APMIS 114: 480–91, 2006 C 2006 The Authors Printed in Denmark . All rights reserved Journal Compilation C 2006 APMIS ISSN 0903-4641 Effect of adamantylamide dipeptide as adjuvant therapy to praziquantel in mice infected with different S. mansoni isolates SANAA BOTROS, 1 MADIHA MAHMOUD, 1 OLFAT HAMMAM, 2 FATEN SALAH, 3 ZDENEK ZIDEK 4 and KAREL MASEK 4 Departments of 1 Pharmacology, 2 Pathology & 3 Immunology, Theodor Bilharz Research Institute, Warrak El-Hadar, Imbaba, Giza, Egypt, and, 4 Institute of Pharmacology, Academy of Sciences of the Czech Republic Botros S, Mahmoud M, Hammam O, Salah F , Zidek Z, Masek K . Effect of adamantylamide dipeptide as adjuvant therapy to praziquantel in mice infected with different S. mansoni isolates. APMIS 2006;114:480–91. This work investigated the possible use of AdDP as adjuvant therapy to praziquantel (PZQ) in mice infected with PZQ-insusceptible Schistosoma mansoni isolate in a trial to increase the susceptibility of this isolate to the drug. Two batches of C57 BL/6 mice were infected with PZQ-susceptible and -insusceptible S. mansoni isolates, and each batch was divided into five groups. Seven weeks postinfec- tion, the experimental group received AdDP (5 mg/kg) in addition to PZQ in reduced dose (3¿100 mg/kg). Three of the remaining four groups were treated controls; they received AdDP, PZQ in re- duced dose and in full dose (2¿500 mg/kg), and the fourth group was infected untreated. In mice infected with PZQ-susceptible or -insusceptible S. mansoni isolate, praziquantel alone, and in addition to AdDP, reduced worm and egg loads and increased percentage dead eggs. Also, they improved the histopathological changes (reduction in granuloma diameter, percentage fibrotic area with increased percentage degenerated eggs). Inducible nitric oxide synthase (iNOS), nitric oxide (NO) in culture of peritoneal macrophages, and number of CD68-positive cells were decreased with improved alanine amino transaminase. In mice receiving combined therapy AdDPπPZQ, the antischistosomal efficacy and the reductions in the inflammatory granulomatous reactions, NO in cultured peritoneal macro- phages, percentage fibrotic areas recorded, were comparable to that in mice receiving full dose of PZQ, with significantly higher reduction in CD68 cells denoting enhanced antischistosomal efficacy and healing of the inflammatory reactions in the liver. Key words: S. manson; nitric oxide (NO); inducible nitric oxide synthase (iNOS); adamantylamide dipeptide (AdDP); praziquantel (PZQ). Sanaa Botros, Pharmacology Department, Theodor Bilharz Research Institute, El-Nile road Warrak El-Hadar, P.O. Box 30 Imbaba, Giza 12411, Egypt. e-mail: sanaabotros/link.net Severe disease caused by schistosomiasis results from the chronic granulomatous response in- duced by eggs trapped in the liver. The magni- tude of the egg- induced inflammatory response as well as the mechanism leading to severe he- Received 10 November 2005. Accepted 22 February 2006. 480 patic fibrosis appears to be regulated by the relative dominance of type-1 versus type-2 cyto- kines (1). Th1 cells secrete IL-2, INF-g and TNF-a, which play a major role in granuloma formation, while IL-4 secreted by Th2 plays an important role in the development and modu- lation of the granulomatous response (2). At the onset of egg deposition, when the host immune

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

APMIS 114: 480–91, 2006 C 2006 The Authors
Printed in Denmark . All rights reservedJournal Compilation C 2006 APMIS
ISSN 0903-4641
Effect of adamantylamide dipeptide as adjuvant therapyto praziquantel in mice infected with different
S. mansoni isolates
SANAA BOTROS,1 MADIHA MAHMOUD,1 OLFAT HAMMAM,2 FATEN SALAH,3
ZDENEK ZIDEK4 and KAREL MASEK4
Departments of 1Pharmacology, 2Pathology & 3Immunology, Theodor Bilharz Research Institute,Warrak El-Hadar, Imbaba, Giza, Egypt, and, 4Institute of Pharmacology,
Academy of Sciences of the Czech Republic
Botros S, Mahmoud M, Hammam O, Salah F , Zidek Z, Masek K . Effect of adamantylamidedipeptide as adjuvant therapy to praziquantel in mice infected with different S. mansoni isolates.APMIS 2006;114:480–91.
This work investigated the possible use of AdDP as adjuvant therapy to praziquantel (PZQ) in miceinfected with PZQ-insusceptible Schistosoma mansoni isolate in a trial to increase the susceptibility ofthis isolate to the drug. Two batches of C57 BL/6 mice were infected with PZQ-susceptible and-insusceptible S. mansoni isolates, and each batch was divided into five groups. Seven weeks postinfec-tion, the experimental group received AdDP (5 mg/kg) in addition to PZQ in reduced dose (3¿100mg/kg). Three of the remaining four groups were treated controls; they received AdDP, PZQ in re-duced dose and in full dose (2¿500 mg/kg), and the fourth group was infected untreated. In miceinfected with PZQ-susceptible or -insusceptible S. mansoni isolate, praziquantel alone, and in additionto AdDP, reduced worm and egg loads and increased percentage dead eggs. Also, they improved thehistopathological changes (reduction in granuloma diameter, percentage fibrotic area with increasedpercentage degenerated eggs). Inducible nitric oxide synthase (iNOS), nitric oxide (NO) in culture ofperitoneal macrophages, and number of CD68-positive cells were decreased with improved alanineamino transaminase. In mice receiving combined therapy AdDPπPZQ, the antischistosomal efficacyand the reductions in the inflammatory granulomatous reactions, NO in cultured peritoneal macro-phages, percentage fibrotic areas recorded, were comparable to that in mice receiving full dose ofPZQ, with significantly higher reduction in CD68 cells denoting enhanced antischistosomal efficacyand healing of the inflammatory reactions in the liver.
Key words: S. manson; nitric oxide (NO); inducible nitric oxide synthase (iNOS); adamantylamidedipeptide (AdDP); praziquantel (PZQ).
Sanaa Botros, Pharmacology Department, Theodor Bilharz Research Institute, El-Nile road WarrakEl-Hadar, P.O. Box 30 Imbaba, Giza 12411, Egypt. e-mail: sanaabotros/link.net
Severe disease caused by schistosomiasis resultsfrom the chronic granulomatous response in-duced by eggs trapped in the liver. The magni-tude of the egg- induced inflammatory responseas well as the mechanism leading to severe he-
Received 10 November 2005.Accepted 22 February 2006.
480
patic fibrosis appears to be regulated by therelative dominance of type-1 versus type-2 cyto-kines (1). Th1 cells secrete IL-2, INF-g andTNF-a, which play a major role in granulomaformation, while IL-4 secreted by Th2 plays animportant role in the development and modu-lation of the granulomatous response (2). At theonset of egg deposition, when the host immune

IMMUNOMODULATORY DRUG AS ADJUVANT THERAPY IN S. MANSONI
response is predominantly a Th1 response, theincrease in NO has been suggested to play a rolein limiting hepatocyte damage caused by the ar-rival of the eggs in the liver (3). Inducible NOsynthase (iNOS) mRNA was found in schisto-somal liver along with TNF-a and IFN-g; theknown promotor of iNOS expression (4). NOcan have a cytoprotective or cytotoxic effect de-pending not only on the initial insult but alsoon the source, rate of production, and concen-tration of NO in the biological milieu (4).
NO, an endogenously secreted short-livedlipophilic free radical, is the byproduct of theconversion of arginine and oxygen into citrul-line through an enzymatic reaction mediated byNO synthase (5). Three isoforms of this enzymeencoding distinct genes were identified (6). Thethird isoform, the inducible NOS (iNOS, NOS-2), present in virtually all cells (7), is expressedin response to proinflammatory cytokines and/or microbial products, resulting in the pro-longed production of large amounts of NO (8).
In the liver, NO plays a unique role and itsinvolvement in a variety of hepatic processes issupported by the finding that the majority ofcells found in the liver, such as hepatocytes,Kupffer cells, fat-storing cells, and endothelialcells, are able to produce it under adequatestimulation (9). However, whether NO is cyto-protective or cytotoxic is still controversial (4).As host protective, NO acts as effector moleculeof macrophage cytotoxicity against tumor cellsand invading pathogens (10, 11), and as a regu-lator of inflammatory responses (12, 13).
Adamantylamide dipeptide (AdDP) is a syn-thetic immunomodulatory substance from thegroup of desmuramyl peptides with minimalside effects (14). It was found to have hepatop-rotective activity (15) and to stimulate humoralimmunity (16). Previous work demonstrated theefficacy of a combined regimen of AdDP andPZQ in maintenance of the state of resistanceto reinfection with S. mansoni after primary in-fection eradication (17).
Bearing in mind that AdDP stimulates hu-moral immunity and that PZQ acts in synergywith the immune response, this study investi-gates the use of AdDP as adjuvant therapy topraziquantel in mice infected with praziquantel-insusceptible S. mansoni isolate in a trial to in-crease the susceptibility of this isolate to thedrug. A comparison with mice harboring pra-
481
ziquantel-susceptible S. mansoni isolate wasmade. The effect of this treatment regimen onliver disease severity and NO production wasalso examined.
MATERIALS AND METHODS
Animals and infectionMale C57BL/6 mice (18–20 g), bred and main-
tained at the Schistosome Biology Supply Center(SBSC), Theodor Bilharz Research Institute (TBRI),Giza, Egypt, were used in this study. Animals wereinfected (80∫10 cercariae/mouse) using the body im-mersion technique (18) with either PZQ-susceptible(CD-S) or PZQ-insusceptible (EE10-I) S. mansoniisolates. Susceptibility and insusceptibility to PZQwas considered on the basis of PZQ ED50 estimatedin hosts infected with the previously mentioned iso-lates where the drug ED50 was 77 mg/kg in suscep-tible and 236 mg/kg in insusceptible (19).
Drugs and dosagesPraziquantel (Distocide, Epico Pharmaceuticals,
Cairo, Egypt) was given orally as aqueous suspensionin 2% Cremophor-EL (Sigma Chemical Co., St.Louis, MO, USA) 7 weeks postinfection either in re-duced dose (100¿3 mg/kg) or in full curative dose(500¿2 mg/ kg) (20).
Adamantylamide dipeptide, (kindly provided byDr. Zdenek Zidek) was injected i.p. in a single doseof 5 mg/kg one day before infection with S. mansonicercariae.
Animal groupsEach of the two batches of C57 BL/ 6 mice infected
with either PZQ-susceptible (CD-S) or PZQ-insuscep-tible (EE10-I) S. mansoni isolate was divided into fivegroups 7 weeks postinfection. The experimental groupreceived AdDP (5 mg/kg) and PZQ in reduced dose(100 mg/kg ¿3) and the remaining four groups werecontrol groups. Three of them were treated controls,receiving AdDP in the same dose, PZQ in reduced andin full dose. The fourth control group was the infecteduntreated control receiving only the vehicle.
Parasitological studiesAfter sacrifice, animals were perfused (21) using a
MasterFlex pump (Cole-Parmer Instrument Com-pany). Worms from hepatic and portomesentericveins were collected and counted. Parts of both thelivers and intestines were removed and kept frozenfor subsequent study of tissue egg load (22). The re-maining parts of the small intestine were collectedin 2 ¿ saline to study percentage egg developmentalstages (23). Parts of the livers were fixed in 10% form-alin for subsequent preparation of hematoxylin andeosin (H&E) and Masson trichrome-stained sections

BOTROS et al.
to study the state of hepatic granuloma and associ-ated histopathological changes.
Biochemical and immunological parametersAlanine aminotransferase (ALT). Two weeks after
the last dose of PZQ, animals in all groups werekilled by decapitation and blood was collected.Serum was separated by centrifugation at 3000 ¿gfor 10 min and stored at ª70 æC for assay of serumALT (Boehringer reagent kit, Mannheim, Germany).
NO in culture of peritoneal macrophagesMice were killed by cervical dislocation and were in-
jected intraperitoneally with 8 ml of sterile saline. Peri-toneal cells were collected and pooled, washed, and re-suspended in complete RPMI 1640 culture medium(SEVAC, Prague CZ). The cells were counted andseeded into 96-well flat-bottom microplates (Costar,Cambridge, MA) in volumes of 100 ml (2¿106 ml). Ad-herent peritoneal cells (macrophages) were isolated byincubating the cells in 5% CO2 for 2 h at 37 æC. Theplates were then vigorously shaken and the wellswashed three times to remove non-adherent cells. Cul-tured cells were maintained in a 5% CO2 incubator at37 æC, in a humidified Heraeus incubator for 24 h. Allexperimental variants were set in triplicate. The con-centration of nitrites in the supernatants of the tripli-cate wells was taken as a measure of NO production.Supernate cell-free samples (50 ml) were incubated for10 min at 37 æC with an aliquot of Griess reagent. Theabsorbance at 540 nm was recorded using a microplatereader. A nitrite calibration curve was used to convertabsorbance into mM of nitrite (24).
Histopathological studiesGranuloma diameter and percentage fibrotic area.
Approximately half of the livers were collected andimmediately fixed in 10% formalin and embedded inparaffin. Histological sections were processed andstained with hematoxylin and eosin (H&E) and Mas-son trichrome. Diameters of the granuloma (30/mouse) were measured using an ocular micrometer.Only granuloma around a single egg containing a mi-racidium was measured. Egg viability was assessedmicroscopically in the same liver sections and cellcomposition of granulomas was also investigated.
The collagen content of the liver tissue sections wasevaluated after staining with picrosirius red (Histo-Path of America). Semiquantitation of percentageareas of collagen fibers using computer-controlledImage Analysis (Kontrone Image Analysis System)was conducted. This essentially consists of a com-puter-controlled microscope (Zeiss axioskope, Ger-many), video camera, two monitors, computer unit(P.C., IBM compatible, 486D¿100) and a desk jetcolored printer. Image analysis was performed usingthe software program KS 400. Assessment of thetotal areas of fibrosis was analyzed, in five consecu-tive microscopic fields, at ¿100 magnification.
482
Immunohistochemical detection of iNOS and CD68antigens
Standard avidin-biotin immunoperoxidase tech-nique was used (25). Paraffin sections (5 mm in thick-ness) were dewaxed in xylene and hydrated in de-scending grades of ethanol. Endogenous peroxidaseactivity was quenched by incubation in 3% hydrogenperoxide in 100% methanol for 20 min. Antigen re-trieval was performed by microwaving of the sectionsin citrate buffer (PH 6.0) for 15 min at 700 W. Sec-tions were incubated overnight at 4 æC with the pri-mary antibodies against iNOS (Santa Cruz Biotech-nology, USA) and CD68 mouse anti-macrophagesmonoclonal antibody (DAKO, Denmark). Both anti-bodies were diluted 1:50 in PBS. After further rinsingin PBS, sections were incubated at room temperaturefor 15 min with biotinylated secondary anti-mouseantibody (DAKO, Denmark) and after a furtherwash in PBS, the slides were incubated with avidin-biotin complex horseradish peroxidase solution(DAKO, Denmark). After 10 min incubation, theperoxidase reaction was developed with 0.01% hydro-gen peroxide in 0.05% diaminobenzidine tetrahy-drochoride. Sections were counterstained with hema-toxylin and dehydrated in methanol prior tomounting.
Liver sections with voluntary omission of the pri-mary antibody and its replacement by PBS served asnegative control. iNOS-positive controls were sec-tions of colonic cancer specimen, where macrophagesand neutrophils labeling very strongly in every slideserved as an internal positive control.
Evaluation of immunostainingSections were examined by light microscopy
(¿400) for both iNOS and CD68 markers; two fea-tures of the immune reaction were assessed separatelyon a semiquantitative basis:
1. The extent of staining is the mean of the percen-tage of positively stained cells. The number ofpositively stained cells with the highest expressionwas recorded in five consecutive fields and the finalvalue represents the mean. Zero percentage wasgiven to unstained sections. The patterns of bothiNOS and CD68 expressions were cytoplasmicstaining (26).
2. The intensity of staining was evaluated relative tonormal liver section expression where no expres-sion for CD68 and iNOS was recorded. The inten-sity of the immunostaining was designated asnegative (ª), mild or weak (1), moderate (2), andmarked or strong (3). Negative staining was de-fined as the absence of cells expressing the marker.
Statistical analysisStatistical analysis was performed using the SPSS
9 software program. Data were summarized as meansfor values and percentages. Means of groups were

IMMUNOMODULATORY DRUG AS ADJUVANT THERAPY IN S. MANSONI
compared using unpaired t-test. For comparing morethan two groups, ANOVA test was used. The meandifference was significant at p�0.05.
RESULTS
Parasitological parametersWorms were recovered from mice infected with
PZQ-susceptible (CD-S) S. mansoni isolate insignificantly higher numbers (24.2∫1.3) thanthose recovered from mice infected with PZQ-in-susceptible (EE10-I) S. mansoni isolate (20.25∫0.82). Meanwhile, the total tissue egg load wassignificantly higher in the group infected with thePZQ-susceptible (38.84∫1.93) vs PZQ-insuscep-tible S. mansoni isolate (28.56∫1.30).
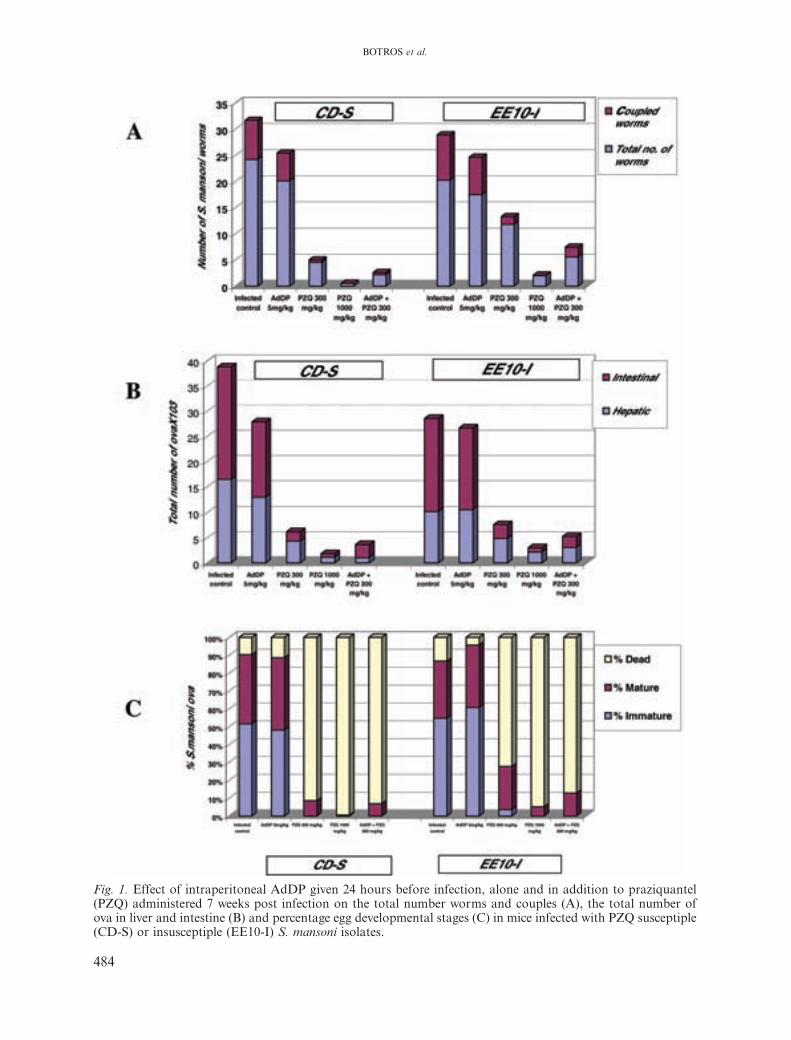
In mice infected with the PZQ-susceptibleand -insusceptible S. mansoni isolates and com-pared to infected untreated controls, intraper-itoneal (i.p) injection of AdDP alone did notsignificantly reduce the total number of worms(Fig. 1A). A significant reduction was recordedwhen AdDP was combined with PZQ in re-duced dose in mice infected with CD-S (91% &93%) and EE10-I (72% & 82%) S. mansoni iso-lates. In CD-S S. mansoni-infected mice, the re-duction in worm and egg loads was comparableto that in mice treated with full dose of PZQ(98% & 95%) and was significantly higher thanin mice treated with PZQ in reduced dose(81% & 84%). In the group infected with EE10-I S. mansoni isolate, the profile of reduction inworm and egg loads of mice receiving combinedAdDP and PZQ was almost the same as thatrecorded in mice infected with the CD-S re-ceiving the same treatment, but to a lesser ex-tent (72% & 82%). The reduction recorded wassignificantly less than in mice treated with fulldose PZQ (91% & 89%), but was significantlyhigher than that recorded in mice receiving re-duced dose (42% & 73%) of PZQ.
Concerning percentage egg developmentalstages, mice infected with the PZQ-susceptibleS. mansoni isolate and treated with AdDP inaddition to PZQ showed a higher percentage ofdead eggs (93.1%), yet this was not significantlydifferent from mice receiving PZQ in reduceddose alone (91%), The same profile was ob-served in mice infected with PZQ-insusceptibleS. mansoni isolate treated with AdDP in ad-dition to PZQ in reduced dose, where there was
483
a significantly higher number of dead eggs(87%) than was seen in mice receiving PZQ inreduced dose alone (72%), yet the difference wasstatistically significant. Meanwhile, in the groupreceiving combined treatment, all immature eggstages disappeared, compared to 4% immatureeggs in mice receiving PZQ in reduced dosealone (Fig. 1B).
Histopathological parametersGranuloma diameter and percentage fibrotic
area. All treatment regimens reduced granu-loma diameter significantly when compared toinfected untreated control (p�0.05) whether theanimals were infected with CD-S or EE10-I S.mansoni isolate. In both hosts infected withCD-S or EE10-I, S. mansoni isolate and com-pared to infected untreated controls, the highestreductions in granuolma diameters and percen-tage fibrotic areas recorded were in the groupstreated with combined AdDP and praziquantel.The reduction recorded in granuloma diametersand percentage fibrotic areas in both CD-S(21% & 59%) and EE10-I (18% & 64%) infectedgroups was comparable to that in mice receivingPZQ in full dose whether the animals were in-fected with CD-S (21% & 51%) or EE10-I(17% & 59%) S. mansoni isolate. Meanwhile, thepercentage degenerated eggs in the same groupswas comparable to that in mice receiving PZQin full dose (Table 1).
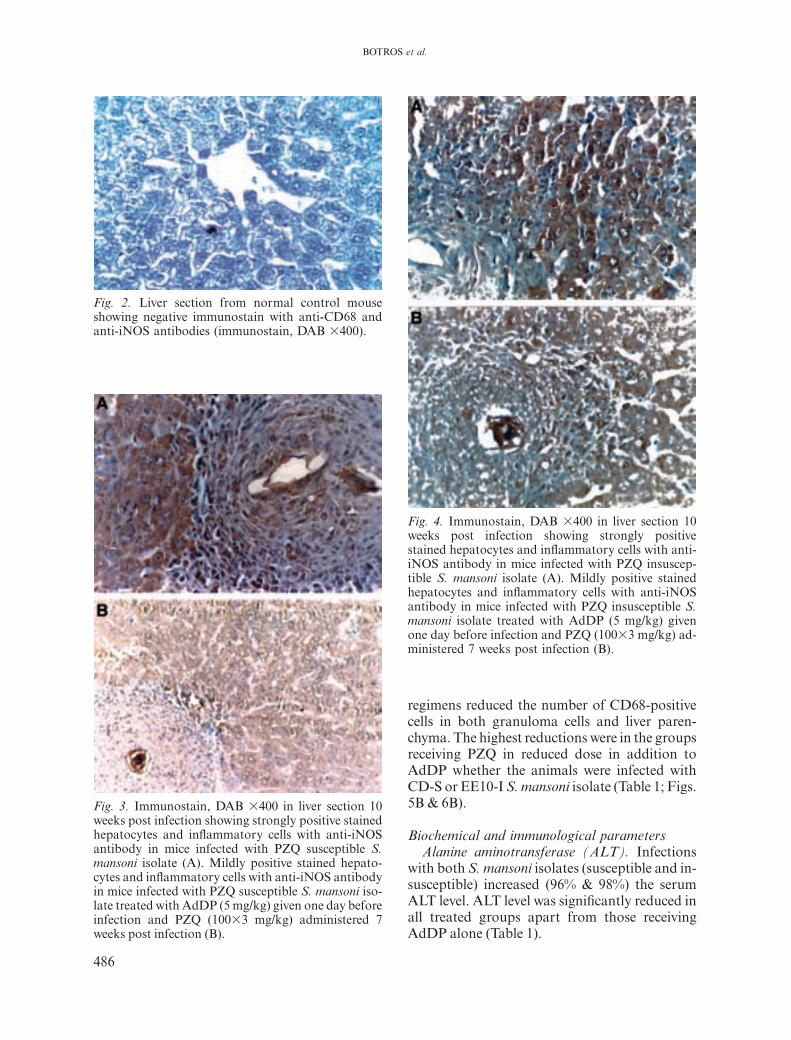
iNOS and CD68-positive cellsNormal hepatocytes were negative for iNOS
stain (Fig. 2). In S. manson-infected mice, iNOSwas expressed as brownish intracytoplasmicgranules in the hepatocytes and in the inflam-matory cells around the granuloma as well asthe sinusoidal and endothelial cells. In bothhosts infected with the PZQ-susceptible and -insusceptible S. mansoni isolates, the best ex-pression of iNOS was in the cytoplasm ofhepatocytes (Figs. 3A & 4A). All treatment regi-mens reduced iNOS in granuloma cells andhepatocytes. The highest reduction was ingranuloma cells of mice treated with PZQ in fulldose whether the animals were infected withCD-S (26.1∫2.0) or EE10-I (29.2∫2.1) S. man-soni isolate. Meanwhile, in EE10-I S. mansoni-infected animals the reduction in hepatocyteiNOS recorded in mice treated with AdDP inaddition to PZQ in reduced dose was compar-
BOTROS et al.
Fig. 1. Effect of intraperitoneal AdDP given 24 hours before infection, alone and in addition to praziquantel(PZQ) administered 7 weeks post infection on the total number worms and couples (A), the total number ofova in liver and intestine (B) and percentage egg developmental stages (C) in mice infected with PZQ susceptiple(CD-S) or insusceptiple (EE10-I) S. mansoni isolates.
484
IMMUNOMODULATORY DRUG AS ADJUVANT THERAPY IN S. MANSONI
TABLE 1. Effect of adamantylamide dipeptide (AdDP), alone and in addition to praziquantel (PZQ), on serumalanine aminotransferase (ALT) and hepatic histopathological changes in C57BL/6 mice infected with the PZQ-
susceptible (CD-S) and -insusceptible (EE10-I) S. mansoni isolates 10 weeks postinfection (nΩ7–10)
Hepatic granuloma Hepatic number of CD68π cells
Dose Diameter % Fibrotic % DegeneratedAnimal groups mg/kg ALT (U/L) (mm) areas ova Granuloma Parenchyma
Normal control (8) 63.13∫3.94 – – –Infected control (9) S 123.56∫3.34. 258.13∫3.47 16.47∫1.3 17.33∫1.42 36.66∫2.64 29.44∫2.54
I 124.75∫3.39. 245.4∫3.52* 19.72∫0.92 17.37∫1.61 35.00∫2.11 28.75∫2.45AdDP (7) S 5 117.43∫3.30.a 222.85∫2.86* 11.12∫1.88* 60.71∫4.14* 31.42∫2.83* 21.43∫2.86**
I 118.88∫4.02.a 228.12∫2.97* 11.72∫1.45* 55.25∫6.53* 32.5∫1.88 22.50∫1.88*PZQ (9) S 100¿3 83.89∫2.59* 209.44∫5.08* 11.35∫0.33* 66.66∫3.43* 10.33∫0.68** 7.56∫0.73**
I 84.55∫2.36* 212.00∫3.95* 11.22∫0.82* 65.00∫3.49* 9.8∫1.21** 7.40∫0.94**PZQ (9) S 500¿2 72.67∫3.21* 205.00∫11.00* 8.02∫0.87* 78.88∫1.62* 13.88∫1.11** 10.44∫0.74**
I 76.42∫2.93* 202.91∫2.08* 8.03∫0.55* 72.08∫2.57* 16.08∫1.21** 11.08∫0.77**AdDP & PZQ (10) S 5 79.2∫2.66* 204.06∫3.39*c 6.69∫0.68*c,a 73.00∫3.08*c 9.80∫1.5**d,b,e 8.70∫0.82**d
I 100¿3 80.14∫3.06* 200.08∫2.29*a,c 7.10∫0.77*b,c 72.85∫2.64*a 9.71∫1.91**c,b,e 7.85∫0.50**c
SΩPZQ-susceptible S. mansoni isolate. IΩPZQ-insusceptible S. mansoni isolate.Values are means∫SE. AdDP was given intraperitoneally 24 h before infection and PZQ was given orally 7 weeks postinfection. () numberof animals/group. Significant difference versus infected control at *p�0.05 and **p�0.01, versus PZQ (100¿3 mg/kg) at ap�0.05 andbp�0.01, versus AdDP (5 mg/ kg mg/kg) at cp�0.05 and dp�0.01, and versus PZQ (500¿2 mg/kg) at ep�0.05. % fibrotic area was assessedin five successive fields (¿100) and % degenerated ovaΩno. of degenerated eggs ¿100/total eggs in 30 granulomata.
able to that in the group treated with PZQ infull dose (Figs. 3B & 4B). The reduced expres-sion of iNOS in granuloma cells and hepato-cytes was more manifest (p�0.05) in animals in-fected with PZQ-insusceptible than in animalsinfected with PZQ-susceptible S. mansoni iso-lates (Table 2).
TABLE 2. Effect of adamantylamide dipeptide (AdDP), alone and in addition to praziquantel (PZQ), on theexpression of iNOS in hepatic granuloma and hepatocytes and nitric oxide in culture of peritoneal macrophagesin C57BL/6 mice infected with PZQ-susceptible (CD-S) and -insusceptible (EE10-I) S. mansoni isolates 10
weeks post infection (nΩ7–10)
NO in culture of peritonealHepatic iNOS (no. of cells) macrophages
% Change fromDose corresponding
Animal groups mg/kg Granuloma Hepatocytes infected controlNormal control (8) – 2.95∫0.68 –Infected control (9) S 70.55∫3.86 80.00∫3.73 11.9∫2.28. –
I 64.37∫3.71 73.75∫2.79 15.3∫0.41. –AdDP (7) S 5 49.28∫4.14* 58.57∫4.72** 7.50∫2.29. 36.9
I 51.87∫4.42* 63.12∫4.99 8.03∫0.66.* 47.6PZQ (9) S 100¿3 34.44∫3.86** 32.77∫4.79** 6.40∫0.94.* 46.2
I 31.00∫4.87** 33.5∫3.54** 11.56∫1.97. 24.4PZQ (9) S 500¿2 26.11∫2.01** 28.88∫2.17** 4.68∫0.96* 60.7
I 29.16∫2.11** 31.66∫1.97** 8.31∫1.43.* 45.7AdDPπ S 5 37.50∫3.09**c,e 35.5∫1.66**d 2.21∫0.61*a,c 81.4PZQ (10) I 100¿3 42.14∫1.84**e,a 39.28∫5.71**c 5.80∫0.70.,a* 62.1SΩPZQ-susceptible S. mansoni isolate. IΩPZQ-insusceptible S. mansoni isolate. Values are means∫SE.AdDP was given intraperitoneally 24 h before infection and PZQ was given orally 7 weeks postinfection. ()number of animals/group. Significant difference versus infected control at *p�0.05 and **p�0.01, versusPZQ (100¿3 mg/kg) at ap�0.05 and bp�0.01, versus AdDP (5 mg/ kg mg/kg) at cp�0.05 and dP�0.01,and versus PZQ (500¿2 mg/kg) at ep�0.05 and fp�0.01.
485
CD68-positive cells in granuloma cells andliver parenchyma were comparable in CD-S andEE10-I S. mansoni-infected animals; they appearas a brownish cytoplasmic color staining themacrophages, which were distributed through-out the granuloma but with greater localizationat the periphery (Figs. 5A & 6A). All treatment
BOTROS et al.
Fig. 2. Liver section from normal control mouseshowing negative immunostain with anti-CD68 andanti-iNOS antibodies (immunostain, DAB ¿400).
Fig. 3. Immunostain, DAB ¿400 in liver section 10weeks post infection showing strongly positive stainedhepatocytes and inflammatory cells with anti-iNOSantibody in mice infected with PZQ susceptible S.mansoni isolate (A). Mildly positive stained hepato-cytes and inflammatory cells with anti-iNOS antibodyin mice infected with PZQ susceptible S. mansoni iso-late treated with AdDP (5 mg/kg) given one day beforeinfection and PZQ (100¿3 mg/kg) administered 7weeks post infection (B).
486
Fig. 4. Immunostain, DAB ¿400 in liver section 10weeks post infection showing strongly positivestained hepatocytes and inflammatory cells with anti-iNOS antibody in mice infected with PZQ insuscep-tible S. mansoni isolate (A). Mildly positive stainedhepatocytes and inflammatory cells with anti-iNOSantibody in mice infected with PZQ insusceptible S.mansoni isolate treated with AdDP (5 mg/kg) givenone day before infection and PZQ (100¿3 mg/kg) ad-ministered 7 weeks post infection (B).
regimens reduced the number of CD68-positivecells in both granuloma cells and liver paren-chyma. The highest reductions were in the groupsreceiving PZQ in reduced dose in addition toAdDP whether the animals were infected withCD-S or EE10-I S. mansoni isolate (Table 1; Figs.5B & 6B).
Biochemical and immunological parametersAlanine aminotransferase (ALT). Infections
with both S. mansoni isolates (susceptible and in-susceptible) increased (96% & 98%) the serumALT level. ALT level was significantly reduced inall treated groups apart from those receivingAdDP alone (Table 1).

IMMUNOMODULATORY DRUG AS ADJUVANT THERAPY IN S. MANSONI
Fig. 5. Immunostain, DAB ¿200 in liver section 10weeks post infection showing CD68 positive cells atthe periphery of liver granuloma in mice infectedwith PZQ susceptible S. mansoni isolate (A). Reducednumber of CD68 positive cells at the periphery ofliver granuloma in mice infected with PZQ suscep-tible S. mansoni isolate treated with AdDP (5 mg/kg)given one day before infection and PZQ (100¿3 mg/kg) administered 7 weeks post infection (B).
NO in culture of peritoneal macrophages. NOproduction in culture of peritoneal macro-phages was increased upon S. mansoni infection,whether this infection was with CD-S or EE 10-I, S. mansoni isolate. All treatment regimens sig-nificantly reduced the NO level in culture ofperitoneal macrophages. The highest reductionwas in mice treated with AdDP in addition toPZQ in reduced dose whether the animals wereinfected with CD-S or EE10-I S. mansoni isolate(81.4% & 62.1%, respectively). This reductionwas not significantly different from that re-corded in the groups receiving full dose of PZQ(60.7% & 45.7%, respectively) (Table 2).
487
Fig. 6. Immunostain, DAB ¿400 in liver section 10weeks post infection showing CD68 positive cells atthe periphery of liver granuloma in mice infectedwith PZQ insusceptible S. mansoni isolate. (A). Re-duced number of CD68 positive cells at the peripheryof liver granuloma in mice infected with PZQ insus-ceptible S. mansoni isolate treated with AdDP (5 mg/kg) given one day before infection and PZQ (100¿3mg/kg) administered 7 weeks post infection (B).
DISCUSSION
In this work S. mansoni worm and tissue eggloads recovered from mice infected with PZQ-susceptible ‘‘CD-S’’ S. mansoni isolate were sig-nificantly higher than parallel values in mice in-fected with PZQ-insusceptible EE10-I S. man-soni isolate, suggesting a compromised biologi-cal development for the PZQ-insusceptible S.mansoni isolate. Similar findings were reportedby others who suggested that there may be somebiological cost associated with the relative in-sensitivity to PZQ (19, 27).
As regards infected treated animal groups,AdDP was given in a dose of 5 mg/kg i.p. one daybefore S. mansoni infection. A dose comparable(100 mg/mouse) to that tested in this work when

BOTROS et al.
given i.p. increased ovalbumin-specific antibodytiters when AdDP was tested as adjuvant (28).Time of injection of immunomodulatory drugsin relation to infection decides the fate of thesedrugs (29). Injection before sensitization by theinvading cercariae proved successful in main-taining a state of resistance to reinfection in amodel of murine schistosomiasis (17). AdDPalone slightly reduced the total number of S.mansoni worms and eggs, yet the reduction wasonly significant in the egg load. The reduction intissue egg and worm loads in mice treated withPZQ in its full dose was within the range reportedby other authors (30, 31), and it was significantlyhigher than that in mice receiving reduced doseof PZQ. Combined treatment with AdDP andPZQ in reduced dose enhanced the killing effectof PZQ on worms and consequently the egg loadwas less. The efficacy of this treatment protocolwas significantly higher than that recorded inmice treated with PZQ in reduced dose whetherthe animals were infected with PZQ- susceptibleor -insusceptible S. mansoni isolate, and wascomparable to that in the group receiving PZQ infull dose in mice infected with the PZQ-suscep-tible S. mansoni isolate. An immunostimulatoryactivity of AdDP on humoral immune responseof the host has been reported (14). Meanwhile,PZQ is known to act in synergy with the immuneresponse of the host (32). PZQ induces tegu-mental damage of adult worms (33) and disruptsthe integrity of the worm surface membrane (34),which is a target of antibody attack (35). At thisstage, AdDP, an immunostimulant of the hu-moral immunity of the host, may be enhancingthe antibody interaction at the surface of the ex-posed, damaged schistosome tegument as a re-sult of treatment with PZQ (36). Meanwhile, thedrug may be activating B cells to secrete specificantibodies, IgG & IgE, leading to sensitization ofneutrophils, macrophages, platelets and eosino-phils, which are known to attack the wormsthrough an antibody-dependent cell-mediatedcytotoxicity mechanism (37).
Concerning hepatic granulomata in mice re-ceiving combined treatment with AdDP andPZQ in reduced dose, they were significantlysmaller in diameter and percentage fibroticareas with higher percentage of degeneratedeggs. The macrophages forming the granuloma-expressing CD68 antibody were more in theouter zone of the granuloma; they expressed
488
CD68 antibody with the same intensity allthrough the granuloma but with greater localiz-ation at the periphery. This pattern may reflectthe distribution of phagocytes, particularly acti-vated ones within the granuloma (38). Numberof CD68-positive cells was significantly less inall treated groups than in infected untreatedcontrols, yet was least in the group receivingPZQ in reduced dose in addition to AdDPwhether the animals were infected with CD-Sor EE10-I S. mansoni isolate. Zidek et al. (39)reported that AdDP is able to induce mild andfully reversible paw edema in experimental ani-mals. The role played by AdDP in modulatingthe interplay between stimulating cytokines andtheir cellular sources has been reported (40). Inthis work, granulomata were reduced in sizewith fewer CD68-positive cells, a main constitu-ent of the inflammatory granulomatous reac-tion. It may be suggested that treatment withthe immunomodulatory AdDP and PZQ wouldhave induced faster downregulation of the in-flammatory granulomatous reaction. This re-duction in the size of the hepatic granulomatawas recorded hand in hand with significantlylower levels of NO in culture of peritonealmacrophages compared to the increased levelsrecorded in S. mansoni-infected untreated mice.Peak levels of NO in sera by Ag-stimulatedsplenocytes upon infection with S. mansoni werereported to be between days 37 and 45 of infec-tion ‘‘coincident with egg deposition’’ and thaton day 70 of infection NO levels returned tolevels comparable to that in naıve uninfectedmice (4). In this work, NO in culture of peri-toneal macrophages showed a significant in-crease 9 weeks postinfection (63 days postinfec-tion), which may suggest that levels of NO notonly depend on the initial insult but also on thesource, which may reflect different productionand period rates (4). The increase of NO as aresult of infection has been suggested to be asort of protection at the time the liver is firstexposed to eggs (41–45). Brunet et al. (4) re-ported that, absence of NO may lead to severemorbidity with increased hepatocellular damagecharacterized by hepatocyte apoptosis and in-creased hepatocellular enzyme release. More-over, in the course of liver regeneration, for ex-ample after partial hepatectomy, NOS2 is ex-pressed at moderate levels and contributes tothe inhibition of apoptosis, favoring pro-

IMMUNOMODULATORY DRUG AS ADJUVANT THERAPY IN S. MANSONI
gression in the cell cycle until organ size andfunction are restored (46).
In this work, hepatocytes showing positivityfor iNOS recorded around the central veins,periportal area and in the inflammatory cellsforming granuloma (lymphocytes and macro-phages) supports the view that NO is made inreasonable quantity within the liver itself (4) asa result of exposure to parasite eggs and thatiNOS mRNA transcripts are upregulated andcell recruited to form granulomas aroundtrapped eggs. Mohammed et al. (47) reportednegative iNOS in livers of the control groupwith high expression in the livers from the cir-rhotic group. iNOS was seen mainly in the in-flammatory cells infiltrating the portal tracts,blood monocyte-like cells, hepatocytes, sinus-oidal cells and endothelial cells.
The modulated hepatic granulomas in treatedmice with significantly less NO in culture ofperitoneal macrophages, iNOS in hepatocytesand granulomas may support the notion thatthese treatment regimens favor faster downregu-lation of the inflammatory granulomatous reac-tion, making NO no longer required to protecthepatocytes. In the group receiving treatmentswith both AdDP and PZQ in reduced doseshowing the least NO levels, alanine amino-transaminase showed the best tendency to nor-malization. This was contrary to the elevatedlevels recorded in S. mansoni-infected animals.It has been stated that NO can have cytoprotec-tive or cytotoxic effects; hence it is the balancebetween NO and other inflammatory cytokinemediators that is crucial in determining its ef-fect on the host (4). Findings from this worksupport the view that it may be playing a cyto-protective role at time of egg deposition andthat if one interferes with the proper chemo-therapeutic agent at the proper time, thus con-trolling the inflammatory reaction, then it is nolonger needed. This is because, despite the lowNO levels recorded, together with the modu-lated inflammatory granulomatous reaction, in-cluding one of its main cell constituents, themacrophages, liver function showed a tendencyto normalization. Compared to infected con-trols, animals treated with combined AdDP andPZQ in reduced dose showing the lowest NOlevel in culture of peritoneal macrophages;iNOS expression was not diminished to thesame extent in granuloma or hepatocytes. These
489
results may be due to the presence of differentisoforms of inducible NOS [iNOS, NOS2] (7).
In CD-S and EE10-I S. manson-infected micetreated with AdDP and PZQ in reduced dose,the antischistosomal efficacy obtained was sig-nificantly higher than that in mice receiving re-duced dose of PZQ, showing that AdDP re-duced the effective dose of PZQ. This efficacy –obtained together with granuloma diameter re-duction, diminution in percentage fibrotic areasand NO level in cultured peritoneal macro-phages – was comparable to that in mice re-ceiving full dose of PZQ. Moreover, CD68-posi-tive cells were significantly fewer than in micetreated with full dose of PZQ, revealing fewestassociated histopathological changes. Thesefindings may stimulate further studies on theuse of a cocktail of drugs as long as there are noalternatives to the presently and solely availableantischistosomal.
REFERENCES
1. Stadecker MJ. The role of T cell energy in theimmunomodulation of schistosomiasis. ParasitolToday 1992;8:199–204.
2. Yamashita T, Boros DL. IL-4 influences IL-2production and granulomatous inflammation inmurine S. mansoni. J Immunol 1992;149:3659–64.
3. Pearce EL, Cspar PJM, Lewis FA, Sher A. Downregulation of Th1 cytokine production accom-panies induction of Th2 responses by a parasitichelminth, S. mansoni. J Exp Med 1991;173:159–69.
4. Brunet LR, Beall M, Dunne DW, Pearce EJ. Ni-tric oxide and the Th2 response combine to pre-vent severe hepatic damage during schistosomalmansoni infection. J Immunol 1999;163:4976–84.
5. Michel T, Feron O. Nitric oxide synthase: which,where, how and why? J Clin Invest 1997;100:2146–52.
6. Griffith OW, Stueher DJ. Nitric oxide synthaseproperties and catalytic mechanism. Annu RevPhysiol 1995;57:707–36.
7. Forstermann U, Kleinert H, Gath I, Schwarz P,Closs E, Dun N. Expression and expressionalcontrol of nitric oxide syntheses in various celltypes. Adv Pharmacol 1995;34:171–4.
8. Nathan C. Inducible nitric oxide synthesis: whatdifference does it make? Review. J Clin Invest1997;100:2417–23.
9. Spitzer JS. Cytokine stimulation of nitric oxideformation and differential regulation in hepato-

BOTROS et al.
cytes and nonparenchymal cells of endotoxemicrats. Hepatol 1994;19:217–28.
10. Nathan C. Nitric oxide a secretory product ofmammalian cells. FASEB J 1992;6:3051–64.
11. Fang F. Mechanisms of nitric oxide related anti-microbial activity. J Clin Invest 1997;99:2818–22.
12. Lyons CR. The role of nitric oxide in inflam-mation. Adv Immunol 1995;50:323–71.
13. Bingisser RM, Tilbrook PA, Holt PA, Kees RU.Macrophage-derived nitric oxide regulates T cellactivation via reversible disruption of the Jak3/STAT5 signaling pathway. J Immunol 1998;160:5729–34.
14. Masek K, Seifert J, Flegel M, Krojidlo M, Kol-insky J. The immunomodulatory property of anovel synthetic compound adamantylamide di-peptide. Methods Find Exp Clin Pharmacol 1984;6:667–9.
15. Farghali H, Buchar E, Jiricka Z, Kamaenikova L,Janku I, Masek K. The hepatoprotective effect ofa new immunomodulator adamntylamide dippep-tide on carbon tertrachloride toxicity in rats.Highlights Modern Biochem 1989;2:1539–42.
16. Zidek Z, Louckova M, Sula K, Nouza K, MasekK. Effects of adamantylamide dipeptide (AdDP)on humoral immunity in mice. Int J Immunother1991;8:15.
17. Botros S, Mahmoud S, Hassan S, Ibrahim A, Zi-dek Z, Masek K. Effect of adamantylamide di-peptide on reinfection resistance after primaryinfection eradication in experimental schistoso-miasis mansoni. Arzneimitt Forsch Drug Res1996;46:74–8.
18. Liang YS, John BI, Boyed DA. Laboratory culti-vation of schistosome vector snails and mainten-ance of schistosome life cycles. Proc of the FirstSino-American Symposium 1987;1:34–48.
19. Hanallah S, El Lakkany N, Mahmoud S, MousaM, Botros S. Altered immunoglobulin isotypeprofile and anti-immature worm surface immuno-globulins in mice harboring a praziquantel-resis-tant Schistosoma mansoni isolate. APMIS 2003;111:1125–32.
20. Gonnert R, Andrews P. Praziquantel, a newbroad-spectrum antischistosomal agent. Zeit fürParasitenk 1977;52:129–50.
21. Duvall RH, DeWitt WB. Technique for re-covering adult schistosomes from laboratory ani-mals. Am J Trop Med Hyg 1967;16:438–86.
22. Kloetzel K. A suggestion for the prevention ofsevere clinical forms of schistosomiasis mansoni.Bull World Health Organ 1967;37:686–7.
23. Pellegrino J, Oliveria CA, Faria J, Cunha A. Newapproach to the screening of drugs in experimen-tal Schistosoma mansoni in mice. Am J Trop MedHyg 1962;11:201–15.
24. Tracey WR, Tse J, Carter G. Lipolysaccharide-induced changes in plasma nitric and nitric con-centration in rats and mice: pharmacological
490
evaluation of nitric oxide synthase inhibitors. JPhamacol Exp Ther 1995;272:1011–5.
25. Hsu SM, Raine L. Protein A, avidin and biotinin immunohistochemistry J Histochem Cyto-chem 1981;29:1349–53.
26. Vakkala M, Kahlos K, Lakari E, et al. Induciblenitric oxide synthase expression, apoptosis, andangiogenesis in in situ and invasive breast carci-nomas. Clin Cancer Res 2000;6:2408–16.
27. William S, Sabra A, Ramzy F, Mousa M, Demer-dash Z, Bennett L, Day TA, Botros SS. Stabilityand reproductive fitness of schistosomiasis man-soni isolate with decreased sensitivity to pra-ziquantel. Int J Parasitol 2001;31:1093–100.
28. Becker PD, Corral RS, Gurman CA, GrinsteinS. Adamantylamide dipeptide as effective immu-noadjuvant in rabbits and mice. Vaccine 2001;19:4603–9.
29. Perper RJ, Wood D, Zimmerman M. Immuno-pharmacology. In: Rosenthale ME, Mansmann,HC, eds. Spectrum Publications, Inc.: New York,1975, pp. 15–29.
30. Giboda M, Smith JM, Prichard RK. Reductionin tissue egg load and maintenance of resistanceto challenge in mice infected with Schistosomamansoni, following combined treatment withpraziquantel and an antifibrotic agent. Ann TropMed Parasitol 1994;88:385–95.
31. Mahmoud MR, Botros SS. Artemether as adju-vant therapy to praziquantel in murine Egyptianschistosomiasis mansoni. J Parasitol 2005;91:175–8.
32. Piper KP, Mott RF, Hockley DL, Mc Laren DJ.A new view of PGF regulation of the immuneresponse. Immunol Today 1991;12:349–52.
33. Fallon PG, Fooker RE, Whartin GA. Temporaldifferences in praziquantel- and oxamniquine-in-duced tegumental damage to adult Schistosomamansoni: implications for drug-antibody synergy.Parasitology 1996;112:47–58.
34. Fallon PG, Hamilton JV, Doenhoff MT. Efficacyof treatment of murine S. mansoni infection withpraziquantel and oxaminquine correlate with in-fection intensity : role of host antibody. Parasit-ology 1994;111:59–66.
35. Fallon PG, Cooper RO, Robert AJ, Doenhoff MT.Immune-dependent chemotherapy of schistoso-miasis. Parasitology 1992;105(Suppl);541–48.
36. Brindly PJ, Strand M, Norden AP, Sher A. Roleof host antibody in chemotherapeutic action ofpraziquantel against S. mansoni: identification oftarget antigens. Mol Biochem Parasitol 1989;34:99–108.
37. Dunne DW, Butterworth AE, Fulford AJ, et al.Immunity after treatment of human S mansoni:association between IgE antibodies to adultworm antigen and resistance to reinfection. EurJ Immunol 1995;22:1483–94.
38. El-Ahwany EG, Hanallah SB, Zada S, El Ghor-

IMMUNOMODULATORY DRUG AS ADJUVANT THERAPY IN S. MANSONI
ab NM, Badir B, Badawy A. Immunolocaliz-ation of macrophage adhesion molecule-1 andmacrophage inflammatory protein-1 in schistoso-mal soluble egg antigen-induced granulomatoushyporesponsiveness. Int J Parasitol 2000;30:837–42.
39. Zidek Z, Masek K, Frankova D, Flegel M. T-cell-dependent immunobiological activity of a de-smuramyl analogue of muramyl dipeptide, ada-mantylamide dipeptide (AdDP), and D-isogluta-mine. Int J Immunopharmacol 1993;15:631–7.
40. Vacek A, Hofer M, Weiterova L, Hoverova Z,Pipalova I, Masek K. Hemopoiesis-stimulatingaction of adamantylamide dipeptide: kinetics ofincrease of GM-CFC in femur and co-stimulat-ing activity of serum, role of bone marrowstromal cells. Immunopharmacol Immunotoxicol2001;23:505–17.
41. Kim Y, Talanian RV, Billar TR. Nitric oxide in-hibits apoptosis by preventing increase in caspa-se-3 like activity via two distinct mechanisms. JBiol Chem 1997;49:31138–48.
42. Harbrecht BG, Billiar TR, Stadler J, DemetrisAJ, Ochoa J, Curran RD, Simmons RL. Inhi-bition of nitric oxide synthesis during endotoxe-
491
mia promotes intrahepatic thrombosis and anoxygen radical-mediated hepatic injury. J LeukocBiol 1992;52:390–4.
43. Wynn TA, Eltoum I, Cheever AW, Lewis FA,Gause WC, Sher A. Analysis of cytokine mRNAexpression during primary granuloma formationinduced by eggs of Schistosoma mansoni. J Im-munol 1993;151:1430–40.
44. Wink D, Cook J, Pacelli R, Liebmann J, KrishnaM, Mitchell J. Nitrogen oxide protects againstcellular damage by reactive oxygen species. Re-view. Toxicol lett 1995;221:82–3.
45. McCormick M, Metwali A, Railsback M, Wein-stock J, Britigan B. Eosinophils from schisto-some-induced hepatic granulomas produce su-peroxide and hydroxyl radical. J Immunol 1996;157:5009–15.
46. Martin Sanz P, Horetelano S, Callejas NA, Gor-en N, Casado M, Zeini M, Bosca I. Nitric oxidein liver inflammation and regeneration. MetabBrain Dis 2002;17:325–34.
47. Mohammed NA, Abd El-Aleem S, Appleton I,Maklouf MM, Said M, McMahon RF. Expres-sion of nitric oxide synthase isoform in humanliver cirrhosis. J Pathol 2003;200:647–55.
Related Documents











