temp 1 40.53 40.68 31.69 31.82 4 Skin Temp 2 42.43 42.45 30.77 30.79 4 Skin Temp 3 28.68 28.78
3.00 4.00 4 Bursts 6.50 5.00 551.82 626.93 4 Total Activity 1495.54 1742.00
34.93 35.02 4 Muscle Temp 38.30 38.31 0 0 4 RPE 6 0
70.72 73.60 4 REN TAP 71.75 73.15 0.83 0.82 4 REN CON 0.97 0.95 1.46 1.29 4 CBF 1.35 1.66 0.02 0.01 4 CVC 0.018 0.022
156.60 148.30 5 SBP 142.27 145.10 84.66 80.37 5 DBP 75.38 79.31
109.00 104.00 5 MAP 96.00 102.00 54.00 54.00 5 HR 63.33 83.00 31.21 31.29 5 Skin Temp 1 38.85 38.69 30.66 30.72 5 Skin Temp 2 39.13 38.90 29.50 29.52 5 Skin Temp 3 29.87 29.85
3.00 1.50 5 Bursts 0.50 0.00 1012.35 345.75 5 Total Activity 87.93 0.00
34.65 34.74 5 Muscle Temp 38.28 38.31 0 0 5 RPE 6 0
49.90 50.77 5 REN TAP 55.81 57.00 0.46 0.49 5 REN CON 0.58 0.56 1.42 1.65 5 CBF 1.83 2.56 0.01 0.02 5 CVC 0.019 0.025
148.40 146.40 6 SBP 108.80 114.70 81.08 78.73 6 DBP 60.41 63.65
102.00 100.00 6 MAP 75.33 78.00 61.00 59.00 6 HR 57.00 61.00 31.97 32.19 6 Skin Temp 1 39.26 38.88 30.83 30.83 6 Skin Temp 2 42.80 41.64 29.94 29.89 6 Skin Temp 3 30.97 30.74
5.00 3.50 6 Bursts 1.00 1.00 848.29 324.58 6 Total Activity 163.89 78.53
34.43 34.51 6 Muscle Temp 38.92 38.90 0 0 6 RPE 6 0
51.63 54.40 6 REN TAP 54.55 50.17 0.51 0.54 6 REN CON 0.72 0.64 1.18 1.56 6 CBF 1.78 2.26 0.01 0.02 6 CVC 0.024 0.029

141
YOUNG HEAT Subject Variable 20-40% 40-60% 60-80% 80-100%
4 SBP 124.50 130.70 137.20 143.70 4 DBP 71.56 79.87 84.40 90.89 4 MAP 85.00 94.00 101.00 107.00 4 HR 62.00 62.00 65.00 74.00 4 Skin Temp 1 41.33 41.47 41.56 41.58 4 Skin Temp 2 41.33 41.47 41.56 41.58 4 Skin Temp 3 28.81 28.79 28.82 28.83 4 Bursts 6.25 15.00 20.00 23.75 4 Total Activity 1987.25 3359.13 7635.15 9069.00 4 Muscle Temp 38.39 38.48 38.56 38.64 4 RPE 14 17 18 20 4 REN TAP 70.00 75.40 71.80 71.80 4 REN CON 0.82 0.80 0.71 0.67 4 CBF 1.41 1.10 1.36 1.29 4 CVC 0.017 0.012 0.013 0.012 5 SBP 166.00 185.90 197.40 210.90 5 DBP 97.63 110.80 118.70 131.10 5 MAP 120.00 138.00 146.00 161.00 5 HR 92.00 97.00 101.00 98.00 5 Skin Temp 1 38.63 38.57 38.54 38.54 5 Skin Temp 2 39.01 39.07 39.10 39.11 5 Skin Temp 3 29.87 29.88 29.88 29.91 5 Bursts 0.77 2.31 3.08 3.08 5 Total Activity 199.51 493.57 621.62 678.69 5 Muscle Temp 38.51 38.61 38.68 38.73 5 REN TAP 49.10 51.57 47.40 46.57 5 REN CON 0.41 0.37 0.32 0.29 5 CBF 2.36 1.53 1.98 1.61 5 CVC 0.020 0.011 0.014 0.010 6 SBP 123.70 135.70 141.90 154.60 6 DBP 70.15 78.12 82.73 90.96 6 MAP 88.00 99.00 104.00 117.00 6 HR 79.00 80.00 87.00 89.00 6 Skin Temp 1 39.38 39.51 39.62 39.73 6 Skin Temp 2 41.54 41.61 41.67 41.74 6 Skin Temp 3 30.61 30.70 30.66 30.56 6 Bursts 2.88 2.88 8.64 7.20 6 Total Activity 435.27 675.50 2036.92 1426.09 6 Muscle Temp 39.00 39.03 39.03 39.06 6 RPE 10 15 15 19 6 REN TAP 55.80 45.00 28.50 34.20 6 REN CON 0.63 0.45 0.27 0.29 6 CBF 2.21 1.21 1.15 0.98 6 CVC 0.025 0.012 0.011 0.008

142
YOUNG NORMOTHERMIA Subject Variable BL AV ISCH 0-20% 20-40% 40-60%
7 SBP 120.20 125.60 131.60 134.60 147.30 7 DBP 61.36 65.60 73.28 78.48 86.13 7 MAP 76.00 81.00 89.00 95.00 106.00 7 HR 63.33 62.00 70.00 73.00 78.00 7 Skin Temp 1 29.04 29.67 29.81 29.93 30.03 7 Skin Temp 2 29.69 30.13 30.19 30.27 30.32 7 Skin Temp 3 31.45 31.35 31.34 31.32 31.29 7 Bursts 1.17 8.00 9.43 7.71 7.71 7 Total Activity 534.43 2430.30 5379.93 4630.57 4924.81 7 Muscle Temp 34.50 34.44 34.53 34.57 34.60 7 RPE 6 0 12 14 16 7 REN TAP 50.97 50.97 49.92 50.31 48.86 7 REN CON 0.67 0.48 0.56 0.53 0.46 7 CBF 1.65 1.68 2.17 2.03 2.19 7 CVC 0.022 0.021 0.024 0.021 0.021 8 SBP 130.80 141.36 142.06 150.57 161.14 8 DBP 77.66 86.23 86.33 92.66 99.68 8 MAP 94.12 103.91 105.50 112.57 121.79 8 HR 60.55 64.45 78.00 77.00 80.00 8 Skin Temp 1 29.83 30.15 30.31 30.41 30.51 8 Skin Temp 2 28.79 29.00 29.08 29.13 29.19 8 Skin Temp 3 27.74 27.73 27.74 27.79 27.78 8 Bursts 4.50 12.00 4.50 11.25 18.00 8 Total Activity 700.41 1712.42 768.98 2093.89 3733.29 8 Muscle Temp 34.50 34.66 34.65 34.75 34.79 8 RPE 6 0 9 15 18 8 REN TAP 54.55 55.68 53.40 55.47 54.53 8 REN CON 0.54 0.54 0.51 0.49 0.45 8 CBF 1.19 1.09 1.29 1.04 0.92 8 CVC 0.013 0.011 0.012 0.009 0.008 9 SBP 132.66 114.30 143.87 152.23 161.87 9 DBP 77.28 82.50 88.39 94.77 101.25 9 MAP 94.90 73.00 107.54 114.26 122.84 9 HR 48.00 57.00 78.00 78.00 81.00 9 Skin Temp 1 30.49 33.28 30.74 30.74 30.74 9 Skin Temp 2 29.51 13.35 26.93 29.74 20.85 9 Skin Temp 3 31.94 31.98 31.92 31.92 31.92 9 Bursts 0.50 2.00 0.75 3.75 8.25 9 Total Activity 63.93 238.53 137.93 939.60 2574.38 9 Muscle Temp 34.73 35.04 35.23 35.30 35.30 9 RPE 6 0 11 13 15 9 REN TAP 52.09 53.00 49.35 48.07 49.93 9 REN CON 0.84 0.73 0.46 0.42 0.41 9 CBF 1.44 1.55 1.79 2.11 2.24 9 CVC 0.015 0.021 0.017 0.018 0.018

143
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
7 SBP 154.50 166.40 168.70 145.80 130.60 7 DBP 88.24 93.93 95.45 76.26 61.49 7 MAP 110.00 119.00 121.00 97.00 80.00 7 HR 82.00 81.00 83.00 63.00 61.00 7 Skin Temp 1 30.10 30.16 30.17 30.20 30.30 7 Skin Temp 2 30.36 30.42 30.43 30.44 30.52 7 Skin Temp 3 31.26 31.25 31.27 31.39 31.35 7 Bursts 12.00 13.71 11.97 5.50 6.00 7 Total Activity 6779.49 9287.62 11163.22 643.97 405.26 7 Muscle Temp 34.61 34.63 34.64 34.57 34.74 7 RPE 18 19 19 0 0 7 REN TAP 47.40 47.49 47.49 41.40 53.80 7 REN CON 0.43 0.40 0.39 0.43 0.67 7 CBF 2.51 2.62 2.62 2.04 2.22 7 CVC 0.023 0.022 0.022 0.02 0.03 8 SBP 169.48 175.80 175.80 145.80 130.60 8 DBP 105.66 108.67 108.60 76.26 61.49 8 MAP 129.29 134.36 135.35 97.00 80.00 8 HR 78.00 76.00 75.00 63.00 61.00 8 Skin Temp 1 30.58 30.64 30.66 30.20 30.30 8 Skin Temp 2 29.27 29.34 29.35 30.44 30.52 8 Skin Temp 3 27.78 27.81 27.82 31.39 31.35 8 Bursts 18.75 28.50 27.00 5.50 6.00 8 Total Activity 3977.06 7289.71 8337.42 643.97 405.26 8 Muscle Temp 34.89 34.95 34.95 34.57 34.74 8 RPE 19 20 20 0 0 8 REN TAP 43.00 45.70 39.60 41.40 53.80 8 REN CON 0.33 0.34 0.29 0.43 0.67 8 CBF 1.41 1.80 1.73 2.04 2.22 8 CVC 0.011 0.013 0.013 0.02 0.03 9 SBP 169.63 176.34 176.34 150.90 108.20 9 DBP 107.39 110.94 110.94 60.66 61.56 9 MAP 130.23 135.88 135.88 91.00 62.00 9 HR 79.00 76.00 74.00 47.00 44.00 9 Skin Temp 1 30.75 30.77 30.78 30.78 30.77 9 Skin Temp 2 10.12 22.18 20.07 23.20 9.48 9 Skin Temp 3 31.97 32.01 32.01 31.97 31.69 9 Bursts 10.25 12.75 15.00 13.50 8.00 9 Total Activity 3225.00 3876.30 4915.05 5537.95 2372.45 9 Muscle Temp 35.31 35.30 35.29 35.05 35.78 9 RPE 17 17 18 0 0 9 REN TAP 47.15 51.00 51.00 47.70 47.43 9 REN CON 0.36 0.38 0.38 0.52 0.76 9 CBF 2.13 2.63 1.75 1.61 1.98 9 CVC 0.016 0.019 0.013 0.02 0.03

144
YOUNG HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
125.60 124.70 7 SBP 100.15 112.50 118.60 57.99 58.49 7 DBP 48.64 54.62 62.45 75.00 74.00 7 MAP 62.33 69.00 77.00 60.00 60.00 7 HR 61.00 62.00 70.00 30.35 30.43 7 Skin Temp 1 40.97 40.84 41.18 30.76 30.95 7 Skin Temp 2 40.81 40.76 41.12 31.34 31.30 7 Skin Temp 3 31.15 31.12 31.15
3.50 4.00 7 Bursts 4.00 11.00 9.03 287.48 358.92 7 Total Activity 1197.70 5481.90 5283.84
35.04 35.21 7 Muscle Temp 37.27 37.31 37.32 0 0 7 RPE 6 0 13
53.27 57.67 7 REN TAP 52.58 49.95 48.90 0.71 0.78 7 REN CON 0.79 0.72 0.64 1.88 1.99 7 CBF 2.11 2.31 2.78 0.03 0.03 7 CVC 0.034 0.033 0.036
125.60 124.70 8 SBP 137.20 134.70 144.60 57.99 58.49 8 DBP 81.45 83.18 92.37 75.00 74.00 8 MAP 100.67 101.00 110.00 60.00 60.00 8 HR 65.67 67.00 73.00 30.35 30.43 8 Skin Temp 1 39.43 39.62 39.96 30.76 30.95 8 Skin Temp 2 40.42 40.39 40.83 31.34 31.30 8 Skin Temp 3 27.06 27.16 27.07
3.50 4.00 8 Bursts 6.17 12.00 10.30 287.48 358.92 8 Total Activity 717.45 1319.39 1160.51
35.04 35.21 8 Muscle Temp 38.49 38.51 38.51 0 0 8 RPE 6 0 12
53.27 57.67 8 REN TAP 54.77 47.90 46.05 0.71 0.78 8 REN CON 0.54 0.47 0.42 1.88 1.99 8 CBF 1.47 1.55 1.31 0.03 0.03 8 CVC 0.015 0.015 0.012
114.10 106.70 9 SBP 105.53 108.10 117.80 61.67 54.64 9 DBP 67.40 67.24 74.53 61.00 56.00 9 MAP 81.67 81.00 92.00 45.00 45.00 9 HR 55.67 62.00 60.00 30.76 30.69 9 Skin Temp 1 40.48 40.27 40.41
6.54 6.70 9 Skin Temp 2 40.25 40.02 40.09 31.57 31.47 9 Skin Temp 3 32.22 32.23 32.23
6.50 5.50 9 Bursts 0.50 1.00 0.00 1786.77 1259.08 9 Total Activity 83.53 403.20 0.00
35.98 35.97 9 Muscle Temp 38.83 38.76 38.76 0 0 9 RPE 6 0 12
49.35 49.65 9 REN TAP 57.52 57.70 58.80 0.81 0.89 9 REN CON 0.70 0.71 0.64 1.56 1.57 9 CBF 1.87 1.92 2.48 0.03 0.03 9 CVC 0.023 0.024 0.027

145
YOUNG HEAT Subject Variable 20-40% 40-60% 60-80% 80-100%
7 SBP 127.90 130.50 141.80 150.40 7 DBP 68.33 70.89 79.33 82.87 7 MAP 85.00 90.00 99.00 106.00 7 HR 73.00 75.00 76.00 86.00 7 Skin Temp 1 41.35 41.44 41.52 41.61 7 Skin Temp 2 41.27 41.37 41.49 41.65 7 Skin Temp 3 31.18 31.21 31.26 31.32 7 Bursts 9.03 10.32 10.32 15.48 7 Total Activity 7564.17 12033.12 13360.66 25161.97 7 Muscle Temp 37.37 37.43 37.43 37.43 7 RPE 14 18 18 18 7 REN TAP 46.10 45.40 44.00 42.60 7 REN CON 0.54 0.50 0.44 0.40 7 CBF 2.33 2.48 2.83 3.41 7 CVC 0.027 0.028 0.029 0.032 8 SBP 154.80 163.70 169.70 184.10 8 DBP 100.70 104.80 109.70 117.40 8 MAP 122.00 129.00 135.00 145.00 8 HR 75.00 79.00 78.00 71.00 8 Skin Temp 1 40.09 40.19 40.30 40.35 8 Skin Temp 2 41.18 41.30 41.40 41.63 8 Skin Temp 3 27.14 27.06 27.05 27.11 8 Bursts 11.33 20.60 20.60 24.72 8 Total Activity 1524.66 3617.85 4514.05 7395.61 8 Muscle Temp 38.71 38.80 38.85 38.90 8 RPE 15 18 19 19 8 REN TAP 45.50 43.95 46.40 37.20 8 REN CON 0.37 0.34 0.34 0.26 8 CBF 1.41 1.35 1.16 1.59 8 CVC 0.012 0.010 0.009 0.011 9 SBP 121.10 132.00 157.60 177.90 9 DBP 77.56 85.08 97.97 105.00 9 MAP 94.00 103.00 120.00 133.00 9 HR 74.00 75.00 73.00 83.00 9 Skin Temp 1 40.48 40.54 40.60 40.67 9 Skin Temp 2 40.13 40.14 40.14 33.50 9 Skin Temp 3 32.23 32.23 32.25 32.30 9 Bursts 1.28 7.68 11.52 12.80 9 Total Activity 488.06 2782.98 4508.80 4150.91 9 Muscle Temp 38.76 38.82 38.94 39.11 9 RPE 12 14 16 17 9 REN TAP 50.45 51.00 49.90 46.97 9 REN CON 0.54 0.50 0.42 0.35 9 CBF 2.19 1.63 1.64 1.93 9 CVC 0.023 0.016 0.014 0.015

146
YOUNG HEAT Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
7 SBP 156.50 142.80 119.50 115.50 119.10 7 DBP 86.30 72.55 54.02 51.23 53.27 7 MAP 111.00 93.00 71.00 68.00 69.00 7 HR 86.00 63.00 64.00 62.00 63.00 7 Skin Temp 1 41.63 41.72 41.10 40.15 39.66 7 Skin Temp 2 41.67 41.72 40.48 39.54 39.08 7 Skin Temp 3 31.33 31.36 31.20 31.10 31.09 7 Bursts 12.95 18.00 5.50 11.00 5.00 7 Total Activity 21574.70 12183.85 1973.95 3606.95 1908.10 7 Muscle Temp 37.43 37.55 37.55 37.64 37.66 7 RPE 18 0 0 0 0 7 REN TAP 44.70 44.00 49.95 48.43 48.20 7 REN CON 0.40 0.47 0.70 0.71 0.70 7 CBF 3.41 3.95 3.29 2.41 2.31 7 CVC 0.031 0.042 0.046 0.035 0.033 8 SBP 189.30 142.80 119.50 115.50 119.10 8 DBP 120.40 72.55 54.02 51.23 53.27 8 MAP 148.00 93.00 71.00 68.00 69.00 8 HR 67.00 63.00 64.00 62.00 63.00 8 Skin Temp 1 40.34 41.72 41.10 40.15 39.66 8 Skin Temp 2 41.68 41.72 40.48 39.54 39.08 8 Skin Temp 3 27.12 31.36 31.20 31.10 31.09 8 Bursts 22.66 18.00 5.50 11.00 5.00 8 Total Activity 7589.45 12183.85 1973.95 3606.95 1908.10 8 Muscle Temp 38.90 37.55 37.55 37.64 37.66 8 RPE 19 0 0 0 0 8 REN TAP 38.20 44.00 49.95 48.43 48.20 8 REN CON 0.26 0.47 0.70 0.71 0.70 8 CBF 1.72 3.95 3.29 2.41 2.31 8 CVC 0.012 0.042 0.046 0.035 0.033 9 SBP 181.70 166.20 9 DBP 106.40 86.09 9 MAP 136.00 115.00 9 HR 83.00 53.00 9 Skin Temp 1 40.67 40.71 40.79 40.55 40.32 9 Skin Temp 2 29.73 31.07 25.89 26.13 25.95 9 Skin Temp 3 32.32 32.35 32.14 31.98 31.89 9 Bursts 12.80 11.00 1.50 0.50 0.00 9 Total Activity 5524.74 4302.65 408.70 0.00 0.00 9 Muscle Temp 39.11 39.13 38.76 38.54 38.44 9 RPE 17 0 0 0 0 9 REN TAP 42.20 57.43 61.03 61.58 60.30 9 REN CON 0.31 0.50 0.91 0.91 0.87 9 CBF 1.64 1.59 1.41 1.55 1.34 9 CVC 0.012 0.014 0.021 0.023 0.019

147
YOUNG NORMOTHERMIA
Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
10 SBP 130.00 130.30 147.40 155.30 166.10 10 DBP 77.60 101.00 115.20 118.70 128.80 10 MAP 94.98 114.00 130.00 134.00 146.00 10 HR 61.00 62.00 68.00 72.00 67.00 10 Skin Temp 1 29.58 29.69 29.72 29.74 29.76 10 Skin Temp 2 29.04 29.13 29.14 29.14 29.16 10 Skin Temp 3 27.38 27.23 27.18 27.17 27.11 10 Bursts 1.17 0.00 1.55 6.20 10.85 10 Total Activity 284.73 0.00 248.91 1655.11 3968.00 10 Muscle Temp 32.37 32.50 32.56 32.64 32.70 10 RPE 6 0 13 13 14 10 REN TAP 42.67 39.43 38.20 37.60 37.30 10 REN CON 0.37 0.35 0.29 0.28 0.26 10 CBF 1.79 1.90 2.08 1.96 1.58 10 CVC 0.019 0.017 0.016 0.015 0.011 11 SBP 144.37 141.60 140.40 154.90 165.40 11 DBP 94.00 94.45 93.81 106.50 114.40 11 MAP 113.00 112.00 113.00 126.00 135.00 11 HR 42.67 43.00 44.00 45.00 49.00 11 Skin Temp 1 32.10 32.09 32.06 32.06 32.09 11 Skin Temp 2 31.85 31.86 31.89 31.92 31.94 11 Skin Temp 3 31.01 31.05 31.07 31.08 31.10 11 Bursts 0.33 0.00 0.91 0.91 1.82 11 Total Activity 114.47 0.00 278.02 237.72 394.74 11 Muscle Temp 33.64 33.65 33.72 33.73 33.74 11 RPE 6 0 8 11 14 11 REN TAP 39.70 38.98 36.80 36.30 36.80 11 REN CON 0.35 0.35 0.33 0.29 0.27 11 CBF 1.75 1.89 1.68 1.66 1.62 11 CVC 0.016 0.017 0.015 0.013 0.012 12 SBP 148.50 151.00 160.50 163.00 172.20 12 DBP 81.46 86.91 93.84 95.04 105.30 12 MAP 102.00 109.00 117.00 119.00 130.00 12 HR 53.33 56.00 60.00 70.00 72.00 12 Skin Temp 1 33.12 33.08 32.98 32.94 32.89 12 Skin Temp 2 32.96 32.96 32.88 32.81 32.77 12 Skin Temp 3 31.18 31.18 31.08 31.13 31.16 12 Muscle Temp 34.71 34.74 34.77 34.75 34.75 12 RPE 6 13 16 18 12 REN TAP 47.85 43.20 43.00 46.10 45.10 12 REN CON 0.47 0.40 0.37 0.39 0.35 12 CBF 3.04 2.97 3.67 4.69 4.44 12 CVC 0.030 0.027 0.031 0.039 0.034

148
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
10 SBP 167.20 164.20 160.80 158.70 147.10 10 DBP 132.90 125.00 120.70 116.00 99.46 10 MAP 148.00 146.00 140.00 132.00 116.00 10 HR 68.00 76.00 74.00 67.00 63.00 10 Skin Temp 1 29.78 29.80 29.80 29.83 29.92 10 Skin Temp 2 29.20 29.25 29.26 29.33 29.47 10 Skin Temp 3 27.11 27.10 27.11 27.08 27.01 10 Bursts 14.73 18.60 21.71 19.00 10.00 10 Total Activity 6282.18 7073.26 8554.11 7148.65 2352.43 10 Muscle Temp 32.76 32.81 32.87 32.87 32.97 10 RPE 14 17 19 0 0 10 REN TAP 32.50 34.40 34.40 37.57 41.08 10 REN CON 0.22 0.24 0.25 0.28 0.35 10 CBF 1.85 2.31 2.40 2.20 1.78 10 CVC 0.013 0.016 0.017 0.02 0.02 11 SBP 177.70 194.50 196.60 198.10 161.60 11 DBP 124.90 134.00 133.90 125.00 104.30 11 MAP 149.00 159.00 162.00 153.00 126.00 11 HR 60.00 68.00 70.00 49.00 43.00 11 Skin Temp 1 32.12 32.14 32.14 32.19 32.36 11 Skin Temp 2 31.96 31.99 31.99 32.00 32.01 11 Skin Temp 3 31.12 31.14 31.15 31.17 31.21 11 Bursts 1.82 0.91 1.82 4.00 1.00 11 Total Activity 586.32 327.81 655.62 680.35 91.13 11 Muscle Temp 33.74 33.79 33.79 33.77 34.36 11 RPE 16 19 19 0 0 11 REN TAP 36.80 34.88 36.80 38.77 39.45 11 REN CON 0.25 0.22 0.23 0.25 0.31 11 CBF 1.85 2.27 2.43 1.83 1.69 11 CVC 0.012 0.014 0.015 0.01 0.01 12 SBP 172.30 171.60 171.40 169.50 156.80 12 DBP 112.60 116.00 116.70 96.81 81.85 12 MAP 138.00 142.00 143.00 123.00 105.00 12 HR 75.00 76.00 77.00 57.00 56.00 12 Skin Temp 1 32.84 32.81 0.00 32.76 32.84 12 Skin Temp 2 32.74 32.70 32.70 32.66 32.63 12 Skin Temp 3 31.12 31.15 31.16 31.18 31.15 12 Muscle Temp 34.75 34.75 34.75 34.75 34.63 12 RPE 18 19 19 0 0 12 REN TAP 41.45 39.90 40.40 37.67 40.80 12 REN CON 0.30 0.28 0.28 0.31 0.39 12 CBF 3.28 2.85 2.93 2.22 3.19 12 CVC 0.024 0.020 0.020 0.02 0.03

149
YOUNG HEAT
Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20% 134.80 128.70 10 SBP 130.00 157.00 151.90
86.35 81.50 10 DBP 76.00 128.20 126.30 102.00 96.00 10 MAP 90.38 141.00 140.00
65.00 62.00 10 HR 62.33 62.00 70.00 29.99 30.03 10 Skin Temp 1 38.89 39.03 39.25 29.62 29.87 10 Skin Temp 2 39.24 39.26 39.40 26.98 26.98 10 Skin Temp 3 28.39 28.41 28.42
3.50 5.50 10 Bursts 1.17 4.00 1.92 630.05 1300.11 10 Total Activity 232.15 725.94 298.18
33.48 33.98 10 Muscle Temp 39.30 39.24 39.21 0 0 10 RPE 6 0 15
42.33 39.65 10 REN TAP 40.77 39.65 37.30 0.42 0.41 10 REN CON 0.30 0.28 0.27 2.10 1.41 10 CBF 1.39 1.44 1.89 0.02 0.01 10 CVC 0.015 0.010 0.013
148.70 147.40 11 SBP 143.27 152.20 139.90 98.49 96.96 11 DBP 88.03 94.14 91.31
115.00 114.00 11 MAP 108.00 116.00 111.00 42.00 40.00 11 HR 47.67 47.00 55.00 32.70 32.95 11 Skin Temp 1 39.77 39.71 39.90 31.97 32.01 11 Skin Temp 2 40.58 40.46 40.58 31.21 31.20 11 Skin Temp 3 29.59 29.57 29.57
0.00 0.50 11 Bursts 1.33 4.00 1.00 0.00 0.00 11 Total Activity 290.33 814.00 290.00
34.46 34.49 11 Muscle Temp 37.95 37.95 37.99 0 0 11 RPE 6 0 10
40.28 40.68 11 REN TAP 43.88 43.90 42.60 0.35 0.36 11 REN CON 0.41 0.38 0.38 1.49 1.56 11 CBF 2.00 1.89 2.11 0.01 0.01 11 CVC 0.018 0.016 0.019
149.00 154.20 12 SBP 142.17 154.00 165.50 75.48 77.67 12 DBP 79.17 84.84 98.24 97.00 100.00 12 MAP 99.33 106.00 121.00 56.00 54.00 12 HR 54.67 55.00 62.00 33.01 32.90 12 Skin Temp 1 39.70 39.84 40.18 32.74 32.83 12 Skin Temp 2 39.12 39.19 39.44 31.20 31.22 12 Skin Temp 3 30.22 30.08 30.07 34.96 35.25 12 Muscle Temp 39.37 39.37 39.60
0 0 12 RPE 6 0 14 47.00 47.46 12 REN TAP 50.06 46.60 37.30
0.48 0.47 12 REN CON 0.50 0.44 0.31 3.27 3.21 12 CBF 2.21 2.44 2.74 0.03 0.03 12 CVC 0.022 0.023 0.023

150
YOUNG HEAT Subject Variable 20-40% 40-60% 60-80% 80-100%
10 SBP 160.40 163.90 167.10 167.10 10 DBP 133.80 136.20 139.90 142.80 10 MAP 146.00 149.00 153.00 158.00 10 HR 71.00 75.00 78.00 70.00 10 Skin Temp 1 39.32 39.39 39.47 39.53 10 Skin Temp 2 39.45 39.49 39.53 39.57 10 Skin Temp 3 28.42 28.42 28.42 28.42 10 Bursts 1.92 7.68 19.20 26.88 10 Total Activity 1143.55 2416.97 7136.66 9549.00 10 Muscle Temp 39.21 39.25 39.25 39.34 10 RPE 15 17 17 17 10 REN TAP 34.30 35.35 32.00 31.50 10 REN CON 0.23 0.24 0.21 0.20 10 CBF 1.88 1.01 1.40 1.84 10 CVC 0.013 0.007 0.009 0.012 11 SBP 155.10 166.50 183.60 199.10 11 DBP 100.30 110.70 128.90 135.10 11 MAP 120.00 132.00 153.00 166.00 11 HR 57.00 56.00 65.00 76.00 11 Skin Temp 1 40.01 40.09 40.13 40.14 11 Skin Temp 2 40.65 40.68 40.65 40.60 11 Skin Temp 3 29.57 29.54 29.54 29.53 11 Bursts 2.38 8.33 7.14 8.33 11 Total Activity 663.78 2981.07 1789.88 2796.62 11 Muscle Temp 37.99 38.08 38.13 38.19 11 RPE 10 15 17 19 11 REN TAP 42.60 42.60 40.37 39.70 11 REN CON 0.36 0.32 0.26 0.24 11 CBF 1.73 1.50 2.19 2.03 11 CVC 0.014 0.011 0.014 0.012 12 SBP 167.50 170.80 172.20 172.90 12 DBP 101.10 110.00 115.40 118.40 12 MAP 124.00 133.00 140.00 144.00 12 HR 64.00 73.00 79.00 81.00 12 Skin Temp 1 40.29 40.38 40.45 40.47 12 Skin Temp 2 39.55 39.65 39.74 39.80 12 Skin Temp 3 30.08 30.09 30.11 30.12 12 Muscle Temp 39.60 39.76 39.76 39.85 12 RPE 14 19 19 19 12 REN TAP 41.50 43.50 37.30 37.30 12 REN CON 0.33 0.33 0.27 0.26 12 CBF 2.24 1.99 1.72 1.60 12 CVC 0.018 0.015 0.012 0.011

151
YOUNG HEAT Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
10 SBP 167.00 151.10 150.40 148.60 146.40 10 DBP 141.50 119.80 112.40 108.80 105.20 10 MAP 159.00 134.00 127.00 123.00 120.00 10 HR 68.00 72.00 65.00 64.00 64.00 10 Skin Temp 1 39.55 39.51 39.22 38.74 38.62 10 Skin Temp 2 39.59 39.65 39.74 39.59 39.41 10 Skin Temp 3 28.42 28.46 28.45 28.39 28.39 10 Bursts 34.61 17.00 5.00 3.00 4.00 10 Total Activity 12666.53 8848.79 1049.85 771.90 768.00 10 Muscle Temp 39.34 39.50 39.61 39.49 39.32 10 RPE 17 0 0 0 0 10 REN TAP 31.50 31.05 38.93 42.28 40.83 10 REN CON 0.20 0.23 0.31 0.34 0.34 10 CBF 1.84 1.56 1.83 1.59 1.39 10 CVC 0.012 0.012 0.014 0.013 0.012 11 SBP 203.30 196.60 168.70 149.70 151.00 11 DBP 137.20 125.50 102.40 94.92 95.35 11 MAP 168.00 155.00 126.00 116.00 116.00 11 HR 77.00 58.00 48.00 43.00 43.00 11 Skin Temp 1 40.15 40.20 40.00 39.27 38.94 11 Skin Temp 2 40.59 40.55 40.13 39.38 39.11 11 Skin Temp 3 29.54 29.52 29.48 29.46 29.44 11 Bursts 9.52 13.50 3.50 4.50 3.00 11 Total Activity 3298.92 5736.41 1237.00 1384.00 565.50 11 Muscle Temp 38.19 38.35 38.19 38.02 37.96 11 RPE 19 0 0 0 0 11 REN TAP 39.70 40.20 46.48 47.43 44.55 11 REN CON 0.24 0.26 0.37 0.41 0.38 11 CBF 2.06 1.97 1.85 1.33 1.64 11 CVC 0.012 0.013 0.015 0.011 0.014 12 SBP 173.00 160.90 135.80 142.40 151.60 12 DBP 119.30 96.70 74.40 75.65 78.61 12 MAP 146.00 122.00 96.00 97.00 101.00 12 HR 81.00 59.00 58.00 59.00 54.00 12 Skin Temp 1 40.47 40.31 39.31 38.53 38.32 12 Skin Temp 2 39.80 0.00 0.00 0.00 0.00 12 Skin Temp 3 30.12 30.16 30.25 30.28 30.32 12 Muscle Temp 40.06 40.20 39.83 39.38 39.16 12 RPE 19 0 0 0 0 12 REN TAP 37.30 36.80 40.07 44.30 44.36 12 REN CON 0.26 0.30 0.42 0.46 0.44 12 CBF 1.60 1.89 2.53 2.64 2.40 12 CVC 0.011 0.015 0.026 0.027 0.024

152
YOUNG NORMOTHERMIA Subject Variable BL AV ISCH 0-20% 20-40% 40-60%
13 SBP 135.10 144.90 164.80 171.50 176.90 13 DBP 70.03 70.87 90.35 93.85 97.72 13 MAP 88.00 91.00 113.00 117.00 121.00 13 HR 66.00 77.00 114.00 114.00 117.00 13 Skin Temp 1 33.11 33.05 32.96 32.90 32.87 13 Skin Temp 2 32.24 32.12 32.58 32.58 32.59 13 Skin Temp 3 31.26 31.24 31.76 31.77 31.79 13 Bursts 2.00 1.00 1.57 0.00 0.79 13 Total Activity 271.51 65.35 240.79 0.00 163.04 13 Muscle Temp 34.83 34.74 34.68 34.60 34.55 13 RPE 6 0 11 12 13 13 REN TAP 42.27 39.75 44.00 43.23 44.20 13 REN CON 0.48 0.44 0.39 0.37 0.37 13 CBF 3.58 4.35 6.85 6.08 5.89 13 CVC 0.041 0.048 0.061 0.052 0.049 14 SBP 140.70 146.50 152.00 152.80 161.00 14 DBP 88.31 87.13 92.22 92.22 96.47 14 MAP 109.00 110.00 115.00 113.00 121.00 14 HR 70.67 80.00 86.00 87.00 89.00 14 Skin Temp 1 31.61 31.50 31.49 31.48 31.47 14 Skin Temp 2 32.21 32.13 32.12 32.10 32.09 14 Skin Temp 3 31.97 31.90 31.94 31.95 32.00 14 Bursts 2.33 3.00 1.56 4.68 12.48 14 Total Activity 344.28 358.12 335.32 692.16 1872.89 14 Muscle Temp 33.67 33.57 33.61 33.62 33.63 14 RPE 6 0 13 15 17 14 REN TAP 56.94 55.88 52.83 53.67 53.65 14 REN CON 0.52 0.51 0.46 0.47 0.44 14 CBF 2.30 2.14 2.31 2.62 2.69 14 CVC 0.021 0.019 0.020 0.023 0.022 15 SBP 109.77 121.40 126.50 137.80 152.70 15 DBP 62.25 72.32 74.79 82.78 91.57 15 MAP 80.67 91.00 93.00 102.00 112.00 15 HR 82.33 97.00 104.00 108.00 112.00 15 Skin Temp 1 34.18 34.13 34.05 34.03 34.02 15 Skin Temp 2 32.52 32.53 32.50 29.45 16.31 15 Skin Temp 3 32.68 32.68 32.68 32.68 32.67 15 Bursts 3.67 10.00 6.16 6.16 7.92 15 Total Activity 456.67 975.82 720.05 879.19 835.81 15 Muscle Temp 33.94 34.04 34.15 34.18 34.20 15 RPE 6 0 12 12 14 15 REN TAP 55.06 52.13 47.85 46.75 44.40 15 REN CON 0.68 0.57 0.51 0.46 0.40 15 CBF 1.40 1.91 2.18 1.72 1.84 15 CVC 0.017 0.021 0.023 0.017 0.016

153
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
13 SBP 180.70 187.70 191.30 187.60 177.20 13 DBP 101.10 110.00 111.20 103.80 91.83 13 MAP 126.00 135.00 137.00 131.00 115.00 13 HR 120.00 121.00 120.00 102.00 84.00 13 Skin Temp 1 32.85 32.84 32.84 32.82 33.40 13 Skin Temp 2 32.58 32.58 32.58 32.56 32.59 13 Skin Temp 3 31.83 31.89 31.91 31.99 32.16 13 Bursts 0.79 6.28 7.85 5.50 5.00 13 Total Activity 99.50 728.51 933.82 488.78 555.35 13 Muscle Temp 34.51 34.50 34.49 34.37 34.44 13 RPE 13 16 16 0 0 13 REN TAP 42.43 39.30 39.30 43.25 43.98 13 REN CON 0.34 0.29 0.29 0.40 0.46 13 CBF 8.62 8.62 8.62 5.82 3.82 13 CVC 0.068 0.064 0.063 0.04 0.03 14 SBP 173.00 178.60 180.60 170.60 157.10 14 DBP 103.20 105.90 107.90 96.59 81.96 14 MAP 129.00 132.00 136.00 123.00 105.00 14 HR 92.00 93.00 95.00 79.00 77.00 14 Skin Temp 1 31.47 31.46 31.46 31.46 31.46 14 Skin Temp 2 32.08 32.07 32.06 32.05 32.20 14 Skin Temp 3 32.07 32.19 32.21 32.33 32.27 14 Bursts 17.16 17.94 14.04 15.00 3.00 14 Total Activity 2830.21 2841.00 2841.38 3076.04 470.18 14 Muscle Temp 33.63 33.64 33.56 33.56 33.70 14 RPE 19 20 20 0 0 14 REN TAP 51.80 52.40 54.90 55.33 56.48 14 REN CON 0.40 0.40 0.40 0.45 0.54 14 CBF 2.84 3.00 2.86 3.51 2.59 14 CVC 0.022 0.023 0.021 0.03 0.02 15 SBP 163.30 181.60 193.90 165.30 135.70 15 DBP 97.26 105.00 110.00 88.13 68.10 15 MAP 121.00 130.00 135.00 112.00 89.00 15 HR 114.00 120.00 117.00 78.00 70.00 15 Skin Temp 1 34.02 34.01 34.00 33.98 43.02 15 Skin Temp 2 17.65 18.34 19.15 17.78 27.27 15 Skin Temp 3 32.68 32.67 32.67 32.70 32.90 15 Bursts 16.72 22.00 19.36 9.00 0.00 15 Total Activity 2427.15 2804.66 6020.26 1449.81 0.00 15 Muscle Temp 34.23 34.28 34.28 34.21 34.42 15 RPE 18 19 19 0 0 15 REN TAP 48.40 49.00 50.10 47.30 47.85 15 REN CON 0.40 0.38 0.37 0.42 0.54 15 CBF 1.52 1.10 1.25 2.16 1.37 15 CVC 0.013 0.008 0.009 0.02 0.02

154
YOUNG HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
169.80 175.50 13 SBP 133.40 136.80 172.70 85.95 87.54 13 DBP 73.27 72.95 90.68
109.00 110.00 13 MAP 90.67 92.00 114.00 80.00 76.00 13 HR 64.33 70.00 106.00 33.77 33.74 13 Skin Temp 1 39.48 39.57 41.48 32.76 32.76 13 Skin Temp 2 40.03 40.14 42.40 32.13 32.06 13 Skin Temp 3 30.90 30.92 31.68
3.00 2.00 13 Bursts 1.83 2.00 3.40 210.48 323.07 13 Total Activity 204.45 164.38 152.32
34.63 34.72 13 Muscle Temp 37.89 38.24 38.67 0 0 13 RPE 6 0 11
45.23 45.50 13 REN TAP 45.28 43.73 45.40 0.51 0.52 13 REN CON 0.50 0.48 0.40 3.69 3.37 13 CBF 3.99 3.75 6.49 0.03 0.03 13 CVC 0.044 0.041 0.057
155.70 156.10 14 SBP 138.67 139.60 147.50 81.38 82.14 14 DBP 71.92 73.52 82.08
104.00 105.00 14 MAP 92.67 96.00 104.00 74.00 72.00 14 HR 78.33 86.00 95.00 31.57 31.64 14 Skin Temp 1 42.57 42.92 43.34 32.44 32.38 14 Skin Temp 2 40.25 40.81 41.51 32.19 32.13 14 Skin Temp 3 31.80 32.08 32.13
3.50 4.50 14 Bursts 2.33 8.00 5.95 496.58 650.19 14 Total Activity 419.23 1430.25 995.32
33.75 33.75 14 Muscle Temp 37.83 37.90 37.93 0 0 14 RPE 6 0 14
57.75 58.38 14 REN TAP 53.57 51.85 51.20 0.56 0.56 14 REN CON 0.58 0.54 0.49 2.49 2.31 14 CBF 2.88 2.43 2.94 0.02 0.02 14 CVC 0.031 0.025 0.028
131.30 124.70 15 SBP 116.23 124.20 132.90 65.12 61.48 15 DBP 59.16 65.32 72.14 85.00 80.00 15 MAP 75.67 82.00 90.00 70.00 72.00 15 HR 83.67 89.00 106.00 34.36 34.49 15 Skin Temp 1 40.57 40.77 41.18 11.99 12.12 15 Skin Temp 2 40.91 41.00 41.41 32.66 32.65 15 Skin Temp 3 31.45 31.41 31.40
0.00 0.00 15 Bursts 3.00 7.00 2.86 0.00 0.00 15 Total Activity 355.45 980.91 276.14
34.65 34.76 15 Muscle Temp 38.80 38.79 38.89 0 0 15 RPE 6 0 14
53.83 55.07 15 REN TAP 63.79 56.65 47.90 0.63 0.69 15 REN CON 0.84 0.69 0.53 1.07 1.27 15 CBF 1.52 1.67 2.46 0.01 0.02 15 CVC 0.020 0.020 0.027

155
YOUNG HEAT Subject Variable 20-40% 40-60% 60-80% 80-100%
13 SBP 173.80 182.00 190.80 196.30 13 DBP 95.54 105.00 105.50 109.40 13 MAP 120.00 129.00 133.00 140.00 13 HR 101.00 106.00 113.00 114.00 13 Skin Temp 1 41.85 42.18 42.49 42.80 13 Skin Temp 2 42.78 43.10 43.36 43.60 13 Skin Temp 3 31.67 31.67 31.71 31.77 13 Bursts 1.70 4.25 5.10 11.05 13 Total Activity 125.26 358.63 519.13 1178.67 13 Muscle Temp 38.94 39.22 39.47 39.62 13 RPE 11 13 15 17 13 REN TAP 47.10 41.15 38.40 38.40 13 REN CON 0.39 0.32 0.29 0.27 13 CBF 8.48 7.30 7.27 9.20 13 CVC 0.071 0.057 0.055 0.066 14 SBP 159.30 175.30 190.40 195.50 14 DBP 92.72 106.50 114.70 114.60 14 MAP 118.00 133.00 143.00 146.00 14 HR 98.00 108.00 110.00 103.00 14 Skin Temp 1 43.58 43.77 43.94 44.08 14 Skin Temp 2 41.83 42.11 42.37 42.60 14 Skin Temp 3 32.20 32.30 32.46 32.66 14 Bursts 8.33 16.66 20.23 20.23 14 Total Activity 1463.00 2008.81 3171.98 3171.98 14 Muscle Temp 37.93 37.99 38.09 38.27 14 RPE 14 17 18 20 14 REN TAP 44.30 45.55 43.70 43.70 14 REN CON 0.38 0.34 0.31 0.30 14 CBF 3.42 4.14 4.57 3.96 14 CVC 0.029 0.031 0.032 0.027 15 SBP 143.70 155.30 175.20 175.80 15 DBP 79.25 89.03 99.15 101.40 15 MAP 98.00 109.00 122.00 123.00 15 HR 111.00 119.00 123.00 127.00 15 Skin Temp 1 41.33 41.46 41.59 41.71 15 Skin Temp 2 34.97 31.62 31.82 41.58 15 Skin Temp 3 31.40 31.40 31.40 31.40 15 Bursts 4.29 12.88 30.06 28.61 15 Total Activity 277.84 976.36 3290.87 3368.92 15 Muscle Temp 38.89 39.03 39.18 39.18 15 RPE 14 18 19 20 15 REN TAP 47.90 50.70 52.40 50.10 15 REN CON 0.49 0.47 0.43 0.41 15 CBF 1.41 1.36 1.01 0.88 15 CVC 0.014 0.013 0.008 0.007

156
YOUNG HEAT Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
13 SBP 196.70 196.40 167.20 161.10 164.40 13 DBP 110.10 110.00 93.31 87.87 86.21 13 MAP 140.00 140.00 114.00 108.00 106.00 13 HR 116.00 105.00 83.00 77.00 67.00 13 Skin Temp 1 42.88 40.71 40.31 39.39 39.15 13 Skin Temp 2 43.66 41.38 41.08 40.08 39.82 13 Skin Temp 3 31.78 31.05 31.38 31.32 31.23 13 Bursts 13.60 16.00 3.00 4.00 0.50 13 Total Activity 1196.32 1684.67 293.40 362.09 82.60 13 Muscle Temp 39.70 39.93 39.68 38.77 38.09 13 RPE 19 0 0 0 0 13 REN TAP 38.40 38.07 46.10 40.04 44.30 13 REN CON 0.27 0.32 0.46 0.42 0.48 13 CBF 9.20 6.15 4.61 3.86 3.40 13 CVC 0.066 0.044 0.040 0.036 0.032 14 SBP 195.90 182.10 154.30 150.60 150.60 14 DBP 114.80 102.10 80.13 75.78 74.54 14 MAP 146.00 131.00 105.00 100.00 98.00 14 HR 103.00 78.00 74.00 75.00 77.00 14 Skin Temp 1 44.10 44.28 44.48 43.71 42.72 14 Skin Temp 2 42.63 42.83 42.32 41.10 40.36 14 Skin Temp 3 32.68 32.59 32.28 32.15 32.08 14 Bursts 4.76 7.00 1.00 0.50 1.00 14 Total Activity 313.59 1097.53 150.85 89.40 133.88 14 Muscle Temp 38.27 38.49 39.00 38.64 38.39 14 RPE 20 0 0 0 0 14 REN TAP 43.70 57.10 53.05 56.18 55.25 14 REN CON 0.30 0.44 0.51 0.56 0.56 14 CBF 3.36 2.69 2.54 2.63 2.90 14 CVC 0.023 0.021 0.024 0.026 0.030 15 SBP 177.60 173.80 127.30 124.30 124.20 15 DBP 103.40 90.38 65.18 61.82 60.40 15 MAP 125.00 115.00 86.00 82.00 80.00 15 HR 128.00 89.00 78.00 74.00 73.00 15 Skin Temp 1 41.74 41.93 41.74 40.86 40.56 15 Skin Temp 2 41.98 42.17 42.34 41.78 41.48 15 Skin Temp 3 31.40 31.41 31.46 31.48 31.52 15 Bursts 14.31 28.00 8.00 8.00 11.00 15 Total Activity 1721.13 0.00 0.00 0.00 0.00 15 Muscle Temp 39.18 39.38 39.55 39.31 39.10 15 RPE 20 0 0 0 0 15 REN TAP 50.10 52.40 58.93 58.40 56.95 15 REN CON 0.40 0.46 0.69 0.71 0.71 15 CBF 0.88 1.86 1.29 1.18 1.12 15 CVC 0.007 0.016 0.015 0.014 0.014

157
Appendix D
Raw Data For Young Normothermia & Cooling Studies

158
YOUNG NORMOTHERMIA Sbjt Variable BL AVE ISCH 0-20% 20-40% 40-60%
1 SBP 129.43 130.30 137.80 139.00 146.10 1 DBP 59.99 63.28 65.02 69.39 77.69 1 MAP 80.33 84.00 85.00 91.00 101.00 1 HR 66.33 69.00 76.00 82.00 90.00 1 Skin Temp 1 32.53 32.58 32.59 32.57 32.56 1 Skin Temp 2 22.60 22.56 22.52 22.43 22.46 1 Skin Temp 3 29.85 29.82 29.83 29.80 29.79 1 Bursts 5.17 4.00 3.16 9.48 17.38 1 Total Activity 1010.86 798.12 782.70 3710.35 8912.24 1 Muscle Temp 34.06 34.21 34.38 34.39 34.37 1 RPE 6 0 13 16 17 1 REN TAP 67.18 63.85 56.85 62.10 56.67 1 REN CON 0.84 0.76 0.67 0.68 0.56 1 CBF 1.13 1.13 1.21 1.21 1.29 1 CVC 0.014 0.013 0.014 0.013 0.013 2 SBP 148.00 150.50 165.90 181.90 202.50 2 DBP 115.00 117.50 125.80 137.00 153.40 2 MAP 127.67 130.00 141.00 155.00 172.00 2 HR 59.33 75.00 95.00 99.00 103.00 2 Skin Temp 1 33.80 33.72 33.74 33.74 33.71 2 Skin Temp 2 33.70 33.73 33.61 33.54 33.48 2 Skin Temp 3 31.72 31.74 31.74 31.79 31.85 2 Bursts 2.50 1.50 0.88 1.77 4.42 2 Total Activity 522.14 317.25 292.90 588.90 2551.10 2 Muscle Temp 34.07 34.20 34.19 34.21 34.24 2 RPE 6 0 8 11 15 2 REN TAP 43.44 44.43 43.70 43.70 41.05 2 REN CON 0.34 0.34 0.31 0.28 0.24 2 CBF 2.01 1.77 5.45 4.20 2.60 2 CVC 0.016 0.014 0.039 0.027 0.015 3 SBP 139.57 147.00 155.80 143.30 171.30 3 DBP 77.36 80.04 87.80 79.26 92.94 3 MAP 94.67 99.00 108.00 99.00 116.00 3 HR 67.00 68.00 84.00 96.00 87.00 3 Skin Temp 1 33.43 33.43 33.37 33.32 33.33 3 Skin Temp 2 33.89 33.92 33.95 33.97 33.99 3 Skin Temp 3 32.87 32.85 32.83 32.82 32.82 3 Bursts 0.67 0.00 0.00 2.38 5.94 3 Total Activity 248.77 0.00 0.00 1197.86 2824.33 3 Muscle Temp 36.09 36.13 36.12 36.12 36.12 3 RPE 6 0 14 14 17 3 REN TAP 68.91 69.80 62.55 62.95 63.30 3 REN CON 0.73 0.71 0.58 0.64 0.55 3 CBF 2.53 3.16 4.49 5.15 7.29 3 CVC 0.027 0.032 0.042 0.052 0.063

159
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI
1 SBP 156.80 176.70 180.60 159.50 1 DBP 86.24 98.77 101.10 82.13 1 MAP 112.00 129.00 131.00 110.00 1 HR 100.00 110.00 112.00 91.00 1 Skin Temp 1 32.56 32.56 32.56 32.58 1 Skin Temp 2 22.48 22.44 22.43 22.50 1 Skin Temp 3 29.79 29.86 29.88 29.93 1 Bursts 25.28 30.81 12.64 21.50 1 Total Activity 17541.79 28268.64 14466.98 16353.05 1 Muscle Temp 34.34 34.33 34.34 34.16 1 RPE 19 19 19 0 1 REN TAP 50.03 46.10 42.20 61.53 1 REN CON 0.45 0.36 0.32 0.56 1 CBF 1.16 1.46 2.53 1.93 1 CVC 0.010 0.011 0.019 0.018 2 SBP 220.80 248.50 246.60 187.40 2 DBP 167.20 200.50 194.70 134.10 2 MAP 186.00 213.00 207.00 160.00 2 HR 99.00 89.00 103.00 79.00 2 Skin Temp 1 33.68 33.63 33.62 33.32 2 Skin Temp 2 33.42 33.37 33.36 33.57 2 Skin Temp 3 31.93 32.03 32.05 32.12 2 Bursts 2.65 4.42 4.42 18.50 2 Total Activity 2123.50 2551.10 2551.10 16724.70 2 Muscle Temp 34.27 34.29 34.31 34.56 2 RPE 18 18 19 0 2 REN TAP 44.90 39.60 36.20 44.90 2 REN CON 0.24 0.19 0.17 0.28 2 CBF 2.12 1.81 2.06 1.66 2 CVC 0.011 0.008 0.010 0.010 3 SBP 198.50 208.50 210.80 186.90 3 DBP 110.30 113.20 113.50 103.30 3 MAP 138.00 143.00 143.00 131.00 3 HR 89.00 96.00 98.00 87.00 3 Skin Temp 1 33.34 33.36 33.37 19.87 3 Skin Temp 2 34.00 34.02 34.02 18.92 3 Skin Temp 3 32.83 32.85 32.85 31.90 3 Bursts 4.75 4.75 3.57 8.50 3 Total Activity 2265.33 3312.36 2462.64 932.50 3 Muscle Temp 36.18 36.30 36.30 26.84 3 RPE 19 20 20 0 3 REN TAP 65.90 53.65 53.65 59.47 3 REN CON 0.48 0.38 0.38 0.45 3 CBF 6.60 5.44 5.09 4.32 3 CVC 0.048 0.038 0.036 0.033

160
YOUNG COLD Rec 1 Rec 2 Rec 3 Subject Variable BL AV ISCH
124.20 128.70 143.60 1 SBP 127.47 142.30 61.42 64.39 71.48 1 DBP 60.62 69.13 81.00 84.00 94.00 1 MAP 82.00 93.00 72.00 67.00 68.00 1 HR 62.33 64.00 32.68 32.86 32.95 1 Skin Temp 1 19.59 19.43 22.56 22.58 22.54 1 Skin Temp 2 21.88 21.86 30.00 29.93 29.89 1 Skin Temp 3 30.52 30.44 18.00 16.50 15.50 1 Bursts 4.83 2.50
8489.30 7096.75 4269.03 1 Total Activity 979.95 398.69 34.64 34.35 34.61 1 Muscle Temp 27.44 27.33
0 0 0 1 RPE 6 0 70.90 71.10 68.20 1 REN TAP 66.08 60.90
0.88 0.85 0.73 1 REN CON 0.81 0.65 1.08 0.72 1.01 1 CBF 1.38 1.47
0.013 0.009 0.011 1 CVC 0.017 0.016 180.40 165.70 164.90 2 SBP 155.23 158.30 125.90 127.10 132.80 2 DBP 116.90 118.10 142.00 134.00 136.00 2 MAP 130.67 132.00
50.00 49.00 49.00 2 HR 54.67 70.00 33.46 33.54 33.53 2 Skin Temp 1 16.31 16.22 33.24 32.88 33.16 2 Skin Temp 2 21.91 21.73 32.02 31.98 31.89 2 Skin Temp 3 31.81 31.83 12.50 11.50 10.00 2 Bursts 2.50 1.50
8917.20 5861.85 4872.15 2 Total Activity 522.14 317.25 34.85 35.14 35.40 2 Muscle Temp 24.74 24.63
0 0 0 2 RPE 6 0 38.58 38.60 39.50 2 REN TAP 44.93 49.13
0.27 0.29 0.29 2 REN CON 0.34 0.37 1.17 1.33 1.29 2 CBF 1.94 4.33
0.008 0.010 0.009 2 CVC 0.015 0.033 165.30 158.90 153.80 3 SBP 147.10 152.70
88.12 81.85 78.44 3 DBP 80.94 82.05 111.00 104.00 100.00 3 MAP 100.33 103.00
72.00 62.00 61.00 3 HR 64.00 76.00 20.61 21.29 22.11 3 Skin Temp 1 19.96 19.70 19.65 20.05 20.21 3 Skin Temp 2 18.10 17.96 31.94 31.91 31.91 3 Skin Temp 3 31.87 31.88
5.50 3.00 1.50 3 Bursts 2.17 1.50 271.30 160.00 79.27 3 Total Activity 475.58 242.25
29.71 29.23 28.94 3 Muscle Temp 26.51 26.38 0 0 0 3 RPE 6 0
57.88 58.28 59.43 3 REN TAP 62.04 59.13 0.52 0.56 0.59 3 REN CON 0.62 0.57 3.29 3.15 2.92 3 CBF 2.57 3.89
0.030 0.030 0.029 3 CVC 0.026 0.038

161
YOUNG COLD Subject Variable 0-20% 20-40% 40-60% 60-80% 80-100%
1 SBP 147.50 147.00 146.70 146.40 146.90 1 DBP 72.66 74.69 77.05 76.33 75.69 1 MAP 97.00 97.00 100.00 102.00 100.00 1 HR 67.00 76.00 89.00 95.00 94.00 1 Skin Temp 1 19.38 19.35 19.34 19.34 19.35 1 Skin Temp 2 21.82 21.84 21.88 21.94 22.01 1 Skin Temp 3 30.40 30.39 30.38 30.38 30.38 1 Bursts 2.46 2.46 7.38 4.92 9.84 1 Total Activity 714.34 1857.05 4296.15 2793.20 4963.03 1 Muscle Temp 28.10 28.10 28.10 28.01 28.01 1 RPE 20 20 20 20 20 1 REN TAP 60.90 57.40 53.90 52.75 51.60 1 REN CON 0.63 0.59 0.54 0.52 0.52 1 CBF 1.05 1.53 1.35 1.18 1.18 1 CVC 0.011 0.016 0.014 0.012 0.012 2 SBP 142.30 150.30 164.40 184.70 187.70 2 DBP 109.00 114.80 127.90 154.00 145.90 2 MAP 119.00 129.00 141.00 166.00 161.00 2 HR 62.00 72.00 80.00 72.00 103.00 2 Skin Temp 1 21.68 21.63 21.67 21.66 21.66 2 Skin Temp 2 16.38 16.54 16.71 16.84 16.96 2 Skin Temp 3 31.85 31.83 31.85 31.90 31.95 2 Bursts 0.88 1.77 4.42 2.65 4.42 2 Total Activity 292.90 588.90 2551.10 2123.50 2551.10 2 Muscle Temp 24.64 24.57 24.63 24.72 24.70 2 RPE 8 8 14 17 18 2 REN TAP 44.90 44.45 44.00 42.00 44.90 2 REN CON 0.38 0.34 0.31 0.25 0.28 2 CBF 3.02 3.63 2.53 3.48 1.71 2 CVC 0.025 0.028 0.018 0.021 0.011 3 SBP 164.20 163.10 178.00 192.30 203.00 3 DBP 91.20 89.17 97.73 107.40 112.00 3 MAP 113.00 112.00 122.00 134.00 141.00 3 HR 84.00 89.00 86.00 91.00 98.00 3 Skin Temp 1 19.36 19.06 19.06 19.22 19.49 3 Skin Temp 2 18.05 18.14 18.26 18.41 18.58 3 Skin Temp 3 31.89 31.88 31.88 31.87 31.87 3 Bursts 0.00 3.57 3.57 4.75 10.70 3 Total Activity 0.00 471.05 416.97 571.71 1730.82 3 Muscle Temp 27.07 27.07 27.15 27.14 27.15 3 RPE 13 13 16 19 20 3 REN TAP 54.30 54.70 55.20 56.90 47.83 3 REN CON 0.48 0.49 0.45 0.42 0.34 3 CBF 7.45 6.61 6.98 6.10 4.99 3 CVC 0.066 0.059 0.057 0.046 0.035

162
YOUNG COLD Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
1 SBP 147.40 146.70 156.10 149.90 139.80 1 DBP 76.09 69.77 71.49 71.25 70.36 1 MAP 100.00 96.00 98.00 97.00 95.00 1 HR 95.00 79.00 68.00 65.00 67.00 1 Skin Temp 1 19.36 19.40 19.24 19.13 19.05 1 Skin Temp 2 22.02 22.02 22.13 22.15 22.16 1 Skin Temp 3 30.38 30.38 30.34 30.33 30.30 1 Bursts 4.92 10.50 12.00 11.50 8.00 1 Total Activity 2744.51 4631.30 3666.25 2881.85 1558.10 1 Muscle Temp 28.01 27.43 27.09 26.78 26.63 1 RPE 20 0 0 0 0 1 REN TAP 51.60 62.70 61.83 61.77 62.98 1 REN CON 0.52 0.65 0.63 0.64 0.66 1 CBF 1.18 0.88 1.12 1.15 1.10 1 CVC 0.012 0.009 0.011 0.012 0.012 2 SBP 185.20 159.90 174.40 158.80 151.60 2 DBP 137.80 110.40 122.40 120.00 115.70 2 MAP 155.00 133.00 136.00 125.00 120.00 2 HR 102.00 69.00 57.00 51.00 50.00 2 Skin Temp 1 21.68 17.16 17.98 18.30 18.62 2 Skin Temp 2 17.00 21.78 22.08 21.84 21.90 2 Skin Temp 3 31.97 32.00 31.90 31.84 31.81 2 Bursts 4.42 0.00 0.00 0.00 0.00 2 Total Activity 2551.10 0.00 0.00 0.00 0.00 2 Muscle Temp 24.73 24.62 24.65 25.26 25.64 2 RPE 20 0 0 0 0 2 REN TAP 44.90 54.48 48.40 46.23 47.23 2 REN CON 0.29 0.41 0.36 0.37 0.39 2 CBF 0.53 3.39 1.25 1.48 1.35 2 CVC 0.003 0.025 0.009 0.012 0.011 3 SBP 204.90 196.60 155.50 150.50 158.33 3 DBP 114.30 106.60 80.02 77.36 81.52 3 MAP 144.00 134.00 101.00 98.00 103.00 3 HR 101.00 89.00 67.00 61.00 67.67 3 Skin Temp 1 19.60 33.53 33.87 34.11 33.89 3 Skin Temp 2 18.63 33.99 34.14 34.25 34.14 3 Skin Temp 3 31.87 32.84 32.82 32.79 32.82 3 Bursts 5.94 4.50 0.50 2.00 1.67 3 Total Activity 1266.68 1860.00 189.10 930.80 679.12 3 Muscle Temp 27.15 36.26 36.84 36.83 36.74 3 RPE 20 0 0 0 0 3 REN TAP 49.80 70.67 71.88 74.13 74.50 3 REN CON 0.35 0.53 0.71 0.76 0.72 3 CBF 4.99 5.81 3.17 2.30 2.99 3 CVC 0.035 0.043 0.031 0.023 0.029

163
YOUNG NORMOTHERMIA Sbjt Variable BL AVE ISCH 0-20% 20-40% 40-60%
4 SBP 119.93 127.00 133.50 136.40 139.10 4 DBP 69.40 71.11 75.61 78.76 80.91 4 MAP 81.00 84.00 90.00 93.00 98.00 4 HR 51.00 62.00 66.00 66.00 67.00 4 Skin Temp 1 31.40 31.34 31.29 31.30 31.31 4 Skin Temp 2 31.87 31.80 0.00 0.00 0.00 4 Skin Temp 3 30.89 30.84 30.81 30.85 30.82 4 Bursts 1.67 1.50 1.96 1.96 6.54 4 Total Activity 296.05 224.25 422.92 261.67 1169.82 4 Muscle Temp 34.87 34.87 34.86 34.85 34.84 4 RPE 6 0 12 12 14 4 REN TAP 72.00 70.00 62.80 62.80 60.15 4 REN CON 0.89 0.83 0.70 0.68 0.61 4 CBF 1.58 2.51 1.91 1.80 1.54 4 CVC 0.019 0.030 0.021 0.019 0.016 5 SBP 116.63 113.00 123.10 128.60 144.70 5 DBP 86.07 84.48 92.03 95.93 102.60 5 MAP 100.00 97.00 106.00 111.00 121.00 5 HR 63.00 71.00 78.00 91.00 94.00 5 Skin Temp 1 31.67 31.66 31.62 31.58 31.58 5 Skin Temp 2 31.91 31.94 31.96 31.99 32.04 5 Skin Temp 3 32.35 32.24 32.19 32.18 32.17 5 Bursts 0.67 1.00 0.00 0.00 0.66 5 Total Activity 87.23 82.82 0.00 0.00 239.14 5 Muscle Temp 33.41 33.49 33.55 33.55 33.59 5 RPE 6 0 11 11 12 5 REN TAP 47.73 51.23 45.90 45.13 43.70 5 REN CON 0.48 0.53 0.43 0.41 0.36 5 CBF 2.51 3.25 2.77 3.05 2.38 5 CVC 0.025 0.034 0.026 0.027 0.020 6 SBP 110.47 114.10 119.20 132.40 149.10 6 DBP 79.34 80.85 84.29 92.20 101.30 6 MAP 93.33 96.00 101.00 111.00 123.00 6 HR 56.67 63.00 83.00 73.00 73.00 6 Skin Temp 1 30.06 30.07 30.08 30.09 30.09 6 Skin Temp 2 29.99 30.03 30.04 30.03 30.04 6 Skin Temp 3 29.92 29.91 29.94 29.94 29.92 6 Bursts 1.00 1.00 0.82 0.82 1.64 6 Total Activity 175.64 181.00 65.13 109.80 239.33 6 Muscle Temp 33.09 33.18 33.45 33.56 33.59 6 RPE 6 0 12 13 14 6 REN TAP 69.89 64.23 67.00 68.40 66.35 6 REN CON 0.75 0.67 0.66 0.62 0.54 6 CBF 2.13 2.52 2.69 2.45 2.28 6 CVC 0.023 0.026 0.027 0.022 0.019

164
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI
4 SBP 146.20 150.90 153.20 147.50 4 DBP 85.92 86.37 88.71 76.15 4 MAP 106.00 110.00 110.00 97.00 4 HR 70.00 75.00 75.00 59.00 4 Skin Temp 1 31.31 31.32 31.32 31.36 4 Skin Temp 2 0.00 0.00 31.76 31.77 4 Skin Temp 3 30.83 30.86 30.85 30.84 4 Bursts 5.89 9.81 3.92 5.00 4 Total Activity 1029.80 1832.30 973.72 794.59 4 Muscle Temp 34.84 34.84 34.84 34.76 4 RPE 17 18 19 0 4 REN TAP 71.80 79.00 79.90 72.70 4 REN CON 0.68 0.72 0.73 0.75 4 CBF 1.14 1.22 1.52 1.63 4 CVC 0.011 0.011 0.014 0.017 5 SBP 149.90 154.40 161.60 161.30 5 DBP 105.80 109.70 115.90 111.30 5 MAP 125.00 129.00 136.00 132.00 5 HR 99.00 98.00 98.00 86.00 5 Skin Temp 1 31.62 31.66 31.68 31.76 5 Skin Temp 2 32.08 32.11 32.12 32.16 5 Skin Temp 3 32.18 32.19 32.18 32.19 5 Bursts 2.64 2.64 1.32 2.50 5 Total Activity 183.38 202.49 105.85 351.85 5 Muscle Temp 33.60 33.63 33.64 33.70 5 RPE 14 15 16 0 5 REN TAP 44.30 43.70 43.70 44.33 5 REN CON 0.35 0.34 0.32 0.34 5 CBF 2.58 2.43 2.36 3.07 5 CVC 0.021 0.019 0.017 0.023 6 SBP 158.50 159.10 158.20 145.40 6 DBP 102.40 105.10 102.50 85.80 6 MAP 127.00 129.00 126.00 108.00 6 HR 77.00 82.00 70.00 52.00 6 Skin Temp 1 30.07 30.05 30.05 30.08 6 Skin Temp 2 30.07 30.08 30.08 30.10 6 Skin Temp 3 29.93 29.95 29.95 29.89 6 Bursts 8.19 13.92 5.73 9.50 6 Total Activity 1099.97 3252.01 1360.40 2203.73 6 Muscle Temp 33.62 33.62 33.62 33.36 6 RPE 15 16 16 0 6 REN TAP 64.73 67.03 69.80 82.33 6 REN CON 0.51 0.52 0.55 0.76 6 CBF 2.35 2.82 1.70 1.90 6 CVC 0.018 0.022 0.013 0.018

165
YOUNG COLD Rec 1 Rec 2 Rec 3 Subject Variable BL AV ISCH
134.40 137.20 141.90 4 SBP 114.07 114.00 62.16 67.18 73.43 4 DBP 83.61 84.02 80.00 85.00 90.00 4 MAP 95.33 94.00 56.00 53.00 56.00 4 HR 49.67 55.00 31.44 31.58 31.75 4 Skin Temp 1 14.77 14.54 31.72 31.69 31.82 4 Skin Temp 2 15.79 15.67 30.80 30.77 30.79 4 Skin Temp 3 29.47 29.51
1.50 3.00 4.00 4 Bursts 1.33 3.00 148.40 551.82 626.93 4 Total Activity 277.42 341.20
34.76 34.93 35.02 4 Muscle Temp 24.56 24.34 0 0 0 4 RPE 6 0
74.80 70.72 73.60 4 REN TAP 71.95 74.68 0.94 0.83 0.82 4 REN CON 0.75 0.79 1.81 1.46 1.29 4 CBF 1.19 1.33
0.023 0.017 0.014 4 CVC 0.012 0.014 147.80 137.50 133.00 5 SBP 128.93 116.50
96.51 90.13 89.51 5 DBP 87.14 84.22 114.00 108.00 106.00 5 MAP 104.33 98.00
78.00 77.00 78.00 5 HR 61.33 76.00 32.01 32.16 32.40 5 Skin Temp 1 24.30 23.87 32.24 32.27 32.32 5 Skin Temp 2 24.38 23.43 32.20 32.21 32.20 5 Skin Temp 3 31.12 31.05
1.50 0.50 0.00 5 Bursts 0.33 0.50 83.74 0.00 0.00 5 Total Activity 25.96 20.04 34.25 34.37 34.46 5 Muscle Temp 27.54 27.54
0 0 0 5 RPE 6 0 42.55 45.40 45.57 5 REN TAP 46.98 50.67
0.37 0.42 0.43 5 REN CON 0.45 0.52 3.11 3.43 3.22 5 CBF 2.90 3.41
0.027 0.032 0.030 5 CVC 0.028 0.035 110.10 99.70 96.50 6 SBP 107.93 111.60
63.44 61.54 61.24 6 DBP 80.77 84.83 82.00 78.00 78.00 6 MAP 94.67 98.00 48.00 48.00 46.00 6 HR 61.33 68.00 30.19 30.24 30.30 6 Skin Temp 1 14.08 14.14 30.18 30.30 30.32 6 Skin Temp 2 14.21 14.28 29.74 29.68 29.64 6 Skin Temp 3 29.91 30.10
1.50 2.50 1.00 6 Bursts 1.17 1.00 189.65 370.75 34.51 6 Total Activity 203.35 254.20
33.42 33.50 33.60 6 Muscle Temp 26.35 26.38 0 0 0 6 RPE 6 0
77.73 76.30 69.28 6 REN TAP 67.45 62.70 0.95 0.98 0.89 6 REN CON 0.71 0.64 2.06 1.88 2.06 6 CBF 2.69 2.60
0.025 0.024 0.026 6 CVC 0.028 0.027

166
YOUNG COLD Subject Variable 0-20% 20-40% 40-60% 60-80% 80-100%
4 SBP 114.20 118.40 123.60 128.20 134.60 4 DBP 85.35 88.89 93.52 96.82 99.50 4 MAP 96.00 100.00 106.00 109.00 113.00 4 HR 71.00 63.00 62.00 66.00 68.00 4 Skin Temp 1 14.35 14.36 14.44 14.51 14.58 4 Skin Temp 2 0.00 0.00 0.00 0.00 0.00 4 Skin Temp 3 29.53 29.48 29.49 29.46 0.94 4 Bursts 0.00 3.64 4.85 7.28 8.49 4 Total Activity 0.00 683.43 1064.43 1087.01 1820.41 4 Muscle Temp 24.31 24.19 24.12 24.09 24.08 4 RPE 13 14 17 19 19 4 REN TAP 71.80 71.80 76.20 81.70 77.20 4 REN CON 0.75 0.72 0.72 0.75 0.68 4 CBF 1.70 1.42 1.26 0.96 1.20 4 CVC 0.018 0.014 0.012 0.009 0.011 5 SBP 136.90 144.80 147.40 152.40 154.90 5 DBP 96.45 97.56 95.59 100.20 105.00 5 MAP 115.00 116.00 116.00 121.00 126.00 5 HR 85.00 92.00 93.00 98.00 102.00 5 Skin Temp 1 23.62 23.56 23.58 23.60 23.63 5 Skin Temp 2 23.22 23.28 23.26 23.20 23.18 5 Skin Temp 3 31.06 31.05 31.06 31.08 31.11 5 Bursts 0.52 0.52 1.05 0.52 1.57 5 Total Activity 51.22 24.50 60.55 49.45 125.24 5 Muscle Temp 27.54 27.64 27.62 27.61 27.67 5 RPE 10 10 10 12 16 5 REN TAP 51.60 46.90 47.35 47.80 43.87 5 REN CON 0.45 0.40 0.41 0.40 0.35 5 CBF 3.28 3.21 2.81 3.00 3.08 5 CVC 0.028 0.028 0.024 0.025 0.024 6 SBP 114.30 121.80 133.00 148.70 161.10 6 DBP 87.51 90.83 94.06 100.60 101.60 6 MAP 101.00 104.00 111.00 120.00 126.00 6 HR 40.00 30.00 30.00 34.00 39.00 6 Skin Temp 1 14.44 14.67 14.87 15.04 15.21 6 Skin Temp 2 14.43 14.57 14.73 14.92 15.16 6 Skin Temp 3 30.15 30.19 30.21 30.23 30.26 6 Bursts 0.80 1.61 5.62 7.22 10.43 6 Total Activity 107.78 258.59 658.07 954.93 2112.58 6 Muscle Temp 27.21 27.48 27.59 27.70 27.74 6 RPE 11 12 14 15 16 6 REN TAP 65.50 65.50 63.60 61.33 67.40 6 REN CON 0.65 0.63 0.57 0.51 0.53 6 CBF 2.73 2.88 2.22 3.27 2.90 6 CVC 0.027 0.028 0.020 0.027 0.023

167
YOUNG COLD Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
4 SBP 134.80 126.30 122.60 117.70 115.60 4 DBP 96.87 89.66 86.58 82.69 81.46 4 MAP 112.00 103.00 97.00 92.00 91.00 4 HR 69.00 57.00 52.00 49.00 48.00 4 Skin Temp 1 14.58 14.62 14.55 14.63 14.89 4 Skin Temp 2 15.48 15.48 15.23 15.25 15.82 4 Skin Temp 3 29.50 29.54 29.51 29.55 29.50 4 Bursts 6.06 8.00 5.00 3.50 0.50 4 Total Activity 1439.98 1655.46 932.70 450.15 0.00 4 Muscle Temp 24.08 23.81 23.61 23.54 23.59 4 RPE 19 0 0 0 0 4 REN TAP 77.20 86.56 79.60 74.05 77.20 4 REN CON 0.69 0.84 0.82 0.80 0.85 4 CBF 1.20 1.13 1.21 1.02 1.19 4 CVC 0.011 0.011 0.012 0.011 0.013 5 SBP 155.30 158.80 160.30 146.90 137.50 5 DBP 105.90 97.72 99.68 92.42 91.20 5 MAP 127.00 120.00 121.00 111.00 109.00 5 HR 100.00 82.00 67.00 58.00 56.00 5 Skin Temp 1 23.63 23.70 24.01 24.17 24.41 5 Skin Temp 2 23.17 23.43 23.80 23.87 23.66 5 Skin Temp 3 31.13 31.10 31.05 31.03 31.04 5 Bursts 1.05 2.50 3.50 1.50 2.00 5 Total Activity 79.93 298.38 386.88 177.37 164.67 5 Muscle Temp 27.67 27.72 27.91 28.21 28.30 5 RPE 16 0 0 0 0 5 REN TAP 43.80 45.20 45.57 46.00 43.35 5 REN CON 0.34 0.38 0.38 0.41 0.40 5 CBF 2.98 2.64 2.81 2.56 2.49 5 CVC 0.023 0.022 0.023 0.023 0.023 6 SBP 162.80 141.90 115.00 95.83 89.75 6 DBP 99.30 86.87 70.38 63.64 63.46 6 MAP 124.00 107.00 88.00 77.00 75.00 6 HR 76.00 50.00 47.00 47.00 45.00 6 Skin Temp 1 15.26 15.53 16.11 16.27 16.39 6 Skin Temp 2 15.23 15.48 15.91 16.11 16.31 6 Skin Temp 3 30.27 30.31 30.22 30.16 30.17 6 Bursts 6.42 12.50 5.50 1.50 3.00 6 Total Activity 1533.46 3266.20 901.94 158.21 361.00 6 Muscle Temp 27.66 26.75 26.57 26.65 26.98 6 RPE 18 0 0 0 0 6 REN TAP 67.40 72.37 67.85 70.68 64.08 6 REN CON 0.54 0.68 0.77 0.92 0.85 6 CBF 2.95 2.65 2.12 2.24 1.98 6 CVC 0.024 0.025 0.024 0.029 0.026

168
YOUNG NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
7 SBP 150.27 155.40 154.40 154.70 152.70 7 DBP 80.72 85.23 84.56 87.79 89.63 7 MAP 104.00 109.00 108.00 112.00 114.00 7 HR 56.33 60.00 63.00 63.00 65.00 7 Skin Temp 1 30.64 30.64 30.59 30.54 30.51 7 Skin Temp 2 31.89 31.90 31.90 31.86 31.85 7 Skin Temp 3 29.94 29.90 29.89 29.89 29.90 7 Bursts 2.83 2.00 2.91 4.85 3.88 7 Total Activity 376.62 400.50 514.87 616.11 1148.71
7 Muscle Temp 33.31 33.04 33.27 33.06 33.04
7 RPE 6 0 15 14 14 7 REN TAP 60.86 57.60 55.90 57.03 55.90 7 REN CON 0.59 0.53 0.52 0.51 0.49 7 CBF 1.72 1.86 2.02 2.11 1.96 7 CVC 0.017 0.017 0.019 0.019 0.017 8 SBP 120.93 121.90 134.70 140.70 148.60 8 DBP 70.42 71.73 79.70 83.69 91.64 8 MAP 87.67 89.00 99.00 104.00 115.00 8 HR 52.00 54.00 71.00 63.00 62.00 8 Skin Temp 1 30.11 30.10 30.09 30.08 30.07 8 Skin Temp 2 30.90 30.92 30.99 31.07 31.08 8 Skin Temp 3 30.71 30.72 30.70 30.69 30.67
8 Muscle Temp 33.61 33.57 33.64 33.64 33.71
8 RPE 6 0 11 16 18 8 REN TAP 46.18 42.63 38.13 39.70 38.15 8 REN CON 0.53 0.48 0.39 0.38 0.33 8 CBF 2.27 2.51 2.78 2.14 2.26 8 CVC 0.026 0.028 0.028 0.021 0.020 9 SBP 146.80 153.40 164.30 167.90 171.80 9 DBP 72.42 75.52 84.32 88.12 94.48 9 MAP 92.00 97.00 107.00 111.00 119.00 9 HR 56.00 59.00 64.00 71.00 72.00 9 Skin Temp 1 33.91 33.92 33.76 33.66 33.59 9 Skin Temp 2 35.31 35.25 34.99 34.74 34.59 9 Skin Temp 3 31.69 31.61 22.14 22.11 22.10
9 Muscle Temp 35.17 35.42 35.49 35.51 35.53
9 RPE 6 0 10 13 15 9 REN TAP 52.56 55.03 50.50 50.80 50.30 9 REN CON 0.57 0.57 0.47 0.46 0.42 9 CBF 2.86 3.02 3.45 3.79 3.36 9 CVC 0.031 0.031 0.032 0.034 0.028

169
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI
7 SBP 163.20 175.00 176.80 163.60 7 DBP 97.78 104.60 105.80 96.37 7 MAP 123.00 132.00 133.00 123.00 7 HR 68.00 76.00 78.00 66.00 7 Skin Temp 1 30.49 30.47 30.47 30.46 7 Skin Temp 2 31.84 31.83 31.83 31.82 7 Skin Temp 3 29.90 29.88 29.87 29.84 7 Bursts 4.85 3.88 0.97 11.00 7 Total Activity 1198.45 1002.78 172.11 2193.70 7 Muscle Temp 33.04 32.96 32.96 32.64 7 RPE 14 18 18 0 7 REN TAP 56.75 47.45 42.40 54.28 7 REN CON 0.46 0.36 0.32 0.44 7 CBF 1.96 2.20 2.20 1.82 7 CVC 0.016 0.017 0.017 0.015 8 SBP 160.80 167.40 170.80 161.20 8 DBP 100.20 106.20 110.00 95.03 8 MAP 123.00 130.00 134.00 119.00 8 HR 67.00 66.00 65.00 51.00 8 Skin Temp 1 30.07 30.06 30.06 30.12 8 Skin Temp 2 31.11 31.12 31.11 30.82 8 Skin Temp 3 30.66 30.66 30.66 30.66 8 Muscle Temp 33.73 33.76 33.76 33.89 8 RPE 19 19 19 0 8 REN TAP 42.30 42.85 42.30 53.63 8 REN CON 0.34 0.33 0.32 0.45 8 CBF 2.57 2.06 2.13 2.50 8 CVC 0.021 0.016 0.016 0.021 9 SBP 176.80 182.20 183.30 157.20 9 DBP 102.00 106.50 108.90 87.48 9 MAP 127.00 136.00 138.00 113.00 9 HR 76.00 71.00 74.00 57.00 9 Skin Temp 1 33.52 33.43 33.41 33.34 9 Skin Temp 2 34.44 34.30 34.27 34.27 9 Skin Temp 3 22.09 22.11 22.11 31.45 9 Muscle Temp 35.56 35.57 35.57 35.44 9 RPE 18 19 19 0 9 REN TAP 44.95 45.00 45.00 36.00 9 REN CON 0.35 0.33 0.33 0.32 9 CBF 2.19 2.06 2.06 2.45 9 CVC 0.017 0.015 0.015 0.022

170
YOUNG COLD
Rec 1 Rec 2 Rec 3 Subject Variable BL AVE ISCH
148.00 148.30 140.80 7 SBP 145.80 145.00 79.55 81.90 72.74 7 DBP 79.37 76.53
103.00 104.00 96.00 7 MAP 102.00 100.00 60.00 56.00 56.00 7 HR 54.33 59.00 30.55 30.76 30.90 7 Skin Temp 1 19.02 19.59 31.91 32.17 32.33 7 Skin Temp 2 23.20 23.43 29.73 29.70 29.72 7 Skin Temp 3 30.05 30.07
4.50 6.50 5.00 7 Bursts 1.67 2.00 815.96 1043.26 540.14 7 Total Activity 218.31 272.69
32.85 33.10 33.10 7 Muscle Temp 24.61 25.73 0 0 0 7 RPE 6 0
54.63 59.30 59.30 7 REN TAP 61.00 60.15 0.53 0.57 0.62 7 REN CON 0.60 0.60 1.69 1.37 1.43 7 CBF 1.44 1.71
0.016 0.013 0.015 7 CVC 0.014 0.017 146.20 138.00 135.60 8 SBP 116.60 116.70
78.24 72.52 70.23 8 DBP 70.21 69.87 100.00 93.00 91.00 8 MAP 86.33 86.00
47.00 47.00 47.00 8 HR 45.33 45.00 30.22 30.22 30.23 8 Skin Temp 1 11.39 13.51 30.77 30.86 30.90 8 Skin Temp 2 17.33 18.04 30.63 30.60 30.57 8 Skin Temp 3 30.02 30.02 33.90 34.28 34.48 8 Muscle Temp 25.68 25.35
0 0 0 8 RPE 6 0 43.03 43.13 47.10 8 REN TAP 42.42 40.75
0.43 0.46 0.52 8 REN CON 0.49 0.47 2.09 2.13 2.26 8 CBF 1.66 1.66
0.021 0.023 0.025 8 CVC 0.019 0.019 142.10 142.80 142.60 9 SBP 130.90 137.60
69.37 66.41 66.49 9 DBP 64.37 68.36 91.00 88.00 87.00 9 MAP 82.00 87.00 57.00 57.00 56.00 9 HR 57.67 57.00 33.42 33.66 33.69 9 Skin Temp 1 18.05 18.17 34.47 34.81 34.87 9 Skin Temp 2 19.51 19.82 31.03 30.96 30.92 9 Skin Temp 3 31.69 31.61 35.41 35.47 35.41 9 Muscle Temp 24.31 24.42
0 0 0 9 RPE 6 0 45.00 48.70 49.07 9 REN TAP 46.07 52.90
0.49 0.55 0.56 9 REN CON 0.56 0.61 2.92 3.02 3.07 9 CBF 3.02 3.11
0.032 0.034 0.035 9 CVC 0.037 0.036

171
YOUNG COLD Subject Variable 0-20% 20-40% 40-60% 60-80% 80-100%
7 SBP 148.40 152.40 151.80 159.00 161.30 7 DBP 79.85 86.84 87.39 93.89 97.18 7 MAP 103.00 110.00 112.00 119.00 122.00 7 HR 57.00 60.00 62.00 68.00 72.00 7 Skin Temp 1 19.91 19.97 20.01 20.07 20.19 7 Skin Temp 2 23.64 23.72 23.71 23.68 23.67 7 Skin Temp 3 30.09 30.10 30.11 30.11 30.12 7 Bursts 2.11 2.11 4.21 7.37 6.32 7 Total Activity 561.68 242.12 832.84 957.01 1180.00 7 Muscle Temp 25.30 25.22 25.22 25.24 25.24 7 RPE 15 17 19 19 19 7 REN TAP 59.30 59.30 61.00 60.15 60.15 7 REN CON 0.58 0.54 0.54 0.51 0.49 7 CBF 1.55 1.58 1.65 1.86 1.75 7 CVC 0.015 0.014 0.015 0.016 0.014 8 SBP 122.70 128.20 140.40 150.60 155.10 8 DBP 75.79 80.36 88.80 98.59 103.90 8 MAP 92.00 100.00 107.00 118.00 126.00 8 HR 47.00 47.00 54.00 59.00 76.00 8 Skin Temp 1 14.72 15.69 16.54 17.28 17.84 8 Skin Temp 2 18.55 18.95 19.33 19.68 19.93 8 Skin Temp 3 29.99 30.00 30.00 29.99 29.97 8 Muscle Temp 25.28 25.38 25.45 25.50 25.59 8 RPE 9 13 14 17 18 8 REN TAP 40.75 40.75 42.85 43.40 43.93 8 REN CON 0.44 0.41 0.40 0.37 0.35 8 CBF 1.78 1.96 1.97 1.59 1.56 8 CVC 0.019 0.020 0.018 0.014 0.012 9 SBP 145.70 149.80 157.20 171.60 177.10 9 DBP 73.58 76.70 83.52 96.22 104.80 9 MAP 93.00 97.00 104.00 119.00 130.00 9 HR 61.00 63.00 66.00 75.00 88.00 9 Skin Temp 1 18.12 17.91 17.77 17.60 17.47 9 Skin Temp 2 19.62 19.28 19.05 18.83 18.74 9 Skin Temp 3 31.62 31.59 31.61 31.60 31.56 9 Muscle Temp 24.51 24.66 24.71 24.76 24.79 9 RPE 9 11 13 14 16 9 REN TAP 50.10 41.27 44.47 46.05 42.88 9 REN CON 0.54 0.43 0.43 0.39 0.33 9 CBF 3.25 3.00 3.36 2.60 2.07 9 CVC 0.035 0.031 0.032 0.022 0.016

172
YOUNG COLD Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
7 SBP 161.50 153.80 137.20 137.80 142.30 7 DBP 97.37 92.33 76.98 72.91 73.14 7 MAP 123.00 117.00 100.00 96.00 98.00 7 HR 72.00 67.00 57.00 52.00 55.00 7 Skin Temp 1 20.20 20.49 21.18 21.54 21.86 7 Skin Temp 2 23.67 23.74 24.12 24.42 24.74 7 Skin Temp 3 30.13 30.14 30.08 30.01 29.97 7 Bursts 3.16 7.00 6.00 2.00 1.50 7 Total Activity 289.68 1497.20 1263.22 393.95 184.50 7 Muscle Temp 25.24 25.12 26.19 26.21 26.09 7 RPE 19 0 0 0 0 7 REN TAP 57.60 58.45 57.60 62.70 62.28 7 REN CON 0.47 0.50 0.58 0.65 0.64 7 CBF 1.75 1.72 1.38 1.36 1.47 7 CVC 0.014 0.015 0.014 0.014 0.015 8 SBP 160.00 148.70 138.30 127.20 115.00 8 DBP 106.80 85.81 76.94 70.99 66.75 8 MAP 130.00 106.00 97.00 89.00 82.00 8 HR 79.00 47.00 45.00 43.00 45.00 8 Skin Temp 1 17.94 18.33 18.73 18.88 19.12 8 Skin Temp 2 19.97 20.11 20.49 20.66 20.90 8 Skin Temp 3 29.97 29.96 29.95 29.94 29.91 8 Muscle Temp 25.83 25.44 25.68 26.64 27.39 8 RPE 19 0 0 0 0 8 REN TAP 45.00 44.25 39.45 41.55 42.85 8 REN CON 0.35 0.42 0.41 0.47 0.52 8 CBF 1.26 1.82 1.62 1.80 1.82 8 CVC 0.010 0.017 0.017 0.020 0.022 9 SBP 178.60 160.10 142.10 138.90 142.40 9 DBP 107.10 84.34 68.71 62.91 63.68 9 MAP 134.00 109.00 91.00 83.00 85.00 9 HR 94.00 58.00 52.00 53.00 56.00 9 Skin Temp 1 17.44 17.53 18.30 18.86 19.17 9 Skin Temp 2 18.72 18.89 19.78 21.02 21.94 9 Skin Temp 3 31.56 31.45 31.03 30.96 30.92 9 Muscle Temp 24.76 24.56 24.38 24.64 24.85 9 RPE 17 0 0 0 0 9 REN TAP 41.30 39.20 41.25 52.90 48.70 9 REN CON 0.31 0.36 0.45 0.64 0.57 9 CBF 2.10 2.63 2.70 2.36 2.36 9 CVC 0.016 0.024 0.030 0.028 0.028

173
YOUNG NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
10 SBP 168.43 169.00 164.80 171.50 176.90 10 DBP 89.96 93.86 90.35 93.85 97.72 10 MAP 113.00 116.00 113.00 117.00 121.00 10 HR 75.00 89.00 114.00 114.00 117.00 10 Skin Temp 1 33.13 33.08 32.96 32.90 32.87 10 Skin Temp 2 32.56 32.56 32.58 32.58 32.59 10 Skin Temp 3 31.78 31.75 31.76 31.77 31.79 10 Bursts 2.00 0.50 1.58 0.00 0.79 10 Total Activity 271.51 32.68 242.05 0.00 163.89
10 Muscle Temp 34.83 34.74 34.68 34.60 34.55
10 RPE 6 0 11 12 13 10 REN TAP 42.27 39.75 44.00 43.23 44.20 10 REN CON 0.48 0.44 0.39 0.37 0.37 11 SBP 138.07 142.90 148.90 151.70 155.70 11 DBP 75.98 81.65 85.09 87.60 88.43 11 MAP 97.33 105.00 110.00 111.00 113.00 11 HR 77.67 86.00 92.00 98.00 96.00 11 Skin Temp 1 32.32 32.84 32.99 33.06 33.12 11 Skin Temp 2 31.98 32.40 32.52 32.58 32.63 11 Skin Temp 3 31.78 31.73 31.72 31.72 31.73 11 Bursts 1.83 8.50 8.16 6.34 9.06 11 Total Activity 380.85 1829.50 2048.79 1299.28 3249.70
11 Muscle Temp 35.49 35.41 35.45 35.44 35.46
11 RPE 6 0 12 14 17 11 REN TAP 45.15 43.45 44.30 43.95 43.45 11 REN CON 0.43 0.41 0.40 0.40 0.38 11 CBF 2.59 1.44 2.07 2.47 1.71 11 CVC 0.027 0.014 0.019 0.022 0.015 12 SBP 119.43 118.00 126.80 132.40 135.80 12 DBP 69.12 67.74 75.92 81.57 85.18 12 MAP 87.67 87.00 96.00 103.00 106.00 12 HR 54.00 55.00 66.00 71.00 77.00 12 Skin Temp 1 31.56 31.77 31.77 31.75 31.75 12 Skin Temp 2 32.28 32.38 32.40 32.40 32.38 12 Skin Temp 3 31.77 31.71 31.69 31.67 31.65
12 Muscle Temp 35.37 35.30 35.33 35.33 35.31
12 RPE 6 0 13 13 15 12 REN TAP 62.77 55.28 63.77 61.15 63.73 12 REN CON 0.72 0.64 0.66 0.59 0.60 12 CBF 2.41 3.34 2.21 2.30 2.34

174
YOUNG NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI
10 SBP 180.70 187.70 191.30 187.60 10 DBP 101.10 110.00 111.20 103.80 10 MAP 126.00 135.00 137.00 131.00 10 HR 120.00 121.00 120.00 102.00 10 Skin Temp 1 32.85 32.84 32.84 32.82 10 Skin Temp 2 32.58 32.58 32.58 32.56 10 Skin Temp 3 31.83 31.89 31.91 31.99 10 Bursts 0.79 6.32 4.74 5.50 10 Total Activity 100.03 732.34 581.55 488.78 10 Muscle Temp 34.51 34.50 34.49 34.37 10 RPE 13 16 16 0 10 REN TAP 42.43 39.30 39.30 43.25 10 REN CON 0.34 0.29 0.29 0.40 11 SBP 165.50 174.90 175.50 164.90 11 DBP 93.13 96.60 96.32 85.73 11 MAP 119.00 125.00 124.00 113.00 11 HR 100.00 99.00 98.00 82.00 11 Skin Temp 1 33.15 33.17 33.17 33.18 11 Skin Temp 2 32.66 32.70 32.71 32.74 11 Skin Temp 3 31.74 31.76 31.76 31.76 11 Bursts 13.60 21.75 9.97 17.50 11 Total Activity 4898.16 8185.77 4270.72 6527.89 11 Muscle Temp 35.47 35.46 35.46 35.44 11 RPE 19 20 20 0 11 REN TAP 43.95 40.50 39.00 39.98 11 REN CON 0.37 0.32 0.31 0.35 11 CBF 2.08 2.15 1.95 2.10 11 CVC 0.017 0.017 0.016 0.019 12 SBP 151.70 155.60 157.30 144.40 12 DBP 97.94 97.15 98.55 84.38 12 MAP 120.00 122.00 122.00 106.00 12 HR 83.00 85.00 83.00 60.00 12 Skin Temp 1 31.76 31.77 31.77 31.81 12 Skin Temp 2 32.37 32.36 32.36 32.37 12 Skin Temp 3 31.64 31.64 31.64 31.67 12 Muscle Temp 35.31 35.31 35.31 35.06 12 RPE 17 20 20 0 12 REN TAP 60.73 63.30 63.30 67.13 12 REN CON 0.51 0.52 0.52 0.63 12 CBF 2.09 2.49 1.93 2.08 12 CVC 0.017 0.020 0.016 0.020

175
YOUNG COLD
Rec 1 Rec 2 Rec 3 Subject Variable BL AVE ISCH
177.20 169.80 175.50 10 SBP 167.70 173.00 91.83 85.95 87.54 10 DBP 98.86 98.61
115.00 109.00 110.00 10 MAP 117.67 120.00 84.00 80.00 76.00 10 HR 72.33 75.00 33.40 33.77 33.74 10 Skin Temp 1 16.26 16.71 32.59 32.76 32.76 10 Skin Temp 2 15.55 16.50 32.16 32.13 32.06 10 Skin Temp 3 30.54 30.50
5.00 3.00 2.00 10 Bursts 1.67 1.00 555.35 210.48 323.07 10 Total Activity 198.21 117.50
34.44 34.63 34.72 10 Muscle Temp 23.77 23.74 0 0 0 10 RPE 6 0
43.98 45.23 45.50 10 REN TAP 65.50 62.23 0.46 0.51 0.52 10 REN CON 0.56 0.52
145.20 145.20 148.40 11 SBP 133.77 136.00 70.40 71.32 73.45 11 DBP 71.99 75.79 93.00 93.00 95.00 11 MAP 92.33 98.00 71.00 70.00 67.00 11 HR 78.67 95.00 33.25 33.41 33.50 11 Skin Temp 1 20.48 20.41 32.88 33.00 33.08 11 Skin Temp 2 18.07 18.01 31.66 31.59 31.53 11 Skin Temp 3 32.84 32.81
3.00 4.50 4.50 11 Bursts 1.67 0.50 823.45 1184.85 1142.85 11 Total Activity 344.47 195.95
35.92 35.96 35.95 11 Muscle Temp 26.69 26.92 0 0 0 11 RPE 6 0
39.00 43.95 45.65 11 REN TAP 44.37 43.23 0.42 0.47 0.48 11 REN CON 0.48 0.44 2.44 1.75 2.46 11 CBF 3.60 3.21
0.026 0.019 0.026 11 CVC 0.039 0.033 130.90 126.50 120.90 12 SBP 116.97 117.90
69.58 69.24 67.54 12 DBP 73.85 72.40 89.00 88.00 85.00 12 MAP 90.33 87.00 59.00 54.00 55.00 12 HR 61.33 67.00 31.83 31.77 31.80 12 Skin Temp 1 16.87 16.71 32.49 32.58 32.67 12 Skin Temp 2 17.63 17.46 31.68 31.68 31.67 12 Skin Temp 3 32.42 32.50 35.51 35.81 35.93 12 Muscle Temp 24.63 24.97
0 0 0 12 RPE 6 0 58.90 62.00 64.28 12 REN TAP 66.23 63.63
0.66 0.70 0.76 12 REN CON 0.73 0.73 2.65 2.43 2.31 12 CBF 2.25 2.06
0.030 0.028 0.027 12 CVC 0.025 0.024

176
YOUNG COLD Subject Variable 0-20% 20-40% 40-60% 60-80% 80-100%
10 SBP 168.90 175.90 171.20 177.00 194.40 10 DBP 101.20 99.53 104.30 104.20 115.40 10 MAP 122.00 124.00 126.00 128.00 142.00 10 HR 94.00 91.00 100.00 106.00 106.00 10 Skin Temp 1 17.09 17.34 17.52 17.61 17.64 10 Skin Temp 2 17.18 17.52 17.77 17.93 18.05 10 Skin Temp 3 30.56 30.55 30.58 30.58 30.59 10 Bursts 0.00 0.87 0.87 0.87 1.74 10 Total Activity 0.00 131.88 36.97 56.90 96.64 10 Muscle Temp 23.81 23.84 23.93 24.03 24.14 10 RPE 11 11 13 15 17 10 REN TAP 71.80 71.20 70.90 70.00 70.00 10 REN CON 0.59 0.57 0.56 0.55 0.49 11 SBP 147.60 149.10 152.10 156.20 164.90 11 DBP 81.60 81.70 83.46 91.86 102.30 11 MAP 102.00 103.00 105.00 114.00 123.00 11 HR 95.00 94.00 100.00 109.00 111.00 11 Skin Temp 1 20.39 20.35 20.31 20.25 20.20 11 Skin Temp 2 18.12 18.16 18.19 18.21 18.21 11 Skin Temp 3 32.80 32.81 32.84 32.86 32.96 11 Bursts 0.00 3.76 1.25 5.01 5.01 11 Total Activity 0.00 499.08 239.40 917.45 919.16 11 Muscle Temp 27.23 27.23 27.15 27.06 26.81 11 RPE 11 11 13 17 20 11 REN TAP 40.00 42.95 38.00 38.00 39.00 11 REN CON 0.39 0.42 0.36 0.33 0.32 11 CBF 3.33 3.48 4.79 3.51 3.60 11 CVC 0.033 0.034 0.046 0.031 0.029 12 SBP 120.50 124.90 132.00 133.80 139.00 12 DBP 77.90 82.43 88.09 89.02 93.87 12 MAP 95.00 99.00 105.00 108.00 115.00 12 HR 69.00 75.00 75.00 83.00 89.00 12 Skin Temp 1 16.69 16.69 16.76 16.84 16.96 12 Skin Temp 2 17.44 17.43 17.43 17.45 17.50 12 Skin Temp 3 32.52 32.54 32.55 32.56 32.57 12 Muscle Temp 24.97 24.97 25.11 25.12 25.12 12 RPE 10 13 14 17 20 12 REN TAP 61.35 62.03 63.30 64.00 63.30 12 REN CON 0.65 0.63 0.60 0.59 0.55 12 CBF 1.53 1.92 2.15 1.60 1.91 12 CVC 0.016 0.019 0.020 0.015 0.017

177
YOUNG COLD Subject Variable fatigue PEMI Rec 1 Rec 2 Rec 3
10 SBP 196.50 184.10 179.90 175.10 172.50 10 DBP 117.80 110.20 101.10 95.16 93.66 10 MAP 144.00 132.00 122.00 116.00 114.00 10 HR 107.00 85.00 65.00 58.00 62.00 10 Skin Temp 1 17.64 17.72 18.70 19.22 19.09 10 Skin Temp 2 18.07 18.19 17.87 17.44 17.02 10 Skin Temp 3 30.61 30.68 30.65 30.65 30.63 10 Bursts 4.36 4.50 6.50 4.00 1.00 10 Total Activity 398.07 422.76 814.16 328.29 146.10 10 Muscle Temp 24.14 24.43 24.54 24.84 25.05 10 RPE 19 0 0 0 0 10 REN TAP 70.00 70.00 63.75 60.90 65.50 10 REN CON 0.49 0.53 0.52 0.53 0.57 11 SBP 164.70 145.60 144.70 143.50 147.20 11 DBP 100.50 80.94 68.64 69.21 69.95 11 MAP 123.00 102.00 91.00 91.00 92.00 11 HR 107.00 87.00 81.00 80.00 75.00 11 Skin Temp 1 20.19 20.08 19.71 19.45 19.34 11 Skin Temp 2 18.20 18.17 17.97 17.89 17.91 11 Skin Temp 3 33.00 33.07 33.00 32.93 32.90 11 Bursts 2.50 2.50 1.00 1.50 1.00 11 Total Activity 613.27 469.20 277.75 238.35 361.85 11 Muscle Temp 26.81 26.57 27.72 27.79 27.79 11 RPE 20 0 0 0 0 11 REN TAP 39.00 38.25 39.00 41.75 42.00 11 REN CON 0.32 0.38 0.43 0.46 0.46 11 CBF 3.60 4.17 3.75 3.81 3.90 11 CVC 0.029 0.041 0.041 0.042 0.042 12 SBP 139.00 134.20 127.70 121.00 117.60 12 DBP 93.77 84.06 76.06 70.94 69.45 12 MAP 114.00 103.00 94.00 89.00 86.00 12 HR 92.00 67.00 62.00 56.00 59.00 12 Skin Temp 1 16.98 17.16 17.32 17.18 17.14 12 Skin Temp 2 17.51 17.61 17.75 17.89 18.15 12 Skin Temp 3 32.57 32.60 32.63 32.64 32.66 12 Muscle Temp 25.12 24.47 25.75 25.75 26.38 12 RPE 20 0 0 0 0 12 REN TAP 63.30 68.83 60.70 59.45 60.40 12 REN CON 0.56 0.67 0.65 0.67 0.70 12 CBF 1.91 2.51 2.23 1.82 1.90 12 CVC 0.017 0.024 0.024 0.020 0.022

178
Appendix E
Raw Data For Older Normothermia & Heating Studies

179
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
1 SBP 127.90 123.70 131.70 130.70 132.20 1 ∆ SBP 0.00 -4.20 3.80 2.80 4.30 1 DBP 58.13 57.66 66.24 64.52 66.49 1 ∆ DBP 0.00 -0.47 8.11 6.39 8.36 1 MAP 82.67 82.00 92.00 90.00 94.00 1 ∆MAP 0.00 -0.67 9.33 7.33 11.33 1 HR 57.33 57.00 70.00 74.00 76.00 1 ∆ HR 0.00 -0.33 12.67 16.67 18.67 1 Skin temp 1 30.25 30.23 30.24 30.25 30.28 1 ∆ Skin temp 1 0.00 -0.02 -0.01 0.00 0.03 1 Skin temp 2 31.30 31.21 31.15 31.13 31.12 1 ∆ Skin temp 2 0.00 -0.09 -0.15 -0.17 -0.18 1 Skin temp 3 31.85 31.76 31.75 31.72 31.71 1 ∆ Skin temp 3 0.00 -0.09 -0.10 -0.13 -0.14 1 bursts 1 ∆bursts 1 %∆bursts 1 total act 1 ∆total act 1 % ∆ total act 1 muscle temp 34.12 34.12 34.09 34.06 34.03 1 ∆mus temp 0.00 0.00 -0.03 -0.06 -0.09 1 RPE 6 0 15 17 19 1 ∆RPE 0 0 9 11 13 1 REN TAP 61.99 60.88 59.40 50.30 59.40 1 ∆RENAL TAP 0.00 -1.11 -2.59 -11.69 -2.59 1 %∆REN TAP 0.00 -1.70 -4.17 -18.85 1.47 1 REN CON 0.75 0.74 0.65 0.56 0.63 1 ∆REN CON 0.00 -0.01 -0.10 -0.19 -0.12 1 %∆ REN CON 0.00 -0.99 -13.89 -25.46 -15.73 1 CBF 1 ∆CBF 1 %∆CBF 1 cvc 1 ∆cvc 1 %∆cvc

180
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
1 SBP 136.00 145.10 148.20 139.00 132.30 1 ∆ SBP 8.10 17.20 20.30 11.10 4.40 1 DBP 63.87 68.77 66.22 62.56 58.37 1 ∆ DBP 5.74 10.64 8.09 4.43 0.24 1 MAP 96.00 100.00 100.00 91.00 85.00 1 ∆MAP 13.33 17.33 17.33 8.33 2.33 1 HR 76.00 74.00 73.00 62.00 62.00 1 ∆ HR 18.67 16.67 15.67 4.67 4.67 1 Skin temp 1 30.31 30.33 30.34 30.36 30.43 1 ∆ Skin temp 1 0.06 0.08 0.09 0.11 0.18 1 Skin temp 2 31.12 31.13 31.14 31.13 31.18 1 ∆ Skin temp 2 -0.18 -0.17 -0.16 -0.17 -0.12 1 Skin temp 3 31.70 31.69 31.69 31.64 31.54 1 ∆ Skin temp 3 -0.15 -0.16 -0.16 -0.21 -0.31 1 bursts 1 ∆bursts 1 %∆bursts 1 total act 1 ∆total act 1 % ∆ total act 1 muscle temp 34.03 34.03 34.03 34.01 33.99 1 ∆mus temp -0.09 -0.09 -0.09 -0.11 -0.13 1 RPE 20 20 20 0 0 1 ∆RPE 14 14 14 0 0 1 REN TAP 55.43 51.23 48.20 48.55 56.88 1 ∆RENAL TAP -6.55 -10.75 -13.79 -13.44 -5.11 1 %∆REN TAP -11.32 -17.35 -22.24 -20.69 -7.87 1 REN CON 0.56 0.51 0.48 0.53 0.67 1 ∆REN CON -0.19 -0.24 -0.27 -0.22 -0.08 1 %∆ REN CON -25.33 -31.67 -35.72 -28.85 -10.76 1 CBF 1 ∆CBF 1 %∆CBF 1 cvc 1 ∆cvc 1 %∆cvc

181
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
134.30 131.80 1 SBP 123.10 123.90 123.50 6.40 3.90 1 ∆ SBP 0.00 0.80 0.40
57.56 55.90 1 DBP 66.93 78.89 87.74 -0.57 -2.23 1 ∆ DBP 0.00 11.96 20.81 84.00 83.00 1 MAP 90.00 99.00 103.00
1.33 0.33 1 ∆MAP 0.00 9.00 13.00 57.00 60.00 1 HR 72.00 75.00 79.00 -0.33 2.67 1 ∆ HR 0.00 3.00 7.00 30.62 30.90 1 Skin temp 1 40.49 40.52 40.78
0.37 0.65 1 ∆ Skin temp 1 0.00 0.03 0.29 31.39 31.45 1 Skin temp 2 39.01 39.10 39.35
0.09 0.15 1 ∆ Skin temp 2 0.00 0.09 0.34 31.50 31.46 1 Skin temp 3 31.15 31.08 31.06 -0.35 -0.39 1 ∆ Skin temp 3 0.00 -0.07 -0.09
1 bursts 1 ∆bursts 1 %∆bursts 1 total act 1 ∆total act 1 % ∆ total act
34.22 34.33 1 muscle temp 38.48 38.52 38.74 0.10 0.21 1 ∆mus temp 0.00 0.04 0.26
0 0 1 RPE 6 0 13 0 0 1 ∆RPE 0 0 7
59.40 59.63 1 REN TAP 59.11 56.30 56.10 -2.59 -2.35 1 ∆RENAL TAP 0.00 -2.81 -3.01 -3.98 -3.62 1 %∆REN TAP 0.00 -4.76 -5.09 0.71 0.72 1 REN CON 0.66 0.57 0.54
-0.04 -0.03 1 ∆REN CON 0.00 -0.09 -0.11 -5.69 -4.18 1 %∆ REN CON 0.00 -13.41 -17.07
1 CBF 1 ∆CBF 1 %∆CBF 1 cvc 1 ∆cvc 1 %∆cvc

182
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
1 SBP 127.00 138.30 142.10 154.70 154.20 1 ∆ SBP 3.90 15.20 19.00 31.60 31.10 1 DBP 88.21 91.66 96.41 109.80 108.60 1 ∆ DBP 21.28 24.73 29.48 42.87 41.67 1 MAP 108.00 120.00 119.00 129.00 127.00 1 ∆MAP 18.00 30.00 29.00 39.00 37.00 1 HR 76.00 70.00 82.00 80.00 79.00 1 ∆ HR 4.00 -2.00 10.00 8.00 7.00 1 Skin temp 1 40.90 41.02 41.10 41.15 41.17 1 ∆ Skin temp 1 0.41 0.53 0.61 0.66 0.68 1 Skin temp 2 39.47 39.60 39.71 39.82 39.85 1 ∆ Skin temp 2 0.46 0.59 0.70 0.81 0.84 1 Skin temp 3 31.05 31.04 31.03 31.00 31.00 1 ∆ Skin temp 3 -0.10 -0.11 -0.12 -0.15 -0.15 1 bursts 1 ∆bursts 1 %∆bursts 1 total act 1 ∆total act 1 % ∆ total act 1 muscle temp 38.74 38.82 38.90 39.00 39.00 1 ∆mus temp 0.26 0.34 0.42 0.52 0.52 1 RPE 13 18 18 20 20 1 ∆RPE 7 12 12 14 14 1 REN TAP 57.30 58.00 50.30 50.30 50.30 1 ∆RENAL TAP -1.81 -1.11 -8.81 -8.81 -8.81 1 %∆REN TAP -3.06 -1.88 -14.91 -14.91 -14.91 1 REN CON 0.53 0.48 0.42 0.39 0.40 1 ∆REN CON -0.13 -0.17 -0.23 -0.27 -0.26 1 %∆ REN CON -19.22 -26.41 -35.64 -40.63 -39.70 1 CBF 1 ∆CBF 1 %∆CBF 1 cvc 1 ∆cvc 1 %∆cvc

183
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
1 SBP 143.60 139.10 138.50 133.20 1 ∆ SBP 20.50 16.00 15.40 10.10 1 DBP 97.87 81.96 85.46 90.80 1 ∆ DBP 30.94 15.03 18.53 23.87 1 MAP 114.00 99.00 105.00 110.00 1 ∆MAP 24.00 9.00 15.00 20.00 1 HR 68.00 73.00 73.00 75.00 1 ∆ HR -4.00 1.00 1.00 3.00 1 Skin temp 1 41.27 40.42 38.88 38.19 1 ∆ Skin temp 1 0.78 -0.07 -1.61 -2.30 1 Skin temp 2 39.92 39.32 38.15 37.51 1 ∆ Skin temp 2 0.91 0.31 -0.86 -1.50 1 Skin temp 3 30.94 30.80 30.76 30.74 1 ∆ Skin temp 3 -0.21 -0.35 -0.39 -0.41 1 bursts 1 ∆bursts 1 %∆bursts 1 total act 1 ∆total act 1 % ∆ total act 1 muscle temp 39.20 39.39 39.21 38.92 1 ∆mus temp 0.72 0.91 0.73 0.44 1 RPE 0 0 0 0 1 ∆RPE 0 0 0 0 1 REN TAP 52.63 55.43 55.20 58.70 1 ∆RENAL TAP -6.48 -3.68 -3.91 -0.41 1 %∆REN TAP -10.96 -6.22 -6.62 -0.70 1 REN CON 0.46 0.56 0.53 0.53 1 ∆REN CON -0.20 -0.10 -0.13 -0.12 1 %∆ REN CON -29.70 -14.75 -19.96 -18.75 1 CBF 1 ∆CBF 1 %∆CBF 1 cvc 1 ∆cvc 1 %∆cvc

184
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
2 SBP 164.63 184.50 198.00 201.60 202.00 2 ∆ SBP 0.00 19.87 33.37 36.97 37.37 2 DBP 79.42 92.00 100.00 98.53 100.60 2 ∆ DBP 0.00 12.58 20.58 19.11 21.18 2 MAP 111.67 126.00 136.00 137.00 139.00 2 ∆MAP 0.00 14.33 24.33 25.33 27.33 2 HR 64.00 65.00 69.00 72.00 74.00 2 ∆ HR 0.00 1.00 5.00 8.00 10.00 2 Skin temp 1 30.32 30.27 30.24 30.22 30.20 2 ∆ Skin temp 1 0.00 -0.05 -0.08 -0.10 -0.12 2 Skin temp 2 30.63 30.60 30.59 30.59 30.58 2 ∆ Skin temp 2 0.00 -0.03 -0.04 -0.04 -0.05 2 Skin temp 3 30.63 30.62 30.58 30.58 30.56 2 ∆ Skin temp 3 0.00 -0.01 -0.05 -0.05 -0.07 2 bursts 0.00 0.00 2 ∆bursts 0.00 0.00 2 %∆bursts 0.00 0.00 2 total act 0.00 0.00 2 ∆total act 0.00 0.00 2 % ∆ total act 0.00 0.00 2 muscle temp 33.04 33.10 33.24 33.26 33.27 2 ∆mus temp 0.00 0.06 0.20 0.22 0.23 2 RPE 6 0 12 14 16 2 ∆RPE 0 0 6 8 10 2 REN TAP 43.84 37.08 38.10 36.00 34.90 2 ∆RENAL TAP 0.00 -6.77 -5.74 -7.84 -8.94 2 %∆REN TAP 0.00 -15.43 -13.10 -17.89 -20.40 2 REN CON 0.39 0.29 0.28 0.26 0.25 2 ∆REN CON 0.00 -0.10 -0.11 -0.13 -0.14 2 %∆ REN CON 0.00 -25.05 -28.65 -33.07 -36.05 2 CBF 2.05 2.40 2.75 2.98 3.19 2 ∆CBF 0.00 0.35 0.70 0.93 1.14 2 %∆CBF 0.00 16.87 34.18 45.33 55.63 2 cvc 0.018 0.019 0.020 0.022 0.023 2 ∆cvc 0.000 0.001 0.002 0.003 0.005 2 %∆cvc 0.00 3.57 10.17 18.45 25.03

185
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
2 SBP 209.20 208.10 207.30 183.50 184.10 2 ∆ SBP 44.57 43.47 42.67 18.87 19.47 2 DBP 100.30 89.00 82.55 96.25 89.98 2 ∆ DBP 20.88 9.58 3.13 16.83 10.56 2 MAP 137.00 135.00 128.00 130.00 123.00 2 ∆MAP 25.33 23.33 16.33 18.33 11.33 2 HR 75.00 79.00 82.00 71.00 68.00 2 ∆ HR 11.00 15.00 18.00 7.00 4.00 2 Skin temp 1 30.17 30.15 30.15 30.13 30.22 2 ∆ Skin temp 1 -0.15 -0.17 -0.17 -0.19 -0.10 2 Skin temp 2 30.57 30.56 30.56 30.55 30.56 2 ∆ Skin temp 2 -0.06 -0.07 -0.07 -0.08 -0.07 2 Skin temp 3 30.54 30.51 30.50 30.47 30.42 2 ∆ Skin temp 3 -0.09 -0.12 -0.13 -0.16 -0.21 2 bursts 2 ∆bursts 2 %∆bursts 2 total act 2 ∆total act 2 % ∆ total act 2 muscle temp 33.29 33.33 33.36 33.34 33.59 2 ∆mus temp 0.25 0.29 0.32 0.30 0.55 2 RPE 18 19 20 0 0 2 ∆RPE 12 13 14 0 0 2 REN TAP 28.60 25.40 25.40 34.15 36.33 2 ∆RENAL TAP -15.24 -18.44 -18.44 -9.69 -7.51 2 %∆REN TAP -34.77 -42.06 -42.06 -21.67 -16.79 2 REN CON 0.21 0.19 0.20 0.26 0.30 2 ∆REN CON -0.18 -0.20 -0.19 -0.13 -0.10 2 %∆ REN CON -46.83 -52.08 -49.46 -33.09 -24.76 2 CBF 3.08 2.79 2.79 3.90 2.36 2 ∆CBF 1.02 0.73 0.73 1.85 0.31 2 %∆CBF 49.94 35.80 35.80 90.35 14.91 2 cvc 0.022 0.021 0.022 0.030 0.019 2 ∆cvc 0.004 0.002 0.003 0.010 0.000 2 %∆cvc 22.21 12.32 18.47 52.90 -2.44

186
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
176.70 173.20 2 SBP 171.20 180.70 195.40 12.07 8.57 2 ∆ SBP 0.00 9.50 24.20 87.12 87.01 2 DBP 82.42 89.32 95.22
7.70 7.59 2 ∆ DBP 0.00 6.90 12.80 119.00 118.00 2 MAP 110.33 119.00 130.00
7.33 6.33 2 ∆MAP 0.00 8.67 19.67 63.00 62.00 2 HR 62.00 65.00 67.00 -1.00 -2.00 2 ∆ HR 0.00 3.00 5.00 30.34 30.34 2 Skin temp 1 40.73 40.66 41.09
0.02 0.02 2 ∆ Skin temp 1 0.00 -0.07 0.36 30.58 30.61 2 Skin temp 2 40.23 40.12 40.29 -0.05 -0.02 2 ∆ Skin temp 2 0.00 -0.11 0.06 30.42 30.39 2 Skin temp 3 29.20 29.17 29.16 -0.21 -0.24 2 ∆ Skin temp 3 0.00 -0.03 -0.04
2 bursts 0.00 2 ∆bursts 0.00 2 %∆bursts 0.00 2 total act 0.00 2 ∆total act 0.00 2 % ∆ total act 0.00
33.89 34.12 2 muscle temp 37.61 37.58 37.69 0.85 1.08 2 ∆mus temp 0.00 -0.03 0.08
0 0 2 RPE 6 0 12 0 0 2 ∆RPE 0 0 6
41.80 42.88 2 REN TAP 56.39 46.10 44.00 -2.04 -0.97 2 ∆RENAL TAP 0.00 -10.29 -12.39 -4.56 -2.16 2 %∆REN TAP 0.00 -18.24 -21.97 0.35 0.36 2 REN CON 0.51 0.39 0.34
-0.04 -0.03 2 ∆REN CON 0.00 -0.12 -0.17 -10.53 -7.45 2 %∆ REN CON 0.00 -24.20 -33.77
2.51 1.91 2 CBF 1.60 1.67 1.78 0.45 -0.14 2 ∆CBF 0.00 0.07 0.18
22.15 -6.76 2 %∆CBF 0.00 4.17 10.94 0.021 0.016 2 cvc 0.015 0.014 0.014 0.001 -0.003 2 ∆cvc 0.000 0.000 -0.001
7.19 -17.49 2 %∆cvc 0.00 -3.42 -5.84

187
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
2 SBP 202.20 208.60 217.00 229.40 233.00 2 ∆ SBP 31.00 37.40 45.80 58.20 61.80 2 DBP 106.20 113.40 114.60 124.40 128.00 2 ∆ DBP 23.78 30.98 32.18 41.98 45.58 2 MAP 142.00 151.00 152.92 164.30 168.06 2 ∆MAP 31.67 40.67 42.59 53.97 57.73 2 HR 71.00 73.00 73.00 78.00 78.00 2 ∆ HR 9.00 11.00 11.00 16.00 16.00 2 Skin temp 1 41.25 41.40 41.51 41.63 41.65 2 ∆ Skin temp 1 0.52 0.67 0.78 0.90 0.92 2 Skin temp 2 40.36 40.42 40.49 40.56 40.57 2 ∆ Skin temp 2 0.13 0.19 0.26 0.33 0.34 2 Skin temp 3 29.15 29.13 29.13 29.11 29.11 2 ∆ Skin temp 3 -0.05 -0.07 -0.07 -0.09 -0.09 2 bursts 2 ∆bursts 2 %∆bursts 2 total act 2 ∆total act 2 % ∆ total act 2 muscle temp 37.77 37.83 37.88 38.10 38.10 2 ∆mus temp 0.16 0.21 0.27 0.49 0.49 2 RPE 16 17 18 19 19 2 ∆RPE 10 11 12 13 13 2 REN TAP 34.00 32.80 28.60 23.30 23.30 2 ∆RENAL TAP -22.39 -23.59 -27.79 -33.09 -33.09 2 %∆REN TAP -39.70 -41.83 -49.28 -58.68 -58.68 2 REN CON 0.24 0.22 0.19 0.14 0.14 2 ∆REN CON -0.27 -0.29 -0.32 -0.37 -0.37 2 %∆ REN CON -53.15 -57.50 -63.40 -72.25 -72.87 2 CBF 1.21 2.27 2.14 2.01 2.01 2 ∆CBF -0.40 0.66 0.54 0.41 0.41 2 %∆CBF -24.92 41.26 33.34 25.37 25.37 2 cvc 0.008 0.015 0.014 0.012 0.012 2 ∆cvc -0.006 0.000 -0.001 -0.002 -0.003 2 %∆cvc -41.66 3.21 -3.79 -15.81 -17.69

188
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
2 SBP 180.40 164.70 161.90 165.10 2 ∆ SBP 9.20 -6.50 -9.30 -6.10 2 DBP 93.71 81.74 82.58 82.97 2 ∆ DBP 11.29 -0.68 0.16 0.55 2 MAP 126.00 106.00 106.00 107.00 2 ∆MAP 15.67 -4.33 -4.33 -3.33 2 HR 71.00 64.00 63.00 62.00 2 ∆ HR 9.00 2.00 1.00 0.00 2 Skin temp 1 41.82 41.61 41.00 40.66 2 ∆ Skin temp 1 1.09 0.88 0.27 -0.07 2 Skin temp 2 40.67 40.84 40.64 40.38 2 ∆ Skin temp 2 0.44 0.61 0.41 0.15 2 Skin temp 3 29.08 29.09 29.11 29.13 2 ∆ Skin temp 3 -0.12 -0.11 -0.09 -0.07 2 bursts 2 ∆bursts 2 %∆bursts 2 total act 2 ∆total act 2 % ∆ total act 2 muscle temp 38.22 38.42 38.20 37.75 2 ∆mus temp 0.61 0.81 0.59 0.14 2 RPE 0 0 0 0 2 ∆RPE 0 0 0 0 2 REN TAP 40.57 48.95 52.93 56.90 2 ∆RENAL TAP -15.82 -7.44 -3.46 0.51 2 %∆REN TAP -28.06 -13.19 -6.14 0.91 2 REN CON 0.32 0.46 0.50 0.53 2 ∆REN CON -0.19 -0.05 -0.01 0.02 2 %∆ REN CON -37.00 -9.64 -2.30 4.05 2 CBF 4.23 2.03 1.70 1.53 2 ∆CBF 2.63 0.42 0.09 -0.08 2 %∆CBF 163.73 26.21 5.77 -4.77 2 cvc 0.034 0.019 0.016 0.014 2 ∆cvc 0.019 0.005 0.001 0.000 2 %∆cvc 130.94 31.37 10.10 -1.81

189
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
3 SBP 101.68 94.18 113.10 129.40 147.30 3 ∆ SBP 0.00 -7.50 11.42 27.72 45.62 3 DBP 70.20 69.87 82.58 90.71 101.30 3 ∆ DBP 0.00 -0.33 12.38 20.51 31.10 3 MAP 85.67 80.00 96.00 107.00 121.00 3 ∆MAP 0.00 -5.67 10.33 21.33 35.33 3 HR 56.33 56.00 59.00 61.00 65.00 3 ∆ HR 0.00 -0.33 2.67 4.67 8.67 3 Skin temp 1 30.31 30.24 30.25 30.24 30.19 3 ∆ Skin temp 1 0.00 -0.07 -0.06 -0.07 -0.12 3 Skin temp 2 30.57 30.56 30.58 30.58 30.54 3 ∆ Skin temp 2 0.00 -0.01 0.01 0.01 -0.03 3 Skin temp 3 30.03 29.93 29.91 29.91 29.87 3 ∆ Skin temp 3 0.00 -0.10 -0.12 -0.12 -0.16 3 bursts 11.00 13.00 8.53 9.19 21.00 3 ∆bursts 0.00 2.00 -2.47 -1.81 10.00 3 %∆bursts 0.00 18.18 -22.46 -16.50 90.87 3 peak amp 1293.00 1502.10 853.58 1379.52 3081.60 3 ∆ peak amp 0.00 208.47 -439.42 86.52 1788.60 3 %∆ peak amp 0.00 16.11 -33.98 6.69 138.33 3 muscle temp 33.82 33.79 33.86 33.82 33.82 3 ∆muscle temp 0.00 -0.03 0.04 0.00 0.00 3 RPE 6 0 12 14 17 3 ∆RPE 0 0 6 8 11 3 REN TAP 83.87 73.64 73.80 72.80 71.40 3 ∆RENAL TAP 0.00 -10.23 -10.07 -11.07 -10.47 3 %∆REN TAP 0.00 -12.19 -12.00 -13.20 -12.48 3 REN CON 0.98 0.92 0.77 0.68 0.61 3 ∆REN CON 0.00 -0.06 -0.21 -0.30 -0.37 3 %∆ REN CON 0.00 -5.97 -21.48 -30.50 -38.04 3 CBF 0.88 0.87 1.03 1.12 1.34 3 ∆CBF 0.00 0.00 0.15 0.24 0.46 3 %∆CBF 0.00 -0.54 17.14 27.68 52.60 3 cvc 0.010 0.011 0.011 0.010 0.011 3 ∆cvc 0.000 0.001 0.000 0.000 0.001 3 %∆cvc 0.00 6.50 4.53 2.22 8.04

190
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
3 SBP 159.10 174.20 174.10 146.50 135.20 3 ∆ SBP 57.42 72.52 72.42 44.82 33.52 3 DBP 107.10 117.60 119.00 95.58 85.85 3 ∆ DBP 36.90 47.40 48.80 25.38 15.65 3 MAP 129.00 141.00 142.00 116.00 105.00 3 ∆MAP 43.33 55.33 56.33 30.33 19.33 3 HR 67.00 72.00 74.00 59.00 57.00 3 ∆ HR 10.67 15.67 17.67 2.67 0.67 3 Skin temp 1 30.12 30.08 30.08 30.21 30.29 3 ∆ Skin temp 1 -0.19 -0.23 -0.23 -0.10 -0.02 3 Skin temp 2 30.50 30.43 30.43 30.40 29.93 3 ∆ Skin temp 2 -0.07 -0.14 -0.14 -0.17 -0.64 3 Skin temp 3 29.85 29.85 29.84 29.84 29.77 3 ∆ Skin temp 3 -0.18 -0.18 -0.19 -0.19 -0.26 3 bursts 20.34 24.28 19.68 15.00 25.00 3 ∆bursts 9.34 13.28 8.68 18.00 14.00 3 %∆bursts 84.90 120.69 78.91 150.00 127.27 3 peak amp 3793.35 4511.11 4332.46 2467.42 3122.47 3 ∆ peak amp 2500.35 3218.11 3039.46 3516.98 1828.84 3 %∆ peak amp 193.38 248.89 235.07 248.05 141.37 3 muscle temp 33.82 33.81 33.79 33.94 34.12 3 ∆muscle temp 0.00 -0.01 -0.03 0.12 0.30 3 RPE 18 19 19 0 0 3 ∆RPE 12 13 13 0 0 3 REN TAP 72.20 72.20 69.80 103.90 97.87 3 ∆RENAL TAP -12.47 -11.67 -14.07 20.03 14.00 3 %∆REN TAP -14.86 -13.91 -16.77 24.86 17.37 3 REN CON 0.55 0.51 0.49 0.90 0.93 3 ∆REN CON -0.43 -0.47 -0.49 -0.08 -0.05 3 %∆ REN CON -43.46 -47.70 -49.79 -8.51 -4.79 3 CBF 1.19 1.42 1.29 1.33 3.80 3 ∆CBF 0.31 0.54 0.42 0.45 2.92 3 %∆CBF 35.70 62.00 47.28 51.30 332.35 3 cvc 0.009 0.010 0.009 0.011 0.036 3 ∆cvc -0.001 0.000 -0.001 0.001 0.026 3 %∆cvc -9.88 -1.57 -11.15 10.01 247.29

191
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
133.10 128.30 3 SBP 117.93 125.90 139.70 31.42 26.62 3 ∆ SBP 0.00 7.97 21.77 83.65 81.23 3 DBP 79.64 80.86 89.41 13.45 11.03 3 ∆ DBP 0.00 1.22 9.77
103.00 100.00 3 MAP 95.00 98.00 109.00 17.33 14.33 3 ∆MAP 0.00 3.00 14.00 56.00 56.00 3 HR 59.33 61.00 65.00 -0.33 -0.33 3 ∆ HR 0.00 1.67 5.67 30.36 30.43 3 Skin temp 1 38.35 38.30 38.26
0.05 0.12 3 ∆ Skin temp 1 0.00 -0.05 -0.09 29.94 29.93 3 Skin temp 2 39.58 39.45 39.40 -0.63 -0.64 3 ∆ Skin temp 2 0.00 -0.13 -0.18 29.72 29.68 3 Skin temp 3 30.96 31.08 31.12 -0.31 -0.35 3 ∆ Skin temp 3 0.00 0.12 0.16 19.00 13.00 3 bursts 9.00 17.00 15.11
8.00 2.00 3 ∆bursts 0.00 -1.00 6.11 72.73 18.18 3 %∆bursts 0.00 -5.56 67.93
2479.41 1742.89 3 total act 1357.83 2676.78 2784.53 1185.78 449.26 3 ∆total act 0.00 -38.88 1426.70
91.66 34.73 3 % ∆ total act 0.00 -1.43 105.07 34.20 34.24 3 muscle temp 38.75 38.65 38.59
0.38 0.42 3 ∆mus temp 0.00 -0.10 -0.16 0 0 3 RPE 6 0 12 0 0 3 ∆RPE 0 0 6
127.25 114.00 3 REN TAP 117.03 102.18 96.20 43.38 30.13 3 ∆RENAL TAP 0.00 -14.86 -20.83 53.83 37.39 3 %∆REN TAP 0.00 -12.70 -17.80
1.24 1.14 3 REN CON 1.23 1.04 0.88 0.26 0.16 3 ∆REN CON 0.00 -0.19 -0.35
26.20 16.45 3 %∆ REN CON 0.00 -15.37 -28.36 3.27 3.27 3 CBF 1.24 1.28 1.25 2.39 2.39 3 ∆CBF 0.00 0.03 0.01
271.74 271.74 3 %∆CBF 0.00 2.60 0.71 0.032 0.033 3 cvc 0.013 0.013 0.011 0.021 0.022 3 ∆cvc 0.000 0.000 -0.002
204.41 213.54 3 %∆cvc 0.00 -0.55 -12.23

192
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
3 SBP 159.30 176.90 177.90 185.60 186.80 3 ∆ SBP 41.37 58.97 59.97 67.67 68.87 3 DBP 101.80 110.70 108.50 115.40 117.60 3 ∆ DBP 22.16 31.06 28.86 35.76 37.96 3 MAP 125.00 139.00 138.00 145.00 147.00 3 ∆MAP 30.00 44.00 43.00 50.00 52.00 3 HR 72.00 76.00 72.00 79.00 79.00 3 ∆ HR 12.67 16.67 12.67 19.67 19.67 3 Skin temp 1 38.17 38.02 37.95 37.93 37.92 3 ∆ Skin temp 1 -0.18 -0.33 -0.40 -0.42 -0.43 3 Skin temp 2 39.31 39.18 39.10 39.03 39.02 3 ∆ Skin temp 2 -0.27 -0.40 -0.48 -0.55 -0.56 3 Skin temp 3 31.18 31.24 31.31 31.34 31.35 3 ∆ Skin temp 3 0.22 0.28 0.35 0.38 0.39 3 bursts 24.05 21.98 19.92 24.05 24.75 3 ∆bursts 15.05 12.98 10.92 15.05 15.75 3 %∆bursts 167.17 144.27 121.37 167.17 174.96 3 total act 5118.03 4913.69 4404.50 5425.14 4910.85 3 ∆total act 3760.21 3555.86 3046.67 4067.31 3553.03 3 % ∆ total act 276.93 261.88 224.38 299.55 261.67 3 muscle temp 38.59 38.56 38.56 38.53 38.51 3 ∆mus temp -0.16 -0.19 -0.19 -0.22 -0.24 3 RPE 14 17 19 19 19 3 ∆RPE 8 11 13 13 13 3 REN TAP 76.55 65.00 63.75 62.57 58.95 3 ∆RENAL TAP -40.48 -52.03 -53.28 -54.47 -58.08 3 %∆REN TAP -34.59 -44.46 -45.53 -46.54 -49.63 3 REN CON 0.61 0.47 0.46 0.43 0.40 3 ∆REN CON -0.62 -0.76 -0.77 -0.80 -0.83 3 %∆ REN CON -50.29 -62.04 -62.50 -64.97 -67.45 3 CBF 1.14 1.36 1.34 1.81 1.81 3 ∆CBF -0.11 0.11 0.10 0.56 0.56 3 %∆CBF -8.70 9.01 7.94 45.39 45.39 3 cvc 0.009 0.010 0.010 0.012 0.012 3 ∆cvc -0.004 -0.003 -0.003 -0.001 -0.001 3 %∆cvc -30.61 -25.50 -25.69 -4.74 -6.04

193
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
3 SBP 147.60 133.40 135.00 134.40 3 ∆ SBP 29.67 15.47 17.07 16.47 3 DBP 86.02 81.78 82.05 82.19 3 ∆ DBP 6.38 2.14 2.41 2.55 3 MAP 110.00 102.00 103.00 103.00 3 ∆MAP 15.00 7.00 8.00 8.00 3 HR 64.00 60.00 59.00 60.00 3 ∆ HR 4.67 0.67 -0.33 0.67 3 Skin temp 1 37.95 38.00 38.01 38.01 3 ∆ Skin temp 1 -0.40 -0.35 -0.34 -0.34 3 Skin temp 2 38.98 38.81 38.86 38.90 3 ∆ Skin temp 2 -0.60 -0.77 -0.72 -0.68 3 Skin temp 3 31.37 31.39 31.41 31.45 3 ∆ Skin temp 3 0.41 0.43 0.45 0.49 3 bursts 22.00 15.00 23.00 16.00 3 ∆bursts 2.00 -3.00 5.00 -2.00 3 %∆bursts 4.00 -16.67 27.78 -11.11 3 total act 44.44 3155.58 3350.94 2129.85 3 ∆total act 226.91 439.92 635.28 -585.81 3 % ∆ total act 453.82 16.20 23.39 -21.57 3 muscle temp 33.42 38.78 38.80 38.80 3 ∆mus temp 0.00 0.03 0.05 0.05 3 RPE 0 0 0 0 3 ∆RPE 0 0 0 0 3 REN TAP 82.40 83.40 89.63 89.00 3 ∆RENAL TAP -34.63 -33.63 -27.41 -28.03 3 %∆REN TAP -29.59 -28.74 -23.42 -23.95 3 REN CON 0.75 0.82 0.87 0.86 3 ∆REN CON -0.48 -0.41 -0.36 -0.37 3 %∆ REN CON -39.19 -33.63 -29.37 -29.86 3 CBF 1.32 0.87 0.84 0.74 3 ∆CBF 0.07 -0.37 -0.40 -0.50 3 %∆CBF 5.95 -29.73 -32.14 -40.37 3 cvc 0.012 0.009 0.008 0.007 3 ∆cvc -0.001 -0.005 -0.005 -0.006 3 %∆cvc -8.50 -34.56 -37.41 -45.01

194
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
4 SBP 140.63 145.90 161.20 169.00 181.00 4 ∆ SBP 0.00 5.27 20.57 28.37 40.37 4 DBP 94.01 94.39 101.90 106.50 113.20 4 ∆ DBP 0.00 0.38 7.89 12.49 19.19 4 MAP 114.00 118.00 126.00 136.00 143.00 4 ∆MAP 0.00 4.00 12.00 22.00 29.00 4 HR 64.33 67.00 69.00 71.00 77.00 4 ∆ HR 0.00 2.67 4.67 6.67 12.67 4 Skin temp 1 28.69 29.52 29.72 29.81 29.88 4 ∆ Skin temp 1 0.00 0.83 1.03 1.12 1.19 4 Skin temp 2 28.86 29.52 29.67 29.76 29.84 4 ∆ Skin temp 2 0.00 0.66 0.81 0.90 0.98 4 Skin temp 3 29.40 29.35 29.35 29.36 29.35 4 ∆ Skin temp 3 0.00 -0.05 -0.05 -0.04 -0.05 4 bursts 1.83 3.00 5.01 7.02 6.02 4 ∆bursts 0.00 -0.67 3.18 5.18 4.18 4 %∆bursts 0.00 -18.18 173.44 282.81 228.12 4 total act 234.37 508.65 576.82 1075.99 870.83 4 ∆total act 0.00 39.90 342.44 841.62 636.45 4 % ∆ total act 0.00 8.51 146.11 359.09 271.56 4 muscle temp 33.82 33.95 34.05 34.08 34.07 4 ∆mus temp 0.00 0.13 0.23 0.26 0.25 4 RPE 6 0 12 15 18 4 ∆RPE 0 0 6 9 12 4 REN TAP 59.04 57.70 55.05 53.70 48.95 4 ∆RENAL TAP 0.00 -1.34 -3.99 -5.34 -10.09 4 %∆REN TAP 0.00 -2.27 -6.76 -9.05 -17.09 4 REN CON 0.52 0.49 0.44 0.39 0.34 4 ∆REN CON 0.00 -0.03 -0.08 -0.12 -0.18 4 %∆ REN CON 0.00 -5.59 -15.64 -23.76 -33.91 4 CBF 1.00 0.90 1.11 1.94 1.94 4 ∆CBF 0.00 -0.11 0.11 0.94 0.94 4 %∆CBF 0.00 -10.70 10.60 93.62 93.22 4 cvc 0.009 0.008 0.009 0.014 0.014 4 ∆cvc 0.000 -0.001 0.000 0.005 0.005 4 %∆cvc 0.00 -13.73 0.07 62.30 54.03
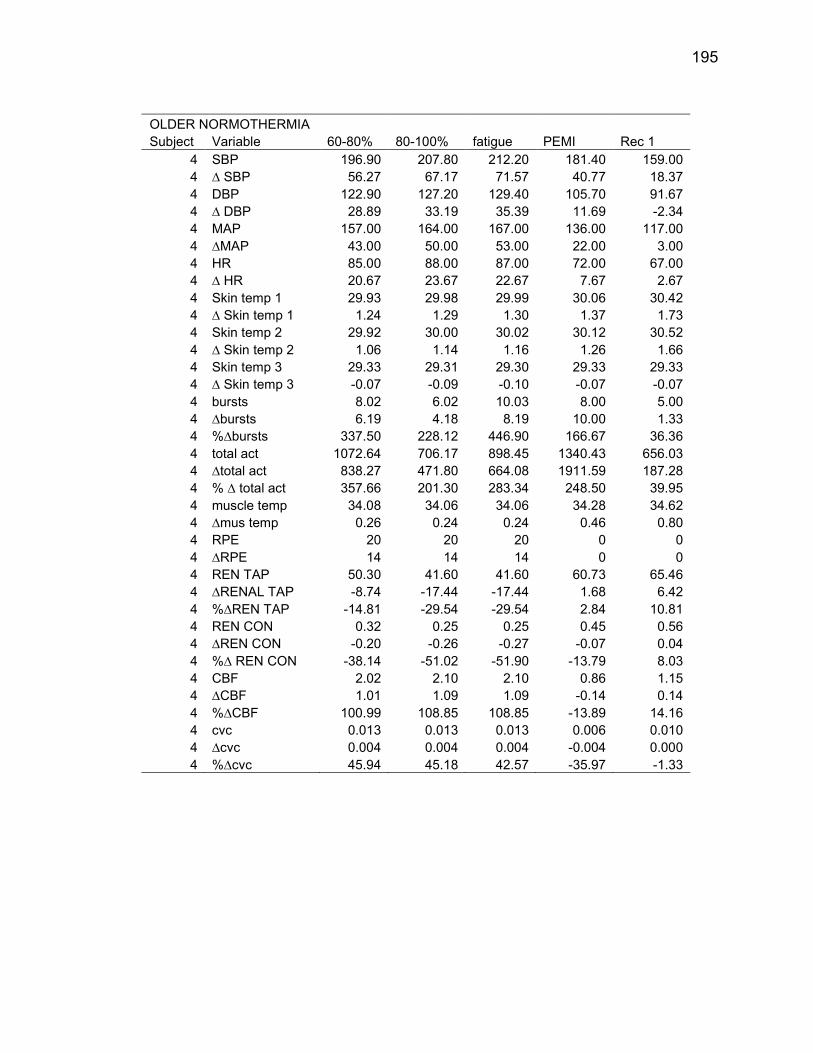
195
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
4 SBP 196.90 207.80 212.20 181.40 159.00 4 ∆ SBP 56.27 67.17 71.57 40.77 18.37 4 DBP 122.90 127.20 129.40 105.70 91.67 4 ∆ DBP 28.89 33.19 35.39 11.69 -2.34 4 MAP 157.00 164.00 167.00 136.00 117.00 4 ∆MAP 43.00 50.00 53.00 22.00 3.00 4 HR 85.00 88.00 87.00 72.00 67.00 4 ∆ HR 20.67 23.67 22.67 7.67 2.67 4 Skin temp 1 29.93 29.98 29.99 30.06 30.42 4 ∆ Skin temp 1 1.24 1.29 1.30 1.37 1.73 4 Skin temp 2 29.92 30.00 30.02 30.12 30.52 4 ∆ Skin temp 2 1.06 1.14 1.16 1.26 1.66 4 Skin temp 3 29.33 29.31 29.30 29.33 29.33 4 ∆ Skin temp 3 -0.07 -0.09 -0.10 -0.07 -0.07 4 bursts 8.02 6.02 10.03 8.00 5.00 4 ∆bursts 6.19 4.18 8.19 10.00 1.33 4 %∆bursts 337.50 228.12 446.90 166.67 36.36 4 total act 1072.64 706.17 898.45 1340.43 656.03 4 ∆total act 838.27 471.80 664.08 1911.59 187.28 4 % ∆ total act 357.66 201.30 283.34 248.50 39.95 4 muscle temp 34.08 34.06 34.06 34.28 34.62 4 ∆mus temp 0.26 0.24 0.24 0.46 0.80 4 RPE 20 20 20 0 0 4 ∆RPE 14 14 14 0 0 4 REN TAP 50.30 41.60 41.60 60.73 65.46 4 ∆RENAL TAP -8.74 -17.44 -17.44 1.68 6.42 4 %∆REN TAP -14.81 -29.54 -29.54 2.84 10.81 4 REN CON 0.32 0.25 0.25 0.45 0.56 4 ∆REN CON -0.20 -0.26 -0.27 -0.07 0.04 4 %∆ REN CON -38.14 -51.02 -51.90 -13.79 8.03 4 CBF 2.02 2.10 2.10 0.86 1.15 4 ∆CBF 1.01 1.09 1.09 -0.14 0.14 4 %∆CBF 100.99 108.85 108.85 -13.89 14.16 4 cvc 0.013 0.013 0.013 0.006 0.010 4 ∆cvc 0.004 0.004 0.004 -0.004 0.000 4 %∆cvc 45.94 45.18 42.57 -35.97 -1.33

196
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
151.10 150.80 4 SBP 136.50 137.30 151.00 10.47 10.17 4 ∆ SBP 0.00 0.80 14.50 90.63 89.38 4 DBP 85.47 86.43 93.15 -3.38 -4.63 4 ∆ DBP 0.00 0.96 7.68
115.00 114.00 4 MAP 105.67 106.00 116.00 1.00 0.00 4 ∆MAP 0.00 0.33 10.33
65.00 64.00 4 HR 67.67 68.00 72.00 0.67 -0.33 4 ∆ HR 0.00 0.33 4.33
30.61 30.78 4 Skin temp 1 39.36 39.45 39.92 1.92 2.09 4 ∆ Skin temp 1 0.00 0.09 0.56
30.73 30.83 4 Skin temp 2 38.74 38.82 39.16 1.87 1.97 4 ∆ Skin temp 2 0.00 0.08 0.42
29.34 29.35 4 Skin temp 3 29.93 29.82 29.80 -0.06 -0.05 4 ∆ Skin temp 3 0.00 -0.11 -0.13 3.00 4.00 4 bursts 2.83 2.00 4.62
-0.67 0.33 4 ∆bursts 0.00 -3.67 1.78 -18.18 9.09 4 %∆bursts 0.00 -64.71 62.99 433.00 356.06 4 total act 411.05 368.90 750.70 -35.75 -112.69 4 ∆total act 0.00 -453.20 339.65 -7.63 -24.04 4 % ∆ total act 0.00 -55.13 82.63 34.68 34.74 4 muscle temp 38.40 38.42 38.42
0.86 0.92 4 ∆mus temp 0.00 0.02 0.02 0 0 4 RPE 6 0 12 0 0 4 ∆RPE 0 0 6
63.08 65.40 4 REN TAP 59.62 54.33 56.35 4.03 6.36 4 ∆RENAL TAP 0.00 -5.29 -3.27 6.79 10.71 4 %∆REN TAP 0.00 -8.88 -5.48 0.55 0.57 4 REN CON 0.56 0.51 0.49 0.03 0.06 4 ∆REN CON 0.00 -0.05 -0.08 5.90 10.77 4 %∆ REN CON 0.00 -9.16 -13.90 1.25 0.81 4 CBF 1.06 1.48 1.19 0.25 -0.20 4 ∆CBF 0.00 0.42 0.13
24.46 -19.71 4 %∆CBF 0.00 39.24 12.30 0.011 0.007 4 cvc 0.010 0.014 0.010 0.001 -0.003 4 ∆cvc 0.000 0.004 0.000
9.45 -28.77 4 %∆cvc 0.00 38.80 2.29

197
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
4 SBP 154.40 167.70 178.60 205.40 215.90 4 ∆ SBP 17.90 31.20 42.10 68.90 79.40 4 DBP 94.52 104.40 112.90 129.10 138.00 4 ∆ DBP 9.05 18.93 27.43 43.63 52.53 4 MAP 118.00 131.00 144.00 166.00 176.00 4 ∆MAP 12.33 25.33 38.33 60.33 70.33 4 HR 75.00 76.00 87.00 93.00 93.00 4 ∆ HR 7.33 8.33 19.33 25.33 25.33 4 Skin temp 1 40.18 40.41 40.62 40.78 40.81 4 ∆ Skin temp 1 0.82 1.05 1.26 1.42 1.45 4 Skin temp 2 39.36 39.54 39.71 39.84 39.87 4 ∆ Skin temp 2 0.62 0.80 0.97 1.10 1.13 4 Skin temp 3 29.79 29.76 29.75 29.76 29.76 4 ∆ Skin temp 3 -0.14 -0.17 -0.18 -0.17 -0.17 4 bursts 7.39 10.16 11.08 11.08 11.08 4 ∆bursts 4.56 7.33 8.25 8.25 8.25 4 %∆bursts 160.78 258.57 291.17 291.17 291.13 4 total act 1504.82 1099.95 1936.70 1463.63 1583.78 4 ∆total act 1093.77 688.90 1525.65 1052.58 1172.73 4 % ∆ total act 266.09 167.59 371.16 256.07 285.30 4 muscle temp 38.44 38.46 38.49 38.53 38.53 4 ∆mus temp 0.04 0.06 0.09 0.13 0.13 4 RPE 15 17 18 20 20 4 ∆RPE 9 11 12 14 14 4 REN TAP 55.67 44.45 45.60 36.20 36.20 4 ∆RENAL TAP -3.95 -15.17 -14.02 -23.42 -23.42 4 %∆REN TAP -6.63 -25.44 -23.51 -39.28 -39.28 4 REN CON 0.47 0.34 0.32 0.22 0.21 4 ∆REN CON -0.09 -0.22 -0.25 -0.35 -0.36 4 %∆ REN CON -16.39 -39.86 -43.87 -61.35 -63.54 4 CBF 1.22 1.11 1.11 1.32 1.32 4 ∆CBF 0.16 0.05 0.05 0.26 0.26 4 %∆CBF 15.20 5.04 4.71 24.36 24.36 4 cvc 0.010 0.009 0.008 0.008 0.007 4 ∆cvc 0.000 -0.002 -0.002 -0.002 -0.003 4 %∆cvc 3.16 -15.27 -23.16 -20.84 -25.34

198
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
4 SBP 180.30 148.50 153.00 152.50 4 ∆ SBP 43.80 12.00 16.50 16.00 4 DBP 103.50 85.92 88.30 88.56 4 ∆ DBP 18.03 0.45 2.83 3.09 4 MAP 134.00 109.00 112.00 112.00 4 ∆MAP 28.33 3.33 6.33 6.33 4 HR 77.00 72.00 71.00 70.00 4 ∆ HR 9.33 4.33 3.33 2.33 4 Skin temp 1 40.92 40.83 40.14 39.73 4 ∆ Skin temp 1 1.56 1.47 0.78 0.37 4 Skin temp 2 40.00 40.21 39.85 39.48 4 ∆ Skin temp 2 1.26 1.47 1.11 0.74 4 Skin temp 3 29.74 29.72 29.69 29.66 4 ∆ Skin temp 3 -0.19 -0.21 -0.24 -0.27 4 bursts 5.00 6.00 3.00 8.00 4 ∆bursts 7.17 0.33 -2.67 2.33 4 %∆bursts 252.94 5.88 -47.06 41.18 4 total act 619.27 800.82 393.82 1036.89 4 ∆total act 469.28 -21.28 -428.28 214.79 4 % ∆ total act 114.17 -2.59 -52.10 26.13 4 muscle temp 27.77 38.78 38.75 38.75 4 ∆mus temp 0.24 0.38 0.35 0.35 4 RPE 0 0 0 0 4 ∆RPE 0 0 0 0 4 REN TAP 61.38 75.48 73.15 70.13 4 ∆RENAL TAP 1.76 15.86 13.53 10.51 4 %∆REN TAP 2.95 26.60 22.70 17.63 4 REN CON 0.46 0.69 0.65 0.63 4 ∆REN CON -0.11 0.13 0.09 0.06 4 %∆ REN CON -18.82 22.73 15.76 10.98 4 CBF 1.63 1.14 0.94 1.24 4 ∆CBF 0.56 0.08 -0.12 0.18 4 %∆CBF 53.19 7.25 -11.55 17.07 4 cvc 0.012 0.010 0.008 0.011 4 ∆cvc 0.002 0.000 -0.002 0.001 4 %∆cvc 20.80 3.97 -16.55 10.45

199
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
5 SBP 143.13 160.70 160.00 169.50 182.50 5 ∆ SBP 0.00 17.57 16.87 26.37 39.37 5 DBP 98.22 114.20 115.60 124.00 130.00 5 ∆ DBP 0.00 15.98 17.38 25.78 31.78 5 MAP 117.00 136.00 136.00 143.00 151.00 5 ∆MAP 0.00 19.00 19.00 26.00 34.00 5 HR 62.67 68.00 69.00 71.00 73.17 5 ∆ HR 0.00 5.33 6.33 8.33 10.50 5 Skin temp 1 30.19 30.17 30.22 30.28 30.32 5 ∆ Skin temp 1 0.00 -0.02 0.03 0.09 0.13 5 Skin temp 2 30.26 30.26 30.31 30.34 30.35 5 ∆ Skin temp 2 0.00 0.00 0.05 0.08 0.09 5 Skin temp 3 29.88 29.91 29.94 29.96 29.97 5 ∆ Skin temp 3 0.00 0.03 0.06 0.08 0.09 5 bursts 9.83 16.00 10.78 16.58 15.75 5 ∆bursts 0.00 -3.67 0.94 6.75 5.92 5 %∆bursts 0.00 -18.64 9.60 68.61 60.18 5 total act 1447.13 2321.01 1655.71 1881.73 1916.78 5 ∆total act 0.00 -573.25 208.58 434.60 469.65 5 % ∆ total act 0.00 -19.81 14.41 30.03 32.45 5 muscle temp 33.97 33.68 33.64 33.57 33.48 5 ∆mus temp 0.00 -0.29 -0.33 -0.40 -0.49 5 RPE 6 0 13 15 17 5 ∆RPE 0 0 7 9 11 5 REN TAP 40.39 42.21 38.10 40.23 41.00 5 ∆RENAL TAP 0.00 1.82 -2.29 -0.16 0.61 5 %∆REN TAP 0.00 4.51 -5.67 -0.39 1.50 5 REN CON 0.35 0.31 0.28 0.28 0.27 5 ∆REN CON 0.00 -0.03 -0.07 -0.06 -0.07 5 %∆ REN CON 0.00 -10.09 -18.85 -18.50 -21.35 5 CBF 1.77 2.15 2.43 2.48 2.37 5 ∆CBF 0.00 0.38 0.66 0.72 0.60 5 %∆CBF 0.00 21.52 37.26 40.48 34.00 5 cvc 0.015 0.016 0.018 0.017 0.016 5 ∆cvc 0.000 0.001 0.003 0.002 0.001 5 %∆cvc 0.00 4.55 18.08 14.94 3.83

200
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
5 SBP 191.50 201.30 199.10 180.60 145.80 5 ∆ SBP 48.37 58.17 55.97 37.47 2.67 5 DBP 134.40 144.50 144.20 127.40 101.90 5 ∆ DBP 36.18 46.28 45.98 29.18 3.68 5 MAP 158.00 169.00 170.00 148.00 119.00 5 ∆MAP 41.00 52.00 53.00 31.00 2.00 5 HR 76.00 79.33 80.50 72.00 68.00 5 ∆ HR 13.33 16.67 17.83 9.33 5.33 5 Skin temp 1 30.34 30.34 30.34 30.26 30.12 5 ∆ Skin temp 1 0.15 0.15 0.15 0.07 -0.07 5 Skin temp 2 30.37 30.38 30.37 30.24 30.18 5 ∆ Skin temp 2 0.11 0.12 0.11 -0.02 -0.08 5 Skin temp 3 29.96 29.96 29.96 29.91 29.82 5 ∆ Skin temp 3 0.08 0.08 0.08 0.03 -0.06 5 bursts 16.58 14.92 14.92 13.00 0.00 5 ∆bursts 6.75 5.09 5.09 4.00 -19.67 5 %∆bursts 68.61 51.75 51.75 18.18 -100.00 5 total act 1916.00 1916.00 1916.00 2765.90 0.00 5 ∆total act 468.87 468.87 468.87 2449.36 -2894.26 5 % ∆ total act 32.40 32.40 32.40 79.46 -100.00 5 muscle temp 33.44 33.39 33.39 33.30 33.59 5 ∆mus temp -0.53 -0.58 -0.58 -0.67 -0.38 5 RPE 20 20 20 0 0 5 ∆RPE 14 14 14 0 0 5 REN TAP 42.20 44.93 44.55 46.85 49.53 5 ∆RENAL TAP 1.81 4.54 4.16 6.46 9.13 5 %∆REN TAP 4.48 11.24 10.29 16.71 23.63 5 REN CON 0.27 0.27 0.26 0.32 0.42 5 ∆REN CON -0.08 -0.08 -0.08 -0.03 0.07 5 %∆ REN CON -22.64 -22.99 -24.09 -8.31 20.55 5 CBF 2.58 2.70 1.85 4.28 2.40 5 ∆CBF 0.82 0.94 0.08 2.52 0.63 5 %∆CBF 46.18 52.99 4.43 142.45 35.66 5 cvc 0.016 0.016 0.011 0.029 0.020 5 ∆cvc 0.001 0.001 -0.004 0.015 0.006 5 %∆cvc 8.25 5.92 -28.13 111.60 47.25

201
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
146.80 147.70 5 SBP 126.73 127.30 148.70 3.67 4.57 5 ∆ SBP 0.00 0.57 21.97
101.10 101.50 5 DBP 77.39 76.38 86.47 2.88 3.28 5 ∆ DBP 0.00 -1.01 9.08
120.00 122.00 5 MAP 96.00 99.00 114.00 3.00 5.00 5 ∆MAP 0.00 3.00 18.00
64.00 65.00 5 HR 64.00 71.00 71.00 1.33 2.33 5 ∆ HR 0.00 7.00 7.00
30.17 30.38 5 Skin temp 1 39.12 39.24 39.36 -0.02 0.19 5 ∆ Skin temp 1 0.00 0.12 0.24 30.33 30.53 5 Skin temp 2 39.40 39.56 39.67
0.07 0.27 5 ∆ Skin temp 2 0.00 0.16 0.27 29.80 29.82 5 Skin temp 3 29.79 29.75 29.72 -0.08 -0.06 5 ∆ Skin temp 3 0.00 -0.04 -0.07 0.00 0.00 5 bursts 9.83 23.00 10.23
-19.67 -19.67 5 ∆bursts 0.00 3.33 0.40 -100.00 -100.00 5 %∆bursts 0.00 16.95 4.03
0.00 0.00 5 total act 1447.13 4938.00 2771.98 -2894.26 -2894.26 5 ∆total act 0.00 2043.74 1324.85 -100.00 -100.00 5 % ∆ total act 0.00 70.61 91.55
34.31 34.59 5 muscle temp 38.53 38.67 38.97 0.34 0.62 5 ∆mus temp 0.00 0.14 0.44
0 0 5 RPE 6 0 11 0 0 5 ∆RPE 0 0 5
52.43 51.55 5 REN TAP 54.15 52.98 46.47 12.03 11.16 5 ∆RENAL TAP 0.00 -1.17 -7.68 31.13 28.87 5 %∆REN TAP 0.00 -2.16 -14.19
0.44 0.42 5 REN CON 0.56 0.54 0.41 0.09 0.08 5 ∆REN CON 0.00 -0.03 -0.16
26.55 22.39 5 %∆ REN CON 0.00 -5.13 -27.74 2.02 2.10 5 CBF 1.84 2.60 2.59 0.25 0.34 5 ∆CBF 0.00 0.76 0.75
14.22 19.07 5 %∆CBF 0.00 41.27 40.90 0.017 0.017 5 cvc 0.019 0.026 0.023 0.003 0.004 5 ∆cvc 0.000 0.007 0.004 22.95 26.07 5 %∆cvc 0.00 36.99 18.65

202
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
5 SBP 162.80 178.70 187.60 203.00 209.90 5 ∆ SBP 36.07 51.97 60.87 76.27 83.17 5 DBP 96.35 103.00 111.50 119.30 124.10 5 ∆ DBP 18.96 25.61 34.11 41.91 46.71 5 MAP 125.00 135.00 143.00 153.00 158.00 5 ∆MAP 29.00 39.00 47.00 57.00 62.00 5 HR 77.00 78.00 87.00 93.00 94.00 5 ∆ HR 13.00 14.00 23.00 29.00 30.00 5 Skin temp 1 39.44 39.52 39.60 39.66 39.67 5 ∆ Skin temp 1 0.32 0.40 0.48 0.54 0.55 5 Skin temp 2 39.74 39.83 39.93 40.03 40.06 5 ∆ Skin temp 2 0.34 0.43 0.53 0.63 0.66 5 Skin temp 3 29.70 29.68 29.66 29.64 29.63 5 ∆ Skin temp 3 -0.09 -0.11 -0.13 -0.15 -0.16 5 bursts 21.39 25.11 31.62 34.41 37.20 5 ∆bursts 11.56 15.28 21.78 24.57 27.36 5 %∆bursts 117.51 155.34 221.54 249.91 278.28 5 total act 5943.37 7954.46 11513.08 14949.32 15281.96 5 ∆total act 4496.24 6507.33 10065.95 13502.19 13834.83 5 % ∆ total act 310.70 449.67 695.58 933.03 956.02 5 muscle temp 39.02 39.10 39.13 39.21 39.21 5 ∆mus temp 0.49 0.57 0.60 0.68 0.68 5 RPE 13 17 19 20 20 5 ∆RPE 7 11 13 14 14 5 REN TAP 46.90 50.40 45.75 38.10 38.10 5 ∆RENAL TAP -7.25 -3.75 -8.40 -16.05 -16.05 5 %∆REN TAP -13.39 -6.93 -15.51 -29.64 -29.64 5 REN CON 0.38 0.37 0.32 0.25 0.24 5 ∆REN CON -0.19 -0.19 -0.24 -0.32 -0.32 5 %∆ REN CON -33.48 -33.81 -43.28 -55.85 -57.25 5 CBF 2.77 2.23 2.32 2.36 2.36 5 ∆CBF 0.93 0.39 0.48 0.52 0.52 5 %∆CBF 50.61 21.08 25.97 28.23 28.23 5 cvc 0.022 0.016 0.016 0.015 0.015 5 ∆cvc 0.003 -0.003 -0.003 -0.004 -0.004 5 %∆cvc 15.67 -13.90 -15.43 -19.54 -22.09

203
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
5 SBP 182.80 148.90 144.90 134.30 5 ∆ SBP 56.07 22.17 18.17 7.57 5 DBP 107.90 85.24 79.24 72.51 5 ∆ DBP 30.51 7.85 1.85 -4.88 5 MAP 137.00 108.00 102.00 94.00 5 ∆MAP 41.00 12.00 6.00 -2.00 5 HR 79.00 70.00 66.00 66.00 5 ∆ HR 15.00 6.00 2.00 2.00 5 Skin temp 1 39.71 39.53 38.69 38.32 5 ∆ Skin temp 1 0.59 0.41 -0.43 -0.80 5 Skin temp 2 40.21 40.26 39.68 39.22 5 ∆ Skin temp 2 0.81 0.86 0.28 -0.18 5 Skin temp 3 29.57 29.35 29.22 29.16 5 ∆ Skin temp 3 -0.22 -0.44 -0.57 -0.63 5 bursts 26.00 26.00 35.00 31.00 5 ∆bursts 42.17 6.33 15.33 11.33 5 %∆bursts 428.81 32.20 77.97 57.63 5 total act 5531.80 4398.18 6199.90 5784.18 5 ∆total act 7981.15 1503.92 3305.64 2889.92 5 % ∆ total act 551.52 51.96 114.21 99.85 5 muscle temp 38.11 39.30 39.00 39.50 5 ∆mus temp 0.80 0.77 0.47 0.97 5 RPE 0 0 0 0 5 ∆RPE 0 0 0 0 5 REN TAP 51.55 54.23 58.60 57.73 5 ∆RENAL TAP -2.60 0.07 4.45 3.58 5 %∆REN TAP -4.80 0.14 8.22 6.60 5 REN CON 0.38 0.50 0.57 0.61 5 ∆REN CON -0.19 -0.06 0.01 0.05 5 %∆ REN CON -33.29 -10.99 1.85 8.87 5 CBF 4.41 2.71 2.09 1.64 5 ∆CBF 2.57 0.87 0.26 -0.20 5 %∆CBF 139.77 47.20 13.89 -10.86 5 cvc 0.032 0.025 0.021 0.017 5 ∆cvc 0.013 0.006 0.001 -0.002 5 %∆cvc 68.01 30.84 7.19 -8.96

204
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
6 SBP 93.12 94.93 109.70 124.40 133.50 6 ∆ SBP 0.00 1.81 16.58 31.28 40.38 6 DBP 43.26 50.54 66.57 73.32 75.65 6 ∆ DBP 0.00 7.28 23.31 30.06 32.39 6 MAP 62.00 68.00 87.00 96.00 102.00 6 ∆MAP 0.00 6.00 25.00 34.00 40.00 6 HR 58.33 57.00 59.00 62.00 62.00 6 ∆ HR 0.00 -1.33 0.67 3.67 3.67 6 Skin temp 1 32.26 32.25 32.20 32.18 32.18 6 ∆ Skin temp 1 0.00 -0.01 -0.06 -0.08 -0.08 6 Skin temp 2 33.15 33.15 33.14 33.12 33.11 6 ∆ Skin temp 2 0.00 0.00 -0.01 -0.03 -0.04 6 Skin temp 3 31.24 31.24 31.26 31.25 31.25 6 ∆ Skin temp 3 0.00 0.00 0.02 0.01 0.01 6 bursts 3.17 8.00 5.34 6.52 16.01 6 ∆bursts 0.00 1.67 2.17 3.36 12.84 6 %∆bursts 0.00 26.32 68.50 105.95 405.51 6 total act 556.90 1163.48 784.91 1003.62 3130.09 6 ∆total act 0.00 49.68 228.01 446.71 2573.18 6 % ∆ total act 0.00 4.46 40.94 80.21 462.05 6 muscle temp 35.05 35.01 35.04 35.04 35.05 6 ∆mus temp 0.00 -0.04 -0.01 -0.01 0.00 6 RPE 6 0 11 14 16 6 ∆RPE 0 0 5 8 10 6 REN TAP 142.00 138.75 134.40 135.00 128.25 6 ∆RENAL TAP 0.00 -3.25 -7.60 -7.00 -13.75 6 %∆REN TAP 0.00 -2.29 -5.35 -4.93 -9.68 6 REN CON 2.29 2.04 1.54 1.41 1.26 6 ∆REN CON 0.00 -0.25 -0.75 -0.88 -1.03 6 %∆ REN CON 0.00 -10.91 -32.55 -38.60 -45.10 6 CBF 1.67 1.17 1.45 1.84 1.76 6 ∆CBF 0.00 -0.51 -0.23 0.17 0.09 6 %∆CBF 0.00 -30.24 -13.50 10.25 5.37 6 cvc 0.027 0.017 0.017 0.019 0.017 6 ∆cvc 0.000 -0.010 -0.010 -0.008 -0.010 6 %∆cvc 0.00 -36.40 -38.36 -28.80 -35.95

205
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
6 SBP 148.00 157.20 157.20 142.30 120.30 6 ∆ SBP 54.88 64.08 64.08 49.18 27.18 6 DBP 82.87 87.45 87.42 73.19 61.64 6 ∆ DBP 39.61 44.19 44.16 29.93 18.38 6 MAP 110.00 117.00 117.00 103.00 89.00 6 ∆MAP 48.00 55.00 55.00 41.00 27.00 6 HR 68.00 71.00 72.00 63.00 63.00 6 ∆ HR 9.67 12.67 13.67 4.67 4.67 6 Skin temp 1 32.16 32.15 32.15 32.17 32.20 6 ∆ Skin temp 1 -0.10 -0.11 -0.11 -0.09 -0.06 6 Skin temp 2 33.10 33.09 33.09 33.08 33.10 6 ∆ Skin temp 2 -0.05 -0.06 -0.06 -0.07 -0.05 6 Skin temp 3 31.24 31.23 31.23 31.22 31.18 6 ∆ Skin temp 3 0.00 -0.01 -0.01 -0.02 -0.06 6 bursts 19.57 20.16 17.70 17.50 8.00 6 ∆bursts 16.40 16.99 14.53 30.00 1.67 6 %∆bursts 517.84 536.57 458.95 600.00 26.32 6 total act 4589.45 4608.15 4560.70 2570.18 877.72 6 ∆total act 4032.55 4051.25 4003.80 4244.16 -236.08 6 % ∆ total act 724.10 727.46 718.94 473.57 -21.20 6 muscle temp 35.05 35.06 35.09 35.05 35.44 6 ∆mus temp 0.00 0.01 0.04 0.00 0.39 6 RPE 19 19 0 0 0 6 ∆RPE 13 13 0 0 0 6 REN TAP 123.00 98.54 93.10 120.75 131.00 6 ∆RENAL TAP -19.00 -43.46 -48.90 -21.25 -11.00 6 %∆REN TAP -13.38 -30.61 -34.44 -14.31 -7.41 6 REN CON 1.12 0.84 0.80 1.17 1.47 6 ∆REN CON -1.17 -1.45 -1.49 -1.12 -0.82 6 %∆ REN CON -51.18 -63.23 -65.26 -48.81 -35.73 6 CBF 2.04 1.80 2.06 1.92 1.58 6 ∆CBF 0.36 0.13 0.38 0.25 -0.09 6 %∆CBF 21.71 7.85 22.89 14.89 -5.44 6 cvc 0.019 0.015 0.018 0.019 0.018 6 ∆cvc -0.008 -0.012 -0.009 -0.008 -0.009 6 %∆cvc -31.40 -42.85 -34.88 -29.68 -33.02

206
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
112.40 118.10 6 SBP 93.12 114.70 108.70 19.28 24.98 6 ∆ SBP 0.00 21.58 15.58 60.54 64.36 6 DBP 43.26 46.91 56.67 17.28 21.10 6 ∆ DBP 0.00 3.65 13.41 85.00 88.00 6 MAP 62.00 77.00 80.00 23.00 26.00 6 ∆MAP 0.00 15.00 18.00 56.00 58.00 6 HR 51.00 53.00 56.00 -2.33 -0.33 6 ∆ HR 0.00 2.00 5.00 32.33 32.47 6 Skin temp 1 40.34 40.24 40.27
0.07 0.21 6 ∆ Skin temp 1 0.00 -0.10 -0.07 33.23 33.33 6 Skin temp 2 39.61 39.57 39.66
0.08 0.18 6 ∆ Skin temp 2 0.00 -0.04 0.05 31.16 31.13 6 Skin temp 3 30.58 30.57 30.56 -0.08 -0.11 6 ∆ Skin temp 3 0.00 -0.01 -0.02 12.00 16.00 6 bursts 2.50 6.00 2.76
5.67 9.67 6 ∆bursts 0.00 1.00 0.26 89.47 152.63 6 %∆bursts 0.00 20.00 10.40
1708.05 2157.59 6 total act 390.13 832.46 306.00 594.25 1043.79 6 ∆total act 0.00 52.21 -84.13 53.35 93.71 6 % ∆ total act 0.00 6.69 -21.56 35.70 35.75 6 muscle temp 39.12 39.08 39.19
0.65 0.70 6 ∆mus temp 0.00 -0.04 0.07 0 0 6 RPE 6 0 8 0 0 6 ∆RPE 0 0 2
142.80 141.00 6 REN TAP 133.60 121.50 131.00 0.80 -1.00 6 ∆RENAL TAP 0.00 -12.10 -2.60 0.54 -0.67 6 %∆REN TAP 0.00 -9.06 -1.95 1.68 1.60 6 REN CON 2.15 1.58 1.64
-0.61 -0.69 6 ∆REN CON 0.00 -0.58 -0.52 -26.65 -30.04 6 %∆ REN CON 0.00 -26.77 -24.01
1.62 1.48 6 CBF 1.24 1.23 1.32 -0.05 -0.19 6 ∆CBF 0.00 -0.01 0.08 -3.14 -11.57 6 %∆CBF 0.00 -0.97 6.60 0.019 0.017 6 cvc 0.020 0.016 0.017
-0.007 -0.010 6 ∆cvc 0.000 -0.004 -0.003 -28.16 -36.65 6 %∆cvc 0.00 -20.26 -17.39

207
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
6 SBP 118.50 154.40 172.40 187.60 192.00 6 ∆ SBP 25.38 61.28 79.28 94.48 98.88 6 DBP 67.03 81.50 87.22 99.25 102.80 6 ∆ DBP 23.77 38.24 43.96 55.99 59.54 6 MAP 90.00 109.00 120.00 134.00 138.00 6 ∆MAP 28.00 47.00 58.00 72.00 76.00 6 HR 60.00 66.00 69.00 76.00 78.00 6 ∆ HR 9.00 15.00 18.00 25.00 27.00 6 Skin temp 1 40.28 40.29 40.30 40.29 40.29 6 ∆ Skin temp 1 -0.06 -0.05 -0.04 -0.05 -0.05 6 Skin temp 2 39.71 39.74 39.78 39.80 39.80 6 ∆ Skin temp 2 0.10 0.13 0.17 0.19 0.19 6 Skin temp 3 30.56 30.56 30.56 30.55 30.55 6 ∆ Skin temp 3 -0.02 -0.02 -0.02 -0.03 -0.03 6 bursts 11.04 18.40 25.76 27.60 16.56 6 ∆bursts 8.54 15.90 23.26 25.10 14.06 6 %∆bursts 341.60 636.00 930.40 1004.00 562.40 6 total act 1585.66 3155.79 5529.71 8554.89 4793.94 6 ∆total act 1195.53 2765.67 5139.58 8164.77 4403.81 6 % ∆ total act 306.45 708.92 1317.41 2092.85 1128.82 6 muscle temp 39.22 39.26 39.32 39.40 39.40 6 ∆mus temp 0.10 0.14 0.20 0.28 0.28 6 RPE 9 11 15 19 19 6 ∆RPE 3 5 9 13 13 6 REN TAP 126.00 111.00 114.00 86.10 72.15 6 ∆RENAL TAP -7.60 -22.60 -19.60 -47.50 -61.45 6 %∆REN TAP -5.69 -16.92 -14.67 -35.55 -46.00 6 REN CON 1.40 1.02 0.95 0.64 0.52 6 ∆REN CON -0.75 -1.14 -1.20 -1.51 -1.63 6 %∆ REN CON -35.03 -52.74 -55.91 -70.18 -75.74 6 CBF 1.52 1.32 1.41 1.35 1.35 6 ∆CBF 0.28 0.08 0.16 0.11 0.11 6 %∆CBF 22.22 6.28 13.12 9.05 9.01 6 cvc 0.017 0.012 0.012 0.010 0.010 6 ∆cvc -0.003 -0.008 -0.008 -0.010 -0.010 6 %∆cvc -15.81 -39.55 -41.55 -49.54 -51.02

208
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
6 SBP 168.80 129.60 129.40 123.10 6 ∆ SBP 75.68 36.48 36.28 29.98 6 DBP 78.51 61.25 68.47 65.30 6 ∆ DBP 35.25 17.99 25.21 22.04 6 MAP 115.00 90.00 94.00 90.00 6 ∆MAP 53.00 28.00 32.00 28.00 6 HR 65.00 67.00 73.00 70.00 6 ∆ HR 14.00 16.00 22.00 19.00 6 Skin temp 1 40.33 40.52 40.27 39.88 6 ∆ Skin temp 1 -0.01 0.18 -0.07 -0.46 6 Skin temp 2 39.83 39.95 39.60 39.26 6 ∆ Skin temp 2 0.22 0.34 -0.01 -0.35 6 Skin temp 3 30.54 30.52 30.52 30.52 6 ∆ Skin temp 3 -0.04 -0.06 -0.06 -0.06 6 bursts 20.50 15.00 7.00 10.00 6 ∆bursts 38.50 10.00 2.00 5.00 6 %∆bursts 1540.00 200.00 40.00 100.00 6 total act 4262.62 3052.90 1292.36 1080.42 6 ∆total act 7629.03 2272.65 512.10 300.17 6 % ∆ total act 1955.53 291.27 65.63 38.47 6 muscle temp 39.43 39.01 38.60 38.43 6 ∆mus temp 0.31 -0.11 -0.52 -0.69 6 RPE 0 0 0 0 6 ∆RPE 0 0 0 0 6 REN TAP 102.13 112.50 120.00 134.25 6 ∆RENAL TAP -31.47 -21.10 -13.60 0.65 6 %∆REN TAP -23.55 -15.79 -10.18 0.49 6 REN CON 0.89 1.25 1.28 1.49 6 ∆REN CON -1.27 -0.90 -0.88 -0.66 6 %∆ REN CON -58.79 -41.99 -40.76 -30.78 6 CBF 2.20 1.52 1.36 1.50 6 ∆CBF 0.95 0.28 0.12 0.25 6 %∆CBF 76.80 22.40 9.66 20.47 6 cvc 0.019 0.017 0.014 0.017 6 ∆cvc -0.001 -0.003 -0.006 -0.003 6 %∆cvc -4.68 -15.68 -27.67 -17.01

209
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
7 SBP 161.20 166.80 177.90 190.90 194.80 7 ∆ SBP 0.00 5.60 16.70 29.70 33.60 7 DBP 58.53 66.39 72.58 75.26 79.07 7 ∆ DBP 0.00 7.86 14.05 16.73 20.54 7 MAP 92.00 102.00 109.00 114.00 121.00 7 ∆MAP 0.00 10.00 17.00 22.00 29.00 7 HR 83.67 88.00 89.00 89.00 88.00 7 ∆ HR 0.00 4.33 5.33 5.33 4.33 7 Skin temp 1 32.06 31.98 31.94 31.91 31.87 7 ∆ Skin temp 1 0.00 -0.08 -0.12 -0.15 -0.19 7 Skin temp 2 32.30 32.18 32.12 32.09 32.04 7 ∆ Skin temp 2 0.00 -0.12 -0.18 -0.21 -0.26 7 Skin temp 3 30.55 30.56 30.58 30.59 30.61 7 ∆ Skin temp 3 0.00 0.01 0.03 0.04 0.06 7 bursts 5.17 11.00 9.33 11.20 15.86 7 ∆bursts 0.00 0.67 4.16 6.03 10.70 7 %∆bursts 0.00 6.45 80.60 116.72 207.02 7 peak amp 622.20 1098.41 1029.80 1376.30 1911.18 7 ∆ peak amp 0.00 -145.99 407.60 754.10 1288.98 7 %∆ peak amp 0.00 -11.73 65.51 121.20 207.17 7 muscle temp 35.40 35.39 35.65 35.68 35.65 7 ∆muscle temp 0.00 -0.01 0.25 0.28 0.25 7 RPE 6 0 11 14 17 7 ∆RPE 0 0 5 8 11 7 REN TAP 75.64 59.73 61.00 61.00 55.90 7 ∆RENAL TAP 0.00 -15.92 -14.64 -14.64 -19.74 7 REN CON 0.82 0.59 0.56 0.54 0.46 7 ∆REN CON 0.00 -0.24 -0.26 -0.29 -0.36 7 %∆REN TAP 0.00 -21.04 -19.36 -19.36 -26.10 7 CBF 1.60 2.14 1.88 2.23 2.04 7 ∆CBF 0.00 0.54 0.28 0.63 0.44 7 %∆CBF 0.00 33.45 17.55 39.08 27.39 7 cvc 0.017 0.021 0.017 0.020 0.017 7 ∆cvc 0.000 0.004 0.000 0.002 -0.001 7 %∆cvc 0.00 20.37 -0.78 12.24 -3.14

210
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
7 SBP 198.30 193.00 191.50 169.40 162.10 7 ∆ SBP 37.10 31.80 30.30 8.20 0.90 7 DBP 80.47 77.01 78.16 65.88 55.72 7 ∆ DBP 21.94 18.48 19.63 7.35 -2.81 7 MAP 125.00 121.00 122.00 103.00 90.00 7 ∆MAP 33.00 29.00 30.00 11.00 -2.00 7 HR 86.00 84.00 84.00 71.00 81.00 7 ∆ HR 2.33 0.33 0.33 -12.67 -2.67 7 Skin temp 1 31.84 31.81 31.81 31.78 31.68 7 ∆ Skin temp 1 -0.22 -0.25 -0.25 -0.28 -0.38 7 Skin temp 2 32.00 31.97 31.96 31.92 31.85 7 ∆ Skin temp 2 -0.30 -0.33 -0.34 -0.38 -0.45 7 Skin temp 3 30.63 30.66 30.67 30.71 30.81 7 ∆ Skin temp 3 0.08 0.11 0.12 0.16 0.26 7 bursts 17.73 17.73 22.39 11.00 7 ∆bursts 12.56 12.56 17.23 5.83 7 %∆bursts 243.14 243.14 333.44 112.90 7 peak amp 2034.33 1994.67 2433.60 1098.41 7 ∆ peak amp 1412.13 1372.47 1811.40 476.21 7 %∆ peak amp 226.96 220.58 291.13 76.54 7 muscle temp 35.62 35.62 35.62 35.23 35.23 7 ∆muscle temp 0.22 0.22 0.22 -0.17 -0.17 7 RPE 19 20 20 0 0 7 ∆RPE 13 14 14 0 0 7 REN TAP 52.50 45.70 45.50 60.58 66.70 7 ∆RENAL TAP -23.14 -29.94 -30.14 -15.07 -8.94 7 REN CON 0.42 0.38 0.37 0.59 0.74 7 ∆REN CON -0.40 -0.44 -0.45 -0.23 -0.08 7 %∆REN TAP -30.59 -39.58 -39.85 -20.00 -11.87 7 CBF 2.40 2.17 2.04 2.11 1.60 7 ∆CBF 0.79 0.57 0.44 0.51 0.00 7 %∆CBF 49.64 35.80 27.27 32.08 0.01 7 cvc 0.019 0.018 0.017 0.021 0.018 7 ∆cvc 0.002 0.001 -0.001 0.002 -0.001 7 %∆cvc 10.13 3.25 -4.03 8.43 -6.04

211
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
165.80 169.00 7 SBP 145.37 163.70 169.60 4.60 7.80 7 ∆ SBP 0.00 18.33 24.23
55.91 57.87 7 DBP 52.85 61.05 69.39 -2.62 -0.66 7 ∆ DBP 0.00 8.20 16.54 90.00 93.00 7 MAP 86.33 99.00 107.00 -2.00 1.00 7 ∆MAP 0.00 12.67 20.67 83.00 84.00 7 HR 90.00 92.00 93.00 -0.67 0.33 7 ∆ HR 0.00 2.00 3.00 31.64 31.60 7 Skin temp 1 39.08 39.13 39.26 -0.42 -0.46 7 ∆ Skin temp 1 0.00 0.05 0.18 31.88 31.91 7 Skin temp 2 38.47 38.54 38.69 -0.42 -0.39 7 ∆ Skin temp 2 0.00 0.07 0.22 30.85 30.88 7 Skin temp 3 30.87 30.85 30.85
0.30 0.33 7 ∆ Skin temp 3 0.00 -0.02 -0.02 7 bursts 5.17 14.00 9.79 7 ∆bursts 0.00 3.67 4.62 7 %∆bursts 0.00 35.48 89.48 7 total act 622.20 1450.16 722.33 7 ∆total act 0.00 205.76 100.13 7 % ∆ total act 0.00 16.53 16.09
35.29 35.29 7 muscle temp 39.06 39.07 39.02 -0.11 -0.11 7 ∆mus temp 0.00 0.01 -0.04
0 0 7 RPE 6 0 12 0 0 7 ∆RPE 0 0 6
71.18 73.25 7 REN TAP 80.73 71.58 61.00 -4.47 -2.39 7 ∆RENAL TAP 0.00 -9.16 -19.73 0.79 0.79 7 REN CON 0.94 0.72 0.57
-0.03 -0.03 7 ∆REN CON 0.00 -0.21 -0.37 -5.93 -3.17 7 %∆REN TAP 0.00 -11.34 -24.44 1.30 1.43 7 CBF 1.69 1.94 2.34
-0.30 -0.17 7 ∆CBF 0.00 0.25 0.65 -18.72 -10.91 7 %∆CBF 0.00 14.73 38.63 0.014 0.015 7 cvc 0.020 0.020 0.022
-0.004 -0.004 7 ∆cvc 0.000 0.000 0.002 -23.63 -19.00 7 %∆cvc 0.00 0.05 11.85

212
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
7 SBP 197.50 197.60 197.70 194.50 192.20 7 ∆ SBP 52.13 52.23 52.33 49.13 46.83 7 DBP 80.69 80.72 80.75 78.85 78.40 7 ∆ DBP 27.84 27.87 27.90 26.00 25.55 7 MAP 118.00 121.00 124.00 122.00 120.00 7 ∆MAP 31.67 34.67 37.67 35.67 33.67 7 HR 91.00 91.00 91.00 91.00 91.00 7 ∆ HR 1.00 1.00 1.00 1.00 1.00 7 Skin temp 1 39.36 39.46 39.54 39.61 39.63 7 ∆ Skin temp 1 0.28 0.38 0.46 0.53 0.55 7 Skin temp 2 38.80 38.91 39.00 39.08 39.10 7 ∆ Skin temp 2 0.33 0.44 0.53 0.61 0.63 7 Skin temp 3 30.86 30.85 30.86 30.86 30.86 7 ∆ Skin temp 3 -0.01 -0.02 -0.01 -0.01 -0.01 7 bursts 16.91 12.46 16.91 16.91 19.58 7 ∆bursts 11.74 7.29 11.74 11.74 14.41 7 %∆bursts 227.29 141.16 227.29 227.29 278.97 7 total act 876.14 818.03 1633.47 1513.41 2019.90 7 ∆total act 253.94 195.83 1011.27 891.21 1397.70 7 % ∆ total act 40.81 31.47 162.53 143.23 224.64 7 muscle temp 39.04 39.07 39.09 39.16 39.25 7 ∆mus temp -0.02 0.01 0.03 0.10 0.19 7 RPE 14 17 19 20 20 7 ∆RPE 8 11 13 14 14 7 REN TAP 64.40 59.30 55.90 44.00 45.70 7 ∆RENAL TAP -16.33 -21.43 -24.83 -36.73 -35.03 7 REN CON 0.55 0.49 0.45 0.36 0.38 7 ∆REN CON -0.39 -0.45 -0.48 -0.57 -0.55 7 %∆REN TAP -20.23 -26.55 -30.76 -45.50 -43.39 7 CBF 2.37 2.16 2.10 2.14 1.91 7 ∆CBF 0.68 0.47 0.41 0.45 0.22 7 %∆CBF 40.47 27.57 24.08 26.39 13.16 7 cvc 0.020 0.018 0.017 0.018 0.016 7 ∆cvc 0.001 -0.002 -0.003 -0.002 -0.004 7 %∆cvc 2.78 -8.98 -13.61 -10.56 -18.59

213
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
7 SBP 158.90 150.80 145.60 151.10 7 ∆ SBP 13.53 5.43 0.23 5.73 7 DBP 62.68 52.68 54.15 54.96 7 ∆ DBP 9.83 -0.17 1.30 2.11 7 MAP 100.00 86.00 86.00 88.00 7 ∆MAP 13.67 -0.33 -0.33 1.67 7 HR 82.00 86.00 86.00 86.00 7 ∆ HR -8.00 -4.00 -4.00 -4.00 7 Skin temp 1 39.71 39.95 39.91 39.78 7 ∆ Skin temp 1 0.63 0.87 0.83 0.70 7 Skin temp 2 39.19 39.39 39.30 39.17 7 ∆ Skin temp 2 0.72 0.92 0.83 0.70 7 Skin temp 3 30.86 30.85 30.85 30.83 7 ∆ Skin temp 3 -0.01 -0.02 -0.02 -0.04 7 bursts 15.00 5.00 11.00 9.00 7 ∆bursts 24.83 -5.33 0.67 -1.33 7 %∆bursts 480.65 -51.61 6.45 -12.90 7 total act 1894.31 596.46 1330.55 972.28 7 ∆total act 2016.41 -647.94 86.15 -272.12 7 % ∆ total act 324.08 -52.07 6.92 -21.87 7 muscle temp 39.39 39.65 39.64 39.64 7 ∆mus temp 0.33 0.59 0.58 0.58 7 RPE 0 0 0 0 7 ∆RPE 0 0 0 0 7 REN TAP 65.68 72.30 78.33 80.45 7 ∆RENAL TAP -15.06 -8.43 -2.41 -0.28 7 REN CON 0.66 0.84 0.91 0.91 7 ∆REN CON -0.28 -0.09 -0.02 -0.02 7 %∆REN TAP -18.65 -10.45 -2.98 -0.35 7 CBF 2.63 1.77 1.51 1.67 7 ∆CBF 0.94 0.08 -0.18 -0.02 7 %∆CBF 55.89 4.61 -10.90 -1.09 7 cvc 0.026 0.021 0.018 0.019 7 ∆cvc 0.007 0.001 -0.002 -0.001 7 %∆cvc 34.59 5.01 -10.55 -2.96

214
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
8 SBP 128.80 131.20 139.50 143.10 154.00 8 ∆ SBP 0.00 2.40 10.70 14.30 25.20 8 DBP 62.57 62.52 68.78 73.58 79.83 8 ∆ DBP 0.00 -0.05 6.21 11.01 17.26 8 MAP 83.33 84.00 92.00 98.00 107.00 8 ∆MAP 0.00 0.67 8.67 14.67 23.67 8 HR 66.67 66.00 69.00 71.00 73.00 8 ∆ HR 0.00 -0.67 2.33 4.33 6.33 8 Skin temp 1 32.14 32.12 32.11 32.14 32.15 8 ∆ Skin temp 1 0.00 -0.02 -0.03 0.00 0.01 8 Skin temp 2 32.65 32.58 32.54 32.53 32.51 8 ∆ Skin temp 2 0.00 -0.07 -0.11 -0.12 -0.14 8 Skin temp 3 31.89 31.85 31.83 31.83 31.83 8 ∆ Skin temp 3 0.00 -0.04 -0.06 -0.06 -0.06 8 bursts 11.33 18.00 11.14 10.44 12.53 8 ∆bursts 0.00 -4.67 -0.20 -0.89 1.19 8 %∆bursts 0.00 -20.59 -1.74 -7.88 10.54 8 total act 1572.45 1796.16 1433.95 2877.19 2993.82 8 ∆total act 0.00 -1348.74 -138.50 1304.74 1421.37 8 % ∆ total act 0.00 -42.89 -8.81 82.98 90.39 8 muscle temp 35.04 35.03 35.10 35.09 35.08 8 ∆mus temp 0.00 -0.01 0.06 0.05 0.04 8 RPE 6 0 9 13 17 8 ∆RPE 0 0 3 7 11 8 REN TAP 0.00 8 ∆RENAL TAP 0.00 8 %∆REN TAP 0.00 8 REN CON 0.00 8 ∆REN CON 0.00 8 %∆ REN CON 0.00 8 CBF 1.74 1.80 1.88 1.63 1.49 8 ∆CBF 0.00 0.06 0.14 -0.11 -0.25 8 %∆CBF 0.00 3.50 8.03 -6.11 -14.41 8 cvc 0.021 0.021 0.020 0.017 0.014 8 ∆cvc 0.000 0.001 0.000 -0.004 -0.007 8 %∆cvc 0.00 2.68 -2.15 -20.17 -33.34

215
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
8 SBP 168.90 184.10 191.30 177.20 147.60 8 ∆ SBP 40.10 55.30 62.50 48.40 18.80 8 DBP 89.96 99.18 103.30 80.51 63.93 8 ∆ DBP 27.39 36.61 40.73 17.94 1.36 8 MAP 121.00 134.00 138.00 116.00 91.00 8 ∆MAP 37.67 50.67 54.67 32.67 7.67 8 HR 75.00 82.00 84.00 67.00 67.00 8 ∆ HR 8.33 15.33 17.33 0.33 0.33 8 Skin temp 1 32.15 32.14 32.15 32.18 32.16 8 ∆ Skin temp 1 0.01 0.00 0.01 0.04 0.02 8 Skin temp 2 32.47 32.42 32.42 32.41 32.57 8 ∆ Skin temp 2 -0.18 -0.23 -0.23 -0.24 -0.08 8 Skin temp 3 31.84 31.86 31.86 31.87 31.84 8 ∆ Skin temp 3 -0.05 -0.03 -0.03 -0.02 -0.05 8 bursts 14.62 16.70 22.30 12.00 0.00 8 ∆bursts 3.28 5.37 10.96 3.00 -22.67 8 %∆bursts 28.96 47.39 96.75 14.29 -100.00 8 total act 3285.88 3577.95 4776.21 0.00 0.00 8 ∆total act 1713.43 2005.49 3203.76 -2657.99 -3144.90 8 % ∆ total act 108.97 127.54 203.74 -100.00 -100.00 8 muscle temp 35.07 35.07 35.06 34.96 34.93 8 ∆mus temp 0.03 0.03 0.02 -0.08 -0.11 8 RPE 18 19 19 0 0 8 ∆RPE 12 13 13 0 0 8 REN TAP 8 ∆RENAL TAP 8 %∆REN TAP 8 REN CON 8 ∆REN CON 8 %∆ REN CON 8 CBF 1.16 1.11 1.22 2.08 2.23 8 ∆CBF -0.58 -0.63 -0.52 0.34 0.50 8 %∆CBF -33.36 -36.08 -29.76 19.47 28.61 8 cvc 0.010 0.008 0.009 0.018 0.025 8 ∆cvc -0.011 -0.013 -0.012 -0.004 0.003 8 %∆cvc -54.11 -60.25 -57.58 -17.13 13.71

216
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
145.40 150.20 8 SBP 128.20 130.90 140.90 16.60 21.40 8 ∆ SBP 0.00 2.70 12.70 66.25 70.31 8 DBP 66.11 67.11 72.35
3.68 7.74 8 ∆ DBP 0.00 1.00 6.24 94.00 98.00 8 MAP 85.67 87.00 94.00 10.67 14.67 8 ∆MAP 0.00 1.33 8.33 64.00 67.00 8 HR 65.33 67.00 69.00 -2.67 0.33 8 ∆ HR 0.00 1.67 3.67 32.18 32.20 8 Skin temp 1 39.88 39.69 39.83
0.04 0.06 8 ∆ Skin temp 1 0.00 -0.19 -0.05 32.93 33.24 8 Skin temp 2 40.64 40.46 40.69
0.28 0.59 8 ∆ Skin temp 2 0.00 -0.18 0.05 31.81 31.79 8 Skin temp 3 31.29 31.30 31.29 -0.08 -0.10 8 ∆ Skin temp 3 0.00 0.01 0.00 0.00 0.00 8 bursts 10.67 19.00 14.77
-22.67 -22.67 8 ∆bursts 0.00 -2.33 4.11 -100.00 -100.00 8 %∆bursts 0.00 -10.94 38.51
0.00 0.00 8 total act 2077.12 3107.28 2479.08 -3144.90 -3144.90 8 ∆total act 0.00 -1046.96 401.96 -100.00 -100.00 8 % ∆ total act 0.00 -25.20 19.35
35.02 35.05 8 muscle temp 38.59 38.42 38.38 -0.02 0.01 8 ∆mus temp 0.00 -0.17 -0.21
0 0 8 RPE 6 0 9 0 0 8 ∆RPE 0 0 3
8 REN TAP 0.00 8 ∆RENAL TAP 0.00 8 %∆REN TAP 0.00 8 REN CON 0.00 8 ∆REN CON 0.00 8 %∆ REN CON 0.00
1.22 1.54 8 CBF 1.60 1.49 1.40 -0.51 -0.20 8 ∆CBF 0.00 -0.11 -0.20
-29.61 -11.38 8 %∆CBF 0.00 -6.73 -12.28 0.013 0.016 8 cvc 0.019 0.017 0.015
-0.009 -0.006 8 ∆cvc 0.000 -0.002 -0.004 -39.75 -27.24 8 %∆cvc 0.00 -8.16 -20.05

217
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
8 SBP 146.40 196.60 182.80 206.80 218.40 8 ∆ SBP 18.20 68.40 54.60 78.60 90.20 8 DBP 75.80 85.80 100.30 114.00 121.80 8 ∆ DBP 9.69 19.69 34.19 47.89 55.69 8 MAP 99.00 121.00 131.00 150.00 159.00 8 ∆MAP 13.33 35.33 45.33 64.33 73.33 8 HR 70.00 72.00 79.00 85.00 88.00 8 ∆ HR 4.67 6.67 13.67 19.67 22.67 8 Skin temp 1 39.89 39.93 39.96 40.00 40.01 8 ∆ Skin temp 1 0.01 0.05 0.08 0.12 0.13 8 Skin temp 2 40.81 40.89 40.96 41.02 41.02 8 ∆ Skin temp 2 0.17 0.25 0.32 0.38 0.38 8 Skin temp 3 31.29 31.29 31.29 31.31 31.31 8 ∆ Skin temp 3 0.00 0.00 0.00 0.02 0.02 8 bursts 15.64 21.73 30.42 36.50 36.50 8 ∆bursts 4.98 11.06 19.75 25.83 25.83 8 %∆bursts 46.65 103.69 185.16 242.19 242.19 8 total act 3445.99 5905.18 11763.68 20999.18 24885.88 8 ∆total act 1368.87 3828.06 9686.56 18922.07 22808.76 8 % ∆ total act 65.90 184.30 466.35 910.98 1098.10 8 muscle temp 38.43 38.47 38.52 38.59 38.73 8 ∆mus temp -0.16 -0.12 -0.07 0.00 0.14 8 RPE 12 17 19 20 20 8 ∆RPE 6 11 13 14 14 8 REN TAP 8 ∆RENAL TAP 8 %∆REN TAP 8 REN CON 8 ∆REN CON 8 %∆ REN CON 8 CBF 1.25 1.14 1.16 1.13 1.13 8 ∆CBF -0.35 -0.46 -0.44 -0.47 -0.47 8 %∆CBF -22.03 -28.92 -27.78 -29.61 -29.62 8 cvc 0.013 0.009 0.009 0.008 0.007 8 ∆cvc -0.006 -0.009 -0.010 -0.011 -0.012 8 %∆cvc -32.53 -49.68 -52.77 -59.80 -62.08

218
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
8 SBP 197.30 158.30 148.40 144.20 8 ∆ SBP 69.10 30.10 20.20 16.00 8 DBP 92.85 69.69 67.20 66.75 8 ∆ DBP 26.74 3.58 1.09 0.64 8 MAP 131.00 99.00 94.00 92.00 8 ∆MAP 45.33 13.33 8.33 6.33 8 HR 69.00 67.00 67.00 67.00 8 ∆ HR 3.67 1.67 1.67 1.67 8 Skin temp 1 39.98 40.00 39.72 39.45 8 ∆ Skin temp 1 0.10 0.12 -0.16 -0.43 8 Skin temp 2 41.04 40.55 39.87 39.64 8 ∆ Skin temp 2 0.40 -0.09 -0.77 -1.00 8 Skin temp 3 31.31 31.28 31.25 31.24 8 ∆ Skin temp 3 0.02 -0.01 -0.04 -0.05 8 bursts 25.00 27.00 31.00 34.00 8 ∆bursts 39.33 5.67 9.67 12.67 8 %∆bursts 368.75 26.56 45.31 59.38 8 total act 9273.60 4066.61 4217.94 4527.05 8 ∆total act 15889.21 -87.63 63.70 372.81 8 % ∆ total act 764.96 -2.11 1.53 8.97 8 muscle temp 38.85 38.92 38.82 38.82 8 ∆mus temp 0.26 0.33 0.23 0.23 8 RPE 0 0 0 0 8 ∆RPE 0 0 0 0 8 REN TAP 8 ∆RENAL TAP 8 %∆REN TAP 8 REN CON 8 ∆REN CON 8 %∆ REN CON 8 CBF 3.40 1.74 1.37 1.31 8 ∆CBF 1.80 0.14 -0.23 -0.29 8 %∆CBF 112.58 8.55 -14.38 -18.13 8 cvc 0.026 0.018 0.015 0.014 8 ∆cvc 0.007 -0.001 -0.004 -0.004 8 %∆cvc 39.02 -6.07 -21.97 -23.76

219
OLDER NORMOTHERMIA Subject Variable BL AVE ISCH 0-20% 20-40% 40-60%
9 SBP 132.23 134.10 142.60 151.00 158.60 9 ∆ SBP 0.00 1.87 10.37 18.77 26.37 9 DBP 55.38 58.42 62.47 66.75 71.90 9 ∆ DBP 0.00 3.04 7.09 11.37 16.52 9 MAP 80.33 82.00 90.00 95.00 101.00 9 ∆MAP 0.00 1.67 9.67 14.67 20.67 9 HR 73.67 75.00 83.00 83.00 81.00 9 ∆ HR 0.00 1.33 9.33 9.33 7.33 9 Skin temp 1 33.28 33.35 33.29 33.24 33.19 9 ∆ Skin temp 1 0.00 0.07 0.01 -0.04 -0.09 9 Skin temp 2 33.24 33.28 33.26 33.25 33.23 9 ∆ Skin temp 2 0.00 0.04 0.02 0.01 -0.01 9 Skin temp 3 31.63 31.54 31.56 31.52 31.53 9 ∆ Skin temp 3 0.00 -0.09 -0.07 -0.11 -0.10 9 bursts 9.83 13.00 7.64 6.79 6.79 9 ∆bursts 0.00 3.17 -2.19 -3.04 -3.04 9 %∆bursts 0.00 32.20 -22.28 -30.91 -30.91 9 total act 1023.52 1287.53 783.47 720.07 739.31 9 ∆total act 0.00 264.01 -240.05 -303.46 -284.21 9 % ∆ total act 0.00 25.79 -23.45 -29.65 -27.77 9 RPE 6 0 12 15 18 9 ∆RPE 0 0 6 9 12 9 Muscle temp 36.54 36.56 36.55 36.55 36.53 9 ∆Muscle temp 0.00 0.02 0.01 0.01 -0.01 9 RBV 61.92 65.50 52.10 52.70 56.27 9 ∆RBV 0.00 3.58 -9.82 -9.22 -5.65 9 %∆RBV 0.00 5.78 -15.86 -14.89 -9.13 9 RVC 1.86 1.97 1.57 1.58 1.69 9 ∆RVC 0.00 0.11 -0.30 -0.28 -0.17 9 %∆RVC 0.00 5.64 -15.92 -14.92 -9.11 9 CBF 1.91 2.13 2.57 3.84 3.14 9 ∆CBF 0.00 0.21 0.66 1.93 1.22 9 %∆CBF 0.00 11.13 34.54 100.82 63.99 9 CVC 0.024 0.026 0.029 0.040 0.031 9 ∆CVC 0.000 0.002 0.005 0.017 0.007 9 %∆CVC 0.00 8.87 20.09 69.82 30.43

220
OLDER NORMOTHERMIA Subject Variable 60-80% 80-100% fatigue PEMI Rec 1
9 SBP 173.90 185.00 188.80 155.50 133.80 9 ∆ SBP 41.67 52.77 56.57 23.27 1.57 9 DBP 79.58 85.05 87.08 70.26 64.07 9 ∆ DBP 24.20 29.67 31.70 14.88 8.69 9 MAP 112.00 119.00 122.00 97.00 84.00 9 ∆MAP 31.67 38.67 41.67 16.67 3.67 9 HR 83.00 83.00 82.00 71.00 69.00 9 ∆ HR 9.33 9.33 8.33 -2.67 -4.67 9 Skin temp 1 33.14 33.10 33.09 33.07 33.32 9 ∆ Skin temp 1 -0.14 -0.18 -0.19 -0.21 0.04 9 Skin temp 2 33.22 33.19 33.19 33.18 33.29 9 ∆ Skin temp 2 -0.02 -0.05 -0.05 -0.06 0.05 9 Skin temp 3 31.54 31.46 31.44 31.46 31.47 9 ∆ Skin temp 3 -0.09 -0.17 -0.19 -0.17 -0.16 9 bursts 7.64 12.74 18.68 12.00 9.50 9 ∆bursts -2.19 2.90 8.85 2.17 -0.33 9 %∆bursts -22.28 29.54 89.99 22.03 -3.39 9 total act 886.74 825.50 521.02 1699.08 967.32 9 ∆total act -136.79 -198.02 -502.50 675.56 -56.21 9 % ∆ total act -13.36 -19.35 -49.09 66.00 -5.49 9 RPE 19 20 20 0 0 9 ∆RPE 13 14 14 0 0 9 Muscle temp 36.51 36.49 36.49 36.45 36.52 9 ∆Muscle temp -0.03 -0.05 -0.05 -0.09 -0.02 9 RBV 58.83 53.90 52.10 58.63 68.80 9 ∆RBV -3.09 -8.02 -9.82 -3.29 6.88 9 %∆RBV -5.00 -12.95 -15.86 -5.31 11.11 9 RVC 1.77 1.62 1.57 1.77 2.07 9 ∆RVC -0.09 -0.24 -0.29 -0.10 0.20 9 %∆RVC -4.95 -12.83 -15.74 -5.15 10.93 9 CBF 2.29 2.48 2.58 2.85 2.57 9 ∆CBF 0.37 0.57 0.66 0.94 0.65 9 %∆CBF 19.59 29.63 34.75 49.02 34.11 9 CVC 0.020 0.021 0.021 0.029 0.031 9 ∆CVC -0.003 -0.003 -0.003 0.006 0.007 9 %∆CVC -14.22 -12.49 -11.27 23.42 28.25

221
OLDER HEAT Rec 2 Rec 3 Subject Variable BL AVE ISCH 0-20%
127.70 131.50 9 SBP 130.23 130.00 136.80 -4.53 -0.73 9 ∆ SBP 0.00 -0.23 6.57 62.53 65.42 9 DBP 62.52 65.01 69.48
7.15 10.04 9 ∆ DBP 0.00 2.49 6.96 82.00 85.00 9 MAP 84.67 86.00 93.00
1.67 4.67 9 ∆MAP 0.00 1.33 8.33 71.00 73.00 9 HR 69.00 69.00 75.00 -2.67 -0.67 9 ∆ HR 0.00 0.00 6.00 33.72 33.73 9 Skin temp 1 39.70 39.89 40.34
0.44 0.45 9 ∆ Skin temp 1 0.00 0.19 0.64 33.42 33.38 9 Skin temp 2 39.74 39.81 40.12
0.18 0.14 9 ∆ Skin temp 2 0.00 0.07 0.38 31.48 31.44 9 Skin temp 3 31.10 31.09 31.07 -0.15 -0.19 9 ∆ Skin temp 3 0.00 -0.01 -0.03 8.50 10.50 9 bursts 4.50 13.00 7.58
-1.33 0.67 9 ∆bursts 0.00 8.50 3.08 -13.56 6.78 9 %∆bursts 0.00 188.89 68.49 858.10 1364.60 9 total act 476.64 1152.73 800.70
-165.43 341.08 9 ∆total act 0.00 676.10 324.06 -16.16 33.32 9 % ∆ total act 0.00 141.85 67.99
0 0 9 RPE 6 0 13 0 0 9 ∆RPE 0 0 7
36.70 36.75 9 Muscle temp 39.36 39.33 39.53 0.16 0.21 9 ∆Muscle temp 0.00 -0.03 0.17
70.00 58.40 9 RBV 62.04 55.70 51.20 8.08 -3.52 9 ∆RBV 0.00 -6.34 -10.84
13.05 -5.68 9 %∆RBV 0.00 -10.22 -17.47 2.09 1.75 9 RVC 1.56 1.40 1.28 0.23 -0.11 9 ∆RVC 0.00 -0.16 -0.29
12.43 -6.09 9 %∆RVC 0.00 -10.38 -18.26 2.02 1.89 9 CBF 1.76 1.92 1.84 0.10 -0.03 9 ∆CBF 0.00 0.16 0.08 5.40 -1.32 9 %∆CBF 0.00 8.97 4.47
0.025 0.022 9 CVC 0.021 0.022 0.020 0.001 -0.002 9 ∆CVC 0.000 0.002 -0.001
3.26 -6.74 9 %∆CVC 0.00 7.29 -4.89

222
OLDER HEAT Subject Variable 20-40% 40-60% 60-80% 80-100% fatigue
9 SBP 156.60 182.30 190.40 187.00 188.20 9 ∆ SBP 26.37 52.07 60.17 56.77 57.97 9 DBP 73.91 79.57 85.62 87.53 87.77 9 ∆ DBP 11.39 17.05 23.10 25.01 25.25 9 MAP 99.00 112.00 121.00 124.00 125.00 9 ∆MAP 14.33 27.33 36.33 39.33 40.33 9 HR 74.00 76.00 73.00 73.00 72.00 9 ∆ HR 5.00 7.00 4.00 4.00 3.00 9 Skin temp 1 40.55 40.74 40.89 41.03 41.07 9 ∆ Skin temp 1 0.85 1.04 1.19 1.33 1.37 9 Skin temp 2 40.27 40.38 40.49 40.57 40.59 9 ∆ Skin temp 2 0.53 0.64 0.75 0.83 0.85 9 Skin temp 3 31.09 31.08 31.09 31.07 31.05 9 ∆ Skin temp 3 -0.01 -0.02 -0.01 -0.03 -0.05 9 bursts 8.85 13.90 17.69 16.43 17.69 9 ∆bursts 4.35 9.40 13.19 11.93 13.19 9 %∆bursts 96.57 208.90 293.15 265.07 293.15 9 total act 1110.44 1575.51 1911.96 2135.45 1052.75 9 ∆total act 633.81 1098.87 1435.33 1658.81 576.12 9 % ∆ total act 132.98 230.55 301.14 348.03 120.87 9 RPE 13 15 19 20 20 9 ∆RPE 7 9 13 14 14 9 Muscle temp 39.59 39.64 39.77 39.91 39.91 9 ∆Muscle temp 0.23 0.28 0.41 0.55 0.55 9 RBV 53.90 54.80 54.80 50.30 50.30 9 ∆RBV -8.14 -7.24 -7.24 -11.74 -11.74 9 %∆RBV -13.12 -11.67 -11.67 -18.92 -18.92 9 RVC 1.34 1.36 1.35 1.24 1.24 9 ∆RVC -0.22 -0.20 -0.21 -0.32 -0.32 9 %∆RVC -14.27 -13.08 -13.31 -20.59 -20.63 9 CBF 1.97 2.26 2.63 2.44 2.50 9 ∆CBF 0.20 0.50 0.86 0.68 0.73 9 %∆CBF 11.56 28.37 49.04 38.44 41.56 9 CVC 0.020 0.020 0.022 0.020 0.020 9 ∆CVC -0.001 -0.001 0.001 -0.001 -0.001 9 %∆CVC -4.60 -2.96 4.29 -5.48 -4.12

223
OLDER HEAT Subject Variable PEMI Rec 1 Rec 2 Rec 3
9 SBP 159.10 125.30 128.20 130.30 9 ∆ SBP 28.87 -4.93 -2.03 0.07 9 DBP 72.04 62.67 67.34 67.58 9 ∆ DBP 9.52 0.15 4.82 5.06 9 MAP 101.00 82.00 86.00 87.00 9 ∆MAP 16.33 -2.67 1.33 2.33 9 HR 66.00 67.00 72.00 72.00 9 ∆ HR -3.00 -2.00 3.00 3.00 9 Skin temp 1 41.24 40.00 37.79 36.99 9 ∆ Skin temp 1 1.54 0.30 -1.91 -2.71 9 Skin temp 2 40.71 39.69 37.71 36.77 9 ∆ Skin temp 2 0.97 -0.05 -2.03 -2.97 9 Skin temp 3 31.10 31.07 31.07 31.02 9 ∆ Skin temp 3 0.00 -0.03 -0.03 -0.08 9 bursts 15.00 8.50 5.00 5.00 9 ∆bursts 10.50 4.00 0.50 0.50 9 %∆bursts 233.33 88.89 11.11 11.11 9 total act 1552.45 797.42 558.33 611.80 9 ∆total act 1075.82 320.78 81.69 135.16 9 % ∆ total act 225.71 67.30 17.14 28.36 9 RPE 0 0 0 0 9 ∆RPE 0 0 0 0 9 Muscle temp 39.99 39.78 39.63 39.47 9 ∆Muscle temp 0.63 0.42 0.27 0.11 9 RBV 58.10 64.67 62.83 59.30 9 ∆RBV -3.94 2.63 0.79 -2.74 9 %∆RBV -6.35 4.24 1.28 -4.41 9 RVC 1.43 1.63 1.67 1.61 9 ∆RVC -0.13 0.07 0.10 0.05 9 %∆RVC -8.59 4.36 6.72 3.30 9 CBF 2.12 1.71 1.36 1.74 9 ∆CBF 0.35 -0.05 -0.40 -0.02 9 %∆CBF 20.12 -2.81 -22.94 -1.06 9 CVC 0.021 0.021 0.016 0.020 9 ∆CVC 0.000 0.000 -0.005 -0.001 9 %∆CVC 0.69 0.35 -24.14 -3.72

224
Appendix F
Raw Data For ACE-inhibitor Study: Drug and Placebo Trials During
Normothermia and Heating

225
DRUG NORMOTHERMIA Subject condition variable BL AVE EX 20% EX 40% EX 60%
1 drug SBP 102.95 110.70 117.80 126.90 1 drug DBP 67.02 74.27 79.19 85.32 1 drug MAP 81.40 89.00 95.00 102.00 1 drug HR 57.40 75.00 82.00 90.00 1 drug Skin Temp 1 32.80 32.75 32.74 32.73 1 drug Skin Temp 2 31.69 31.76 31.76 31.77 1 drug Skin Temp 3 31.79 31.96 31.98 32.01 1 drug Bursts 0.70 0.00 0.75 0.75 1 drug Total Activity 99.60 0.00 92.34 140.02 1 drug CBF 2.36 2.92 2.78 2.76 1 drug CVC 0.029 0.033 0.029 0.027 1 drug RBV 43.58 38.80 39.93 41.65 1 drug RVC 0.54 0.44 0.42 0.41 1 drug RPE 6 11 13 16 1 drug Muscle temp 33.79 33.50 33.48 33.41 2 drug SBP 134.34 147.70 154.80 160.70 2 drug DBP 60.09 70.04 75.24 80.45 2 drug MAP 79.20 93.00 100.00 105.00 2 drug HR 65.60 98.00 107.00 104.00 2 drug Skin Temp 1 30.54 30.17 29.95 29.80 2 drug Skin Temp 2 27.64 27.57 27.58 27.58 2 drug Skin Temp 3 33.85 33.92 33.94 33.97 2 drug Bursts 1.10 0.00 2.34 4.69 2 drug Total Activity 150.99 0.00 175.86 761.25 2 drug CBF 1.69 1.52 2.43 1.29 2 drug CVC 0.021 0.016 0.024 0.012 2 drug RBV 80.86 80.20 80.20 80.43 2 drug RVC 1.02 0.86 0.80 0.77 2 drug RPE 6 12 15 17 2 drug Muscle temp 36.21 36.10 36.01 35.99 3 drug SBP 113.40 118.90 119.80 125.80 3 drug DBP 49.42 54.28 55.39 58.92 3 drug MAP 67.00 73.00 75.00 80.00 3 drug HR 65.00 73.00 79.00 80.00 3 drug Skin Temp 1 31.66 31.65 31.67 31.69 3 drug Skin Temp 2 31.73 31.52 31.54 31.54 3 drug Skin Temp 3 22.18 22.21 22.21 22.20 3 drug Bursts 9.00 4.82 4.82 14.47 3 drug Total Activity 1133.74 712.83 498.36 1548.34 3 drug CBF 1.73 2.11 1.73 1.29 3 drug CVC 0.026 0.029 0.023 0.016 3 drug RBV 47.20 45.75 46.53 45.70 3 drug RVC 0.70 0.63 0.62 0.57 3 drug RPE 6 10 14 17 3 drug Muscle temp 35.56 35.72 35.72 35.72

226
DRUG NORMOTHERMIA Subject condition variable EX 80% EX 100% PEMI AVE
1 drug SBP 139.40 153.40 154.70 1 drug DBP 90.52 98.30 103.14 1 drug MAP 110.00 120.00 117.00 1 drug HR 95.00 90.00 64.67 1 drug Skin Temp 1 32.72 32.73 32.70 1 drug Skin Temp 2 31.77 31.77 31.82 1 drug Skin Temp 3 32.05 32.10 32.18 1 drug Bursts 1.50 2.25 6.00 1 drug Total Activity 147.28 332.88 1332.68 1 drug CBF 2.21 1.67 2.31 1 drug CVC 0.020 0.014 0.020 1 drug RBV 39.36 38.80 39.90 1 drug RVC 0.36 0.32 0.34 1 drug RPE 18 19 1 drug Muscle temp 33.38 33.37 33.36 2 drug SBP 173.60 193.80 178.20 2 drug DBP 88.35 98.89 91.27 2 drug MAP 115.00 129.00 120.00 2 drug HR 107.00 105.00 71.33 2 drug Skin Temp 1 29.70 29.62 29.65 2 drug Skin Temp 2 27.57 27.56 27.51 2 drug Skin Temp 3 34.00 34.03 34.47 2 drug Bursts 3.52 9.38 6.50 2 drug Total Activity 663.08 1203.77 1547.14 2 drug CBF 0.81 1.06 0.84 2 drug CVC 0.007 0.008 0.007 2 drug RBV 83.25 81.00 87.10 2 drug RVC 0.72 0.63 0.73 2 drug RPE 19 20 2 drug Muscle temp 35.93 35.93 35.90 3 drug SBP 137.30 145.50 135.23 3 drug DBP 66.03 68.96 63.02 3 drug MAP 89.00 94.00 85.33 3 drug HR 81.00 82.00 67.00 3 drug Skin Temp 1 31.70 31.69 31.65 3 drug Skin Temp 2 31.54 31.53 31.48 3 drug Skin Temp 3 22.23 22.27 22.25 3 drug Bursts 16.40 22.19 17.50 3 drug Total Activity 2141.75 3065.75 2747.13 3 drug CBF 1.39 1.71 1.69 3 drug CVC 0.016 0.018 0.020 3 drug RBV 46.50 46.90 52.70 3 drug RVC 0.52 0.50 0.62 3 drug RPE 19 19 3 drug Muscle temp 35.73 35.60 35.58

227
DRUG NORMOTHERMIA Subject condition variable REC 1 Rec 2 Rec 3
1 drug SBP 138.90 124.70 121.70 1 drug DBP 77.83 66.82 66.48 1 drug MAP 96.00 86.00 86.00 1 drug HR 60.00 61.00 56.00 1 drug Skin Temp 1 32.72 32.80 32.92 1 drug Skin Temp 2 31.86 31.98 32.02 1 drug Skin Temp 3 32.20 32.19 32.15 1 drug Bursts 1.50 1.00 0.50 1 drug Total Activity 185.55 102.96 48.45 1 drug CBF 2.85 2.75 2.74 1 drug CVC 0.030 0.032 0.032 1 drug RBV 41.43 42.90 41.47 1 drug RVC 0.43 0.50 0.48 1 drug RPE 1 drug Muscle temp 33.44 33.63 33.67 2 drug SBP 149.90 145.60 142.60 2 drug DBP 68.51 61.89 59.72 2 drug MAP 92.00 84.00 80.00 2 drug HR 62.00 56.00 58.00 2 drug Skin Temp 1 29.81 29.65 29.78 2 drug Skin Temp 2 27.45 27.38 27.51 2 drug Skin Temp 3 34.33 33.96 33.82 2 drug Bursts 1.00 0.50 1.00 2 drug Total Activity 135.54 90.85 175.35 2 drug CBF 1.56 1.15 1.54 2 drug CVC 0.017 0.014 0.019 2 drug RBV 88.60 98.83 95.20 2 drug RVC 0.96 1.18 1.19 2 drug RPE 2 drug Muscle temp 36.00 35.92 35.83 3 drug SBP 122.70 118.80 120.50 3 drug DBP 52.72 50.72 50.83 3 drug MAP 73.00 70.00 70.00 3 drug HR 69.00 66.00 67.00 3 drug Skin Temp 1 31.65 31.89 32.08 3 drug Skin Temp 2 31.51 31.71 31.81 3 drug Skin Temp 3 22.23 22.23 22.21 3 drug Bursts 15.50 9.50 9.00 3 drug Total Activity 1928.79 1414.78 1623.85 3 drug CBF 1.99 1.69 1.86 3 drug CVC 0.027 0.024 0.027 3 drug RBV 55.87 57.03 52.15 3 drug RVC 0.77 0.81 0.75 3 drug RPE 3 drug Muscle temp 35.73 35.81 35.83

228
DRUG HEAT Subject condition variable BL AVE H EX 20%H EX 40%H
1 drug SBP 102.55 94.21 113.30 1 drug DBP 64.02 63.87 76.46 1 drug MAP 77.80 77.00 91.00 1 drug HR 62.60 84.00 89.00 1 drug Skin Temp 1 40.14 39.95 40.03 1 drug Skin Temp 2 40.69 40.39 40.51 1 drug Skin Temp 3 31.83 31.81 31.83 1 drug Bursts 1.00 1.69 0.00 1 drug Total Activity 103.06 172.76 0.00 1 drug CBF 2.48 2.92 3.58 1 drug CVC 0.032 0.038 0.039 1 drug RBV 53.00 50.50 46.23 1 drug RVC 0.68 0.66 0.51 1 drug RPE 6 11 12 1 drug Muscle temp 37.42 37.52 37.56 2 drug SBP 139.10 141.80 160.50 2 drug DBP 58.18 64.16 73.56 2 drug MAP 78.60 88.00 99.00 2 drug HR 62.00 97.00 96.00 2 drug Skin Temp 1 39.66 39.61 39.66 2 drug Skin Temp 2 41.27 40.98 41.05 2 drug Skin Temp 3 32.76 32.75 32.77 2 drug Bursts 0.90 2.30 2.30 2 drug Total Activity 117.60 447.47 198.81 2 drug CBF 1.82 1.95 2.10 2 drug CVC 0.023 0.022 0.021 2 drug RBV 80.03 71.05 79.45 2 drug RVC 1.02 0.81 0.80 2 drug RPE 6 12 16 2 drug Muscle temp 38.12 38.19 38.21 3 drug SBP 98.93 103.00 113.00 3 drug DBP 42.84 45.23 52.58 3 drug MAP 57.60 61.00 71.00 3 drug HR 71.40 90.00 95.00 3 drug Skin Temp 1 39.18 39.47 39.54 3 drug Skin Temp 2 39.56 39.86 39.92 3 drug Skin Temp 3 22.25 22.30 22.27 3 drug Bursts 4.60 3.14 10.47 3 drug Total Activity 467.28 305.28 846.22 3 drug CBF 2.60 4.19 3.31 3 drug CVC 0.045 0.069 0.047 3 drug RBV 35.53 34.95 34.60 3 drug RVC 0.62 0.57 0.49 3 drug RPE 6 10 14 3 drug Muscle temp 39.41 39.55 39.58

229
DRUG HEAT Subject condition variable EX 60%H EX 80%H EX 100%H
1 drug SBP 122.80 136.80 153.50 1 drug DBP 81.09 86.46 96.06 1 drug MAP 97.00 104.00 115.00 1 drug HR 94.00 97.00 100.00 1 drug Skin Temp 1 40.12 40.17 40.27 1 drug Skin Temp 2 40.61 40.69 40.77 1 drug Skin Temp 3 31.87 31.91 31.97 1 drug Bursts 1.69 0.85 5.92 1 drug Total Activity 70.51 136.48 812.82 1 drug CBF 3.16 2.53 1.97 1 drug CVC 0.033 0.024 0.017 1 drug RBV 46.60 46.23 44.95 1 drug RVC 0.48 0.44 0.39 1 drug RPE 14 18 19 1 drug Muscle temp 37.62 37.76 37.88 2 drug SBP 168.40 184.80 198.30 2 drug DBP 79.65 87.91 98.95 2 drug MAP 109.00 117.00 131.00 2 drug HR 100.00 101.00 107.00 2 drug Skin Temp 1 39.73 39.78 39.82 2 drug Skin Temp 2 41.09 41.12 41.16 2 drug Skin Temp 3 32.80 32.83 32.90 2 drug Bursts 2.30 4.61 8.06 2 drug Total Activity 196.37 478.88 855.41 2 drug CBF 1.78 1.17 1.31 2 drug CVC 0.016 0.010 0.010 2 drug RBV 77.10 77.40 63.00 2 drug RVC 0.71 0.66 0.48 2 drug RPE 18 19 20 2 drug Muscle temp 38.26 38.30 38.30 3 drug SBP 121.50 135.00 149.60 3 drug DBP 56.70 62.51 70.00 3 drug MAP 78.00 86.00 97.00 3 drug HR 93.00 92.00 93.00 3 drug Skin Temp 1 39.56 39.57 39.63 3 drug Skin Temp 2 39.85 39.82 39.87 3 drug Skin Temp 3 22.26 22.29 22.31 3 drug Bursts 9.42 21.98 20.94 3 drug Total Activity 930.84 2669.16 4045.70 3 drug CBF 1.92 1.78 2.63 3 drug CVC 0.025 0.021 0.027 3 drug RBV 34.15 34.95 32.20 3 drug RVC 0.44 0.41 0.33 3 drug RPE 16 18 19 3 drug Muscle temp 39.63 39.64 39.77

230
DRUG HEAT Subject condition variable PEMI AVE Rec 1H Rec 2H Rec 3H
1 drug SBP 153.97 138.20 125.30 121.90 1 drug DBP 97.64 78.86 68.93 64.93 1 drug MAP 119.67 99.00 86.00 83.00 1 drug HR 74.67 67.00 59.00 57.00 1 drug Skin Temp 1 40.35 40.29 40.09 39.85 1 drug Skin Temp 2 40.90 40.90 40.58 40.32 1 drug Skin Temp 3 32.04 32.05 32.05 32.04 1 drug Bursts 9.50 3.50 0.50 0.00 1 drug Total Activity 1822.20 640.75 102.85 0.00 1 drug CBF 1.79 2.47 2.70 2.55 1 drug CVC 0.015 0.025 0.031 0.031 1 drug RBV 46.60 48.43 51.80 50.30 1 drug RVC 0.39 0.49 0.60 0.61 1 drug RPE 1 drug Muscle temp 38.05 37.88 37.65 37.53 2 drug SBP 193.77 152.20 149.30 142.00 2 drug DBP 96.97 70.76 65.20 61.95 2 drug MAP 127.00 95.00 88.00 83.00 2 drug HR 73.33 64.00 58.00 55.00 2 drug Skin Temp 1 39.82 39.69 38.91 38.09 2 drug Skin Temp 2 41.24 41.23 40.68 40.10 2 drug Skin Temp 3 33.21 33.08 32.71 32.53 2 drug Bursts 8.50 0.00 1.00 0.50 2 drug Total Activity 1440.85 0.00 105.47 14.00 2 drug CBF 1.29 2.09 1.14 1.45 2 drug CVC 0.010 0.022 0.013 0.017 2 drug RBV 89.35 87.57 92.20 86.00 2 drug RVC 0.70 0.92 1.05 1.04 2 drug RPE 2 drug Muscle temp 38.37 38.39 38.40 38.29 3 drug SBP 144.20 124.40 108.40 110.00 3 drug DBP 67.15 51.85 44.17 44.69 3 drug MAP 91.67 74.00 62.00 63.00 3 drug HR 73.67 74.00 71.00 72.00 3 drug Skin Temp 1 39.78 39.98 39.69 39.49 3 drug Skin Temp 2 40.16 40.42 40.24 40.06 3 drug Skin Temp 3 22.29 22.30 22.27 22.21 3 drug Bursts 17.50 8.50 7.50 8.00 3 drug Total Activity 3380.22 1448.37 872.82 1120.56 3 drug CBF 2.74 2.75 2.04 2.44 3 drug CVC 0.030 0.037 0.033 0.039 3 drug RBV 41.65 44.23 43.77 39.05 3 drug RVC 0.45 0.60 0.71 0.62 3 drug RPE 3 drug Muscle temp 39.83 39.78 39.72 39.67

231
PLACEBO NORMOTHERMIA Subject condition variable BL AVE EX 20% EX 40%
1 placebo SBP 114.90 117.80 123.60 1 placebo DBP 55.76 59.20 65.86 1 placebo MAP 73.40 79.00 85.00 1 placebo HR 63.00 88.00 95.00 1 placebo Skin Temp 1 33.71 33.53 33.48 1 placebo Skin Temp 2 33.07 32.86 32.72 1 placebo Skin Temp 3 32.20 32.17 32.16 1 placebo Bursts 1.50 0.00 4.55 1 placebo Total Activity 125.92 0.00 306.42 1 placebo CBF 3.26 3.80 4.68 1 placebo CVC 0.044 0.048 0.055 1 placebo RBV 43.99 42.55 44.03 1 placebo RVC 0.60 0.54 0.52 1 placebo RPE 6 11 12 1 placebo Muscle temp 34.22 33.96 33.93 2 placebo SBP 158.14 165.00 175.70 2 placebo DBP 81.51 87.75 92.45 2 placebo MAP 100.60 110.00 117.00 2 placebo HR 77.40 110.00 109.00 2 placebo Skin Temp 1 33.56 33.66 33.69 2 placebo Skin Temp 2 33.55 33.51 33.50 2 placebo Skin Temp 3 31.98 32.03 32.00 2 placebo Bursts 1.70 2.25 4.49 2 placebo Total Activity 178.25 242.92 735.00 2 placebo CBF 2.81 5.48 4.70 2 placebo CVC 0.028 0.050 0.040 2 placebo RBV 59.42 58.70 58.65 2 placebo RVC 0.59 0.53 0.50 2 placebo RPE 6 13 15 2 placebo Muscle temp 34.34 34.29 34.26 3 placebo SBP 121.62 131.80 136.90 3 placebo DBP 59.96 64.29 69.15 3 placebo MAP 76.80 83.00 90.00 3 placebo HR 70.20 79.00 80.00 3 placebo Skin Temp 1 30.64 30.28 30.22 3 placebo Skin Temp 2 30.99 30.47 30.39 3 placebo Skin Temp 3 30.28 30.16 30.18 3 placebo Bursts 8.20 7.58 14.07 3 placebo Total Activity 723.96 547.82 1190.56 3 placebo CBF 2.37 2.82 2.22 3 placebo CVC 0.031 0.034 0.025 3 placebo RBV 42.75 42.20 42.20 3 placebo RVC 0.56 0.51 0.47 3 placebo RPE 6 10 14 3 placebo Muscle temp 34.99 34.90 34.88

232
PLACEBO NORMOTHERMIA Subject condition variable EX 60% EX 80 % EX 100 %
1 placebo SBP 131.40 149.20 163.90 1 placebo DBP 70.43 81.12 89.32 1 placebo MAP 91.00 107.00 119.00 1 placebo HR 104.00 106.00 102.00 1 placebo Skin Temp 1 33.41 33.39 33.40 1 placebo Skin Temp 2 32.60 32.59 32.65 1 placebo Skin Temp 3 32.16 32.18 32.21 1 placebo Bursts 10.01 17.29 14.56 1 placebo Total Activity 921.47 1325.71 1392.30 1 placebo CBF 4.09 3.08 4.03 1 placebo CVC 0.045 0.029 0.034 1 placebo RBV 44.40 41.63 36.05 1 placebo RVC 0.49 0.39 0.30 1 placebo RPE 15 17 19 1 placebo Muscle temp 33.87 33.79 33.56 2 placebo SBP 190.20 202.50 214.10 2 placebo DBP 98.52 107.40 117.70 2 placebo MAP 125.00 136.00 149.00 2 placebo HR 99.00 98.00 98.00 2 placebo Skin Temp 1 33.70 33.69 33.69 2 placebo Skin Temp 2 33.49 33.48 33.47 2 placebo Skin Temp 3 31.98 31.97 31.93 2 placebo Bursts 2.25 8.99 7.87 2 placebo Total Activity 145.57 872.26 1306.18 2 placebo CBF 2.83 2.31 2.65 2 placebo CVC 0.023 0.017 0.018 2 placebo RBV 66.30 67.00 70.30 2 placebo RVC 0.53 0.49 0.47 2 placebo RPE 18 19 20 2 placebo Muscle temp 34.24 34.21 34.20 3 placebo SBP 145.10 159.00 169.10 3 placebo DBP 76.84 86.56 88.87 3 placebo MAP 98.00 110.00 114.00 3 placebo HR 81.00 81.00 85.00 3 placebo Skin Temp 1 30.18 30.13 30.09 3 placebo Skin Temp 2 30.42 30.45 30.44 3 placebo Skin Temp 3 30.21 30.22 30.26 3 placebo Bursts 10.82 18.40 28.14 3 placebo Total Activity 1174.11 2214.61 3653.06 3 placebo CBF 2.03 2.45 1.84 3 placebo CVC 0.021 0.022 0.016 3 placebo RBV 42.20 42.20 42.20 3 placebo RVC 0.43 0.38 0.37 3 placebo RPE 16 18 19 3 placebo Muscle temp 34.88 34.88 34.87

233
PLACEBO NORMOTHERMIA
Subject condition variable PEMI AVE REC 1 Rec 2 Rec 3
1 placebo SBP 164.70 138.30 124.60 123.40 1 placebo DBP 86.88 63.38 51.27 49.60 1 placebo MAP 116.33 88.00 73.00 72.00 1 placebo HR 78.33 68.00 70.00 61.00 1 placebo Skin Temp 1 33.49 33.63 33.73 33.80 1 placebo Skin Temp 2 32.89 33.18 33.43 33.52 1 placebo Skin Temp 3 32.25 32.23 32.25 32.25 1 placebo Bursts 16.00 4.00 2.50 3.00 1 placebo Total Activity 2257.49 531.35 190.96 205.87 1 placebo CBF 2.69 3.61 3.82 3.96 1 placebo CVC 0.023 0.041 0.052 0.055 1 placebo RBV 39.35 38.80 41.43 40.67 1 placebo RVC 0.34 0.44 0.57 0.56 1 placebo RPE 1 placebo Muscle temp 34.56 33.52 34.08 34.11 2 placebo SBP 205.83 178.40 165.30 150.20 2 placebo DBP 111.83 93.50 86.89 81.32 2 placebo MAP 141.00 116.00 108.00 101.00 2 placebo HR 79.67 69.00 65.00 71.00 2 placebo Skin Temp 1 33.55 33.32 33.38 33.68 2 placebo Skin Temp 2 33.41 33.30 33.16 33.32 2 placebo Skin Temp 3 30.84 30.28 29.56 29.72 2 placebo Bursts 5.50 1.00 1.00 1.50 2 placebo Total Activity 566.40 113.28 53.09 148.67 2 placebo CBF 2.96 3.38 2.33 3.12 2 placebo CVC 0.021 0.029 0.022 0.031 2 placebo RBV 70.50 51.70 48.90 48.43 2 placebo RVC 0.50 0.45 0.45 0.48 2 placebo RPE 2 placebo Muscle temp 34.16 34.15 34.17 34.17 3 placebo SBP 161.80 139.70 126.80 128.50 3 placebo DBP 84.58 68.95 65.30 65.20 3 placebo MAP 108.33 89.00 83.00 83.00 3 placebo HR 67.33 71.00 68.00 68.00 3 placebo Skin Temp 1 30.10 30.12 30.23 30.37 3 placebo Skin Temp 2 30.44 30.53 31.05 31.53 3 placebo Skin Temp 3 30.24 30.18 30.16 30.08 3 placebo Bursts 19.50 13.00 9.50 6.50 3 placebo Total Activity 2350.80 1115.10 890.33 585.48 3 placebo CBF 2.49 2.63 2.07 2.31 3 placebo CVC 0.023 0.030 0.025 0.028 3 placebo RBV 45.70 43.95 46.30 43.40 3 placebo RVC 0.42 0.49 0.56 0.52 3 placebo RPE 3 placebo Muscle temp 34.72 35.02 35.02 35.19

234
PLACEBO HEAT Subject condition variable BL AVE H EX 20%H EX 40%H
1 placebo SBP 116.83 129.20 132.80 1 placebo DBP 59.73 68.21 70.85 1 placebo MAP 76.33 87.00 90.00 1 placebo HR 65.67 80.00 91.00 1 placebo Skin Temp 1 39.92 40.01 40.07 1 placebo Skin Temp 2 40.25 40.42 40.56 1 placebo Skin Temp 3 32.66 32.62 32.61 1 placebo Bursts 2.50 3.26 0.00 1 placebo Total Activity 217.47 340.00 0.00 1 placebo CBF 3.82 3.00 3.87 1 placebo CVC 0.05 0.035 0.043 1 placebo RBV 47.22 47.33 42.53 1 placebo RVC 0.62 0.54 0.47 1 placebo RPE 6 10 12 1 placebo Muscle temp 36.29 36.55 36.67 2 placebo SBP 184.32 199.70 214.50 2 placebo DBP 100.53 118.80 130.00 2 placebo MAP 123.40 142.00 154.00 2 placebo HR 79.60 111.00 115.00 2 placebo Skin Temp 1 40.77 40.54 40.54 2 placebo Skin Temp 2 40.62 40.41 40.43 2 placebo Skin Temp 3 31.98 31.84 31.83 2 placebo Bursts 1.60 2.86 1.43 2 placebo Total Activity 184.30 378.07 103.26 2 placebo CBF 3.27 4.55 5.18 2 placebo CVC 0.03 0.032 0.034 2 placebo RBV 72.13 72.60 72.60 2 placebo RVC 0.58 0.51 0.47 2 placebo RPE 6 14 15 2 placebo Muscle temp 37.24 37.34 37.38 3 placebo SBP 119.42 121.60 135.70 3 placebo DBP 55.83 57.48 66.66 3 placebo MAP 73.00 75.00 87.00 3 placebo HR 72.40 101.00 102.00 3 placebo Skin Temp 1 39.31 39.41 39.52 3 placebo Skin Temp 2 39.14 39.56 39.74 3 placebo Skin Temp 3 31.16 31.12 31.11 3 placebo Bursts 9.65 14.07 11.90 3 placebo Total Activity 1032.61 1393.66 1114.45 3 placebo CBF 3.94 8.66 7.57 3 placebo CVC 0.05 0.115 0.087 3 placebo RBV 46.76 39.25 36.13 3 placebo RVC 0.64 0.52 0.42 3 placebo RPE 6 11 14 3 placebo Muscle temp 39.28 39.35 39.42

235
PLACEBO HEAT Subject condition variable EX 60%H EX80%H EX 100%H
1 placebo SBP 138.80 152.70 164.70 1 placebo DBP 77.97 86.14 93.54 1 placebo MAP 98.00 109.00 118.00 1 placebo HR 100.00 102.00 102.00 1 placebo Skin Temp 1 40.08 40.03 39.99 1 placebo Skin Temp 2 40.46 40.25 40.15 1 placebo Skin Temp 3 32.61 32.62 32.65 1 placebo Bursts 3.26 9.79 22.84 1 placebo Total Activity 234.92 732.49 1827.97 1 placebo CBF 2.84 3.13 3.79 1 placebo CVC 0.029 0.029 0.032 1 placebo RBV 42.17 44.40 41.90 1 placebo RVC 0.43 0.41 0.36 1 placebo RPE 14 17 19 1 placebo Muscle temp 36.81 36.93 37.15 2 placebo SBP 227.10 234.90 254.70 2 placebo DBP 137.50 139.60 154.20 2 placebo MAP 164.00 168.00 187.00 2 placebo HR 117.00 110.00 105.00 2 placebo Skin Temp 1 40.55 40.55 40.55 2 placebo Skin Temp 2 40.46 40.46 40.47 2 placebo Skin Temp 3 31.90 32.01 32.06 2 placebo Bursts 5.72 8.58 5.72 2 placebo Total Activity 386.02 1148.78 517.98 2 placebo CBF 5.08 4.13 3.03 2 placebo CVC 0.031 0.025 0.016 2 placebo RBV 71.90 71.20 71.20 2 placebo RVC 0.44 0.42 0.38 2 placebo RPE 16 18 19 2 placebo Muscle temp 37.40 37.44 37.47 3 placebo SBP 146.40 162.50 179.50 3 placebo DBP 75.92 83.72 89.31 3 placebo MAP 98.00 110.00 118.00 3 placebo HR 101.00 95.00 99.00 3 placebo Skin Temp 1 39.63 39.73 39.83 3 placebo Skin Temp 2 39.89 40.01 40.13 3 placebo Skin Temp 3 31.10 31.14 31.22 3 placebo Bursts 24.89 31.39 32.47 3 placebo Total Activity 2920.98 5408.60 5897.68 3 placebo CBF 4.15 2.87 3.15 3 placebo CVC 0.042 0.026 0.027 3 placebo RBV 37.30 41.37 41.63 3 placebo RVC 0.38 0.38 0.35 3 placebo RPE 16 18 19 3 placebo Muscle temp 39.49 39.58 39.73

236
PLACEBO HEAT Subject condition variable PEMI H Rec 1H Rec 2H Rec 3H
1 placebo SBP 168.83 139.00 122.20 116.50 1 placebo DBP 96.37 73.91 61.81 57.88 1 placebo MAP 123.33 95.00 81.00 76.00 1 placebo HR 75.00 69.00 64.00 63.00 1 placebo Skin Temp 1 39.97 39.99 39.88 39.75 1 placebo Skin Temp 2 40.23 40.34 40.04 39.80 1 placebo Skin Temp 3 32.73 32.74 32.67 32.65 1 placebo Bursts 14.50 5.50 5.00 2.50 1 placebo Total Activity 1821.27 619.66 324.37 332.54 1 placebo CBF 2.64 3.23 3.13 3.89 1 placebo CVC 0.021 0.034 0.039 0.051 1 placebo RBV 47.70 44.10 44.95 45.87 1 placebo RVC 0.39 0.46 0.55 0.60 1 placebo RPE 1 placebo Muscle temp 37.29 37.28 36.63 36.46 2 placebo SBP 245.27 211.30 195.50 190.20 2 placebo DBP 143.17 113.80 101.40 97.68 2 placebo MAP 174.67 140.00 126.00 121.00 2 placebo HR 82.67 68.00 71.00 74.00 2 placebo Skin Temp 1 40.63 40.14 39.23 39.13 2 placebo Skin Temp 2 40.56 40.18 39.73 39.65 2 placebo Skin Temp 3 32.04 32.01 32.06 32.06 2 placebo Bursts 7.00 1.50 1.50 0.50 2 placebo Total Activity 675.63 107.85 153.82 36.49 2 placebo CBF 3.23 4.37 3.88 3.94 2 placebo CVC 0.018 0.031 0.031 0.033 2 placebo RBV 55.90 53.10 63.33 65.60 2 placebo RVC 0.32 0.38 0.50 0.54 2 placebo RPE 2 placebo Muscle temp 37.56 37.71 37.62 37.45 3 placebo SBP 170.77 138.20 125.80 125.90 3 placebo DBP 82.00 62.57 60.41 58.29 3 placebo MAP 107.67 83.00 79.00 77.00 3 placebo HR 72.33 76.00 72.00 72.00 3 placebo Skin Temp 1 39.96 39.99 39.67 39.21 3 placebo Skin Temp 2 40.32 39.89 39.03 38.53 3 placebo Skin Temp 3 31.20 31.12 31.12 31.13 3 placebo Bursts 26.00 18.50 20.50 15.00 3 placebo Total Activity 4438.22 2404.44 2407.68 1752.62 3 placebo CBF 3.92 3.02 3.03 3.38 3 placebo CVC 0.036 0.036 0.038 0.044 3 placebo RBV 50.80 52.75 54.50 52.75 3 placebo RVC 0.47 0.64 0.69 0.69 3 placebo RPE 3 placebo Muscle temp 39.89 39.65 39.57 39.57

237
DRUG NORMOTHERMIA Subject condition variable BL AVE EX 20% EX 40% EX 60%
4 drug SBP 102.61 109.20 118.30 137.40 4 drug DBP 43.18 46.66 53.52 64.34 4 drug MAP 57.80 62.00 70.00 83.00 4 drug HR 75.20 89.00 86.00 85.00 4 drug Skin Temp 1 32.42 32.53 32.60 32.58 4 drug Skin Temp 2 32.42 32.53 32.60 32.58 4 drug Skin Temp 3 32.42 32.53 32.60 32.58 4 drug Bursts 2.40 1.08 5.41 6.49 4 drug Total Activity 285.51 60.48 511.83 794.93 4 drug CBF 3.01 3.14 2.16 2.61 4 drug CVC 0.052 0.051 0.031 0.031 4 drug RBV 40.21 44.10 35.55 36.33 4 drug RVC 0.70 0.71 0.51 0.44 4 drug RPE 6 12 15 17 4 drug Muscle temp 33.83 33.89 33.86 33.83 5 drug SBP 137.70 139.50 142.50 145.70 5 drug DBP 54.73 59.50 61.50 63.36 5 drug MAP 76.60 81.00 84.00 87.00 5 drug HR 81.20 106.00 107.00 112.00 5 drug Skin Temp 1 33.88 33.91 33.89 33.87 5 drug Skin Temp 2 33.39 33.40 33.37 33.35 5 drug Skin Temp 3 32.52 32.50 32.50 32.50 5 drug Bursts 4.70 8.20 7.17 8.20 5 drug Total Activity 419.39 411.17 293.72 449.06 5 drug CBF 4.64 7.66 8.85 7.21 5 drug CVC 0.061 0.095 0.105 0.083 5 drug RBV 39.83 39.13 40.95 40.70 5 drug RVC 0.52 0.48 0.49 0.47 5 drug RPE 6 12 15 15 5 drug Muscle temp 34.47 34.45 34.42 34.40 6 drug SBP 110.32 131.50 123.40 141.00 6 drug DBP 54.39 64.72 61.40 69.79 6 drug MAP 70.40 86.00 80.00 92.00 6 drug HR 54.40 60.00 75.00 69.00 6 drug Skin Temp 1 31.77 32.08 32.10 32.12 6 drug Skin Temp 2 31.66 31.81 31.81 31.81 6 drug Skin Temp 3 29.53 27.83 27.77 27.73 6 drug Bursts 1.00 1.16 2.32 1.16 6 drug Total Activity 172.76 405.37 422.18 124.38 6 drug CBF 3.41 3.21 2.99 5.81 6 drug CVC 0.048 0.037 0.037 0.063 6 drug RPE 6 12 15 17 6 drug Muscle temp 34.27 34.43 34.46 34.48

238
DRUG NORMOTHERMIA Subject condition variable EX 80% EX 100% PEMI AVE
4 drug SBP 144.40 160.60 149.17 4 drug DBP 71.89 82.26 52.89 4 drug MAP 91.00 63.00 83.00 4 drug HR 88.00 58.00 58.00 4 drug Skin Temp 1 32.60 32.61 32.67 4 drug Skin Temp 2 32.60 32.61 32.67 4 drug Skin Temp 3 32.60 32.61 32.67 4 drug Bursts 8.65 9.19 10.00 4 drug Total Activity 961.28 1197.49 1768.09 4 drug CBF 2.74 3.17 1.83 4 drug CVC 0.030 0.050 0.022 4 drug RBV 31.30 31.11 34.20 4 drug RVC 0.34 0.49 0.41 4 drug RPE 18 19 4 drug Muscle temp 33.80 33.85 33.93 5 drug SBP 155.60 156.20 170.23 5 drug DBP 68.08 70.25 74.29 5 drug MAP 93.00 97.00 102.67 5 drug HR 117.00 120.00 102.67 5 drug Skin Temp 1 33.83 33.81 33.73 5 drug Skin Temp 2 33.35 33.34 33.37 5 drug Skin Temp 3 32.50 32.51 32.60 5 drug Bursts 7.17 12.30 30.50 5 drug Total Activity 384.86 797.73 2177.54 5 drug CBF 8.48 9.06 6.95 5 drug CVC 0.091 0.093 0.068 5 drug RBV 37.75 37.20 38.17 5 drug RVC 0.41 0.38 0.37 5 drug RPE 18 18 5 drug Muscle temp 34.39 34.39 34.41 6 drug SBP 170.30 170.30 143.80 6 drug DBP 49.31 49.31 67.87 6 drug MAP 6.00 6.00 92.00 6 drug HR 9.00 9.00 54.67 6 drug Skin Temp 1 32.13 32.13 32.16 6 drug Skin Temp 2 31.80 31.80 31.79 6 drug Skin Temp 3 27.76 27.76 27.64 6 drug Bursts 11.59 15.07 8.00 6 drug Total Activity 2578.25 2453.91 1987.42 6 drug CBF 3.10 2.69 2.96 6 drug CVC 0.517 0.449 0.032 6 drug RPE 18 20 6 drug Muscle temp 34.50 34.51 34.54

239
DRUG NORMOTHERMIA Subject condition variable REC 1 Rec 2 Rec 3
4 drug SBP 126.90 119.90 115.80 4 drug DBP 53.19 50.60 48.77 4 drug MAP 71.00 67.00 65.00 4 drug HR 70.00 69.00 71.00 4 drug Skin Temp 1 32.73 32.74 32.79 4 drug Skin Temp 2 32.73 32.74 32.79 4 drug Skin Temp 3 32.73 32.74 32.79 4 drug Bursts 6.00 4.00 3.50 4 drug Total Activity 851.45 618.82 499.31 4 drug CBF 3.37 1.69 2.69 4 drug CVC 0.047 0.025 0.041 4 drug RBV 41.18 45.40 45.40 4 drug RVC 0.58 0.68 0.70 4 drug RPE 4 drug Muscle temp 34.01 34.14 34.29 5 drug SBP 139.70 141.90 141.90 5 drug DBP 57.15 55.46 55.46 5 drug MAP 78.00 76.00 76.00 5 drug HR 76.00 70.00 70.00 5 drug Skin Temp 1 34.04 34.01 34.01 5 drug Skin Temp 2 33.87 33.86 33.86 5 drug Skin Temp 3 32.84 32.80 32.80 5 drug Bursts 6.50 6.00 6.00 5 drug Total Activity 887.60 536.62 536.62 5 drug CBF 4.39 3.77 3.77 5 drug CVC 0.056 0.050 0.050 5 drug RBV 40.55 41.40 41.40 5 drug RVC 0.52 0.54 0.54 5 drug RPE 5 drug Muscle temp 34.43 34.43 34.58 6 drug SBP 124.80 110.20 110.00 6 drug DBP 60.40 54.97 54.15 6 drug MAP 80.00 72.00 71.00 6 drug HR 58.00 58.00 57.00 6 drug Skin Temp 1 32.21 32.22 32.20 6 drug Skin Temp 2 31.87 32.01 32.03 6 drug Skin Temp 3 27.66 27.66 27.71 6 drug Bursts 1.50 1.00 1.00 6 drug Total Activity 268.17 138.56 158.03 6 drug CBF 2.93 3.14 3.31 6 drug CVC 0.037 0.044 0.047 6 drug RPE 6 drug Muscle temp 33.98 33.99 33.99

240
DRUG HEAT Subject condition variable BL AVE H EX 20%H EX 40%H
4 drug SBP 111.86 118.70 122.30 4 drug DBP 52.57 61.55 63.85 4 drug MAP 68.00 78.00 80.00 4 drug HR 75.60 92.00 93.00 4 drug Skin Temp 1 39.92 39.78 39.91 4 drug Skin Temp 2 40.06 39.80 39.77 4 drug Skin Temp 3 32.42 32.53 32.60 4 drug Bursts 1.80 4.68 5.62 4 drug Total Activity 186.99 547.28 417.23 4 drug CBF 2.15 2.32 1.90 4 drug CVC 0.032 0.030 0.024 4 drug RBV 44.50 33.30 35.10 4 drug RVC 0.65 0.43 0.44 4 drug RPE 6 12 15 4 drug Muscle temp 37.67 37.51 37.52 5 drug SBP 131.08 147.10 140.90 5 drug DBP 53.26 57.87 54.21 5 drug MAP 72.00 80.00 78.00 5 drug HR 84.80 104.00 116.00 5 drug Skin Temp 1 41.11 41.23 41.38 5 drug Skin Temp 2 41.00 41.35 41.55 5 drug Skin Temp 3 32.82 32.86 32.86 5 drug Bursts 4.00 9.72 9.72 5 drug Total Activity 295.30 380.31 698.24 5 drug CBF 4.72 7.17 8.18 5 drug CVC 0.066 0.090 0.105 5 drug RBV 38.50 31.60 34.90 5 drug RVC 0.53 0.40 0.45 5 drug RPE 6 12 12 5 drug Muscle temp 37.41 37.49 37.49 6 drug SBP 109.04 127.90 123.90 6 drug DBP 56.81 67.40 65.69 6 drug MAP 72.40 86.00 84.00 6 drug HR 49.80 59.00 67.00 6 drug Skin Temp 1 39.68 39.35 39.41 6 drug Skin Temp 2 40.24 39.98 40.05 6 drug Skin Temp 3 31.23 31.14 31.12 6 drug Bursts 1.10 4.08 9.51 6 drug Total Activity 233.24 483.33 1870.30 6 drug CBF 2.61 2.38 1.99 6 drug CVC 0.036 0.028 0.024 6 drug RPE 6 13 15 6 drug Muscle temp 37.44 37.31 37.31

241
DRUG HEAT Subject condition variable EX 60%H EX 80%H EX 100%H
4 drug SBP 136.80 146.30 160.60 4 drug DBP 73.30 57.17 82.26 4 drug MAP 92.00 26.00 63.00 4 drug HR 97.00 37.00 58.00 4 drug Skin Temp 1 40.03 40.08 40.11 4 drug Skin Temp 2 39.76 39.80 39.88 4 drug Skin Temp 3 32.58 32.60 32.61 4 drug Bursts 13.12 13.12 18.74 4 drug Total Activity 1804.30 2115.76 3275.01 4 drug CBF 1.68 2.16 2.15 4 drug CVC 0.018 0.083 0.034 4 drug RBV 33.30 32.40 36.30 4 drug RVC 0.36 1.25 0.58 4 drug RPE 17 19 19 4 drug Muscle temp 37.56 37.60 37.63 5 drug SBP 151.30 168.10 175.10 5 drug DBP 59.36 69.79 78.06 5 drug MAP 84.00 98.00 106.00 5 drug HR 120.00 121.00 119.00 5 drug Skin Temp 1 41.51 41.58 41.63 5 drug Skin Temp 2 41.71 41.81 41.87 5 drug Skin Temp 3 32.85 32.85 32.88 5 drug Bursts 8.33 20.83 25.00 5 drug Total Activity 866.07 1555.07 1786.51 5 drug CBF 9.91 9.36 9.28 5 drug CVC 0.118 0.096 0.088 5 drug RBV 33.30 35.50 38.25 5 drug RVC 0.40 0.36 0.36 5 drug RPE 16 16 20 5 drug Muscle temp 37.61 37.61 37.71 6 drug SBP 135.80 152.60 167.90 6 drug DBP 75.06 85.32 94.83 6 drug MAP 96.00 108.00 120.00 6 drug HR 70.00 69.00 68.00 6 drug Skin Temp 1 39.46 39.51 39.55 6 drug Skin Temp 2 40.10 40.13 40.17 6 drug Skin Temp 3 30.99 31.00 31.10 6 drug Bursts 4.08 13.59 20.38 6 drug Total Activity 773.94 3949.46 5134.62 6 drug CBF 2.92 3.03 2.61 6 drug CVC 0.030 0.028 0.022 6 drug RPE 17 18 19 6 drug Muscle temp 37.32 37.32 37.34

242
DRUG HEAT
Subject condition variable PEMI AVE H Rec 1H Rec 2H Rec 3H
4 drug SBP 149.17 126.90 119.90 115.80 4 drug DBP 52.89 53.19 50.60 48.77 4 drug MAP 83.00 71.00 67.00 65.00 4 drug HR 58.00 70.00 69.00 71.00 4 drug Skin Temp 1 40.28 40.48 40.34 40.08 4 drug Skin Temp 2 40.05 40.21 39.91 39.65 4 drug Skin Temp 3 32.67 32.73 32.74 32.79 4 drug Bursts 14.00 4.00 2.50 1.00 4 drug Total Activity 2145.13 576.03 419.15 184.70 4 drug CBF 1.75 2.44 1.89 2.36 4 drug CVC 0.021 0.034 0.028 0.036 4 drug RBV 40.05 44.07 49.10 44.70 4 drug RVC 0.48 0.62 0.73 0.69 4 drug RPE 4 drug Muscle temp 37.67 37.81 37.74 37.65 5 drug SBP 166.63 147.70 137.20 143.10 5 drug DBP 73.64 63.09 62.54 62.20 5 drug MAP 100.67 86.00 83.00 83.00 5 drug HR 102.00 88.00 84.00 76.00 5 drug Skin Temp 1 41.83 41.59 40.56 40.10 5 drug Skin Temp 2 42.06 41.56 40.23 39.78 5 drug Skin Temp 3 33.25 33.61 33.59 33.54 5 drug Bursts 28.50 16.00 15.50 13.00 5 drug Total Activity 2034.36 957.40 573.37 571.27 5 drug CBF 7.47 5.47 5.34 4.57 5 drug CVC 0.074 0.064 0.064 0.055 5 drug RBV 42.20 33.85 34.47 33.97 5 drug RVC 0.42 0.39 0.42 0.41 5 drug RPE 5 drug Muscle temp 37.83 37.92 37.66 37.48 6 drug SBP 159.23 132.40 124.00 124.90 6 drug DBP 87.50 71.86 69.90 68.25 6 drug MAP 113.67 92.00 89.00 88.00 6 drug HR 62.00 61.00 59.00 57.00 6 drug Skin Temp 1 39.64 39.31 38.78 38.64 6 drug Skin Temp 2 40.30 39.49 38.83 38.91 6 drug Skin Temp 3 31.10 31.13 31.14 31.17 6 drug Bursts 12.00 6.00 2.50 2.50 6 drug Total Activity 2708.14 932.55 494.15 447.45 6 drug CBF 2.42 3.03 2.54 2.92 6 drug CVC 0.021 0.033 0.029 0.033 6 drug RPE 6 drug Muscle temp 37.38 37.39 37.35 37.32

243
PLACEBO NORMOTHERMIA
Subject condition variable BL AVE EX 20% EX 40%
4 placebo SBP 122.60 122.60 121.60 4 placebo DBP 63.84 67.52 71.67 4 placebo MAP 77.20 81.00 85.00 4 placebo HR 75.80 88.00 94.00 4 placebo Skin Temp 1 31.92 31.87 31.87 4 placebo Skin Temp 2 33.00 32.94 32.91 4 placebo Skin Temp 3 33.50 33.57 33.58 4 placebo Bursts 2.20 5.90 3.37 4 placebo Total Activity 128.19 287.95 282.93 4 placebo CBF 2.68 3.09 2.65 4 placebo CVC 0.035 0.038 0.031 4 placebo RBV 45.00 40.70 41.60 4 placebo RVC 0.58 0.50 0.49 4 placebo RPE 6 13 15 4 placebo Muscle temp 33.98 33.98 33.95 5 placebo SBP 146.10 162.60 161.60 5 placebo DBP 66.02 74.31 74.94 5 placebo MAP 86.40 97.00 98.00 5 placebo HR 68.20 84.00 86.00 5 placebo Skin Temp 1 34.45 34.59 34.60 5 placebo Skin Temp 2 33.94 34.09 34.09 5 placebo Skin Temp 3 31.70 31.58 31.54 5 placebo Bursts 0.50 0.00 0.82 5 placebo Total Activity 68.03 0.00 58.77 5 placebo CBF 3.42 4.28 4.10 5 placebo CVC 0.040 0.044 0.042 5 placebo RBV 55.94 52.70 52.33 5 placebo RVC 0.65 0.54 0.53 5 placebo RPE 6 13 14 5 placebo Muscle temp 35.03 35.10 35.09 6 placebo SBP 109.42 131.20 130.10 6 placebo DBP 57.51 69.60 70.07 6 placebo MAP 74.60 91.00 90.00 6 placebo HR 54.40 65.00 76.00 6 placebo Skin Temp 1 34.49 34.42 34.39 6 placebo Skin Temp 2 34.72 34.71 34.70 6 placebo Skin Temp 3 34.28 34.20 34.19 6 placebo Bursts 0.60 2.13 6.39 6 placebo Total Activity 106.89 157.06 1024.51 6 placebo CBF 2.55 3.32 3.24 6 placebo CVC 0.034 0.037 0.036 6 placebo RPE 6 13 16 6 placebo Muscle temp 36.56 36.60 36.60

244
PLACEBO NORMOTHERMIA Subject condition variable EX 60% EX 80 % EX 100 %
4 placebo SBP 135.60 151.30 156.70 4 placebo DBP 77.66 86.15 100.10 4 placebo MAP 93.00 103.00 115.00 4 placebo HR 92.00 95.00 99.00 4 placebo Skin Temp 1 31.85 31.78 31.72 4 placebo Skin Temp 2 32.91 32.90 32.89 4 placebo Skin Temp 3 33.55 33.57 33.64 4 placebo Bursts 9.27 14.33 10.96 4 placebo Total Activity 623.84 964.73 1142.17 4 placebo CBF 2.87 2.73 5.80 4 placebo CVC 0.031 0.027 0.050 4 placebo RBV 47.63 39.83 38.55 4 placebo RVC 0.51 0.39 0.34 4 placebo RPE 17 18 19 4 placebo Muscle temp 33.93 33.90 33.91 5 placebo SBP 169.70 176.80 189.60 5 placebo DBP 78.09 82.86 88.69 5 placebo MAP 101.00 108.00 116.00 5 placebo HR 86.00 93.00 98.00 5 placebo Skin Temp 1 34.59 34.59 34.59 5 placebo Skin Temp 2 34.10 34.10 34.10 5 placebo Skin Temp 3 31.52 31.53 31.53 5 placebo Bursts 0.82 0.00 3.26 5 placebo Total Activity 43.04 0.00 263.18 5 placebo CBF 3.88 4.32 4.58 5 placebo CVC 0.038 0.040 0.039 5 placebo RBV 53.45 50.40 47.30 5 placebo RVC 0.53 0.47 0.41 5 placebo RPE 15 17 19 5 placebo Muscle temp 35.07 35.06 35.04 6 placebo SBP 145.20 161.50 177.90 6 placebo DBP 78.71 89.21 102.00 6 placebo MAP 103.00 113.00 128.00 6 placebo HR 72.00 69.00 69.00 6 placebo Skin Temp 1 34.36 34.35 34.35 6 placebo Skin Temp 2 34.69 34.68 34.68 6 placebo Skin Temp 3 34.17 34.20 34.23 6 placebo Bursts 9.58 9.58 12.78 6 placebo Total Activity 1268.52 2056.67 3005.26 6 placebo CBF 2.62 2.08 2.09 6 placebo CVC 0.025 0.018 0.016 6 placebo RPE 18 19 20 6 placebo Muscle temp 36.59 36.57 36.57

245
PLACEBO NORMOTHERMIA
Subject condition variable PEMI AVE REC 1 Rec 2 Rec 3
4 placebo SBP 155.27 142.80 138.30 129.00 4 placebo DBP 84.59 71.70 68.34 65.60 4 placebo MAP 102.00 87.00 83.00 80.00 4 placebo HR 73.67 73.00 70.00 76.00 4 placebo Skin Temp 1 31.71 31.82 31.89 31.99 4 placebo Skin Temp 2 32.93 32.97 33.01 33.09 4 placebo Skin Temp 3 33.63 33.46 33.40 33.39 4 placebo Bursts 11.00 7.50 4.00 3.50 4 placebo Total Activity 1109.68 453.91 341.58 404.47 4 placebo CBF 4.47 2.95 2.52 2.75 4 placebo CVC 0.044 0.034 0.030 0.034 4 placebo RBV 42.85 41.57 43.30 40.70 4 placebo RVC 0.42 0.48 0.52 0.51 4 placebo RPE 4 placebo Muscle temp 34.03 34.04 34.24 34.40 5 placebo SBP 183.30 155.70 149.40 151.50 5 placebo DBP 87.71 71.55 68.83 68.85 5 placebo MAP 112.33 93.00 89.00 89.00 5 placebo HR 72.00 64.00 66.00 66.00 5 placebo Skin Temp 1 34.57 34.59 34.71 34.76 5 placebo Skin Temp 2 34.09 34.11 34.23 34.33 5 placebo Skin Temp 3 31.53 31.54 31.55 31.53 5 placebo Bursts 7.00 1.00 1.00 0.00 5 placebo Total Activity 866.77 152.70 111.60 0.00 5 placebo CBF 5.17 3.79 3.37 3.50 5 placebo CVC 0.046 0.041 0.038 0.039 5 placebo RBV 48.10 43.80 55.00 51.93 5 placebo RVC 0.43 0.47 0.62 0.58 5 placebo RPE 5 placebo Muscle temp 34.95 34.93 34.97 35.06 6 placebo SBP 150.60 127.40 120.70 113.90 6 placebo DBP 84.89 68.36 64.89 61.51 6 placebo MAP 109.67 89.00 84.00 81.00 6 placebo HR 53.33 56.00 57.00 54.00 6 placebo Skin Temp 1 34.33 34.32 34.39 34.44 6 placebo Skin Temp 2 34.64 34.70 34.84 34.84 6 placebo Skin Temp 3 34.27 34.24 34.22 34.19 6 placebo Bursts 11.00 2.00 4.00 2.00 6 placebo Total Activity 2511.81 245.62 382.30 186.87 6 placebo CBF 2.88 2.96 2.74 2.66 6 placebo CVC 0.026 0.033 0.033 0.033 6 placebo RPE 6 placebo Muscle temp 36.56 36.56 36.58 36.57

246
PLACEBO HEAT Subject condition variable BL AVE H EX 20%H EX 40%H
4 placebo SBP 130.82 133.20 137.40 4 placebo DBP 65.30 66.21 73.84 4 placebo MAP 81.40 84.00 92.00 4 placebo HR 78.20 95.00 101.00 4 placebo Skin Temp 1 43.80 43.93 44.07 4 placebo Skin Temp 2 39.88 39.93 40.00 4 placebo Skin Temp 3 33.77 33.82 33.82 4 placebo Bursts 1.25 1.34 2.67 4 placebo Total Activity 148.28 294.65 838.37 4 placebo CBF 3.20 2.88 6.04 4 placebo CVC 0.04 0.034 0.066 4 placebo RBV 47.25 39.40 36.40 4 placebo RVC 0.58 0.47 0.40 4 placebo RPE 6 13 14 4 placebo Muscle temp 37.00 36.83 36.89 5 placebo SBP 142.55 157.90 150.90 5 placebo DBP 68.85 75.48 74.23 5 placebo MAP 87.00 95.00 96.00 5 placebo HR 68.75 81.00 87.00 5 placebo Skin Temp 1 40.87 41.23 41.41 5 placebo Skin Temp 2 39.95 40.28 40.45 5 placebo Skin Temp 3 26.92 27.09 27.14 5 placebo Bursts 0.60 1.08 3.25 5 placebo Total Activity 67.39 46.27 343.41 5 placebo CBF 3.49 3.81 3.90 5 placebo CVC 0.04 0.040 0.041 5 placebo RBV 50.29 48.90 46.83 5 placebo RVC 0.58 0.51 0.49 5 placebo RPE 6 13 13 5 placebo Muscle temp 37.28 37.35 37.47 6 placebo SBP 116.57 129.50 135.30 6 placebo DBP 60.67 66.52 72.65 6 placebo MAP 79.33 88.00 94.00 6 placebo HR 56.67 66.00 78.00 6 placebo Skin Temp 1 38.14 37.96 37.99 6 placebo Skin Temp 2 38.54 38.33 38.38 6 placebo Skin Temp 3 33.27 33.18 33.18 6 placebo Bursts 2.00 6.38 8.94 6 placebo Total Activity 312.07 858.23 1272.57 6 placebo CBF 2.58 2.69 2.27 6 placebo CVC 0.03 0.031 0.024 6 placebo RPE 6 15 16 6 placebo Muscle temp 38.01 37.98 38.00

247
PLACEBO HEAT Subject condition variable EX 60%H EX80%H EX 100%H
4 placebo SBP 144.30 174.70 176.20 4 placebo DBP 79.83 81.84 93.79 4 placebo MAP 97.00 113.00 118.00 4 placebo HR 98.00 97.00 89.00 4 placebo Skin Temp 1 44.23 44.39 44.58 4 placebo Skin Temp 2 40.06 40.12 40.16 4 placebo Skin Temp 3 33.82 33.89 34.09 4 placebo Bursts 10.70 14.71 21.39 4 placebo Total Activity 2016.54 2768.24 4919.83 4 placebo CBF 6.95 4.27 3.47 4 placebo CVC 0.072 0.038 0.029 4 placebo RBV 35.05 34.60 35.70 4 placebo RVC 0.36 0.31 0.30 4 placebo RPE 15 17 19 4 placebo Muscle temp 36.94 36.94 39.94 5 placebo SBP 154.00 162.90 168.10 5 placebo DBP 76.74 82.35 79.73 5 placebo MAP 99.00 106.00 109.00 5 placebo HR 93.00 95.00 88.00 5 placebo Skin Temp 1 41.56 41.69 41.81 5 placebo Skin Temp 2 40.60 40.72 40.82 5 placebo Skin Temp 3 27.13 27.04 26.96 5 placebo Bursts 1.08 5.42 10.83 5 placebo Total Activity 161.05 570.80 1462.79 5 placebo CBF 4.21 4.21 5.30 5 placebo CVC 0.043 0.040 0.049 5 placebo RBV 48.37 44.77 40.30 5 placebo RVC 0.49 0.42 0.37 5 placebo RPE 15 17 20 5 placebo Muscle temp 37.57 37.70 37.70 6 placebo SBP 143.60 165.70 185.40 6 placebo DBP 78.40 92.88 104.70 6 placebo MAP 101.00 118.00 133.00 6 placebo HR 81.00 77.00 73.00 6 placebo Skin Temp 1 38.02 38.04 38.04 6 placebo Skin Temp 2 38.42 38.44 38.43 6 placebo Skin Temp 3 33.20 33.23 33.26 6 placebo Bursts 10.21 19.15 21.70 6 placebo Total Activity 1309.76 3671.62 6323.43 6 placebo CBF 3.34 2.12 1.62 6 placebo CVC 0.033 0.018 0.012 6 placebo RPE 17 18 20 6 placebo Muscle temp 38.00 38.02 38.06

248
PLACEBO HEAT
Subject condition variable PEMI H Rec 1H Rec 2H Rec 3H
4 placebo SBP 162.47 141.40 293.90 146.80 4 placebo DBP 80.80 61.37 24.50 37.80 4 placebo MAP 102.00 83.00 80.00 39.00 4 placebo HR 86.67 78.00 62.00 13.00 4 placebo Skin Temp 1 44.68 43.84 43.02 42.48 4 placebo Skin Temp 2 40.28 40.18 39.74 39.45 4 placebo Skin Temp 3 34.24 34.02 33.91 33.90 4 placebo Bursts 14.50 3.50 2.50 2.50 4 placebo Total Activity 2767.44 694.32 594.85 485.65 4 placebo CBF 4.64 3.90 2.76 2.33 4 placebo CVC 0.046 0.047 0.035 0.060 4 placebo RBV 43.30 43.30 45.60 46.45 4 placebo RVC 0.42 0.52 0.57 1.19 4 placebo RPE 4 placebo Muscle temp 37.08 37.18 37.03 36.91 5 placebo SBP 178.97 156.50 148.40 141.50 5 placebo DBP 90.84 74.62 70.33 67.25 5 placebo MAP 117.33 96.00 91.00 87.00 5 placebo HR 72.33 69.00 64.00 62.00 5 placebo Skin Temp 1 41.86 40.49 39.61 39.48 5 placebo Skin Temp 2 40.88 39.41 38.36 38.32 5 placebo Skin Temp 3 27.10 27.36 27.24 27.12 5 placebo Bursts 12.00 1.00 0.50 1.50 5 placebo Total Activity 1695.43 125.42 76.85 182.02 5 placebo CBF 5.06 3.27 3.08 3.14 5 placebo CVC 0.043 0.034 0.034 0.036 5 placebo RBV 41.20 50.40 50.40 51.95 5 placebo RVC 0.35 0.53 0.55 0.60 5 placebo RPE 5 placebo Muscle temp 37.95 38.18 37.75 37.45 6 placebo SBP 159.17 135.20 130.60 131.00 6 placebo DBP 87.08 73.61 71.08 69.66 6 placebo MAP 112.00 96.00 92.00 91.00 6 placebo HR 59.33 58.00 54.00 53.00 6 placebo Skin Temp 1 38.06 38.09 37.92 37.78 6 placebo Skin Temp 2 38.49 38.45 37.93 37.77 6 placebo Skin Temp 3 33.34 33.29 33.28 33.27 6 placebo Bursts 11.00 3.00 4.00 3.50 6 placebo Total Activity 2920.67 555.32 707.23 565.31 6 placebo CBF 2.58 2.75 2.31 2.40 6 placebo CVC 0.023 0.029 0.025 0.026 6 placebo RPE 6 placebo Muscle temp 38.09 38.12 38.12 38.12

249
DRUG NORMOTHERMIA Subject condition variable BL AVE EX 20% EX 40% EX 60%
7 drug SBP 116.12 134.00 134.10 141.00 7 drug DBP 59.43 69.19 72.76 79.29 7 drug MAP 76.40 90.00 93.00 102.00 7 drug HR 65.00 77.00 80.00 90.00 7 drug Skin Temp 1 33.74 33.76 33.75 33.74 7 drug Skin Temp 2 30.96 30.96 30.97 30.98 7 drug Skin Temp 3 31.17 31.13 31.16 31.16 7 drug Bursts 0.80 3.25 0.00 2.17 7 drug Total Activity 53.54 293.99 0.00 126.88 7 drug CBF 1.39 1.64 3.07 1.54 7 drug CVC 0.018 0.018 0.033 0.015 7 drug RBV 58.13 54.40 55.97 53.40 7 drug RVC 0.76 0.60 0.60 0.52 7 drug RPE 6 11 15 15 7 drug Muscle temp 35.39 35.27 35.30 35.31 8 drug SBP 122.76 130.10 138.70 146.70 8 drug DBP 60.67 67.62 72.40 77.11 8 drug MAP 78.80 88.00 94.00 100.00 8 drug HR 66.80 79.00 78.00 82.00 8 drug Skin Temp 1 33.89 34.07 33.95 33.84 8 drug Skin Temp 2 32.47 32.83 32.78 32.75 8 drug Skin Temp 3 33.22 33.12 33.14 33.15 8 drug Bursts 5.30 6.15 12.30 12.30 8 drug Total Activity 605.54 726.44 1322.72 2059.23 8 drug CBF 2.43 2.09 2.46 2.78 8 drug CVC 0.031 0.024 0.026 0.028 8 drug RBV 102.72 91.40 532.45 90.20 8 drug RVC 1.30 1.04 5.66 0.90 8 drug RPE 6 12 13 15 8 drug Muscle temp 33.87 33.94 33.92 33.90

250
DRUG NORMOTHERMIA Subject condition variable EX 80% EX 100% PEMI AVE
7 drug SBP 148.10 159.70 146.20 7 drug DBP 83.15 92.07 80.01 7 drug MAP 106.00 117.00 104.00 7 drug HR 84.00 84.00 71.33 7 drug Skin Temp 1 33.74 33.75 33.79 7 drug Skin Temp 2 30.99 31.02 31.09 7 drug Skin Temp 3 31.17 31.19 31.19 7 drug Bursts 15.18 13.01 8.50 7 drug Total Activity 2526.36 2266.26 1378.16 7 drug CBF 1.67 1.09 1.83 7 drug CVC 0.016 0.009 0.018 7 drug RBV 51.80 48.20 61.45 7 drug RVC 0.49 0.41 0.59 7 drug RPE 18 20 7 drug Muscle temp 35.33 35.31 35.31 8 drug SBP 161.90 167.80 148.83 8 drug DBP 86.98 90.89 77.60 8 drug MAP 113.00 119.00 103.33 8 drug HR 86.00 85.00 76.00 8 drug Skin Temp 1 33.74 33.68 33.58 8 drug Skin Temp 2 32.71 32.68 32.63 8 drug Skin Temp 3 33.16 33.17 33.08 8 drug Bursts 15.38 19.22 17.50 8 drug Total Activity 2136.58 2679.89 2661.38 8 drug CBF 5.08 2.74 3.32 8 drug CVC 0.045 0.023 0.032 8 drug RBV 83.00 84.20 111.00 8 drug RVC 0.73 0.71 1.07 8 drug RPE 18 20 8 drug Muscle temp 33.87 33.84 33.65

251
DRUG NORMOTHERMIA Subject condition variable REC 1 Rec 2 Rec 3
7 drug SBP 125.40 109.50 105.90 7 drug DBP 64.83 56.55 53.96 7 drug MAP 84.00 74.00 70.00 7 drug HR 76.00 67.00 62.00 7 drug Skin Temp 1 33.83 33.88 33.90 7 drug Skin Temp 2 31.17 31.20 31.12 7 drug Skin Temp 3 31.12 31.11 31.09 7 drug Bursts 3.50 2.50 4.50 7 drug Total Activity 541.42 426.35 492.87 7 drug CBF 1.56 0.86 0.95 7 drug CVC 0.019 0.012 0.014 7 drug RBV 55.17 60.77 56.87 7 drug RVC 0.66 0.82 0.81 7 drug RPE 7 drug Muscle temp 35.31 35.38 35.40 8 drug SBP 136.30 127.80 126.80 8 drug DBP 65.99 61.81 60.90 8 drug MAP 89.00 83.00 82.00 8 drug HR 71.00 71.00 67.00 8 drug Skin Temp 1 33.57 33.69 33.81 8 drug Skin Temp 2 32.68 32.83 32.87 8 drug Skin Temp 3 32.97 32.93 32.90 8 drug Bursts 11.50 13.50 9.50 8 drug Total Activity 1401.77 1369.00 1245.69 8 drug CBF 3.60 2.69 3.04 8 drug CVC 0.040 0.032 0.037 8 drug RBV 91.40 102.25 95.43 8 drug RVC 1.03 1.23 1.16 8 drug RPE 8 drug Muscle temp 33.78 33.81 33.83

252
DRUG HEAT Subject condition variable BL AVE H EX 20%H EX 40%H
7 drug SBP 105.32 141.20 131.10 7 drug DBP 54.20 74.54 67.99 7 drug MAP 70.80 96.00 89.00 7 drug HR 65.40 76.00 95.00 7 drug Skin Temp 1 40.61 40.83 40.95 7 drug Skin Temp 2 39.41 39.21 39.26 7 drug Skin Temp 3 31.25 31.30 31.29 7 drug Bursts 1.20 0.00 1.58 7 drug Total Activity 158.47 0.00 83.02 7 drug CBF 0.85 1.13 0.75 7 drug CVC 0.012 0.012 0.008 7 drug RBV 60.65 51.65 52.50 7 drug RVC 0.86 0.54 0.59 7 drug RPE 6 12 15 7 drug Muscle temp 38.88 38.65 38.71 8 drug SBP 125.36 140.30 138.50 8 drug DBP 57.27 68.52 68.82 8 drug MAP 77.00 91.00 92.00 8 drug HR 75.20 82.00 83.00 8 drug Skin Temp 1 39.23 39.47 39.62 8 drug Skin Temp 2 41.07 41.53 41.76 8 drug Skin Temp 3 33.39 33.34 33.35 8 drug Bursts 6.00 8.38 10.06 8 drug Total Activity 735.87 1209.23 1109.36 8 drug CBF 3.01 2.97 2.82 8 drug CVC 0.039 0.033 0.031 8 drug RBV 73.48 62.23 60.15 8 drug RVC 0.95 0.68 0.65 8 drug RPE 6 12 15 8 drug Muscle temp 36.73 36.96 37.01

253
DRUG HEAT Subject condition variable EX 60%H EX 80%H EX 100%H
7 drug SBP 142.70 158.00 166.60 7 drug DBP 77.07 86.02 92.15 7 drug MAP 100.00 111.00 122.00 7 drug HR 99.00 97.00 97.00 7 drug Skin Temp 1 41.05 41.13 41.19 7 drug Skin Temp 2 39.29 39.33 39.37 7 drug Skin Temp 3 31.28 31.26 31.26 7 drug Bursts 6.32 15.79 23.68 7 drug Total Activity 781.88 2043.30 4500.73 7 drug CBF 3.55 3.65 1.48 7 drug CVC 0.036 0.033 0.012 7 drug RBV 53.70 55.00 50.53 7 drug RVC 0.54 0.50 0.41 7 drug RPE 17 17 20 7 drug Muscle temp 38.71 38.76 38.81 8 drug SBP 150.70 163.60 170.60 8 drug DBP 75.59 83.23 85.61 8 drug MAP 101.00 111.00 115.00 8 drug HR 87.00 93.00 94.00 8 drug Skin Temp 1 39.74 39.81 39.86 8 drug Skin Temp 2 41.96 42.14 42.30 8 drug Skin Temp 3 33.37 33.41 33.45 8 drug Bursts 12.58 14.25 20.96 8 drug Total Activity 1489.12 2017.42 3043.77 8 drug CBF 3.51 2.59 2.20 8 drug CVC 0.035 0.023 0.019 8 drug RBV 59.27 56.27 52.10 8 drug RVC 0.59 0.51 0.45 8 drug RPE 17 19 20 8 drug Muscle temp 37.11 37.22 37.33

254
DRUG HEAT
Subject condition variable PEMI AVE H Rec 1H Rec 2H Rec 3H
7 drug SBP 162.07 117.40 103.40 105.30 7 drug DBP 86.20 62.58 55.30 55.40 7 drug MAP 113.67 81.00 72.00 73.00 7 drug HR 76.00 67.00 65.00 61.00 7 drug Skin Temp 1 41.31 41.01 40.03 39.50 7 drug Skin Temp 2 39.51 39.31 38.26 37.30 7 drug Skin Temp 3 31.26 31.25 31.27 31.23 7 drug Bursts 18.50 8.00 9.00 6.50 7 drug Total Activity 4755.17 2074.38 1695.21 1395.05 7 drug CBF 0.91 1.35 0.97 1.02 7 drug CVC 0.008 0.017 0.013 0.014 7 drug RBV 69.10 63.07 63.50 62.83 7 drug RVC 0.61 0.78 0.88 0.86 7 drug RPE 7 drug Muscle temp 38.89 38.90 38.75 38.61 8 drug SBP 164.33 140.00 135.10 126.10 8 drug DBP 80.04 62.27 61.42 57.59 8 drug MAP 108.67 85.00 83.00 78.00 8 drug HR 80.67 78.00 79.00 75.00 8 drug Skin Temp 1 40.04 39.45 38.70 38.39 8 drug Skin Temp 2 42.44 40.56 39.36 39.25 8 drug Skin Temp 3 33.44 33.27 33.18 33.10 8 drug Bursts 22.00 8.50 8.00 10.50 8 drug Total Activity 3284.06 1036.62 1147.07 1525.27 8 drug CBF 3.36 3.35 2.12 2.30 8 drug CVC 0.031 0.039 0.026 0.029 8 drug RBV 77.20 73.60 74.80 74.80 8 drug RVC 0.71 0.87 0.90 0.96 8 drug RPE 8 drug Muscle temp 37.60 37.61 37.33 37.16

255
PLACEBO NORMOTHERMIA
Subject condition variable BL AVE EX 20% EX 40%
7 placebo SBP 108.23 129.70 126.80 7 placebo DBP 61.44 72.80 72.46 7 placebo MAP 76.40 89.00 89.00 7 placebo HR 55.00 78.00 86.00 7 placebo Skin Temp 1 33.36 33.30 33.32 7 placebo Skin Temp 2 32.22 32.18 32.17 7 placebo Skin Temp 3 32.47 32.63 32.60 7 placebo Bursts 1.80 0.00 1.27 7 placebo Total Activity 187.89 0.00 10.72 7 placebo CBF 1.27 1.34 2.04 7 placebo CVC 0.017 0.015 0.023 7 placebo RBV 48.92 47.75 49.60 7 placebo RVC 0.64 0.54 0.56 7 placebo RPE 6 16 16 7 placebo Muscle temp 36.52 36.46 36.46 8 placebo SBP 117.30 133.30 134.20 8 placebo DBP 50.83 60.26 62.68 8 placebo MAP 69.80 82.00 86.00 8 placebo HR 59.60 67.00 69.00 8 placebo Skin Temp 1 30.97 30.64 30.63 8 placebo Skin Temp 2 28.85 28.75 28.84 8 placebo Skin Temp 3 33.22 33.12 33.14 8 placebo Bursts 0.70 1.95 0.98 8 placebo Total Activity 109.17 675.59 183.79 8 placebo CBF 1.26 1.09 1.08 8 placebo CVC 0.018 0.013 0.013 8 placebo RBV 101.20 99.85 93.83 8 placebo RVC 1.45 1.22 1.09 8 placebo RPE 6 15 17 8 placebo Muscle temp 35.32 35.11 35.07

256
PLACEBO NORMOTHERMIA Subject condition variable EX 60% EX 80 % EX 100 %
7 placebo SBP 150.60 159.20 159.20 7 placebo DBP 86.37 108.20 108.20 7 placebo MAP 105.00 116.00 116.00 7 placebo HR 79.00 83.00 83.00 7 placebo Skin Temp 1 33.32 33.32 33.32 7 placebo Skin Temp 2 32.18 32.18 32.18 7 placebo Skin Temp 3 32.58 32.57 32.57 7 placebo Bursts 1.27 1.27 10.18 7 placebo Total Activity 88.52 45.71 981.87 7 placebo CBF 3.69 2.59 1.27 7 placebo CVC 0.035 0.022 0.011 7 placebo RBV 49.60 42.10 34.90 7 placebo RVC 0.47 0.36 0.30 7 placebo RPE 18 19 20 7 placebo Muscle temp 36.44 36.44 36.42 8 placebo SBP 138.70 143.60 152.40 8 placebo DBP 65.94 67.80 77.82 8 placebo MAP 90.00 93.00 105.00 8 placebo HR 72.00 73.00 76.00 8 placebo Skin Temp 1 30.59 30.56 30.53 8 placebo Skin Temp 2 28.81 28.76 28.69 8 placebo Skin Temp 3 33.15 33.16 33.17 8 placebo Bursts 5.86 4.88 7.81 8 placebo Total Activity 572.63 777.15 1367.22 8 placebo CBF 1.15 0.93 1.27 8 placebo CVC 0.013 0.010 0.012 8 placebo RBV 90.80 91.40 91.40 8 placebo RVC 1.01 0.98 0.87 8 placebo RPE 19 20 20 8 placebo Muscle temp 34.98 34.97 34.95

257
PLACEBO NORMOTHERMIA
Subject condition variable PEMI AVE REC 1 Rec 2 Rec 3
7 placebo SBP 143.73 116.20 101.70 128.60 7 placebo DBP 90.42 65.79 55.48 67.80 7 placebo MAP 99.67 80.00 71.00 85.00 7 placebo HR 52.33 55.00 55.00 59.00 7 placebo Skin Temp 1 33.31 33.35 33.42 33.49 7 placebo Skin Temp 2 32.17 32.28 32.32 32.38 7 placebo Skin Temp 3 32.59 32.59 32.56 32.54 7 placebo Bursts 10.50 3.50 3.50 0.50 7 placebo Total Activity 2270.07 387.41 362.10 34.22 7 placebo CBF 0.37 1.98 1.37 1.63 7 placebo CVC 0.004 0.025 0.019 0.019 7 placebo RBV 61.75 59.03 59.50 55.00 7 placebo RVC 0.62 0.74 0.84 0.65 7 placebo RPE 7 placebo Muscle temp 36.43 36.44 36.51 36.70 8 placebo SBP 141.40 131.10 131.70 129.60 8 placebo DBP 66.23 54.44 56.69 56.36 8 placebo MAP 91.33 76.00 78.00 78.00 8 placebo HR 63.67 66.00 63.00 63.00 8 placebo Skin Temp 1 30.44 30.42 30.55 30.82 8 placebo Skin Temp 2 28.60 28.83 29.61 29.97 8 placebo Skin Temp 3 33.08 32.97 32.93 32.90 8 placebo Bursts 3.00 3.00 0.50 0.50 8 placebo Total Activity 544.14 708.55 21.70 232.80 8 placebo CBF 1.49 1.36 1.37 1.46 8 placebo CVC 0.016 0.018 0.018 0.019 8 placebo RBV 101.35 92.20 97.03 98.03 8 placebo RVC 1.11 1.21 1.24 1.26 8 placebo RPE 8 placebo Muscle temp 34.82 34.81 34.81 34.84

258
PLACEBO HEAT Subject condition variable BL AVE H EX 20%H EX 40%H
7 placebo SBP 111.12 133.50 136.10 7 placebo DBP 62.23 70.98 73.94 7 placebo MAP 78.20 93.00 96.00 7 placebo HR 57.60 65.00 66.00 7 placebo Skin Temp 1 39.71 39.77 39.81 7 placebo Skin Temp 2 39.72 39.77 39.81 7 placebo Skin Temp 3 33.57 33.65 33.65 7 placebo Bursts 3.30 0.00 0.00 7 placebo Total Activity 432.38 0.00 0.00 7 placebo CBF 1.12 1.05 1.50 7 placebo CVC 0.01 0.011 0.016 7 placebo RBV 53.20 51.00 51.00 7 placebo RVC 0.68 0.55 0.53 7 placebo RPE 6 17 17 7 placebo Muscle temp 39.00 39.13 39.13 8 placebo SBP 104.30 130.20 129.30 8 placebo DBP 46.14 60.14 61.95 8 placebo MAP 64.00 82.00 85.00 8 placebo HR 62.67 68.00 74.00 8 placebo Skin Temp 1 39.73 39.83 39.95 8 placebo Skin Temp 2 40.29 40.20 40.33 8 placebo Skin Temp 3 33.41 33.34 33.35 8 placebo Bursts 1.30 0.95 3.79 8 placebo Total Activity 224.33 37.68 1063.55 8 placebo CBF 1.54 1.31 1.12 8 placebo CVC 0.02 0.016 0.013 8 placebo RBV 85.13 74.50 55.70 8 placebo RVC 1.33 0.91 0.66 8 placebo RPE 6 14 18 8 placebo Muscle temp 39.41 38.72 38.42

259
PLACEBO HEAT Subject condition variable EX 60%H EX80%H EX 100%H
7 placebo SBP 135.00 143.10 151.50 7 placebo DBP 77.27 81.73 89.71 7 placebo MAP 98.00 106.00 113.00 7 placebo HR 87.00 99.00 104.00 7 placebo Skin Temp 1 39.84 39.87 39.90 7 placebo Skin Temp 2 39.85 39.89 39.93 7 placebo Skin Temp 3 33.63 33.61 33.59 7 placebo Bursts 4.50 4.50 27.03 7 placebo Total Activity 596.04 382.77 3050.95 7 placebo CBF 1.13 0.92 0.92 7 placebo CVC 0.012 0.009 0.008 7 placebo RBV 50.03 49.60 37.60 7 placebo RVC 0.51 0.47 0.33 7 placebo RPE 19 19 20 7 placebo Muscle temp 39.18 39.22 39.22 8 placebo SBP 134.00 140.90 148.50 8 placebo DBP 64.23 71.46 74.10 8 placebo MAP 87.00 96.00 100.00 8 placebo HR 76.00 77.00 77.00 8 placebo Skin Temp 1 40.03 40.03 40.01 8 placebo Skin Temp 2 40.32 40.16 40.06 8 placebo Skin Temp 3 33.37 33.41 33.45 8 placebo Bursts 9.48 4.74 7.58 8 placebo Total Activity 1925.37 1479.44 2360.49 8 placebo CBF 1.33 1.35 0.76 8 placebo CVC 0.015 0.014 0.008 8 placebo RBV 56.60 61.67 63.75 8 placebo RVC 0.65 0.64 0.64 8 placebo RPE 18 20 20 8 placebo Muscle temp 38.65 38.57 38.31

260
PLACEBO HEAT Subject condition variable PEMI H Rec 1H Rec 2H Rec 3H
7 placebo SBP 159.43 131.40 136.00 135.90 7 placebo DBP 98.51 65.74 -0.41 -0.31 7 placebo MAP 116.33 55.00 67.00 67.00 7 placebo HR 64.67 32.00 67.00 67.00 7 placebo Skin Temp 1 39.88 39.32 38.60 38.41 7 placebo Skin Temp 2 39.89 39.29 38.49 38.33 7 placebo Skin Temp 3 33.55 33.55 33.59 33.64 7 placebo Bursts 11.00 10.50 10.00 8.00 7 placebo Total Activity 4411.13 1962.78 1299.26 1082.05 7 placebo CBF 0.40 0.95 0.91 1.28 7 placebo CVC 0.003 0.017 0.014 0.019 7 placebo RBV 61.05 55.47 55.03 56.37 7 placebo RVC 0.52 1.01 0.82 0.84 7 placebo RPE 7 placebo Muscle temp 39.30 39.46 38.97 38.86 8 placebo SBP 133.17 112.80 107.20 103.80 8 placebo DBP 64.19 52.20 50.32 49.16 8 placebo MAP 88.00 72.00 69.00 67.00 8 placebo HR 69.67 70.00 67.00 65.00 8 placebo Skin Temp 1 40.19 40.34 40.10 39.91 8 placebo Skin Temp 2 40.42 40.38 39.48 39.24 8 placebo Skin Temp 3 33.44 33.27 33.18 33.10 8 placebo Bursts 4.50 5.00 5.50 1.00 8 placebo Total Activity 906.34 1357.61 1238.56 387.55 8 placebo CBF 1.29 1.59 1.50 1.96 8 placebo CVC 0.015 0.022 0.022 0.029 8 placebo RBV 87.80 83.40 82.60 78.20 8 placebo RVC 1.00 1.16 1.20 1.17 8 placebo RPE 8 placebo Muscle temp 38.75 39.43 39.52 39.50

261
Appendix G
Blood Data For ACE-inhibitor Study: Drug and Placebo Trials During
Normothermia and Heating

262
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
1 drug AnGap 7.5 5.2 8.9 1.7 1 drug BE(B) -0.9 -2.2 -0.5 -6.9 1 drug BE(ecf) -0.6 -2 -0.3 -7.9 1 drug BO2 9.2 8.4 9.5 7.1 1 drug CA++ 1.16 1.08 1.2 0.86 1 drug CA++(7.4) 1.14 1.06 1.19 0.85 1 drug Cl- 111 114 109 124 1 drug ctCO2 26.4 25.1 26.2 18.4 1 drug ctHb 15 13.7 15.5 11.7 1 drug ctO2 6.3 5.3 8.7 5.3 1 drug FCOHb 0.7 0.7 0.3 0.7 1 drug FHHb 31.4 37.2 9 25.5 1 drug FMetHb 0.8 0.5 0.6 0.9 1 drug FO@Hb 67.1 61.6 90.1 72.9 1 drug Glucose 4.3 4.2 4.7 3.5 1 drug HCO3 std 23 21.9 23.9 18.4 1 drug HCO3-act 25 23.7 24.9 17.4 1 drug Hct 44 40 46 34 1 drug K+ 3.6 3.37 3.85 2.29 1 drug Lactate 1.2 1.07 0.95 0.73 1 drug Na++ 139.9 139 138.9 140.8 1 drug P02 36.9 32.9 59.7 38.4 1 drug p50 26.3 25.6 26.3 25.4 1 drug pCO2 46 44.6 43.3 31.1 1 drug pH 7.353 7.343 7.377 7.366 1 drug pO2 36.9 32.9 59.7 38.4 1 drug sO2 68.1 62.3 90.9 74.1

263
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
1 placebo AnGap 8.3 8.1 8.5 8.8 1 placebo BE(B) 1.1 -0.1 1.1 0.7 1 placebo BE(ecf) 1.8 0.4 1.7 1.3 1 placebo BO2 9 8.7 9.2 8.2 1 placebo CA++ 1.22 1.19 1.22 1.19 1 placebo CA++(7.4) 1.2 1.17 1.21 1.17 1 placebo Cl- 106 108 106 107 1 placebo ctCO2 28.6 27.3 28.3 28.1 1 placebo ctHb 14.6 14.2 15 13.4 1 placebo ctO2 7.5 6.6 8.3 6.3 1 placebo FCOHb 0.3 0.3 0.2 0.1 1 placebo FHHb 17.1 24.9 10.2 24.3 1 placebo FMetHb 0.7 0.7 0.9 0.8 1 placebo FO@Hb 81.9 74.1 88.7 74.8 1 placebo Glucose 4.6 4.5 4.9 4.7 1 placebo HCO3 std 25.1 23.9 25.3 24.6 1 placebo HCO3-act 27.2 25.9 26.8 26.6 1 placebo Hct 43 42 44 39 1 placebo K+ 3.89 3.77 3.9 3.8 1 placebo Lactate 1.07 0.98 0.51 0.58 1 placebo Na++ 137.9 138.2 137.4 138.6 1 placebo P02 44.6 38.7 53.2 39.5 1 placebo p50 24.1 24.5 24 24.9 1 placebo pCO2 48.4 47.2 46.4 47.7 1 placebo pH 7.367 3.357 7.38 7.365 1 placebo pO2 44.6 38.7 53.2 39.5 1 placebo sO2 82.7 74.8 89.7 75.5

264
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
2 drug AnGap 6 9.1 1.9 7.6 2 drug BE(B) -0.7 -1.2 1.6 0.2 2 drug BE(ecf) -0.9 -1.4 2.1 0.3 2 drug BO2 8.5 8.2 8.9 6.5 2 drug CA++ 1.09 1.11 1.19 1.12 2 drug CA++(7.4) 1.09 1.11 1.19 1.11 2 drug Cl- 109 109 105 108 2 drug ctCO2 25.1 24.8 28.3 26.7 2 drug ctHb 13.9 13.4 14.4 10.6 2 drug ctO2 7.9 5.7 7.7 4.6 2 drug FCOHb 0.7 0.8 0.2 0.2 2 drug FHHb 8.6 30.9 14.1 31 2 drug FMetHb 0.5 0.4 0.7 0.3 2 drug FO@Hb 90.2 67.9 85 68.5 2 drug Glucose 4.6 4.3 5 4.7 2 drug HCO3 std 23.8 22.8 25.6 24.2 2 drug HCO3-act 23.9 23.5 26.9 25.3 2 drug Hct 41 39 42 31 2 drug K+ 3.27 3.36 3.69 3.39 2 drug Lactate 0.6 0.77 0.67 0.67 2 drug Na++ 135.6 138.3 130.1 137.5 2 drug P02 54.7 34.3 48.1 36.6 2 drug p50 91.3 25.2 24.8 26.6 2 drug pCO2 39.3 39.7 44.9 43.1 2 drug pH 7.402 7.391 7.396 7.387 2 drug pO2 54.7 34.3 48.1 36.6 2 drug sO2 91.3 68.7 85.8 68.8

265
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
2 placebo AnGap 8.1 9.3 7.5 10.1 2 placebo BE(B) 1.7 -0.7 2.5 -0.4 2 placebo BE(ecf) 2 -0.7 3 -0.3 2 placebo BO2 4.4 4.9 4.2 2 placebo CA++ 1.17 1.12 1.2 1.14 2 placebo CA++(7.4) 1.16 1.11 1.18 1.12 2 placebo Cl- 16 110 106 107 2 placebo ctCO2 28.6 25.6 29.9 26.6 2 placebo ctHb 7.1 8 6.9 2 placebo ctO2 3.8 3.6 3.7 2 placebo FCOHb 0.6 0.6 0.5 2 placebo FHHb 13.1 28 13.5 2 placebo FMetHb 0.3 0.2 0.3 2 placebo FO@Hb 86 71.2 85.7 2 placebo Glucose 4.1 3.6 4.4 3.7 2 placebo HCO3 std 25.9 23.6 26.4 24.1 2 placebo HCO3-act 27.2 24.3 28.4 25.2 2 placebo Hct 21 24 20 2 placebo K+ 4.14 3.94 3.55 3.48 2 placebo Lactate 1.02 1.06 0.79 0.88 2 placebo Na++ 132.5 133.8 133.6 134.1 2 placebo P02 44.7 54.3 34.3 44.5 2 placebo p50 21.9 22.9 21.7 2 placebo pCO2 47.4 41.6 51 46 2 placebo pH 7.376 7.385 7.363 7.356 2 placebo pO2 44.7 34.3 44.5 2 placebo sO2 86.8 71.8 86.4

266
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
3 drug AnGap 7 8.3 7 6.7 3 drug BE(B) 1.6 0.3 1.8 0.7 3 drug BE(ecf) 2.1 0.5 2.5 1 3 drug BO2 8.5 8.2 8.1 8.1 3 drug CA++ 1.17 1.15 1.19 1.14 3 drug CA++(7.4) 1.17 1.14 1.18 1.13 3 drug Cl- 108 109 108 111 3 drug ctCO2 28.3 26.9 29.1 27.5 3 drug ctHb 13.9 13.4 13.4 13.1 3 drug ctO2 4.9 5.5 5.9 6.4 3 drug FCOHb 0.5 0.3 1.6 0.3 3 drug FHHb 42.7 34 26.6 21.5 3 drug FMetHb 0.5 0.6 1.1 0.6 3 drug FO@Hb 56.3 65.1 70.7 77.6 3 drug Glucose 4.7 4.7 4.9 48 3 drug HCO3 std 24.9 24 25.5 24.7 3 drug HCO3-act 26.9 25.6 27.6 26.1 3 drug Hct 41 39 39 39 3 drug K+ 3.82 3.7 3.91 3.61 3 drug Lactate 0.94 1.04 0.52 0.61 3 drug Na++ 138.1 139.2 138.7 140.2 3 drug P02 28 34.1 39 41.8 3 drug p50 24.8 26.1 26.2 25.4 3 drug pCO2 45 34.1 48 44.8 3 drug pH 7.395 7.384 7.378 7.383 3 drug pO2 28 34.1 39 41.8 3 drug sO2 56.9 65.7 72.7 78.3

267
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
3 placebo AnGap 10.5 9.6 8.7 4.9 3 placebo BE(B) 1.1 1.2 -0.5 -2.3 3 placebo BE(ecf) 1.1 1.4 -0.6 -2.7 3 placebo BO2 9.2 8.5 8.7 7.8 3 placebo CA++ 1.17 1.17 1.1 1.03 3 placebo CA++(7.4) 1.18 1.17 1.1 1.03 3 placebo Cl- 107 108 111 116 3 placebo ctCO2 26.8 27.4 25.5 23.3 3 placebo ctHb 15 13.9 14.1 12.7 3 placebo ctO2 7.2 5.8 7.9 6.9 3 placebo FCOHb 0.2 0.4 0.9 0.2 3 placebo FHHb 22.9 31.8 3.55 12.7 3 placebo FMetHb 0.6 0.5 9.7 0.9 3 placebo FO@Hb 76.3 67.3 0 86.2 3 placebo Glucose 5 4.9 4.4 4.1 3 placebo HCO3 std 24.9 24.8 23.9 22.3 3 placebo HCO3-act 25.5 26.1 24.2 22.2 3 placebo Hct 44 41 41 37 3 placebo K+ 3.85 3.85 3.55 3.23 3 placebo Lactate 0.63 0.67 0.46 0.37 3 placebo Na++ 139.2 139.8 140.4 139.8 3 placebo P02 38.7 33.3 54.2 51.8 3 placebo p50 25.2 25.1 90.2 26 3 placebo pCO2 40 42.4 40.4 37.1 3 placebo pH 7.423 7.424 7.396 7.394 3 placebo pO2 38.7 33.3 54.2 51.8 3 placebo sO2 76.9 67.9 89.4 87.2

268
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
4 drug AnGap 10.3 9.1 9.3 8.6 4 drug BE(B) 0.6 0.9 0.4 -0.4 4 drug BE(ecf) 0.9 1.4 0.5 -0.5 4 drug BO2 8.7 8.7 8.9 8.5 4 drug CA++ 1.16 1.15 1.13 1.11 4 drug CA++(7.4) 1.15 1.14 1.13 1.11 4 drug Cl- 107 108 108 110 4 drug ctCO2 27.3 27.9 26.6 25.7 4 drug ctHb 14.2 14.1 14.4 13.8 4 drug ctO2 6.4 6.5 7.5 7 4 drug FCOHb 0.4 0.4 0.6 0.5 4 drug FHHb 27.6 25.3 15.8 18.5 4 drug FMetHb 0.4 0.5 0.3 0.4 4 drug FO@Hb 71.6 73.8 83.3 80.6 4 drug Glucose 6.4 6.4 5.8 5.4 4 drug HCO3 std 24.4 24.7 24.5 23.8 4 drug HCO3-act 26 26.5 25.3 24.4 4 drug Hct 42 41 42 41 4 drug K+ 3.58 3.64 3.55 3.43 4 drug Lactate 1.01 1.18 0.97 1.08 4 drug Na++ 139.7 139.9 139.1 139.6 4 drug P02 38.4 38.7 46.6 43.9 4 drug p50 26.3 25.3 25.3 25.4 4 drug pCO2 44.3 45.7 41.9 40.8 4 drug pH 7.386 7.381 7.399 7.395 4 drug pO2 38.4 38.7 46.6 43.9 4 drug sO2 72.2 74.5 84.1 81.3

269
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
4 placebo AnGap 3.5 6.6 9.5 3.7 4 placebo BE(B) 1.2 0.1 0.2 0.1 4 placebo BE(ecf) 1.2 0 0.1 0 4 placebo BO2 8.3 8.2 8.4 8.7 4 placebo CA++ 1.13 1.09 1.14 1.15 4 placebo CA++(7.4) 1.14 1.1 1.15 1.16 4 placebo Cl- 108 111 107 108 4 placebo ctCO2 26.9 25.9 25.9 25.7 4 placebo ctHb 13.4 13.2 13.6 14.1 4 placebo ctO2 5.8 5.7 6.6 7.8 4 placebo FCOHb 0.2 0.2 0.1 0.4 4 placebo FHHb 30.5 31.1 21.6 11.4 4 placebo FMetHb 0.4 0.2 0.4 0.3 4 placebo FO@Hb 68.9 68.5 77.9 87.9 4 placebo Glucose 5.8 5.5 6 6 4 placebo HCO3 std 24.9 24 24.2 24.4 4 placebo HCO3-act 25.7 24.7 24.7 24.5 4 placebo Hct 39 39 40 41 4 placebo K+ 3.81 3.64 3.7 3.79 4 placebo Lactate 1.04 1.14 1 1.03 4 placebo Na++ 133.4 138.6 137.5 132.4 4 placebo P02 34.4 34.5 40.2 50.9 4 placebo p50 25.8 25.8 25.3 24.9 4 placebo pCO2 40.2 39.6 39.7 38.9 4 placebo pH 7.423 7.412 7.412 7.417 4 placebo pO2 34.4 34.5 40.2 50.9 4 placebo sO2 69.3 68.8 78.3 88.5

270
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
5 drug AnGap 9 12.3 10 10.7 5 drug BE(B) 0.3 0.9 -0.8 -0.7 5 drug BE(ecf) 0.1 1 -0.9 -0.8 5 drug BO2 8.8 9.2 8.6 8.7 5 drug CA++ 1.16 1.22 1.2 1.21 5 drug CA++(7.4) 1.17 1.22 1.2 1.21 5 drug Cl- 111 108 111 110 5 drug ctCO2 25.9 27.1 25.2 25.3 5 drug ctHb 14.3 15 13.9 14.2 5 drug ctO2 6.9 7.3 7 7.5 5 drug FCOHb 0.3 0.4 0.3 0.3 5 drug FHHb 22.01 21.11 19.1 15 5 drug FMetHb 0.4 0.3 0.5 0.5 5 drug FO@Hb 77.3 78.2 80.1 84.2 5 drug Glucose 5.6 6.2 7.6 8.11 5 drug HCO3 std 24.2 24.7 23.4 23.6 5 drug HCO3-act 24.7 25.8 23.9 24.1 5 drug Hct 42 44 41 42 5 drug K+ 3.82 4.19 3.74 3.87 5 drug Lactate 0.62 0.77 1 1.16 5 drug Na++ 140.9 141.9 141.2 140.9 5 drug P02 39.6 39.3 41.1 44.2 5 drug p50 25.2 24.2 24.1 23.5 5 drug pCO2 39.4 42.3 40.1 40 5 drug pH 7.415 7.403 7.394 7.397 5 drug pO2 39.6 39.3 41.1 44.2 5 drug sO2 77.8 78.8 80.7 84.9

271
PLACEBO
Subject Visit Variable BL NORM FAT NORM
BL HEAT
FAT HEAT
5 placebo AnGap 8.2 7.9 4.5 6 5 placebo BE(B) 5.4 1.6 -4 0.1 5 placebo BE(ecf) 4.5 2.1 -4.7 0.2 5 placebo BO2 8.8 8.5 8.1 8.1 5 placebo CA++ 0.92 1.1 0.97 1.11 5 placebo CA++(7.4) 0.99 1.1 0.96 1.11 5 placebo Cl- 112 109 119 113 5 placebo ctCO2 27.3 28.4 21.7 26.4 5 placebo ctHb 14.3 14 13.3 13.2 5 placebo ctO2 6.9 6.7 5.5 6.5 5 placebo FCOHb 0.3 1.4 1.1 1.1 5 placebo FHHb 22.01 21.7 32 20.3 5 placebo FMetHb 0.4 0.3 0.2 0.2 5 placebo FO@Hb 77.3 76.6 66.7 78.4 5 placebo Glucose 6.1 5.3 4.2 4.9 5 placebo HCO3 std 29.4 25.4 20.5 24.2 5 placebo HCO3-act 26.4 27 20.6 25.1 5 placebo Hct 42 41 39 39 5 placebo K+ 4.14 4.50 2.9 3.43 5 placebo Lactate 1.42 1.25 0.88 1.22 5 placebo Na++ 132.7 136.2 141.2 140.7 5 placebo P02 41.47 38.4 32.6 38.8 5 placebo p50 25.2 23.6 24.1 23.3 5 placebo pCO2 28.8 45.6 36.4 42 5 placebo pH 7.58 7.391 7.37 7.394 5 placebo pO2 39.6 38.4 32.6 38.8 5 placebo sO2 77.8 77.9 67.6 79.4

272
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
6 drug AnGap 8.4 7 7.2 6.9 6 drug BE(B) 4.9 3.8 2.5 4.3 6 drug BE(ecf) 6.2 4.9 3.1 5.3 6 drug BO2 8.3 8.3 8.1 8.5 6 drug CA++ 1.23 1.19 1.14 1.18 6 drug CA++(7.4) 1.23 1.18 1.14 1.18 6 drug Cl- 105 106 109 107 6 drug ctCO2 32.7 31.5 29.3 31.7 6 drug ctHb 13.6 13.5 13.2 13.9 6 drug ctO2 6.3 5.8 6.9 6.3 6 drug FCOHb 0.8 0.7 0.6 0.7 6 drug FHHb 24.81 30.5 15 26.6 6 drug FMetHb 0.5 0.4 0.5 0.5 6 drug FO@Hb 73.9 68.4 83.9 72.2 6 drug Glucose 5.4 5 5 5 6 drug HCO3 std 28.3 27.2 26.4 27.7 6 drug HCO3-act 31.1 29.9 27.9 30.2 6 drug Hct 40 40 39 41 6 drug K+ 4.22 3.96 3.52 3.77 6 drug Lactate 1.01 1.02 0.87 0.98 6 drug Na++ 140.3 138.9 140.5 140.3 6 drug P02 38.9 35.7 46 37 6 drug p50 25.2 25.5 24.4 25 6 drug pCO2 52.3 51 45.9 50.2 6 drug pH 7.392 7.386 7.4 7.397 6 drug pO2 38.9 35.7 46 37 6 drug sO2 74.9 69.2 84.8 73.1

273
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
6 placebo AnGap 10.3 14.7 2.5 16.2 6 placebo BE(B) 3 2.1 0.2 1.9 6 placebo BE(ecf) 3.7 4 0.1 3.6 6 placebo BO2 8.7 9.6 8.2 9.9 6 placebo CA++ 1.21 1.27 1.06 1.24 6 placebo CA++(7.4) 1.21 1.22 1.06 1.2 6 placebo Cl- 104 103 112 104 6 placebo ctCO2 30 32.1 26 31.5 6 placebo ctHb 14.1 15.6 13.3 16.1 6 placebo ctO2 8 3.7 7.6 4 6 placebo FCOHb 0 0.6 0.3 0.5 6 placebo FHHb 8.7 61 8.5 59.8 6 placebo FMetHb 0.7 0.6 0.6 0.5 6 placebo FO@Hb 90.6 37.8 90.6 39.2 6 placebo Glucose 5.2 5.1 4.5 5 6 placebo HCO3 std 27 24.7 24.5 24.5 6 placebo HCO3-act 28.5 30.2 24.8 29.7 6 placebo Hct 14 46 39 47 6 placebo K+ 3.8 5 3.16 5.12 6 placebo Lactate 0.79 1.06 0.74 0.87 6 placebo Na++ 139 142.8 136.1 144.8 6 placebo P02 55.5 23.1 57 26.2 6 placebo p50 55.5 25 28 6 placebo pCO2 47.1 61.2 40 58.8 6 placebo pH 7.4 7.31 7.41 7.321 6 placebo pO2 55.5 23.1 57 26.2 6 placebo sO2 91.6 38.3 91.4 39.6

274
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
7 drug AnGap 12.5 8.9 11.2 10.5 7 drug BE(B) 1.6 -0.3 0.4 0.3 7 drug BE(ecf) 2.6 -0.1 0.7 0.7 7 drug BO2 9.8 9.4 9.7 9.5 7 drug CA++ 1.25 1.14 1.21 1.21 7 drug CA++(7.4) 1.23 1.13 1.2 1.2 7 drug Cl- 105 111 107 108 7 drug ctCO2 29.4 26.5 27.1 27.2 7 drug ctHb 15.9 15.3 15.8 15.5 7 drug ctO2 6 7 8.1 8.3 7 drug FCOHb 0.1 0.2 0.2 0.2 7 drug FHHb 39.1 26.2 17.1 13.8 7 drug FMetHb 0.5 0.6 0.6 0.6 7 drug FO@Hb 60.3 73 82.1 85.4 7 drug Glucose 5.1 4.7 5 5 7 drug HCO3 std 24.9 23.6 24.4 24.5 7 drug HCO3-act 27.9 25.2 25.7 25.8 7 drug Hct 47 45 46 46 7 drug K+ 4.05 3.59 3.93 3.94 7 drug Lactate 0.32 0.47 0.27 0.42 7 drug Na++ 141.3 141.5 140 140.4 7 drug P02 30.7 37.3 43.5 46 7 drug p50 24.9 24.6 24.1 23.2 7 drug pCO2 49.4 44.1 43.9 44.8 7 drug pH 7.369 7.374 7.386 7.379 7 drug pO2 30.7 37.3 43.5 46 7 drug sO2 60.7 73.6 82.8 86.1

275
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
7 placebo AnGap 10 9.3 8.7 8 7 placebo BE(B) -0.6 -1.1 -1.1 -1.8 7 placebo BE(ecf) -0.5 -1.2 -1.2 -2.1 7 placebo BO2 9.5 9.1 8.7 8.7 7 placebo CA++ 1.119 1.14 1.14 1.1 7 placebo CA++(7.4) 1.18 1.13 1.14 1.1 7 placebo Cl- 110 111 111 112 7 placebo ctCO2 26.1 25.1 25 24.2 7 placebo ctHb 15.4 14.8 14.2 14.1 7 placebo ctO2 6.8 7.3 6 7.4 7 placebo FCOHb 0.4 0.3 0.6 0.3 7 placebo FHHb 28.1 20.5 31.7 139.6 7 placebo FMetHb 0.4 0.6 0.3 0.4 7 placebo FO@Hb 71.1 78.6 67.4 84 7 placebo Glucose 5 4.9 4.5 4.6 7 placebo HCO3 std 23.3 23.1 22.9 22.6 7 placebo HCO3-act 24.8 23.9 23.7 23 7 placebo Hct 45 44 42 41 7 placebo K+ 3.53 3.42 3.4 1.11 7 placebo Lactate 1.13 1.22 1.1 1.18 7 placebo Na++ 141.2 140.8 140 139.6 7 placebo P02 37.1 43.9 34.5 46.6 7 placebo p50 25.4 26.3 25.7 24.7 7 placebo pCO2 43.3 40.9 40 39.6 7 placebo pH 7.375 7.384 7.391 7.382 7 placebo pO2 37.1 43.9 34.5 46.6 7 placebo sO2 71.7 79.3 68 84.6

276
DRUG
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
8 drug AnGap 6.2 8.6 11.2 7.5 8 drug BE(B) 0.1 -0.6 1.3 -2.4 8 drug BE(ecf) 0.1 -0.6 1.6 -3 8 drug BO2 9.3 9 9.5 8.1 8 drug CA++ 1.12 1.08 1.14 0.99 8 drug CA++(7.4) 1.12 1.08 1.14 0.99 8 drug Cl- 110 111 106 114 8 drug ctCO2 26.1 25.6 27.7 23 8 drug ctHb 15.1 14.6 15.5 13.2 8 drug ctO2 8.1 7.3 8.4 7 8 drug FCOHb 0.4 0.4 0.3 0.2 8 drug FHHb 13.5 19 12.1 14.7 8 drug FMetHb 0.7 0.7 0.7 0.8 8 drug FO@Hb 85.4 79.9 86.9 84.3 8 drug Glucose 5.3 5 6 4.8 8 drug HCO3 std 24.3 23.6 25.3 22.2 8 drug HCO3-act 24.9 24.3 26.3 21.9 8 drug Hct 44 43 46 39 8 drug K+ 3.48 3.32 3.85 3.06 8 drug Lactate 1.1 1.12 1.05 0.93 8 drug Na++ 137.6 140.6 139.7 140.3 8 drug P02 47.3 42.9 49 45.9 8 drug p50 24.3 25 24.1 24.4 8 drug pCO2 40.8 41 43.3 36.3 8 drug pH 7.403 7.391 7.402 7.398 8 drug pO2 47.3 42.9 49 45.9 8 drug sO2 86.3 80.8 87.8 85.2

277
PLACEBO
Subject Visit Variable BL NORM
FAT NORM
BL HEAT
FAT HEAT
8 placebo AnGap 9.3 9.9 4.5 6.2 8 placebo BE(B) 1.5 0.8 1.9 -2.7 8 placebo BE(ecf) 2.5 1.7 2.8 -3 8 placebo BO2 9.4 9.3 9.3 7.9 8 placebo CA++ 1.19 1.13 1.22 0.98 8 placebo CA++(7.4) 1.17 1.11 1.2 0.97 8 placebo Cl- 107 108 107 116 8 placebo ctCO2 29.6 28.7 29.6 23.6 8 placebo ctHb 15.3 15.1 15.2 12.8 8 placebo ctO2 6.6 7 7 6.6 8 placebo FCOHb 0.3 0.2 0.6 0.3 8 placebo FHHb 30.4 24.9 25.6 17.3 8 placebo FMetHb 0.7 0.6 0.5 0.06 8 placebo FO@Hb 68.6 74.3 73.3 81.8 8 placebo Glucose 6.2 6 6 4.7 8 placebo HCO3 std 25 24.6 25.5 21.9 8 placebo HCO3-act 28.1 27.2 28.1 22.3 8 placebo Hct 45 44 45 38 8 placebo K+ 5.94 3.936 4.24 3.9 8 placebo Lactate 1.04 1.12 0.87 0.89 8 placebo Na++ 138.4 137.8 135.3 140.6 8 placebo P02 35.3 38 37.5 46 8 placebo p50 24.5 23.9 24.2 25.3 8 placebo pCO2 51.5 49.8 49.8 39.7 8 placebo pH 7.354 7.355 7.369 7.368 8 placebo pO2 35.3 38 37.5 46 8 placebo sO2 69.3 74.9 74.1 82.5

Nathan T. Kuipers
VITA
Education Institution Degree Discipline Date Completed Calvin College B.S. Biology 1999 Slippery Rock University
M.S. Exercise Science 2001
Pennsylvania State University
Ph.D. I.B.I.O.S. 2008
Publications
S.C. Newcomer, C. L. Sauder, N.T. Kuipers, M.H. Laughlin, and C.A. Ray (2008). Effects of posture on shear rates in human brachial and superficial femoral arteries. Am J Physiol H 294, H1833-H1839. N.T. Kuipers, C. L. Sauder, and C. A. Ray (2008). Neurovascular responses to mental stress in the supine and upright postures. J Appl Physiol 104:1129-1136. N.T. Kuipers, C.L. Sauder, M.L. Kearney, and C. A. Ray. (2007). Changes in forearm muscle temperature alter renal vascular responses to isometric handgrip. Am J Physio H 293: H3432-H3439. T. E. Wilson, C. L. Sauder, M.L. Kearney, N. T. Kuipers, U.A. Leuenberger, K.D. Monahan and C. A. Ray (2007). Skin-surface cooling elicits peripheral and visceral vasoconstriction in humans. J Appl Physiol 103, 1257-1262. N.T. Kuipers, C.L. Sauder, and C.A. Ray. (2007). Influence of static magnetic fields on pain perception and sympathetic nerve activity in humans. J Appl Physiol 102, 1410-1415. J. R. Carter, N. T. Kuipers, and C. A. Ray (2005). Neurovascular responses to mental stress. J Physiol 564: 321-327. T. E. Wilson, N. T. Kuipers, E. A. McHugh, and C. A. Ray (2004). Vestibular activation does not influence skin sympathetic nerve responses during whole body heating. J Appl Physiol 97: 540-544. N. T. Kuipers, C. L. Sauder, and C. A. Ray (2003). Aging attenuates the vestibulorespiratory reflex in humans. J Physiol 548: 955-961.
Related Documents











