Molecular and Cellular Pathobiology Dysregulation of RUNX2/Activin-A Axis upon miR-376c Downregulation Promotes Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma Wei-Min Chang 1,2,3 , Yuan-Feng Lin 4 , Chia-Yi Su 3 , Hsuan-Yu Peng 2 , Yu-Chan Chang 3 , Tsung-Ching Lai 3 , Guan-Hsun Wu 2 , Yuan-Ming Hsu 2 , Li-Hsing Chi 3,5,6 , Jenn-Ren Hsiao 7 , Chi-Long Chen 8,9 , Jang-Yang Chang 2,10 , Yi-Shing Shieh 1,11 , Michael Hsiao 3,12 , and Shine-Gwo Shiah 2,11 Abstract Epigenetic correlates of the head and neck cancer may illumi- nate its pathogenic roots. Through a gene set enrichment analysis, we found that the oncogenic transcription factor RUNX2 is widely upregulated in the head and neck squamous cell carcinoma (HNSCC) with lymph node metastasis, where it also predicts poor prognosis in patients with HNSCC. Enforced expression of ectopic RUNX2 promoted the metastatic capabilities of HNSCC, whereas RUNX2 silencing inhibited these features. Mechanistic investigations showed that manipulating levels of activin A (INHBA) could rescue or compromise the RUNX2-mediated metastatic capabilities of HNSCC cells. Furthermore, we found that miR-376c-3p encoded within the 3 0 -untranslated region of RUNX2 played a pivotal role in regulating RUNX2 expression in highly metastatic HNSCC cells, where it was downregulated commonly. Restoring miR-376c expression in this setting sup- pressed expression of RUNX2/INHBA axis along with metastatic capability. Clinically, we observed an inverse relationship between miR-376c-3p expression and the RUNX2/INHBA axis in HNSCC specimens. In summary, our results defined a novel pathway in which dysregulation of the RUNX2/INHBA axis due to miR-376c downregulation fosters lymph node metastasis in HNSCC. Cancer Res; 76(24); 1–11. Ó2016 AACR. Introduction Head and neck squamous cell carcinoma (HNSCC) is one of highly prevalent and lethal cancers in the world, including Taiwan (1, 2). More than 90% of HNSCCs are oral squamous cell carcinomas, which arise in the oral cavity (3). The initiation and progression of HNSCCs are highly correlated with the exposure to carcinogens, such as cigarette and alcohol. In southeast Asia and Taiwan, betel chewing has been considered as a major risk factor for HNSCC (3). Although cancer therapies have tremendously improved over the past 20 years, the prognosis of patients with HNSCC remains unfavorable (4, 5). Several recent studies have reported single-molecule biomarkers, such as IGF1R (6), Axl (7), and EpCAM (8) for predicting HNSCC progression; however, their practical usefulness remains to be determined. By using genome-wide analysis, this study was thus attempted to identify the critical molecule(s) as a diagnostic biomarker or even ther- apeutic target for metastatic HNSCC. RUNX2, a Runt family transcription factor, is a key regulator physiologically associated with osteoblast differentiation and chondrocyte maturation (9, 10), as well as cytoskeleton remo- deling and cellular movement via activating its downstream genes. Recently, RUNX2 plays as a pivotal molecule in forcing breast and prostate cancer progression, for example, bone metastasis, through the transcriptional regulation of MMPs, VEGFs, SNAIL, SLUG, and PTHrP expression (11). Neverthe- less, the functional consequences of RUNX2 upregulation and its downstream transcriptome are still unclear in HNSCC progression. 1 Graduate Institute of Medical Sciences, National Defense Medical Center, Taipei, Taiwan. 2 National Institute of Cancer Research, National Health Research Institutes, Miaoli, Taiwan. 3 Genomics Research Center, Academia Sinica, Taipei, Taiwan. 4 Graduate Institute of Clinical Medicine, College of Medicine, Taipei Medical University, Taipei, Taiwan. 5 The PhD Program for Translational Medicine, Taipei Medical University and Academia Sinica, Taipei, Taiwan. 6 Division of Oral and Maxillofacial Surgery, Department of Dentistry, Taipei Medical University Hospital, Taipei, Taiwan. 7 Department of Otolaryngology, Head and Neck Col- laborative Oncology Group and National Cheng-Kung University Hospital, College of Medicine, National Cheng-Kung University, Tainan, Taiwan. 8 Department of Pathology, College of Medicine, Taipei Medical University, Taipei, Taiwan. 9 Department of Pathology, Taipei Medical University Hospital, Taipei Medical University, Taipei, Taiwan. 10 Department of Internal Medicine, National Cheng- Kung University Hospital, College of Medicine, National Cheng-Kung University, Tainan, Taiwan. 11 Department of Dentistry, Tri-Service General Hospital, National Defense Medical Center, Taipei, Taiwan. 12 Department of Biochemistry, College of Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan. Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). Corresponding Authors: Shine-Gwo Shiah, National Institute of Cancer Research, National Health Research Institutes, Miaoli, Taiwan, Keyan Road, Zhunan, Miaoli County 35053, Taiwan. Phone: 886-3724-6166, ext. 31707; Fax: 886-3758-6463; E-mail: [email protected]; Michael Hsiao, Genomics Research Center, Academia Sinica, 128 Academia Road, Section 2, Nankang, Taipei 115, Taiwan. Phone: 886-2278-71243; Fax: 886-2278-99931; E-mail: [email protected]; and Yi-Shing Shieh, Department of Dentistry, Tri- Service General Hospital, National Defense Medical Center, Taipei 161, Taiwan. Phone: 886-2879-23100, ext. 18138; Fax: 886-2879-2-2911; E-mail: [email protected] doi: 10.1158/0008-5472.CAN-16-1188 Ó2016 American Association for Cancer Research. Cancer Research www.aacrjournals.org OF1 Research. on February 14, 2018. © 2016 American Association for Cancer cancerres.aacrjournals.org Downloaded from Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular and Cellular Pathobiology
Dysregulation of RUNX2/Activin-A Axis uponmiR-376c Downregulation Promotes LymphNodeMetastasis in Head and Neck Squamous CellCarcinomaWei-Min Chang1,2,3, Yuan-Feng Lin4, Chia-Yi Su3, Hsuan-Yu Peng2, Yu-Chan Chang3,Tsung-Ching Lai3, Guan-Hsun Wu2, Yuan-Ming Hsu2, Li-Hsing Chi3,5,6, Jenn-Ren Hsiao7,Chi-Long Chen8,9, Jang-Yang Chang2,10, Yi-Shing Shieh1,11, Michael Hsiao3,12, andShine-Gwo Shiah2,11
Abstract
Epigenetic correlates of the head and neck cancer may illumi-nate its pathogenic roots. Through a gene set enrichment analysis,we found that the oncogenic transcription factor RUNX2 is widelyupregulated in the head and neck squamous cell carcinoma(HNSCC) with lymph node metastasis, where it also predictspoor prognosis in patients with HNSCC. Enforced expression ofectopic RUNX2 promoted the metastatic capabilities of HNSCC,whereas RUNX2 silencing inhibited these features. Mechanisticinvestigations showed that manipulating levels of activin A(INHBA) could rescue or compromise the RUNX2-mediatedmetastatic capabilities of HNSCC cells. Furthermore, we found
that miR-376c-3p encoded within the 30-untranslated region ofRUNX2 played a pivotal role in regulating RUNX2 expression inhighly metastatic HNSCC cells, where it was downregulatedcommonly. Restoring miR-376c expression in this setting sup-pressed expression of RUNX2/INHBA axis along with metastaticcapability. Clinically, we observed an inverse relationshipbetween miR-376c-3p expression and the RUNX2/INHBA axisin HNSCC specimens. In summary, our results defined a novelpathway inwhich dysregulation of the RUNX2/INHBA axis due tomiR-376c downregulation fosters lymph node metastasis inHNSCC. Cancer Res; 76(24); 1–11. �2016 AACR.
IntroductionHead and neck squamous cell carcinoma (HNSCC) is one of
highly prevalent and lethal cancers in theworld, including Taiwan(1, 2). More than 90% of HNSCCs are oral squamous cellcarcinomas, which arise in the oral cavity (3). The initiation andprogression ofHNSCCs are highly correlated with the exposure tocarcinogens, such as cigarette and alcohol. In southeast Asia andTaiwan, betel chewing has been considered as a major risk factorfor HNSCC (3). Although cancer therapies have tremendouslyimproved over the past 20 years, the prognosis of patients withHNSCC remains unfavorable (4, 5). Several recent studies havereported single-molecule biomarkers, such as IGF1R (6), Axl (7),and EpCAM (8) for predicting HNSCC progression; however,their practical usefulness remains to be determined. By usinggenome-wide analysis, this study was thus attempted to identifythe critical molecule(s) as a diagnostic biomarker or even ther-apeutic target for metastatic HNSCC.
RUNX2, a Runt family transcription factor, is a key regulatorphysiologically associated with osteoblast differentiation andchondrocyte maturation (9, 10), as well as cytoskeleton remo-deling and cellular movement via activating its downstreamgenes. Recently, RUNX2 plays as a pivotal molecule in forcingbreast and prostate cancer progression, for example, bonemetastasis, through the transcriptional regulation of MMPs,VEGFs, SNAIL, SLUG, and PTHrP expression (11). Neverthe-less, the functional consequences of RUNX2 upregulation andits downstream transcriptome are still unclear in HNSCCprogression.
1Graduate Institute ofMedical Sciences, National DefenseMedical Center, Taipei,Taiwan. 2National Institute of Cancer Research, National Health ResearchInstitutes, Miaoli, Taiwan. 3Genomics Research Center, Academia Sinica, Taipei,Taiwan. 4Graduate Institute of Clinical Medicine, College of Medicine, TaipeiMedical University, Taipei, Taiwan. 5The PhDProgram for Translational Medicine,Taipei Medical University and Academia Sinica, Taipei, Taiwan. 6Division of Oraland Maxillofacial Surgery, Department of Dentistry, Taipei Medical UniversityHospital, Taipei, Taiwan. 7Department of Otolaryngology, Head and Neck Col-laborativeOncologyGroupandNationalCheng-KungUniversityHospital, Collegeof Medicine, National Cheng-Kung University, Tainan, Taiwan. 8Department ofPathology, College of Medicine, Taipei Medical University, Taipei, Taiwan.9Department of Pathology, Taipei Medical University Hospital, Taipei MedicalUniversity, Taipei, Taiwan. 10Department of Internal Medicine, National Cheng-Kung University Hospital, College of Medicine, National Cheng-Kung University,Tainan, Taiwan. 11Department of Dentistry, Tri-Service General Hospital, NationalDefenseMedical Center, Taipei, Taiwan. 12Department of Biochemistry, College ofMedicine, Kaohsiung Medical University, Kaohsiung, Taiwan.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Authors: Shine-Gwo Shiah, National Institute of CancerResearch, National Health Research Institutes, Miaoli, Taiwan, Keyan Road,Zhunan, Miaoli County 35053, Taiwan. Phone: 886-3724-6166, ext. 31707; Fax:886-3758-6463; E-mail: [email protected]; Michael Hsiao, GenomicsResearch Center, Academia Sinica, 128 Academia Road, Section 2, Nankang,Taipei 115, Taiwan. Phone: 886-2278-71243; Fax: 886-2278-99931; E-mail:[email protected]; and Yi-Shing Shieh, Department of Dentistry, Tri-Service General Hospital, National Defense Medical Center, Taipei 161, Taiwan.Phone: 886-2879-23100, ext. 18138; Fax: 886-2879-2-2911; E-mail:[email protected]
doi: 10.1158/0008-5472.CAN-16-1188
�2016 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org OF1
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

Homodimer INHBA forms a polypeptide hormone, known asactivin A, which is a pluripotent growth and differentiation factor(12). In addition to its physiologic functions, activin A appears todisplay multiple roles in tumorigenesis and cancer progression.Upregulation of activin Ahas been reported in esophageal, gastric,colon, and lung cancer (13–16) and promotes in vitro invasionand epithelial–mesenchymal transition (EMT) in the oral cancer(17). However, the mechanism for INHBA regulation remaineduncertain in HNSCC.
miRNAs are small noncoding RNAs that posttranscriptionallycontrol gene expression. miRNAs use seeding regions to matchmiRNA response elements (MRE) of its target mRNAs and fine-tune target expression throughmRNA degradation or translation-al inhibition (18). In mammals, approximately 30% of protein-coding genes are regulated by miRNAs and a single miRNA canregulate hundreds to thousands of targets (19). AlthoughmiRNAshave fine-tuning roles in cell, it can make a huge change whentargeted to key biological molecules, such as transcription factors,upstream receptors, or hub proteins. Aberrant miRNA expressionwas considered to be a common phenotype in tumorigenesis andcancer progression. More than 50 miRNAs are located in14q32.31 behind the long intervening noncodingRNA (lincRNA)meg3 and are regulated coordinately (20). Here we find that miR-376c-3p, which is located in chromosome 14q32.31, is profound-ly downregulated in HNSCC and thereby leads to the dysregula-tion of RUNX2/activin-A axis. Significantly, this scenario stronglycorrelates with the mechanism for lymph node metastasis andpoor prognosis in HNSCC.
Materials and MethodsHead and neck squamous cell carcinoma cell lines
Cell cultures were prepared and maintained according to astandard protocol. HOKs were obtained from ScienCell andpassaged according to the manufacturer's instructions. 293T andCal-27 cells were obtained fromATCC. Ca9-22 and SAS cells wereobtained from JCRBCell Bank in 2014 andmaintained accordingto the manufacturer's instructions. All culture media were sup-plementedwith 10%FBS (Invitrogen), 1,000U/mLpenicillin and100 mg/mL streptomycin, and 2 mmol/L GlutaMAX at 37�C in ahumidified atmosphere of 95% air and 5% CO2. All cells wereroutinely authenticated on the basis of morphologic and growthcharacteristics as well as by short tandem repeat (STR) analysisand confirmed to be free of mycoplasma.
Patient samplesPaired RNAs from tumor specimens and adjacent noncancer-
ous epithelia were obtained from surgeries performed between1999 and 2010 at the National Cheng Kung University Hospital(Tainan, Taiwan). Fresh/frozen tissues were preserved in liquidnitrogen until use. The study protocol was reviewed and approvedby the Institutional Human Experiment and Ethics Committee ofNational Health Research Institutes (HR-97-100). HNSCC tissuemicroarrays were collected from Taipei Medical University Hos-pital (Taipei, Taiwan) with Institutional Review Board (IRB)approval (TMU-IRB 99049), and then fixed in formalin andembedded in paraffin before archiving. Archived specimens werespotted onto tissue microarrays on Dako-coated slides beforebeing used for immunohistochemical staining or in situ hybrid-ization. The histologic diagnosis of the type of head and neckcancer was performed according to the recommendations of the
WHO classification. Tumor size, local invasion, lymph nodeinvolvement, distal metastasis, and the final disease stage weredetermined according to the definition of the American JointCommittee on Cancer TNM staging system of HNSCC (21).Follow-up was done for up to 100 months.
HNSCC patient microarray analysisHead and neck cancer cDNA microarrays were performed
according to our previous studies (20) and analyzed with thecore transcription factor of gene set enrichment analysis (GSEA)and upstream regulator of Ingenuity Pathway Analysis (IPA).Briefly, the HNSCC patient cDNA microarrays were divided intotwo groups, adjacent normal tissues or HNSCC tumors, andanalyzed by C3 TFT module of GSEA; then, the core oncogenictranscription factors were ranked by normalized enrichment score(NES). The upstream regulators of IPA were analyzed from thesignificant differently expressed genes (>2-fold) between adjacentnormal tissues and HNSCC tumors, and then analyzed by theupstream regulator of IPA. The prognostic value of single gene orpredefined gene set among The Cancer Genome Atlas (TCGA)HNSCC cohort was analyzed through SurvExpress database (23).
Lentiviral viral gene delivery or knockdown and RUNX2 andmicroarray assay
The miR-376c-3p expressing, shRUNX2, shINHBA, and anontargeting silencing miRNA or shRNA control vector werepurchased from OpenBiosystems and Gateway donor cDNAs,such as RUNX2, INHBA, and PTHLH, were purchased fromDNasu and then recombinated into plenti6.3-DEST (Invitrogen)vector by Clonase LR (Invitrogen). All lentiviral vectors weretransfected into the packaging cell line 293T along with thepMD.G and pCMVDR8.91 plasmids using a calcium phosphatetransfection kit (Invitrogen). After 48-hour incubation, the viralsupernatants were transferred to the target cells, and the infectedcells were cultured in the presence of different concentrations ofpuromycin or blasticidin (Calbiochem), depending on the cellline and vector backbone. The RUNX2 regulation targets inHNSCC were performed by Affymetrix U133 microarray assaysand uploaded onto GEO database (GSE74137).
Real-time PCR and chromatin immunoprecipitationquantitative PCR
All of the gene detection assays were conducted as described inour previous publication (20). In the pairwise miR-376c-3p andRUNX2 expression assay, patients' RNAs were reverse-transcribedusing the miScript II RT-Kit (Qiagen). Chromatin immunopre-cipitation (ChIP) was performed using both the EZ-Zyme chro-matin prep kit (Millipore) and ab500 ChIP kit (Abcam) with10 mg primary RUNX2 antibodies (sc-10758X, Santa Cruz Bio-technology) and equal level of rabbit IgG controls (Millipore).The specific gene and amplicon expression was detected withOmicsGreen. The primer sequences and antibodies are shown inSupplementary Table S1.
In vitro migration and invasion assayThe migration and invasion ability of HNSCC cells was deter-
mined in a Boyden Chamber Invasion assay (GEHealthcare). Thepolyvinylidene difluoride (PVDF)membrane pore size was 8 mm.For the invasion assay, PVDF was coated with 1 mg/mL MatrigelBasement Membrane Matrix (BD Biosciences) on the upper side,
Chang et al.
Cancer Res; 76(24) December 15, 2016 Cancer ResearchOF2
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

and 10mg/mLfibronectinwas used to coat the opposite site. In themigration assay, where only fibronectin coating was used, 1.5 �104 cells were seeded into the uppermedium. After incubation for16hours at 37�C, the invaded cells were stainedwithGiemsa stain(Sigma-Aldrich). At least five phase-contrast images were finallyobtained through microscopy, and the invasive cells were quan-tified. For each treatment, the cells were seeded in at least trip-licate, and the experiments were repeated at least twice.
Ago2 immunoprecipitation assayAgo2 immunoprecipitation (IP) was performed following the
protocol described in ref. 20.
Vector construction and reporter assayThe RUNX2-30UTR-reporter was cloned into the pmiRGLO
vector (Promega) at the SacI and XhoI sites, and the 1.5-kbINHBA promoter was cloned into the pGL4.22 vector at theXhoI and HindIII sites. The miRNA-binding site mutationreporter pmiRGLO-RUNX2-mt-376c, which carries the seedingsequence mutation of miRNA-binding sites and harbors adeletion in the RUNX2-binding site, was produced throughsite-directed mutagenesis, and its sequence was subsequentlyconfirmed.
In vivo experiments of HNSCCAll animal experiments were performed in strict accordance
with the recommendations in the guidelines for the Care and Useof Laboratory Animals of the National Health Research Institutes(Miaoli, Taiwan). The protocol was approved by the InstitutionalAnimal Care and Use Committee of the Genomic ResearchCenter, Academia Sinica (Taipei, Taiwan; protocol no.: AS-IACUC-15-06-833). Male NOD-SCID mice aged 5–6 weeks werebred in theGenomicResearchCenter. The animalswere housed ina climate-controlled room (12:12 dark–light cycle, with constanthumidity and temperature) with food and water provided adlibitum. All efforts were made to minimize suffering.
For the lymph node metastasis assay, 1.5 � 103 HNSCC GL-subline cells were injected intomice through an intrabuccal route.In vivo tumor images were captured with an IVIS Imaging System(Caliper Life Sciences) to measure the signal intensity from theGFPþ luciferase vector.
Chromogenic in situ hybridization assayIn situ hybridization was performed with the Ishyb In Situ
Hybridization Kit (Biochain) following the manufacturer'sprotocol.
Statistical analysisA paired t test was performed to compare the RUNX2 and
INHBA IHC expression levels andmiR-376c-3p or RUNX2mRNAlevel in cancer tissues, and in the corresponding normal adjacenttissues. The association between clinicopathologic categoricalvariables and the IHC expression levels were analyzed by Pearsonc2 test. Estimates of the survival rates were calculated using theKaplan–Meier method and compared using the log-rank test.Patient follow-up time was censored until the patient was lostduring follow-up. For all experiments, bar graphs represent themean (� SEM) from three independent experiments and statis-tical analyses were performed using SPSS (Statistical Package forthe Social Sciences; SPSS) 17.0 software. Unless otherwise stated,statistical differences between means were determined using a
Student t test. A P value of <0.05 was considered significant for allof our analyses.
ResultsGSEA identifies RUNX2 as a key transcription factor inpromoting HNSCC progression
To investigate the key transcription factor in HNSCC progres-sion, we used the c3 transcription factor targets (TFT) module ofGSEA (22) to dissect the 40 pairwise HNSCC cDNA microarrays(GSE37991; ref. 23). We identified 10 key transcription factorsthat are highly upregulated in patients with HNSCC (Fig. 1A andB). We further analyzed the prognostic significance of the wholepredefined gene set of those 10 transcription factors from Molec-ular Signatures Database in Taiwanese cohort and TCGA HNSCCcohort. SPI1, BACH2, TFCP2, NFE2, JUN, and BACH2 had higherHRs and significant P values in both cohorts (Table 1). We furtherused the IPA to analyze upstream regulators in TaiwaneseHNSCCpatients, and we found that RUNX2 was the most activatedupstream regulator in patients with HNSCC (SupplementaryTable S2). RUNX2 was also the only transcription factor that waspresented in both analyses. Thus, the activation of RUNX2 mightbe important for HNSCC progression. Immunocytochemistry(ICC) revealed that RUNX2massively accumulates in the nucleusprobably implying a constitutively activated status of RUNX2 inHNSCC cells (Supplementary Fig. S1). Remarkably, our resultsshowed that most of HNSCC cells express RUNX2 much higherthan primary oral keratinocytes (HOKs; Fig. 1C and D). Kaplan–Meier analysis revealed that RUNX2 protein serves as a poorprognostic marker in an independent validation cohort (Fig.1E). Clinical pathologic results also elucidated that RUNX2 sig-nificantly correlates with perineural invasion (P ¼ 0.001) andmarginally associates with N status (N1–3 vs. N0, P ¼ 0.06;Supplementary Table S3). Accordingly, RUNX2 mRNA level alsoappeared to be a poor prognostic marker in TCGAHNSCC cohortdeposited in the SurvExpress database (24) (SupplementaryFig. S2). These results demonstrated that RUNX2 is likely hyper-activated/upregulated and acts as a poor prognostic marker inHNSCC.
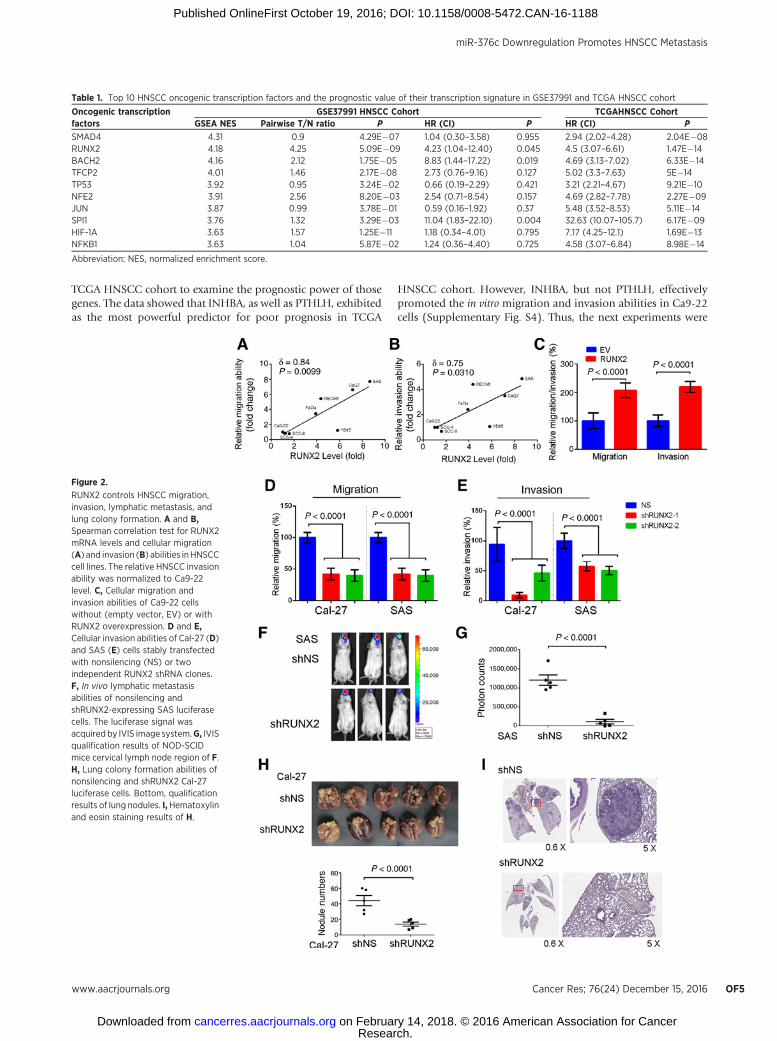
RUNX2 expression causally correlates with the metastaticpotential of HNSCC cells
To understand the correlation between RUNX2 expression andmetastatic potential in HNSCC cells, we determined the endog-enous RUNX2 levels and cellular migration/invasion abilities in apanel ofHNSCC cells. The data showed that RUNX2 level causallyassociates with in vitromigration and invasion abilities of HNSCCcells (Fig. 2A and B) and lung colony formation ability (Supple-mentary Fig. S3; Supplementary Table S4). Next, we enforcedlyexpressed the exogenous RUNX2 gene in Ca9-22 cells with lessendogenous RUNX2 to validate its effects on promoting HNSCCmetastatic progression. Our results revealed that RUNX2 effec-tively promotes cellular migration/invasion abilities in Ca9-22cells (Fig. 2C). In contrast, we performed RUNX2 knockdown inCal-27 andSAS cells that highly express RUNX2. SilencingRUNX2significantly inhibited in vitro cellularmigration/invasion abilitiesin bothCal-27 and SAS cells (Fig. 2D and E) and suppressed the invivo lymph node metastasis judged by the reduced luminescentsignals in tumor-bearing mice (Fig. 2F and G). Moreover, RUNX2knockdown also decreased the lung colonization ability ofHNSCC cells in mice (Fig. 2H and I). These results demonstrated
miR-376c Downregulation Promotes HNSCC Metastasis
www.aacrjournals.org Cancer Res; 76(24) December 15, 2016 OF3
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

that RUNX2 overexpression fosters the metastatic potential ofHNSCC cells.
RUNX2 regulates INHBA expression and RUNX2–INHBA axispromotes HNSCC metastasis
To further investigate the role of RUNX2 transcription targets inHNSCC metastasis progression. We used three differentapproaches to identify specific RUNX2 targets in HNSCC (Fig.3A). First, to analyze the transcriptomeofRUNX2,weperformed a
microarray analysis against RUNX2-overexpressing Ca9-22 cellsand then found 538 genes with 2-fold changes after RUNX2overexpression. In addition, we employed the RUNX2 ChIPsequence data obtained from ChEA, in which 3,423 genes werevalidated tobe regulatedbyRUNX2 (25).Wealso identified2,884genes that display 2-fold upregulation with a high concordance(>50%) in HNSCC tumors. Combining these approaches, weidentified 26 RUNX2 downstream genes that are obviously upre-gulated in HNSCC (Supplementary Table S5). Then, we used the
Figure 1.
RUNX2 upregulation is predominant and serves as a poor prognostic marker in HNSCC. A, The flowchart of identifying the possibly activated transcriptionfactor in HNSCC by GSEA analysis. B, The top ten candidates for possibly activated transcription factors from GSEA analysis. C and D, Expressions of RUNX2 wereassessed by qRT-PCR (C) and immunoblotting (D) in the indicated cell lines. E, Top, immunohistochemical images of RUNX2 staining in HNSCC tissuemicroarray; bottom, Kaplan–Meier survival plot for RUNX2 expression in HNSCC patients.
Chang et al.
Cancer Res; 76(24) December 15, 2016 Cancer ResearchOF4
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188
TCGA HNSCC cohort to examine the prognostic power of thosegenes. The data showed that INHBA, as well as PTHLH, exhibitedas the most powerful predictor for poor prognosis in TCGA
HNSCC cohort. However, INHBA, but not PTHLH, effectivelypromoted the in vitro migration and invasion abilities in Ca9-22cells (Supplementary Fig. S4). Thus, the next experiments were
Table 1. Top 10 HNSCC oncogenic transcription factors and the prognostic value of their transcription signature in GSE37991 and TCGA HNSCC cohort
Oncogenic transcriptionfactors
GSE37991 HNSCC Cohort TCGAHNSCC CohortGSEA NES Pairwise T/N ratio P HR (CI) P HR (CI) P
SMAD4 4.31 0.9 4.29E�07 1.04 (0.30–3.58) 0.955 2.94 (2.02–4.28) 2.04E�08RUNX2 4.18 4.25 5.09E�09 4.23 (1.04–12.40) 0.045 4.5 (3.07–6.61) 1.47E�14BACH2 4.16 2.12 1.75E�05 8.83 (1.44–17.22) 0.019 4.69 (3.13–7.02) 6.33E�14TFCP2 4.01 1.46 2.17E�08 2.73 (0.76–9.16) 0.127 5.02 (3.3–7.63) 5E�14TP53 3.92 0.95 3.24E�02 0.66 (0.19–2.29) 0.421 3.21 (2.21–4.67) 9.21E�10NFE2 3.91 2.56 8.20E�03 2.54 (0.71–8.54) 0.157 4.69 (2.82–7.78) 2.27E�09JUN 3.87 0.99 3.78E�01 0.59 (0.16–1.92) 0.37 5.48 (3.52–8.53) 5.11E�14SPI1 3.76 1.32 3.29E�03 11.04 (1.83–22.10) 0.004 32.63 (10.07–105.7) 6.17E�09HIF-1A 3.63 1.57 1.25E�11 1.18 (0.34–4.01) 0.795 7.17 (4.25–12.1) 1.69E�13NFKB1 3.63 1.04 5.87E�02 1.24 (0.36–4.40) 0.725 4.58 (3.07–6.84) 8.98E�14
Abbreviation: NES, normalized enrichment score.
Figure 2.
RUNX2 controls HNSCC migration,invasion, lymphatic metastasis, andlung colony formation. A and B,Spearman correlation test for RUNX2mRNA levels and cellular migration(A) and invasion (B) abilities inHNSCCcell lines. The relative HNSCC invasionability was normalized to Ca9-22level. C, Cellular migration andinvasion abilities of Ca9-22 cellswithout (empty vector, EV) or withRUNX2 overexpression. D and E,Cellular invasion abilities of Cal-27 (D)and SAS (E) cells stably transfectedwith nonsilencing (NS) or twoindependent RUNX2 shRNA clones.F, In vivo lymphatic metastasisabilities of nonsilencing andshRUNX2-expressing SAS luciferasecells. The luciferase signal wasacquired by IVIS image system.G, IVISqualification results of NOD-SCIDmice cervical lymph node region of F.H, Lung colony formation abilities ofnonsilencing and shRUNX2 Cal-27luciferase cells. Bottom, qualificationresults of lung nodules. I,Hematoxylinand eosin staining results of H.
miR-376c Downregulation Promotes HNSCC Metastasis
www.aacrjournals.org Cancer Res; 76(24) December 15, 2016 OF5
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

Figure 3.
RUNX2 regulates INHBA to promote HNSCCmigration, invasion, andmetastasis.A, The flowchart of identifying the RUNX2-regulated target genes in HNSCC.B andC,RUNX2 and INHBA protein (B) and mRNA (C) expression after the enforced expression of ectopic RUNX2 in Ca9-22 and SCC-9 cells. D, INHBA promoterassay of thewild-type INHBApromoterwithout orwith a deletion of the consensusRUNX2-binding sequence.E,TheChIP after theectopicRUNX2expression inCa9-22cells. F and G, In vitro migration and invasion assay for the RUNX2/INHBA axis. The restoration and knockdown of INHBA was performed in RUNX2-silencingSAS cells (F) and RUNX2-overexpressing Ca9-22 cells (G). H, The cellular migration and invasion abilities in Cal-27 and SAS cells with high endogenousRUNX2 levels after INHBA knockdown. I, The cellular migration and invasion abilities in Ca9-22 cells after the treatment with recombinant activin A (100 ng/mL).J, The lymphatic metastatic abilities of SAS cells after INHBA knockdown in NOD-SCID mice. K, IVIS qualification results for cervical lymph node region ofNOD-SCID mice (n ¼ 5) in J. In D–G, data from three independent experiments are presented as mean � SEM. The statistical significance was analyzed byStudent t test.
Chang et al.
Cancer Res; 76(24) December 15, 2016 Cancer ResearchOF6
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

designed to examine the metastasis-promoting effect ofRUNX2/INHBA axis in HNSCC cells. Enforced ectopic expres-sion of RUNX2 gene was found to promote INHBA expressionin both Ca9-22 and SCC-9 cells (Fig. 3B and C) and alsoregulated INHBA promoter activity assay in Ca9-22 cells (Fig.3D). Chromatin immunoprecipitation quantitative PCR (ChIP-qPCR) experiment was performed and the results showed thedirect binding of RUNX2 to INHBA promoter (Fig. 3E). Thedeletion of RUNX2-binding site in the promoter region ofINHBA suppressed the INHBA promoter activity, which indi-cates a direct regulation of RUNX2 on the transcriptionalactivity of INHBA gene at the RNA level (Fig. 3D). RestoringINHBA expression in RUNX2-silencing SAS cells rescued thecellular migration and invasion abilities (Fig. 3F). Conversely,INHBA knockdown in RUNX2-overexpressing Ca9-22 cellscompromised the RUNX2-promoted cellular migration andinvasion abilities (Fig. 3G). In Ca9-22 cells, the activin/inhibinfamily genes, only INHBA, not INHA or INHBB, were stimu-lated by RUNX2 overexpression. Thus, we used the homodimerINHBA, activin-A recombinant protein, which is able to stim-ulate Ca9-22 migration and invasion (Fig. 3I). Moreover, inCal-27 and SAS cells that have high endogenous RUNX2expression, INHBA knockdown prevented the in vitromigrationand invasion (Fig. 3H), as well as in vivo cervical lymph nodemetastasis of SAS cells (Fig. 3J and K). Furthermore, we found apositive correlation of RUNX2 with INHBA in the 2016 HNSCCTCGA dataset that high-level of RUNX2 and INHBA expressionstrongly predicted a worst clinical outcome (SupplementaryFig. S5) and might correlate to HPV-negative head and neckcancer (Supplementary Table S6). These findings demonstrated
that RUNX2/INHBA axis potentially dominates the metastaticprogression in HNSCC.
miR-376c-3p controls RUNX2/activin-A axis in HNSCCTo ascertain the possiblemechanism for RUNX2dysregulation,
we surveyed the configuration of miRNA-binding sites within the30-untranslated region (30UTR). We found that miR-376c-3p issignificantly silenced in clinical HNSCC samples (SupplementaryTable S7; refs. 20, 26) and alignedwithin the 30UTRof theRUNX2gene (Fig. 4A). Dramatically, the miR-376c-3p expression wasshown tonegatively correlatewith themalignancy ofHNSCCcells(Fig. 4B). Moreover, the mutation of miR-376c-3p–binding sitepredominantly restored the transcriptional activity of RUNX2 inSAS cells (Fig. 4C; Supplementary Fig. S6). Other HNSCC tran-scription factors, such as SMAD4, BACH2, and HIF-1A, also havethe miR-376c–binding sequences in its 30-UTR regions (Supple-mentary Table S8). The ago2-dependent immunoprecipitationresults also revealed that the Ago2–RUNX2–miR-376c-3p com-plex was enriched in miR-376c–expressing cells compared withscramble miRNA–expressing cells (61.15 � 1.97-fold, P < 0.001)and HIF-1A was also significant enriched (2.08 � 0.05-fold, P <0.01) compared with SMAD4 and BACH2. Moreover, miR-376c-3p expression showed an inverse correlation with the RUNX2mRNA levels in HNSCC patients (Fig. 4E). These findings dem-onstrated that miR-376c-3p directly regulates RUNX2 expressionin HNSCC cells.
Next, we enforcedly expressed an ectopic miR-376c and thenfound a dramatic decrease in RUNX2 (Fig. 5A) and INHBA (Fig.5B) expression, as well as the cellular migration/invasion abilities(Fig. 5C and D) in Cal-27 and SAS cells. Notably, miR-376c
Figure 4.
RUNX2 is a direct target of miR-376c-3p in HNSCC. A, Presentation of thesequence of wild-type RUNX2 30-UTR,miR-376c-3p and generated mutationin RUNX2 30-UTR reporter plasmid.B, miR-376c-3p expression level inHNSCC cells. C, The reporter assay forwild-type andmutant RUNX2 30UTR inmiR-376c–overexpressing SAS cells.D, Ago2-dependentimmunoprecipitation assay in miR-376c-3p–overexpressing cells andcontrol cells. E, miR-376c-3p andRUNX2 levels in in 46 paired nontumorand tumor tissues from HNSCCpatients. cDNAs in an independentvalidation cohort were analyzed byOmicsGreen-based qRT-PCR analysis.The expression levels of RUNX2 andmiR-376c-3p were normalized toendogenous GAPDH. In B and C, datafrom three independent experimentsare presented as mean � SEM. Thestatistical significance was analyzedby Student t test. N.S., nonsignificant.
miR-376c Downregulation Promotes HNSCC Metastasis
www.aacrjournals.org Cancer Res; 76(24) December 15, 2016 OF7
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

overexpression prevented the lymph node metastasis of SAS cellsin an orthotopic animal model (Fig. 5E and F). In contrast, aftertreating Ca9-22 cells with miR-376c antisense oligonucleotides,we found significantly enhanced expression levels of RUNX2 andHIF-1A proteins at 24 hours and the levels sustained till 72 hours.However, INHBAprotein expressionwas found to be subsequent-ly expressed 48 hours after the anti-miR-376c treatment (Fig. 5G).
The cellular migration/invasion abilities in Ca9-22 cells were alsoaffected correspondingly (Fig. 5H and I). Taken together, our datashowed that enforced expression of 30UTR-free ectopic RUNX2gene strongly restores the miR-376c–suppressed cellular migra-tion/invasion abilities in SAS cells (Fig. 5J and K). Anti-miR-376ctreatment could increase the cellular migration/invasion abilitiesin Ca9-22 cells and then subsequent knockdown of RUNX2 in
Figure 5.
miR-376c-3p regulates RUNX2/INHBA axis and serves as metastasissuppressor in HNSCC. A and B,RUNX2 (A) and INHBA (B)expression in Cal-27 and SAS cellsafter the enforced expression ofectopic miR-376c. NS, nonsilencing.C and D, The cellular migration andinvasion abilities in Cal-27 and SAScells after the enforced expression ofectopic miR-376c. E, The lymphaticmetastatic abilities of SAS cells inNOD-SCID mice after the enforcedexpression of ectopic miR-376c. F,IVIS qualification results for cervicallymph node region of NOD-SCIDmice (n¼ 5) in E. Data are presentedas mean � SEM. The statisticalsignificancewas analyzed by Studentt test. G, RUNX2 and INHBAexpression in Ca9-22 cells after thetreatment with anti-miR-376c-3poligonucleotide. H and I, Cellularmigration and invasion abilities inCa9-22 cells after the treatment withanti-miR-376c-3p oligonucleotide.J and K, The cellular migration andinvasion abilities in miR-376c–overexpressing SAS cells without orwith the transfection of 30UTR-freeRUNX2. In C, D, H–K, data from threeindependent experiments arepresented as mean � SEM. Thestatistical significance was analyzedby Student t test.
Chang et al.
Cancer Res; 76(24) December 15, 2016 Cancer ResearchOF8
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

those cells could deplete the migration/invasion abilities fromanti-miR-376c treatments (Supplementary Fig. S7). These resultsindicated that RUNX2 is an important downstream target of miR-376c–induced HNSCC progression.
Significantly, IHC results revealed that INHBA expressioninversely correlateswithmiR-376c-3p levels (Fig. 6A) andpredictspoor prognosis in HNSCC (Fig. 6B). Therefore, miR-376c down-regulation likely leads to the dysregulation of RUNX2/INHBA axisand eventually promotes HNSCC metastatic progression.
DiscussionHere we demonstrate that the loss of miR-376c-3p in HNSCC
promotes lymph node metastasis because of the dysregulation ofRUNX2/INHBA axis. Although the posttranscriptional regulationof miR-376c toward RUNX2 in malignancies other than HNSCCremains to be verified, RUNX2 has been shown to be the keytranscription factor in controlling the cancer metastasis in breastand prostate cancer (11, 27). Other well-known RUNX2 down-stream genes, such as MMP-9, TGFB1, and Snail, appeared toenhance cellular metastatic ability; however, these genes were notmarkedly stimulated by RUNX2 in Ca9-22 cells. This indicatedthat the RUNX2may have specific transcription target in HNSCC.
On the other hand, we determined that INHBA acts as a masterdownstreamgeneof RUNX2 inpromoting cancermetastasis, eventhough PTHLH also displayed as a stronger poor prognosticmarker in HNSCC. RUNX2 is a multipotent transcription factorthat regulates hundreds of genes (25) and also has direct protein–protein interaction with other HNSCC oncogenic transcriptionfactors, such as JUN (28), HIF-1A (29), and TP53 (30) in othercancer cell types. In HNSCC, RUNX2 may also have a cooperatedrole or cross-talk with other transcription factors to controloncogene functions and cancer progression (SupplementaryTable S9). Our study provides a prometastatic function of RUNX2that may contribute to HNSCC cervical metastasis but also iden-tifies its novel transcriptional targets INHBA in HNSCC.
INHBA has been found to interplay both oncogenic and tumorsuppressor roles in cancer (31). INHBAwas appeared to associatewith a mechanism for promoting cancer initiation, cachexia, anddrug resistance (32–34). Upregulation of INHBA also promotesEMT process (35, 36) via suppressing E-cadherinmRNA level, butstimulating mesenchymal molecule expression, such as N-cad-herin and Vimentin mRNA expression in HNSCC cells (17). Herewe further show that INHBA acts as a key effector in HNSCClymphaticmetastasis. A similar aspect that INHBA is referred to asa predictor for lymphnodemetastasis and poor overall survival in
Figure 6.
The expression of miR-376c-3p andINHBA is negatively correlated andmiR-376c-3p/activin A are poorprognostic markers in HNSCCpatients. A, Chromogenic in situhybridization assay analysis for theexpression of miR-376c-3p and IHCstaining for INHBA protein in HNSCCtumors. Pearson correlation test wasused to evaluate the statisticalsignificance of correlation betweenmiR-376c and INHBA expression. B,The Kaplan–Meier survival plot forINHBA in HNSCC patients. C,Hypothetical model of the miR-376c-3p-RUNX2/INHBAaxis thatmayplay amajor role in the lymphatic metastasisof HNSCC while HIF-1A may play aminor role.
miR-376c Downregulation Promotes HNSCC Metastasis
www.aacrjournals.org Cancer Res; 76(24) December 15, 2016 OF9
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

HNSCC was reported recently by Kelner and colleagues (37).Although the practical application of activins has been proposedto treat cancers previously, none of activin agents are available incurrent cancer therapy (38). Our findingsmay offer a feasibility ofactivin-A agents for combating the metastatic HNSCC.
Previously, miRNAs have been developed for several types oftranslational applications, such as prognostic, diagnostic, andpredictive biomarkers (39). Recently, a miR-34a–based cancertherapywas entered into a phase I clinical trial for the treatment ofliver cancer (40). In addition to act as a negative regulator ofRUNX2/INHBA axis, miR-376c-3p also controls the cellularexpression of activin receptors, ALK-5, and co-SMADs, Smad4(41, 42). It demonstrates that downregulation of miR-376c-3ppromotes the cancer progression in HNSCC probably due to theloss of regulation in several oncogenic signaling cascades.Although, RUNX2, HIF-1A, SMAD4, and BACH2 have the miR-376c–binding elements on its 30-UTR regions, however, RUNX2was the most predominant transcription factor in HNSCC cells.Ago2–HIF-1A–miR-376c complex was found to have a significantincrease in HNSCC cells (Fig. 4D). HIF-1A has been reported topromote INHBA expression under hypoxia in chondrocytes (43).There is a possibility that HIF1A can transactivate INHBA expres-sion in HNSCC. Indeed, in our experimental results, similar toRUNX2,HIF-1Aproteinwas found to increase, to a lesser extent, at24 hours after anti-miR-376c treatment. These results suggest thatboth RUNX2 and HIF1A transcription factors may be able toincrease the transcriptions of INHBA RNA (Fig. 5G).
Betel nut exposure induces meg-3 promoter hypermethylationin chromosome 14q32.31 and coordinately downregulates theexpression ofmiRNAs includingmiR-376c (20).Herewefind thatthe loss of miR-376c-3p expression increases RUNX2 and itstranscriptional target, INHBA, in HNSCC. The overexpression ofmiR-376c-3p was capable of inhibiting the in vitro cellular migra-tion/invasion abilities and in vivo lymphaticmetastasis inHNSCCcells. Moreover, the downregulation of miR-376c was extensivelyfound in HNSCC tumor compared with the adjacent normaltissues asdeterminedby IHCanalysis andRNA-sequencing resultsfrom the TCGAdatabase (44). Similar viewswere also observed inmetastatic melanoma and prostate cancer cells (45, 46). There-fore, these findings demonstrate the tumor-suppressive functionof miR-376c, particularly in preventing metastatic progression inHNSCC.
Directly targeting transcriptional dysregulation is an emergingapproach bypassing cancer clonal heterogeneity inmodern cancer
therapy (47). On the basis of our findings, we suggest that thereplenishment of miR-376c, therapeutic targeting of RUNX2transcriptional activity or the immunoneutralization of extracel-lular activin A could be a new strategy to combat metastaticevolution in HNSCC.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: W.-M. Chang, J.-Y. Chang, Y.-S. Shieh, M. Hsiao,S.-G. ShiahDevelopment of methodology: W.-M. Chang, C.-Y. Su, T.-C. Lai, J.-R. Hsiao,Y.-S. ShiehAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): W.-M. Chang, C.-Y. Su, Y.-C. Chang, J.-R. Hsiao,C.-L. ChenAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis):W.-M. Chang, Y.-F. Lin, C.-Y. Su, H.-Y. Peng, T.-C. Lai,G.-H. Wu, Y.-M. Hsu, L.-H. Chi, Y.-S. Shieh, M. Hsiao, S.-G. ShiahWriting, review, and/or revision of the manuscript:W.-M. Chang, Y.-S. Shieh,M. Hsiao, S.-G. ShiahAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): C.-L. ChenStudy supervision: Y.-S. Shieh, M. Hsiao, S.-G. Shiah
AcknowledgmentsThe authors would like to acknowledge the great help and assistance of
Experimental Animal Imaging and Molecular Pathology Core Facilities ofGenomic Research Center, Academia Sinica (Taipei, Taiwan).
Grant SupportThis work was supported by National Health Research Institutes (NHRI)
grants from Taiwan (NHRI-CA103-PP-02, NHRI-CA104-PP-03) and the Min-istry of Science and Technology (MOST) grant from Taiwan (MOST-104-2314-B-400-018 to S.-G. Shiah). This study was also supported by Academia Sinicaand Ministry of Science and Technology grants MOST 104-0210-01-09-02 andMOST 105-0210-01-13-01 to M. Hsiao. The head and neck cancer tissue arrayconstruction and relatedworkswere supported byHealth andWelfare surchargeon tobacco products (DOH102-TD-C-111-008) from the Ministry of Healthand Welfare to Comprehensive Cancer Center of Taipei Medical University.
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
ReceivedMay 2, 2016; revised September 11, 2016; acceptedOctober 6, 2016;published OnlineFirst October 19, 2016.
References1. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of
worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer2010;127:2893–917.
2. Su CC, Yang HF, Huang SJ, Lian Ie B. Distinctive features of oral cancer inChanghua County: high incidence, buccal mucosa preponderance, and aclose relation to betel quid chewing habit. J Formos Med Assoc 2007;106:225–33.
3. Warnakulasuriya S.Global epidemiology of oral andoropharyngeal cancer.Oral Oncol 2009;45:309–16.
4. Forastiere A, KochW, Trotti A, Sidransky D. Head and neck cancer. N Engl JMed 2001;345:1890–900.
5. Beenken SW, Krontiras H, Maddox WA, Peters GE, Soong S, Urist MM.T1 and T2 squamous cell carcinoma of the oral tongue: prognosticfactors and the role of elective lymph node dissection. Head Neck1999;21:124–30.
6. Yen YC, Shiah SG, Chu HC, Hsu YM, Hsiao JR, Chang JY, et al. Reciprocalregulation of microRNA-99a and insulin-like growth factor I receptorsignaling in oral squamous cell carcinoma cells. Mol Cancer 2014;13:6.
7. Lee CH, Yen CY, Liu SY, Chen CK, Chiang CF, Shiah SG, et al. Axl is aprognostic marker in oral squamous cell carcinoma. Ann Surg Oncol2012;19:S500–8.
8. Shiah SG, Chang LC, Tai KY, LeeGH,WuCW, Shieh YS. The involvement ofpromoter methylation and DNA methyltransferase-1 in the regulation ofEpCAM expression in oral squamous cell carcinoma. Oral Oncol 2009;45:e1–8.
9. Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, et al.Targeted disruption of Cbfa1 results in a complete lack of bone formationowing to maturational arrest of osteoblasts. Cell 1997;89:755–64.
10. Yoshida CA, Yamamoto H, Fujita T, Furuichi T, Ito K, Inoue K, et al. Runx2and Runx3 are essential for chondrocyte maturation, and Runx2 regulates
Cancer Res; 76(24) December 15, 2016 Cancer ResearchOF10
Chang et al.
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

limb growth through induction of Indian hedgehog. Genes Dev 2004;18:952–63.
11. Baniwal SK, Khalid O, Gabet Y, Shah RR, Purcell DJ, Mav D, et al. Runx2transcriptome of prostate cancer cells: insights into invasiveness and bonemetastasis. Mol Cancer 2010;9:258.
12. Luisi S, Florio P, Reis FM, Petraglia F. Expression and secretion of activin A:possible physiological and clinical implications. Eur J Endocrinol 2001;145:225–36.
13. Yoshinaga K, Mimori K, Yamashita K, Utsunomiya T, Inoue H, Mori M.Clinical significance of the expressionof activinA in esophageal carcinoma.Int J Oncol 2003;22:75–80.
14. Seder CW, Hartojo W, Lin L, Silvers AL, Wang Z, Thomas DG, et al.Upregulated INHBA expression may promote cell proliferation and isassociated with poor survival in lung adenocarcinoma. Neoplasia 2009;11:388–96.
15. WangQ,WenYG, LiDP, Xia J, ZhouCZ, YanDW, et al. Upregulated INHBAexpression is associated with poor survival in gastric cancer. Med Oncol2012;29:77–83.
16. Wildi S, Kleeff J, Maruyama H, Maurer CA, Buchler MW, Korc M. Over-expression of activin A in stage IV colorectal cancer. Gut 2001;49:409–17.
17. Bufalino A, Cervigne NK, de Oliveira CE, Fonseca FP, Rodrigues PC,Macedo CC, et al. Low miR-143/miR-145 cluster levels induce activin Aoverexpression in oral squamous cell carcinomas, which contributes topoor prognosis. PLoS One 2015;10:e0136599.
18. Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNAbiogenesis, function and decay. Nat Rev Genet 2010;11:597–610.
19. Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-tran-scriptional regulation by microRNAs: are the answers in sight? Nat RevGenet 2008;9:102–14.
20. Shiah SG, Hsiao JR, Chang WM, Chen YW, Jin YT, Wong TY, et al. Down-regulated miR329 and miR410 promote the proliferation and invasion oforal squamous cell carcinoma by targeting Wnt-7b. Cancer Res2014;74:7560–72.
21. Greene FL.The American Joint Committee on Cancer: updating the strat-egies in cancer staging. Bull Am Coll Surg 2002;87:13–5.
22. SubramanianA, TamayoP,Mootha VK,Mukherjee S, Ebert BL,GilletteMA,et al. Gene set enrichment analysis: a knowledge-based approach forinterpreting genome-wide expression profiles. Proc Natl Acad Sci U S A2005;102:15545–50.
23. Lee CH, Wong TS, Chan JY, Lu SC, Lin P, Cheng AJ, et al. Epigeneticregulation of the X-linked tumour suppressors BEX1 and LDOC1 in oralsquamous cell carcinoma. J Pathol 2013;230:298–309.
24. Aguirre-Gamboa R, Gomez-Rueda H, Martinez-Ledesma E, Martinez-Tor-teya A, Chacolla-Huaringa R, Rodriguez-Barrientos A, et al. SurvExpress: anonline biomarker validation tool and database for cancer gene expressiondata using survival analysis. PLoS One 2013;8:e74250.
25. Lachmann A, Xu H, Krishnan J, Berger SI, Mazloom AR, Ma'ayan A. ChEA:transcription factor regulation inferred from integrating genome-wideChIP-X experiments. Bioinformatics 2010;26:2438–44.
26. Soga D, Yoshiba S, Shiogama S, Miyazaki H, Kondo S, Shintani S. micro-RNA expression profiles in oral squamous cell carcinoma. Oncol Rep2013;30:579–83.
27. Little GH, Noushmehr H, Baniwal SK, Berman BP, Coetzee GA, Frenkel B.Genome-wide Runx2 occupancy in prostate cancer cells suggests a role inregulating secretion. Nucleic Acids Res 2012;40:3538–47.
28. D'Alonzo RC, Selvamurugan N, Karsenty G, Partridge NC. Physical inter-action of the activator protein-1 factors c-Fos and c-Jun with Cbfa1 forcollagenase-3 promoter activation. J Biol Chem 2002;277:816–22.
29. Lee SH, Che X, Jeong JH, Choi JY, Lee YJ, Lee YH, et al. Runx2 proteinstabilizes hypoxia-inducible factor-1alpha through competition with vonHippel-Lindau protein (pVHL) and stimulates angiogenesis in growthplate hypertrophic chondrocytes. J Biol Chem 2012;287:14760–71.
30. Ozaki T, Wu D, Sugimoto H, Nagase H, Nakagawara A. Runt-relatedtranscription factor 2 (RUNX2) inhibits p53-dependent apoptosis throughthe collaborationwithHDAC6 in response toDNAdamage. CellDeathDis2013;4:e610.
31. Loomans HA, Andl CD. Intertwining of activin A and TGFbeta signaling:dual roles in cancer progression and cancer cell invasion. Cancers2014;7:70–91.
32. Chen JL, Walton KL, Winbanks CE, Murphy KT, Thomson RE, Makanji Y,et al. Elevated expression of activins promotes muscle wasting and cachex-ia. FASEB J 2014;28:1711–23.
33. Zhou X, Wang JL, Lu J, Song Y, Kwak KS, Jiao Q, et al. Reversal of cancercachexia and muscle wasting by ActRIIB antagonism leads to prolongedsurvival. Cell 2010;142:531–43.
34. Lonardo E,HermannPC,MuellerMT,Huber S, Balic A,Miranda-Lorenzo I,et al. Nodal/Activin signaling drives self-renewal and tumorigenicity ofpancreatic cancer stem cells and provides a target for combined drugtherapy. Cell Stem Cell 2011;9:433–46.
35. Wamsley JJ, Kumar M, Allison DF, Clift SH, Holzknecht CM, Szymura SJ,et al. Activin upregulation by NF-kappaB is required to maintain mesen-chymal features of cancer stem-like cells in non-small cell lung cancer.Cancer Res 2015;75:426–35.
36. Park CY, KimDK, Sheen YY. EW-7203, a novel small molecule inhibitor oftransforming growth factor-beta (TGF-beta) type I receptor/activin recep-tor-like kinase-5, blocks TGF-beta1-mediated epithelial-to-mesenchymaltransition in mammary epithelial cells. Cancer Sci 2011;102:1889–96.
37. Kelner N, Rodrigues PC, Bufalino A, Fonseca FP, Santos-Silva AR, MiguelMC, et al. Activin A immunoexpression as predictor of occult lymph nodemetastasis and overall survival in oral tongue squamous cell carcinoma.Head Neck 2015;37:479–86.
38. Fields SZ, Parshad S, Anne M, Raftopoulos H, Alexander MJ, Sherman ML,et al. Activin receptor antagonists for cancer-related anemia and bonedisease. Expert Opin Investig Drugs 2013;22:87–101.
39. Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functionsand therapy. Trends Mol Med 2014;20:460–9.
40. Bouchie A.First microRNA mimic enters clinic. Nat Biotechnol 2013;31:577.
41. Fu G, Ye G, Nadeem L, Ji L, Manchanda T, Wang Y, et al. MicroRNA-376cimpairs transforming growth factor-beta and nodal signaling to promotetrophoblast cell proliferation and invasion. Hypertension 2013;61:864–72.
42. Liu J, Wang L, Su Z, Wu W, Cai X, Li D, et al. A reciprocal antagonismbetween miR-376c and TGF-beta signaling regulates neural differentiationof human pluripotent stem cells. FASEB J 2014;28:4642–56.
43. Lafont JE, Talma S, Hopfgarten C, Murphy CL. Hypoxia promotesthe differentiated human articular chondrocyte phenotype throughSOX9-dependent and -independent pathways. J Biol Chem 2008;283:4778–86.
44. The Cancer Genome Atlas Network. Comprehensive genomic characteri-zation of head and neck squamous cell carcinomas. Nature 2015;517:576–82.
45. Formosa A, Markert EK, Lena AM, Italiano D, Finazzi-Agro E, Levine AJ,et al. MicroRNAs, miR-154, miR-299-5p, miR-376a, miR-376c, miR-377, miR-381, miR-487b, miR-485-3p, miR-495 and miR-654-3p,mapped to the 14q32.31 locus, regulate proliferation, apoptosis, migra-tion and invasion in metastatic prostate cancer cells. Oncogene2014;33:5173–82.
46. Zehavi L, AvrahamR, Barzilai A, Bar-IlanD, Navon R, Sidi Y, et al. Silencingof a large microRNA cluster on human chromosome 14q32 in melanoma:biological effects of mir-376a and mir-376c on insulin growth factor 1receptor. Mol Cancer 2012;11:44.
47. Gonda TJ, Ramsay RG. Directly targeting transcriptional dysregulation incancer. Nat Rev Cancer 2015;15:686–94.
www.aacrjournals.org Cancer Res; 76(24) December 15, 2016 OF11
miR-376c Downregulation Promotes HNSCC Metastasis
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188

Published OnlineFirst October 19, 2016.Cancer Res Wei-Min Chang, Yuan-Feng Lin, Chia-Yi Su, et al. and Neck Squamous Cell CarcinomaDownregulation Promotes Lymph Node Metastasis in Head Dysregulation of RUNX2/Activin-A Axis upon miR-376c
Updated version
10.1158/0008-5472.CAN-16-1188doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2016/10/19/0008-5472.CAN-16-1188.DC1
Access the most recent supplemental material at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/early/2016/12/05/0008-5472.CAN-16-1188To request permission to re-use all or part of this article, use this link
Research. on February 14, 2018. © 2016 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 19, 2016; DOI: 10.1158/0008-5472.CAN-16-1188
Related Documents











