Early Responsiveness to Stimuli Paired With Different Stages Within the State of Alcohol Intoxication Ricardo M. Pautassi, Juan Carlos Godoy, Norman E. Spear, and Juan Carlos Molina Background: Infant rats quickly learn to avoid a sensory cue paired with alcohol as an unconditioned stimulus, particularly when the drug reaches peak blood concentrations. In this study, a tactile cue paired with the onset of alcohol intoxication preceded subsequent presentations of a gustatory conditioned stim- ulus (CS). The goal was to address the possibility of differential conditioning depending on when stimuli were introduced during the course of the toxic state. Methods: In experiment 1, rat pups received sequential presentations of a salient texture (sandpaper) and a gustatory cue (saccharin) while intoxicated with a 2.5 g/kg alcohol dose or after receiving saline. Texture location tests and saccharin intake assessments were then performed. A third modality of assess- ment was defined by a saccharin intake test while pups simultaneously experienced sandpaper. In experi- ment 2, alcohol-mediated conditioning was followed by tests similar to those of experiment 1, but after pups were re-exposed to either the tactile CS or the alcohol-unconditioned stimulus. Results: Conditioned taste aversions, due to pairing saccharin and the later stage of alcohol intoxication, were reliably established in both experiments. Also in both experiments, this excitatory aversive response was dramatically inhibited when the association between the texture CS and the earlier stage of alcohol intoxication was activated. There were no indications of conditioned motor responses to the tactile CS that might compete with intake behavior of saccharin or distort measurement of an appetitive memory derived from pairing the texture and the earlier stage of intoxication. Conclusions: Rat pups’ expression of an association between a taste signaling aversive consequences of alcohol was eliminated by the presence of a tactile stimulus that originally had signaled the absence of aversive consequences of alcohol intoxication. The results suggest the interaction of inhibitory and excita- tory conditioning involving the aversive properties of alcohol. Key Words: Alcohol Intoxication, Infant Rats, Taste Aversion Learning, Tactile Conditioning, Inhibi- tory Conditioning. A LCOHOL HAS BEEN observed to act as an uncondi- tioned stimulus (US) capable of supporting associa- tive learning that shapes seeking and intake behaviors of this drug (Cunningham et al., 2000; US Department of Health and Human Services, 2000). In heterogeneous strains of rats, an aversive component of the state of alco- hol intoxication is easily detected with various tests of learning. For example, alcohol has been demonstrated to act as an aversive US capable of establishing conditioned taste aversion (CTA) learning (Cappel and Le Blanc, 1973; Cunningham et al., 1988, 1992; Dominguez et al., 1994; Hunt et al., 1990, 1991). Rats generally do not express appetitive contents of memories generated by contingent experiences between specific visual or tactile cues and post- absorptive effects of alcohol. On the contrary, the predom- inant motivational effects of this associative process seem to be aversive. Conditioned place aversions are readily established when alcohol doses are used that exceed 1 g/kg of ethanol (EtOH) (Cunningham et al., 1993; Gauvin and Holloway, 1992; Schechter and Krimmer, 1992). Lower doses rarely exert significant aversive or appetitive condi- tioning (Stewart and Grupp, 1985; Van der Kooy et al., 1983). Very extensive training procedures (Bozarth, 1990), long-term pre-exposure to the drug’s interoceptive effects (Reid et al., 1985), concurrent presentation of other rein- forcers (Marglin et al., 1988), or a combination of these have been necessary to attain alcohol-mediated condi- tioned place preferences in heterogeneous rats. In these animals, failure to find evidence of appetitive reinforcing properties of alcohol has frequently been reported when From the Instituto de Investigación Médica M.M. Ferreryra (RMP, JCG, JCM), Consejo Nacional de Investigaciones Científicas y Técnicas, Cordoba, Argentina; the Center for Developmental Psychobiology (NES, JCM), De- partment of Psychology, Binghamton University-State University of New York, Binghamton, New York; and Facultad de Psicología (JCG, JCM), Univer- sidad Nacional de Córdoba, Argentina. Received for publication November 2, 2001; accepted February 25, 2002. This research, a collaborative project between the Research Foundation of SUNY Binghamton and Instituto Ferreyra, was supported by Grants 1R21AA12762 and 1RO1AA11960 from the NIAAA (NES), Agencia Na- cional de Promoción Científica y Tecnológica Beca and Carrillo-Onativia, Ministerio de Salud, Argentina (JCM), and fellowships of the Fundación Interior Argentina and Consejo Nacional de Investigaciones Científicas y Técnicas, (RMP). Reprint requests: Juan Carlos Molina, PhD, Instituto Ferreyra, Casilla de Correo 389, 5000 Córdoba, Argentina; Fax: 54-351-469-5163; E-mail: [email protected] Copyright © 2002 by the Research Society on Alcoholism. 0145-6008/02/2605-0644$03.00/0 ALCOHOLISM:CLINICAL AND EXPERIMENTAL RESEARCH Vol. 26, No. 5 May 2002 644 Alcohol Clin Exp Res, Vol 26, No 5, 2002: pp 644–654

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Early Responsiveness to Stimuli Paired With DifferentStages Within the State of Alcohol Intoxication
Ricardo M. Pautassi, Juan Carlos Godoy, Norman E. Spear, and Juan Carlos Molina
Background: Infant rats quickly learn to avoid a sensory cue paired with alcohol as an unconditionedstimulus, particularly when the drug reaches peak blood concentrations. In this study, a tactile cue pairedwith the onset of alcohol intoxication preceded subsequent presentations of a gustatory conditioned stim-ulus (CS). The goal was to address the possibility of differential conditioning depending on when stimuliwere introduced during the course of the toxic state.
Methods: In experiment 1, rat pups received sequential presentations of a salient texture (sandpaper)and a gustatory cue (saccharin) while intoxicated with a 2.5 g/kg alcohol dose or after receiving saline.Texture location tests and saccharin intake assessments were then performed. A third modality of assess-ment was defined by a saccharin intake test while pups simultaneously experienced sandpaper. In experi-ment 2, alcohol-mediated conditioning was followed by tests similar to those of experiment 1, but after pupswere re-exposed to either the tactile CS or the alcohol-unconditioned stimulus.
Results: Conditioned taste aversions, due to pairing saccharin and the later stage of alcohol intoxication,were reliably established in both experiments. Also in both experiments, this excitatory aversive responsewas dramatically inhibited when the association between the texture CS and the earlier stage of alcoholintoxication was activated. There were no indications of conditioned motor responses to the tactile CS thatmight compete with intake behavior of saccharin or distort measurement of an appetitive memory derivedfrom pairing the texture and the earlier stage of intoxication.
Conclusions: Rat pups’ expression of an association between a taste signaling aversive consequences ofalcohol was eliminated by the presence of a tactile stimulus that originally had signaled the absence ofaversive consequences of alcohol intoxication. The results suggest the interaction of inhibitory and excita-tory conditioning involving the aversive properties of alcohol.
Key Words: Alcohol Intoxication, Infant Rats, Taste Aversion Learning, Tactile Conditioning, Inhibi-tory Conditioning.
ALCOHOL HAS BEEN observed to act as an uncondi-tioned stimulus (US) capable of supporting associa-
tive learning that shapes seeking and intake behaviors ofthis drug (Cunningham et al., 2000; US Department ofHealth and Human Services, 2000). In heterogeneousstrains of rats, an aversive component of the state of alco-hol intoxication is easily detected with various tests of
learning. For example, alcohol has been demonstrated toact as an aversive US capable of establishing conditionedtaste aversion (CTA) learning (Cappel and Le Blanc, 1973;Cunningham et al., 1988, 1992; Dominguez et al., 1994;Hunt et al., 1990, 1991). Rats generally do not expressappetitive contents of memories generated by contingentexperiences between specific visual or tactile cues and post-absorptive effects of alcohol. On the contrary, the predom-inant motivational effects of this associative process seemto be aversive. Conditioned place aversions are readilyestablished when alcohol doses are used that exceed 1 g/kgof ethanol (EtOH) (Cunningham et al., 1993; Gauvin andHolloway, 1992; Schechter and Krimmer, 1992). Lowerdoses rarely exert significant aversive or appetitive condi-tioning (Stewart and Grupp, 1985; Van der Kooy et al.,1983). Very extensive training procedures (Bozarth, 1990),long-term pre-exposure to the drug’s interoceptive effects(Reid et al., 1985), concurrent presentation of other rein-forcers (Marglin et al., 1988), or a combination of thesehave been necessary to attain alcohol-mediated condi-tioned place preferences in heterogeneous rats. In theseanimals, failure to find evidence of appetitive reinforcingproperties of alcohol has frequently been reported when
From the Instituto de Investigación Médica M.M. Ferreryra (RMP, JCG,JCM), Consejo Nacional de Investigaciones Científicas y Técnicas, Cordoba,Argentina; the Center for Developmental Psychobiology (NES, JCM), De-partment of Psychology, Binghamton University-State University of New York,Binghamton, New York; and Facultad de Psicología (JCG, JCM), Univer-sidad Nacional de Córdoba, Argentina.
Received for publication November 2, 2001; accepted February 25, 2002.This research, a collaborative project between the Research Foundation of
SUNY Binghamton and Instituto Ferreyra, was supported by Grants1R21AA12762 and 1RO1AA11960 from the NIAAA (NES), Agencia Na-cional de Promoción Científica y Tecnológica Beca and Carrillo-Onativia,Ministerio de Salud, Argentina (JCM), and fellowships of the FundaciónInterior Argentina and Consejo Nacional de Investigaciones Científicas yTécnicas, (RMP).
Reprint requests: Juan Carlos Molina, PhD, Instituto Ferreyra, Casilla deCorreo 389, 5000 Córdoba, Argentina; Fax: 54-351-469-5163; E-mail:[email protected]
Copyright © 2002 by the Research Society on Alcoholism.
0145-6008/02/2605-0644$03.00/0ALCOHOLISM: CLINICAL AND EXPERIMENTAL RESEARCH
Vol. 26, No. 5May 2002
644 Alcohol Clin Exp Res, Vol 26, No 5, 2002: pp 644–654

EtOH oral self-administration (Stewart et al., 1988; Stewartand Li, 1997) or EtOH intracranial self-administration(Arregui-Aguirre et al., 1987; Schaefer and Michael, 1987)is examined. Nevertheless, it is important to emphasize thatalcohol preferences or aversions are strongly dependent onfactors such as the dosage of the drug and the nature ofprior experience with the psychotropic agent in terms ofchronic or acute exposure (Hine and Figueroa, 1991).
Appetitive properties of EtOH have been most fre-quently encountered when genetically selected lines of ratsand mice are used (Chester and Cunningham, 1999; Cic-occiopo et al., 1999; Cunningham et al., 1991). The use ofthese animal models has indicated biphasic hedonic effectsof the drug during the course of the toxic episode. The drugseems to exert appetitive effects during the early stages ofthe toxic process, when blood alcohol concentrations areincreasing (Risinger and Cunningham, 1992). Aversive ef-fects seem to prevail during later stages of the toxic course.Alcohol-mediated aversive conditioning seems to be easilyestablished when blood alcohol concentrations reach peaklevels. Other drugs, such as morphine and amphetamine,also induce differential hedonic effects during the develop-ment of the toxic episode (Reicher and Holman, 1977;Wise et al., 1976). It has been observed that taste-aversionlearning supported by amphetamine facilitates subsequentplace-preference learning involving this psychotropic agent(Lett, 1988). Apparently, taste conditioning is capable ofblocking the association between other sensory stimuli andamphetamine-induced aversive effects. It is still a matter ofdiscussion whether opposite hedonic effects of drugs ofabuse reflect independent neurobehavioral mechanisms(Cunningham and Ignatoff, 2000; Cunningham and Tull,2000) or whether they are regulated by common physiolog-ical processes (Grigson, 1997, 2000).
During early ontogeny, rat pups can learn about chemo-sensory properties of alcohol (Molina and Chotro, 1989).EtOH-related olfactory and taste memories can modulatelater EtOH consumption. For example, infantile experi-ences characterized by contingent presentations of alcoholodor and lithium or apomorphine toxicosis decrease alco-hol consumption even during adulthood (Molina et al.1985, 1986a,b).
Rat pups are also able to acquire associative memoriesinvolving alcohol as a US. Alcohol-mediated CTAs werefound in 16-day-old pups even with an alcohol dose as lowas 0.4 g/kg (Hunt et al., 1990). Tactile conditioned aver-sions also were found when a salient texture was pairedwith peak blood and brain alcohol concentrations derivedfrom a 2.0 g/kg EtOH dose (Molina et al., 1996). Theauthors also found that the hedonic content of the associa-tive memory was malleable. Specifically, the magnitude ofthe conditioned responses was affected when, after texture-alcohol pairings, the state of intoxication was associatedeither with an appetitive or an aversive stimulus. In theformer case (devaluation procedure in accordance with thelearning literature), tactile conditioned aversions were dra-
matically inhibited, whereas in the latter case (inflationprocedure), the magnitude of the aversive response wasstrengthened.
EtOH can also act as a US capable of supporting asso-ciative learning during prenatal life. Behavioral condi-tioned responses to an odor experienced in utero in conti-guity with alcohol intoxication have been recently reported(Abate et al., 2000). Abate et al. (2001) also found thatpups given prenatal pairings between the flavor of cineoleand alcohol intoxication later avoided consumption of acineole solution. As can be observed, the literature indi-cates with few exceptions that during early ontogeny, thehedonic content of the memory acquired when alcohol isused as a US is aversive [Molina et al., 1999; Pepino et al.,1999; but also see Cheslock et al. (2001)].
The aim of this study was to determine whether, duringearly stages of the rat’s ontogenetic development, the ef-fectiveness of EtOH in generating conditioned aversion tospecific stimuli might depend on when the stimuli arepresented during the course of the toxic episode. The strat-egy was to present a taste conditioned stimulus (CS) withina stage of intoxication that generally leads to a conditionedaversion. A completely different CS (a novel texture) waspresented during the onset of the stage of intoxication, aperiod in which we have failed to observe the generation ofa conditioned aversion (Brasser et al., 1993) and duringwhich appetitive effects of alcohol have been sometimesreported (Cunningham and Prather, 1992; Risinger andCunningham, 1992). It was uncertain whether this proce-dure would yield tactile appetitive and taste-aversive con-ditioned responses or whether EtOH’s aversive propertieswould prevail and, hence, both sensory cues would later berejected. It also seemed possible that the tactile and tastestimuli might act as cues that signal either absence (inhib-itory conditioning) or presence (excitatory conditioning) ofthe aversive effects of alcohol, respectively. Inhibitory con-ditioning is certainly a possibility when excitatory learningis being developed (Domjan and Burkhard, 1986; LoLordoand Fairless, 1985). In other words, acquisition of a saccha-rin CTA can provide the basis for inhibitory learning as afunction of alternative contingencies between a texture CSand EtOH as a US.
EXPERIMENT 1
Prior studies and the results of extensive pilot experi-ments were taken into account to determine optimal con-ditioning parameters for establishing alcohol-mediatedCTAs in infant rats. Fourteen-day-old pups were found toexhibit reliable taste aversions when two conditioning trialswere used that were defined by contingent presentations ofsaccharin (CS) and alcohol (US) (Pautassi et al., 2001). Theflavor CS was presented during an alcohol postadministra-tion time interval characterized by peak blood alcohol lev-els (Lopez et al., 1996).
INHIBITION OF ALCOHOL-MEDIATED TASTE AVERSIONS 645

In this experiment, a salient tactile cue was also pre-sented during the course of the toxic episode. The textureCS was presented early during the state of intoxication(5–10 min alcohol postadministration time). Prior studies(Brasser et al., 1993) found that when tactile cues werepaired with the onset of EtOH intoxication, conditionedaversion to the texture was not observed in the infant rat.
In summary, a tactile stimulus was presented relativelysoon after EtOH administration, and a gustatory stimuluswas presented later on. We expected that the organismwould learn to avoid the gustatory stimulus and prefer thetactile cue or to obtain behavioral evidence indicative thattaste signaled the presence of aversive consequences ofEtOH, whereas texture signaled the absence of these effects.
Methods
Subjects. Sixty-seven Wistar-derived pups representativeof 12 litters born and reared at the vivarium of the InstitutoFerreyra were used. Births were examined daily, and theday of parturition was considered as postnatal day 0 (PD0).During this day, pups were housed with their biologicalparents in standard maternity cages. Unless specified, allanimals had free access to water and food. Maternal cageswere filled with wood shavings that served as bedding.Temperature in the colony room was kept at 22 to 24°C,and a 12-hr light:dark cycle was used (artificial light onset,0800 hr). Animals were 14 days old (PD14) at the beginningof the experiment.
Experimental Design. The experimental approach was de-fined by two independent factors. One factor was the phar-macological treatment received by the pups during condi-tioning. They were intragastrically administered eitherEtOH or saline. The second factor consisted of the mannerin which animals were tested. There were three alternativetesting procedures: (a) a saccharin intake test took placebefore tactile preference assessments (I-T); (b) the tactiletest was executed before the intake test (T-I); or (c) sac-charin consumption was evaluated when infants were si-multaneously exposed to the texture CS (simultaneous con-dition; S).
Conditioning Procedures. One daily conditioning sessiontook place during PD14 and PD15. On each conditioningday (1200 hr), pups were withdrawn from their maternalcages, and a polyethylene cannula was implanted into thepup’s cheek. This cannulation procedure was similar to thatdescribed by various authors (Abate et al., 2000; Hall andRosenblatt, 1977; Hoffman and Spear, 1988). Briefly, can-nulae were made from 7-cm sections of polyethylene tubing(PE-10, Clay Adams, Parsippany, NJ). One end of thesedevices was heated to form a small flange. The unflangedend was attached to a curved 27-gauge 1⁄2 precision glideneedle (Becton Dickinson, Rutherford, NJ). The needlewas pulled through the medial internal surface of the rightcheek of the subject. Consequently, the flanged end of thecannulae rested over the oral mucosae, and the remainder
exited from the mouth cavity. Each animal was cannulatedin approximately 7 sec. This procedure has been shown togenerate minimal stress to the pups (Spear et al., 1989) andto represent a useful technique in studies involving orosen-sory stimulation (Pepino et al., 1999). In addition, previousexperiments have shown that during early postnatal life, ratpups are able to control ingestion of fluids delivered throughthis cannula (Domínguez et al., 1998; Pepino et al. 1998).
Immediately after the cannulation procedures, pups wereplaced in holding Plexiglas (Rohm and Haas Co., Philadel-phia, PA) cages (18 � 18 � 15 cm). Throughout theexperiment (conditioning and testing procedures), ambientroom temperature was maintained at 23 � 0.2°C. To avoidhypothermia derived from maternal separation, the holdingchambers were placed over a heating pad. Extensive pilotstudies conducted in this laboratory indicate that with thesethermal arrangements, the core temperatures of the infantsin the holding chambers are similar to those recorded in thenursing environment.
Four hours after the cannulation procedure, half of thesubjects received an intragastric administration of a 2.5 g/kgEtOH dose. This dose was achieved by administering 0.017ml of an 18.53% v/v alcohol solution (vehicle was physio-logical saline) per gram of body weight (190-proof alcohol,Porta Hnos). The remaining subjects received an equalvolume of the 0.89% w/v saline solution.
Immediately after EtOH or saline administration, sub-jects were returned to maintenance cages. Five minuteslater, pups were placed into opaque individual Plexiglasboxes equipped with a sandpaper floor (Doble A, BuenosAires, Argentina; waterproof, 50 granules per square cen-timeter), where they remained for 10 consecutive minutes.Pups were then returned to their maintenance cages and 10min later (25 min after drug administration) were placedinto clear individual cotton floor–covered boxes (15 � 7 �14 cm). In these chambers, pups were stimulated for 10 minwith intraoral infusions of a saccharin solution [0.05% w/v;commercially available Parker Davis Co. (Charlotte, NC)saccharin; vehicle, distilled water; temperature, 23°C]. Dur-ing this postadministration interval, EtOH reached peakblood alcohol levels in 15-day-old pups (Lopez et al., 1996).The infusion procedure took place after the anogenitalregion of the preweanling was gently stroked with a cottonswab to stimulate defecation and void the subject’s blad-ders. Pup body weights were registered to the nearest 0.01 g(Dial-O-Gram balance, Ohaus, Florham Park, NJ). Saccha-rin infusions were performed with a 10-way peristaltic in-fusion pump (Cassette R Pump, Manostat Corp, NewYork). Total administration volume was equivalent to 2.2%of the subject’s preinfusion body weight. After the infusionprocedure, subjects were again weighed to estimate saccha-rin consumption scores [percentage body weight gain (%BWG) was calculated as follows: postinfusion weight �preinfusion weight/preinfusion weight � 100]. After com-pletion of saccharin stimulation procedures, oral cannulae
646 PAUTASSI ET AL.
were removed, and pups were returned to their respectivematernity cages.
Testing Procedures. On PD16, oral cannulae were againimplanted into the cheeks of the animals. After this proce-dure, pups remained deprived of maternal care, as well asof food and water, for 8 hr. Testing procedures took placeimmediately after completion of this time interval.
Subjects that were administered EtOH or saline duringconditioning were randomly assigned to three groups ac-cording to the procedure of testing. One group was firstevaluated in terms of saccharin intake and subsequently ona two-way tactile location preference test (I-T; tests wereseparated by a 5-min period), and a second group wasevaluated by using the opposite order of testing procedures(T-I; tests were separated by a 5-min period). Finally, athird group of animals was tested for saccharin consump-tion patterns in a compartment with a sandpaper floorsurface (S). Detailed characteristics of the testing proce-dures are presented in the following paragraphs.
Tactile Location Preference Test. Tactile preference loca-tion tests were performed by individually placing the sub-jects into an opaque Plexiglas split-floor box (28 � 12 � 15cm). Half of the floor was lined with the tactile stimuluspreviously used during conditioning procedures (sandpa-per). The remaining floor surface was covered with theback side of a piece of sandpaper. Both textures werereplaced in each new test. The evaluation procedure lasted5 min. The test started by gently placing the animal in themiddle section of the apparatus. The activity of the animalwas recorded with an automatic system for data acquisitionand analysis of motor behavior (Ethovision, video trackingmotion analysis and behavior recognition system, NoldusInformation Technology, Wageningen, Holland). The de-pendent variable under consideration was the time spentover each particular tactile section of the apparatus. Toensure that the animal was over a given surface, the com-puterized program for recording behavior was set to con-sider the middle section of the apparatus (approximately15% of the entire surface) as a neutral area.
Saccharin Intake Test. The cannulae, surgically implanted8 hr before, were connected to the peristaltic infusionpump. Saccharin solution was infused during a 10-minperiod. Total volume infused was equivalent to 2.2% of thesubject’s preinfusion body weight. The test took place inclear individual cotton-floored Plexiglas cages. Postinfusionweights were recorded to establish the amount of saccharinintake (% BWG).
Saccharin Intake Test in the Presence of the Tactile (Sand-paper) CS. Under this testing condition, the intake test wassimilar to the one previously described. The only differencewas that that the test took place in a sandpaper-flooredPlexiglas cage (28 � 12 � 15 cm). Locomotive patterns(distance traveled) were recorded during the 10-min testthrough the use of the Ethovision automated system.
Results and Discussion
Relative increases in body weights (% BWG) during thesaccharin intake test were analyzed by using a two-wayanalysis of variance (ANOVA). The analysis recognizedtwo independent factors: drug treatment (EtOH or saline)and testing condition (IT, TI, or S).
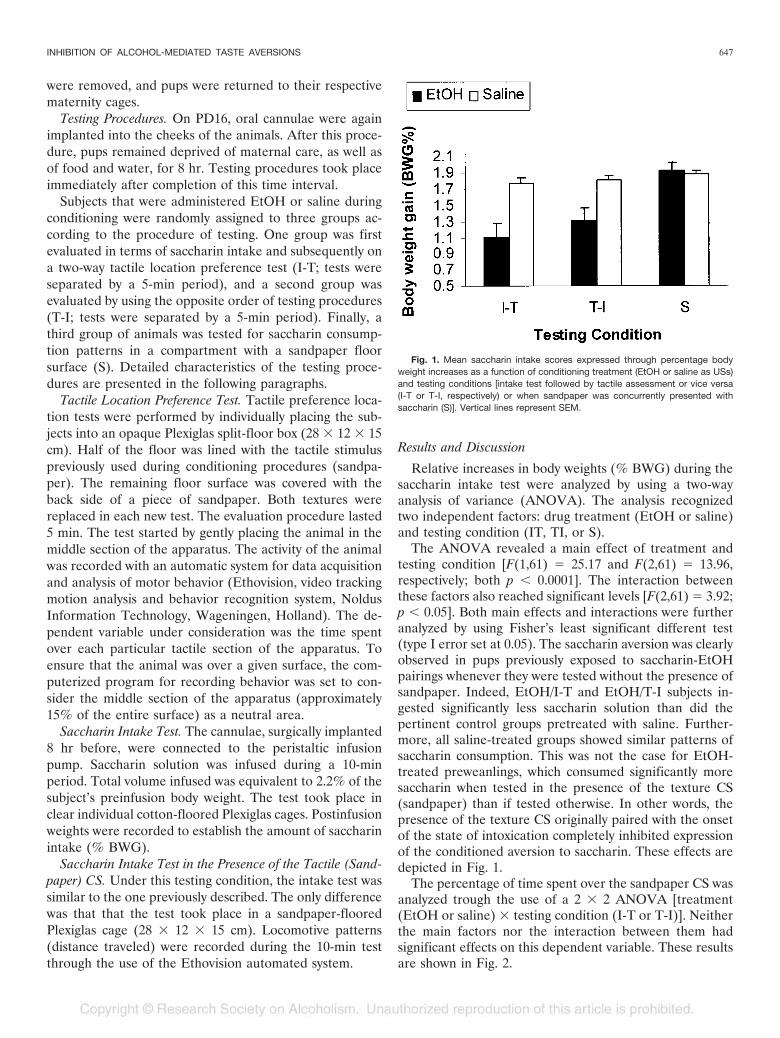
The ANOVA revealed a main effect of treatment andtesting condition [F(1,61) � 25.17 and F(2,61) � 13.96,respectively; both p � 0.0001]. The interaction betweenthese factors also reached significant levels [F(2,61) � 3.92;p � 0.05]. Both main effects and interactions were furtheranalyzed by using Fisher’s least significant different test(type I error set at 0.05). The saccharin aversion was clearlyobserved in pups previously exposed to saccharin-EtOHpairings whenever they were tested without the presence ofsandpaper. Indeed, EtOH/I-T and EtOH/T-I subjects in-gested significantly less saccharin solution than did thepertinent control groups pretreated with saline. Further-more, all saline-treated groups showed similar patterns ofsaccharin consumption. This was not the case for EtOH-treated preweanlings, which consumed significantly moresaccharin when tested in the presence of the texture CS(sandpaper) than if tested otherwise. In other words, thepresence of the texture CS originally paired with the onsetof the state of intoxication completely inhibited expressionof the conditioned aversion to saccharin. These effects aredepicted in Fig. 1.
The percentage of time spent over the sandpaper CS wasanalyzed trough the use of a 2 � 2 ANOVA [treatment(EtOH or saline) � testing condition (I-T or T-I)]. Neitherthe main factors nor the interaction between them hadsignificant effects on this dependent variable. These resultsare shown in Fig. 2.
Fig. 1. Mean saccharin intake scores expressed through percentage bodyweight increases as a function of conditioning treatment (EtOH or saline as USs)and testing conditions [intake test followed by tactile assessment or vice versa(I-T or T-I, respectively) or when sandpaper was concurrently presented withsaccharin (S)]. Vertical lines represent SEM.
INHIBITION OF ALCOHOL-MEDIATED TASTE AVERSIONS 647
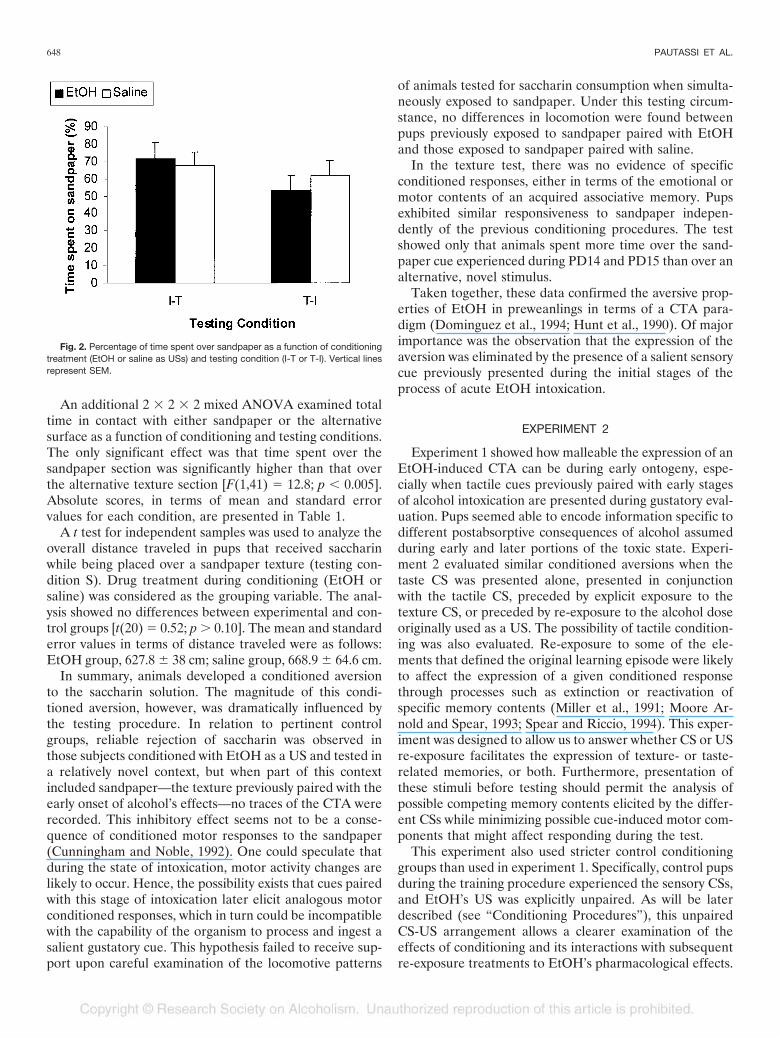
An additional 2 � 2 � 2 mixed ANOVA examined totaltime in contact with either sandpaper or the alternativesurface as a function of conditioning and testing conditions.The only significant effect was that time spent over thesandpaper section was significantly higher than that overthe alternative texture section [F(1,41) � 12.8; p � 0.005].Absolute scores, in terms of mean and standard errorvalues for each condition, are presented in Table 1.
A t test for independent samples was used to analyze theoverall distance traveled in pups that received saccharinwhile being placed over a sandpaper texture (testing con-dition S). Drug treatment during conditioning (EtOH orsaline) was considered as the grouping variable. The anal-ysis showed no differences between experimental and con-trol groups [t(20) � 0.52; p � 0.10]. The mean and standarderror values in terms of distance traveled were as follows:EtOH group, 627.8 � 38 cm; saline group, 668.9 � 64.6 cm.
In summary, animals developed a conditioned aversionto the saccharin solution. The magnitude of this condi-tioned aversion, however, was dramatically influenced bythe testing procedure. In relation to pertinent controlgroups, reliable rejection of saccharin was observed inthose subjects conditioned with EtOH as a US and tested ina relatively novel context, but when part of this contextincluded sandpaper—the texture previously paired with theearly onset of alcohol’s effects—no traces of the CTA wererecorded. This inhibitory effect seems not to be a conse-quence of conditioned motor responses to the sandpaper(Cunningham and Noble, 1992). One could speculate thatduring the state of intoxication, motor activity changes arelikely to occur. Hence, the possibility exists that cues pairedwith this stage of intoxication later elicit analogous motorconditioned responses, which in turn could be incompatiblewith the capability of the organism to process and ingest asalient gustatory cue. This hypothesis failed to receive sup-port upon careful examination of the locomotive patterns
of animals tested for saccharin consumption when simulta-neously exposed to sandpaper. Under this testing circum-stance, no differences in locomotion were found betweenpups previously exposed to sandpaper paired with EtOHand those exposed to sandpaper paired with saline.
In the texture test, there was no evidence of specificconditioned responses, either in terms of the emotional ormotor contents of an acquired associative memory. Pupsexhibited similar responsiveness to sandpaper indepen-dently of the previous conditioning procedures. The testshowed only that animals spent more time over the sand-paper cue experienced during PD14 and PD15 than over analternative, novel stimulus.
Taken together, these data confirmed the aversive prop-erties of EtOH in preweanlings in terms of a CTA para-digm (Dominguez et al., 1994; Hunt et al., 1990). Of majorimportance was the observation that the expression of theaversion was eliminated by the presence of a salient sensorycue previously presented during the initial stages of theprocess of acute EtOH intoxication.
EXPERIMENT 2
Experiment 1 showed how malleable the expression of anEtOH-induced CTA can be during early ontogeny, espe-cially when tactile cues previously paired with early stagesof alcohol intoxication are presented during gustatory eval-uation. Pups seemed able to encode information specific todifferent postabsorptive consequences of alcohol assumedduring early and later portions of the toxic state. Experi-ment 2 evaluated similar conditioned aversions when thetaste CS was presented alone, presented in conjunctionwith the tactile CS, preceded by explicit exposure to thetexture CS, or preceded by re-exposure to the alcohol doseoriginally used as a US. The possibility of tactile condition-ing was also evaluated. Re-exposure to some of the ele-ments that defined the original learning episode were likelyto affect the expression of a given conditioned responsethrough processes such as extinction or reactivation ofspecific memory contents (Miller et al., 1991; Moore Ar-nold and Spear, 1993; Spear and Riccio, 1994). This exper-iment was designed to allow us to answer whether CS or USre-exposure facilitates the expression of texture- or taste-related memories, or both. Furthermore, presentation ofthese stimuli before testing should permit the analysis ofpossible competing memory contents elicited by the differ-ent CSs while minimizing possible cue-induced motor com-ponents that might affect responding during the test.
This experiment also used stricter control conditioninggroups than used in experiment 1. Specifically, control pupsduring the training procedure experienced the sensory CSs,and EtOH’s US was explicitly unpaired. As will be laterdescribed (see “Conditioning Procedures”), this unpairedCS-US arrangement allows a clearer examination of theeffects of conditioning and its interactions with subsequentre-exposure treatments to EtOH’s pharmacological effects.
Fig. 2. Percentage of time spent over sandpaper as a function of conditioningtreatment (EtOH or saline as USs) and testing condition (I-T or T-I). Vertical linesrepresent SEM.
648 PAUTASSI ET AL.

Methods
Subjects. Seventy-five pups representative of nine litterswere used. Pups were 14 days old when the experimentbegan. Genetic and housing conditions of these animalswere similar to those described for experiment 1.
Experimental Design. A 2 (conditioning treatment) � 4(testing procedure) design was used. In terms of condition-ing treatments, paired (P) animals were those that experi-enced tactile and gustatory cues while intoxicated withalcohol. An explicitly unpaired control group (UP) was alsoincluded. In this UP group, pups experienced the state ofalcohol intoxication, and not until 6 hr later were theyexposed to the same sequential presentation of sandpaperand saccharin as used for P animals. In regard to theevaluation procedures, pups representative of P and UPconditions were assigned to one of four groups. A firstgroup received their saccharin intake test before beingevaluated for tactile preference (I-T group). A secondgroup was equivalent to the simultaneous testing conditiondescribed in experiment 1; i.e., saccharin intake was re-corded when sandpaper as the floor’s texture was concur-rently presented (S group), and these subjects were latertested for tactile preference. The two remaining test con-ditions also received a saccharin test followed by the test fortactile preference. One of these groups was exposed tosandpaper immediately before measurement of saccharinintake (sandpaper exposed; SE), whereas the second groupwas reintroduced to acute alcohol intoxication before thetests for intake and tactile preference (EtOH exposed; EE).
Conditioning Procedures. Conditioning procedures for Panimals replicated those used for the analogous group inexperiment 1. Briefly, during PD14 and PD15, pups wereexposed to a 2.5 g/kg alcohol dose (intragastric administra-tion). During postadministration time 5 to 15 min, theywere placed over a sandpaper surface, and during postad-ministration interval 25 to 35 min, they were intraorallystimulated with a 0.05% w/v saccharin solution. UP pupswere intragastrically administered the 2.5 g/kg EtOH dose6 hr before exposure to sandpaper and saccharin. Totaldurations of exposure to each stimulus, as well as durationsof the interstimulus intervals, were equivalent to thosedescribed in P pups. UP rats also received these treatmentsduring PD14 and PD15. The UP group [also referred to aslearning control in prior literature (Cunningham, 1993)] hascertain advantages relative to nondrug controls when un-conditioned effects of pharmacological agents capable ofsupporting type I associative learning are focused on. Anobvious advantage is the possibility of assessing the effect
of temporal contiguity between CSs and USs because P andUP groups are given identical sensory and pharmacologicalexperience but differ in terms of the temporal relationshipbetween them. Furthermore, changes in performance thatmight be due to the effect of the drug on various cognitiveprocesses (e.g., attention, memory encoding, and so on) areunder control when differences between P and UP groupsare examined. Another specific advantage for this experi-ment is control over the possibility of differential toleranceto the drug, because P and UP animals share similar his-tories in regard to alcohol’s effects.
Testing Procedures. Testing was conducted on PD16. Allgroups were evaluated in a saccharin intake test and, 5 minlater, in the texture preference assessment. The tests usedthe same instruments and procedures used in experiment 1.Hence, I-T pups were exposed to saccharin alone in thefirst test and later to the tactile preference test, for whichone section of the compartment was lined with sandpaperand the remaining section with the back side (soft card-board) of a piece of sandpaper. Preweanlings in the S groupexperienced saccharin while they were in contact with asandpaper floor. The tactile test was similar to the one usedin all the remaining groups. Subjects corresponding to theSE group were positioned over sandpaper for 10 min andthen were immediately placed into clear individual Plexi-glas chambers fitted with a cotton floor and given thesaccharin intake test. Subjects assigned to the EE testingcondition received an intragastric administration of a 2.5g/kg EtOH dose, 4 hr before assessment of saccharin in-take. With this alcohol dose and animals of a similar strainand age, we have observed complete clearance of alcohol inblood after a 4-hr postadministration interval (Lopez et al.,1996).
Results and Discussion
A 2 � 4 ANOVA [treatment (P or UP) � testing con-dition (I-T, SE, EE, or S)] was used to analyze % BWG inthe saccharin consumption test. The ANOVA showed sig-nificant main effects of treatment and testing condition[F(1,67) � 26.05, p � 0.001; and F(3,67) � 5.10, p � 0.005,respectively]. The interaction between these factors wasalso significant [F(3,67) � 4.00; p � 0.025]. Fisher post hoctests with an � set at 0.05 revealed that P/I-T pups dranksignificantly less saccharin than did the corresponding UP/I-T control group, establishing the CTA. All UP animalsdrank similar amounts of the taste CS.
The CTA was not manifested, however, when P pupswere exposed to sandpaper either before (SE) or during (S)
Table 1. Performance in the Tactile Location Test Regarding Absolute Time Spent Over Sandpaper and the Alternative Surface as a Function of Conditioning andTesting Procedures
Dependent variables
Conditioning and testing procedure
Saline/I-T Saline/T-I EtOH/I-T EtOH/T-I
Total time over sandpaper (sec) 169.4 � 25.4 160.2 � 11.4 195.3 � 26.2 135.2 � 15.9Total time over alternative surface (sec) 82.6 � 24.3 98.3 � 11.5 76.8 � 25.6 120.3 � 15.5
Data are presented as means � SEM.
INHIBITION OF ALCOHOL-MEDIATED TASTE AVERSIONS 649
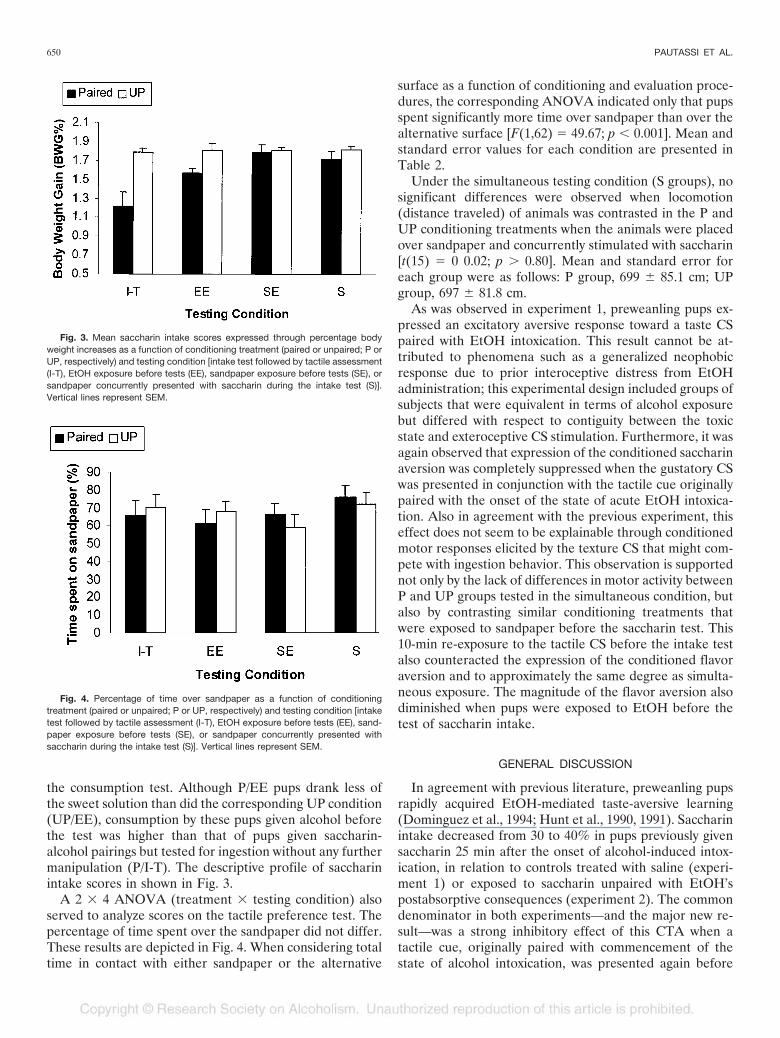
the consumption test. Although P/EE pups drank less ofthe sweet solution than did the corresponding UP condition(UP/EE), consumption by these pups given alcohol beforethe test was higher than that of pups given saccharin-alcohol pairings but tested for ingestion without any furthermanipulation (P/I-T). The descriptive profile of saccharinintake scores in shown in Fig. 3.
A 2 � 4 ANOVA (treatment � testing condition) alsoserved to analyze scores on the tactile preference test. Thepercentage of time spent over the sandpaper did not differ.These results are depicted in Fig. 4. When considering totaltime in contact with either sandpaper or the alternative
surface as a function of conditioning and evaluation proce-dures, the corresponding ANOVA indicated only that pupsspent significantly more time over sandpaper than over thealternative surface [F(1,62) � 49.67; p � 0.001]. Mean andstandard error values for each condition are presented inTable 2.
Under the simultaneous testing condition (S groups), nosignificant differences were observed when locomotion(distance traveled) of animals was contrasted in the P andUP conditioning treatments when the animals were placedover sandpaper and concurrently stimulated with saccharin[t(15) � 0 0.02; p � 0.80]. Mean and standard error foreach group were as follows: P group, 699 � 85.1 cm; UPgroup, 697 � 81.8 cm.
As was observed in experiment 1, preweanling pups ex-pressed an excitatory aversive response toward a taste CSpaired with EtOH intoxication. This result cannot be at-tributed to phenomena such as a generalized neophobicresponse due to prior interoceptive distress from EtOHadministration; this experimental design included groups ofsubjects that were equivalent in terms of alcohol exposurebut differed with respect to contiguity between the toxicstate and exteroceptive CS stimulation. Furthermore, it wasagain observed that expression of the conditioned saccharinaversion was completely suppressed when the gustatory CSwas presented in conjunction with the tactile cue originallypaired with the onset of the state of acute EtOH intoxica-tion. Also in agreement with the previous experiment, thiseffect does not seem to be explainable through conditionedmotor responses elicited by the texture CS that might com-pete with ingestion behavior. This observation is supportednot only by the lack of differences in motor activity betweenP and UP groups tested in the simultaneous condition, butalso by contrasting similar conditioning treatments thatwere exposed to sandpaper before the saccharin test. This10-min re-exposure to the tactile CS before the intake testalso counteracted the expression of the conditioned flavoraversion and to approximately the same degree as simulta-neous exposure. The magnitude of the flavor aversion alsodiminished when pups were exposed to EtOH before thetest of saccharin intake.
GENERAL DISCUSSION
In agreement with previous literature, preweanling pupsrapidly acquired EtOH-mediated taste-aversive learning(Dominguez et al., 1994; Hunt et al., 1990, 1991). Saccharinintake decreased from 30 to 40% in pups previously givensaccharin 25 min after the onset of alcohol-induced intox-ication, in relation to controls treated with saline (experi-ment 1) or exposed to saccharin unpaired with EtOH’spostabsorptive consequences (experiment 2). The commondenominator in both experiments—and the major new re-sult—was a strong inhibitory effect of this CTA when atactile cue, originally paired with commencement of thestate of alcohol intoxication, was presented again before
Fig. 3. Mean saccharin intake scores expressed through percentage bodyweight increases as a function of conditioning treatment (paired or unpaired; P orUP, respectively) and testing condition [intake test followed by tactile assessment(I-T), EtOH exposure before tests (EE), sandpaper exposure before tests (SE), orsandpaper concurrently presented with saccharin during the intake test (S)].Vertical lines represent SEM.
Fig. 4. Percentage of time over sandpaper as a function of conditioningtreatment (paired or unpaired; P or UP, respectively) and testing condition [intaketest followed by tactile assessment (I-T), EtOH exposure before tests (EE), sand-paper exposure before tests (SE), or sandpaper concurrently presented withsaccharin during the intake test (S)]. Vertical lines represent SEM.
650 PAUTASSI ET AL.

(experiment 2) or during (experiments 1 and 2) the saccha-rin intake test. Expression of saccharin-conditioned aver-sions was also partially inhibited when pups were re-exposed to the toxic effects of EtOH before the saccharinintake test (experiment 2).
There are several questions about these results thatshould be considered. Are they due to nonspecific interfer-ence exerted by processing of competing stimuli before orduring the saccharin test? This alternative seems to receivelittle support when features of the experimental designs areconsidered. Whenever a tactile test was given before thetest for saccharin consumption (experiment 1), the CTAwas not affected in terms of its magnitude relative to that ofpups first evaluated in the consumption test. Furthermore,experience with EtOH’s US properties before the test forsaccharin consumption (experiment 2) did not suppress theexpression of the taste aversion response.
Is it possible that differential affective properties of al-cohol during the course of the toxic state determine tactilepreferences as well as saccharin aversions that competeduring memory retrieval processes? A negative answerseems to be provided by the tactile assessments in bothexperiments. We completely failed to observe any behav-ioral evidence of appetitive learning mediated by EtOHwhen it was paired with a salient tactile cue. As shown indifferent studies performed with rats, subnarcoleptic alco-hol doses have more often supported aversive than appet-itive associative learning in regard to locations character-ized by specific tactile features, visual features, or both(Cunningham et al., 1993; Cunningham and Niehus, 1993;Schechter and Krimmer, 1992). Despite these observations,it is important to note that under these experimental cir-cumstances, sandpaper preference scores in control ani-mals in both experiments ranged from 60 to 70%. Do thesevalues represent a ceiling effect that does not allow us toobserve appetitive conditioning in pertinent experimentalgroups? Although this possibility cannot be completely dis-regarded, a recent set of experiments seems to indicate thatthe absence of appetitive tactile conditioning does not obeyparametric issues related with baseline preference scores.When using baseline tactile preference scores that rangedfrom 25 to 45%, we have also failed to encounter reliableexpression of learned tactile preferences when using con-ditioning treatments similar to those used here (Pautassi etal., 2000).
Conditioned locomotor activation induced by exterocep-tive contextual stimuli, as well as by specific tactile cues
paired with EtOH, has been reported previously (Cunning-ham and Noble, 1992). Such an effect could conceivablyexplain the lack of evidence for tactile conditioning, dis-ruptions in the expression of saccharin-conditioned aver-sions, or both when a tactile cue was simultaneously pre-sented. We could find no experimental evidence to supportthis hypothesis. No changes in locomotive patterns wereobserved in the response of animals to a sandpaper texturewhen they were simultaneously infused with saccharin. In-dependently of prior conditioning history, all pups testedunder simultaneous tactile-flavor conditions showed verysimilar rates of motor activity. Furthermore, the inhibitoryeffect of sandpaper exposure on the expression of saccharinaversions also occurred when the texture cue was presentedbefore and not during the intake test.
These effects seem to be explained better in terms ofinhibitory and excitatory conditioning associated withEtOH’s support of aversive conditioning. In other words, itseems that the results argue in favor of recruitment ofmemories that differ in relation to the information theyprovide regarding EtOH’s aversive properties. Due to dif-ferential temporal arrangements between the CSs and aver-sive effects of the drug that are more readily perceivedwhen the intensity of the US is maximal, it is likely that thetactile cue and the saccharin taste predicted the absenceand presence, respectively, of such effects. Beyond thecontiguity factor, biological affinity between chemosensorycues and interoceptive distress might aid in the occurrenceof excitatory aversive conditioning with saccharin ratherthan a tactile cue (Domjan, 1985; Gemberling and Domjan,1982; Klein and Mowrer, 1989). Under this hypothesis it islogical to expect that the magnitude of the conditionedsaccharin aversion will be higher when the taste is pre-sented alone than when it is simultaneously presented witha tactile cue that originally signaled the absence of aversiveEtOH-derived consequences. This indeed was the case; i.e.,simultaneous testing conditions that involved concurrentpresentation of the tactile and gustatory CSs resulted incomplete inhibition of aversive conditioning.
In other words, subjects seem to learn that wheneversandpaper is present, saccharin-EtOH association is not inforce (Rescorla, 1985). The simultaneous testing conditionresembles a summation test, a procedure normally used todetermine whether a given stimulus has acquired the capa-bility to signal the forthcoming absence of a biologicallyrelevant stimulus (Domjan and Burkhard, 1986; LoLordoand Fairless, 1985). Conditioned inhibition is measured in
Table 2. Performance in the Tactile Location Test Regarding Absolute Time Spent Over Sandpaper and the Alternative Surface as a Function of Conditioning andTesting Procedures
Dependent variables
Conditioning and testing procedure
UP/IT UP/EE UP/SE UP/S P/IT P/EE P/SE P/S
Total time over sandpaper(sec)
196.6 � 22.1 183.5 � 15.8 163.3 � 20.8 181.7 � 20.4 166.3 � 31.7 162.7 � 25.4 172.4 � 18.6 206.8 � 23.7
Total time over alternativesurface (sec)
82.44 � 20.9 86.2 � 14.8 111.3 � 18.2 65.9 � 14.1 76.6 � 20.9 93.2 � 18.8 85.9 � 15.5 57.6 � 13.3
Data are presented as means � SEM.
INHIBITION OF ALCOHOL-MEDIATED TASTE AVERSIONS 651

terms of reduction of the magnitude of an excitatorylearned response when a CS that signals the absence ratherthan presence of a US is introduced in the test situation.
The inhibitory effect of the texture CS also occurredwhen this stimulus was perceived alone for a relatively longperiod immediately before the saccharin test. This suggeststhe reactivation of memories involving cues that signal theabsence of aversive consequences of EtOH. Notice thatwhen the texture CS was presented for a shorter period oftime and 5 min elapsed between that CS and the intake test,and an alternative tactile stimulus was available during thetexture preference tests (group T-I, experiment 1), no ef-fects were obtained in terms of the magnitude of the taste-aversive response. The amount of exposure to a given cueoriginally involved in the learning situation, the delay ex-isting between re-exposure to this cue and the testing situ-ation, and the similarity between the nature of the elementsthat define the learning situation and the re-exposure treat-ment represent critical variables in terms of determiningreactivation processes of the memory under analysis (Mill-er et al., 1991; Moore Arnold and Spear, 1993; Richardsonet al. 1993).
It is uncertain whether consideration of conditioned in-hibition is the most useful way to view these phenomenaand whether conditioned inhibition will be verified in thesecircumstances in further experiments that include a retar-dation test. Briefly, this procedure implies that a cue (CS)will consistently be presented when a given US is absent.Subsequently, this CS is explicitly paired with the US underconsideration. Inhibitory conditioning is inferred wheneverthe rate of acquisition of excitatory conditioning is retardedin relation to organisms that never experienced the CS inabsence of the US. It is important to observe that sometheorists have argued that the retardation test is not nec-essary to establish conditioned inhibition (Holland, 1985).
If inhibitory and excitatory conditioning processes occuras a function of the course of the state of intoxication, thenit is conceivable that reactivation of competing memoriesencoded during such a state could also lead to attenuationof the excitatory conditioned response under analysis. Post-conditioning presentation of the US, as well as presentationof an apparently inhibitory CS, has been shown to reacti-vate associative learning processes that otherwise seem tobe forgotten (Miller et al., 1991). Memories under analysisin these experiments—in particular, the conditioned aver-sion to saccharin—seemed to be readily accessible in termsof retrieval. As previously mentioned, when this is the case,presentation of an inhibitory CS tends to attenuate themagnitude of the excitatory conditioned response. Beyondthese considerations, it should not be discarded that theattenuation exerted by pretest exposure to the US (groupP/EE, experiment 2) could also be due to a decrease in thecontingency between the US and the excitatory CS causedby presentation of the US alone (Williams, 2001).
Further experiments are required to determine whetherthe inhibitory capability of the tactile CS is determined by
experiencing this sensory stimulus within a given stage ofthe process of intoxication or whether it is simply caused bythe fact that the CS is presented in the absence of EtOH-induced aversive effects. This question implies analysis ofthe inhibitory capabilities of a given stimulus when it isperceived just before alcohol exposure or during differentpostabsorptive time intervals. This experimental approachshould help determine whether there are specific temporalwindows within the process of intoxication that aid in theestablishment of differential memories concerning a par-ticular postabsorptive effect. In addition, taking into ac-count ontogenetic variations in alcohol metabolism and interms of sensitivity to different sensorimotor and cognitiveeffects of the drug (Hollstedt and Rydberg, 1985; Lopez etal., 1996; Silveri and Spear, 2000), it seems important toinvestigate whether these findings are observed during peri-adolescent or adult stages of development.
These results argue in favor of the malleability of EtOH-related memories established through the drug’s aversiveproperties in infant rats. Postconditioning manipulations ofthe elements that define the original learning situationseem likely to modify expression of the hedonic contents ofsuch a memory. To our knowledge, this represents the firstset of results showing that a discrete sensory stimulus,experienced during the onset of the state of intoxication,will later completely prevent expression of the aversiverepresentation of the drug’s effects. Does this eliminationof memory expression emerge from a temporal contiguityfactor that allows learning about the absence of EtOH’saversive consequences, a conditioned inhibition, or arethere specific early postabsorptive consequences of thestate of intoxication that aid in the establishment of theeffect? The need to address this particular question isrelevant to the following issues. Differential motivationalproperties of alcohol, as is the case with most substances ofabuse, represent critical factors in the modulation of drug-seeking and -taking behaviors. Moreover, it is generallyadmitted that aversive consequences of drugs decrease thesubsequent probability of the occurrence of such behaviors(Cunningham, 1998). Hence, comprehension of factors thatmight inhibit or exacerbate the hedonic representation ofalcohol seems necessary to understand the mechanismsthat determine seeking and self-administration processes.The development of experimental procedures for control ofthe acquisition, expression, or both of memories defined byalcohol’s aversive components could aid in the analysis ofpossible positive- and negative-reinforcement capabilitiesof alcohol. This strategy should also provide a tool that caneasily be used across different ontogenetic stages of devel-opment—an opportunity that is particularly importantwhen considering recent studies that suggest substantialappetitive reinforcing properties of alcohol during veryearly postnatal life (Cheslock et al. 2000, 2001; Petrov et al.,1999).
652 PAUTASSI ET AL.

ACKNOWLEDGMENTS
The authors thank Teri Tanenhaus and Anahí Bulacio for theirtechnical assistance.
REFERENCES
Abate P, Pepino MY, Domínguez HD, Spear NE, Molina JC (2000) Fetalassociative learning mediated through maternal alcohol administration.Alcohol Clin Exp Res 24:39–47.
Abate P, Spear NE, Molina JC (2001) Fetal and infantile alcohol-mediated associative learning in the rat. Alcohol Clin Exp Res 25:989–998.
Arregui-Aguirre A, Claro-Izaguirre F, Goni-Garrido MJ, Zarate-OleagaJA, Morgado-Bernal I (1987) Effects of acute nicotine and ethanol onmedial prefrontal cortex self-stimulation in rats. Pharmacol BiochemBehav 27:15–20.
Bozarth M (1990) Evidence for the rewarding effects of ethanol using theconditioned place preference method. Pharmacol Biochem Behav 35:485–487.
Brasser SM, Mckinzie DL, Molina JC, Spear NE (1993) Ethanol intoxi-cation promotes odor and texture aversions in the infant rat. Presentedat the 65th annual meeting of the Midwestern Psychological Associa-tion, p. 56. Chicago, IL., April 29. May 1.
Cappel H, Le Blanc A (1973) Aversive conditioning by psychoactive drugs:effects of morphine, alcohol and chlordiazepoxide. Psychopharmacol-ogy 29:239–246.
Cheslock S, Varlinskaya E, Petrov ES, Silveri MM, Spear LP, Spear NE(2001) Ethanol as a reinforcer in the newborn’s first suckling experi-ence. Alcohol Clin Exp Res 25:395–402.
Cheslock SJ, Varlinskaya EI, Silveri MM, Petrov ES, Spear LP, Spear NE(2000) Acute effects of ethanol and the first suckling episode in thenewborn rat. Alcohol Clin Exp Res 24:996–1002.
Chester JA, Cunningham CL (1999) Baclofen alters ethanol-stimulatedactivity but not conditioned place preference or taste aversion in mice.Pharmacol Biochem Behav 63:325–331.
Cicocciopo R, Panocka I, Froldi R, Quitadamo E, Mazzi M (1999) Eth-anol induces conditioned place preference in genetically selectedalcohol-preferring rats. Psychopharmacology 141:235–241.
Cunningham CL (1993) Pavlovian drug conditioning, in Methods in Be-havioral Pharmacology (Vaan Haaren F ed), chapt 14, pp 349–381.Elsevier, Amsterdam.
Cunningham CL (1998) Drug conditioning and seeking behavior, inLearning and Behavior Therapy (O’Donohue W ed), pp 518–540. Allynand Bacon, Boston.
Cunningham CL, Fidler TL, Hill K (2000) Animal models of alcohol’smotivational effects. Alcohol Res Health 24:85–92.
Cunningham CL, Hallet C, Niehus D, Hunter J, Nouth L, Risinger F(1991) Assessment of ethanol’s hedonic effects in mice selectively bredfor sensitivity to ethanol-induced hypothermia. Psychopharmacology105:84–92.
Cunningham CL, Hawks D, Niehus D (1988) Role of hypothermia inethanol-induced conditioned taste aversion. Psychopharmacology 95:318–322.
Cunningham CL, Ignatoff DL (2000) Genetic differences in ethanol in-duced conditioned place aversion (abstract). Alcohol Clin Exp Res24:57a.
Cunningham CL, Niehus J, Bachtold J (1992) Ambient temperature ef-fects on taste aversion conditioned by ethanol: contribution of ethanol-induced hypothermia. Alcohol Clin Exp Res 16:117–124.
Cunningham CL, Niehus JS, Noble D (1993) Species difference in sensi-tivity to ethanol’s hedonic effects. Alcohol 10:97–102.
Cunningham CL, Noble D (1992) Conditioned activation induced byethanol: role in sensitization and conditioned place preference. Phar-macol Biochem Behav 43:307–313.
Cunningham CL, Prather LK (1992) Conditioning trial duration affectsethanol-induced conditioned place preference in mice. Anim LearnBehav 20:187–194.
Cunningham CL, Tull LE (2000) Tolerance to ethanol’s aversive effect butnot sensitization to its rewarding effect (abstract). Alcohol Clin Exp Res24:57a.
Dominguez HD, Bocco G, Chotro MG, Spear NE, Molina JC (1994)Aversions to alcohol’s orosensory cues in infant rats: generalization tocompounds of alcohol with sucrose or sodium chloride. Alcohol 2:225–233.
Domínguez HD, López MF, Molina JC (1998) Neonatal responsiveness toalcohol odor and infant alcohol intake as a function of alcohol experi-ence during late gestation. Alcohol 16:109–117.
Domjan M (1985) Cue-consequence specificity and long-delay learning.Ann NY Acad Sci 443:54–66.
Domjan M, Burkhard B (1986) The Principles of Learning and Behavior.2nd ed. Brooks/Cole, Belmont, CA.
Gauvin DV, Holloway FA (1992) Historical factors in the development ofEtOH-conditioned place preference. Alcohol 9:1–7.
Gemberling G, Domjan M (1982) Selective associations in one-day-oldrats: taste-toxicosis and texture-shock aversion learning. J Comp PhysiolPsychol 96:105–113.
Grigson PS (1997) Conditioned taste aversions and drugs of abuse: areinterpretation. Behav Neurosci 111:129–136.
Grigson PS (2000) Drugs of abuse and reward comparison: a brief review.Appetite 35:89–91.
Hall WA, Rosenblatt JS (1977) Suckling behavior and intake control inthe developing rat pup. J Comp Physiol Psychol 91:1232–1247.
Hine B, Figueroa B (1991) Low-dose, rapid facilitation of brain stimula-tion reward by ethanol. Res Commun Subst Abuse 12:59–62.
Hoffman H, Spear NS (1988) Ontogenetic differences in conditioning ofan aversion to a gustatory CS with a peripheral US. Behav Neural Biol50:16–23.
Holland PC (1985) The nature of condition inhibition in serial and simul-taneous feature negative discriminations, in Information Processing inAnimals: Conditioned Inhibition (Miller RR, Spear NE eds). Erlbaum,Hillsdale, NJ.
Hollstedt C, Rydberg U (1985) Postnatal effect of ethanol on the devel-oping rat, in Alcohol and the Developing Brain (Rydberg U, Engel J eds),pp 69–84. Raven Press, New York.
Hunt PS, Molina JC, Spear LP, Spear LE (1990) Ethanol-mediated tasteaversions and state dependency in preweanling (16-day-old-rats). BehavNeural Biol 54:300–322.
Hunt PS, Spear LP, Spear NE (1991) An ontogenetic comparison ofethanol-mediated taste aversion learning and ethanol-induced hypo-thermia in preweanling rats. Behav Neurosci 105:971–983.
Klein SB, Mowrer RR (1989) Contemporary Learning Theories: Instrumen-tal Conditioning Theory and the Impact of Biological Constraints onLearning. Erlbaum, Mahwah, NJ.
Lett BT (1988) Enhancement of conditioned preference for a place pairedwith amphetamine produced by blocking the association between placeand amphetamine-induced sickness. Psychopharmacology 95:390–394.
LoLordo VM, Fairless JL (1985) Pavlovian conditioned inhibition: theliterature since 1969, in Information Processing in Animals: ConditionedInhibition (Miller RR, Spear NE eds), pp 1–149. Erlbaum, Hillsdale, NJ.
Lopez MF, Spear NE, Molina JC (1996) Ontogenetic differences in theexpression of olfactory-conditioned aversions resulting from a state ofacute alcohol intoxication in the rat. Alcohol 13:473–481.
Marglin SH, Mackchenie DK, Mattie M, Hui Y, Reid LD (1988) Ethanolwith small doses of morphine establishes a conditioned place prefer-ence. Alcohol 5:309–313.
Miller JS, Jagielo JA, Spear NE (1991) Differential effectiveness of vari-ous prior-cuing treatments in the reactivation and maintenance ofmemory. J Exp Psychol Anim Behav Process 17:249–258.
Molina JC, Bannoura MD, Chotro MG, Mckinzie DL, Moore A, SpearNE (1996) Alcohol-mediated tactile conditioned aversions in infants
INHIBITION OF ALCOHOL-MEDIATED TASTE AVERSIONS 653

rats: devaluation of conditioning through alcohol-sucrose associations.Neurobiol Learn Mem 66:121–132.
Molina JC, Chotro MG (1989) Acute alcohol intoxication paired withaversive reinforcement: ethanol odor as a conditioned reinforcer in ratspups. Behav Neural Biol 52:1–19.
Molina JC, Domínguez HD, López MF, Pepino MY, Faas AE (1999) Therole of fetal and infantile experience with alcohol in later recognitionand acceptance patterns of the drug, in Alcohol and Alcoholism. Effectson Brain and Development (Hanningan JH, Spear LP, Spear NS,Goodlett CR eds), pp 199–228. Erlbaum, Mahwah, NJ.
Molina JC, Hoffman H, Spear NE (1986a) Conditioning of aversion toalcohol orosensory cues in 5- and 10-day-old rats: subsequent reductionin alcohol ingestion. Dev Psychobiol 19:175–183.
Molina JC, Serwatka J, Spear N (1986b) Alcohol drinking patterns ofyoung adult rats as a function of infantile aversive experiences withalcohol odor. Behav Neural Biol 46:257–271.
Molina JC, Serwatka J, Spear L, Spear N (1985) Differential ethanololfactory experiences affect ethanol ingestion in preweanlings but not inolder rats. Behav Neural Biol 44:90–100.
Moore Arnold H, Spear NE (1993) Order and duration of stimuli areimportant determinants of reactivation. Anim Learn Behav 21:391–398.
Pautassi RM, Godoy JC, Molina JC (2000) Generación de asociacionesmúltiples durante el rranscurso de un mismo episodio tóxico en latemprana ontogenia de la rata. Prceedings of the 3rd Congress of theSpanish Society of Experimental Psychology, p. 18. Barcelona, Spain,March 2–4.
Pautassi RM, Godoy JC, Molina JC (2001) Unconditional properties ofethanol during the course of an acute intoxication: effects of prenatalethanol exposure (abstract). Cell Mol Neurobiol 21:154.
Pepino MY, Kraebel KS, López MF, Spear NE, Molina JC (1998) Behav-ioral detection of low concentrations of alcohol in milk in the prewean-ling rat. Alcohol 15:337–353.
Pepino MY, López MF, Spear NE, Molina JC (1999) Infant rats responddifferentially to alcohol after nursing from an alcohol-intoxicated dam.Alcohol 18:189–201.
Petrov ES, Varlinskaya EI, Bregman K, Smotherman WP (1999) Sus-tained attachment in the newborn rat depends on experience with thenipple and expression of oral grasping. Behav Neurosci 113:211–221.
Reicher MA, Holman EW (1977) Location preference and flavor aversionreinforced by amphetamine in rats. Anim Learn Behav 5:343–346.
Reid LD, Hunter GA, Beaman CW, Hubbel CL (1985) Toward under-standing ethanol’s capacity to be reinforcing: a conditioned place pref-
erence following injections of alcohol. Pharmacol Biochem Behav 22:483–487.
Rescorla R (1985) Conditioned inhibition and facilitation, in InformationProcessing in Animals: Conditioned Inhibition (Miller RR, Spear NEeds), pp 1–149. Erlbaum, Hillsdale, NJ.
Richardson R, Wang P, Campbell BA (1993) Reactivation of nonassocia-tive memories. Dev Psychobiol 26:1–23.
Risinger FO, Cunningham CL (1992) Ethanol produces rapid biphasichedonic effects, in The Neurobiology of Drug and Alcohol Addiction, Vol.654 (Kalivas P, Samson H, eds), pp 506–508. Annals of the New YorkAcademy of Sciences. New York, NY.
Schaefer GJ, Michael RP (1987) Ethanol and current thresholds for brainself-stimulation in the lateral hypothalamus of the rat. Alcohol 4:209–213.
Schechter MD, Krimmer EC (1992) Difference in response to the aversiveproperties and activity effects of low doses of ethanol in LAS and HASselectively bred rats. Psychopharmacology 107:564–568.
Silveri MM, Spear LP (2000) Ontogeny of ethanol elimination andethanol-induced hypothermia. Alcohol 20:45–53.
Spear NE, Riccio DC (1994) Memory: Phenomena and Principles. Allynand Bacon, Boston.
Spear LP, Specht SM, Kirstein CL, Kuhn CM (1989) Anterior and pos-terior, but not cheek, intraoral cannulation procedures elevate serumcorticosterone levels in neonatal rat pups. Dev Psychobiol 22:401–411.
Stewart RB, Grupp L (1985) Some determinants of the motivationalproperties of ethanol in the rat: concurrent administration of food orsocial stimuli. Psychopharmacology 87:43–50.
Stewart RB, Li T-K (1997) The neurobiology of alcoholism in geneticallyselected rat models. Alcohol Health Res World 21:169–176.
Stewart RB, Perlanski E, Tech D, Grupp LA (1988) Ethanol as a rein-forcer for rats: factors of facilitation and constraint. Alcohol Clin ExpRes 12:599–608.
US Department of Health and Human Services (2000) 10th Special Reportto the U.S Congress on Alcohol and Health. National Institutes of Health,National Institute of Alcohol Abuse and Alcoholism, Bethesda, MA.
Van der Kooy D, O’Shaughnessy M, Mucha RF, Kalant H (1983) Moti-vational properties of ethanol in naive rats as studied by place condi-tioning. Pharmacol Biochem Behav 19:441–445.
Williams BA (2001) The critical dimensions of the response-reinforcercontingency. Behav Processes 54:111–126.
Wise RA, Yokel RA, De Witt H (1976) Both positive reinforcement andconditioned aversion from amphetamine and from apomorphine in rats.Science 191:1273–1275.
654 PAUTASSI ET AL.
Related Documents











