Journal of Plant Ecology VOLUME 9, NUMBER 4, PAGES 402–409 AUGUST 2016 doi:10.1093/jpe/rtv071 Advance Access publication 5 November 2015 available online at www.jpe.oxfordjournals.org © The Author 2015. Published by Oxford University Press on behalf of the Institute of Botany, Chinese Academy of Sciences and the Botanical Society of China. All rights reserved. For permissions, please email: [email protected] Ecological succession following forest decline in a xeric oak forest of south-central United States Devin P. Bendixsen 1,2, *, Stephen W. Hallgren 1 and Jesse A. Burton 1,3 1 Department of Natural Resource Ecology and Management, Oklahoma State University, 008 Ag Hall, Stillwater, OK 74074, USA 2 Present address: Department of Biomolecular Sciences, Boise State University, 1357 S. Brady St., Boise, ID 83725, USA 3 Present address: National Park Service, 2680 Natchez Trace Pkwy, Tupelo, MS 38804, USA *Correspondence address. Department of Natural Resource Ecology and Management, Oklahoma State University, 008 Ag Hall, Stillwater, OK 74074, USA. Tel: +1 575 725 0163; Fax: +1 208 392 1430; E-mail: [email protected] Abstract Aims The loss of canopy trees associated with forest decline can greatly influence the species composition and structure of a forest and have major impacts on the ecosystem. We studied the changes in forest composition and structure 1 and 5 years following nearly total canopy mortality on several hundreds of hectares of xeric oak forests in south-central United States. Because the forests were within an ecotonal vegetation type composed of a mosaic of forest, savanna and grassland, we sought to learn whether forest decline areas would recover to forest or change to more open savanna and grassland conditions in the landscape pattern of vegetation. Because low intensity fire shaped the vegetation type, we sought to learn whether fire would keep the decline areas open. Methods The study was conducted in a xeric oak forest in east-central Oklahoma, USA. Randomly located vegetation and regeneration surveys were conducted in decline and non-decline stands 1 and 5 years following nearly total canopy mortality. Diameter at breast height (DBH), regeneration and sprout origin were recorded for all woody species. Important Findings The major canopy species post oak (Quercus stellata Wangenh.), black- jack oak (Quercus marilandica Muenchh.) and black hickory (Carya texana Buckl.) suffered 85–92% mortality; however, minor canopy components experienced limited mortality. Mortality affected all size classes of canopy trees except those below 5 cm breast height diam- eter. There was abundant regeneration of all species and fire seemed to maintain a high level of sprouting. Decline appeared to decrease the relative importance of stump sprouting and increase other types including root sprouts. Decline areas had abundant true seedlings, with stem origin from a root with the same diameter as the stem, which is very unusual for xeric oak forests. Regeneration height in decline areas was twice that of non-decline forests. Our findings suggest that forest decline may lead to: (i) reduced oak dominance and species change in the canopy, (ii) change in reproduction type to increase success of true seedlings and maintain genetic diversity of oaks. Keywords: oak decline, prescribed fire, seedling, stump sprouts, drought Received: 3 June 2015, Revised: 9 October 2015, Accepted: 31 October 2015 INTRODUCTION Forest decline is a disease complex that affects oak or mixed oak forests which represent over 50% of forests in eastern United States (Spetich et al. 2002). It has been documented in USA for over 100 years (Houston 1987; Law and Gott 1987) and in Europe for over 300 years (Thomas et al. 2002). Due to the frequency and damage of these mortality events, forest decline has been determined to be one of the most serious forest disease problems in the southern and eastern United States (Oak 2002). Forest decline in oak forests causes a loss of tree and stand vigor and health without obvious evi- dence of a single clearly identifiable causal factor (Ciesla and Donaubauer 1994). It has been reported that tree death can occur in as short as a few months, but usually develops in the span of 2–4 years (Tomiczek 1993) and can range from a few trees to hundreds of hectares (Oak 2002). Species in the red oak group (Quercus section Lobatae) have been shown Downloaded from https://academic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Plant EcologyVolumE 9, NumbEr 4, PagEs 402–409
august 2016
doi:10.1093/jpe/rtv071
advance access publication 5 November 2015
available online at www.jpe.oxfordjournals.org
© The Author 2015. Published by Oxford University Press on behalf of the Institute of Botany, Chinese Academy of Sciences and the Botanical Society of China.
All rights reserved. For permissions, please email: [email protected]
Ecological succession following forest decline in a xeric oak forest of south-central united states
Devin P. Bendixsen1,2,*, Stephen W. Hallgren1 and
Jesse A. Burton1,3
1 Department of Natural Resource Ecology and Management, Oklahoma State University, 008 Ag Hall, Stillwater, OK 74074, USA2 Present address: Department of Biomolecular Sciences, Boise State University, 1357 S. Brady St., Boise, ID 83725, USA3 Present address: National Park Service, 2680 Natchez Trace Pkwy, Tupelo, MS 38804, USA*Correspondence address. Department of Natural Resource Ecology and Management, Oklahoma State University, 008 Ag Hall, Stillwater, OK 74074, USA. Tel: +1 575 725 0163; Fax: +1 208 392 1430; E-mail: [email protected]
Abstract
Aimsthe loss of canopy trees associated with forest decline can greatly influence the species composition and structure of a forest and have major impacts on the ecosystem. We studied the changes in forest composition and structure 1 and 5 years following nearly total canopy mortality on several hundreds of hectares of xeric oak forests in south-central united states. because the forests were within an ecotonal vegetation type composed of a mosaic of forest, savanna and grassland, we sought to learn whether forest decline areas would recover to forest or change to more open savanna and grassland conditions in the landscape pattern of vegetation. because low intensity fire shaped the vegetation type, we sought to learn whether fire would keep the decline areas open.
Methodsthe study was conducted in a xeric oak forest in east-central oklahoma, usa. randomly located vegetation and regeneration surveys were conducted in decline and non-decline stands 1 and 5 years following nearly total canopy mortality. Diameter at breast height (DbH), regeneration and sprout origin were recorded for all woody species.
Important Findingsthe major canopy species post oak (Quercus stellata Wangenh.), black-jack oak (Quercus marilandica muenchh.) and black hickory (Carya texana buckl.) suffered 85–92% mortality; however, minor canopy components experienced limited mortality. mortality affected all size classes of canopy trees except those below 5 cm breast height diam-eter. there was abundant regeneration of all species and fire seemed to maintain a high level of sprouting. Decline appeared to decrease the relative importance of stump sprouting and increase other types including root sprouts. Decline areas had abundant true seedlings, with stem origin from a root with the same diameter as the stem, which is very unusual for xeric oak forests. regeneration height in decline areas was twice that of non-decline forests. our findings suggest that forest decline may lead to: (i) reduced oak dominance and species change in the canopy, (ii) change in reproduction type to increase success of true seedlings and maintain genetic diversity of oaks.
Keywords: oak decline, prescribed fire, seedling, stump sprouts, drought
received: 3 June 2015, revised: 9 october 2015, accepted: 31 october 2015
INtroDuCtIoNForest decline is a disease complex that affects oak or mixed oak forests which represent over 50% of forests in eastern United States (Spetich et al. 2002). It has been documented in USA for over 100 years (Houston 1987; Law and Gott 1987) and in Europe for over 300 years (Thomas et al. 2002). Due to the frequency and damage of these mortality events, forest decline has been determined to be one of the most serious
forest disease problems in the southern and eastern United States (Oak 2002). Forest decline in oak forests causes a loss of tree and stand vigor and health without obvious evi-dence of a single clearly identifiable causal factor (Ciesla and Donaubauer 1994). It has been reported that tree death can occur in as short as a few months, but usually develops in the span of 2–4 years (Tomiczek 1993) and can range from a few trees to hundreds of hectares (Oak 2002). Species in the red oak group (Quercus section Lobatae) have been shown
August
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022

Bendixsen et al. | Ecological succession following forest decline 403
to be particularly susceptible (Greenberg 2011; Starkey and Oak 1989). The loss of oak trees following forest decline can greatly influence ecosystem processes and functions due to changes in species composition and structure of a forest. Oak trees are a ‘keystone’ to biological diversity, mostly because their mast production plays a vital role in the food webs of oak forests (Feldhamer 2002). A reduction of oak trees in a for-est can have a negative effect on wildlife dependent on oak, due to habitat and food loss (McShea et al. 2007). Even before tree death, declining trees in Hungary have been shown to decrease their growth rate to almost half of healthy trees of the same size (Standovar and Somogyi 1998).
Several hundred hectares of old growth xeric oak forest in the south-central United States suffered nearly 100% canopy mortality in 2008. This near complete canopy mortality was determined to be caused by a complex of stress factors includ-ing: distance to water, slope, elevation, aspect, drought, a false spring and three possible plant pathogens (Bendixsen et al. 2015). Although these forests are on shallow drought-prone soils, where drought-caused mortality has been docu-mented (Albertson and Weaver 1945; DeSantis et al. 2010; Johnson and Risser 1975; Rice and Penfound 1959), there have been no prior reports of complete canopy mortality over large areas. The forests are part of an ecotonal vegetation type called the Cross Timbers, characterized by a mosaic of forest, savanna and grassland. Our first question was whether forest decline could contribute to the vegetation pattern of this eco-tone by rapidly converting closed forest to more open savanna or grassland. Furthermore, because frequent low intensity prescribed fire has maintained this vegetation type recently (Burton et al. 2010) and for thousands of years (Delcourt 2002; Guyette et al. 2002), another question was whether fire would keep decline areas open. The purpose of this study was to determine the overall ecological succession of the decline stands to learn whether the forest would recover and whether the species composition of the overstory would change.
matErIals aND mEtHoDsStudy area
Research was conducted at the Okmulgee Game Management Area (OGMA), a heavily forested reserve of 3700 ha in eastern Oklahoma (35° 38′ N, 96° 02′ W) managed by the Oklahoma Department of Wildlife Conservation (ODWC). The climate was subtropical with a mean annual temperature of 16°C and mean annual precipitation of 104 cm (OCS, http://climate.ok.gov/index.php/climate/). In the surveyed upland sites, the dominant soil was Hector–Endsaw complex characterized as well drained, shallow, stony fine sandy loam with bedrock at 30 cm. The land ranged from level to short steep topography with 5–30% slopes (Sparwasser et al. 1968).
The study area was ~95% wooded. An earlier study found a mean diameter at breast height (1.4 m) of 16 cm, dominant tree height of 13 m, stand density of 1450 trees ha−1, and mean basal area of 26 m2 ha−1 (Karki and Hallgren 2015). The
forest was dominated by post oak (Quercus stellata Wangenh., 77%), blackjack oak (Quercus marilandica Muenchh., 9%) and black hickory (Carya texana Buckl., 8%). OGMA has been managed by ODWC since the 1950s and regularly prescribe burned since 1988. Low intensity dormant season burns were conducted in well-defined management units (Burton et al. 2010).
In 2008, forest decline was found to have killed nearly 100% of canopy trees on several hundred hectares of OGMA (Fig. 1). This study was conducted in three man-agement units where the majority of the decline occurred. Units ranged in size from 150 to 220 ha and had been burned three times in the 12 years prior to measurement. Two of the study units were treated with an early spring prescribed burn following forest decline, one in 2011 and the other in 2010. The third unit was burned by wildfire in August 2011.
Measurements
Measurements were conducted in two management units in August 2009 and three management units in summer 2013. Study plots were randomly located in both decline and non-decline stands using the random point tool in ArcCatalog (ESRI 2011). In 2009, there were 10 plots in each unit, 5 in decline areas and 5 in non-decline areas, for a total of 20 plots. In 2013, there were 30 plots in each unit, 15 in decline areas and 15 in non-decline areas, for a total of 90 plots. Decline areas were clearly distinguished by nearly 100% can-opy mortality and the boundary between decline and non-decline stands was abrupt and easily identified. No plots were located within 10 m of man-made clearings, such as wildlife food plots or roads.
Vegetation surveys were conducted at each randomly located point on a 100 m2 plot. The diameter at breast height (dbh) was measured for all live and dead woody plants. At each randomly located plot in 2013, a round regenera-tion survey plot with a diameter of 3 m was conducted. All woody regeneration ≤2 m tall were identified by species and measured for height. In 2013 surveys, sprout origin was also determined according to the following definitions (Clark and Hallgren 2003; DeSantis and Hallgren 2011):
• Root sprout:Stemoriginwas root>25cm fromproximalend of the root to insure that sprouts originated from lateral root and not root collar.
• Stumpsprout:Stemoriginwasrootcollaroflivetreewithdiameter>5cm.
• Seedlingsprout:Stemoriginwasrootcollarlivetreewithdiameter <5 cm.
• Seedling: Stem origin was root of same diameter as thestem with no evidence of dieback and sprouting.
Data analysis
Stem density was broken into four categories; large trees (dbh >10cm),smalltrees(10cm≥dbh≥5cm),saplings(dbh<5cmandheight> breastheight) and regeneration (woodyplants
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022

404 Journal of Plant Ecology
< 2 m tall). Two-way analysis of variance (ANOVA) was used to test for significance of decline effect and year effect using SPSS 21.1 statistical software (IBM Corp. 2012, Version 21.0., Armonk, NY, USA). The independent variables in our analy-sis were mortality (decline and non-decline) and year (2009 and 2013). The dependent variables were live and dead basal area, stem density (sprouts, saplings, small trees, large trees) and sprout height. Two-tailed Student’s t-test was used to ana-lyze regeneration origin because this data was only collected in 2013. T-test was also used to test for differences between two species within a given year. Percentage data was arcsine transformed prior to statistical analysis. The experimental unit was the decline or non-decline stand condition within each management unit. Sample size for analyses was two in 2009 and three in 2013. Within each decline treatment sample, there were 5 subsamples in 2009 and 15 subsamples in 2013. Effects are reported only when the statistical test produced a P ≤ 0.05.
rEsultsOverstory
Decline stands had much higher mortality of total basal area ha−1 (84–91%) than non-decline stands (10–17%) in both 2009 and 2013 (Fig. 2, F = 60.041, P < 0.001). In decline stands,
mortality as a percent of total basal area ha−1 was equally high for both post oak and blackjack oak which comprised 75% and 15% of the total basal area, respectively (data not shown, P = 0.71). In non-decline stands, post oak mortality as a per-cent of total basal area ha−1 was much lower (3%) compared to blackjack oak (22%) (data not shown, P = 0.024). Mortality in terms of stems ha−1 in decline stands affected all size classes (Fig. 3) except the smallest size class, <5cm dbh.
Post oak and blackjack oak dominated relative basal area of live trees in non-decline stands in both 2009 and 2013 (Fig. 4, P < 0.03). Winged elm (Ulmus alata Michx.) relative basal area showed a large increase in decline stands in 2009 and 2013 (F = 46.056, P = 0.01). Black hickory relative basal area increased sharply in decline stands in 2009 but not in 2013 (F = 49.365, P < 0.001).
Total large tree density in terms of stem ha−1 was 80% lower in decline stands than non-decline stands in 2013 (Fig. 5, F = 39.6, P = 0.001). Forest decline resulted in the reduction in density of large trees of blackjack oak by 80% (F = 14.745, P = 0.009) and of post oak by 65–80% (F = 7.615, P = 0.033) in both 2009 and 2013. Forest decline caused a reduction in the density of small trees of black hickory by 80% in 2009 and 2013 (F = 8.093, P = 0.033). Post oak and blackjack oak small trees remained dominant after decline in both 2009 and 2013 (P < 0.001). Winged elm increased in small trees from 2009 to 2013 to become a major component of the stand in both non-decline and decline stands (F = 21.110, P = 0.004). Forest decline caused an increase of post oak saplings of 14-fold in 2009 and 3-fold in 2013 (F = 5.203, P = 0.063). Winged elm dominated sprout density for both non-decline and decline stands in both 2009 and 2013 (F = 0.566, P = 0.657).
Regeneration
The number of stems per hectare for sprouts did not vary sig-nificantly in decline and non-decline areas and between 2009 and 2013 (Fig. 5, F = 3.013, P = 0.161). Average sprout height was approximately double in decline areas compared to non-decline areas for 2009 and 2013 (data not shown, F = 16.933, P = 0.006) and was approximately the same in 2009 and 2013 for both decline and non-decline stands (data not shown, F = 0.068, P = 0.802).
The relative importance of stump sprouts decreased in decline stands in 2013 for black hickory (P = 0.033), post oak (P = 0.013), other species (P = 0.029) and total sprouts (P = 0.004) (Fig. 6). The proportion of root sprouts increased for black hickory in decline stands (P = 0.018). The proportion of seedling sprouts increased in decline stands for post oak (P = 0.012), other spe-cies (P = 0.034) and total species (P = 0.008). The proportion of seedlings in decline stands increased for blackjack oak (P = 0.02), other species (P = 0.01) and total species (P = 0.010).
DIsCussIoNThe major findings of this study were forest decline in xeric oak forests had the potential to change composition of the
Figure 1: map of Okmulgee Game Management Area and areas affected by oak decline. Decline areas represent stands with nearly 100% overstory mortality.
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022
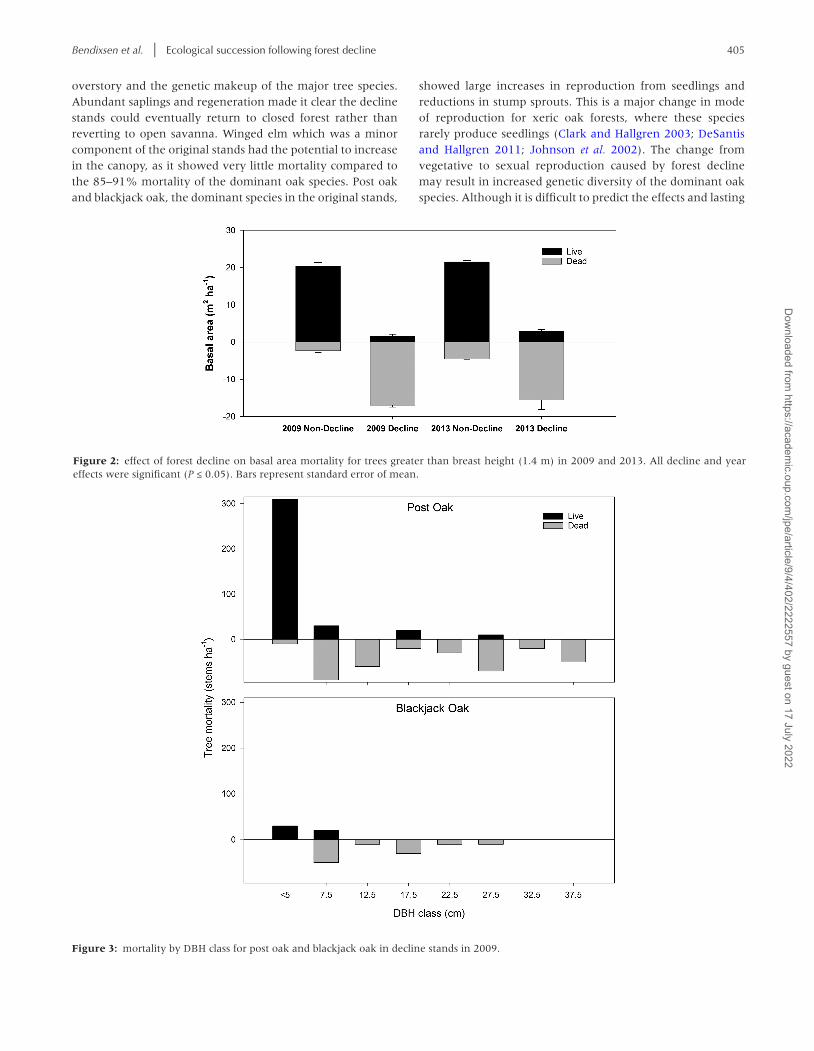
Bendixsen et al. | Ecological succession following forest decline 405
overstory and the genetic makeup of the major tree species. Abundant saplings and regeneration made it clear the decline stands could eventually return to closed forest rather than reverting to open savanna. Winged elm which was a minor component of the original stands had the potential to increase in the canopy, as it showed very little mortality compared to the 85–91% mortality of the dominant oak species. Post oak and blackjack oak, the dominant species in the original stands,
showed large increases in reproduction from seedlings and reductions in stump sprouts. This is a major change in mode of reproduction for xeric oak forests, where these species rarely produce seedlings (Clark and Hallgren 2003; DeSantis and Hallgren 2011; Johnson et al. 2002). The change from vegetative to sexual reproduction caused by forest decline may result in increased genetic diversity of the dominant oak species. Although it is difficult to predict the effects and lasting
Figure 2: effect of forest decline on basal area mortality for trees greater than breast height (1.4 m) in 2009 and 2013. All decline and year effects were significant (P ≤ 0.05). Bars represent standard error of mean.
Figure 3: mortality by DBH class for post oak and blackjack oak in decline stands in 2009.
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022
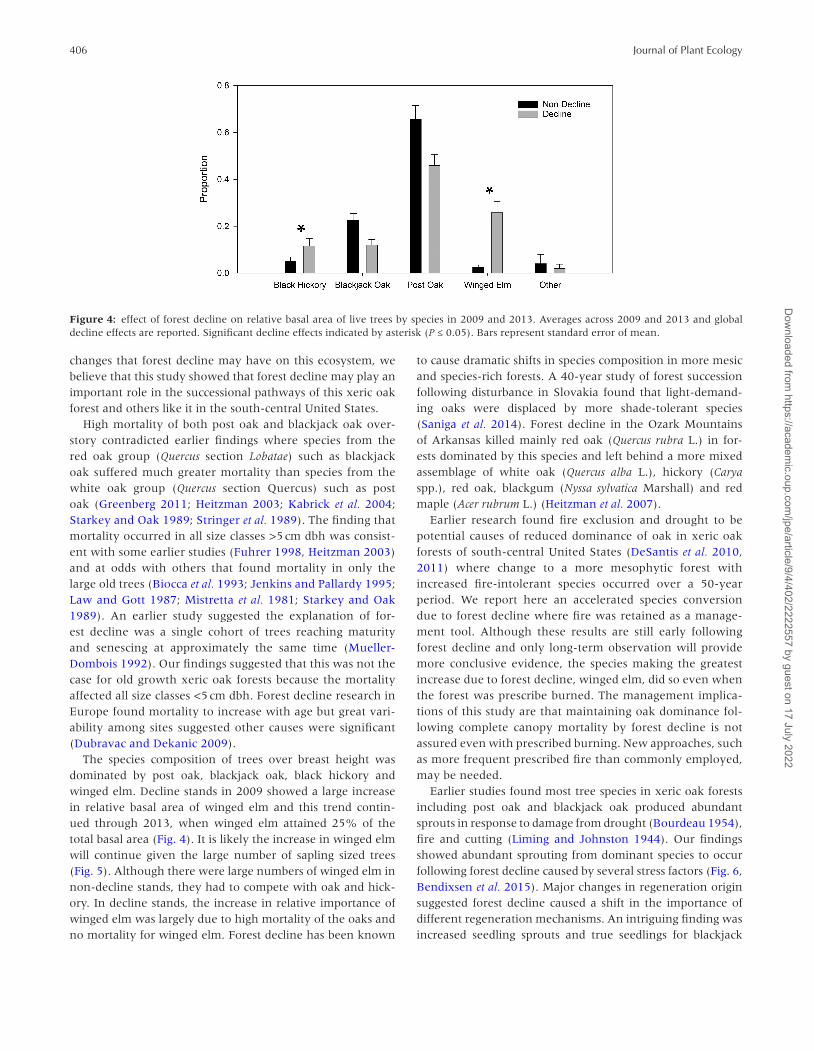
406 Journal of Plant Ecology
changes that forest decline may have on this ecosystem, we believe that this study showed that forest decline may play an important role in the successional pathways of this xeric oak forest and others like it in the south-central United States.
High mortality of both post oak and blackjack oak over-story contradicted earlier findings where species from the red oak group (Quercus section Lobatae) such as blackjack oak suffered much greater mortality than species from the white oak group (Quercus section Quercus) such as post oak (Greenberg 2011; Heitzman 2003; Kabrick et al. 2004; Starkey and Oak 1989; Stringer et al. 1989). The finding that mortalityoccurredinallsizeclasses>5cmdbhwasconsist-ent with some earlier studies (Fuhrer 1998, Heitzman 2003) and at odds with others that found mortality in only the large old trees (Biocca et al. 1993; Jenkins and Pallardy 1995; Law and Gott 1987; Mistretta et al. 1981; Starkey and Oak 1989). An earlier study suggested the explanation of for-est decline was a single cohort of trees reaching maturity and senescing at approximately the same time (Mueller-Dombois 1992). Our findings suggested that this was not the case for old growth xeric oak forests because the mortality affected all size classes <5 cm dbh. Forest decline research in Europe found mortality to increase with age but great vari-ability among sites suggested other causes were significant (Dubravac and Dekanic 2009).
The species composition of trees over breast height was dominated by post oak, blackjack oak, black hickory and winged elm. Decline stands in 2009 showed a large increase in relative basal area of winged elm and this trend contin-ued through 2013, when winged elm attained 25% of the total basal area (Fig. 4). It is likely the increase in winged elm will continue given the large number of sapling sized trees (Fig. 5). Although there were large numbers of winged elm in non-decline stands, they had to compete with oak and hick-ory. In decline stands, the increase in relative importance of winged elm was largely due to high mortality of the oaks and no mortality for winged elm. Forest decline has been known
to cause dramatic shifts in species composition in more mesic and species-rich forests. A 40-year study of forest succession following disturbance in Slovakia found that light-demand-ing oaks were displaced by more shade-tolerant species (Saniga et al. 2014). Forest decline in the Ozark Mountains of Arkansas killed mainly red oak (Quercus rubra L.) in for-ests dominated by this species and left behind a more mixed assemblage of white oak (Quercus alba L.), hickory (Carya spp.), red oak, blackgum (Nyssa sylvatica Marshall) and red maple (Acer rubrum L.) (Heitzman et al. 2007).
Earlier research found fire exclusion and drought to be potential causes of reduced dominance of oak in xeric oak forests of south-central United States (DeSantis et al. 2010, 2011) where change to a more mesophytic forest with increased fire-intolerant species occurred over a 50-year period. We report here an accelerated species conversion due to forest decline where fire was retained as a manage-ment tool. Although these results are still early following forest decline and only long-term observation will provide more conclusive evidence, the species making the greatest increase due to forest decline, winged elm, did so even when the forest was prescribe burned. The management implica-tions of this study are that maintaining oak dominance fol-lowing complete canopy mortality by forest decline is not assured even with prescribed burning. New approaches, such as more frequent prescribed fire than commonly employed, may be needed.
Earlier studies found most tree species in xeric oak forests including post oak and blackjack oak produced abundant sprouts in response to damage from drought (Bourdeau 1954), fire and cutting (Liming and Johnston 1944). Our findings showed abundant sprouting from dominant species to occur following forest decline caused by several stress factors (Fig. 6, Bendixsen et al. 2015). Major changes in regeneration origin suggested forest decline caused a shift in the importance of different regeneration mechanisms. An intriguing finding was increased seedling sprouts and true seedlings for blackjack
Figure 4: effect of forest decline on relative basal area of live trees by species in 2009 and 2013. Averages across 2009 and 2013 and global decline effects are reported. Significant decline effects indicated by asterisk (P ≤ 0.05). Bars represent standard error of mean.
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022
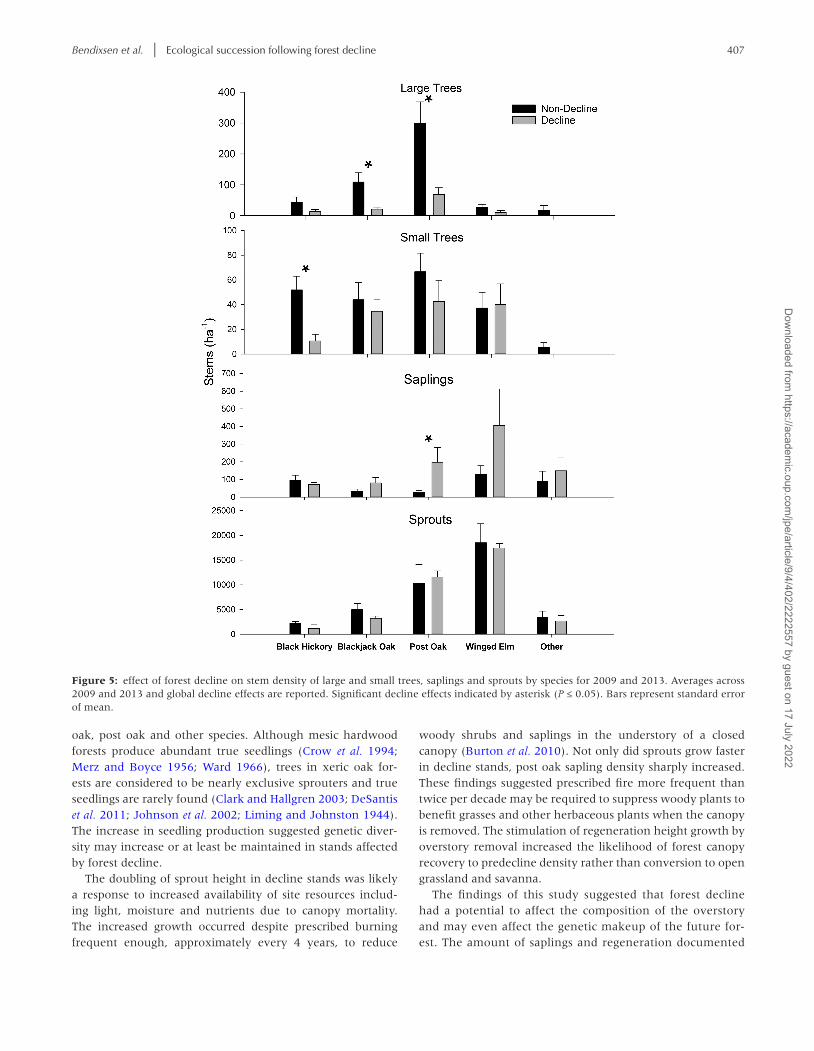
Bendixsen et al. | Ecological succession following forest decline 407
oak, post oak and other species. Although mesic hardwood forests produce abundant true seedlings (Crow et al. 1994; Merz and Boyce 1956; Ward 1966), trees in xeric oak for-ests are considered to be nearly exclusive sprouters and true seedlings are rarely found (Clark and Hallgren 2003; DeSantis et al. 2011; Johnson et al. 2002; Liming and Johnston 1944). The increase in seedling production suggested genetic diver-sity may increase or at least be maintained in stands affected by forest decline.
The doubling of sprout height in decline stands was likely a response to increased availability of site resources includ-ing light, moisture and nutrients due to canopy mortality. The increased growth occurred despite prescribed burning frequent enough, approximately every 4 years, to reduce
woody shrubs and saplings in the understory of a closed canopy (Burton et al. 2010). Not only did sprouts grow faster in decline stands, post oak sapling density sharply increased. These findings suggested prescribed fire more frequent than twice per decade may be required to suppress woody plants to benefit grasses and other herbaceous plants when the canopy is removed. The stimulation of regeneration height growth by overstory removal increased the likelihood of forest canopy recovery to predecline density rather than conversion to open grassland and savanna.
The findings of this study suggested that forest decline had a potential to affect the composition of the overstory and may even affect the genetic makeup of the future for-est. The amount of saplings and regeneration documented
Figure 5: effect of forest decline on stem density of large and small trees, saplings and sprouts by species for 2009 and 2013. Averages across 2009 and 2013 and global decline effects are reported. Significant decline effects indicated by asterisk (P ≤ 0.05). Bars represent standard error of mean.
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022

408 Journal of Plant Ecology
in decline stands make a strong argument that areas affected by decline could return to closed canopy forests with time even when burned with low intensity dormant season pre-scribed fire two times per decade. Due to the low impact that decline had on winged elm this species may play a big-ger role in the future forest than previously seen. Genetic diversity of the dominant oak species, post oak and black-jack oak, might also be changed due to the large increases in reproduction from seedlings and the corresponding reduc-tion in stump sprout reproduction. The findings of this study suggested that forest decline may play an important role in the successional pathways seen in xeric oak forests in south-central United States and in similar forests through-out the USA.
aCKNoWlEDgEmENtsWe would like to thank the Oklahoma Department of Wildlife Conservation for access to the Okmulgee Game Management Area. We would also like to thank Dr Michael Palmer and Dr Rod Will for their assistance in analysis and manuscript preparation.Conflict of interest statement. None declared.
rEFErENCEsAlbertson FW, Weaver JE (1945) Injury and death or recovery of trees
in prairie climate. Ecol Monogr 15:393–433.
Bendixsen DP, Hallgren SW, Frazier AE (2015) Stress factors associ-
ated with forest decline in xeric oak forests of south-central United
States. For Ecol Manag 347:40–8.
Biocca M, Tainter FH, Starkey DA, et al (1993) The persistence of
oak decline in the western North Carolina Nantahala Mountains.
Castanea 58:178–84.
Bourdeau P (1954) Oak seedling ecology determining segrega-
tion of species in Piedmont oak-hickory forests. Ecol Monogr
24:297–320.
Burton JA, Hallgren SW, Palmer MW (2010) Fire frequency affects
structure and composition of xeric forests of eastern Oklahoma.
Nat Area J 30:370–9.
Ciesla WM, Donaubauer E (1994) Decline and Dieback of Trees and
Forests: A Global Overview. Food and Agriculture Organization of
the United Nations.
Clark SL, Hallgren SW (2003) Dynamics of oak (Quercus marilandica
and Q. stellata) reproduction in an old-growth Cross Timbers forest.
Southeastern Nat 2:559–74.
Figure 6: effect of oak decline on regeneration origin by species. ST = stump sprout, RO = root sprout, SS = seedling spout, and SE = true seed-ling. Significant decline effect indicated by asterisk (P ≤ 0.05). Bars represent standard error of mean.
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022

Bendixsen et al. | Ecological succession following forest decline 409
Crow TR, Johnson WC, Adkisson CS (1994) Fire and recruitment of
Quercus in a postagricultural field. Am Midl Nat 131:84–97.
Delcourt HR (2002) Forests in Peril: Tracking Deciduous Trees from Ice-Age
Refuges into the Greenhouse World. Blacksburg VA, USA: McDonald
and Woodward Publishing Company.
DeSantis RD, Hallgren SW, Lynch TB, et al (2010) Long-term direc-
tional changes in upland Quercus forests throughout Oklahoma,
USA. J Veg Sci 21:606–15.
DeSantis RD, Hallgren SW (2011) Prescribed burning frequency
affects post oak and blackjack oak regeneration. South J Appl For
35:193–8.
Dubravac T, Dekanic S (2009) Structure and dynamics of the harvest
of dead and declining trees of penduculate oak in the stands of
Spacva forest from 1996 to 2006. Sumarski List 133:391–405.
ESRI (2011). ArcGIS Desktop: Release 10. Redlands, CA: Environmental
Systems Research Institute.
Feldhamer GA (2002) Acorns and white-tailed deer: interrelation-
ships in forest ecosystems. In McShea WJ, Healy WM (eds). Oak
Forest Ecosystems: Ecology and Management for Wildlife. Baltimore,
MD: John Hopkins Press, 215–23.
Fuhrer E (1998) Oak Decline in Central Europe: A Synopsis of Hypotheses.
Population Dynamics, Impacts, and Integrated Management of Forest
Defoliating Insects. USDA Forest Service General Technical Report
NE-247, 7–24.
Greenberg CH (2011) Temporal patterns of oak mortality in a south-
ern Appalachian forest (1991–2006). Nat Area J 31:131–7.
Guyette RP, Muzika RM, Dey DC (2002) Dynamics of an anthropo-
genic fire regime. Ecosystems 5:472–86.
Heitzman E (2003) Effects of oak decline on species composition in a
northern Arkansas forest. South J Appl For 27:264–8.
Heitzman E, Grell A, Spetich M, et al. (2007) Changes in forest struc-
ture associated with oak decline in severely impacted areas of
northern Arkansas. South J Appl For 31:17–22.
Houston DR (1987) Forest tree declines of past and present – Current
understanding. Can J Plant Path 9:349–60.
Jenkins MA, Pallardy SG (1995) The influence of drought on red oak
group species growth and mortality in the Missouri Ozarks. Can J
For Res 25:1119–27.
Johnson FL, Risser PG (1975) A quantitative comparison between an
oak forest and an oak savanna in central Oklahoma. Southwestern
Nat 20:75–84.
Johnson PS, Shifley SR, Rogers R (2002) The Ecology and Silviculture of
Oaks. New York, NY: CABI Publishing.
Kabrick JM, Shifley SR, Jensen RG, et al. (2004) Factors associated
with oak mortality in Missouri Ozark forests. In Yaussy DA, Hix
DM, Long RP, et al. (eds). Proceedings of the 14th Central Hardwood
Forest Conference. USDA Forest Service [General Technical
Report no. NE-316], Northeastern Research Station, Newton
Square, PA.
Karki L, Hallgren SW (2015) Tree-fall gaps and regeneration in old-
growth cross timbers forests. Nat Area J 35:533–41.
Law JR, Gott JD (1987) Oak mortality in the Missouri Ozarks. In Hay
RL, Woods FW, DeSelm H (eds). Proceeding of 6th Central Hardwoods
Forest Conference. Knoxville, TN: University of Tennessee, 427–36.
Liming FG, Johnston JP (1944) Reproduction in oak-hickory forest
stands of the Missouri Ozarks. J For 42:175–80.
McShea WJ, Healy WM, Devers P, et al. (2007) Forestry matters:
Decline of oaks will impact wildlife in hardwood forests. J Wildl
Manag 71:1717–28.
Merz RW, Boyce SG (1956) Age of oak seedlings. J For 54:774–5.
Mistretta PA, Affeltranger CE, Starkey DA, et al. (1981) Evaluation of oak
mortality on the Ozark National Forest, Arkansas. USDA Forest Service
Forest Pest Management [Report no. 81-2-26], Pineville, LA.
Mueller-Dombois D (1992) A Natural Dieback Theory, cohort senescence
as an alternative to the decline disease theory. In PD Manion, Lachance
D (eds). Forest Decline Concepts. St. Paul, MN: APS Press, 26–37.
Oak SW (2002) Native diseases and insects that impact oaks. In
McShea WJ, Healy WM (eds). Oak Forest Ecosystems. Baltimore,
MD: Johns Hopkins University Press, 80–99.
Rice EL, Penfound WT (1959) The upland forest of Oklahoma. Ecology
40:593–608.
Saniga M, Balanda M, Kucbel S, et al. (2014) Four decades of for-
est succession in the oak-dominated forest reserves in Slovakia.
IFOREST 7:324–32.
Sparwasser WA, Bogard VA, Henson OG (1968) Soil Survey. Okmulgee
County, OK: U.S. Government Printing Office, Washington DC.
Spetich MA, Dey DC, Johnson PS, et al. (2002) Competitive capacity
of Quercus rubra L. planted in Arkansas’ Boston Mountains. For Sci
48:504–17.
Standovar T, Somgyi Z (1998) Corresponding patterns of site qual-
ity, decline and tree growth in a sessile oak stand. Eur J For Pathol
28:133–44.
Starkey DA, Oak SW (1989) Site factors and stand conditions asso-
ciated with oak decline in southern upland hardwood forests.
In Rink G, Budelsky CA (eds). Proceedings of the Seventh Central
Hardwood Conference. St Paul, MN: USDA Forest Service [General
Technical Report no. NC-132], North Central Forest Experiment
Station, 95–102.
Stringer JW, Kimmerer TW, Overstreet JC, et al. (1989) Oak mortality
in eastern Kentucky. South J Appl For 13: 86–91.
Thomas FM, Blank R, Hartmann G (2002) Abiotic and biotic factors
and their interactions as causes of oak decline in Central Europe.
For Pathol 32:277–307.
Tomiczek C (1993) Oak decline in Austria and Europe. J Arboric
19:71–3.
Ward WW (1966) Oak-hardwood reproduction in central Pennsy-
lvania. J For 64:744–9.
Dow
nloaded from https://academ
ic.oup.com/jpe/article/9/4/402/2222557 by guest on 17 July 2022
Related Documents











