EFFECT OF CORTICOSTEROID ON LUNG PARENCHYMA REMODELING AT AN EARLY PHASE OF ACUTE LUNG INJURY PATRICIA R.M. ROCCO 1 , ALBA B. SOUZA 1 , DEBORA S. FAFFE 1 , CAROLINE P. PÁSSARO 1 , FLÁVIA B. SANTOS 1 , ELNARA M. NEGRI 2 , JANUÁRIO G.M. LIMA 1 , RENATA S. CONTADOR 1 , VERA L. CAPELOZZI 3 , WALTER A. ZIN 1 1 Laboratory of Respiration Physiology, Carlos Chagas Filho Biophysics Institute, Federal University of Rio de Janeiro, Centro de Ciências da Saúde, Ilha do Fundão, 21949-900, Rio de Janeiro, Brazil; 2 LIM59 and 3 Department of Pathology and Department of Clinical Emergencies, University of São Paulo, São Paulo, Brazil. Correspondence address: Walter Araujo Zin Universidade Federal do Rio de Janeiro Instituto de Biofísica Carlos Chagas Filho - C.C.S. Laboratório de Fisiologia da Respiração Ilha do Fundão 21949-900 - Rio de Janeiro - RJ Brazil e-mail: [email protected] tel: (+5521) 2562-6557 fax: (+5521) 2280-8193 Supported by: Centers of Excellence Program (PRONEX-MCT), Brazilian Council for Scientific and Technological Development (CNPq), Financing for Studies and Projects (FINEP), Rio de Janeiro State Research Supporting Foundation (FAPERJ), São Paulo State Research Support Foundation (FAPESP). Running head: Steroid effects on lung parenchyma remodeling Subject of the manuscript: 1 Word count: 4253 This article has an online data supplement, which is accessible from this issue’s table content online at www.atsjournals.org. Copyright (C) 2003 by the American Thoracic Society. AJRCCM Articles in Press. Published on July 3, 2003 as doi:10.1164/rccm.200302-256OC

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EFFECT OF CORTICOSTEROID ON LUNG PARENCHYMA
REMODELING AT AN EARLY PHASE OF ACUTE LUNG INJURY
PATRICIA R.M. ROCCO1, ALBA B. SOUZA1, DEBORA S. FAFFE1, CAROLINE P.
PÁSSARO1, FLÁVIA B. SANTOS1, ELNARA M. NEGRI2, JANUÁRIO G.M. LIMA1,
RENATA S. CONTADOR1, VERA L. CAPELOZZI3, WALTER A. ZIN1
1Laboratory of Respiration Physiology, Carlos Chagas Filho Biophysics Institute, Federal
University of Rio de Janeiro, Centro de Ciências da Saúde, Ilha do Fundão, 21949-900, Rio
de Janeiro, Brazil; 2LIM59 and 3Department of Pathology and Department of Clinical
Emergencies, University of São Paulo, São Paulo, Brazil.
Correspondence address: Walter Araujo Zin
Universidade Federal do Rio de Janeiro Instituto de Biofísica Carlos Chagas Filho - C.C.S. Laboratório de Fisiologia da Respiração Ilha do Fundão 21949-900 - Rio de Janeiro - RJ Brazil e-mail: [email protected] tel: (+5521) 2562-6557 fax: (+5521) 2280-8193
Supported by: Centers of Excellence Program (PRONEX-MCT), Brazilian Council for
Scientific and Technological Development (CNPq), Financing for Studies and Projects
(FINEP), Rio de Janeiro State Research Supporting Foundation (FAPERJ), São Paulo State
Research Support Foundation (FAPESP).
Running head: Steroid effects on lung parenchyma remodeling
Subject of the manuscript: 1
Word count: 4253
This article has an online data supplement, which is accessible from this issue’s table
content online at www.atsjournals.org.
Copyright (C) 2003 by the American Thoracic Society.
AJRCCM Articles in Press. Published on July 3, 2003 as doi:10.1164/rccm.200302-256OC

Blue-200302-2560C-R1-Clean version 2
ABSTRACT
In vivo [lung resistive and viscoelastic pressures, and static elastance] and in vitro [tissue
resistance, elastance, and hysteresivity] respiratory mechanics were analyzed 1 and 30 days
after saline (control) or paraquat [P (10 and 25 mg/kg, i.p.)] injection in rats. Additionally,
P10 and P25 were treated with methylprednisolone (2 mg/kg, i.v.) at 1 or 6 hours after
acute lung injury (ALI) induction. Collagen and elastic fibers were quantified. Lung
resistive and viscoelastic pressures, and static elastance were higher in P10 and P25 than in
control. Tissue elastance and resistance augmented from control to P10 (1 and 30 days) and
P25. Hysteresivity increased only in P25. Methylprednisolone at 1 or 6 hours attenuated in
vivo and in vitro mechanical changes in P25, while P10 parameters were similar to control.
Collagen increment was dose- and time-dependent. Elastic fibers increased in P25, and at
30 days in P10. Corticosteroid prevented collagen increment, and avoided elastogenesis. In
conclusion, methylprednisolone led to a complete maintenance of in vivo and in vitro
respiratory mechanics in mild lesion, whereas minimized the changes in tissue impedance
and extracellular matrix in severe ALI. The beneficial effects of the early use of steroids in
ALI remained unaltered at day 30.
Word count: 200
Key words: Tissue mechanics, acute lung injury, corticosteroid, hysteresivity, elastin

Blue-200302-2560C-R1-Clean version 3
The use of corticosteroid in the treatment of acute lung injury (ALI)/acute
respiratory distress syndrome (ARDS) has been subject of great controversy and debate
over the years. Although the exact mechanism of action remains unknown, corticosteroids
inhibit a host of potent inflammatory mediators and have been shown to improve morbidity
and mortality in animal models of ALI (1-4). Because of the great difficulties in designing
and interpreting studies in humans, much of our knowledge about ALI/ARDS
pathophysiology comes from animal experimentation.
Corticosteroid therapy in ARDS has been studied in three main different situations:
a) prevention in high risk patients (5), b) early treatment with high dose, short course
therapy (6), and c) prolonged therapy in unresolved cases (7-10). Unfortunately, trials of
short term, high dose steroid therapy failed to show an improvement in mortality of patients
at risk of or with early ARDS (5-6). Despite the unfavorable experience with steroids at the
early phase of ALI, there has been a recent resurgence in enthusiasm for their use in late
ALI (fibroproliferative phase) (8). The late phase of ALI is characterized by progressive
pulmonary fibrosis and lung restriction (11). In addition, pulmonary fibrosis contributes to
the unremitting respiratory failure and death in a significant proportion of patients with ALI
(12). Several investigators have suggested that the use of corticosteroids in the late phase
improved lung function and survival (7-10). Steroids prevent excessive collagen deposition
and increase collagen breakdown (9, 13), however, its effect on the elastic system in ALI is
much less understood.
Some authors have recently observed an increased number of myofibroblasts and
cells producing procollagen types I and III in the early course of ALI (14-18), suggesting
that the proliferative phase begins much sooner than had been previously appreciated.

Blue-200302-2560C-R1-Clean version 4
Complementary, we observed pronounced mechanical changes at tissue level and
fibroelastogenesis at an early phase of acute lung injury, even in mild abnormal lung
parenchyma (19). Thus, in the present study we tested the hypothesis that low-dose
corticosteroid when used at an early phase of acute lung injury could modify: a) in vivo and
in vitro respiratory mechanics, b) lung histology, and c) the structural remodeling of lung
parenchyma in two different degrees of paraquat-induced ALI. These parameters were
studied at 1 and 30 days after the induction of lung injury. We also examined whether
oscillatory tissue mechanical data were correlated with collagen or elastic system fiber
contents.
Some of the results of this study have been previously reported in the form of
abstract (20, 21).
METHODS (Word count: 497)
Animal Preparation
Seventy-eight Wistar rats (250-300 g) were divided into 13 groups of 6 animals each. In the
control group (C) saline (5 ml/kg BW) was administered intraperitoneally (i.p.). In P
groups two different doses of paraquat were injected i.p. (10 and 25 mg/kg BW), 1 or 30
days prior to the measurements. P10 and P25 groups were treated with methylprednisolone
(M, 2 mg/kg, i.v.) at 1 or 6 hours after paraquat administration. Animals were sedated
(diazepam 5 mg i.p.), anesthetized (pentobarbital sodium 20 mg/kg i.p.), and a snugly
fitting cannula (1.7 mm ID) was introduced into the trachea. Airflow, volume, tracheal,
transpulmonary, and esophageal pressures were registered. Respiratory system (rs), lung
(L), and chest wall (w) resistive (∆P1), and viscoelastic/inhomogeneous pressures (∆P2),

Blue-200302-2560C-R1-Clean version 5
∆Ptot (∆P1+∆P2), and static elastance (Est) were computed by end-inflation occlusion
method (22, 23). Lungs were removed en bloc, and placed in a modified Krebs-Henseleith
(K-H) solution [in mM: 118.4 NaCl, 4.7 KCl, 1.2 K3PO4, 25 NaHCO3, 2.5 CaCl2.H2O, 0.6
MgSO4.H2O, and 11.1 glucose] at pH = 7.40 and 6°C (11-13), bubbled with 95% O2-5%
CO2. Strips (3x3x10 mm) were cut from the periphery of the left lung, and suspended
vertically in a K-H organ bath maintained at 37°C and continuously bubbled with 95% O2-
5% CO2. One clip was attached to a force transducer, while the other one was fastened to a
vertical rod. This fiberglass stick was connected to a woofer cone, which was driven by a
waveform generator. A side arm of the rod was linked to a second force transducer by
means of a silver spring of known Young’s modulus, allowing the measurement of
displacement (19, 24).
Strips were preconditioned by sinusoidally oscillating the tissue during 30 min
(frequency = 1 Hz; amplitude large enough to reach a final force of 1x10-2 N). Thereafter
the amplitude was adjusted to 5% of the strip’s resting length (L0), the strips were unloaded
to a force of 8x10-3 N, and the oscillation maintained for another 30 min, or until a stable
length-force loop was reached. The final basal force was approximately 5x10-3 N. After
stress adaptation, strips were oscillated at a frequency = 1 Hz.
Tissue resistance (R), elastance (E), and hysteresivity (η) were calculated from the
oscillatory recordings according to Fredberg and Stamenovic (25).
Morphometric analysis
The right lung was fixed at the end of expiration with glutaraldeyde and submitted to
ultramicrotomy for transmission electron microscopy. Left lung and parenchymal strips
were quick-frozen by immersion in liquid nitrogen and fixed with Carnoy's solution (26,

Blue-200302-2560C-R1-Clean version 6
27). Slices 4-µm-thick were cut and underwent hematoxylin-eosin and specific staining
methods to quantify the collagenous [Picrosirius-polarization method (28)] and elastic
system [Weigert's resorcin fuchsin method modified with oxidation (29)] fibers in the
alveolar septa.
Statistical analysis
SigmaStat 2.0 statistical software package (Jandel Corporation, CA, USA) was used.
Differences among groups were assessed by one-way analysis of variance and the Tukey
test. Correlation between mechanical and histological data was determined by Spearman
correlation test. A p value < 0.05 was considered significant.
RESULTS
The survival rate in P10 and P25 groups was 85% and 50%, respectively, within the first
day after injection. At day 30, survival was 33% in P10. Steroid led to 100% survival in
P10 group, but presented no beneficial effect in rats from P25 group, which died within 48
hours after ALI induction (Table 1).
In vivo mechanics
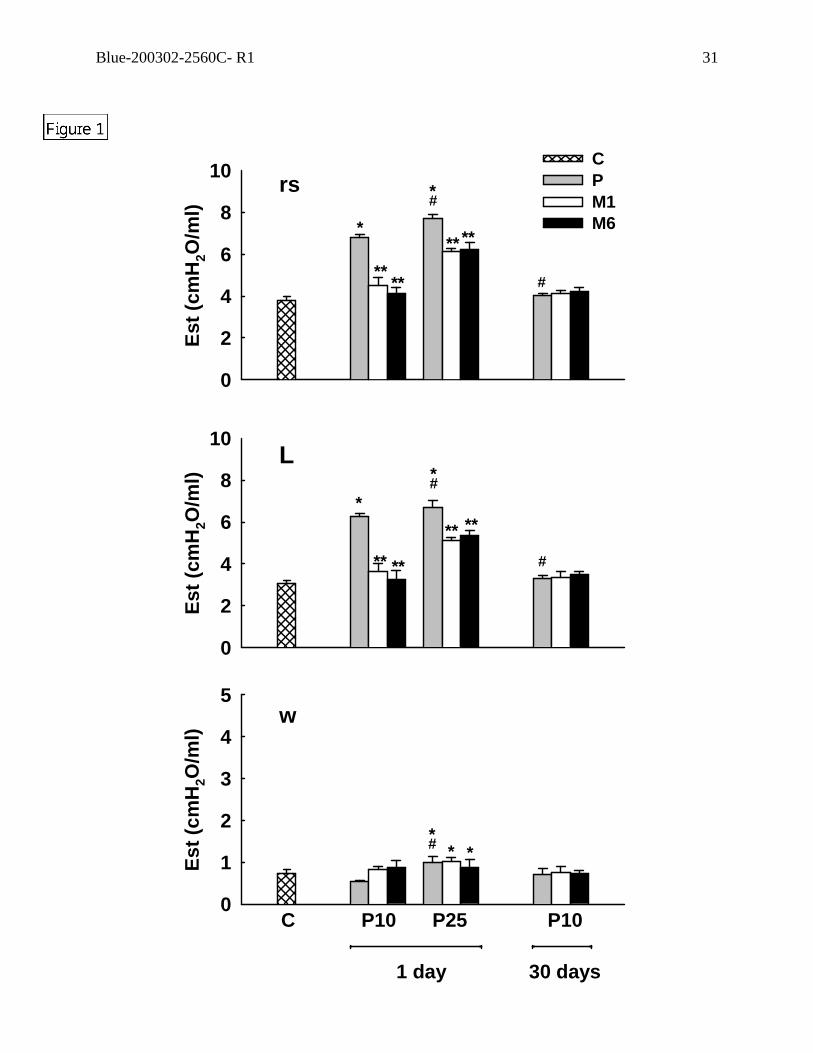
Respiratory system and lung static elastances (Figure 1), resistive,
viscoelastic/inhomogeneous, and total pressures (Figure 2) increased significantly with the
severity of lung injury (from P10 to P25). Respiratory system and lung static elastances and
resistive pressures returned to control values at 30 days in P10, while respiratory system
and lung viscoelastic and total pressure variations remained higher than control. P10 group
treated with steroid presented values of mechanical parameters similar to control group.
The beneficial effects of methylprednisolone were similar when given 1 or 6 hours after

Blue-200302-2560C-R1-Clean version 7
ALI induction, and remained unaltered at day 30. In P25 group, corticosteroid prevented
the modifications in Est,rs, Est,L, ∆P1,rs, and ∆P1,L, and attenuated ∆P2,rs, ∆P2,L,
∆Ptot,rs, and ∆Ptot,L changes. Chest wall total and resistive pressures increased in P25
group, and steroid did not modify these changes (Figures 1 and 2).
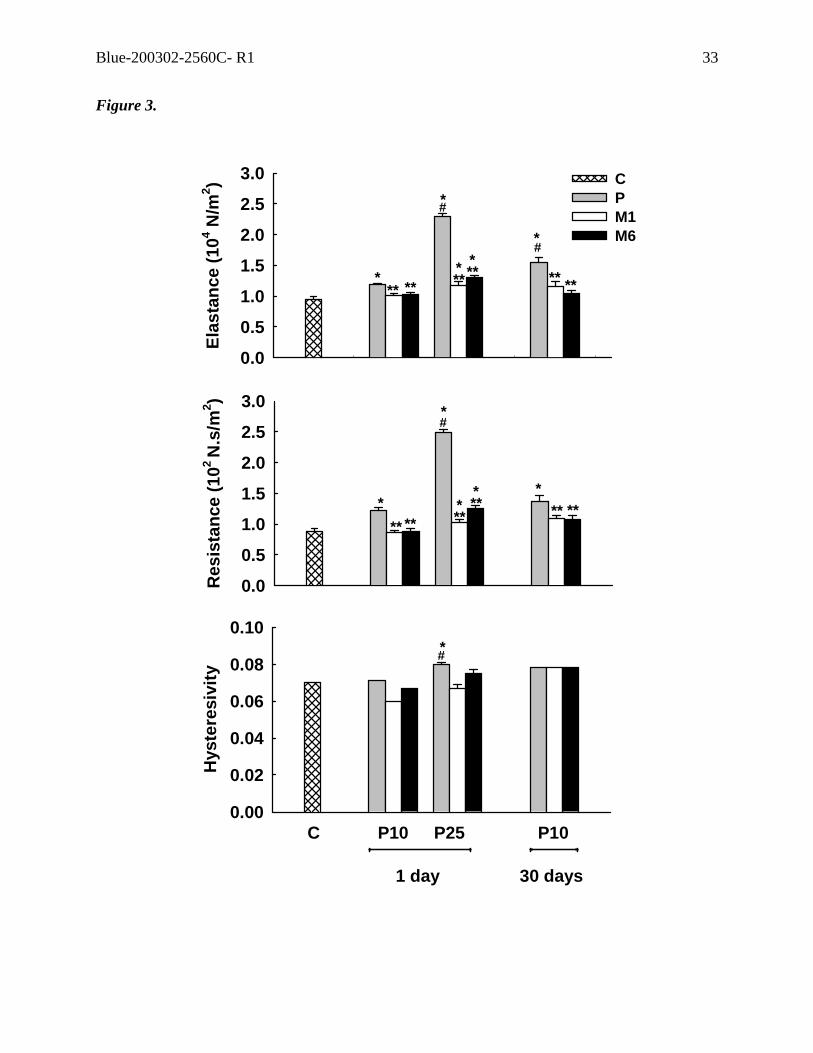
In vitro mechanics
E and R augmented from C to P10 and P25. In P10, E was higher at 30 days than at the first
day after ALI induction, while R was similar independently of the time course (Figure 3).
The use of methylprednisolone, either 1 or 6 hours after ALI induction, yielded E and R
values similar to control in P10 group, and attenuated the changes in P25. η increased only
in P25 group (Figure 3).
Lung histology
Typical photomicrographs of lung parenchyma from control, P10 at 1 and 30 days, and P25
groups with and without steroid 6 hours after ALI induction are shown in Figure 4. Because
lung histology was similar in groups M1 and M6, we depicted only one treatment (6 hours).
Lung histological changes included atelectasis, interstitial edema, and inflammation with
polymorphonuclear (PMN) cells in P10 group (Figure 4B). Lung parenchyma in P25
presented an increment in all histological changes described above, together with alveolar
edema and intra-alveolar hemorrhage (Figure 4D). P10 lung parenchyma at day 30 showed
only thickened alveolar membranes and increased cellularity in comparison to control
group (Figure 4F). Both P10 groups treated with steroid presented less interstitial edema
and tissue cellularity than P10 independent of the time of analysis (Figures 4C and G). In
P25 group, alveolar modifications were prevented by corticosteroid, while interstitial
edema and inflammatory cells, although lessened, remained increased (Figure 4E).

Blue-200302-2560C-R1-Clean version 8
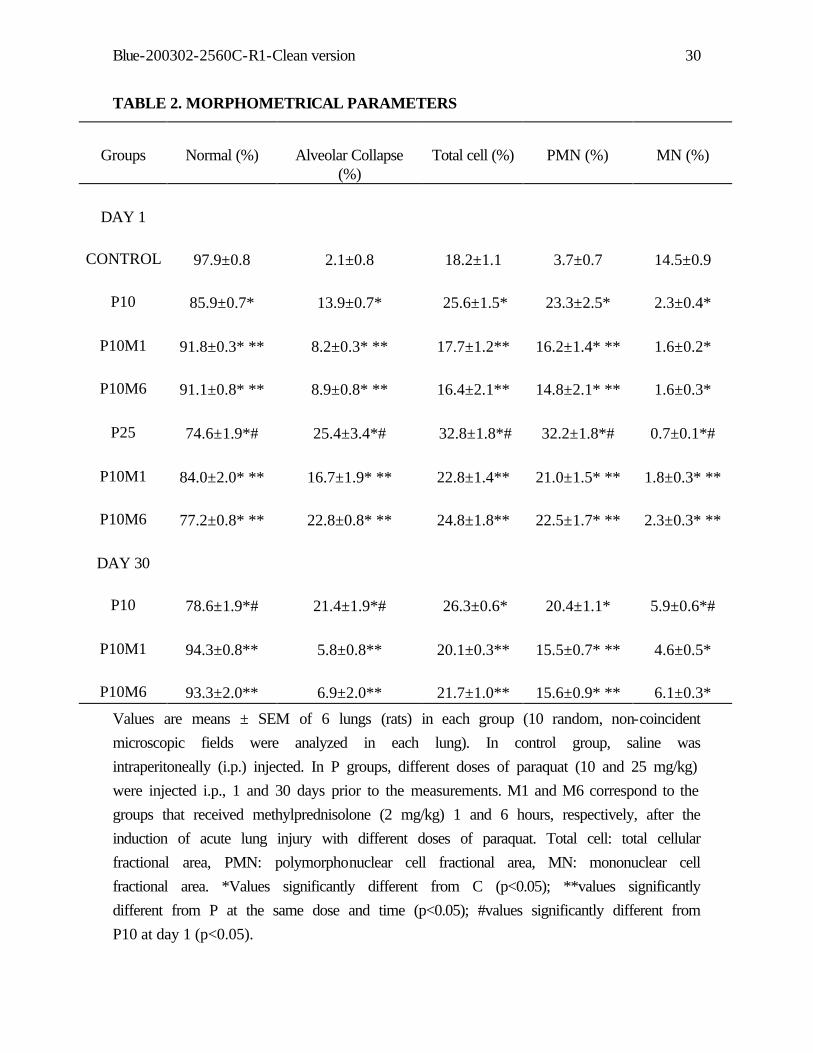
Morphometrical examination of the lung demonstrated significant increase in alveolar
collapse in all P groups in relation to control (Table 2). Steroid reduced alveolar collapse in
P10 and P25 groups, although they remained higher than control at day 1 and similar to
control group at day 30. Alveolar hyperinflation was not present in any group. Total and
polymorphonuclear cell contents augmented from control to P10 and P25, but there was no
difference in cell count between P10 at days 1 and 30 (Table 2). Steroid presented anti-
inflammatory effects on acute lung injury independently its severity. Mononuclear cell
content was smaller in P25 and P10 than in control group. Mononuclear cells increased
with the time course of the lesion (Table 1).
Figure 5 shows the amounts of collagen and elastic system fibers in the alveolar
septa from lung tissue strips of control (C) and acute lung injury groups (P10 and P25), at
day 1 and 30, and after methylprednisolone (2 mg/kg) injection 1 or 6 hours after ALI
induction. Collagen fiber content increased with the severity of lung injury and with the
temporal evolution of ALI (Figure 5). Corticosteroid attenuated the increment in collagen
content independently of ALI degree. Additionally, its beneficial effects were present until
30 days after steroid injection. Elastic system fiber content increased only at 30 days after
ALI induction in P10, augmenting earlier in the more severe lesion (P25). Steroid
administration maintained the elastic fiber content equal to control value independently of
the dose (severity of the lesion) at day 1 and time (P10, 30 days) (Figure 5).
Figure 6 shows the ultramicroscopy of lung parenchyma. In P10 group
inflammatory cells including lymphocytes, plasma cells, macrophages, fibroblasts, and
primitive mesenchymal cell (PMC) were seen in the interstitium. The alveolar interstitium
was thickened due to both edema and increased amounts of matrix elements, such as
collagen fibers. The alveolar spaces had a mixed inflammatory infiltrate consisting of PMN

Blue-200302-2560C-R1-Clean version 9
and alveolar macrophages. Fibroblasts were frequently found together with types I and III
collagen fibers and elastin (Figure 6B). Steroid decreased the number of fibroblasts and
type III collagen fiber content. At day 1, elastic fiber content was normal (Figure 6C).
However, in P10 group 30 days after ALI induction and in P25 there was an increased
number of types I and III collagen fibers and elastin, thus characterizing fibroelastosis
(Figures 6D and F). Steroid treatment inhibited the aggregation process of type III collagen
fibers into type I (Figures 6C, E, and G). In addition, the amount of elastic fibers was also
normal (Figures 6E and G).
Considering all groups together, tissue elastance and resistance were correlated
significantly with total cell count [p=0.007 (r=0.37) and p=0.001 (r=0.44), respectively],
and with polymorphonuclear cell count [p=0.002 (r=0.42) and p=0.002 (r=0.42),
respectively]. Additionally, tissue elastance and resistance values were correlated
significantly with collagen [p<0.001 (r=0.65) and p<0.001 (r=0.57), respectively] and
elastic fiber content [p=0.01 (r=0.36) and p=0.01 (r=0.34), respectively). Hysteresivity was
correlated only with elastic fiber content [p=0.02 (r=0.33)].
DISCUSSION
In the present study, low-dose corticosteroid administered at an early phase of acute lung
injury kept unaltered in vivo and in vitro mechanical parameters in mildly abnormal lung
parenchyma, whereas it minimized the changes in tissue viscoelastic properties in a severe
lesion. Paraquat-induced ALI led to a time- and dose-dependent increase in the amount of
collagen and elastic fibers. Furthermore, the amount of elastic system fibers augmented
later in the course of mild ALI. Corticosteroid treatment acted on the remodeling process,
significantly reducing collagen fibers deposition in mild and severe ALI, and also

Blue-200302-2560C-R1-Clean version 10
preventing elastogenesis. In addition, the use of steroids at the early phase of ALI showed
beneficial effects 24 hours after lung injury induction, which lasted until the 30th day post-
injection in the mild form.
It has been previously described that corticosteroid, when given in large doses
before or shortly after endotoxin administration in sheep, prevent the subsequent increase in
lung vascular permeability, but do not reverse the abnormality once established (30). In
addition, these abnormalities resolve within 24 hours even if no treatment is given. These
results differ from ours, where 50% of the animals with 25 mg/kg paraquat-induced ALI
died at the first 24 hours and 100% at 48 hours. At autopsy, these animals presented acute
tubular necrosis, esteatosis, carditis, and diffuse alveolar damage. The use of corticosteroid
did not change survival rate in the severe lesion. On the other hand, steroid increased
survival in 10 mg/kg paraquat-induced ALI (Table 1).
Methylprednisolone was chosen as steroid therapy because it is commonly used as
an anti-inflammatory agent in the treatment of human pulmonary fibrosis and acute
respiratory distress syndrome (7-10). Corticosteroids modulate the host defence response at
virtually all levels, protecting the host from immune system overreaction. Corticosteroids
inhibit nuclear factor (NF)-κB and activator protein (AP)-1, blocking NF−κB dependent
pro-inflammatory gene expression (31, 32), and the transcription of several cytokines
relevant to ARDS pathology. In addition to its anti-inflammatory properties,
methylprednisolone is also known to inhibit proliferation of fibroblasts and synthesis of
collagen by fibroblasts in tissue culture (13, 33).
The initial enthusiasm for the use of corticosteroids to prevent and treat ARDS was
based on animal and human studies (13, 34). Several investigators suggested that the use of

Blue-200302-2560C-R1-Clean version 11
corticosteroids in the late phase of ARDS improved lung function and survival (7-10),
while trials of short term, high dose steroid therapy failed to show an improvement in
mortality in patients at risk of or with early ARDS (5-6). However, the studies on early
ARDS showed some potential limitations: the population studied, which varied
heterogeneously in terms of case mix and patient management, and the use of high doses of
steroids, leading to negative effects due to profound immunosuppression or other side
effects counterbalancing positive effects. Corticosteroid therapy could also be ineffective if
many of the patients, who were considered to have ARDS based on clinical definitions, did
not develop activation of inflammatory cascades in their lungs. Furthermore, the use of
steroid at the late phase of ARDS was based on the assumption that the fibroproliferative
phase began 7-10 days after the onset of the insult. However, different experiments
reported that the proliferative phase begins much sooner than had been previously
appreciated (14-19). Thus, inflammatory and repair mechanisms occur simultaneously
rather than subsequently.
In the present study, a controlled model of ALI induced by two different doses of
paraquat was used, allowing the analysis of different reproducible degrees of severity. In
addition, the ALI model selected was confirmed by light and electronic microscopy, with
great potential to induce fibrogenesis (19, 35). The single low dose of methylprednisolone
(2 mg/kg) administered early in the course of ALI (1 or 6 hours after lung injury induction)
minimized the negative side effects related to immunodepression. This study design could
potentially bypass the drawbacks observed in previous reports (5, 6). It is interesting to note
that histological lung changes observed at 24 hours in mild and severe ALI were already
present as early as 6 hours after ALI induction, although less intense. Thus,

Blue-200302-2560C-R1-Clean version 12
methylprednisolone was used before the beginning of functional changes (1 hour) and after
the establishment of the lung injury process (6 hours).
Our results showed that corticosteroid treatment at the early phase of ALI improved
in vivo and in vitro respiratory mechanics (Figures 1-3). The method used for determination
of in vivo respiratory mechanics allows the identification of its elastic, resistive, and
viscoelastic and/or inhomogeneous components (22, 23). Respiratory system and lung
static elastances, resistive, viscoelastic/inhomogeneous pressures increased significantly
with the severity of lung injury at day 1 (Figures 1 and 2), but only respiratory system and
lung viscoelastic pressures remained higher than control values at day 30 in P10. Prior
studies described changes in lung resistance and elastance in ALI, resulting from surfactant
dysfunction and/or loss of functional capacity due to alveolar flooding (36, 37). Actually,
mechanical dysfunction can result from air-liquid interface and/or tissue changes (38). In
the present work, the increase in lung resistive pressure probably reflects a reduction in
bronchial caliber caused by fluid in the airways, reflex bronchoconstriction, and/or reduced
lung volume. The augment in lung viscoelastic and/or inhomogeneous pressure, suggests
the presence of heterogeneities that can be due to many different factors, e.g., alveoli
collapse and overdistension, distortion of patent alveoli, edema, inflammation with
neutrophils and mononuclear cells infiltration (Table 2, Figure 4), and changes in collagen
and elastic fiber contents (Figure 5). The maintenance of the changes in lung viscoelastic
pressure at day 30 suggests that parenchymal dysfunction dominates the later ALI, since
alveolar inhomogeneities and edema were less remarkable in this period (Figure 4F). Lung
static elastance in P10 and P25 groups was significantly increased compared to control, in
accordance with previous results (39, 40).

Blue-200302-2560C-R1-Clean version 13
Corticosteroid prevented the in vivo mechanical changes in P10 group
independently of the time of injection and of the moment of analysis (1 or 30 days after
ALI induction), while in P25 group corticosteroid prevented the modifications in lung static
elastance and resistive pressure, and attenuated viscoelastic changes (Figures 1 and 2).
These modifications could be attributed to the effect of corticosteroid on inflammatory (41-
43) and fibroproliferative processes (9, 13, 44, 45), leading to less atelectasis, cellular
influx, and fluid transudation in the groups treated with steroid (Figure 4, Table 2). It has
been demonstrated that in experimental acute lung injury corticosteroid treatment is
effective in decreasing lung collagen content and edema formation as long as treatment is
prolonged, whereas steroid withdrawal rapidly reverses this positive effect (13, 44, 45).
However, recent evidence suggests that the inflammatory and fibrotic processes appear to
be separately regulated (46), thus offering the possibility for early-directed treatments
against fibrosis independently of the effects on inflammation. Recently, the beneficial
effects of early low-dose steroid treatment was also observed in patients with septic shock,
where low-dose hydrocortisone treatment inhibited systemic inflammation and prevented
overwhelming compensatory anti-inflammatory response (47).
Parallel to the in vivo analysis, oscillatory tissue mechanics was also addressed. The
importance of alterations in the biophysical properties of the surfactant system in
ARDS/ALI pathophysiology (36, 37), as well as the relevant role of tissue inhomogeneities
secondary to alveoli collapse/hyperdistension are well established. The method used to
determine lung tissue mechanics avoids the influence of kinetics of surface-active molecule
absorption-desorption to the surface film and of recruitment-derecruitment (25, 48, 49).
This method specifically allows the analysis of tissue resistance, elastance, and
hysteresivity after the induction of ALI in the absence of surfactant and interdependence

Blue-200302-2560C-R1-Clean version 14
effects, providing a direct assessment of tissue physiology (38). Additionally, a direct
analysis of the role of fiber-fiber network within the connective tissue matrix on tissue
mechanical properties is ensured (25, 48, 49). The present study is the first analysis of
oscillatory tissue mechanical properties of lung parenchyma in animals with acute lung
injury treated with corticosteroid. Elastance and resistance of lung parenchyma of paraquat-
treated rats were significantly increased in relation to control (Figure 3), supporting
previous findings that parenchymal mechanical dysfunction plays an important role in ALI
pathophysiology (19, 38). Hysteresivity increased only in P25 group.
The changes in tissue mechanics were accompanied by deposition of collagen and
elastic fibers in the alveolar septa (Figure 5). The kinetics of paraquat-induced fibrogenesis
showed a continuous transition among C, P10 at day 1, and P10 at day 30, as previously
reported (50). In addition, the present results disclosed that collagen content was already
elevated 1 day after tissue damage whatever the dose of paraquat used, indicating that the
biochemical processes implicated in collagen synthesis are indeed able to react very quickly
to the aggression. Conversely, several studies (9, 14-16, 18) have reported that elevated
procollagen aminoterminal propeptide levels in the BALF reflect collagen synthesis at the
site of disease, and may be used as markers of the reparative process. Newly synthesized
procollagen is cleaved by specific endopeptidases at the amino and carboxy termini,
forming collagen molecules (51). The amino and carboxy propeptides are soluble proteins,
sampled relatively ease by BAL, and used as good markers of collagen synthesis without
resorting to invasive tissue sampling (52). Thus, these studies (9, 14-16, 18) demonstrate
the presence of increased collagen synthesis, with no indication of matrix deposition, which
would be possible only through direct histological analysis. In this line, our findings

Blue-200302-2560C-R1-Clean version 15
demonstrated that the increased collagen synthesis previously observed (9, 14-16, 18) was
actually followed by matrix deposition.
Collagen types were identified by electron microscopy (Figure 6). Type I collagen
fibrils closely aggregate to form thick fibers of 74 nm, while type III collagen fibrils present
a mean fibrillary diameter of 45 nm (53). Type III collagen appeared early in mild lesion
(P10), while type I collagen appeared late in P10 group (at 30 day) and in the severe lesion
(P25). In this context, Armstrong et al. (52) hypothesized that an imbalance between
synthesis and degradation may contribute to the net accumulation of type I collagen in
ARDS, and that the profibrotic response occurred early in the course of disease, being
associated with the severity of the lung injury and mortality. In our study steroid treatment
was administered before fibroproliferation advanced to end-stage acellular fibrosis, when
the more resistant type I collagen predominates. Methylprednisolone prevented the
increment in tissue elastance and resistance of paraquat-treated rats (Figure 3). The
improvement in tissue mechanics could be explained by the effect of steroid preventing
excessive collagen deposition, increasing collagen breakdown, or inhibiting the aggregation
process of type III collagen fiber into type I (9). Corticosteroid also minimized epithelial
and endothelial cell damage, fibroproliferation, and extracellular matrix deposition (Figure
6). The beneficial effects of steroid could be attributed to its inhibitory effects on host
defense response, including modulation of macrophage and fibroblast activity (7-10, 43,
54).
The present study also disclosed the effects of corticosteroid on elastic system.
Although there are some studies describing collagen changes after corticosteroid treatment
in ALI, the elastic system is scantly dealt with. Mild ALI was followed by a late increase in
elastic fiber content, while the severe lesion presented early elastogenesis. In bleomycin

Blue-200302-2560C-R1-Clean version 16
models of lung fibrosis, elastic fiber content increased at 14 and 30 days (55, 56). However,
in severe paraquat-induced lung injury, the amount of elastic fibers was augmented as early
as 24 hours after ALI induction.
Methylprednisolone modified elastogenesis both in mild and severe ALI (Figure 5).
Recently, we quantified the amount of different types of elastic fibers in the alveolar septa
(elaunin, oxytalan, and fully developed fibers), and observed that oxytalan fibers were
increased in paraquat-induced ALI (19). Oxytalan fibers appear early in the development of
the elastic system, and consist of microfibrils without elastin, rigid under mechanical stress
(28). In our study, the amount of oxytalan fibers in the presence of steroid did not increase;
supporting the hypothesis that steroid could probably diminish elastase activity,
consequently interfering in the resynthesis of the elastic system fiber.
The anatomic elements that potentially determine tissue viscoelastic behavior
include the network of stress-bearing collagen and elastic fibers, proteoglycan and
glycosaminoglycans, as well as the contractile elements present in parenchymal tissues (25,
57). Other authors (58, 59) have suggested that the pulmonary parenchyma can be modeled
as an interconnected network of elastic elements, presumably composed of collagen and
elastic fibers, which determine the mechanical behavior of the system. In our study, tissue
E and R changes were correlated with collagen and elastic fiber contents, while
hysteresivity was correlated only with elastic fibers. Similarly, Yuan et al reported (48, 49)
that collagen and elastic fibers contribute to tissue elasticity during normal breathing.
According to the structural damping hypothesis, induced changes in hysteresivity
with ALI must reflect changes in the kinetics of the stress-bearing process, e.g., the
extracellular fiber matrix, the surface lining layer, and the contractile apparatus (25). In P25
group, a significant modification of collagen-elastin fiber network was present and was

Blue-200302-2560C-R1-Clean version 17
probably the main determinant of the increase in hysteresivity. We hypothesized that the
increment in the amount of elastic fibers, primarily oxytalan fibers, could be responsible for
a derangement of the tridimensional extracellular matrix organization, which would had a
greater impact on tissue mechanics than the effective amount of each fiber component.
These findings are supported by the observation that tissue hysteresivity is not determined
by collagen and elastic fibers alone, being most likely a network effect (49). It is interesting
to observe that in mild ALI the increment in collagen and tissue cellularity was not large
enough to affect hysteresivity.
In conclusion, single low-dose methylprednisolone led to a complete maintenance
of in vivo and in vitro respiratory mechanics in mild ALI, and minimized the changes in
tissue impedance and extra-cellular matrix components in a severe lesion. In addition, the
beneficial effects of the early use of steroids in ALI were also seen at 30 days after lung
injury. We should be extremely cautious in extrapolating these data to the more complex
clinical situation, since this is an animal model of ALI induced by paraquat, and, thus, the
direct extrapolations to the clinical syndrome is unwarranted. Additionally, our animals
were not ventilated or treated with fluid management, thwarting the comparison of the
animal outcome with the human’s. However, the present study has an important clinical
relevance, i.e., the treatment with a single low dose of methylprednisolone at the early
phase of ALI may help to prevent fibroelastogenesis, avoiding the side effects related to
prolonged and high doses of steroid. Finally, this study may help the design of a clinical
trial focused on diminishing clinical lung injury with steroid.

Blue-200302-2560C-R1-Clean version 18
Acknowledgement: We would like to express our gratitude to Mr. Antonio Carlos de
Souza Quaresma and Mrs. Alaine Prudente for their skillful technical assistance, and to Dr.
Ana Paula B Barbosa for her help with the initial analysis of tissue mechanics.

Blue-200302-2560C-R1-Clean version 19
References
1. Modig J, Borg T. High dose methylprednisolone in a porcine model of ARDS induced
by endotoxemia. Acta Chir Scand (Suppl) 1985;526:94-103
2. Pitcairn M, Schuler J, Erve P, Holzman S, Schumer W. Glucosteroid and antibiotic
effect on experimental gram-negative bacteremic shock. Surgery 1975;110:1012-1015.
3. Thomas Jr CS, Brockman SK. The role of adrenal corticosteroid therapy in Escherichia
coli endotoxin shock. Surg Gynecol Obstet 1968;126:61-69.
4. Toung T, Bordos D, Benson D, Carter D, Zuidema GD, Permutt S, Cameron JL.
Aspiration pneumonia: experimental evaluation of albumin and steroid therapy. Ann
Surg 1976;83:179-184.
5. Luce JM, Montgomery AB, Marks JD, Turner J, Metz CA, Murray JF. Ineffectiveness
of high dose methylprednisolone in preventing parenchymal lung injury and improving
mortality in patients with septic shock. Am Rev Respir Dis 1988;138:62-68.
6. Bernard GR, Luce JM, Sprung CL, Rinaldo JE, Tate RM, Sibbald WJ, Kariman K,
Higgins S, Bradley R, Metz CA, Harris TR, Brigham KL. High dose corticosteroids in
patients with the adult respiratory distress syndrome N Engl J Med 1987;31:1565-1570.
7. Meduri G, Chinn A, Leeper K, Wunderink RG, Tolley E, Viner-Muram HT, Khare V,
ElTorky M. Corticosteroid rescue treatment at progressive fibroproliferation in late
ARDS: Patterns of response and predictors of outcome. Chest 1994;105:1516-1527.
8. Meduri G, Headley A, Golden E, Carson SJ, Umberger RA, Kelso T, Tolley EA. Effect
of prolonged methylprednisolone therapy in unresolving acute respiratory distress
syndrome: a randomized controlled trial. JAMA 1998;280:159-165.
9. Meduri GU, Tolley EA, Chinn A, Stentz F, Postlethwaite A. Procollagen types I and III
aminoterminal propeptide levels during acute respiratory distress syndrome and in

Blue-200302-2560C-R1-Clean version 20
response to methylprednisolone treatment. Am J Respir Crit Care Med 1998;158:1432-
1441.
10. Meduri GU, Tolley EA, Chrousos GP, Stentz F. Prolonged methylprednisolone
treatment suppresses systemic inflammation in patients with unresolving acute
respiratory distress syndrome. Am J Respir Crit Care Med 2002;165:983-991.
11. Pitet J-F, Mackersie RC, Martin TR, Matthay MA. Biological markers of acute lung
injury: prognostic and pathogenetic significance. Am J Respir Crit Care Med
1997;155:1187-1205.
12. Martin C, Papazian L, Paya M-J, Saux P, Gouin F. Pulmonary fibrosis correlates with
outcome in adult respiratory distress syndrome. A study in mechanically ventilated
patients. Chest 1995;107:196-200.
13. Hesterberg TW, Last JÁ. Ozone-induced acute pulmonary fibrosis in rats- Prevention of
increased rates of collagen synthesis by methylprednisolone. Am Rev Respir Dis
1981;123:47-52.
14. Liebler JM, Qu Z, Buckner B, Powers MR, Rosenbaum JT. Fibroproliferation and mast
cells in the acute respiratory distress syndrome. Thorax 1998;53:823-829.
15. Pugin, J, Verghese G, Widmer MC, Matthay MA. The alveolar space is the site of
intense inflammatory and profibrotic reactions in the early phase of acute respiratory
distress syndrome. Crit Care Med 1999;27:304-312.
16. Chesnutt AN, Matthay MA, Tibayan FA, Clark JG. Early detection of type III
procollagen peptide in acute lung injury. Pathogenetic and Prognostic Significance. Am
J Respir Crit Care Med 1997;156:840-845.
17. Clark JG, Milberg JA, Steinberg KP, Hudson LD. Type III procollagen peptide in the
adult respiratory distress syndrome. Ann Intern Med 1995;122:17-23.

Blue-200302-2560C-R1-Clean version 21
18. Marshall RP, Bellingan G, Webb, S, Puddicombe, A, Goldsack N, McAnulty RJ,
Laurent GJ. Fibroproliferation occurs early in the acute respiratory distress syndrome
and impacts on outcome. Am J Respir Crit Care Med 2000;162:1783-1788.
19. Rocco PRM, Negri EM, Kurtz PM, Vasconcellos FP, Silva GH, Capelozzi VL, Zin,
WA Lung tissue mechanics and extracellular matrix in acute lung injury. Am J Respir
Crit Care Med 2001;164:1067-1071.
20. Rocco PRM, Lima JGM, Barros AS, Contador R, Negri E, Capelozzi VL, Mourão
PAS, Faffe DS, Zin WA Effects of corticosteroid on lung parenchyma remodeling at an
early phase of acute lung injury [abstract]. Am J Respir Crit Care Med 2001;164:A451.
21. Negri EM, Souza AB, Pássaro CP, Santos FB, Capelozzi VL, Zin WA, Rocco PRM.
Early corticosteroid treatment in ARDS: late effects on respiratory mechanics and tissue
remodelling [abstract]. Eur Respir J 2002;20:A36.
22. Bates JHT, Rossi A, Milic-Emili J. Analysis of the behavior of the respiratory system
with constant inspiratory flow. J Appl Physiol 1985;58:1840-1848.
23. Bates JHT, Ludwig MS, Sly PD, Brown KA, Martin JG, Fredberg JJ. Interrupter
resistance elucidated by alveolar pressure measurements in open-chest normal dogs. J
Appl Physiol 1988;65:408-414.
24. Faffe DS, Silva GH, Kurtz PMP, Negri EM, Capelozzi VL, Rocco PRM, Zin WA.
Lung tissue mechanics and extracellular matrix composition in a murine model of
silicosis. J Appl Physiol 2001;90:1400-1406.
25. Fredberg JJ, Stamenovic D. On the imperfect elasticity of lung tissue. J Appl Physiol
1989;67:2408-2414.

Blue-200302-2560C-R1-Clean version 22
26. Nagase T, Lei M, Robatto FM, Eidelman DH, Ludwig MS. Tissue viscance during
induced constriction in rabbit lung: morphological-physiological correlation. J Appl
Physiol 1992;73:1900-1907.
27. Weibel ER. Morphometry: stereological theory and practical methods. In: Gil J, editor.
Models of Lung Disease-Microscopy and Structural Methods. New York: Marcel
Dekker; 1990, p. 199-247.
28. Montes GS. Structural biology of the fibers of the collagenous and elastic system. Cell.
Biology International 1996;20:15-27.
29. Weigert C. Über eine methode zur farbung elastischer fasern. Zentralbl. Allg. Pathol.
Anat. 1898;9:289-292.
30. Brigham KL, Bowers RE, McKeen CR. Methylprednisolone prevention of increased
lung vascular permeability following endotoxemia in sheep. J Clin Invest
1981;67:1103-1110.
31. Wissink S, van Heerde EC, van der Burg B, et al. A dual mechanism mediates
repression of NF-kappa B activity by glucocorticoids. Mol Endocrinol 1998;12:355-
363.
32. Jantz MA, Sahn AS. Corticosteroids in acute respiratory failure. Am J Respir Crit Care
Med 1999;160:1079-1100.
33. Thompson BT. Glucocorticoids and acute lung injury. Crit Care Med 2003;31:S253-
S257.
34. Sibbald WJ, Anderson RR, Reid B, Holliday RL, Driedger AA. Alveolocapillary
permeability in human septic ARDS. Chest 1981;79:133-142
35. Smith P, Health D, Kay JM. The pathogenesis and structure of paraquat-induced lung
fibrosis in rats. J Pathol 1974;114:57-67.

Blue-200302-2560C-R1-Clean version 23
36. Grossman RF, Jones JG, Murray JF. Effects of oleic acid-induced pulmonary edema on
lung mechanics. J Appl Physiol 1980;48:1045-1051.
37. Gregory TJ, Longmore WJ, Moxley MA, Whitsett JA, Reed CR, Fowler AA, Hudson
LD, Maunder RJ, Crim C, Hyers TM. Surfactant chemical composition and biophysical
activity in acute respiratory distress syndrome. J Clin Invest 1991;88:1976-1981.
38. Ingenito EP, Mark L, Davison B. Effects of acute lung injury on dynamic tissue
properties. J Appl Physiol 1994;77:2689-2697.
39. Eissa NT, Ranieri VM, Corbeil C, Chassé M, Robatto FM, Braidy J, Milic-Emili J.
Analysis of behavior of the respiratory system in ARDS patients: effects of flow,
volume, and time. J Appl Physiol 1991;70:2719-2729.
40. Barnas GM, Stamenovic D, Lutchen KR. Lung and chest wall impedances in the dog in
normal range of breathing: effects of pulmonary edema. J Appl Physiol 1992;73:1040-
1046.
41. Auphan N., Didonato JA, Rosette C, Helmberg A, Karin M. Immunosuppression by
glicocorticoids: inhibition of NF-kappa B activity through induction of Ikappa B
synthesis. Science 1995;270:286-290.
42. Almawi WY, Beyhum HN, Rahme AA, Rieder MJ. Regulation of cytokine and
cytokine receptor expression by glucocorticoids. J Leukoc Biol 1996;60:563-572.
43. Meduri GU, Headley S, Tolley E, Shelby M, Stentz F, Postlethwaite A.
Plasma and BAL cytokine response to corticosteroid rescue treatment in late ARDS.
Chest 1995;108:1315-1325.
44. Hakkinen PJ, Schmoyer RL, Witschi HP. Potentiation of butylated-hydroxytoluene-
induced acute lung damage by oxygen. Effects of prednisolone and indomethacin. Am
Rev Respir Dis 1983;128:648-651.

Blue-200302-2560C-R1-Clean version 24
45. Kehrer JP, Kleine-Szanto AJP, Sorensen EMB, Pearlman R, Rosner MH. Enhanced
acute lung damage following corticosteroid treatment. Am Rev Respir Dis
1984;130:256-261.
46. Kaminski N, Allard JD, Pittet JF, Zuo F, Griffiths MJ, Morris D, Huang X, Sheppard D,
Heller RA. Global analysis of gene expression in pulmonary fibrosis reveals distinct
programs regulating lung inflammation and fibrosis. Proc Natl Acad Sci USA
2000;97:1778-1783.
47. Keh D, Boehnke T, Weber-Cartens S, Schultz C, Ahlers O, Bercker S, Volk H, Doecke
W, Falke KJ, Gerlach H. Immunologic and hemodynamic effects of “low-dose”
hydrocortisone in septic shock. Am J Respir Crit Care Med 2003;167:512-520.
48. Yuan H, Ingenito EP, Suki B. Dynamic properties of lung parenchyma: mechanical
contributions of fiber network and interstitial cells. J Appl Physiol 1997;83:1420-1431.
49. Yuan H, Kononov S, Cavalcante FSA, Lutchen KR, Ingenito EP, Suki B. Effects of
collagenase and elastase on the mechanical properties of lung tissue strips. J Appl
Physiol 2000;89:3-14.
50. Shinozaki S, Tobayashi T, Kubo K, Sekiguchi M. Pulmonary hemodynamics and lung
function during chronic paraquat poisoning in sheep. Am Rev Respir Dis 1992;146:775-
780.
51. Kessler E, Goldberg B. A method for assaying the activity of the endopeptidases which
excises the nonhelical carboxyterminal extensions from type I procollagen. Anal
Biochem 1978;86:463-469.
52. Armstrong L, Thickett DR, Mansell JP, Ionescu M, Hoyle E, Billinghurst RC, Poole
AP, Millar AB. Changes in collagen turnover in early acute respiratory distress
syndrome. Am J Respir Crit Care Med 1999;160:1910-1915.

Blue-200302-2560C-R1-Clean version 25
53. Junqueira LCU, Montes GS. Biology of collagen-proteoglycan interaction. Arch Histol
Jpn 1983;46:589-629.
54. Wissink S, van Heerde EC, van der Burg B, van der Saag PT.
A dual mechanism mediates repression of NF-kappa B activity by glucocorticoids. Mol
Endocrinol 1998;12:355-363
55. Ebihara T, Venkatesan N, Tanaka R, Ludwig M. Changes in extracellular matrix and
tissue viscoelasticity in bleomycin-induced lung fibrosis. Temporal aspects. Am J
Respir Crit Care Med 2000;162:1569-1576.
56. Goldstein RH, Lucey EC, Frazblau C, Snider GL. Failure of mechanical properties to
parallel changes in lung connective tissue composition in bleomycin-induced
pulmonary fibrosis in hamsters. Am Rev Respir Dis 1979;120:67-73.
57. Mijailovich SM, Stamenovic D, Fredberg JJ. Toward a kinetic theory of connective
tissue micromechanics. J Appl Physiol 1993;74:665-681.
58. Stamenovic D. Micromechanical foundations of pulmonary elasticity. Physiol Rev
1990;70:1117-1134.
59. Stamenovic D, Wilson T. Parenchymal stability. J Appl Physiol 1992;73:596-602.

Blue-200302-2560C-R1-Clean version 26
Legends:
Figure 1. Static elastance (Est) of respiratory system (rs), lung (L), and chest wall (w). In
control group (C) saline (5 ml/kg) was intraperitoneally (i.p.) injected. In P groups,
different doses of paraquat (10 and 25 mg/kg) were injected i.p. 1 and 30 days prior to the
measurements. M1 and M6 correspond to the groups that received methylprednisolone (2
mg/kg) 1 and 6 hours, respectively, after the induction of acute lung injury with different
doses of paraquat. Values are means (+SEM) of 6 animals. *Values significantly different
from C (p<0.05); **values significantly different from P at the same dose and time
(p<0.05); #values significantly different from P10 at day 1 (p<0.05).
Figure 2. Stacked bar chart plot data where the gray bars represent the resistive pressures
(∆P1) and the white bars are the viscoelastic/inhomogeneous (∆P2) pressure dissipations of
the respiratory system (rs), lung (L), and chest wall (w). The whole column represents the
total pressure variation in each group. In control group (C) saline (5 ml/kg) was
intraperitoneally (i.p.) injected. In P groups, different doses of paraquat (10 and 25 mg/kg)
were injected i.p. 1 and 30 days prior to the measurements. M1 and M6 correspond to the
groups that received methylprednisolone (2 mg/kg) 1 and 6 hours, respectively, after the
induction of acute lung injury with different doses of paraquat. Values are means (+SEM)
of 6 animals. *Values significantly different from C (p<0.05); **values significantly
different from P at the same dose and time (p<0.05); #values significantly different from
P10 at 1 day (p<0.05).

Blue-200302-2560C-R1-Clean version 27
Figure 3. Tissue elastance, resistance, and hysteresivity at 1 Hz. In control group (C) saline
(5 ml/kg) was intraperitoneally (i.p.) injected. In P groups, different doses of paraquat (10
and 25 mg/kg) were injected i.p. 1 and 30 days prior to the measurements. M1 and M6
correspond to the groups that received methylprednisolone (2 mg/kg) 1 and 6 hours,
respectively, after the induction of acute lung injury with different doses of paraquat.
Values are means (SEM) of 6 animals.*Values significantly different from C (p<0.05);
**values significantly different from P at the same dose and time (p<0.05); #values
significantly different from P10 at 1 day (p<0.05).
Figure 4. Photomicrographs of lung parenchyma stained with hematoxylin-eosin in control
(A) and paraquat treated lungs [P10 (B) and P25 (D)] at day 1, and P10 at day 30 (F).
Panels C, E, and G correspond to paraquat groups (as in B, D, and F, respectively) treated
with methylprednisolone 6 hours after ALI induction. Scale bars=200 µm.
Figure 5. Amounts of collagen and elastic system fibers in alveolar septa. In control group
(C) saline (5 ml/kg) was intraperitoneally (i.p.) injected. In P groups, different doses of
paraquat (10 and 25 mg/kg) were injected i.p. 1 and 30 days prior to the measurements. M1
and M6 represent the groups that received methylprednisolone (2 mg/kg) 1 and 6 hours,
respectively, after the induction of acute lung injury with different doses of paraquat.
Values are means (+SEM) of 6 animals.*Values significantly different from C (p<0.05);
**values significantly different from P at the same dose and time (p<0.05); #values
significantly different from P10 at 1 day (p<0.05).

Blue-200302-2560C-R1-Clean version 28
Figure 6. Electron microscopy of lung parenchyma in control (A) and paraquat treated
lung at day 1 [P10 (B) and P25 (D)] and 30 (F). In P10 and P25 groups, paraquat (10 and
25 mg/kg, respectively) was injected intraperitoneally. Panels C, E, and G correspond to
paraquat groups treated with methylprednisolone (2 mg/kg, i.v.) 6 hours after the induction
of ALI. Normal ultrastructure in untreated lung (A). Note the type II pneumocyte (PII) with
lamellar bodies (lb) (Panel A). In P10 (Panel B) there were apoptotic changes in PII
characterized by condensation of chromatin (arrows). Type III collagen content (stars) was
augmented and incipient type I collagen fiber (col I) synthesis is also evident. A randomly
orientated, interconnected network of elastic fibers is depicted (arrowheads). PMC:
primitive mesenchymal cell. In panel D, types I and III collagen syntheses are evident in
P25. Arrowheads indicate elastin distributed throughout the interstitium, stars represent
type III collagen fibers. Mφ: macrophage. In P10 group 30 days after ALI induction
fibroelastosis was present (Panel F). In panels C and E, hyaline membranes (HM) are still
evident after methylprednisolone treatment in both P10 and P25 groups, respectively, as
well as edema of the basement membrane (BM) (Panel C). Panels C, E, and G show that
steroid treatment modulates the increment in types I and III collagen fibers (stars) and
elastogenesis. Photomicrographs are representative of data obtained from lung sections
derived from six animals.

Blue-200302-2560C-R1-Clean version 29
TABLE 1. EFFECT OF STEROID ON THE SURVIVAL OF PARAQUAT-
CHALLENGED RATS
Groups Number of surviving rats Survival (%)
DAY 1
CONTROL 6(6) 100
P10 6(7) 85
P10M1 6(6) 100
P10M6 6(6) 100
P25 6(12) 50
P25M1 6(10) 60
P25M6 6(12) 50
DAY 30
P10 6(9) 67
P10M1 6(6) 100
P10M6 6(6) 100
Data represent the number of surviving rats, with the total number of rats in parentheses. In
control group, saline was intraperitoneally (i.p.) injected. In P groups, different doses of
paraquat (10 and 25 mg/kg) were injected i.p. 1 and 30 days prior to the measurements. M1
and M6 correspond to the groups that received methylprednisolone (2 mg/kg) 1 and 6
hours, respectively, after the induction of acute lung injury with different doses of paraquat.
Rats from P25 group independent of the use of steroid died within 48 hours after ALI
induction
Blue-200302-2560C-R1-Clean version 30
TABLE 2. MORPHOMETRICAL PARAMETERS
Groups Normal (%) Alveolar Collapse (%)
Total cell (%) PMN (%) MN (%)
DAY 1
CONTROL 97.9±0.8 2.1±0.8 18.2±1.1 3.7±0.7 14.5±0.9
P10 85.9±0.7* 13.9±0.7* 25.6±1.5* 23.3±2.5* 2.3±0.4*
P10M1 91.8±0.3* ** 8.2±0.3* ** 17.7±1.2** 16.2±1.4* ** 1.6±0.2*
P10M6 91.1±0.8* ** 8.9±0.8* ** 16.4±2.1** 14.8±2.1* ** 1.6±0.3*
P25 74.6±1.9*# 25.4±3.4*# 32.8±1.8*# 32.2±1.8*# 0.7±0.1*#
P10M1 84.0±2.0* ** 16.7±1.9* ** 22.8±1.4** 21.0±1.5* ** 1.8±0.3* **
P10M6 77.2±0.8* ** 22.8±0.8* ** 24.8±1.8** 22.5±1.7* ** 2.3±0.3* **
DAY 30
P10 78.6±1.9*# 21.4±1.9*# 26.3±0.6* 20.4±1.1* 5.9±0.6*#
P10M1 94.3±0.8** 5.8±0.8** 20.1±0.3** 15.5±0.7* ** 4.6±0.5*
P10M6 93.3±2.0** 6.9±2.0** 21.7±1.0** 15.6±0.9* ** 6.1±0.3*
Values are means ± SEM of 6 lungs (rats) in each group (10 random, non-coincident
microscopic fields were analyzed in each lung). In control group, saline was
intraperitoneally (i.p.) injected. In P groups, different doses of paraquat (10 and 25 mg/kg)
were injected i.p., 1 and 30 days prior to the measurements. M1 and M6 correspond to the
groups that received methylprednisolone (2 mg/kg) 1 and 6 hours, respectively, after the
induction of acute lung injury with different doses of paraquat. Total cell: total cellular
fractional area, PMN: polymorphonuclear cell fractional area, MN: mononuclear cell
fractional area. *Values significantly different from C (p<0.05); **values significantly
different from P at the same dose and time (p<0.05); #values significantly different from
P10 at day 1 (p<0.05).
Blue-200302-2560C- R1 31
Est
(cm
H2O
/ml)
0
2
4
6
8
10CPM1M6*
*#
#
rsE
st (
cmH
2O/m
l)
0
2
4
6
8
10
*
*#
#
L
Est
(cm
H2O
/ml)
0
1
2
3
4
5
*# * *
w
P25 P10
30 days
C P10
1 day
****
** **
** **
** **
fling
Figure 1

Blue-200302-2560C- R1 32
Figure 2.
∆∆ ∆∆P (
cmH
2O)
0
1
2
3
4
5
6
7
8 ∆∆∆∆P2∆∆∆∆P1
rs∆∆ ∆∆P
(cm
H2O
)
0
1
2
3
4
5
6
∆∆ ∆∆P (
cmH
2O)
0
1
2
3 w
L
C
**
*
** **** **
** **
*
*
***
#
#
#
*** ****** ***
** ** #
*
*
*
** **** **
** **
*#
*#
*#
***
***
***
***
** **
*#
*
#
** **** **
*#
*#
*#
*#
*#
*#
P10 P25 P10
1 day 30 days
P M1M6 P M1M6 P M1 M6
Blue-200302-2560C- R1 33
Figure 3.
C P10 P25 P10
1 day 30 days
Ela
stan
ce (
104 N
/m2 )
0.0
0.5
1.0
1.5
2.0
2.5
3.0 CPM1M6
Res
ista
nce
(10
2 N
.s/m
2 )
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Hys
tere
sivi
ty
0.00
0.02
0.04
0.06
0.08
0.10
*
*
*#
***
***
*#
***
***
*#
#
** **** **
**
** **** **

Blue-200302-2560C- R1 34
Figure 4.
B
ED
G
C
F
A

Blue-200302-2560C- R1 35
Figure 5.
C P10 P25 P10
1 day 30 days
Co
llag
en f
iber
s/se
ptu
m( µµ µµ
m2 / µµ µµ
m)
0.0
0.1
0.2
0.3
0.4
0.5CPM1M6
*
***
***
*
***
***
Ela
stic
fib
ers/
sep
tum
( µµ µµm
2 / µµ µµm
)
0.0
0.2
0.4
0.6
0.8
*
# *#
# *#
*** *
**
** **** **

Blue-200302-2560C- R1 36
Figure 6.
*
B C
*
Alb
*
**

EFFECT OF CORTICOSTEROID ON LUNG PARENCHYMA REMODELING
AT AN EARLY PHASE OF ACUTE LUNG INJURY
PATRICIA R.M. ROCCO1, ALBA B. SOUZA1, DEBORA S. FAFFE1, CAROLINE P.
PÁSSARO1, FLÁVIA B. SANTOS1, ELNARA M. NEGRI2, JANUÁRIO G.M. LIMA1,
RENATA S. CONTADOR1, VERA L. CAPELOZZI3, WALTER A. ZIN1
ONLINE DATA SUPPLEMENT

Blue-200302-2560C- R1 - Online-only repository
1
METHODS
Animal preparation
Seventy-eight Wistar rats (250-300 g) were randomly divided into 13 groups of 6 animals each.
In the control group (C), saline (0.9% NaCl, 5 ml/kg BW) was injected intraperitoneally (i.p.). In
P groups different doses of paraquat were injected i.p. (10 and 25 mg/kg BW), 1 or 30 days prior
to the measurements. Each group (P10 or P25) was divided into 3 subgroups: animals that
received solely paraquat or were additionally treated with methylprednisolone (M, 2 mg/kg, i.v.)
at 1 or 6 hours after paraquat administration. The animals were sedated with diazepam (5 mg
i.p.), anesthetized with pentobarbital sodium (20 mg/kg i.p.), and a snugly fitting cannula (1.7
mm ID) was introduced into the trachea. Mechanical ventilation (model 683, Harvard Apparatus,
Southnatick, MA, USA) was then started with a frequency of 80 breaths.min-1 and a tidal volume
of 6 ml/kg.
A pneumotachograph (1.5 mm ID, length = 4.2 cm, distance between side ports = 2.1 cm)
was connected to the tracheal cannula for the measurements of airflow (V’) and changes in lung
volume (VT) (1). The pressure gradient across the pneumotachograph was determined by means of a
Validyne MP45-2 differential pressure transducer (Engineering Corp, Northridge, CA, USA). The
flow resistance of the equipment (Req), tracheal cannula included, was constant up to flow rates of
26 mL.s-1, and amounted to 0.12 cmH2O.mL-1.s. Equipment resistive pressure (= Req/V’) was
subtracted from pulmonary resistive pressure so that the present results represent intrinsic values.
Tracheal pressure was measured with a Validyne MP-45 differential pressure transducer
(Engineering Corp, Northridge, CA, USA). Changes in esophageal pressure (Pes), which reflects
chest wall pressure (Pw), were measured with a 30-cm-long water-filled catheter (PE-240) with side
holes at the tip connected to a PR23-2D-300 Statham differential pressure transducer (Hato Rey,
Puerto Rico). The catheter was passed into the stomach and then slowly returned into the

Blue-200302-2560C- R1 - Online-only repository
2
esophagus; its proper positioning was assessed using the "occlusion test" (2). The frequency
responses of the pressure measurement systems (Ptr and Pes) were flat up to 20 Hz, without
appreciable phase shift between the signals. All signals were conditioned and amplified in a
Beckman type R Dynograph (Beckman Instruments, Schiller Park, IL, USA). Flow and pressure
signals were also passed through 8-pole Bessel filters (902LPF, Frequency Devices, Haverhill, MA,
USA) with the corner frequency set at 100 Hz, sampled at 200 Hz with a 12-bit analogue-to-digital
converter (DT2801A, Data Translation, Marlboro, MA, USA), and stored on a PC-compatible
computer. All data were collected using LABDAT software (RHT-InfoData Inc., Montreal, Quebec,
Canada).
Measurement of respiratory mechanics
Muscle relaxation was achieved with gallamine triethyliodide (2 mg/kg i.v.) and artificial
ventilation was provided by a constant flow ventilator (Samay VR15, Universidad de la
Republica, Montevideo, Uruguay). During the test breaths the ventilator was adjusted to generate
a 5-s end-inspiratory pause, whereas during baseline ventilation no pause was used. Special care
was taken to keep tidal volume (VT = 1 ml) and flow (V’= 7 ml/s) constant in all animals in order
to avoid the effects of different flows and volumes (3) and inspiratory duration (4) on the
measured variables.
The experiments did not last more than 40 minutes. Respiratory mechanics were measured by
the end-inflation occlusion method (5, 6). Briefly, after end-inspiratory occlusion, there is an initial
fast drop in tracheal pressure (∆P1,rs) from the preocclusion value down to an inflection point
(Pi,rs) followed by a slow pressure decay (∆P2,rs), until a plateau is reached. This plateau
corresponds to the elastic recoil pressure of the respiratory system (Pel,rs). ∆P1,rs selectively
reflects the combination of airways, pulmonary, and chest wall Newtonian resistances in normal

Blue-200302-2560C- R1 - Online-only repository
3
animals and humans (5, 6), and ∆P2,rs reflects stress relaxation, or viscoelastic properties, of the
lung and chest wall tissues, together with a small contribution of pendelluft (6, 7). The same
procedures apply to the chest wall pressure (Pw) yielding the values of ∆P1,w, Pi,w, ∆P2,w, and
Pel,w, respectively. Transpulmonary pressures (∆P1,L, Pi,L, ∆P2,L, and Pel,L) were calculated by
subtracting the chest wall from the corresponding values pertaining to the respiratory system. Total
pressure drop (∆Ptot) is equal to the sum of ∆P1 and ∆P2 yielding the values of ∆Ptot,rs, ∆Ptot,L,
and ∆Ptot,w. Respiratory system, lung, and chest wall static elastances (Est,rs, Est,L, and Est,w,
respectively) were calculated by dividing Pel,rs, Pel,L, and Pel,w, respectively, by VT. Dynamic
elastances of the respiratory system, lung, and chest wall (Edyn,rs, Edyn,L, and Edyn,w,
respectively) were obtained by dividing Pi,rs, Pi,L, and Pi,w, respectively, by VT. ∆E was calculated
as the difference Edyn - Est, yielding the values of ∆E,rs, ∆E,L, and ∆E,w. Respiratory mechanics
were measured 10 times in each animal.
The delay between the beginning and the end of the valve closure (10 ms) was allowed for by
back-extrapolation of the pressure records to the actual time of occlusion and the corrections in
pressure, although very minute, were performed as previously described (8).
All data were analyzed using ANADAT data analysis software (RHT-InfoData Inc.,
Montreal, Quebec, Canada).
Measurement of tissue mechanics
Tissue preparation
Heparine (1000 IU) was intravenously injected immediately after the determination of
respiratory mechanics. The trachea was clamped ten minutes later at end-expiration, and the
abdominal aorta and vena cava were sectioned, yielding a massive hemorrhage that quickly
killed the animals. The lungs were removed en bloc, and placed in a modified Krebs-Henseleith

Blue-200302-2560C- R1 - Online-only repository
4
(K-H) solution containing (mM) 118.4 NaCl, 4.7 KCl, 1.2 K3PO4, 25 NaHCO3, 2.5 CaCl2.H2O,
0.6 MgSO4.H2O, and 11.1 glucose; at pH = 7.40 and 6°C. A 3x3x10 mm strip of subpleural
parenchyma was cut from the periphery of each left lung and suspended vertically in an organ
bath filled with K-H solution maintained at 37°C, continuously bubbled with a mixture of 95%
O2-5% CO2.
Lung strips were weighed (W), and their unloaded resting lengths (L0) were determined
with a caliper. Lung strip volume was measured by simple densitometry, as: Vol = ∆F/δ, where
∆F is the total change in force before and after strip immersion in K-H solution and δ is the mass
density of K-H solution (9-12).
Apparatus
Parenchymal strips were suspended vertically in a K-H organ bath maintained at 37ºC and
continuously bubbled with of 95% O2-5% CO2. Metal clips made of 0.5 mm-thick music wire
were glued to both ends of the tissue strip with cyanoacrylate. One clip was attached to a force
transducer (FT03, Grass Instruments Co., Quincy, MA, USA), whereas the other one was
fastened to a vertical rod. This fiberglass stick was connected to the cone of a woofer, which was
driven by the amplified sinusoidal signal of a waveform generator (3312A Function Generator,
Hewlett Packard, Beaverton, OR, USA). A sidearm of the rod was linked to a second force
transducer (FT03, Grass Instruments Co., Quincy, MA, USA) by means of a silver spring of
known Young’s modulus, thus allowing the measurement of displacement. Length and force
output signals were conditioned (Gould 5900 Signal Conditioner Frame, Gould Inc., Valley
View, OH, USA), fed through 8-pole Bessel filters (902LPF Frequency Devices, Haverhill, MA,
USA), analogue-to-digital converted (DT2801A, Data Translation Inc., Marlboro, MA, USA),
and stored on a computer. All data were collected using LABDAT software (RHT-InfoData Inc.,

Blue-200302-2560C- R1 - Online-only repository
5
Montreal, Quebec, Canada). The frequency response of the system was dynamically studied by
using calibrated silver springs with different elastic Young’s modulus. Neither amplitude
dependence (<0.1% change in stiffness) nor phase changes with frequency were detected in the
range from 0.01 to 14 Hz. The hysteresivity of the system was independent of frequency and had
a value <0.003 (11, 12).
Preconditioning
Cross-sectional, unstressed area (A0) of the strip was determined from volume and unstressed
length, according to A0 = vol/L0. Basal force (FB) for a stress of 0.1 N/cm2 was calculated as FB
(N) = 0.1 (N/cm2).A0 (cm2) and adjusted by vertical displacement of the force transducer (11-
14). The displacement signal was then set to zero. Once basal force and displacement signals
were adjusted, the length between bindings (LB) was measured by means of a precision caliper.
Instantaneous length during oscillation around LB was determined by adding the value of LB to
the measured value of displacement at any time.
After the basal force was adjusted to 8x10-3 N, each parenchymal strip was
preconditioned by sinusoidal oscillation of the tissue during 30 min (frequency = 1 Hz;
amplitude large enough to reach a final force of 1x10-2 N). Thereafter the amplitude was adjusted
to 5% L0 and the oscillation maintained for another 30 min, or until a stable length-force loop
was reached (12). The isometric stress adaptation period resulted in a final force of 5x10-3 N.
After preconditioning, the strips were oscillated at a frequency (f) = 1 Hz (11, 12). The bath
solution was renewed regularly (every 20 min) with 37° C K-H solution.
Measurements of parenchymal mechanics
To calculate tissue resistance (R), elastance (E), and hysteresivity (η) force-length curves were
analyzed (10-12). Instantaneous average cross sectional area (Ai) was determined as Ai = Vol/Li

Blue-200302-2560C- R1 - Online-only repository
6
(cm2), where Li is instantaneous length. Instantaneous stress (σi) was calculated by dividing
force (g) by Ai (cm2), σi= F/Ai.
All mechanical parameters were measured cycle-by-cycle. Tissue resistance (R) was
determined from the enclosed area of force-length loops:
R = (4⋅H)/ [ π⋅ω⋅(∆ε)2]
where H is the stress-strain hysteresis area, ω is the angular frequency (ω = 2πf, rad/s), and ∆ε is
the normalized strain or peak-to-peak change in length (∆L) divided by LB. Tissue dynamic
elastance was determined as:
E = (∆σi/∆ε) cosθ
where ∆σi represents the peak-to-peak change in force, and θ is the phase lag between force and
displacement [θ = sin-1 (4⋅H/(π⋅∆σi⋅∆ε))]. Hysteresivity, defined by Fredberg and Stamenovic
(15) as a dimensionless variable coupling the dissipative and elastic behaviors was calculated by
using the following equation: η = tan θ.
Histology and morphometry
Lung
Morphometric analysis was performed in excised lungs at end-expiration. Immediately after the
removal of the lungs en bloc, the right lung was quick-frozen by immersion in liquid nitrogen
and fixed with Carnoy's solution (ethanol:cloroform:acetic acid, 70:20:10 by volume) at −70°C
for 24 hours. Progressively increasing concentrations of ethanol at -20°C were then substituted
for Carnoy's solution until 100% ethanol was reached. The tissue was maintained at -20°C for 4
hours, warmed to 4°C for 12 hours, and then allowed to reach and remain at room temperature
for 2 hours (16). After fixation, the tissue was embedded in paraffin. Blocks were cut 4−µm-
thick by a microtome. The slices were stained with hematoxylin-eosin. Two investigators, who

Blue-200302-2560C- R1 - Online-only repository
7
were unaware of the origin of the material, performed the microscopic examination. Morphometric
analysis was performed with an integrating eyepiece with a coherent system made of a 100-point
grid consisting of 50 lines of known length, coupled to a conventional light microscope
(Axioplan, Zeiss, Oberkochen, Germany). The volume fraction of collapsed and normal
pulmonary areas and the fraction of the lung occupied by large-volume gas-exchanging air
spaces (hyperinflated structures with a morphology distinct from that of alveoli and wider than
120 µm) were determined by the point-counting technique (17), made at a magnification of x40
across 10 random, noncoincident microscopic fields.
Polymorpho- and mononuclear cells, and pulmonary tissue were determined in each
sample by the point-counting technique (17) across ten random noncoincident microscopic fields
at x1000 magnification. Points falling on tissue area and not over alveolar spaces were counted
and divided by the total number of points in each microscopic field. Thus, data were reported as
the fractional area of pulmonary tissue (17). The same method was applied to determine
polymorpho- and mononuclear cells.
Parenchymal strips
At the end of the experiments the organ bath was removed and the parenchymal strips were
frozen by rapid immersion in liquid nitrogen at the force maintained during the experiment.
Frozen strips were fixed as aforementioned.
The slices underwent specific staining methods to characterize the fibers of the collagenous
and elastic systems present in the alveolar septa. Collagen: The tissue was stained in a solution
of Sirius Red dissolved in aqueous saturated picric acid, and observed under polarized light
microscopy, as the enhancement of collagen birefringence promoted by the picrosirius-
polarization method is specific for collagenous structures (18). Elastic fibers: Two different

Blue-200302-2560C- R1 - Online-only repository
8
methods were used: Weigert's Resorcin Fuchsin method (RF) (19), which allows the
identification of elaunin and fully developed elastic fibers, and Weigert's Resorcin Fuchsin
method modified with oxidation (ORF) (20), which allows the identification of the three
components of the elastic fiber system (elaunin, oxytalan, and fully developed elastic fibers). The
oxytalan fibers were calculated by subtracting the amount of fibers given by the RF method from
the value provided by the ORF method. In each rat twenty different microscopic fields were
randomly selected in order to quantify collagen or elastic fibers. Quantification was carried out
with the aid of a digital analysis system and specific software (Bioscan-Optimas 5:1, Bioscan
Incorporated, Edmond, WA, USA) under x200 magnification. The images were generated by a
microscope (Axioplan, Zeiss, Oberkochen, Germany) connected to a camera (Sony Trinitron
CCD, Sony, Tokyo, Japan), fed into a computer through a frame grabber (Oculus TCX, Coreco
Inc., St Laurent, PQ, Canada) for off-line processing. The thresholds for fibers of the collagenous
and elastic systems were established after enhancing the contrast up to a point at which the fibers
were easily identified as either black (elastic) or birefringent (collagen) bands. The area occupied
by fibers was determined by digital densitometric recognition. Bronchi and blood vessels were
carefully avoided during the measurements. To avoid any bias due to septal edema or alveolar
collapse the areas occupied by the elastic and collagen fibers, measured in each alveolar septum,
were divided by the length of each studied septum. The results were expressed as the amount of
elastic and collagen fibers per unit of septal length.
Transmission electron microscopy
To obtain a stratified random sample, three slices of 2x2 mm were cut from three different
segments of the right lung (upper, middle, and lower lobes), and then fixed with glutaraldehyde
2.5% and phosphate buffer 0.1M (pH = 7.4) for 60 min at –4°C. The slices were then rinsed in

Blue-200302-2560C- R1 - Online-only repository
9
phosphate buffer, post-fixed with 1% osmic tetroxide in phosphate buffer for 30 min and
rewashed three times in phosphate buffer. Finally, the slices were dehydrated in an acetone
series, and then placed in a mixture of 1:1 acetone:Epon overnight before embedding in Epon for
6 hours. After fixation the material was kept for 48 hours at 60°C before being submitted to
ultramicrotomy for transmission electron microscopy.
Statistical analysis
The normality of the data (Kolmogorov-Smirnov test with Lilliefor’s correction) and the
homogeneity of variances (Levene median test) were tested. In all cases, both conditions were
satisfied, and thus one-way ANOVA was used to determine the possibility of differences among
the groups. If multiple comparisons were then required, Tukey test was applied. The
relationships between mechanical parameters and the amount of collagen and elastic fibers were
evaluated by Spearman correlation. In all tests the significance level was set at 5%. Statistical
analyses were done with Sigmastat 2.0 (Jandel Scientific, San Rafael, CA, USA).

Blue-200302-2560C- R1 - Online-only repository
10
References
E1. Mortola JP, Novoraj A. Two-sidearm tracheal cannula for respiratory airflow measurements
in small animals. J Appl Physiol 1983;55:250-253.
E2. Baydur A, Behrakis PK, Zin WA, Jaeger M, Milic-Emili J. A simple method for assessing
the validity of the esophageal balloon technique. Am Rev Respir Dis 1982;126:788-791.
E3. Kochi T, Okubo S, Zin WA, Milic-Emili J. Flow and volume dependence of pulmonary
mechanics in anesthetized cats. J Appl Physiol 1988;64:441-450.
E4. D'Angelo E, Calderini E, Torri G, Robbato FM, Bono D, Milic-Emili J. Respiratory
mechanics in anesthetized paralyzed humans: effects of flow, volume, and time. J Appl
Physiol 1989;670:2556-2564.
E5. Bates JHT, Baconnier P, and Milic-Emili J. A theoretical analysis of the interrupter technique
for measuring respiratory mechanics. J Appl Physiol 1988;64: 2204-2214.
E6. Bates JHT, Ludwig MS, Sly PD, Brown KA, Martin JG, and Fredberg JJ. Interrupter
resistance elucidated by alveolar pressure measurements in open-chest normal dogs. J Appl
Physiol 1988;65:408-414.
E7. Similowski T, Levy P, Corbeil C, Albala M, Pariente R, Derenne JP, Bates JHT, Jonson B,
and Milic-Emili J. Viscoelastic behavior of lung and chest wall in dogs determined by flow
interruption. J Appl Physiol 1989;67:2219-2229.
E8. Bates JHT, Hunter IW, Sly PD, Okubo S, Filiatrault S, and Milic Emili J. Effect of valve
closure time on the determination of respiratory resistance by flow interruption. Med Biol
Eng Comput 1987;25:136-140.
E9. Salerno FG, Pare P, Ludwig MS. A comparative study of elastic properties of rat and guinea
pig parenchymal strips. Am J Respir Crit Care Med 1998;157:846-852.

Blue-200302-2560C- R1 - Online-only repository
11
E10. Lopez-Aguilar J, Romero PV. Effect of elastase pretreatment on rat lung strip induced
constriction. Respir Physiol 1998;113:239-246.
E11. Faffe DS, Silva GH, Kurtz PMP, Negri EM, Capelozzi VL, Rocco PRM, Zin WA. Lung
tissue mechanics and extracellular matrix composition in a murine model of silicosis. J Appl
Physiol 2001;90:1400-1406.
E12. Rocco PRM, Negri EM, Kurtz PM, Vasconcellos FP, Silva GH, Capelozzi VL, Romero
PV, Zin WA. Lung tissue mechanics and extracellular matrix remodeling in acute lung
injury. Am J Respir Crit Care Med 2001;164:1067-1071.
E13. Moretto A, Dallaire M, Romero PV, Ludwig M. Effect of elastase on oscillation
mechanics of lung parenchymal strips. J Appl Physiol 1994;77:1623-1629.
E14. Yuan H, Ingenito EP, Suki B. Dynamic properties of lung parenchyma: mechanical
contributions of fiber network and interstitial cells. J Appl Physiol 1997;83:1420-1431.
E15. Fredberg JJ, Stamenovic D. On the imperfect elasticity of lung tissue. J Appl Physiol
1989;67:2408-2414.
E16. Nagase T, Dallaire MJ, Ludwig MS. Airway and tissue behavior during early response in
sensitized rats: role of 5-HT and LTD4. J Appl Physiol 1996;80:583-590.
E17. Weibel ER. Morphometry: stereological theory and practical methods. In: Gil J, editor.
Models of Lung Disease-Microscopy and Structural Methods. New York: Marcel Decker;
1990, p.199 -247.
E18. Montes GS. Structural biology of the fibers of the collagenous and elastic system. Cell.
Biology International 1996;20:15-27.
E19. Weigert C. Über eine methode zur farbung elastischer fasern. Zentralbl. Allg. Pathol.
Anat. 1898;9:289-292.

Blue-200302-2560C- R1 - Online-only repository
12
E20. Fullmer HM, Sheetz JH, Narkates AJ. Oxytalan connective tissue fibers: a review. J Oral
Pathol 1974;3:291-316.
Related Documents










![TMZ...2018/12/25 · The lun s are atelectatic C] crepitant C] emphysema tous subcrepi tant The left lung weights The right lung weighs The visceral pleurae The parenchyma is has](https://static.cupdf.com/doc/110x72/60aa6b0304023a14c42ebca0/tmz-20181225-the-lun-s-are-atelectatic-c-crepitant-c-emphysema-tous-subcrepi.jpg)
