Effect of CO 2 supply conditions on lipid production of Chlorella vulgaris from enzymatic hydrolysates of lipid-extracted microalgal biomass residues Hongli Zheng, Zhen Gao, Fengwei Yin, Xiaojun Ji, He Huang ⇑ State Key Laboratory of Materials-Oriented Chemical Engineering, College of Biotechnology and Pharmaceutical Engineering, Nanjing University of Technology, No. 5 Xinmofan Road, Nanjing 210009, People’s Republic of China highlights " CO 2 supply conditions had a significant effect on lipid production. " The most suitable CO 2 concentration for microalgal growth was 5%. " Microalga grew best at a CO 2 aeration rate of 0.5 vvm. " The fatty acids of the microalga are suitable for biodiesel production. article info Article history: Received 25 June 2012 Received in revised form 11 September 2012 Accepted 15 September 2012 Available online 25 September 2012 Keywords: Microalga Biodiesel CO 2 supply conditions Lipid-extracted microalgal biomass residues abstract The hydrolysates from lipid-extracted microalgal biomass residues (LMBRs) were used as a source of nutrients for the cultivation of Chlorella vulgaris for lipid production under various CO 2 supply conditions, including different CO 2 concentrations and aeration rates. Both parameters had a significant effect on lipid production. A CO 2 concentration of 5% was found to be most suitable for microalgal growth. Micro- alga grew best at a CO 2 aeration rate of 0.5 vvm. At this rate, biomass concentration and lipid productivity were at a maximum of 3.83 g L 1 and 157 mg L 1 d 1 , respectively, but decreased at lower or higher aer- ation rates. The present results showed that LMBRs utilization was effective in microalgal lipid produc- tion under suitable CO 2 supply conditions. Crown Copyright Ó 2012 Published by Elsevier Ltd. All rights reserved. 1. Introduction Lipid-extracted microalgal biomass residues (LMBRs) are the residual biomass from microalgal biodiesel production processes. These residues are rich in proteins and carbohydrates, and making use of them is necessary to increase the economic and environ- mental feasibility of microalgal biodiesel production (Ehimen et al., 2011). LMBRs could be converted to products such as amino acids and sugars by enzymatic hydrolysis and utilized as nutrient sources for a new crop of microalgae (Zheng et al., 2012a). Since nutrient supplies have a sizeable effect on cost, sustainability, and site selection for microalgal cultivation (Stephens et al., 2010), the use of LMBRs could contribute to the economy of micro- algal biodiesel production. In a previous study, Chlorella vulgaris using both sugars from the hydrolysates of LMBRs and CO 2 supplied by aeration as carbon sources achieves much higher biomass concentration than without CO 2 aeration (Zheng et al., 2012a). This is because aeration supplies CO 2 as inorganic carbon source for C. vulgaris growth, and/or better mixing, resulting in a sufficient light distribution and better mass transfer (Camacho et al., 2011). Therefore, CO 2 supply conditions, including CO 2 concentration and aeration rate, are very important and deserve more detailed investigation in the mixotrophic culti- vation of C. vulgaris for lipid production using the hydrolysates (mainly amino acids and sugars) of LMBRs as nutrient sources. Lastly, the suitability of the extracted microalgal lipids for biodiesel production was evaluated. 2. Methods 2.1. Microalgal strain and cultivation conditions The microalga C. vulgaris (strain CCTCC M 209256) was obtained from the China Center for Type Culture Collection, Wuhan, China. The strain was preserved in 20% (v/v) glycerol at 80 °C. The 0960-8524/$ - see front matter Crown Copyright Ó 2012 Published by Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.biortech.2012.09.048 ⇑ Corresponding author. Tel./fax: +86 25 83172094. E-mail address: [email protected] (H. Huang). Bioresource Technology 126 (2012) 24–30 Contents lists available at SciVerse ScienceDirect Bioresource Technology journal homepage: www.elsevier.com/locate/biortech

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Bioresource Technology 126 (2012) 24–30
Contents lists available at SciVerse ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Effect of CO2 supply conditions on lipid production of Chlorella vulgaris fromenzymatic hydrolysates of lipid-extracted microalgal biomass residues
Hongli Zheng, Zhen Gao, Fengwei Yin, Xiaojun Ji, He Huang ⇑State Key Laboratory of Materials-Oriented Chemical Engineering, College of Biotechnology and Pharmaceutical Engineering, Nanjing University of Technology, No. 5 XinmofanRoad, Nanjing 210009, People’s Republic of China
h i g h l i g h t s
" CO2 supply conditions had a significant effect on lipid production." The most suitable CO2 concentration for microalgal growth was 5%." Microalga grew best at a CO2 aeration rate of 0.5 vvm." The fatty acids of the microalga are suitable for biodiesel production.
a r t i c l e i n f o
Article history:Received 25 June 2012Received in revised form 11 September2012Accepted 15 September 2012Available online 25 September 2012
Keywords:MicroalgaBiodieselCO2 supply conditionsLipid-extracted microalgal biomass residues
0960-8524/$ - see front matter Crown Copyright � 2http://dx.doi.org/10.1016/j.biortech.2012.09.048
⇑ Corresponding author. Tel./fax: +86 25 83172094E-mail address: [email protected] (H. Huang).
a b s t r a c t
The hydrolysates from lipid-extracted microalgal biomass residues (LMBRs) were used as a source ofnutrients for the cultivation of Chlorella vulgaris for lipid production under various CO2 supply conditions,including different CO2 concentrations and aeration rates. Both parameters had a significant effect onlipid production. A CO2 concentration of 5% was found to be most suitable for microalgal growth. Micro-alga grew best at a CO2 aeration rate of 0.5 vvm. At this rate, biomass concentration and lipid productivitywere at a maximum of 3.83 g L�1 and 157 mg L�1 d�1, respectively, but decreased at lower or higher aer-ation rates. The present results showed that LMBRs utilization was effective in microalgal lipid produc-tion under suitable CO2 supply conditions.
Crown Copyright � 2012 Published by Elsevier Ltd. All rights reserved.
1. Introduction
Lipid-extracted microalgal biomass residues (LMBRs) are theresidual biomass from microalgal biodiesel production processes.These residues are rich in proteins and carbohydrates, and makinguse of them is necessary to increase the economic and environ-mental feasibility of microalgal biodiesel production (Ehimenet al., 2011). LMBRs could be converted to products such as aminoacids and sugars by enzymatic hydrolysis and utilized as nutrientsources for a new crop of microalgae (Zheng et al., 2012a). Sincenutrient supplies have a sizeable effect on cost, sustainability,and site selection for microalgal cultivation (Stephens et al.,2010), the use of LMBRs could contribute to the economy of micro-algal biodiesel production.
In a previous study, Chlorella vulgaris using both sugars from thehydrolysates of LMBRs and CO2 supplied by aeration as carbon
012 Published by Elsevier Ltd. All r
.
sources achieves much higher biomass concentration than withoutCO2 aeration (Zheng et al., 2012a). This is because aeration suppliesCO2 as inorganic carbon source for C. vulgaris growth, and/or bettermixing, resulting in a sufficient light distribution and better masstransfer (Camacho et al., 2011). Therefore, CO2 supply conditions,including CO2 concentration and aeration rate, are very importantand deserve more detailed investigation in the mixotrophic culti-vation of C. vulgaris for lipid production using the hydrolysates(mainly amino acids and sugars) of LMBRs as nutrient sources.Lastly, the suitability of the extracted microalgal lipids for biodieselproduction was evaluated.
2. Methods
2.1. Microalgal strain and cultivation conditions
The microalga C. vulgaris (strain CCTCC M 209256) was obtainedfrom the China Center for Type Culture Collection, Wuhan, China.The strain was preserved in 20% (v/v) glycerol at �80 �C. The
ights reserved.

H. Zheng et al. / Bioresource Technology 126 (2012) 24–30 25
culture medium was composed of instant ocean synthetic sea salt(Aquarium Systems, Inc., USA), 34 g L�1; 200 mL L�1 hydrolysatesof LMBRs. The hydrolysates were sterilized using a sterile 0.45-lm membrane filter (Millipore Corporation, USA). A 1.25-L bubblecolumn photobioreactor (25.0 cm in height, 8.0 cm in diameter, aclosed system) was used with a working volume of 1 L. In all cases,C. vulgaris was inoculated at 1:10 (v/v) ratio into the photobioreac-tor. The initial biomass concentration of 0.10 g L�1 was used in allruns. The culture temperature of 25 �C was regulated by waterrecycled in the outer layer of the photobioreactor. Ten fluorescentlamps were arranged around the photobioreactor to supply contin-uous illumination of 300 lmol photons m�2 s�1 with a 12/12 hlight/dark cycle. A gas sparger was located at the bottom of thereactor. For CO2 concentration experiments, the bioreactor wasaerated at a rate of 1.0 vvm (volume gas per volume medium perminute) with a mixture of compressed air and CO2 in different con-centrations. For CO2 aeration rate experiments, the cultures wereaerated with 5% sterile CO2 at different aeration rates. The initialpH of the medium for all runs was adjusted to 6.5 with 0.5 MHCl or 0.5 M NaOH. The cultivation cycle of all runs was 10 days.The cells were centrifuged at 1,600g for 5 min at room tempera-ture, washed with distilled water three times, and prepared for fur-ther analysis. After biomass harvesting, the culture medium wereused for the analysis of amino acids and sugars.
2.2. Enzymatic hydrolysis of LMBRs
C. vulgaris biomass was obtained from a previous study (Zhenget al., 2011). The methods for preparing and hydrolyzing LMBRsfrom C. vulgaris biomass were the same as outlined by Zhenget al. (2012a). The hydrolysates were centrifuged at 4,800g for5 min, and the supernatants were diluted 1:10 with ethanol priorto HPLC analysis of amino acids and sugars. The samples and mo-bile phase were filtered separately with a 0.45-lm membranefilter.
2.3. Analytical methods
2.3.1. Analysis of sugarsXylose, glucose and arabinose in the hydrolysates and culture
medium were analyzed using the HPLC method according to Yanet al. (2009).
2.3.2. Analysis of amino acidsAmino acids in the hydrolysates and culture medium were ana-
lyzed using a Dionex Ultimate 3,000 HPLC system equipped with aSepax AA column (300 � 4.6 mm, 5 lm, Sepax Technologies, Inc.,Delaware, USA). Amino acid standard solutions and the superna-tants of the hydrolysates and culture medium were derivatizedwith phenylisothiocyanate using a slightly modified version ofthe method described by Heinrikson and Meredith (1984). After1 h of derivatization, 200 lL of hexane was added and mixed,and the mixture was left unagitated for 10 min, then the deriva-tives in the lower layer were filtered with a 0.45-lm membrane fil-ter before injection (20 lL). HPLC analysis was carried out using a4:1 (cyanomethane: water) mobile phase, a flow rate of0.4 mL min�1 and a 36 �C column temperature. The amino acidswere identified by comparison of the retention times with thoseof authentic standards. The quantification was based on the exter-nal standard method.
2.3.3. Measurement of biomass concentrationAfter harvest, the microalgal pellets were lyophilized in a freeze
drier (Model FD-1A-50, Beijing Boyikang Lab Instrument Co. Ltd.,Beijing, China) for dry weight measurements. The biomass concen-tration (dry mass) of C. vulgaris (BC, g L–1) was calculated using
optical density (OD) measurements at 680 nm by a UV/VIS spectro-photometer (Lambda 25, PerkinElmer, Inc., USA) according to thefollowing equation:
BC ¼ 0:548� OD680ðR2 ¼ 0:993Þ ð1Þ
Each sample of 5 mL microalgal suspension was used to mea-sure its OD. Samples were diluted when necessary to give an absor-bance in the range of 0.1–1.0. The optical density measurementswere periodically checked by gravimetry.
Biomass productivity (g L�1 d�1) was calculated according tothe following equation:
P ¼ X2 � X1
t2 � t1ð2Þ
And specific growth rate l (d�1) was calculated from the follow-ing equation:
l ¼ lnðX2=X1Þt2 � t1
ð3Þ
where X1 (g L�1) and X2 (g L�1) are the cell concentration at exper-imental time t1 (d) and t2 (d), respectively.
2.3.4. Determination of chlorophyll-a contentA sample of 10 mL C. vulgaris was centrifuged at 1,600g for
5 min, washed with distilled water three times and the pelletswere resuspended in 10 mL of acetone. The mixture was disruptedusing a 600 W ultrasonic cell disintegrator (GA92-IID, Wuxi Shang-jia Biotechnology Co. Ltd., Jiangsu Province, China) for 30 s with 5 sintervals for a total working time of 10 min followed by centrifuga-tion at 4,800g for 5 min. The amount of chlorophyll-a in the super-natants (Ca mg L�1) and the optical density were correlatedaccording to Wellburn (1994):
Ca ¼ 15:65� OD666 � 7:34� OD653 ð4Þ
The chlorophyll-a content was the ratio of Ca to the biomassconcentration in the 10 mL sample.
2.3.5. Lipid extraction and composition analysis of fatty acidsEach 200 mL microalgal suspension sample based on the
hydrolysates of LMBRs as the culture medium was disrupted bycellulase. The cell slurries were extracted with a mixture of hex-ane–methanol (1:1, v/v) with the samples in a proportion of 1:2using a slightly modified version of Bligh and Dyer’s method (Blighand Dyer, 1959). The mixtures were shaken for 5 min in a separa-tory funnel, and the organic phases were allowed to separate andevaporate at 30 �C using a rotary evaporator to obtain lipids. Theweight of lipids from the organic phases was measured using anelectronic scale. The lipid content was defined as dry weight ratioof extracted lipids to biomass. Lipid productivity (mg L�1 d�1) wascalculated by using the following equation:
PL ¼ ðX2LC2 � X1LC1 Þ=ðt2 � t1Þ ð5Þ
where LC1 and LC2 are the lipid content (%) at time t1 and t2, respec-tively; X1 and X2 are defined same as Eq. (3). Finally, the lipids wereapplied for fatty acids composition analysis.
The composition analysis of fatty acids was established usinggas chromatography–mass spectrometry (GC–MS) (Thermo Finni-gan, USA). Fatty acids were methylated by BF3 in methanol (Metc-halfe and Schmitz, 1961), and dissolved in hexane. Subsequently,1.0 lL of this solution was injected into the GC–MS instrument.The GC–MS method was the same as outlined by Ren et al. (2010).
2.4. Experimental design and data analysis
The experiment was designed and carried out at random. All thetreatments were repeated three times and data are reported as the

26 H. Zheng et al. / Bioresource Technology 126 (2012) 24–30
mean ± SD values. Analysis of variance (ANOVA) and Duncan’smultiple range test (when ANOVA indicated at least one signifi-cantly different result) (Duncan, 1955) were applied for data anal-ysis. Statistical software SPSS 13.0 (SPSS Inc., Chicago, IL, USA) wasapplied to the data generated in CO2 concentration and aerationrate experiments to identify differences in their maximum biomassconcentration (Xmax), maximum biomass productivity (Pmax), max-imum specific growth rate (lmax), total concentrations of aminoacids and sugars, lipid productivity and maximum lipid yield,respectively.
3. Results and discussion
3.1. Effect of CO2 supply conditions on cell growth of C. vulgaris
3.1.1. Effect of CO2 concentration on cell growth of C. vulgarisC. vulgaris was cultivated at an aeration rate of 1.0 vvm using
sterile CO2 in different concentrations. The effect of CO2 concentra-tion on the microalgal growth is shown in Table 1 and Fig. 1. Theoptimal CO2 concentration was 5% with Xmax of 3.76 g L�1, Pmax
of 1.01 g L�1 d�1 and lmax of 1.2 d�1. Xmax, Pmax and lmax of C. vul-garis all increased before they decreased along with increasing CO2
concentration. The culture aerated with air (CO2 concentration wasapproximately 0.03%) reached a plateau stage after 10-day cultiva-tion and its maximum biomass concentration was 2.71 g L�1. Mic-roalgal growth was inhibited with 0.03% CO2 compared with thataerated with 5% CO2. This may be due to the lower concentrationof CO2 in air and, therefore, its supply of carbon source was insuf-ficient for microalgal growth. In our case, the optimal CO2 concen-tration for C. vulgaris was 5%. However, it varies for differentmicroalgal species. The optimal CO2 concentration for Chlorellasp. was 10% (Ramanan et al., 2010). The results in our present studyshowed that CO2 concentration above 5% may be harmful tomicroalgal cells and inhibit microalgal growth. Similar results wereobtained by Chiu et al. (2008) and Yoo et al. (2010). When theC. vulgaris culture was aerated with 15% CO2, Xmax, Pmax and lmax
were substantially decreased compared with those in the cultureaerated with 5% CO2. It indicated that the microalgal growth wascompletely inhibited with 15% CO2. This is because CO2 of highconcentration can result in low pH of the culture medium (Tanget al., 2011), which would cause an activity decrease of ribulose1,5-bisphosphate carboxylase-oxygenase, a key enzyme of photo-synthesis (Jensen and Bahr, 1977), causing inhibition of microalgalgrowth.
Chlorophyll-a content from different CO2 concentrations as afunction of culture time was investigated. Fig. 1A–E show thatchlorophyll-a content increased during the initial 7 days for0.03%, 1% and 5% CO2, respectively, 6 days for 10% CO2, and 4 daysfor 15% CO2, and decreased afterwards. Furthermore, chlorophyll-asynthesis was inhibited with 15% CO2. The total concentrations ofamino acids and sugars decreased with increasing culture time.Amino acids and sugars in the medium of the cultures were ex-hausted except for the culture with 15% CO2. The consumption rateof sugars was faster than that of amino acids, indicating that C. vul-garis consumed more carbon source than nitrogen source whilegrowing. In a previous study (Zheng et al., 2012b), C. vulgaris used
Table 1Cell growth of C. vulgaris under various CO2 supply conditions.
Parameters CO2 concentration (%)
0.03 1 5 10 15
Xmax (g L�1) 2.71 ± 0.12 3.32 ± 0.15 3.76 ± 0.16 2.59 ± 0.10 0.65 ±Pmax (g L�1 d�1) 0.72 ± 0.06 0.90 ± 0.08 1.01 ± 0.11 0.69 ± 0.10 0.15 ±lmax (d�1) 0.91 ± 0.09 1.09 ± 0.08 1.20 ± 0.13 0.86 ± 0.08 0.25 ±
Xmax, the maximum biomass concentration; Pmax, the maximum biomass productivity; a
KNO3 (inorganic nitrogen) to grow, while in the present study, itwas confirmed that C. vulgaris was also able to use amino acids (or-ganic nitrogen) to grow. For each CO2 concentration, the fasterchlorophyll-a content increased, the more amino acids and sugarswere consumed. The results indicated that the microalga grewfastest at this stage. The culture with 5% CO2 had the highest valueof maximum chlorophyll-a content (39.2 mg g�1) at day 7 whenamino acids and sugars were consumed fastest. Changes in sugarsand chlorophyll-a content (Fig. 1) in the cultures indicated that C.vulgaris used sugars and CO2 synchronously as carbon sources forgrowth; thus it was mixotrophic. Yeh and Chang (2012) reportedthat C. vulgaris could grow under autotrophic, heterotrophic andmixotrophic conditions.
3.1.2. Effect of CO2 aeration rate on cell growth of C. vulgarisC. vulgaris used CO2 bubbled into the medium as inorganic car-
bon source for growth. The effect of CO2 aeration rate on the growthof the microalga is shown in Table 1 and Fig. 2. C. vulgaris was cul-tivated with 5% CO2 under different aeration rates. The optimal CO2
aeration rate was 0.5 vvm with Xmax of 3.83 g L�1, Pmax of 1.07 gL�1 d�1 and lmax of 1.22 d�1. Xmax, Pmax and lmax increased signifi-cantly (P < 0.05) with increasing CO2 aeration up to a rate of0.5 vvm. The result is confirmed by Ryu et al. (2009) that the bio-mass concentration of Chlorella sp. was enhanced with increasingaeration rate from 0.06 to 0.4 vvm. However, Xmax, Pmax and lmax
decreased significantly (P < 0.05) when the CO2 aeration rate wasincreased above 0.5 vvm. The results indicated that the CO2 aera-tion rate above 0.5 vvm may be harmful to microalgal cells and in-hibit the microalgal growth. This might be because high aerationrates resulted in microalgal cell damage by shear stress, leadingto reduced cell growth or even cell death (Barbosa et al., 2003; Barb-osa et al., 2004). More detailed investigations are needed to furthervalidate this hypothesis. In the present study, the highest value ofmaximum biomass concentration (3.83 g L�1) was obtained at anaeration rate of 0.5 vvm, which was 1.30 and 3.58 times that with0.1 and 2.0 vvm CO2 aeration, respectively. The results indicatedthat a suitable CO2 aeration rate can promote the microalgalgrowth. The possible reason is that a suitable CO2 aeration ratemight enhance gas–liquid mass transfer (Mirón et al., 2000), replen-ish sufficient CO2 for the microalgal growth, cause proper mixing ofthe culture medium, prevent the microalgal cells from sinking tothe bottom of the reactor, ensure all the microalgal cells in contactwith nutrients, CO2 and light, and ensure produced oxygen removalfrom the culture system. Oxygen may cause inhibition of photosyn-thesis if present in high concentrations (Camacho et al., 2011).
The effect of CO2 aeration rate on chlorophyll-a content as wellas on total concentrations of amino acids and sugars was investi-gated. Fig. 2A–E show that chlorophyll-a content increased duringthe initial 7 days for 0.1, 0.5, 1.0 and 1.5 vvm CO2 aeration, respec-tively, or 6 days for 2.0 vvm CO2 aeration, and decreased after-wards. Similar results were obtained by a previous study (Zhenget al., 2012a). Amino acids and sugars in the culture medium wereused up except for the culture with 2 vvm CO2 aeration. Theseresults indicated that the cultures used both CO2 and sugars as car-bon sources during the initial culture stage and mainly used CO2 ascarbon source during the final culture stage. For each CO2 aeration
Aeration rate (vvm)
0.1 0.5 1.0 1.5 2.0
0.06 2.95 ± 0.15 3.83 ± 0.20 3.51 ± 0.18 2.54 ± 0.16 1.07 ± 0.100.02 0.84 ± 0.07 1.07 ± 0.11 0.95 ± 0.09 0.67 ± 0.07 0.32 ± 0.030.03 1.03 ± 0.10 1.22 ± 0.12 1.13 ± 0.09 0.84 ± 0.07 0.45 ± 0.06
nd lmax, the maximum specific growth rate.
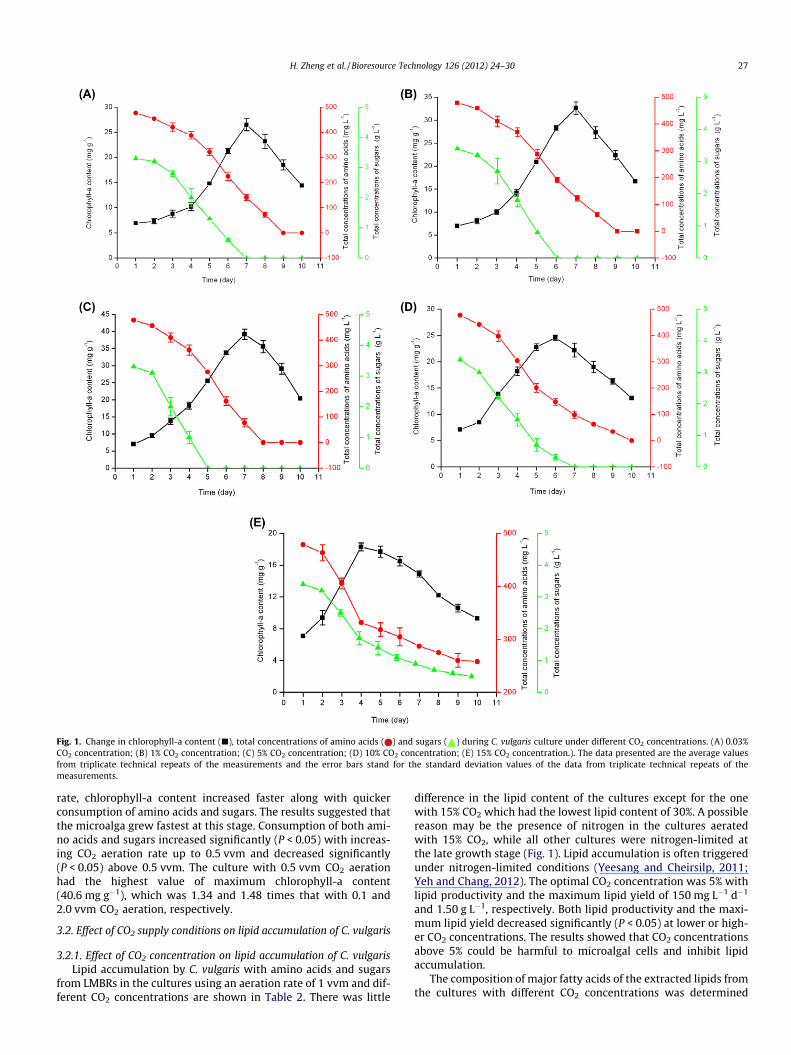
Fig. 1. Change in chlorophyll-a content (j), total concentrations of amino acids ( ) and sugars ( ) during C. vulgaris culture under different CO2 concentrations. (A) 0.03%CO2 concentration; (B) 1% CO2 concentration; (C) 5% CO2 concentration; (D) 10% CO2 concentration; (E) 15% CO2 concentration.). The data presented are the average valuesfrom triplicate technical repeats of the measurements and the error bars stand for the standard deviation values of the data from triplicate technical repeats of themeasurements.
H. Zheng et al. / Bioresource Technology 126 (2012) 24–30 27
rate, chlorophyll-a content increased faster along with quickerconsumption of amino acids and sugars. The results suggested thatthe microalga grew fastest at this stage. Consumption of both ami-no acids and sugars increased significantly (P < 0.05) with increas-ing CO2 aeration rate up to 0.5 vvm and decreased significantly(P < 0.05) above 0.5 vvm. The culture with 0.5 vvm CO2 aerationhad the highest value of maximum chlorophyll-a content(40.6 mg g�1), which was 1.34 and 1.48 times that with 0.1 and2.0 vvm CO2 aeration, respectively.
3.2. Effect of CO2 supply conditions on lipid accumulation of C. vulgaris
3.2.1. Effect of CO2 concentration on lipid accumulation of C. vulgarisLipid accumulation by C. vulgaris with amino acids and sugars
from LMBRs in the cultures using an aeration rate of 1 vvm and dif-ferent CO2 concentrations are shown in Table 2. There was little
difference in the lipid content of the cultures except for the onewith 15% CO2 which had the lowest lipid content of 30%. A possiblereason may be the presence of nitrogen in the cultures aeratedwith 15% CO2, while all other cultures were nitrogen-limited atthe late growth stage (Fig. 1). Lipid accumulation is often triggeredunder nitrogen-limited conditions (Yeesang and Cheirsilp, 2011;Yeh and Chang, 2012). The optimal CO2 concentration was 5% withlipid productivity and the maximum lipid yield of 150 mg L�1 d�1
and 1.50 g L�1, respectively. Both lipid productivity and the maxi-mum lipid yield decreased significantly (P < 0.05) at lower or high-er CO2 concentrations. The results showed that CO2 concentrationsabove 5% could be harmful to microalgal cells and inhibit lipidaccumulation.
The composition of major fatty acids of the extracted lipids fromthe cultures with different CO2 concentrations was determined
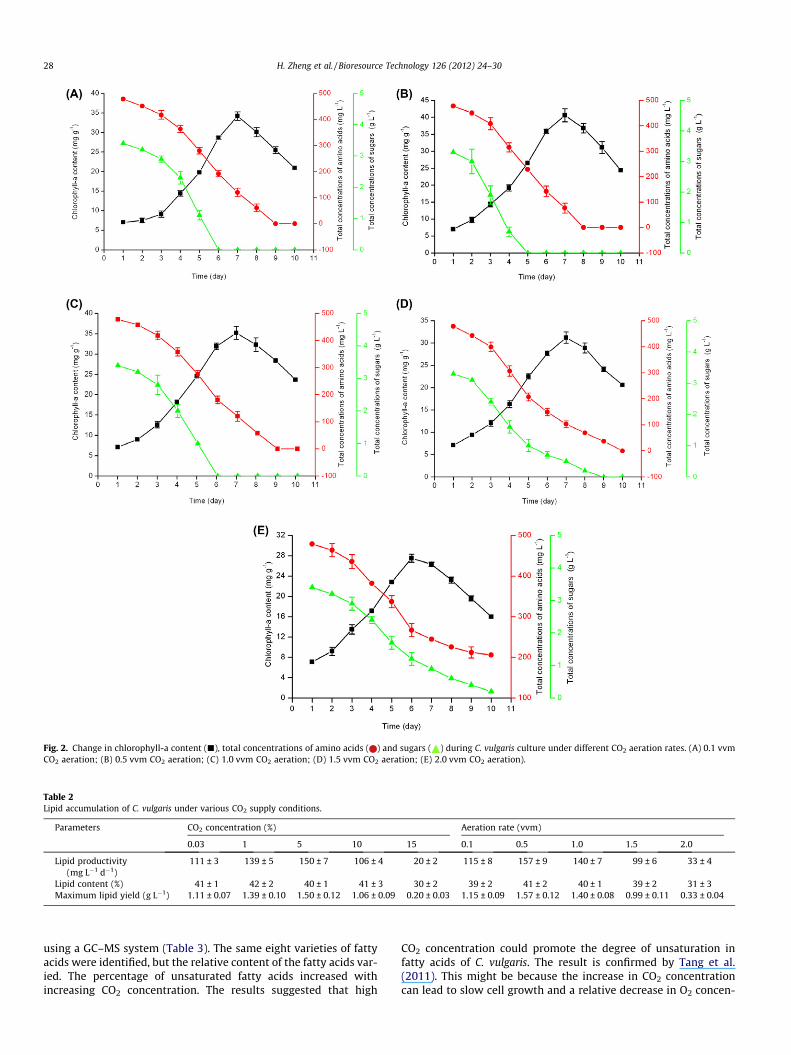
Fig. 2. Change in chlorophyll-a content (j), total concentrations of amino acids ( ) and sugars ( ) during C. vulgaris culture under different CO2 aeration rates. (A) 0.1 vvmCO2 aeration; (B) 0.5 vvm CO2 aeration; (C) 1.0 vvm CO2 aeration; (D) 1.5 vvm CO2 aeration; (E) 2.0 vvm CO2 aeration).
Table 2Lipid accumulation of C. vulgaris under various CO2 supply conditions.
Parameters CO2 concentration (%) Aeration rate (vvm)
0.03 1 5 10 15 0.1 0.5 1.0 1.5 2.0
Lipid productivity(mg L�1 d�1)
111 ± 3 139 ± 5 150 ± 7 106 ± 4 20 ± 2 115 ± 8 157 ± 9 140 ± 7 99 ± 6 33 ± 4
Lipid content (%) 41 ± 1 42 ± 2 40 ± 1 41 ± 3 30 ± 2 39 ± 2 41 ± 2 40 ± 1 39 ± 2 31 ± 3Maximum lipid yield (g L�1) 1.11 ± 0.07 1.39 ± 0.10 1.50 ± 0.12 1.06 ± 0.09 0.20 ± 0.03 1.15 ± 0.09 1.57 ± 0.12 1.40 ± 0.08 0.99 ± 0.11 0.33 ± 0.04
28 H. Zheng et al. / Bioresource Technology 126 (2012) 24–30
using a GC–MS system (Table 3). The same eight varieties of fattyacids were identified, but the relative content of the fatty acids var-ied. The percentage of unsaturated fatty acids increased withincreasing CO2 concentration. The results suggested that high
CO2 concentration could promote the degree of unsaturation infatty acids of C. vulgaris. The result is confirmed by Tang et al.(2011). This might be because the increase in CO2 concentrationcan lead to slow cell growth and a relative decrease in O2 concen-
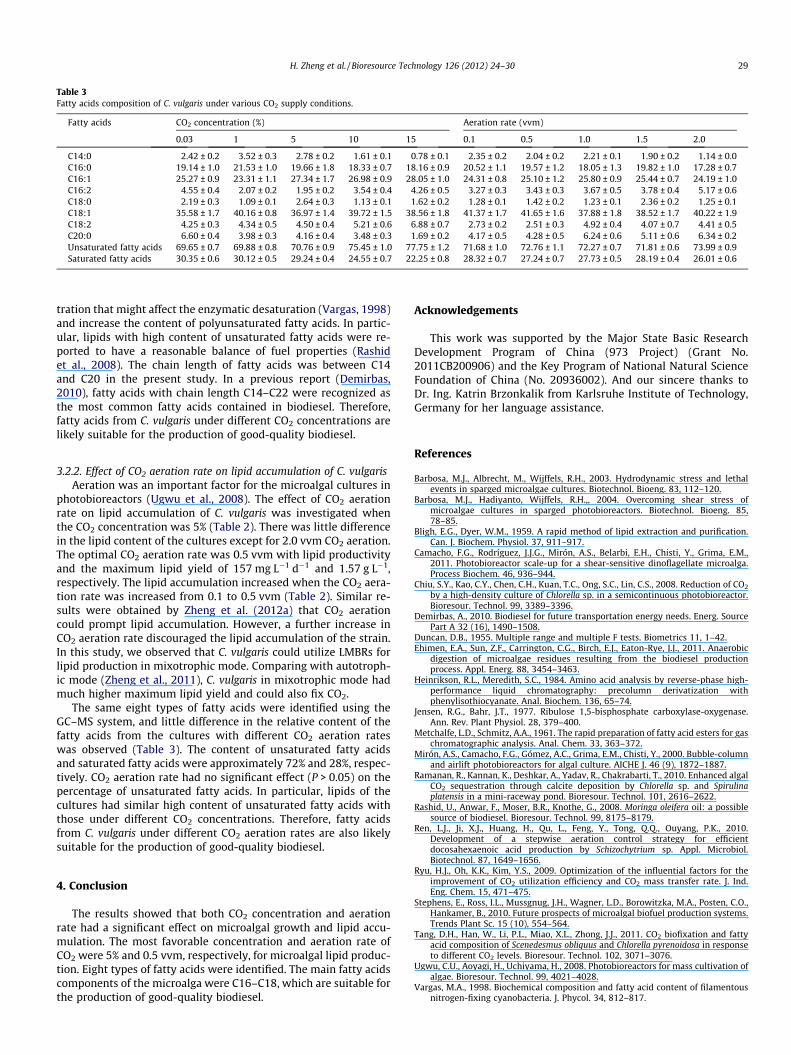
Table 3Fatty acids composition of C. vulgaris under various CO2 supply conditions.
Fatty acids CO2 concentration (%) Aeration rate (vvm)
0.03 1 5 10 15 0.1 0.5 1.0 1.5 2.0
C14:0 2.42 ± 0.2 3.52 ± 0.3 2.78 ± 0.2 1.61 ± 0.1 0.78 ± 0.1 2.35 ± 0.2 2.04 ± 0.2 2.21 ± 0.1 1.90 ± 0.2 1.14 ± 0.0C16:0 19.14 ± 1.0 21.53 ± 1.0 19.66 ± 1.8 18.33 ± 0.7 18.16 ± 0.9 20.52 ± 1.1 19.57 ± 1.2 18.05 ± 1.3 19.82 ± 1.0 17.28 ± 0.7C16:1 25.27 ± 0.9 23.31 ± 1.1 27.34 ± 1.7 26.98 ± 0.9 28.05 ± 1.0 24.31 ± 0.8 25.10 ± 1.2 25.80 ± 0.9 25.44 ± 0.7 24.19 ± 1.0C16:2 4.55 ± 0.4 2.07 ± 0.2 1.95 ± 0.2 3.54 ± 0.4 4.26 ± 0.5 3.27 ± 0.3 3.43 ± 0.3 3.67 ± 0.5 3.78 ± 0.4 5.17 ± 0.6C18:0 2.19 ± 0.3 1.09 ± 0.1 2.64 ± 0.3 1.13 ± 0.1 1.62 ± 0.2 1.28 ± 0.1 1.42 ± 0.2 1.23 ± 0.1 2.36 ± 0.2 1.25 ± 0.1C18:1 35.58 ± 1.7 40.16 ± 0.8 36.97 ± 1.4 39.72 ± 1.5 38.56 ± 1.8 41.37 ± 1.7 41.65 ± 1.6 37.88 ± 1.8 38.52 ± 1.7 40.22 ± 1.9C18:2 4.25 ± 0.3 4.34 ± 0.5 4.50 ± 0.4 5.21 ± 0.6 6.88 ± 0.7 2.73 ± 0.2 2.51 ± 0.3 4.92 ± 0.4 4.07 ± 0.7 4.41 ± 0.5C20:0 6.60 ± 0.4 3.98 ± 0.3 4.16 ± 0.4 3.48 ± 0.3 1.69 ± 0.2 4.17 ± 0.5 4.28 ± 0.5 6.24 ± 0.6 5.11 ± 0.6 6.34 ± 0.2Unsaturated fatty acids 69.65 ± 0.7 69.88 ± 0.8 70.76 ± 0.9 75.45 ± 1.0 77.75 ± 1.2 71.68 ± 1.0 72.76 ± 1.1 72.27 ± 0.7 71.81 ± 0.6 73.99 ± 0.9Saturated fatty acids 30.35 ± 0.6 30.12 ± 0.5 29.24 ± 0.4 24.55 ± 0.7 22.25 ± 0.8 28.32 ± 0.7 27.24 ± 0.7 27.73 ± 0.5 28.19 ± 0.4 26.01 ± 0.6
H. Zheng et al. / Bioresource Technology 126 (2012) 24–30 29
tration that might affect the enzymatic desaturation (Vargas, 1998)and increase the content of polyunsaturated fatty acids. In partic-ular, lipids with high content of unsaturated fatty acids were re-ported to have a reasonable balance of fuel properties (Rashidet al., 2008). The chain length of fatty acids was between C14and C20 in the present study. In a previous report (Demirbas,2010), fatty acids with chain length C14–C22 were recognized asthe most common fatty acids contained in biodiesel. Therefore,fatty acids from C. vulgaris under different CO2 concentrations arelikely suitable for the production of good-quality biodiesel.
3.2.2. Effect of CO2 aeration rate on lipid accumulation of C. vulgarisAeration was an important factor for the microalgal cultures in
photobioreactors (Ugwu et al., 2008). The effect of CO2 aerationrate on lipid accumulation of C. vulgaris was investigated whenthe CO2 concentration was 5% (Table 2). There was little differencein the lipid content of the cultures except for 2.0 vvm CO2 aeration.The optimal CO2 aeration rate was 0.5 vvm with lipid productivityand the maximum lipid yield of 157 mg L�1 d�1 and 1.57 g L�1,respectively. The lipid accumulation increased when the CO2 aera-tion rate was increased from 0.1 to 0.5 vvm (Table 2). Similar re-sults were obtained by Zheng et al. (2012a) that CO2 aerationcould prompt lipid accumulation. However, a further increase inCO2 aeration rate discouraged the lipid accumulation of the strain.In this study, we observed that C. vulgaris could utilize LMBRs forlipid production in mixotrophic mode. Comparing with autotroph-ic mode (Zheng et al., 2011), C. vulgaris in mixotrophic mode hadmuch higher maximum lipid yield and could also fix CO2.
The same eight types of fatty acids were identified using theGC–MS system, and little difference in the relative content of thefatty acids from the cultures with different CO2 aeration rateswas observed (Table 3). The content of unsaturated fatty acidsand saturated fatty acids were approximately 72% and 28%, respec-tively. CO2 aeration rate had no significant effect (P > 0.05) on thepercentage of unsaturated fatty acids. In particular, lipids of thecultures had similar high content of unsaturated fatty acids withthose under different CO2 concentrations. Therefore, fatty acidsfrom C. vulgaris under different CO2 aeration rates are also likelysuitable for the production of good-quality biodiesel.
4. Conclusion
The results showed that both CO2 concentration and aerationrate had a significant effect on microalgal growth and lipid accu-mulation. The most favorable concentration and aeration rate ofCO2 were 5% and 0.5 vvm, respectively, for microalgal lipid produc-tion. Eight types of fatty acids were identified. The main fatty acidscomponents of the microalga were C16–C18, which are suitable forthe production of good-quality biodiesel.
Acknowledgements
This work was supported by the Major State Basic ResearchDevelopment Program of China (973 Project) (Grant No.2011CB200906) and the Key Program of National Natural ScienceFoundation of China (No. 20936002). And our sincere thanks toDr. Ing. Katrin Brzonkalik from Karlsruhe Institute of Technology,Germany for her language assistance.
References
Barbosa, M.J., Albrecht, M., Wijffels, R.H., 2003. Hydrodynamic stress and lethalevents in sparged microalgae cultures. Biotechnol. Bioeng. 83, 112–120.
Barbosa, M.J., Hadiyanto, Wijffels, R.H.,, 2004. Overcoming shear stress ofmicroalgae cultures in sparged photobioreactors. Biotechnol. Bioeng. 85,78–85.
Bligh, E.G., Dyer, W.M., 1959. A rapid method of lipid extraction and purification.Can. J. Biochem. Physiol. 37, 911–917.
Camacho, F.G., Rodríguez, J.J.G., Mirón, A.S., Belarbi, E.H., Chisti, Y., Grima, E.M.,2011. Photobioreactor scale-up for a shear-sensitive dinoflagellate microalga.Process Biochem. 46, 936–944.
Chiu, S.Y., Kao, C.Y., Chen, C.H., Kuan, T.C., Ong, S.C., Lin, C.S., 2008. Reduction of CO2
by a high-density culture of Chlorella sp. in a semicontinuous photobioreactor.Bioresour. Technol. 99, 3389–3396.
Demirbas, A., 2010. Biodiesel for future transportation energy needs. Energ. SourcePart A 32 (16), 1490–1508.
Duncan, D.B., 1955. Multiple range and multiple F tests. Biometrics 11, 1–42.Ehimen, E.A., Sun, Z.F., Carrington, C.G., Birch, E.J., Eaton-Rye, J.J., 2011. Anaerobic
digestion of microalgae residues resulting from the biodiesel productionprocess. Appl. Energ. 88, 3454–3463.
Heinrikson, R.L., Meredith, S.C., 1984. Amino acid analysis by reverse-phase high-performance liquid chromatography: precolumn derivatization withphenylisothiocyanate. Anal. Biochem. 136, 65–74.
Jensen, R.G., Bahr, J.T., 1977. Ribulose 1,5-bisphosphate carboxylase-oxygenase.Ann. Rev. Plant Physiol. 28, 379–400.
Metchalfe, L.D., Schmitz, A.A., 1961. The rapid preparation of fatty acid esters for gaschromatographic analysis. Anal. Chem. 33, 363–372.
Mirón, A.S., Camacho, F.G., Gómez, A.C., Grima, E.M., Chisti, Y., 2000. Bubble-columnand airlift photobioreactors for algal culture. AICHE J. 46 (9), 1872–1887.
Ramanan, R., Kannan, K., Deshkar, A., Yadav, R., Chakrabarti, T., 2010. Enhanced algalCO2 sequestration through calcite deposition by Chlorella sp. and Spirulinaplatensis in a mini-raceway pond. Bioresour. Technol. 101, 2616–2622.
Rashid, U., Anwar, F., Moser, B.R., Knothe, G., 2008. Moringa oleifera oil: a possiblesource of biodiesel. Bioresour. Technol. 99, 8175–8179.
Ren, L.J., Ji, X.J., Huang, H., Qu, L., Feng, Y., Tong, Q.Q., Ouyang, P.K., 2010.Development of a stepwise aeration control strategy for efficientdocosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol.Biotechnol. 87, 1649–1656.
Ryu, H.J., Oh, K.K., Kim, Y.S., 2009. Optimization of the influential factors for theimprovement of CO2 utilization efficiency and CO2 mass transfer rate. J. Ind.Eng. Chem. 15, 471–475.
Stephens, E., Ross, I.L., Mussgnug, J.H., Wagner, L.D., Borowitzka, M.A., Posten, C.O.,Hankamer, B., 2010. Future prospects of microalgal biofuel production systems.Trends Plant Sc. 15 (10), 554–564.
Tang, D.H., Han, W., Li, P.L., Miao, X.L., Zhong, J.J., 2011. CO2 biofixation and fattyacid composition of Scenedesmus obliquus and Chlorella pyrenoidosa in responseto different CO2 levels. Bioresour. Technol. 102, 3071–3076.
Ugwu, C.U., Aoyagi, H., Uchiyama, H., 2008. Photobioreactors for mass cultivation ofalgae. Bioresour. Technol. 99, 4021–4028.
Vargas, M.A., 1998. Biochemical composition and fatty acid content of filamentousnitrogen-fixing cyanobacteria. J. Phycol. 34, 812–817.

30 H. Zheng et al. / Bioresource Technology 126 (2012) 24–30
Wellburn, A.L., 1994. The spectral determination of chlorophylls a and b, as well astotal carotenoids, using various solvents with spectrophotometers of differentresolution. J. Plant Physiol. 144, 307–313.
Yan, L.S., Zhang, H.M., Chen, J.W., Lin, Z.X., Jin, Q., Jia, H.H., Huang, H., 2009. Dilutesulfuric acid cycle spray flow-through pretreatment of corn stover forenhancement of sugar recovery. Bioresour. Technol. 100 (5), 1803–1808.
Yeesang, C., Cheirsilp, B., 2011. Effect of nitrogen, salt, and iron content in thegrowth medium and light intensity on lipid production by microalgae isolatedfrom freshwater sources in Thailand. Bioresour. Technol. 102, 3034–3040.
Yeh, K.L., Chang, J.S., 2012. Effects of cultivation conditions and media compositionon cell growth and lipid productivity of indigenous microalga Chlorella vulgarisESP-31. Bioresour. Technol. 105, 120–127.
Yoo, C., Jun, S.Y., Lee, J.Y., Ahn, C.Y., Oh, H.M., 2010. Selection of microalgae for lipidproduction under high levels carbon dioxide. Bioresour. Technol. 101, S71–S74.
Zheng, H.L., Gao, Z., Yin, F.W., Ji, X.J., Huang, H., 2012a. Lipid production of Chlorellavulgaris from lipid-extracted microalgal biomass residues through two-stepenzymatic hydrolysis. Bioresour. Technol. 117, 1–6.
Zheng, H.L., Gao, Z., Yin, J.L., Tang, X.H., Ji, X.J., Huang, H., 2012b. Harvesting ofmicroalgae by flocculation with poly (c-glutamic acid). Bioresour. Technol. 112,212–220.
Zheng, H.L., Yin, J.L., Gao, Z., Huang, H., Ji, X.J., Dou, C., 2011. Disruption of Chlorellavulgaris cells for the release of biodiesel-producing lipids: a comparison ofgrinding, ultrasonication, bead milling, enzymatic lysis, and microwaves. Appl.Biochem. Biotech. 164, 1215–1224.
Related Documents











