REGULAR ARTICLE Dry matter production in relation to root plastic development, oxygen transport, and water uptake of rice under transient soil moisture stresses Roel Rodriguez Suralta & Yoshiaki Inukai & Akira Yamauchi Received: 15 September 2009 / Accepted: 23 December 2009 # Springer Science+Business Media B.V. 2010 Abstract Drought and waterlogging are important abiotic stresses negatively affecting plant growth and development. They are transiently recurring in rainfed lowlands and in water-saving system prac- ticing intermittent irrigation. This study aimed to determine the contribution of plastic development and associated physiological responses of roots to shoot dry matter production under transient soil moisture stresses. To minimize effect of genetic confounding, a selected line (CSSL47) drawn from 54 chromosome segment substitution lines (CSSL) of Nipponbare (japonica type) carrying an over- lapping chromosome segments of Kasalath (indica type), was used and compared with the recurrent parent Nipponbare. Under transient droughted-to- waterlogged (D–W) conditions, CSSL47 showed greater shoot dry matter production than Nippon- bare. This was due largely to its greater root system development through high induction of aerenchyma formation. Consequently, aerenchyma development effectively facilitated the internal diffusion of oxy- gen (O 2 ) to the root tips under sudden waterlogged condition supporting rapid recovery of stomatal conductance, transpiration, and photosynthesis. Likewise, CSSL47 showed greater shoot dry matter production than Nipponbare under transient waterlogged-to-droughted (W–D) conditions. This was due to CSSL47’ s greater root system develop- ment through more initiation of L type lateral roots that effectively maintained soil water uptake. This in turn sustained higher stomatal conductance, transpi- ration, and photosynthesis. Results implied that utilization of CSSLs could precisely reveal that root plastic development in response to transient soil moisture stresses contributed to the maintenance of shoot dry matter production. Keywords Aerenchyma . Drought . Lateral roots . Oxygen transport . Waterlogging . Water use Abbreviations CSSL chromosome segment substitution line DAS days after sowing D-W droughted-to-waterlogged SMC soil moisture content W–D waterlogged-to-droughted Plant Soil DOI 10.1007/s11104-009-0275-8 Responsible Editor: Len Wade. R. R. Suralta Agronomy, Soils, and Plant Physiology Division, Philippine Rice Research Institute (PhilRice), Science City of Muñoz, Nueva Ecija 3119, Philippines Y. Inukai : A. Yamauchi (*) Graduate School of Bioagricultural Sciences, Nagoya University, Chikusa, Nagoya 464-8601, Japan e-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULAR ARTICLE
Dry matter production in relation to root plasticdevelopment, oxygen transport, and water uptake of riceunder transient soil moisture stresses
Roel Rodriguez Suralta & Yoshiaki Inukai &Akira Yamauchi
Received: 15 September 2009 /Accepted: 23 December 2009# Springer Science+Business Media B.V. 2010
Abstract Drought and waterlogging are importantabiotic stresses negatively affecting plant growthand development. They are transiently recurring inrainfed lowlands and in water-saving system prac-ticing intermittent irrigation. This study aimed todetermine the contribution of plastic developmentand associated physiological responses of roots toshoot dry matter production under transient soilmoisture stresses. To minimize effect of geneticconfounding, a selected line (CSSL47) drawn from54 chromosome segment substitution lines (CSSL)of Nipponbare (japonica type) carrying an over-lapping chromosome segments of Kasalath (indicatype), was used and compared with the recurrentparent Nipponbare. Under transient droughted-to-waterlogged (D–W) conditions, CSSL47 showed
greater shoot dry matter production than Nippon-bare. This was due largely to its greater root systemdevelopment through high induction of aerenchymaformation. Consequently, aerenchyma developmenteffectively facilitated the internal diffusion of oxy-gen (O2) to the root tips under sudden waterloggedcondition supporting rapid recovery of stomatalconductance, transpiration, and photosynthesis.Likewise, CSSL47 showed greater shoot dry matterproduction than Nipponbare under transientwaterlogged-to-droughted (W–D) conditions. Thiswas due to CSSL47’s greater root system develop-ment through more initiation of L type lateral rootsthat effectively maintained soil water uptake. This inturn sustained higher stomatal conductance, transpi-ration, and photosynthesis. Results implied thatutilization of CSSLs could precisely reveal that rootplastic development in response to transient soilmoisture stresses contributed to the maintenance ofshoot dry matter production.
Keywords Aerenchyma . Drought . Lateral roots .
Oxygen transport .Waterlogging .Water use
AbbreviationsCSSL chromosome segment substitution lineDAS days after sowingD-W droughted-to-waterloggedSMC soil moisture contentW–D waterlogged-to-droughted
Plant SoilDOI 10.1007/s11104-009-0275-8
Responsible Editor: Len Wade.
R. R. SuraltaAgronomy, Soils, and Plant Physiology Division,Philippine Rice Research Institute (PhilRice),Science City of Muñoz,Nueva Ecija 3119, Philippines
Y. Inukai :A. Yamauchi (*)Graduate School of Bioagricultural Sciences,Nagoya University,Chikusa,Nagoya 464-8601, Japane-mail: [email protected]

Introduction
Drought and waterlogging are two of the mostimportant abiotic stresses negatively affecting plantgrowth and development. In this aspect, it is impor-tant to understand that these soil moisture stressestransiently recur in the field due to the intermittentnature of rainfall patterns and irrigation systems.Hence, the soil is exposed to frequent episodes andvarying degrees of alternate dry and wet conditionscausing transient drought and O2 deficiency to plants.The inability of plant to acclimate to such changesmay result in reduced growth and dry matterproduction. However, most of the studies in the pastdealt only on the effects of either of the stresses(Wang and Yamauchi 2006). Little attention has beendevoted to their interactive effects.
A series of our previous studies providedevidence that changing soil moisture conditionsadversely affect crop root development mainly byreducing root elongation and branching into lateralroots, and these limit for soil water uptake ability,which then results in reduced stomatal conductance,transpiration and photosynthesis. Ultimately, thisresults in reduced shoot dry matter production(Bañoc et al. 2000a, b; Pardales and Yamauchi2003; Siopongco et al. 2008, 2009; Subere et al.2009; Wade et al. 2000; Yamauchi et al. 1996).Cognizant of this, the ability of roots to respond tothese fluctuating soil moisture regimes was proposedas one of the important developmental traits foradaptation (Ingram et al. 1994; Wang and Yamauchi2006; Yamauchi et al. 1996). Under progressive soildrying, root responses include increased root lengthdensity (Siopongco et al. 2005) as a result of plasticlateral root development (Azhiri-Sigari et al. 2000;Bañoc et al. 2000b; Kamoshita et al. 2000). On theother hand, development of shallow root systemswith enhanced aerenchyma that can provide lessresistant pathway for internal atmospheric O2 diffu-sion to the root tips is a root response to suddenwaterlogging (Colmer 2003; Justin and Armstrong1987; Suralta and Yamauchi 2008). Such ability hasbeen defined as phenotypic plasticity (O’Toole andBland 1987), which plays key roles in plantadaptation to various stress conditions (Wang andYamauchi 2006; Yamauchi et al. 1996).
Under fluctuating soil moistures, we assumedthat rice plants develop their root system adapted to
preceding soil moisture stress (i.e. drought orwaterlogging) prior to encountering the succeedingcontrasting soil moisture stress (i.e. droughted-to-waterlogged or waterlogged-to-droughted transi-tion). Hence, rice plants require different rootsystem development strategies for adaptation. Theextent of the contribution of root plastic develop-ment is difficult to assess under fluctuating soilmoistures, as both the constitutive and adaptive roottrait responses are present and each interacts withboth the existing conditions and the genotypicbackground of the diverse cultivars used (Kamoshitaet al. 2002a). These interactions consequently limitprecise phenotyping for identification of relevantQTLs (Kamoshita et al. 2002b). The constitutiveroot traits are expressed under favorable anaerobicconditions. The adaptive root traits require waterstress for their expressions (e.g., promoted rootelongation rate and plastic lateral root production;Bañoc et al. 2000b, and enhanced aerenchyma undersudden waterlogged conditions; Justin and Armstrong1987).
In our previous study in rice (Suralta et al.2008a), we found that aerobic cultivars have greaterability for plastic lateral root production thanirrigated lowland cultivars under transient moisturestresses. Specifically, greater ability for plasticlateral root production in aerobic than irrigatedlowland cultivars were due to the aerobic cultivars’ability to maintain higher rate of seminal rootelongation and more number of nodal and lateralroots in response to transient O2 deficient-to-droughtconditions, and higher rate of seminal root elonga-tion, more aerenchyma formations in the seminalroot axis, and more number of nodal roots inresponse to transient droughted-to-O2 deficientconditions.
To minimize effect of genetic confounding andincrease precision of the analysis, the above roottrait responses were validated using the chromo-some segment substitution lines (CSSL) of Nippon-bare (japonica) carrying overlapping chromosomesegments of Kasalath (indica) (Suralta et al. 2008b).In that study, we identified and selected one line(CSSL47) with no significant difference in shoot androot growth from Nipponbare under non-stressed(continuously well-aerated) conditions. CSSL47,however, consistently showed more plastic rootdevelopment than Nipponbare in terms of more
Plant Soil

branching of lateral roots and more aerenchymaformation under short-duration of transient moisturestresses in hydroponics such as O2-deficient-to-droughted and vice-versa. Interestingly, root plasticdevelopment responses of CSSL47 did not contrib-ute to greater shoot dry matter production. In thepresent study, therefore, we attempted to test thehypothesis that the plastic responses in root traits areuseful in maintaining crop productivity if plantsare grown under prolonged transient moisturestresses in soils.
The studies presented here examined root plasticdevelopment through aerenchyma formation andlateral root production, and their functions tomaintain atmospheric O2 transport and soil wateruptake under prolonged transient soil moisturestresses. Such responses were also quantified interms of their contribution to the maintenance ofphysiological responses such as stomatal conduc-tance, transpiration, and photosynthesis and ulti-mately, shoot growth in terms of dry matterproduction. To increase precision of the quantitativeanalysis, we used previously selected CSSL47 fromshort-duration transient moisture stresses in hydro-ponics and compared with the recurrent parentNipponbare to minimize the effect of genetic con-founding.
Materials and methods
Plant materials
The two experiments conducted in this studyutilized a selected line CSSL47 drawn from 54chromosome segment substitution lines (CSSL) ofNipponbare (japonica type) carrying an overlappingchromosome segments of Kasalath (indica type) andthe recurrent parent Nipponbare. The CSSL47 havesubstituted segments in 8 of 12 chromosomes inCSSL47 i.e., chr 3 (between loci R1927 and R1925),chr 4 (between R2373 and C734), chr 6 (betweenloci R2147 and C235), chr 7 (between loci C261 andR565), chr 8 (between loci C1107 and R202), chr 10(between loci C701 and R1629 and loci C488 andC223) chr 11 (between loci C447 and C3 and atR1506) and chr 12 (between loci G24B and R617)(http://www.rgrc.dna.affrc.go.jp/ineNKCSSL54.html).
Experiment 1: Transient droughted-to-waterloggedcondition
Seeds of Nipponbare and CSSL47 were soaked inwater and incubated in a seed germinator main-tained at 28°C for 24 h prior to sowing. Three pre-germinated seeds from each genotype were grownin a PVC box (25 cm×2 cm×40 cm, L×W×H)filled with air-dried alluvial loamy sand followingthe method of Kono et al. (1987a). The soil was pre-mixed with fertilizer containing 60 mg nitrogen (N),80 mg phosphorus (P), and 70 mg potassium (K).The plants were grown for 36 days. The seedlingswere later thinned to one seedling per box at 3 daysafter sowing (DAS).
Soil moisture treatments and plant sampling
Two soil moisture treatments were done in thisexperiment: continuously waterlogged (CWL) ascontrol; and transient droughted-to-waterlogged(D-W) conditions. In CWL, water level was increasedand maintained at 2 cm above the soil surface from 3DAS until the end of the experiment (36 DAS). Forthe D–W condition, the soil in the box was firstsubmerged for 24 h and then drained. The soil wasallowed to dry and maintained at 10% soil moisturecontent (SMC; −0.17 MPa) from sowing up to 21DAS. After which, the droughted plants of bothgenotypes were subjected to waterlogged conditionsfor another 15 days. The initial 10% SMC was closeto the critical soil moisture (8% SMC or −0.28 MPa)when most legumes and cereals we have tested startedshowing sign of wilting, especially in the lateafternoon, but recovered the next morning.
All treatments were watered every 2 days to set thedesired SMC. Water was added to replace the waterlost through evapotranspiration and to maintain targetSMC. For lower SMC, the difference in soil moisturebetween the topmost and bottom portions when thesoil was allowed to reach 15% SMC after 7 dayswithout watering was only 1.3%. At 12% SMC after14 days without watering, the difference was 0.3%(Kono et al. 1987b). Since we observed a 2-daywatering interval, differences in moisture between topand bottom soils in the box should have been muchsmaller.
Plant samplings were done 21 DAS (end of initialdrought or waterlogging) and 36 DAS (15 days after
Plant Soil

transient or continuous waterlogging). Three boxes(1 box = 1 replication) from each treatment werecollected from each sampling period.
Physiological measurements
Stomatal conductance, transpiration, and photosyn-thesis of the second-youngest fully-expanded leafwere measured using a portable photosynthesissystem (LI-6400, LI-COR, USA) starting at 1000 h.Measurements were made every 3 days starting at21 DAS (end of initial drought or waterlogging) upto 36 DAS (15 days after transient or continuouswaterlogging).
Root oxygen consumption measurements
Cutting the shoots and sealing the cut portion preventthe entry of atmospheric O2 into the plant’s gas spacesystem (Teal and Kanwisher 1966), allowing aeren-chyma function to be investigated. The difference thenin flux rates between the cut and sealed root systemand intact plant system equates to the amount of O2
transported internally through the plant (Lee 2003;Maricle and Lee 2007; Suralta and Yamauchi 2008).Intact plants with shoots and roots were carefullysampled from the box to prevent damage to rootsystem. The roots were washed to remove soil andother dirt. Root O2 consumptions were determined bysealing the roots of an intact plant into a flask ofnitrogen (N2)-flushed water with initial O2 concentra-tion of 19.0 μmol measured using a dissolved O2 (DO)meter. After 2 h, subsequent decrease in flask O2
concentration was recorded to assess O2 consumptionby the roots. Background rates of O2 flux werelikewise determined in flask with N2-flushed waterwith the same initial O2 concentration as above butwithout suspended plants. A change in O2 concentra-tion in the blank flask was used to correct O2
consumption readings in flask with suspended plants.Shoots were removed by cutting at the stem base
and the cut base was sealed with thick parafilm. O2
consumption from the flask was measured again after2 h to determine the changes in root O2 consumptionswithout shoot. Differences in root O2 consumptionbetween plants with and without shoot indicate thecontribution of aerenchyma to the internal transport ofatmospheric O2 to the roots with O2 deficiency. Theincreases or decreases in respiratory O2 consumption
are expressed as percentages of the total respirationmeasured with the roots excised.
Root porosity measurements
The internal gas space (porosity) may not be identicalwith aerenchyma. For instance, rice roots have rootporosity of ∼9% at 20–25 mm behind the root tipseven with the absence of aerenchyma (Armstrong1971). This means that the estimates of aerenchymabased on porosity may possibly overestimate its size.However, there is no effective method to distinguishthis aerenchyma from total porosity. Thus, we usedtotal porosity as an estimate of aerenchyma develop-ment in this paper.
The coleoptilar nodal root axis was used for rootporosity measurement following the microbalancemethod of Visser and Bögenmann (2003). Coleoptilarnodal roots developed a few days after the emergenceof seminal root axis and, therefore, are assumed to beexposed to both the initial drought and transientwaterlogging. A 2-cm segment from both the basaland apical portions of the roots was cut and excludedfrom the measurements. Briefly, the length of theremaining root axis was divided equally into threeparts and designated as basal (B), middle (M) andapical (A) portions. From each portion, five 1-cmsegments were further sampled. The five 1-cm seg-ments from the basal portion were cut starting fromthe basal proximity. These segments were designatedas B1, B2, B3, B4, and B5.On the other hand, cuttingsfrom the apical portion were done from the apicalproximity and designated as A5, A4, A3, A2, and A1.In the middle portions, five 1-cm segments weresampled using the mid 1-cm segment as reference anddesignated as M3. The two 1-cm segments sampledgoing basally were designated as M2 and M1. Theother 2-cm segments sampled going apically weredesignated as M4 and M5. Each 1-cm root segmentwas cut with a sharp razor blade and gently blotted byrolling it with a small brush on tissue paper for about2 s to remove adherent water. Then, to prevent weightloss by evaporation, the segments were transferredinto a capsule with cover that had been tared on amicrobalance. After closing the capsule, the segmentswere weighed (w1 in µg), transferred to a holder withsmall vials filled with water, and stored for 30 min.Up to 36 samples were weighed before they wereinfiltrated with tap water twice under vacuum for
Plant Soil

30 min each. After water infiltration, the root seg-ments were blotted again on tissue paper for about 2 sand weighed in a capsule (w2 in µg). Using thespecific weight (SW) obtained from larger samples(1.04 g ml−1; Visser and Bögenmann 2003), theporosity was calculated using the formula:
Porosity %; v : vð Þ ¼ 100 � w2� w1ð Þ � SW=w2
with the specific weight of water being 1.00 g ml−1.Furthermore, the number of lateral roots from each
1-cm segment was also counted and expressed aslinear frequency (the number of lateral roots per unitlength root axis; Ito et al. 2006).
Shoot and root development measurements
After O2 consumption and root porosity measure-ments, the main stem leaf number and number ofstems of each plant were counted and then cut, ovendried at 80°C for 48 h, and weighed. Thereafter, theroot system was stored in FAA (formalin: acetic acid:70% ethanol = 1: 1: 18 by volume) solution forfurther measurements. The length of seminal rootswas measured using a ruler and the number of lateralroots along its axis was counted. The number of nodalroots was manually counted. For total root length,each root sample in FAA was rinsed with water andspread on a transparent sheet without overlapping.Digitized images were taken using a scanner with aresolution of 300 dpi and an output format of 256grey scales. The total root length was determinedusing a macro program developed by Kimura et al.(1999) and Kimura and Yamasaki (2001) on the NIHimage software version 1.60 (public domain releasedby the National Institute of Health, U.S.A.).
After scanning, the root samples were oven dried at80°C for 48 h before weighing. The specific rootlength (SRL) was calculated as root length per unitroot dry weight. The variations in SRL can beattributed to the differences in production of lateralroots (Bañoc et al. 2000b), root density (Eissenstat1991), or the combination of both.
Statistical analysis
The treatments were arranged in split-plot design withthree replications. Soil moisture treatment was used asmainplot while the genotypes as subplot.
Experiment 2: Transient waterlogged-to-droughtedcondition
The same set of genotypes used in Experiment 1was used in this experiment. Pre-germinated seedswere sown in rootboxes that were prepared as inExperiment 1.
Soil moisture treatments and plant sampling
Four soil moisture treatments were used in thisexperiment: continuously waterlogged (CWL) ascontrol; and three transient waterlogged-to-droughted(W–D) conditions with varying levels of target SMCs.In CWL, water level was increased and maintained at2 cm above the soil surface from 3 DAS until the end ofthe experiment (38 DAS). In W–D, the soil inside thebox was first waterlogged from 3 to 14DAS. Thereafter,water was drained, allowed to dry and maintained to adesired moisture levels. Water levels were 30% SMC(−0.003 MPa), 25% SMC (−0.014 MPa) and 20% SMC(−0.040 MPa), hereafter referred to as W–D30, W–D25,and W–D20, respectively. Drought treatments imposedthrough progressive soil drying were terminated at 38DAS. These drought stress intensities are well withinthe reported range of soil water potentials (from 0to −0.05 MPa) that significantly reduce rice yields(Bouman and Toung 2001). The decrease in soilmoisture of each box was monitored every 2 days torecord the amount of evapotranspiration. A few boxes(n=3) for both the CWL and transient W–D conditionswere not planted and were used to measure the amountof water lost through evaporation. In this way, theamount of water lost by transpiration alone could beestimated. Watering was done every 2 days orwhenever necessary to maintain the desired SMC.
Samplings were made at 14 DAS (after 14 days ofinitial waterlogging) and at 38 DAS (after 24 days ofcontinuously waterlogged or transient droughtedconditions). Three plants were collected from eachtreatment for each sampling period.
Physiological measurements
Stomatal conductance, transpiration, and photosyn-thesis of the second-youngest fully-expanded leafwere measured using a portable photosynthesissystem (LI-6400, LI-COR, USA) starting at 1000 h.Measurements were done at 14 DAS and 38 DAS.
Plant Soil

Shoot and root development measurements
The main stem leaf number and number of stems ofeach plant were counted, then cut, oven dried at 80°Cfor 48 h before weighing. The whole root system wassampled thereafter, using a pinboard following themethods of Kono et al. (1987a). The extracted rootsystems embedded in plastic sheets were stored inFAA for further measurements.
For total root length, each root sample wasthoroughly rinsed with water to remove FAA andstained in 0.25% Coomassie Brilliant Blue R 250aqueous solution for 48 h. This staining procedurewas indispensable to take high-resolution digitalphotographs of the entire root system including finelateral roots. After staining, the root samples werewashed gently with running water to remove excessstains. Thereafter, each root sample was put in a lightbox and photographed using transmitted light. Digi-tized photographs of the root system profile weretaken using a Canon digital camera at highestresolution (3,456×2,304 pixels). The digital imagesin jpeg format were downloaded in a computer andconverted into tiff format for total root lengthmeasurements. The total root length was measuredusing a macro program on NIH developed by Kimuraet al. (1999) and Kimura and Yamasaki (2001).
The number of nodal roots was manually counted.From each plant, two nodal roots were sampled (oneeach from first and sixth internodes). Each nodal rootwas cut into 4-cm segments keeping the lateral rootsintact. In this way, the number of each type of lateralroot was determined. The lateral roots were classifiedinto L and S types that have been shown to differ inanatomy, morphology, developmental characteristics,carbon and nitrogen dynamics (Yamauchi et al. 1996)and the genetic control of development (Wang et al.2005). The lateral roots that branched into higherorder laterals were regarded as L type and the non-branching roots as S type. Each type of lateral rootwas expressed as linear frequency (the number oflateral roots per unit length of root axis; Ito et al.2006).
Statistical analysis
The treatments were arranged in split-plot design withthree replications. The soil moisture treatments wereused as mainplot while the genotypes as subplot.
Results
Experiment 1: Effects of drought followedby waterlogged (D–W) condition
Shoot and root development
Main stem leaf number as well as shoot and rootgrowth were not significantly different betweenCSSL47 and Nipponbare after initial waterlogging(from 0 to 21 DAS) (Table 1). The removal ofshoots from intact plants significantly increased rootO2 consumption from the hypoxic medium in bothgenotypes (Table 1). The estimated atmospheric O2
contributed by aerenchyma was not significantlydifferent between the two genotypes (Table 1).Likewise, the responses of those measured parame-ters to initial drought were not significantly differentbetween the two genotypes (Table 1). Underdrought, the main stem leaf number was notsignificantly affected in both genotypes (Table 1).Meanwhile, the effects of drought treatment on otherparameters were significant. Specifically, initialdrought significantly reduced the shoot dry weight,number of stems per plant, total root length, numberof nodal roots, and specific root length (SRL), ascompared with their waterlogged controls (Table 1).In contrast, initial drought significantly promoted thelength of seminal roots (Table 1). Furthermore, theremoval of shoots from intact plants of initiallydroughted treatments did not significantly increaseroot O2 consumption from the hypoxic medium inboth genotypes (Table 1). As a result, the atmo-spheric O2 contributed by aerenchyma was signifi-cantly lower in both genotypes, compared with theircontrols (Table 1).
Under CWL condition for 36 days, the mainstem leaf number was not significantly differentbetween CSSL47 and Nipponbare (Table 2). Like-wise, shoot and root development, and other mea-sured parameters were not significantly differentbetween the two genotypes (Table 2). The removalof shoots from an intact plant significantly in-creased O2 consumptions from the hypoxic medium(Table 2). The estimated atmospheric O2 contributedby aerenchyma, however, was not significantlydifferent between the two genotypes (Table 2).Under drought followed by waterlogged (D–W)condition for 15 days, the main stem leaf number
Plant Soil
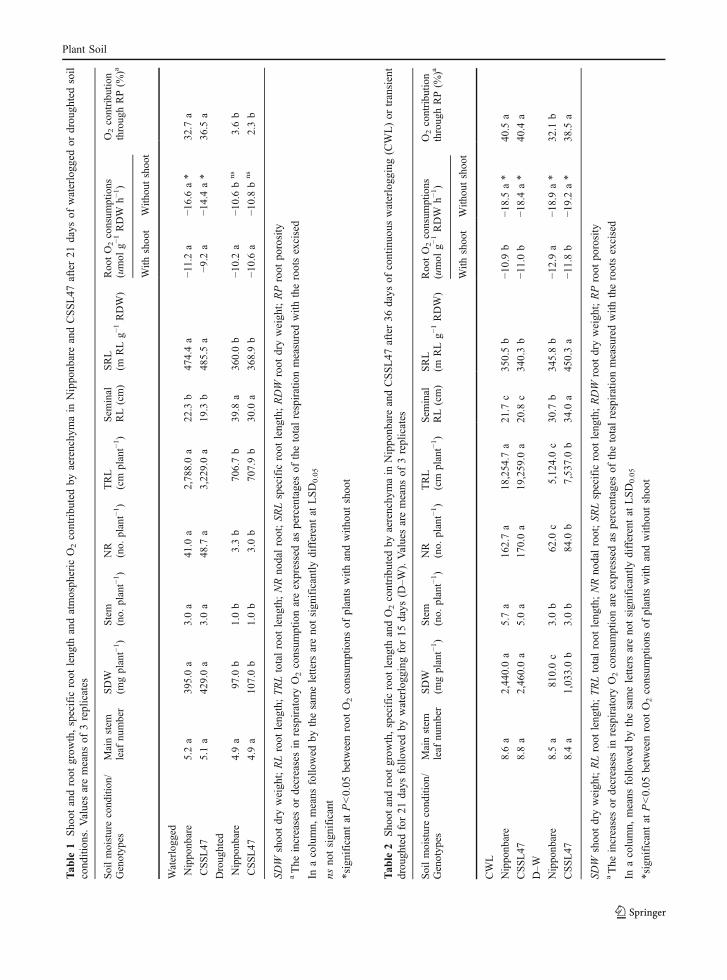
Tab
le1
Sho
otandroot
grow
th,specific
root
leng
thandatmosph
eric
O2contribu
tedby
aerenchy
main
Nippo
nbareandCSSL47
after21
days
ofwaterlogg
edor
drou
ghtedsoil
cond
ition
s.Valuesaremeans
of3replicates
Soilmoisturecondition/
Genotypes
Mainstem
leaf
number
SDW
(mgplant−1)
Stem
(no.
plant−1)
NR
(no.
plant−1)
TRL
(cm
plant−1)
Sem
inal
RL(cm)
SRL
(mRLg−
1RDW)
RootO2consum
ptions
(umol
g−1RDW
h−1)
O2contributio
nthroughRP(%
)a
With
shoot
With
outshoot
Waterlogged
Nipponbare
5.2a
395.0a
3.0a
41.0
a2,788.0a
22.3
b474.4a
−11.2a
−16.6a*
32.7
a
CSSL47
5.1a
429.0a
3.0a
48.7
a3,229.0a
19.3
b485.5a
−9.2
a−1
4.4a*
36.5
a
Droughted
Nipponbare
4.9a
97.0
b1.0b
3.3b
706.7b
39.8
a360.0b
−10.2a
−10.6b
ns
3.6b
CSSL47
4.9a
107.0b
1.0b
3.0b
707.9b
30.0
a368.9b
−10.6a
−10.8b
ns
2.3b
SDW
shoo
tdryweigh
t;RLroot
leng
th;TRLtotalroot
leng
th;NRno
dalroot;SR
Lspecific
root
leng
th;RDW
root
dryweigh
t;RProot
porosity
aThe
increasesor
decreasesin
respiratoryO2consum
ptionareexpressedas
percentagesof
thetotalrespirationmeasuredwith
therootsexcised
Inacolumn,
means
follo
wed
bythesameletters
areno
tsign
ificantly
differentat
LSD0.05
nsno
tsign
ificant
*significant
atP<0.05
betweenroot
O2consum
ptions
ofplantswith
andwith
outshoo
t
Tab
le2
Sho
otandroot
grow
th,specific
root
leng
thandO2contribu
tedby
aerenchy
main
Nippo
nbareandCSSL47
after36
days
ofcontinuo
uswaterlogg
ing(CWL)or
transient
drou
ghtedfor21
days
follo
wed
bywaterlogg
ingfor15
days
(D–W
).Valuesaremeans
of3replicates
Soilmoisturecond
ition
/Genotyp
esMainstem
leaf
number
SDW
(mgplant−1)
Stem
(no.
plant−1)
NR
(no.
plant−1)
TRL
(cm
plant−1)
Sem
inal
RL(cm)
SRL
(mRLg−
1RDW)
Roo
tO2consum
ptions
(umol
g−1RDW
h−1)
O2contribu
tion
throug
hRP(%
)a
With
shoo
tWith
outshoo
t
CWL
Nippo
nbare
8.6a
2,44
0.0a
5.7a
162.7a
18,254
.7a
21.7
c35
0.5b
−10.9b
−18.5a*
40.5
a
CSSL47
8.8a
2,46
0.0a
5.0a
170.0a
19,259
.0a
20.8
c34
0.3b
−11.0b
−18.4a*
40.4
a
D–W
Nippo
nbare
8.5a
810.0c
3.0b
62.0
c5,12
4.0c
30.7
b34
5.8b
−12.9a
−18.9a*
32.1
b
CSSL47
8.4a
1,03
3.0b
3.0b
84.0
b7,53
7.0b
34.0
a45
0.3a
−11.8b
−19.2a*
38.5
a
SDW
shoo
tdryweigh
t;RLroot
leng
th;TRLtotalroot
leng
th;NRno
dalroot;SR
Lspecific
root
leng
th;RDW
root
dryweigh
t;RProot
porosity
aThe
increasesor
decreasesin
respiratoryO2consum
ptionareexpressedas
percentagesof
thetotalrespirationmeasuredwith
therootsexcised
Inacolumn,
means
follo
wed
bythesameletters
areno
tsign
ificantly
differentat
LSD0.05
*significant
atP<0.05
betweenroot
O2consum
ptions
ofplantswith
andwith
outshoo
t
Plant Soil

was not significantly different between the twogenotypes (Table 2). However, W–D significantlyreduced the shoot dry weight, total root length andnumber of nodal roots in both genotypes, but thereduction was significantly less in CSSL47 than inNipponbare (Table 2). The number of stems perplant was also significantly reduced at almostsimilar rates for both genotypes (Table 2). More-over, the length of seminal root was significantlypromoted in both genotypes, but the promotionwas much greater in CSSL47 than in Nipponbare(Table 2).
SRL was not significantly affected by D–Wcondition in Nipponbare while in CSSL47 it wassignificantly increased (Table 2). Furthermore, theremoval of shoots from an intact plant from W–Dtreatments also significantly increased O2 consump-tion from the hypoxic medium in both genotypes(Table 2). However, for Nipponbare, the estimatedatmospheric O2 contributed by aerenchyma under thiscondition was significantly lower than under contin-uous flooding (Table 2).
Stomatal conductance, transpiration,and photosynthesis
At the end of initial drought (21 DAS), stomatalconductance, transpiration, and photosynthesis weresignificantly reduced in both genotypes (Fig. 1).During transient waterlogging period (22 to 36 DAS)under D–W condition, CSSL47 recovered itsstomatal conductance, transpiration, and photosyn-thesis back to the level of its control after 12 daysof waterlogging. In contrast, Nipponbare onlyshowed partial recovery of stomatal conductance,transpiration, and photosynthesis during transientwaterlogging (Fig. 1).
Root porosity in segments along the coleoptilar nodalroot axis
Compared with their CWL controls, mean rootporosity under D–W condition was significantlylower in Nipponbare while it was not affected inCSSL47 (Fig. 2). The lower mean root porosity inNipponbare was particularly attributed to the signif-icantly lower root porosities in segments at the basaland middle portions of the coleoptilar nodal root axis(Fig. 2).
Linear frequency of lateral roots along the coleoptilarnodal root axis
The mean linear frequency of lateral roots on seg-ments along the coleoptilar nodal root axis wassignificantly reduced by D–W condition in Nippon-bare but it was not affected in CSSL47 (Fig. 3). Suchreduction in Nipponbare was attributed to the signif-icant decreases in linear frequency of lateral rootsparticularly on segments along the basal and apicalportions of the coleoptilar nodal root axis (Fig. 3). Incontrast, the linear frequency of lateral roots onsegments all throughout the coleoptilar nodal rootaxis of CSSL47 was not significantly affected bytransient D–W condition.
Experiment 2: Waterlogged followed by droughted(W–D) condition
Main stem leaf number, shoot and root growth,number of stems per plant, stomatal conductance,transpiration, and photosynthesis were not signifi-cantly different between CSSL47 and Nipponbareafter initial waterlogging (14 DAS; Table 3).
Shoot and root growth
The main stem leaf number was not significantlydifferent between the two genotypes at any of the soilmoisture treatments (Table 4). Shoot dry weights underCWL and W–D30 (waterlogging followed by droughtto 30% SMC) conditions were not significantlydifferent between CSSL47 and Nipponbare (Table 4).At W–D25 and W–D20 however, shoot dry weight wassignificantly greater in CSSL47 than in Nipponbare(Table 4). The number of stems per plant was notsignificantly different between the two genotypes atany of the soil moisture treatments (Table 4).
The root system profiles of CSSL47 and Nippon-bare grown under CWL and W–D conditions areshown in Fig. 4. Under CWL, the root systemdevelopment was comparable between the two geno-types. At W–D30, the root system developments ofboth genotypes appeared to be slightly inhibited atalmost the same rates, compared with their water-logged controls. At W–D25 and W–D20 however, theroot system developments of both genotypes wereevidently inhibited (i.e. nodal root productions) withNipponbare exhibiting greater inhibitions than CSSL47
Plant Soil
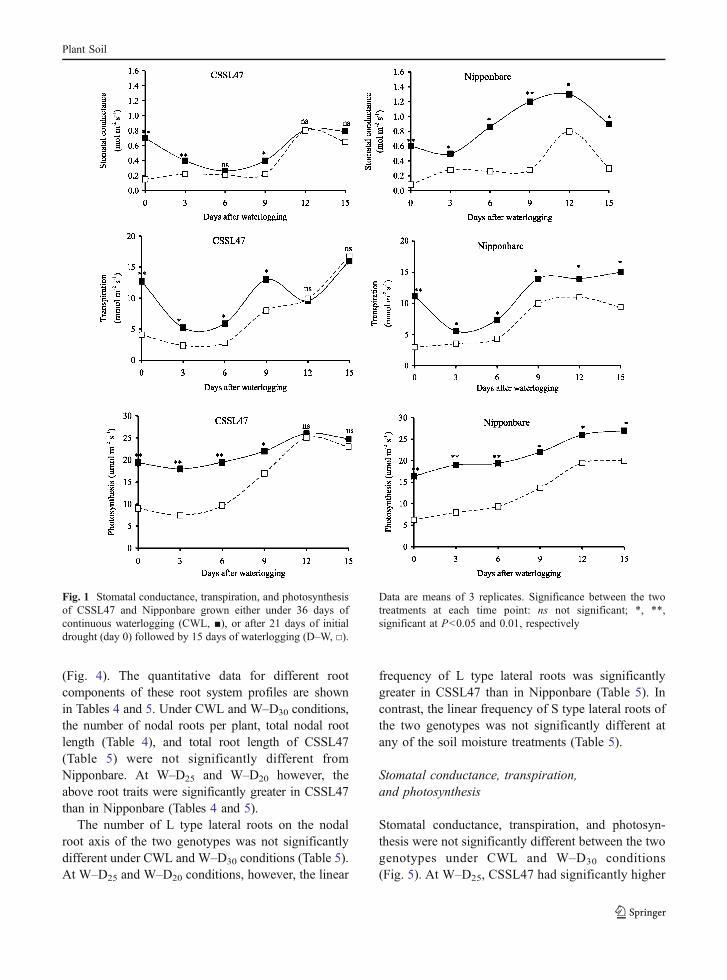
(Fig. 4). The quantitative data for different rootcomponents of these root system profiles are shownin Tables 4 and 5. Under CWL and W–D30 conditions,the number of nodal roots per plant, total nodal rootlength (Table 4), and total root length of CSSL47(Table 5) were not significantly different fromNipponbare. At W–D25 and W–D20 however, theabove root traits were significantly greater in CSSL47than in Nipponbare (Tables 4 and 5).
The number of L type lateral roots on the nodalroot axis of the two genotypes was not significantlydifferent under CWL and W–D30 conditions (Table 5).At W–D25 and W–D20 conditions, however, the linear
frequency of L type lateral roots was significantlygreater in CSSL47 than in Nipponbare (Table 5). Incontrast, the linear frequency of S type lateral roots ofthe two genotypes was not significantly different atany of the soil moisture treatments (Table 5).
Stomatal conductance, transpiration,and photosynthesis
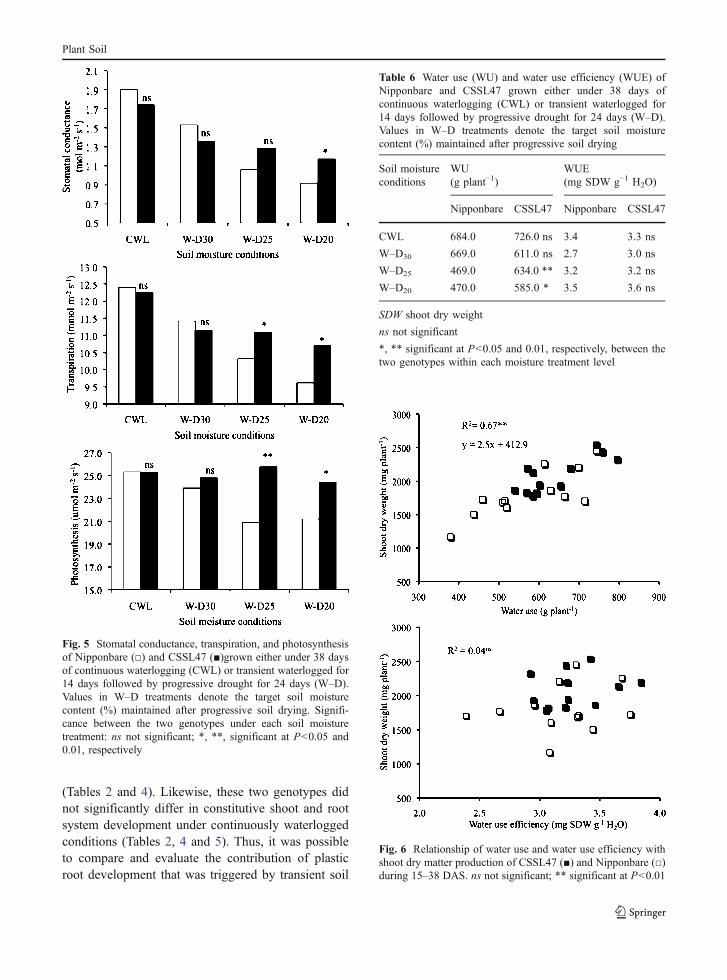
Stomatal conductance, transpiration, and photosyn-thesis were not significantly different between the twogenotypes under CWL and W–D30 conditions(Fig. 5). At W–D25, CSSL47 had significantly higher
Fig. 1 Stomatal conductance, transpiration, and photosynthesisof CSSL47 and Nipponbare grown either under 36 days ofcontinuous waterlogging (CWL, ■), or after 21 days of initialdrought (day 0) followed by 15 days of waterlogging (D–W, □).
Data are means of 3 replicates. Significance between the twotreatments at each time point: ns not significant; *, **,significant at P<0.05 and 0.01, respectively
Plant Soil
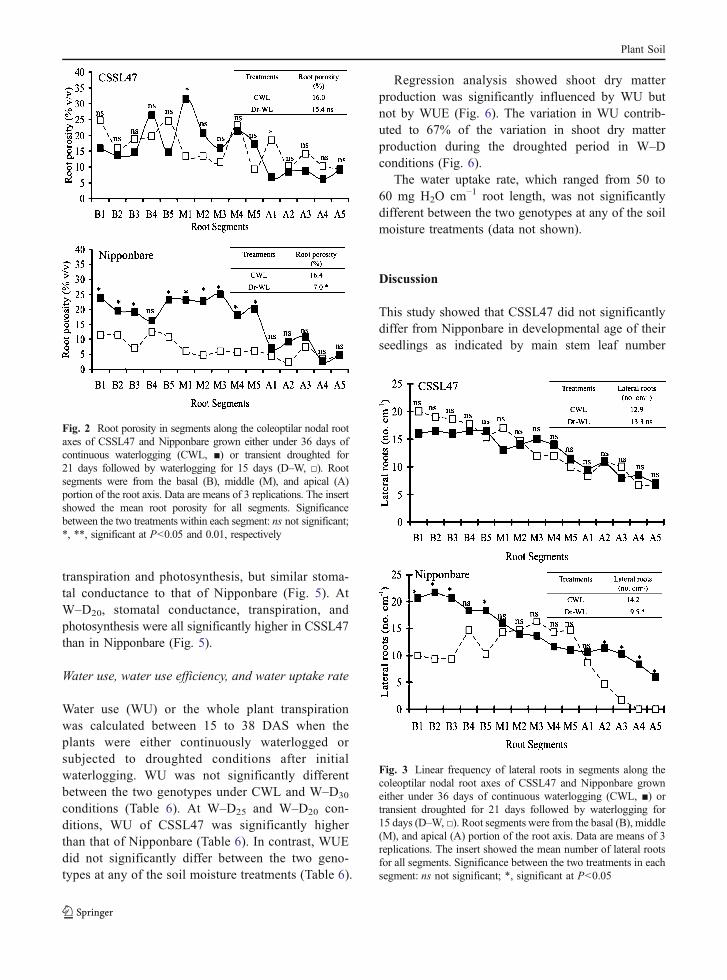
transpiration and photosynthesis, but similar stoma-tal conductance to that of Nipponbare (Fig. 5). AtW–D20, stomatal conductance, transpiration, andphotosynthesis were all significantly higher in CSSL47than in Nipponbare (Fig. 5).
Water use, water use efficiency, and water uptake rate
Water use (WU) or the whole plant transpirationwas calculated between 15 to 38 DAS when theplants were either continuously waterlogged orsubjected to droughted conditions after initialwaterlogging. WU was not significantly differentbetween the two genotypes under CWL and W–D30
conditions (Table 6). At W–D25 and W–D20 con-ditions, WU of CSSL47 was significantly higherthan that of Nipponbare (Table 6). In contrast, WUEdid not significantly differ between the two geno-types at any of the soil moisture treatments (Table 6).
Regression analysis showed shoot dry matterproduction was significantly influenced by WU butnot by WUE (Fig. 6). The variation in WU contrib-uted to 67% of the variation in shoot dry matterproduction during the droughted period in W–Dconditions (Fig. 6).
The water uptake rate, which ranged from 50 to60 mg H2O cm−1 root length, was not significantlydifferent between the two genotypes at any of the soilmoisture treatments (data not shown).
Discussion
This study showed that CSSL47 did not significantlydiffer from Nipponbare in developmental age of theirseedlings as indicated by main stem leaf number
Fig. 3 Linear frequency of lateral roots in segments along thecoleoptilar nodal root axes of CSSL47 and Nipponbare growneither under 36 days of continuous waterlogging (CWL, ■) ortransient droughted for 21 days followed by waterlogging for15 days (D–W, □). Root segments were from the basal (B), middle(M), and apical (A) portion of the root axis. Data are means of 3replications. The insert showed the mean number of lateral rootsfor all segments. Significance between the two treatments in eachsegment: ns not significant; *, significant at P<0.05
Fig. 2 Root porosity in segments along the coleoptilar nodal rootaxes of CSSL47 and Nipponbare grown either under 36 days ofcontinuous waterlogging (CWL, ■) or transient droughted for21 days followed by waterlogging for 15 days (D–W, □). Rootsegments were from the basal (B), middle (M), and apical (A)portion of the root axis. Data are means of 3 replications. The insertshowed the mean root porosity for all segments. Significancebetween the two treatments within each segment: ns not significant;*, **, significant at P<0.05 and 0.01, respectively
Plant Soil
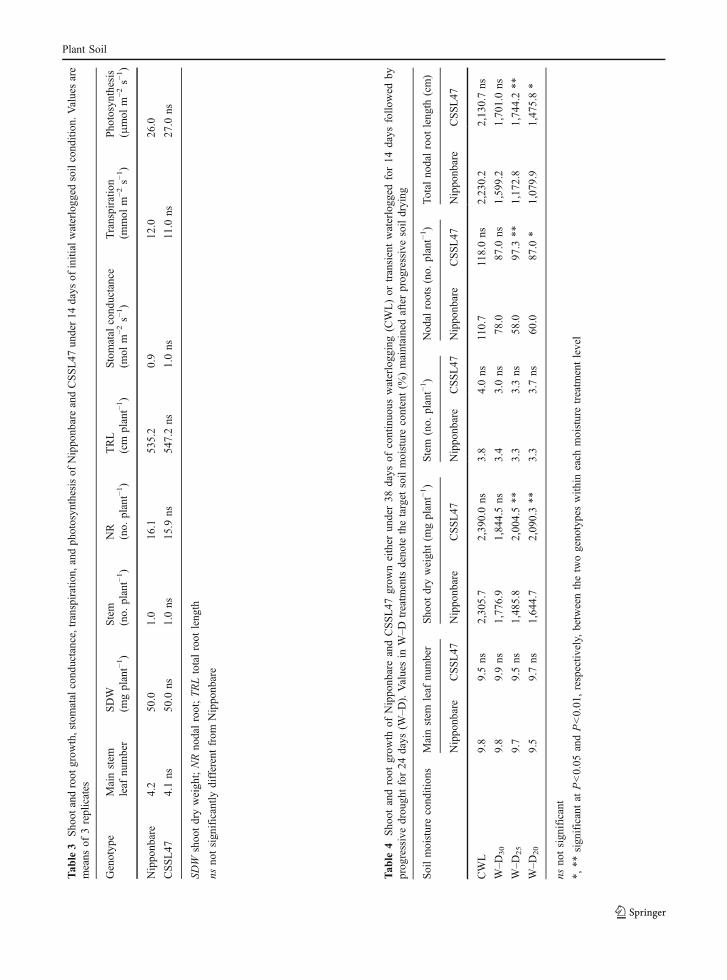
Tab
le3
Sho
otandroot
grow
th,stomatalcond
uctance,transpiration,
andph
otosyn
thesisof
Nippo
nbareandCSSL47
under14
days
ofinitialwaterlogg
edsoilcond
ition
.Valuesare
means
of3replicates
Genotyp
eMainstem
leaf
number
SDW
(mgplant−1)
Stem
(no.
plant−1)
NR
(no.
plant−1)
TRL
(cm
plant−1)
Stomatal
cond
uctance
(mol
m−2s−
1)
Transpiratio
n(m
mol
m−2
s−1)
Pho
tosynthesis
(μmol
m−2
s−1)
Nippo
nbare
4.2
50.0
1.0
16.1
535.2
0.9
12.0
26.0
CSSL47
4.1ns
50.0
ns1.0ns
15.9
ns54
7.2ns
1.0ns
11.0
ns27
.0ns
SDW
shoo
tdryweigh
t;NRno
dalroot;TRLtotalroot
leng
th
nsno
tsign
ificantly
differentfrom
Nippo
nbare
Tab
le4
Sho
otandroot
grow
thof
Nippo
nbareandCSSL47
grow
neither
under38
days
ofcontinuo
uswaterlogg
ing(CWL)or
transientwaterlogg
edfor14
days
follo
wed
byprog
ressivedrou
ghtfor24
days
(W–D
).Valuesin
W–D
treatm
entsdenote
thetarget
soilmoisturecontent(%
)maintainedafterprogressivesoildrying
Soilmoisturecond
ition
sMainstem
leaf
number
Sho
otdryweigh
t(m
gplant−1)
Stem
(no.
plant−1)
Nod
alroots(no.
plant−1)
Total
nodalroot
leng
th(cm)
Nippo
nbare
CSSL47
Nippo
nbare
CSSL47
Nippo
nbare
CSSL47
Nippo
nbare
CSSL47
Nippo
nbare
CSSL47
CWL
9.8
9.5ns
2,30
5.7
2,39
0.0ns
3.8
4.0ns
110.7
118.0ns
2,23
0.2
2,13
0.7ns
W–D
30
9.8
9.9ns
1,77
6.9
1,84
4.5ns
3.4
3.0ns
78.0
87.0
ns1,59
9.2
1,70
1.0ns
W–D
25
9.7
9.5ns
1,48
5.8
2,00
4.5**
3.3
3.3ns
58.0
97.3
**1,17
2.8
1,74
4.2**
W–D
20
9.5
9.7ns
1,64
4.7
2,09
0.3**
3.3
3.7ns
60.0
87.0
*1,07
9.9
1,47
5.8*
nsno
tsign
ificant
*,**
sign
ificantat
P<0.05
andP<0.01
,respectiv
ely,
betweenthetwogeno
typeswith
ineach
moisturetreatm
entlevel
Plant Soil
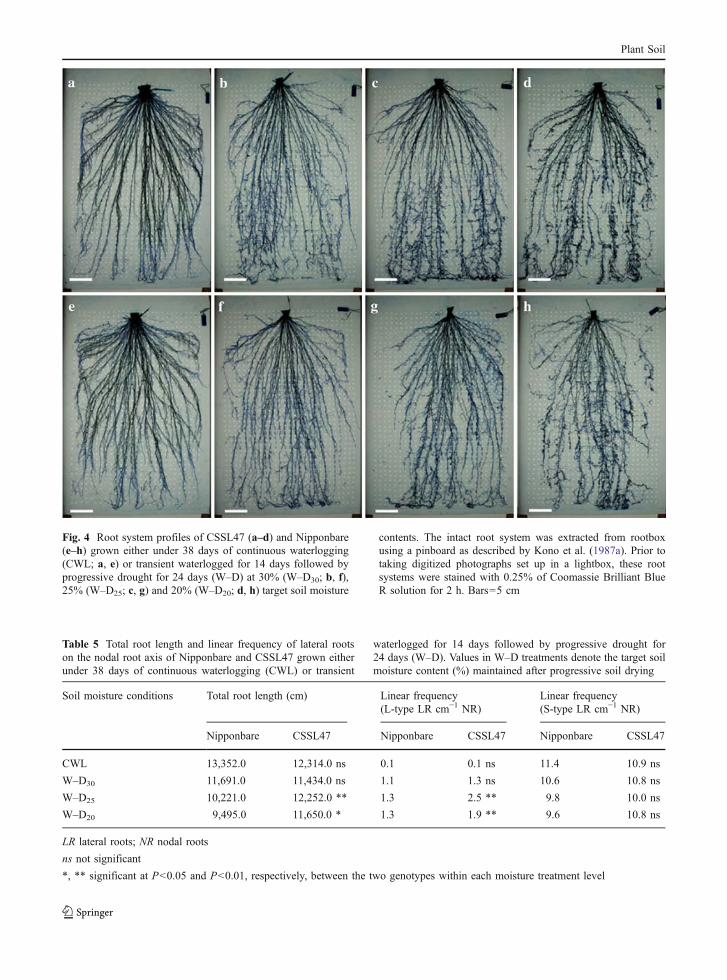
Table 5 Total root length and linear frequency of lateral roots on the nodal root axis of Nipponbare and CSSL47 grown either under38 days of continuous waterlogging (CWL) or transient waterlogged for 14 days followed by progressive drought for 24 days (W–D).Values in W–D treatments denote the target soil moisture content (%) maintained after progressive soil drying
Soil moisture conditions Total root length (cm) Linear frequency(L-type LR cm−1 NR)
Linear frequency(S-type LR cm−1 NR)
Nipponbare CSSL47 Nipponbare CSSL47 Nipponbare CSSL47
CWL 13,352.0 12,314.0 ns 0.1 0.1 ns 11.4 10.9 ns
W–D30 11,691.0 11,434.0 ns 1.1 1.3 ns 10.6 10.8 ns
W–D25 10,221.0 12,252.0 ** 1.3 2.5 ** 9.8 10.0 ns
W–D20 9,495.0 11,650.0 * 1.3 1.9 ** 9.6 10.8 ns
LR lateral roots; NR nodal roots
ns not significant
*, ** significant at P<0.05 and P<0.01, respectively, between the two genotypes within each moisture treatment level
Fig. 4 Root system profiles of CSSL47 (a–d) and Nipponbare(e–h) grown either under 38 days of continuous waterlogging(CWL; a, e) or transient waterlogged for 14 days followed byprogressive drought for 24 days (W–D) at 30% (W–D30; b, f),25% (W–D25; c, g) and 20% (W–D20; d, h) target soil moisture
contents. The intact root system was extracted from rootboxusing a pinboard as described by Kono et al. (1987a). Prior totaking digitized photographs set up in a lightbox, these rootsystems were stained with 0.25% of Coomassie Brilliant BlueR solution for 2 h. Bars=5 cm
Table 5 Total root length and linear frequency of lateral rootson the nodal root axis of Nipponbare and CSSL47 grown eitherunder 38 days of continuous waterlogging (CWL) or transient
waterlogged for 14 days followed by progressive drought for24 days (W–D). Values in W–D treatments denote the target soilmoisture content (%) maintained after progressive soil drying
Plant Soil
(Tables 2 and 4). Likewise, these two genotypes didnot significantly differ in constitutive shoot and rootsystem development under continuously waterloggedconditions (Tables 2, 4 and 5). Thus, it was possibleto compare and evaluate the contribution of plasticroot development that was triggered by transient soil
Fig. 5 Stomatal conductance, transpiration, and photosynthesisof Nipponbare (□) and CSSL47 (■)grown either under 38 daysof continuous waterlogging (CWL) or transient waterlogged for14 days followed by progressive drought for 24 days (W–D).Values in W–D treatments denote the target soil moisturecontent (%) maintained after progressive soil drying. Signifi-cance between the two genotypes under each soil moisturetreatment: ns not significant; *, **, significant at P<0.05 and0.01, respectively
Table 6 Water use (WU) and water use efficiency (WUE) ofNipponbare and CSSL47 grown either under 38 days ofcontinuous waterlogging (CWL) or transient waterlogged for14 days followed by progressive drought for 24 days (W–D).Values in W–D treatments denote the target soil moisturecontent (%) maintained after progressive soil drying
Soil moistureconditions
WU(g plant−1)
WUE(mg SDW g−1 H2O)
Nipponbare CSSL47 Nipponbare CSSL47
CWL 684.0 726.0 ns 3.4 3.3 ns
W–D30 669.0 611.0 ns 2.7 3.0 ns
W–D25 469.0 634.0 ** 3.2 3.2 ns
W–D20 470.0 585.0 * 3.5 3.6 ns
SDW shoot dry weight
ns not significant
*, ** significant at P<0.05 and 0.01, respectively, between thetwo genotypes within each moisture treatment level
Fig. 6 Relationship of water use and water use efficiency withshoot dry matter production of CSSL47 (■) and Nipponbare (□)during 15–38 DAS. ns not significant; ** significant at P<0.01
Plant Soil

moisture stresses and their effects on physiologicalresponses such as stomatal conductance, transpiration,and photosynthesis and ultimately, shoot dry matterproduction with minimal effects of genetic confound-ing. Furthermore, the shoot and root development werenot significantly different between the two genotypesunder the initial drought of D–W treatment (Table 1),and initial waterlogged conditions (Tables 1 and 3).These facts further indicate that better growth inCSSL47 than Nipponbare under both transient soilmoisture stresses can be attributed to more promotedroot development of CSSL47 than Nipponbare inresponse to the second transient stress, which possiblywas influenced by the preceding moisture stress(initially droughted or waterlogged conditions). Then,such plastic root development in CSSL47 in turn isattributable to either one or combinations of thesubstituted segments from Kasalath inserted intoNipponbare genetic background found in this line.Some of the QTL markers contained in the substitutedsegments in CSSL47 coincided with the QTLsreportedly detected in other mapping populations suchas root-to-shoot ratio, maximum root length, and rootthickness at the base and below 90-cm soil depth(Champoux et al. 1995; Price et al. 2002; MacMillan etal. 2006). However, none of the marker-linked QTLscontrolling lateral root production identified in othermapping populations (i.e. Horii et al. 2006; Zheng etal. 2003) coincided with any of the markers containedin the substituted segments of CSSL47.
Droughted followed by waterlogged (D–W) condition
Under D–W condition, although both genotypes werein similar developmental age of their seedlings basedon main stem leaf number, CSSL47 producedsignificantly greater shoot dry matter production thanNipponbare (Table 2). This was attributed to morerapid recovery of stomatal conductance, transpiration,and photosynthesis in CSSL47 than in Nipponbarewhen their droughted plants were subjected to suddenwaterlogged condition (Fig. 1). Rapid recovery instomatal conductance, transpiration, and photosynthe-sis under transient D–W conditions in CSSL47(Fig. 1) was due to its faster root system developmentand recovery than Nipponbare during the transientwaterlogged period (Table 2). It was also noted thatSRL of CSSL47 and Nipponbare was similar underCWL conditions (Table 2). In contrast, SRL of
CSSL47 was significantly higher than that ofNipponbare under D–W condition. An increase inSRL is a common response to limitations in nutrient(Schippers and Olff 2000) and water (Bañoc et al.2000a; Tjoelker et al. 2005). The greater SRL foundin CSSL47 than in Nipponbare under transient D–Wcondition may reflect promoted branching into lateralroots and/or enhanced aerenchyma formation underwaterlogged condition, which was related to theincrease in root porosity (Eissenstat 1991; Raumet etal. 2006). As such, droughted roots of CSSL47subjected to sudden waterlogging exhibited fastergrowth for fast and efficient acquisition of resourcessuch as nutrients, to cope up with its overall growth.
Increased root porosity due to the formation ofaerenchyma is one of the most common adaptiveresponses of plants such as rice to O2 deficiency (Colmer2003; Jackson and Armstrong 1999; Vartapetian andJackson 1997; Wang and Yamauchi 2006). Thus, thegeneral belief is that waterlogging cannot be stressfulfor rice. In this study, however, it is interesting to notethat waterlogging is also stressful to rice especiallywhen this stress is preceded by drought. This wasclearly shown in Nipponbare in which the ability toincrease root porosity under sudden waterlogged con-ditions was lost when roots were previously exposed todrought (Fig. 2). On the other hand, CSSL47 was ableto increase its root porosity under sudden waterloggedconditions even when its roots were previously exposedto drought (Fig. 2). Such increase in root porosity inCSSL47 facilitated higher efficiency of its root systemfor internal diffusion of atmospheric O2 to the root tipsunder transient waterlogging (Table 2). The aboveconditions may in part, explain the higher ability forseminal root elongation of CSSL47 than Nipponbare(Table 2) and more branching of lateral roots on thecoleoptilar nodal root axis (Fig. 3), which were exposedboth to initially droughted and transient waterloggedconditions. Furthermore, the ability of CSSL47’sdroughted roots to continuously grow when subjectedto transient waterlogging may also explain its higherSRL than Nipponbare under D–W condition (Table 2).Such ability is indispensable as it contributed to theincrease in shoot dry matter production (Table 2) viarapid recoveries in stomatal conductance, transpiration,and photosynthesis (Fig. 1). The increased shoot growthin turn, provided materials for production and elonga-tion of new nodal roots (Table 2). The ability toproduce more number of nodal roots, coupled with
Plant Soil

faster elongation, translates to the development of morelateral roots. Since lateral roots comprise the greaterportion of the total length of the whole root system(Wang et al. 2009; Yamauchi et al. 1987), theirresponse to moisture stress directly determines totalroot system development. Under O2 deficiency, thelateral roots serve as effective sites for radial oxygenloss, an essential process for detoxifying phytotoxinsand oxidizing reduced nutrient element to maintainthe viability of roots for efficient nutrient and wateruptake (Armstrong and Armstrong 2005; Revsbechet al. 1999). This eventually support higher shoot drymatter production.
Waterlogged followed by droughted (W–D)conditions
Under W–D condition, both genotypes were insimilar developmental age of their seedlings basedon main stem leaf number in any of the soil moisturetreatments (Table 4). However, CSSL47 showedhigher shoot dry matter production than Nipponbareespecially at W–D25 and W–D20 (Table 4). Suchgreater dry matter production was attributed to thehigher ability of CSSL47 than Nipponbare to main-tain stomatal conductance, transpiration, and photo-synthesis under progressive droughted conditions(Fig. 5).
Dry matter production, especially under water-limited condition, is determined by WU and WUE(Kobata et al. 1996) of the plant. In this study, drymatter production under progressive drought in W–Dcondition was closely related to WU but not to WUE(Fig. 6). This indicates that adaptation of rice underwater-limited conditions in terms of growth andproductivity could be achieved mainly through dehy-dration avoidance by maintaining water uptake ratherthan by tolerance to desiccation, as pointed out byBlum (2005), which was accomplished by maintaininghigher root growth under water-limited soil conditions.
Apparently, CSSL47 had greater ability to main-tain root elongation compared to Nipponbare underW–D25 and W–D20, due to its higher ability tomaintain total nodal root production and plasticlateral root production, especially the L type lateralroots (Table 5). Our result agrees with that of Bañocet al. (2000b), which showed that plasticity in lateralroot production, particularly the L type, plays a keyrole in rice adaptation to waterlogged conditions
followed by drought and rewatering. Such trait inroot plasticity resulted in increased root surface areawhich subsequently increased soil water extractions(Kamoshita et al. 2000, 2004; Siopongco et al. 2005,2006). The greater ability for root growth expressedin CSSL47 than in Nipponbare under progressivedrought, contributed to its higher WU (Table 6),which resulted in the maintenance of stomatalconductance, transpiration, and photosynthesis(Fig. 5). This ultimately led to a higher shoot drymatter production in CSSL47 than in Nipponbare(Table 4).
CSSL47 is unique under transient soil moisturestresses
Shoot and root system development of CSSL47 iscomparable with Nipponbare under constantdroughted condition (Table 1). This indicates thatthey may have similar genetic control for rootresponses to drought. This is consistent with that ofKanou et al. (2007) who showed that CSSL47 didnot exhibit higher root growth than Nipponbareunder constant drought. CSSL47 also showed com-parable shoot and root growth with Nipponbareunder constant waterlogging (Tables 1, 2, 3, 4 and5), which was consistent with the result from ourearlier shorter-duration hydroponics experiment(Suralta et al. 2008b). This indicates that plasticroot system development of CSSL47 was a uniqueresponse to transient soil moisture stress condi-tions. The root system adaptation of CSSL47 tosubsequent moisture stress was not affected by thepreceding contrasting moisture stress, which isdifferent from that of Nipponbare. Field studiesalso confirmed that CSSL47 produced a signifi-cantly higher yield than Nipponbare under inter-mittent irrigations where there was alternaterecurrence of O2 deficiency and progressivedrought. The two genotypes, however, producedthe same yield under continuously waterloggedconditions (Niones et al. 2009; Suralta et al. 2007).These findings clearly show that CSSL47 is betteradapted to the cycle of drought and waterloggingstresses in the field.
Detailed analyses, however, on plastic root devel-opmental responses especially from the upper nodesto different timing of transient water stresses (i.e.progressive drought and sudden waterlogging) at
Plant Soil

different stage of growth (i.e. vegetative and repro-ductive stage) are ongoing. This is done to validatethe plastic root responses found at the seedling stagein the present study and to quantify their contributionto the maintenance of shoot dry matter production andgrain yield.
Conclusion
Our earlier short-duration hydroponics study (Suraltaet al. 2008b) have selected suitable rice line (CSSL47)which showed greater ability for root plastic devel-opmental responses to transient moisture stressesthan Nipponbare. In the present study, however, wehave precisely quantified that more plastic rootdevelopment of CSSL47 than Nipponbare and theirassociated physiological functions under prolongedtransient soil moisture stresses contribute to themaintenance of its shoot dry matter production.Specifically under D-W condition, the root plasticitywas expressed in aerenchyma formation. Namely,CSSL47 showed greater shoot dry matter productionthan Nipponbare due to higher rate of recovery ofthe former genotype in terms of stomatal conduc-tance, transpiration, and photosynthesis in responseto subsequent waterlogging. Such recoveries weresupported by greater root system development ofCSSL47 than Nipponbare, owing to its greaterability to enhance aerenchyma system to facilitateO2 diffusion to the root tips under D–W condition.On the other hand, under W–D condition at lowSMCs, the root plasticity was expressed in lateralroot branching. CSSL47 showed greater shoot drymatter production than Nipponbare due to its greaterability in maintaining stomatal conductance, transpi-ration, and photosynthesis. Such greater ability formaintenance of stomatal conductance, transpiration,and photosynthesis was attributed to CSSL47’sgreater root system development due to promotedbranching, which facilitated higher water extractionthan Nipponbare during the drought period underW–D condition.
Acknowledgment We thank Dr. Abdelbagi M. Ismail (Inter-national Rice Research Institute, Philippines) for a criticalreview and useful comments on our manuscript. This researchwas supported by Grant-in-Aid for Scientific Research (No.19380011) from the Japan Society for the Promotion ofScience.
References
Armstrong W (1971) Radial oxygen losses from intact riceroots as affected by distance from the apex, respiration,and waterlogging. Physiol Plant 25:192–197
Armstrong J, Armstrong W (2005) Rice: sulfide-inducedbarriers to radial oxygen loss, Fe+ and water uptake, andlateral root emergence. Ann Bot 96:625–638
Azhiri-Sigari T, Yamauchi A, Kamoshita A, Wade LJ (2000)Genotypic variation in response of rainfed lowland rice todrought and rewatering. II. Root growth. Plant Prod Sci3:180–188
Bañoc DM, Yamauchi A, Kamoshita A, Wade LJ, Pardales JRJr (2000a) Genotypic variations in response of lateral rootdevelopment to fluctuating soil moisture in rice. PlantProd Sci 3:335–343
Bañoc DM, Yamauchi A, Kamoshita A, Wade LJ, Pardales JRJr (2000b) Genotypic variations in response of lateral rootdevelopment to fluctuating soil moisture in rice. PlantProd Sci 3:335–343
Blum A (2005) Drought resistance, water-use efficiency andyield potential—are they compatible, dissonant and mutu-ally exclusive? Aust J Agr Res 56:1159–1168
Bouman BAM, Toung TP (2001) Field water management tosave water and increased its productivity in irrigatedlowland rice. Agric Water Manag 49:11–30
Champoux MC, Wang G, Sarkarung S, Mackill DJ, O’TooleJC, Huang N, MacCouch SR (1995) Locating genesassociated with root morphology and drought avoidancein rice via linkage to molecular markers. Theor Appl Gene90:969–981
Colmer TD (2003) Aerenchyma and an inducible barrier toradial oxygen loss facilitate root aeration in upland, paddyand deepwater rice (Oryza sativa L.). Ann Bot 91:301–309
Eissenstat D (1991) On the relationship between specific rootlength and the rate of root proliferation: a field study usingcitrus rootstocks. New Phytol 118:63–68
Horii H, Nemoto K, Miyamoto N, Harada J (2006) Quantitativetrait loci for adventitious and lateral roots in rice. PlantBreed 125:198–200
Ingram KT, Bueno FD, Namuco OS, Yambao EB, BeyrouryCA (1994) Rice root traits for drought resistance and theirgenetic variation. In: Kirk CJD (ed) Rice roots: nutrientand water use. International Rice Research Institute,Manila, pp 66–67
Ito K, Tanakamaru K, Morita S, Abe J, Inanaga S (2006)Lateral root development, including responses to soildrying, of maize (Zea mays) and wheat (Triticumaestivum) seminal roots. Physiol Plant 127:260–267
Jackson MB, Armstrong W (1999) Formation of aerenchymaand the process of plant ventilation in relation to soilflooding and submergence. Plant Biol 1:274–287
Justin SHFW, Armstrong W (1987) The anatomical character-istics of roots and plant response to soil flooding. NewPhytol 106:465–495
Kamoshita A, Wade LJ, Yamauchi A (2000) Genotypicvariation in response of rainfed lowland rice to droughtand rewatering. III. Water extraction during the droughtperiod. Plant Prod Sci 3:189–196
Plant Soil

Kamoshita A, Zhang J, Siopongco J, Sarkarung S, Nguyen HT,Wade LJ (2002a) Effects of phenotyping environment onidentification of QTL for rice root morphology underanaerobic conditions. Crop Sci 42:255–265
Kamoshita A, Wade LJ, Ali MM, Pathan MS, Zhang J,Sarkarung S, Nguyen HT (2002b) Mapping QTLs forroot morphology of a rice population adapted to rainfedlowland conditions. Theor Appl Genet 104:880–893
Kamoshita A, Rodriguez R, Yamauchi A, Wade LJ (2004)Genotypic variation in response of rainfed lowland rice toprolonged drought and rewatering. Plant Prod Sci 7:406–420
Kanou M, Inukai Y, Kitano H, Yamauchi A (2007) Quantitativeevaluation of contribution of root plasticity to shoot drymatter production by using chromosome segment substi-tution lines in rice. Jpn J Crop Sci (Extra 1) 76:186–187,in Japanese with English summary
Kimura K, Yamasaki S (2001) Root length and diametermeasurement using NIH Image: application of the line-intercept principle for diameter estimation. Plant Soil234:37–46
Kimura K, Kikuchi S, Yamasaki S (1999) Accurate rootlength measurement by image analysis. Plant Soil 216:117–127
Kobata T, Okuno T, Yamamoto T (1996) Contribution ofcapacity for soil water extraction and water use efficiencyto maintenance of dry matter production in rice subjectedto drought. Jpn J Crop Sci 65:652–662
Kono Y, Yamauchi A, Nonoyama T, Tatsumi J, Kawamura N(1987a) A revised experimental system of root–soil interac-tion for laboratory work. Environ Cont Biol 25:141–151
Kono Y, Tomida K, Tatsumi J, Nonoyama T, Yamauchi A,Kitano J (1987b) Effects of soil moisture conditions on thedevelopment of root systems of soybean plants (Glycinemax Merr.). Jpn J Crop Sci 56:597–607
Lee RW (2003) Physiological adaptations of the invasivecordgrass Spartina anglica to reducing sediments: rhizomemetabolic gas fluxes and enhanced O2 and H2S transport.Mar Biol 143:9–15
MacMillan K, Emrich K, Piepho HP, Mullins CE, Price AH(2006) Assessing the importance of genotype X environmentinteraction for root traits in rice using a mapping populationII: conventional QTL analysis. Theor Appl Genet 113:953–964
Maricle BR, Lee RW (2007) Root respiration and oxygen fluxin salt marsh grasses from different elevational zones.Marine Biol 151:413–423
Niones J, Suralta R, Inukai Y, Yamauchi A (2009) Evaluationof functional roles of plastic responses of root system indry matter production and yield under continuous cycle oftransient soil moisture stresses by using chromosomesegment substitution lines in rice under field conditions.Jpn J Crop Sci (Extra 1) 78:260–261
O’Toole JC, Bland WL (1987) Genotypic variation in cropplant root systems. Adv Agron 41:91–145
Pardales JR Jr, Yamauchi A (2003) Regulation of rootdevelopment in sweetpotato and cassava by soil moistureduring their establishment period. Plant Soil 255:201–208
Price AH, Steele KA, Moore BJ, Jones RGW (2002) Uplandrice grown in soil filled chambers and exposed tocontrasting water-deficit regimes II. Mapping quantitative
trait loci for root morphology and distribution. Field CropRes 76:25–43
Raumet C, Urcelay C, Diaz S (2006) Suites of root traits differbetween annual and perennial species growing in the field.New Phytol 170:357–368
Revsbech NP, Pedersen O, Reichart W, Briones A (1999)Microsensor analysis of oxygen and pH in the ricerhizosphere under field and laboratory conditions. BiolFert Soils 29:379–385
Schippers P, Olff H (2000) Biomass partitioning, architectureand turnover of six herbaceous species from habitats withdifferent nutrient supply. Plant Ecol 149:219–231
Siopongco JDLC, Yamauchi A, Salekdeh H, Bennett J,Wade LJ (2005) Root growth and water extractionresponses of doubled-haploid rice lines to drought andrewatering during the vegetative stage. Plant Prod Sci8:497–508
Siopongco JDLC, Yamauchi A, Salekdeh H, Bennett J, WadeLJ (2006) Growth and water use response of doubledhaploid rice lines to drought and rewatering during thevegetative stage. Plant Prod Sci 9:141–151
Siopongco JDLC, Sekiya K, Yamauchi A, Egdane J, IsmailAM, Wade LJ (2008) Stomatal responses in rainfedlowland rice to partial soil drying; evidence of rootsignals. Plant Prod Sci 11:28–41
Siopongco JDLC, Sekiya K, Yamauchi A, Egdane J, IsmailAM, Wade LJ (2009) Stomatal responses in rainfedlowland rice to partial soil drying; comparison of twolines. Plant Prod Sci 12:17–28
Subere JOQ, Bolatete D, Bergantin R, Pardales A, Belmonte JJ,Mariscal A, Sebidos R, Yamauchi A (2009) Genotypicvariation in responses of cassava (Manihot esculentaCrantz) to drought and rewatering. I. Root systemdevelopment. Plant Prod Sci 12:462–474
Suralta RR, Yamauchi A (2008) Root growth, aerenchymadevelopment, and oxygen transport in rice genotypessubjected to drought and waterlogging. Environ Exp Bot64:75–82
Suralta RR, Inukai Y, Kitano H, Yamauchi A (2007) Responsesin dry matter production, yield and their related develop-mental and physiological traits of rice genotypes to thecycle of waterlogging and drought under field conditions.Jpn J Crop Sci (Extra 2) 76:262–263
Suralta RR, Inukai Y, Yamauchi A (2008a) Genotypicvariations in responses of lateral root development totransient moisture stresses in rice cultivars. Plant Prod Sci11:324–335
Suralta RR, Inukai Y, Yamauchi A (2008b) Utilizing chromo-some segment substitution lines (CSSLs) for evaluation ofroot responses under transient moisture stresses in rice.Plant Prod Sci 11:457–465
Teal JM, Kanwisher JW (1966) Gas transport in the marshgrass, Spartina alterniflora. J Exp Bot 17:355–361
Tjoelker MG, Craine JM, Wedin D, Reich PB, Tilman D (2005)Linking leaf and root trait syndromes among 39 grasslandand savannah species. New Phytol 167:493–508
Vartapetian BB, Jackson MB (1997) Plant adaptation toanaerobic stress. Ann Bot 73:3–20
Visser EJW, Bögenmann GM (2003) Measurement ofporosity in very small samples of plant tissue. PlantSoil 253:81–90
Plant Soil

Wade LJ, Kamoshita A, Yamauchi A, Azhiri-Sigari T (2000)Genotypic variation in response of rainfed rowland rice todrought and rewatering I. Growth and water use. PlantProd Sci 3:173–179
Wang H, Yamauchi A (2006) Growth and function of rootsunder abiotic stress soils. In: Huang B (ed) Plant–environment interactions, 3rd edn. CRC, Taylor andFrancis Group, LLC, New York, pp 271–320
Wang H, Inukai Y, Kamoshita A, Wade LJ, SiopongcoJDLC, Nguyen H, Yamauchi A (2005) QTL analysison plasticity in lateral root development in response towater stress in the rice plant. In: Toriyama K, HeongKL, Hardy B (eds) Rice is life: scientific perspectivesfor the 21st century. The Proceedings of the WorldRice Research Conference, Tsukuba, Japan, pp 464–469
Wang H, Siopongco JDLC, Wade LJ, Yamauchi A (2009)Fractal analysis on root systems of rice plants in responseto drought stress. Environ and Exp Bot 65:338–344
Yamauchi A, Kono Y, Tatsumi J (1987) Quantitative analysison root system structures of upland rice and maize. Jpn JCrop Sci 56:608–617
Yamauchi A, Pardales JR Jr, Kono Y (1996) Root systemstructure and its relation to stress tolerance. In: Ito O,Katayama K, Johansen C, Kumar Rao JVDK, Adu-GyamfiJJ, Rego TJ (eds) Roots and nitrogen in cropping systems ofthe semi-arid tropics. JIRCAS, Tsukuba, pp 211–234
Zheng BS, Yang L, Zhang WP, Mao CZ, Wu YR, Yi KK, LiuFY, Wu P (2003) Mapping QTLs and candidate genes forrice root traits under different water-supply conditions andcomparative analysis across three populations. Theor ApplGenet 107:1505–1515
Plant Soil
Related Documents











