Drug resistance in Leishmania: similarities and differences to other organisms Barbara Papadopoulou, Christoph KOndig, Ajay Singh,Marc Ouellette Centrede Recherche en Infectiotogie du Centre de Recherche du CHULet D~partrnent de Biologie M~dicale~ Division de Microbiologie, Facult~ de M~decine, Universit~ Laval, Quebec, Canada GIV 4G2. Abstract The main line of defense available against parasitic protozoa is chemotherapy. Drug resistance has emerged however, as a primary obstacle to the successful treatment and control of parasitic diseases. Leishmania spp., the causative agents of leishmaniasis, have served as a useful model for studying mechanisms of drug resistance in vitro.Antimonials and amphotericin B are the first line drugs to treat Leishmania followed by pentamidine and a number of other drugs. Parasites resistant against all these classes of drugs have been selected under laboratory conditions.A multiplicity of resistance mechanisms has been detected, the most prevalent being gene amplification and transport mutations.With the tools now available, it should be possible to elucidate the mechanisms that govern drug resistance in field isolates and develop more effective chemotherapeutic agents. LEISHMANIA arasites of the genus Leishmania are distributed worldwide, and 10-15 million people are estimated to be infected with 400 000 new cases each year. ~ Close to a third of the world population lives in endemic areas and is at risk of contracting an infection. Leishmania are endemic from the USA-Mexico border through all of Central America to several of the South American countries; around the Mediterranean sea; in North and East Africa; in Southern Russia, the Middle East, India and China. Cases of Leishmania infections are rising even in developed countries. Increases in travelling and intervention in regional conflicts such as 'Operation Desert Storm '2 have raised the number of Leishmania cases in non-endemic areas. Incidence of leishmaniasis is rising also because of the lack of vaccines, difficult vector control, and increase in resistance to chemotherapy? Leishmania has also emerged as a serious opportunistic pathogen in HIV-infected humans, and several cases have been reported around the Mediterranean littoral) Currently, the number of cases of visceral disease due to L. infantum among HIV-illfected patients in Southern Europe is approximately equal to the number of such cases among non-HIV-infected individuals. 4 Leishmania spp. are responsible for a wide spectrum of diseases in humans and donlestic animals.The clinical mani- festations of leishmaniasis vary depending on the pathogenic species; they encompass cutaneous leishmaniasis (oriental sore), mucocutaneous leishmaniasis (espundia), and visceral leishmaniasis (kala-azar).The latter is the most severe form of the disease, and it is usually fatal if left untreated. The life cycle of the parasite is relatively simple. Flagellated pro- mastigotes are transmitted to humans by the phlebotomine female sandfly. Parasites are engulfed by macrophages, where the promastigotes differentiate into aflagellated amastigotes within the phagolysosomes of the host macrophages. Upon Leishmania infection, several macrophage functions are altered, allowing parasites to replicate and escape the host defense mechanisms, s CURRENT CHEMOTHERAPY The only effective way to control Leishmania infections currently is chemotherapy. The antileishmanial drugs avail- able have recently been reviewed. 6,7The first agents with a favorable therapeutic index, the pentavalent andmonials (SbV), were introduced in the 1940s and are still the main- stay of therapy for all forms of leishmaniasis. Quite remark- ably, the mode of action of antimonials in Leishmania is obscure, although studies on the mechanism(s) of resistance may eventually pinpoint the target(s). 8 Formulations available include sodium stibogluconate (Pentostam) in which antimony is complexed with gluconic acid, and meglumine antimonate (Gincantime) where antimony is complexed with the sugar meglumine. The treatment of visceral leishmaniasis with antimonial regimens is compli- cated by drug toxicity (cardiac and renal) and by drug resis- tance. Unresponsiveness to SbV-containing agents occurs in 5-70% of the patients, and resistance rates as high as 40-50% are found in some endemic regions 3,9particularly in India. Resistance to SbV is so prominent in India that it is often not recommended as a first-line drug. 7 Treatment, therefore, relies on second-line drugs such as pentamidine, which is effective in the treatment of several Leishmania infections. 7 Similarly to SbV, the mechanism of action of pen- tamidine is not well understood. Possibly it interferes with the transport and synthesis of polyamines 1°'" which are required for the synthesis of trypanothione (N1,N8- bisglu- tathionyl spermidine), a metabolite necessary to cope with oxidative stress/2 Long-term regimen of high-dose pentami- dine is probably more toxic than antimonial regimens, and is therefore used only when it is likely to be more effective than antimony: In several regions of the world, amphotericin B (AraB) is now becoming the treatment of choice for visceral leish- maniasis, specially in cases refractory to pentamidine and/or SbV treatment. ~3 The target of this drug is ergosterol-like sterols which, as in fungi, are the major membrane sterols of Leishmania spp. 14 AraB formulations (liposomes, lipid complexes, a colloidal dispersion) maintain potency while reducing toxicity significantly.The lipid formulations of AmB however, have the significant disadvantage of high cost for a disease more prevalent in developing countries. Although these formulations were initially made to treat mycoses, visceral leishmaniasis may be the disease for which AraB formulations have the best therapeutic index, as they are taken up in the reticuloendothelial system where Leishmania reside. Drug ResistanceUpdates (I 998) I, 266-278 © 1998Harcourt Brace & Co. Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Drug resistance in Leishmania: similarities and differences to other organisms
Barbara Papadopoulou, Christoph KOndig, Ajay Singh, Marc Ouellette
Centre de Recherche en Infectiotogie du Centre de Recherche du CHUL et D~partrnent de Biologie M~dicale~ Division de Microbiologie, Facult~ de M~decine, Universit~ Laval, Quebec, Canada G IV 4G2.
A b s t r a c t The main line of defense available against parasitic protozoa is chemotherapy. Drug resistance has emerged however, as a primary obstacle to the successful treatment and control of parasitic diseases. Leishmania spp., the causative agents of leishmaniasis, have served as a useful model for studying mechanisms of drug resistance in vitro.Antimonials and amphotericin B are the first line drugs to treat Leishmania followed by pentamidine and a number of other drugs. Parasites resistant against all these classes of drugs have been selected under laboratory conditions.A multiplicity of resistance mechanisms has been detected, the most prevalent being gene amplification and transport mutations.With the tools now available, it should be possible to elucidate the mechanisms that govern drug resistance in field isolates and develop more effective chemotherapeutic agents.
LEISHMANIA
arasites of the genus L e i s h m a n i a are distributed worldwide, and 10-15 million people are estimated to be infected with 400 000 new cases each year. ~
Close to a third of the world populat ion lives in endemic areas and is at risk of contracting an infection. L e i s h m a n i a are endemic from the USA-Mexico border through all of Central America to several of the South American countries; around the Mediterranean sea; in North and East Africa; in Southern Russia, the Middle East, India and China. Cases of Le i shman ia infections are rising even in developed countries. Increases in travelling and intervention in regional conflicts such as 'Operat ion Desert Storm '2 have raised the number of L e i s h m a n i a cases in non-endemic areas. Incidence of leishmaniasis is rising also because of the lack of vaccines, difficult vector control, and increase in resistance to chemotherapy? L e i s h m a n i a has also emerged as a serious opportunistic pathogen in HIV-infected humans, and several cases have been reported around the Mediterranean littoral) Currently, the number of cases of visceral disease due to L. i n f a n t u m among HIV-illfected patients in Southern Europe is approximately equal to the number of such cases among non-HIV-infected individuals. 4
L e i s h m a n i a spp. are responsible for a wide spectrum of diseases in humans and donlestic animals.The clinical mani- festations of leishmaniasis vary depending on the pathogenic species; they encompass cutaneous leishmaniasis (oriental
sore), mucocutaneous leishmaniasis (espundia), and visceral leishmaniasis (kala-azar).The latter is the most severe form of the disease, and it is usually fatal if left untreated. The life cycle of the parasite is relatively simple. Flagellated pro- mastigotes are transmitted to humans by the phlebotomine female sandfly. Parasites are engulfed by macrophages, where the promastigotes differentiate into aflagellated amastigotes within the phagolysosomes of the host macrophages. Upon L e i s h m a n i a infection, several macrophage functions are altered, allowing parasites to replicate and escape the host defense mechanisms, s
CURRENT CHEMOTHERAPY
The only effective way to control L e i s h m a n i a infections currently is chemotherapy. The antileishmanial drugs avail- able have recently been reviewed. 6,7 The first agents wi th a favorable therapeutic index, the pentavalent andmonials (SbV), were introduced in the 1940s and are still the main- stay of therapy for all forms of leishmaniasis. Quite remark- ably, the mode of action of antimonials in L e i s h m a n i a is obscure, although studies on the mechanism(s) of resistance may eventually pinpoint the target(s). 8 Formulations available include sodium stibogluconate (Pentostam) in which antimony is complexed with gluconic acid, and meglumine antimonate (Gincantime) where antimony is complexed wi th the sugar meglumine. The t reatment of visceral leishmaniasis wi th antimonial regimens is compli- cated by drug toxicity (cardiac and renal) and by drug resis- tance. Unresponsiveness to SbV-containing agents occurs in 5-70% of the patients, and resistance rates as high as 40-50% are found in some endemic regions 3,9 particularly in India. Resistance to SbV is so prominent in India that it is often not r ecommended as a first-line drug. 7 Treatment, therefore, relies on second-line drugs such as pentamidine, which is effective in the t reatment of several L e i s h m a n i a infections. 7 Similarly to SbV, the mechanism of action of pen- tamidine is not well understood. Possibly it interferes wi th the transport and synthesis of polyamines 1°'" which are required for the synthesis of t rypanothione (N1,N8- bisglu- tathionyl spermidine), a metaboli te necessary to cope with oxidative stress/2 Long-term regimen of high-dose pentami- dine is probably more toxic than antimonial regimens, and is therefore used only w h e n it is likely to be more effective than antimony:
In several regions of the world, amphotericin B (AraB) is n o w becoming the treatment of choice for visceral leish- maniasis, specially in cases refractory to pentamidine and/or SbV treatment. ~3 The target of this drug is ergosterol-like sterols which, as in fungi, are the major membrane sterols of L e i s h m a n i a spp. 14 AraB formulations (liposomes, lipid complexes, a colloidal dispersion) maintain potency while reducing toxicity significantly.The lipid formulations of AmB however, have the significant disadvantage of high cost for a disease more prevalent in developing countries. Although these formulations were initially made to treat mycoses, visceral leishmaniasis may be the disease for which AraB formulations have the best therapeutic index, as they are taken up in the reticuloendothelial system where L e i s h m a n i a reside.
Drug Resistance Updates (I 998) I, 266-278 © 1998 Harcourt Brace & Co. Ltd

In addition to SbV, pentamidine, and AmB, there are a number of second line drugs available that have shown good activities for specific clinical manifestations of the disease or against certain species. These drugs include paromomycin, nucleoside analogues, and azoles. Paromomyein is an amino- glycoside that has a broad antiparasitic activity, and it has been used as a monotherapy to treat visceral leishmaniasis. Ribosomes of Leishmania may be the target of paro- momycin, '5 although impairment of mitochondrial functions may also be important. 1~ Although effective for visceral disease, paromomycin is not, as effective as antimonials or amphoter icin B. Paromomycin is useful however, as a local therapy together wi th the methylbenzethonium chloride for some forms of non-disseminated cutaneous diseases."
The three best drugs, SbV,, pentamidine and ARAB, are administered parentally but an oral drug would be most useful for outpatients. Leishmania, as all protozoal parasites, is auxotrophic for purines, TM and thus purine acquisition from the host is essential. Several cytotoxic purine analogs have been tested on protozoa and allopurinol has shown activity against certain forms of leishmaniasis. Several anti- fungal azoles, such as ketoconazole and itraconazole, that can be taken orally have also been used in" the treatment of Leishmania infections. 7
The immunological responses associated with suscep- tibility or resistance to Leishmania infections in experimental systems have been extensively studied. T cell responses are critical in determining the ou tcome of infection with the intracellular protozoan Leishmania. In both murine and human infections, healing has been associated with T Helper type 1 response.19.~° Immunostimulatory cytokines have been used in conjunct ion with SbV to treat patients. ~,22 Several studies have used interferon-gamma (IFN- 7) as an adjunct to antimony therapy for visceral leishmaniasis. Systemic IFN- 7 is not effective by riser" and has side effects, but it does increase the efficacy of SbV therapy. Other cytokines have shown a great potential in animal studies, and some are now in clinical trials.
DRUG RESISTANCE
As indicated in the previous section, a number of drugs are available and used to treat Leishmania infections. Chemotherapy is not always effective, however, and this may be due to the acquisition of drug resistance. Drug resistance has emerged as a major obstacle to the control of parasitic diseases? Unfortunately, limited information is available on the biochemical and molecular mechanisnls that contribute to drug resistance in field isolates. Indeed, our knowledge on the mechanisms of resistance to different type of drugs in Leishmania is based mainly on work carried with mutant strains selected in vitro by stepwise increasing drug concen- trations. Moreover, resistance has more often been induced in the flagellated insect form of the parasite, whereas the drug is directed against amastigotes. Nevertheless, our under- standing of resistance mechanisms in Leishmania has greatly evolved in the last years and it should n o w be feasible to test whe the r resistance mechanisms elucidated in labora- tory strains are also found in field isolates.
In this review, we shall look at resistance mechanisms induced in vitro against antileishmanial drugs used clinically. We will also desc r ibe resistance mechanisms found against other classes of agents (e.g. antifolates) that may fred appli- cation in the treatment of leishmaniasis.When possible, we will contrast resistance mechanisms observed in parasite strains where resistance has been induced in vitro wi th resis- tance mechanisms found in field isolates. Finally, we will try to draw parallels or highlight differences wi th resistance mechanisms for the same class of drugs operating in other organisms.
RESISTANCE TO ANTIMONIAL DRUGS
Several antimony-resistant Leishmania isolated from patients have been described, 9,~3 Since SbV-containing drugs are still the treatment of choice against all forms of leish- maniasis, it is likely that resistance will increase and spread more rapidly. Analysis of mutants selected for resistance to SbIII and to the related metal arsenite has pinpointed the crucial role of drug transporters and of trypanothione. -'4 56 Trypanothione is the major reduced thiol of Leishmania cells consisting of a spermidine and a bis-glutathionel conjugate.72 The role of the transporters, including ABC trans- porters, and of enzymes modulating trypanothione meta- bolism in the context of oxyanion metal resistance have recently been reviewed. 27 Similarly to arsenite resistance, resistance to SbV is multifactorial. 27 Novel genes involved in antimony resistance have recently been isolated, 28 and (Haimeur et al, unpublished). These genes do not exhibit sequence similarities with known sequences, and the exact mechanisms by which they confer resistance are still unknown.
Resistance to antimony and the related metal arsenite has been studied in other organisms, as well. 27,~9 Aromatic arsenicals are important drugs for the treatment of compli- cated eases of African trypanosomiasis. Trypanosoma brucei cells selected in vitro for resistance to melamine-based arsenicals showed reduced drug uptake rather than efflux. This transport defect was due to a mutation in a nucleoside transporter by which melaminophenyl arsenicals are taken up2 ° The route of entry of antimony is unknown in Leishmania.
RESISTANCE TO PENTAMIDINE
Pentamidine (1,5-bis-4'-amidinophenoxy pentane) is the best known aromatic diamidine used in the treatment of early stage Gambian African trypanosomiasis. It is also a useful drug against various Leishmania infections refractory to SbV, and is also used as a second-line drug in the treatment and prophylaxis of Pneumocystis carinii pneumonia in AIDS patientsY The cellular target is unknown, although pentamidine causes a competi t ive inhibition of polyamine transport suggesting that diamidines possibly enter into Leisbmania through the parasite polyamine transporter(s). TM
The uptake of pentamidine in Leishmania is carrier- mediated and energy- dependent . '°,323~ It has been reported that pentamidine inhibits the S-adenosyl-L-methionine
© 1998 Harcourt Brace & Co. Ltd Drug Resistance Updates (I 998) I, 266-278 n

decarboxylase 34 but, at least in T. brucei, inhibi t ion of the latter enzyme does not seem to account for the t rypanocidal act ion of the drug? 5
Resistance to pen tamid ine has b e e n induced in several Leisbmania species, b o t h promast igotes and amast igotes? TM
It can be stably main ta ined th roughou t the life cycle of the parasite, 36 and is associated w i th changes in the intracellular concen t ra t ion of arginine, o rn i th ine and putrescine. 33 This may be due to alterations in polyamine carriers bu t pe rhaps also to modula t ion of the activity of key enzymes in the polyamine biosynthesis pathway, such as orn i th ine decarbo- xylase and spermid ine synthetase, n,3s Analysis of penta- midine-resistant mutan t s indicated addit ional defects in the uptake of several substrates ~3 w h i c h may be due to a pertur- ba t ion of lipid cons t i tuents the reby increasing m e m b r a n e permeability. 37 T h e exact con t r ibu t ion of m e m b r a n e defect to pen tamid ine resis tance needs to be assessed, however. Mternatively, this multiplicity in m e m b r a n e defects may be due to several routes of pen tamid ine en t ry in Leisbmania 33
that are likely to also be p resen t in/7. cruzi? ~
Resistance to pen tamid ine has b e e n repor ted in field strains of Leishmania, but the mechan i sm by w h i c h resis tance is achieved is unknown . Similarly, pen tamid ine resis tance also occurs in field isolates of t rypanosomes but the only resis tance mechan i sms yet character ized were der ived from T. brucei strains in w h i c h resis tance was induced in vi t ro? l African t rypanosomes resistant to me laminopheny l arsenicals are of ten cross-resistant to diamidines? 9 This is due to a decrease in uptake of pentami- dine via the muta ted P2 nucleoside transporter . 4° However, analysis of T. brucei cells se lected for pen tamid ine resis tance showed that they accumula ted pen tamid ine similarly to wild-type cells and that the p roper t i es of the P2 t ranspor te r were unchanged . 4' Thus, several mechan i sms can lead to pen tamid ine resis tance in protozoa.
RESISTANCE TO DMFO
DL-c~-Difluoromethylornithine (DFMO) is an irreversible inhibi tor of o rn i tb ine decarboxylase (ODC), the rate limiting enzyme in polyamine biosynthesis. Polyamines are required for cell proliferat ion and differentiation, and for the synthesis of the spermidine-gluta thione conjugate t rypanoth ione . 42-44 DMFO is the drug of choice in late stage infect ion due to Trypanosoma gambiense. 45 It has s h o w n also good activities against several o the r parasitic p ro tozoa including Leishmania provided that they are g rown in media contain- ing low amounts of polyamines. 46"4s It is possible tha t the lack of h igh affinity polyamine t ranspor te r systems may explain the efficacy of DMFO in T. br~lcei. 49 However, sufficient pu t resc ine is taken up by T. brucei to rescue an ode null mutant , s° Moreover, species-specific differences in ODC characteris t ics of 77. gambiense and T. rbodesiense may con t r ibu te to the differential activity of DFMO. 5~
Leishmania donovani cells se lected s tepwise wi th gradually increasing concent ra t ions of DFMO have demon- strated increased ODC activity 5~'s3 w h i c h was due to an increase in the copy n u m b e r of the odc gene. s4'55 In contrast, the level of ODC activity was found to be similar in wild-type and DFMO-selected t rypanosomes. Instead, an impor t an t
increase in the uptake of orni th ine was observed in the resis- tant mutan t s .The excess of orn i th ine was p roposed to com- pe te wi th DFMO for ODC, h e n c e leading to sufficient polyamines. 56,s7 Analysis of arsenite resistant Leishmania has revealed that t rypanoth ione was increased in these mutants. 24.26 Elevated levels of t rypanoth ione were due to amplification of the 7-glutamylcysteine synthetase gsh I gene 2s and to ode t ranscr ipt overexpress ion (Haimeur et al, manu- script submit ted for publ ica t ion) .The level of put resc ine and spermidine was indeed fotmd to be increased in arsenite resistant L. tarentolae mutants, and DFMO was s h o w n to sensitize mutants to metals (Haimeur et al, submitted).
RESISTANCE TO AMPHOTERICIN B
Amphote r i c in B is a polyene antibiotic p roduced by Streptomyces nodosus tha t preferential ly interacts wi th 24-substi tuted sterols such as ergosterol w h i c h is the major m e m b r a n e sterol in Leishmania 5s,~9 and fungal cells. The in teract ion of AraB wi th ergosterol leads to the format ion of t r ansmembrane AmB channels w h i c h enhance m e m b r a n e permeabi l i ty the reby allowing the leakage of cytoplasmic c o m p o n e n t s tha t eventually leads to cell death. Othe r factors, de t e rmined mainly f rom studies pe r fo rmed in fungi, are likely to play a role in the ant imicrobial activity of AmB. These include oxidative damage to the cell, 6° the inhibi t ion of the anion t ranspor t protein, 61 and the inhibi t ion of various ATPases. 62 AmB and its n e w lipid formulat ions are n o w compe t ing wi th pen tava len t an t imony as p r imary therapy for visceral leishmaniasis. Several AmB lipid formulat ions have been tes ted against b o t h exper imenta l 63 and clinical 6<65
infect ions caused by Leishmania donovani .
Clinical cases of AraB resis tance in Leishmania have no t yet b e e n repor ted, bu t strains have b e e n selected for resis tance to AraB in vitro. Analysis of L. donovani cells se lected forAmB resis tance revealed an ergosterol p recurso r ra ther than ergosterol itself as the p r edominan t m e m b r a n e sterol. This may change m e m b r a n e fluidity and permeabi l i ty result ing in decreased AmB uptake and increased efflux in the mutants . 6~ Collateral resis tance to a n u m b e r of unre la ted drugs was observed in t h e A m B mutan ts and could be attrib- utable to al tered permeability. We have also genera ted AmB resistant mutants in Leishmania tarentolae. Two indepen- den t mutants , II64.4 and II512.1, w h i c h exhib i ted 6- and 50-fold increase in resis tance to AraB over the wild-type pa ren t strain were obtained. The resis tance mechan i sms of these mutan t s are current ly be ing studied. As w i th several o the r Leishmania drug-selected mutan t s (Table 1), gene amplification was observed in AraB resistant cells.
Similarly to the Leishmania cells studied, AmB resistant strains of Candida albicans had a reduced ergosterol con- tent, and a h igher prevalence of o ther sterols. 6~ The relation b e t w e e n ergosterol levels and AmB resistance is not absolute, however; resistant strains of Cryptococcus neoformans, wi th normal ergosterol levels and sensitive strains wi th reduced levels have bo th b e e n observed. 68 Clearly, alterations in sterol con ten t are insufficient to account for drug resistance to AraB. Efflux p u m p s may also be involved inAmB resistance in fungi.69 A C albicans mult idrug resistance major facilitator was shown to be involved ill f luconazole resistance7 °'7.
Drug Resistance Updates (I 998) I, 266-278 © 1998 Harcourt Brace & Co. Ltd

Table I Gene overexpression in drug resistant Leishmania mutants due to gene amplification
Drug Gene Function Resistance mechanism
Heavy metals pgpA ABC transporter drug efflux gshl glutathione biosynthesis drug conjugation odc* polyamine biosynthesis drug conjugation orfSbV unknown unknown
DFMO odc polyamine biosynthesis overproduction of target Mycophenolic acid impdh inosine monophosphate overproduction of target
dehydrogenase Vinblastine mdrl ABC transporter drug efflux Tunicamycin nagt N-acetylglucosaminyl-transferase overproduction of target Methotrexate dhfr-ts dihydrofolate reductase overproduction of target
ptr l pteridine reductase by-pass mechanism btl* biopterin transport increased folate synthesis
Purine analogs tor transcription like factors unknown Amphotericin B (in progress)
*gene overexpressed by a mechanism other than gene amplification.
RESISTANCE TO AZOLES
In addit ion to AraB, o the r drugs are target ing sterol bio- synthesis. In particular, azoles such as ketoconazole , i tracona- zole and f luconazole are widely used for the t r ea tmen t of systemic fungal infections. 72 The same drugs have s h o w n good chemothe rapeu t i c activities against several kineto- plast id parasites including Leishmania.73
The activity of azoles, as in yeast and fungi, is due to the inhibi t ion of the L e i s h m a n i a cy tochrome P-450-dependent 14 a lpha-demethylat ion of lanosterol. ~9,74 The activity of
the current ly used azoles is variable against different L e i s h m a n i a species, ranging f rom h igh efficacy on L. m e x i c a n a and L. m a j o r to little or no activity against L. braz i l iens is infections. 7,7s Differences in the sterol bio- synthet ic pa thways and sterol requ i rements among the various L e i s h m a n i a species could explain the intr insic resis tance to azoles of cer ta in species. 75
Resistance to azoles has no t b e e n e laborated in L e i s h m a n i a bu t has b e e n s tudied extensively in the yeast C. albicans. It can be media ted by overexpress ion of efflux pumps , 7°'76 muta t ions in the lanosterol 14a-demethylase gene 77,78 or overexpress ion of the lat ter gene7 9 Modulat ion in
the activity of enzymes involved in sterol metabol i sm may also induce azole resis tance in fungi. Indeed, azole-resistant mutan t s of C a lb icans had m e m b r a n e sterol changes consis- t en t w i t h a muta t ion in the delta 5,6-sterol desaturase gene. 67 The lack of ergosterol in the cytoplasmic m e m b r a n e of f luconazole-resistant strains also impar ted resis tance to AmB. Similarly, ke toconazole decreases drastically the level of ergosterol in Le i shman ia , w h i c h leads to an increased resis tance to AraB. 8°
RESISTANCE TO PURINE ANALOGS
All parasitic p ro tozoa so far s tudied are auxo t roph ic for purines, whereas the mammal ian cells tha t they infect are capable of synthesizing pur ines de novo .The un ique features
of the pur ine salvage systems of Trypanosomat idae make these pa thogen ic hemoflagel lates selectively suscept ible to the cytotoxic effects of several pyrazolopyr imidine analogs of inosine and hypoxan th ine . Several analogs have b e e n used in vitro and in vivo, some of w h i c h showed good anti- L e i s h m a n i a activities, sl Allopurinol has b e e n tes ted in clini- cal trials, bu t has no t b e e n effective, at least against cer ta in forms of L e i s h m a n i a infections. 82 Biochemical and genet ic studies have identified several pur ine salvage enzymes in Le i shmania , including the hypoxan th ine-guan ine phospho- r ibosyltransferase (HGPRT). TM Inhibi tors or substrates of HGPRT are cons idered as serious candidates for the c h e m o t h e r a p y of parasitic diseases. '8 Recent work has indicated that L e i s h m a n i a cells w i thou t HGPRT and with- ou t genes encod ing for o the r impor t an t enzymes of pur ine metabol i sm such as adenine phosphor ibosyl t ransferase (APRT) and adenos ine kinase (AK) were able to grow in def ined m e d i u m conta in ing any pu r ineY This surpris ing result suggests that o the r enzyme(s) may be involved in pur ine salvage, and this may require the inhib i t ion of a num- ber of enzymes for such therapy to be efficacious. One of these enzymes is likely to be xan th ine phosphor ibosy l transferase (XPRT).83 A Toxoplasma g o n d i i mutan t resistant to the pur ine analog 6- th ioxanth ine lacks HPGRT activity showing that this pa thway is no t essential for the salvage of pur ines in b o t h L e i s h m a n i a and Toxoplasma. 84
Resistance to nucleos ide analogs, as w i th several o ther drugs, has been induced in L e i s h m a n i a in vitro. Two sepa- rate muta t ions leading to decreased accumula t ion of pur ines were character ized in L e i s h m a n i a spp.; one involved the t ranspor t of inosine, guanosine, and the i r analogues and the second implicated in the t ranspor t of adenos ine and its ana- logues. 8s-87 Mutants lacking b o t h t ranspor te rs have b e e n ob ta ined by select ing parasites w i th two toxic analogues, suggesting that o ther pur ine t ranspor te rs mus t also be pre- sent in Le i shmania . Mutations in pur ine t ranspor te rs seem also to be responsible for resis tance to pur ine analogues in T r y p a n o s o m a cruz i s~'89 and in Tr i t r i chomonas foetus. 9° Such
© 1998 Harcourt Brace & Co. Ltd Drug Resistance Updates (I 998) I, 266-278 ~ { ~

mutat ions are also impl icated in resis tance to arsenicals in t rypanosomes. T. brucei resistant to melarsen oxide or T. evansi selected for resis tance to Cymelarsan lack the P2 adenos ine t r anspor te r suggesting that resis tance to arseni- cals can be due t o loss of uptake. 3°'9~ Cells lacking the P2 recep to r were also s h o w n to accumula te less diamidines. ~° Trypanosomes resistant to melarsoprol are of ten highly cross-resistant to diamidines such as pen tamid ine 39 and T. equiperdum cells selected for resis tance to diamidines were s h o w n to conta in a P2 pur ine t ranspor te r w i th reduced substrate affinity. 92
Leishmania can also r e spond to pur ine analogues by amplifying specific por t ions of its genome. Leishmania selected for inosine dialdehyde or tuberc id in resis tance had muta t ions in t r anspor te r activity- bu t had also a 55 kb ampli- fied ex t rachromosomal circle. 93 Character izat ion of the gene involved in resis tance and p resen t on the circle indicated that it has sequence similarities w i th a family of t ranscr ip t ion f ac to r s , 94 however, the mechan i sm by w h i c h this con t r ibu tes to resis tance is still unknown . Resistance to mycopheno l i c acid, an inhibi tor of the inosine m o n o p h o s p h a t e dehydro- genase (IMPDH), an enzyme involved in the synthesis of guanine nucleotides, has b e e n induced in Leishmania. Resistance has b e e n associated to the amplif icat ion of the impdh gene as par t of a l inear stable ampl icon? 5 Selection of T. gambiense for mycopheno l i c acid also led to the ampli- f ication of the impdh gene but in this case, the who le c h r o m o s o m e was amplified. 96
RESISTANCE TO ANTIFOLATES
Folates are a class of cofactors tha t serve as one-carbon donors in b iochemica l reactions. There are many folate- d e p e n d e n t enzymes, several of w h i c h are essential for impor t an t steps in DNA synthesis, including biosynthesis of pur ines and convers ion of deoxyur id ine m o n o p h o s p h a t e (dUMP) to deoxythymidine m o n o p h o s p h a t e (dTMP). Antffolates are inhibi tors of the enzyme dihydrofolate reduc- tase (DHFR), w h i c h toge ther wi th thymidylate synthase (TS) catalyze consecut ive react ions in the de novo synthesis of
dTME In protozoa, these two enzymes are l inked in the bifunct ional p ro te in DHFR-TS? 7 The folate pa thway has b e e n the target of therapeut ic in te rven t ion for a large n u m b e r of neoplasic, bacterial and parasitic diseases28 Inhibi tors of d ihydropteroa te synthetase (DHPS) and dihydrofolate reduc- tase (DHFR) have b e e n exploi ted in the clinical t r ea tment of b o t h malaria and toxoplasmosis . 99a°° Convent ional DHPS and DHFR inhibi tors are ineffective against Leishmania, however. Nevertheless, considerable work has b e e n done on the mechan i sms of resis tance to the antifolate me tho t rexa te (MTX) in Leishmania. 98,~°I
The first mechan i sm of antifolate resis tance descr ibed was the overexpress ion of the target enzyme DHFR-TS. 1°2 This overexpress ion was media ted by dhfr-ts gene amplifi- cation. ~°3 Amplif-ication of dhfr-ts following MTX resis tance has only b e e n descr ibed in L. ma jor mutan ts bu t w h e n a preferred resis tance mechan i sm was d isrupted in L. tarentolae, dhfr-ts was amplified u p o n MTX select ion 8 and (Kiindig et al, in prepara t ion) In at least one case, the DHFR overproduced in L. major had a poin t muta t ion TM in a posi t ion k n o w n to correlate w i th MTX resis tance in mammal ian DHFRs. Amplif icat ion of the dhJ~-ts gene is also observed in bacter ia resistant to t r ime thopr im '°5 and in cancer cells resistant to MTX 98,1°6,'°7 (Table 2). Mutat ions in the target gene cons t i tu te also a c o m m o n mechan i sm of resis tance to MTX in cancer cells and the main m e c h a n i s m of py r ime thamine
98 I08 I09 resistance in the malaria parasite Plasmodium . . (Table 2). In addi t ion to dhfr-ts, ano the r locus was s h o w n to be
f requent ly amplified in different species of Leishmania selected for MTX resistance. "°~'2 It co r responds to the H locus, and the mechan i sm by w h i c h it confers resis tance remained elusive for several years.Transfection studies led to the isolation of the gene, and its character izat ion indicated that its gene product , n a m e d PTR1, be longs to the family of shor t chain dehydrogenases5 ~3a14 Gene d is rupt ion studies and PTR1 b iochemica l character izat ion indicated that PTR1 is involved in the salvage of oxidized p ter ins and is also capable of reduc ing folates into dihydrofolates and into tetrahydrofolates.~0~.,09.,5 ,,7 PTR1, therefore, confers resis tance
by by-passing the need for DHFR. To be effective against
Table 2 Antifolate resistance mechanisms
Mechanism Bacteria Cancer cells Protozoa
Gene amplification rare frequent for dhfr
Target (DHFR) moderate moderate-frequent mutation By-pass mechanism frequent, plasmid-encoded unknown
dhfr
Reduced uptake rare reduced expression of the reduced folate carrier
Modification unknown decreased polyglutamyladon
Uptake of precursors unknown unknown
moderate for dhfr-ts
frequent for pt r l
frequent in Plasrnodium
rare in Leishmania
amplification of ptr I
in Leishmania
loss of activity of the folate transporter in Leishmania
unknown
increased biopterin uptake in Leishmania
Drug Resistance Updates (I 998) I, 266-278 © / 998 Harcourt Brace & Co. Ltd

Le i shman ia , antifolate-based c h e m o t h e r a p y would therefore n e e d to target b o t h DHFR-TS and PTR1, and indeed, such an inhibi tor has recent ly b e e n s h o w n to be effective against Le i shman ia . 118 T r y p a n o s o m a c ruz i was recent ly s h o w n to conta in a p t r I homolog and its overexpress ion also leads to MTX resistance. "9 Interestingly, by-pass of the ch romosomal DHFR is also the main res is tance m e c h a n i s m of gram- negative bacter ia to t r imethopr im, w h e r e resis tance is medi- ated by plasmid encoded prote ins tha t have DHFR activities? 8
L e i s h m a n i a have long b e e n t h o u g h t to be auxo t roph ic for folates, and indeed, u n d e r s tandard laboratory condi t ions L e i s h m a n i a rely mainly on uptake of folates for g r owth? A c o m m o n folate/MTX t ranspor te r has b e e n identified in L e i s h m a n i a and muta t ions wi th in this gene lead to various levels of MTX res is tanceY °,~2°-~23 Two different classes of mutan t s have b e e n obse rved in L. tarentolae; the first one s h o w e d a 2- to 5-fold decrease in MTX accumula t ion and the second one a more than 50-fold decrease. TM Since mutan t cells showing no measurable folate up take can proliferate normal ly u n d e r s tandard g rowth condi t ions, L e i s h m a n i a must be able to synthesize folic acid de novo. Consis tent w i t h this proposal , it has b e e n s h o w n that L e i s h m a n i a is able to conver t radioactively labeled b iop te r in into r educed folates, 125 bu t the exact m e c h a n i s m is u n k n o w n a l though it seems to differ f rom previously descr ibed pathways, sa~6
The surpris ing ability of L. tarento lae cells to thr ive in absence of a folate t r anspor te r has recent ly found a plausible explanat ion whi le pe r fo rming funct ional c loning studies. Transfect ion of L. m e x i c a n a genomic cosmid bank into wild- type parasite ceils and select ion for MTX resis tance led to the isolation of a gene encod ing for a m e m b r a n e pro te in (manusc r ip t in preparat ion) . This m e m b r a n e pro te in was identical to the p roduc t of orfG x27 an o p e n reading frame par t of the LD1/CD1 L e i s h m a n i a region that is f requent ly ampli- fied even in absence of selective pressure. '~ This orfG ( r enamed BT1 by the group of S. Beverley) was found to be a b iop te r in transporter . In every L. tarento lae MTX resistant mu tan t in w h i c h there was no measurable t ranspor t of MTX (or folate), the activity of BT1 was upregulated. Therefore cells c o m p e n s a t e d for the lack of folate uptake by increasing the uptake of biopter in , a p recurso r of folic acid. BT1 is par t of a large gene family and one m e m b e r is consis tent ly dele ted in all MTX resis tant mutan t s in w h i c h there is no measurable uptake. Work is in progress to test w h e t h e r the de le ted copy cor responds to the folate t ransporter .
In mammal ian cells, MTX is t r anspor ted into cells by the carr ier system used for the active t ranspor t of r educed folates. MTX t ranspor t def iciency due to decreased express ion of the r educed folate carr ier was obse rved in MTX clinical res i s tance) 29 Several o the r t ranspor t muta t ions can also lead to MTX resis tance in cance r cells? 8
RESISTANCE TO PAROMOMYCIN
The aminoglycoside antibiotic pa romomyc in (aminosidine) is a potential ly useful anti-leishmanial topical agent in humans ~,7 and is also used for t r ea tment of canine leishmaniasis3 3° Its mode of act ion is no t clearly established, bu t it seems to inter- fere w i th the mi tochondr ia l activity of L e i s h m a n i a since it inhibits cell respirat ion and lowers its associated electric
potent ia l difference across mi tochondr ia l membranes . 16 Paromomycin can also interact wi th L e i s h m a n i a r ibosomes to p romote association w i th the subunits3 5 Previous studies in heterologous systems such as E. coli, Te trahymena, and yeast mi tochondr ia l mutants have demonst ra ted that muta- t ions in r ibosomal RNA is a cause of paramomycin resistance. Resistance to pa romomyc in has b e e n induced in L e i s h m a n i a promastigotes. 13~a32 In contras t to the o ther systems, however, no sequence changes in the L e i s h m a n i a r ibosomal RNA gene b e t w e e n sensitive and resistant isolates were observed. 13~ Possible mechan isms of resistance in L e i s h m a n i a could be related to the effect of pa romomycin on RNA synthesis and to modifications of m e m b r a n e polar lipids and m e m b r a n e fluidity, leading to al tered m e m b r a n e permeability.16 The bac- terial neomyc in phosphot ransferase gene, used as a selectable marker in L e i s h m a n i a transfection, confers resistance to paramomycin, m As several groups are trying to generate live a t tenuated parasite strains for vaccine deve lopmen t using gene targeting technologies, it will be impor tan t that recom- b inan t strains do not conta in markers conferr ing resis tance to drugs possibly used in the t rea tment of L e i s h m a n i a infections.
RESISTANCE MEDIATED BY GENE AMPLIFICATION A N D BY TRANSPORT MUTATIONS
Drug resis tance in L e i s h m a n i a is mainly due to t ranspor t muta t ions or to gene amplification. Amplif icat ion of genes has b e e n observed in mutan t s se lected for res is tance to metals, to AmB, to nucleos ide analogs, to antifolates and to DFMO (Table 1). Gene amplif icat ion was also obse rved in L e i s h m a n i a cells se lected for resis tance to o the r drugs that are no t used for t rea tment .Ampl i f ica t ion of a P-glycoprotein homolog gene was observed in several species se lected for vinblast ine resis tance 134-1~7 and the role of this gene in resis tance was recent ly reviewed. '38,139 L e i s h m a n t a cells se lected for resis tance to tun icamycin have the N-acetylglu- cosamine- l -phospha te transferase gene amplified. '4° Several o the r loci are amplified in resistant or wild-type L e i s h m a n i a species bu t the i r exact role in res is tance remains still unclear.141
The relative ease to de tec t gene amplif icat ion in Le i shman ia , probably due to its relatively low genomic complexity, '°2,139 may explain in par t w h y so many amplified genes have b e e n observed in drug resistant mutants . O the r reasons are likely to con t r ibu te to this h igh f requency of gene amplification, however. Indeed, gene amplif icat ion is rare in T. brucei 8 and in the only example repor ted , the who le c h r o m o s o m e is amplif ied? 6 The amplif icat ion mecha- nisms have b e e n studied in some detail in L e i s h m a n i a and seem to involve r ecombina t ion b e t w e e n repea ted homo- logous sequences to yield ex t rachromosomal circular or l inear amplicons . 142'143 The sequence requ i rements for expression, replication, and ma in t enance of extrachro- mosomal sequences in L e i s h m a n i a is no t well unders tood , bu t bacter ial plasmids w i thou t leishmanial DNA can repl icate in Le i sbman ia . TM This is no t the case in T. bruce i 145 or in several o the r p ro tozoa suggesting that in L e i s b m a n i a several regions can be amplified and maintained, p rovided that they confer a sufficient g rowth advantage to the cell.
© 1998 Harcourt Brace & Co. Ltd Drug Resistance Updates (1998) I, 266-278

In t rachromosomal amplif icat ion is rare in Leishmania ~4~ but is f requent in Plasmod ium w h e r e the size of the chromo- somes increasesJ 4~ In Plasmodium, the increase in copy n u m b e r is small ( 2 - 5 - f o l d ) , ~46 whereas the ex t rachromosomal e lements in Leishmania can be in 100 copies, TM w h i c h permi ts h igh levels of resistance. Gene amplif icat ion is also a c o m m o n mechan i sm of gene overexpress ion in resistant cance r cells, and a l though the mechan i sms seem to differ f rom those found in Leishmania there are a n u m b e r of c o m m o n features.*47
An interest ing variat ion for p ro te in overproduc t ion has recent ly b e e n unraveled in Leishmania drug resistant mutants. Indeed, we obse rved that the level of odc and b t l t ranscripts were increased in Leishmania resistant to arsen- ite and MTX, respect ively (manuscr ip ts in preparat ion) . In ne i the r case was the cor responding gene found to be amplif ied.The mechan i sm by w h i c h RNA is overexpressed is no t unders tood , bu t it should be m e n t i o n e d that no pol II p r o m o t e r has yet b e e n character ized in Leishmania.
Considerable regulat ion occurs at the level of matura t ion ~48 and stability ~49,~5° of RNA transcripts , and it is possible that the increase in RNA in these mutan t s is due to one of those mechanisms.
A second major m e c h a n i s m of resis tance in Leishmania
cells se lected for resis tance to drugs in vitro involves trans- por t muta t ions .These muta t ions can theoret ical ly e i ther lead to efflux of drugs outside the cell; to sequest ra t ion th rough intracellular compar tments ; to r educed uptake of the drug; or to increased uptake of substrates or precursors necessary for the synthesis of key metabol ic in termedia tes (Table 3). Several of the specific impor te rs have recent ly b e e n isolated by funct ional c loning and thei r role in resis tance is current ly be ing established.
STAGE-SPECIFIC EFFECTS O N MECHANISMS OF DRUG RESISTANCE IN L E I S H M A N I A
Most studies on resis tance have so far b e e n done w i th promast igotes and this is explainable by the ease of growing
this form of the parasite. Drugs, however, are d i rected against the amast igote stage, and they possibly could act differently on the two life stages since promast igotes and amastigotes clearly differ on a n u m b e r of cr i ter ia .The ability of cul tur ing axenic amastigotes in vitro ~5I has facilitated the analysis of stage-specific ant i le ishmanial effects of the commonly used anti leishmanial drugs, m-~Ss No significant differences were observed in the susceptibil i ty of b o t h Leishmania life stages to ampho te r i c in B and paromomycin , bu t pen tamid ine was more active against the promast igote than the amastigote stage 153 (Table 4). In contrast , SbV drugs were m u c h more active against the amast igote than the promast igote stage a l though differences varied wi th species f rom 1.8- to 300-fold. .53 The b iochemica l basis for the increased suscepti- bility to SbV in the amastigote stage is present ly unknown . It is possible tha t this increased sensitivity may due e i ther to increased an t imony accumula t ion in the amastigote form or to differences in the convers ion of Sb(V) to Sb(II1). Since axenic free living growing amastigotes are as suscep- tible as ' t rue ' amastigotes/s3 however, it is likely tha t SbV susceptibi l i ty is a trait of the amastigote parasite ra ther than macrophage-mediated. Resistance to SbV in clinical isolates might be be t t e r s tudied in the amastigote stage and the few repor ts on the subject indicated that promast igotes analysis did not correlate w i th the clinical p ic ture whereas a good correlat ion was seen w h e n susceptibil i ty was tes ted in the amastigotes; 156 (D. Sacks, NIH, persona l communica t ion) .
CONCLUSIONS
Leishmaniasis remains endemic in the developing world .The occur rence of co-infection wi th HIV and Zeishmania in Southern Europe is steadily increas ing.The recen t ep idemic of kala-azar in Sudan underscores the explosive increase in the inc idence of leishmaniasis in these regions. Since the beg inn ing of this cen tu ry antimonials have b e e n used to treat h u m a n leishmaniasis. Second line drugs such, as pen tamid ine and ampho te r i c in B, are used in cases refrac- tory to SbV treatment . In addi t ion to the l imitations and the
Table 3 Transport mutations in drug resistant Leishmania
Class Drug Gene Mechanism
Active efflux
Decreased uptake
Sequestration
increased uptake
Heavy metals Unknown Vinblastine mdr l AmB unknown
AmB unknown Nucleosides unknown
Methotrexate unknown Pentamidine unknown
Heavy metals pgpA
plasma membrane metal-thiol pump putative efflux pump (non specific?) increased efflux
(non specific?) reduced uptake reduced uptake of guanosine analogs reduced uptake of adenosine analogs decreased folate/methotrexate uptake alteration in polyamine carrier
putative intracellular X-thiol importer
Methotrexate btl increased biopterin uptake
D Drug Resi~ance Updates (I 998) I, 266-2 78 © 1998 Harcourt Brace & Co. Ltd
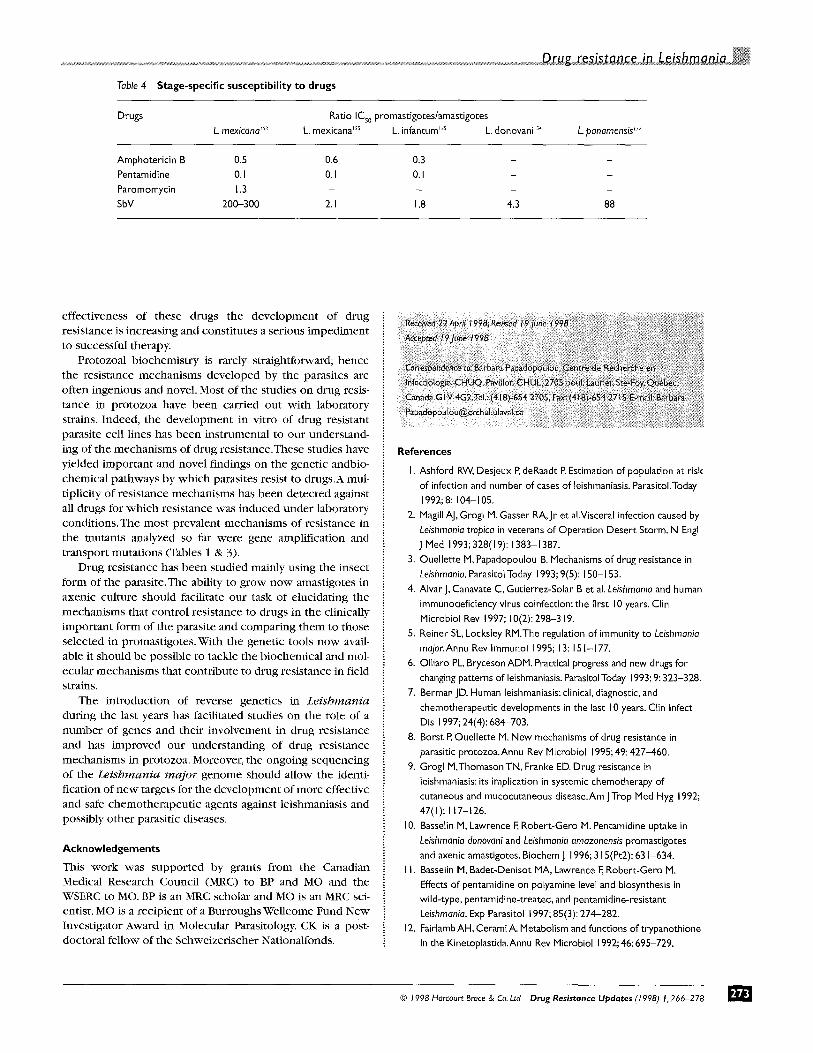
Table 4 Stage-specific susceptibility to drugs
Drugs Ratio ICs0 promastigotes/amastigotes L. mexicana 's3 L. mexicana 'ss L. infantum Lss L. donovani 's4 L. panamensis ~s2
Amphotericin B 0.5 0.6 0.3 - - Pentamidine 0. I 0. I 0. I - - Paromomycin 1.3 . . . . SbV 200-300 2. I 1,8 4.3 88
effect iveness of these drugs the deve l opm en t of drug res is tance is increasing and const i tu tes a serious i m p e d i m e n t to successful therapy.
Protozoal b iochemis t ry is rarely straightforward; h e n c e the resis tance mechan i sms deve loped by the parasites are of ten ingenious and novel. Most of the studies on drug resis- tance in p ro tozoa have b e e n carr ied out w i th laboratory strains. Indeed, the deve l opm en t in vitro of drug resistant parasi te cell lines has b e e n ins t rumenta l to our unders tand- ing of the mechan i sms of drug res is tance.These studies have yielded impor t an t and novel findings on the genet ic andbio- chemical pa thways by w h i c h parasites resist to drugs.A mul- tiplicity of resis tance mechan i sms has b e e n de tec ted against all drugs for w h i c h res is tance was induced u n d e r laboratory condit ions. The mos t prevalent mechan i sms of resis tance in the mutan t s analyzed so far were gene amplif icat ion and t ranspor t muta t ions (Tables 1 & 3).
Drug resis tance has b e e n s tudied mainly using the insect form of the parasi te .The ability to grow n o w amastigotes in axenic cul ture should facilitate our task of elucidat ing the mechan i sms that cont ro l resis tance to drugs in the clinically impor t an t form of the parasi te and compar ing t h e m to those selected in promastigotes. Wi th the genet ic tools n o w avail- able it should be possible to tacMe the b iochemica l and mol- ecular mechan i sms that con t r ibu te to drug resis tance in field strains.
The in t roduc t ion of reverse genet ics in Leishmania dur ing the last years has facilitated studies on the role of a n u m b e r of genes and the i r invo lvement in drug resis tance and has improved our unders t and ing of drug resis tance mechan i sms in protozoa. Moreover, the ongoing sequenc ing of the Leishmania major genome should allow the identi- f ication of n e w targets for the d e v e l o p m e n t of more effective and safe chemothe rapeu t i c agents against leishmaniasis and possibly o the r parasitic diseases.
Acknowledgements
This work was suppo r t ed by grants f rom the Canadian Medical Research Counci l (MRC) to BP and MO and the WSERC to MO. BP is an MRC scholar and MO is an MRC sci- entist. MO is a rec ip ien t of a Burroughs WeUcome Fund New Invest igator Award in Molecular Parasitology. CK is a post- doctoral fel low of the Schweizer ischer Nationalfonds.
References
I. Ashford RW, Desjeux R deRaadt R Estimation of population at risk of infection and number of cases of leishmaniasis. ParasitoI.Today 1992; 8:104-105.
2. Magill AJ, Grogl M, Gasser RA, Jr et aI.Visceral infection caused by Leishmania tropica in veterans of Operation Desert Storm. N Engl J Med 1993;328(19): 1383-1387.
3. Ouellette M, Papadopoulou B. Mechanisms of drug resistance in Leishmania. Parasitol Today 1993; 9(5): 150-153.
4. AIvar J, Canavate C, Gutierrez-Solar Be t al. Leishmania and human immunodeficiency virus coinfection:the first 10 years. Clin Microbiol Rev 1997; 10(2): 298-319.
5. Reiner SL, Locksley RM.The regulation of immunity to Leishmania
major.Annu Rev Immunol 1995; 13: 151-177. 6. Olliaro PL, Bryceson ADM. Practical progress and new drugs for
changing patterns of leishmaniasis. Parasitol Today 1993; 9: 323-328. 7. Berman JD. Human leishmaniasis: clinical, diagnostic, and
chemotherapeutic developments in the last 10 years. Clin Infect Dis 1997; 24(4): 684-703.
8. Borst P, C)uellette M. New mechanisms of drug resistance in parasitic protozoa.Annu Rev Microbiol 1995;49:427-460.
9. Grogl M,ThomasonTN, Franke ED. Drug resistance in leishmaniasis: its implication in systemic chemotherapy of cutaneous and mucocutaneous disease.Am J Trop Med Hyg 1992; 47(I): 117-126.
I 0. Bassdin M, Lawrence F, Robert-Gero M. Pentamidine uptake in Leishmania donovani and Leishmania amazonensis promastigotes and axenic amastigotes. Biochem J 1996; 315(Pt2): 631-634.
I I. Basselin M, Badet-Denisot MA, Lawrence F, Robert-Gero M. Effects of pentamidine on polyamine level and biosynthesis in wild-type, pentamidine-treated, and pentamidine-resistant Leishmania. Exp Parasitol 1997; 85(3): 274-282.
12. Fairlamb AH, Cerami A. Metabolism and Functions of trypanothione in the Kinetoplastida.Annu Rev Microbiol 1992; 46: 695-729.
© 1998 Harcourt Brace & Co. Ltd Drug Resistance Updates (I 998) I, 266-278

13. Mishra M, Biswas UK,JhaAM, KhanAB.Amphotericin versus sodium stibogruconate in first-line treatment of Indian Kata-azar. Lancet 1994; 344(8937): 1599-1600.
14. Berman JD. Chemotherapy for leishmaniasis: biochemical mechanisms, clinical efficacy, and future strategies. Rev Infect Dis 1988; 10(3): 560-586.
15. Maarouf M, Lawrence F, Croft SL, Robert-Gero M. Ribosomes of Leishmania are a target for the aminoglycosides. Parasitol Res 1995; 81 (5): 421-425.
16. Maarouf M, de KouchkovskyY, Brown Set al. In vivo interference of paromomycin with mitochondrial activity of Leishmania. Exp Cell Res 1997; 232(2): 339-348.
17. Soto J, Fuya R Herrera R, Berman J.Topical paromomycin/methylbenzethonium chloride plus parenteral meglumine antimonate as treatment forAmerican cutaneous leishmaniasis: controlled study. Clin Infect Dis 1998; 26(I): 56-58.
18. Ullman B, Carter D: Molecular and biochemical studies on the hypoxanthine-guanine phosphoribosyltransferases of the pathogenic haemoflagellates. Int J Parasitol 1997; 27(2): 203-213.
19. Reed SG, Scott RT-cell and cytokine responses in leishmaniasis. Curr Opin Immunol 1993; 5(4): 524-53 I.
20. Locksley RM,Wakil AE, Corry DB et aI.The development of effector T cell subsets in murine Leishmania major infection. Ciba Found Syrup 1995; 195: I 10-7; discussion I 17-122.
21. Badaro R, Falcoff E, Badaro FS et aI.Treatment of visceral leishmaniasis with pentavalent antimony and interferon gamma [see comments]. N Engl J Med 1990; 322(I): 16-21.
22. Sundar S, Murray HW. Effect of treatment with interferon-gamma alone in visceral leishmaniasis.J Infect Dis 1995; 172(6): 1627-1629.
23. Jackson JE,Tally JD, Ellis WY et al. Quantitative in vitro drug potency and drug susceptibility evaluation of Leishmania ssp. from patients unresponsive to pentavalent antimony therapy.Am J Trop Med Hyg 1990; 43(5): 464-480.
24. Mukhopadhyay R, Dey S, Xu N e t aI.Trypanothione overproduction and resistance to antimonials and arsenicals in Leishmania. Proc Natl Acad Sci U S A 1996; 93(I 9): 10383-10387.
25. Grondin K, HaimeurA, Mukhopadhyay R et al. Co-amplification of the gamma-glutamylcysteine synthetase gene gshl and of the ABC transporter gene pgpA in arsenite-resistant Leishmania tarentolae. EmboJ 1997; 16(I I): 3057-3065.
26. Legare D, Papadopoulou B, Roy Ge t al. Efflux systems and increased trypanothione levels in arsenite-resistant Leishmania. Exp Parasitol 1997; 87(3): 275-282.
27. Ouellette M, L6gar6 D, HaimeurA et aI.ABC transporters in Leishmania and their role in drug resistance, Drug Res. Updates 1998; I : 43-48.
28. Haimeur A, Ouellette M. Gene amplification in Leishmania tarentolae selected for resistance to sodium stibogluconate. Antimicrob.Agents Chemother. 1998; in press.
29. Rosen BR Resistance mechanisms to arsenicals and antimonials. J Basic Clin Physiol Pharmacol 1995; 6(3-4): 251-263.
30. Carter NS, Fairlamb AH.Arsenical-resistant trypanosomes lack an unusual adenosine transporter [published erratum appears in Nature 1993 Jan 28; 361 (6410): 374]. Nature 1993; 361 (6408): 173-176.
3 I. Ouellette M, P6pin J. Pentamidine.Williams and Wilkins, 1998. (Yu VL, ed.Antimicrobial Chemotherapy; vol in press).
32. Calonge M,Johnson R, Balana-Fouce R, Ordonez D. Effects of cationic diamidines on polyamine content and uptake on
Leishmania infantum in in vitro cultures. Biochem Pharmacol 1996; 52(6): 835-84 I.
33. Basselin M, Lawrence F, Robert-Gero M.Altered transport properties of pentamidine-resistant Leishmania donovani and L. amazonensis promastigotes. Parasitol Res 1997; 83(5): 413-418.
34. Bitonti AJ, Dumont JA, McCann PR Characterization of Trypanosoma brucei brucei S-adenosyI-L-methionine decarboxylase and its inhibition by Berenil, pentamidine and methylglyoxal bis(guanylhydrazone). Biochem J 1986; 237(3): 685-689.
35. Berger BJ, Carter NS, FairlambAH. Po~yamine and pentamidine metabolism in African trypanosomes.ActaTrop 1993; 54(3-4): 215-224.
36. Sereno D, Lemesre JL.Axenically cultured amastigote forms as an in vitro model for investigation of antileishmanial agents. Antimicrob Agents Chemother 1997; 41 (5): 972-976.
37. Basselin M, Robert-Gero M.Alterations in membrane fluidity, lipid metabolism, mitochondrial activity, and lipophosphoglycan expression in pentamidine-resistant Leishmania. Parasitol Res 1998; 84(I): 78-83.
38. Le Quesne SA, Fairlamb AH. Regulation of a high-affinity diamine transport system in Trypanosoma cruzi epimastigotes. Biochem J 1996; 316(Pt 2): 481-486.
39. Bacchi CJ,Yarlett N. Effects of antagonists of polyamine metabolism on African trypanosomes.ActaTrop 1993; 54(3-4): 225-236.
40. Carter NS, Berger BJ, FairlambAH. Uptake of diamidine drugs by the P2 nucleoside transporter in melarsen- sensitive and - resistant Trypanosoma bruceibrucei.J Biol Chem 1995; 270(47): 28153-28157.
4 I. Berger BJ, Carter NS, Fairlamb AH. Characterisation of pentamidine-resistant Trypanosoma brucei brucei. Mol Biochem Parasitol 1995; 69(2): 289-298.
42. Tabor CW, Tabor H. Polyamines.Annu Rev Biochem 1984; 53: 749-790.
43. FairlambAH, Carter NS, Cunningham M, Smith K. Characterisation of melarsen-resistant Trypanosoma brucei brucei with respect to cross-resistance to other drugs and trypanothione metabolism. Mol Biochem Parasitol 1992; 53(I-2): 213-222.
44. McCann PP, PeggAE. Ornithine decarboxylase as an enzyme target for therapy. Pharmacol Ther 1992; 54(2): 195-215.
45. Ouellette M, P6pin J. Eflornithine. NewYork:Williams and Wilkins, 1998. (Yu VL, ed.Antimicrobial Chemotherapy; vol in press).
46. Kaur K, Iovannisci DM, UIIman B.Adenine phosphoribosyltransferase-deficient Leishmania donovanL Adv Exp Med Biol 1986; 195 Pt B:553-557.
47. Bitonti AJ, McCann PP, SjoerdsmaA. Plasmodium falciparum and Plasmodium berghei: effects of ornithine decarboxylase inhibitors on erythrocytic schizogony. Exp Parasitol 1987; 64(2): 237-243.
48. Mukhopadhyay R, Madhubala R. Effect of antioxidants on the growth and polyamine levels of Leishmania donovani. Biochem Pharmacol 1994; 47(4): 61 I--615.
49. Fairlamb AH, SA LQ. Polyamine metabolism in Trypanosomes. In: Hide G, Mottram JC, Coombs GH, PH H, eds. CAB International 1997.
50. Li F, Hua SB,Wang CC, Gottesdiener KM. Procyclic Trypanosoma brucei cell lines deficient in ornithine decarboxylase activity. Mol Biochem Parasitol 1996; 78(I-2): 227-236.
5 I. Iten M, Mett H, EvansA et aI.Alterations in ornithine decarboxyiase characteristics account for tolerance of
Drug Resistance Updates (I 998) I, 266-278 © 1998 Harcourt Brace & Ca. Ltd

Trypanosoma brucei rhodesiense to D, L- alpha- difluoromethylornithine.Antimicrob Agents Chemother 1997; 41 (9): 1922-1925.
52. Coons T, Hanson S, Bitonti AJ et aI.Alpha-difluoromethylornithine resistance in Leishmania donovani is associated with increased ornithine decarboxylase activity. Mol Biochem Parasitol 1990; 39(I): 77-89.
53. Mukhopadhyay R, Kapoor P, Madhubala R. Characterization of alpha-difluoromethylornithine resistant Leishmania donovani and its susceptibility to other inhibitors of the polyamine biosynthetic pathway. Pharmacol Res 1996; 34(I-2): 43-46.
54. Hanson S, Beverley SM,Wagner W, UIIman B. Unstable amplification of two extrachromosomal elements in alpha- difluoromethylornithine-resistant Leishmania donovani. Mol Cell Biol 1992; 12(12): 5499-5507.
55. Sanchez CP, Mucci J, Gonzalez NS et al.Alpha- difluoromethylornithine-resistant cell lines obtained after one- step selection of Leishmania mexicana promastigote cultures. Biochem J 1997; 324(Pt 3): 847-853.
56. Phillips MA, Coffino P, Wang CC. Cloning and sequencing of the ornithine decarboxylase gene from Trypanosoma brucei.
Implications for enzyme turnover and selective difluoromethylornithine inhibition.J Biol Chem 1987; 262(I 8): 8721-8727.
57. BellofattoV, Fairlamb AH, Henderson GB, Cross GA. Biochemical changes associated with alpha-difluoromethylornithine uptake and resistance in Trypanosoma brucei. Mol Biochem Parasitol 1987; 25(3): 227-238.
58. Berman JD, Goad LJ, Beach DH, Hoiz GG,Jr. Effects of ketoconazole on sterol biosynthesis by Leishmania mexicana mexicana amastigotes in murine macrophage tumor cells. Mol Biochem Parasitol 1986; 20(I): 85-92.
59. Hart DT, LauwersWJ,Willemsens G et al. Perturbation of sterol biosynthesis by itraconazole and ketoconazole in Leishmania
mexicana mexicana infected macrophages. Mol Biochem Parasitol 1989; 33(2): 123-134.
60. SokoI-Anderson ML, Brajtburg J, Medoff G.Amphotericin B-induced oxidative damage and killing of Candida albicans.J Infect Dis 1986; 154(I): 76-83.
61. Abu-Salah KM, Sedrani SH,TobiaAS, Gambo HA. Influence of amphotericin B on the transport of phosphate, sulphate and potassium ions across the human erythrocyte membrane.Acta Haematol 1988; 79(2): 77-80.
62. Vertut-Doi A, Hannaert R Bolard J.The polyene antibiotic amphotericin B inhibits the Na+/K+ pump of human erythrocytes. Biochem Biophys Res Commun 1988;157(2): 692-697.
63. Vertut-Doi A, Ishiwata H, Miyajima K. Binding and uptake of liposomes containing a poly(ethylene glycol) derivative of cholesterol (stealth 1iposomes) by the macrophage cell line J774: influence of PEG content and its molecular weight. Biochim Biophys Acta 1996; 1278(I): 19-28.
64. Dietze R, Fagundes SM, Brito EF et aI.Treatment of kala-azar in Brazil with Amphocil (amphotericin B cholesterol dispersion) for 5 days.Trans R SocTrop Med Hyg 1995; 89(3): 309-311.
65. Thakur CP, PandeyAK, Sinha GP et al. Comparison of three treatment regimens with liposomal amphotericin B (AmBisome) for visceral leishmaniasis in India: a randomized dose-finding study. Trans R SocTrop Med Hyg 1996; 90(3): 319-322.
66. Mbongo N, Loiseau PM, Billion MA, Robert-Gero M. Mechanism of amphotericin B resistance in Leishmania donovani
promastigotes.Antimicrob Agents Chemother 1998; 42(2): 352-357.
67. Nolte FS, ParkinsonT, Falconer DJ et al. Isolation and characterization of fluconazole- and amphotericin B- resistant Candida albicans from blood of two patients with leukemia. Antimicrob Agents Chemother 1997; 41 (I): 196-199.
68. Warnock DW.Amphotericin B: an introduction.J Antimicrob Chemother 199 I; 28 Suppl B: 27-38.
69. Joseph-HorneT, Hollomon D, Loeffler RS, Kelly SL. Cross- resistance to polyene and azole drugs in Cryptococcus neoformans. Antimicrob Agents Chemother 1995; 39(7): 1526-1529.
70. Sanglard D, Kuchler K, Ischer F et al. Mechanisms of resistance to azole antifungal agents in Candida albicans isolates from AIDS patients involve specific multidrug transporters.Andmicrob Agents Chemother 1995; 39(I I): 2378-2386.
71. Ouellette M, KL~ndig C. Microbial multidrug resistance. Internat J Antimicrob Agents 1997; 8:179-187.
72. Hay RJ.Antifungal therapy and the new azole compounds. J Antimicrob Chemother 199 I; 28 Suppl A: 35-46.
73. Urbina JA. Lipid biosynthesis pathways as chemotherapeutic targets in kinetoplastid parasites. Parasitology 1997; 114 Suppl: $91-99.
74. Beach DH, Goad LJ, Holz GG;Jr. Effects of antimycotic azoles on growth and sterol biosynthesis of Leishmania promastigotes. Mol Biochem Parasitol 1988; 31 (2): 149-162.
75. Rangel H, Dagger F, HernandezA et al. Naturally azole-resistant Leishmania braziliensis promastigotes are rendered susceptible in the presence of terbinafine: comparative study with azole- susceptible Leishmania mexicana promastigotes [published erratum appears in Antimicrob Agents Chemother 1997 Feb; 41 (2): 496].Antimicrob Agents Chemother 1996; 40(I 2): 2785-279 I.
76. White TC. Increased mRNA levels of ERG 16, CDR, and MDRI correlate with increases in azole resistance in Candida albicans isolates from a patient infected with human immunodeflciency virus.Antimicrob Agents Chemother 1997; 41 (7): 1482-1487.
77. WhiteTC.The presence of an R467K amino acid substitution and loss of allelic variation correlate with an azole-resistant lanosterol 14alpha demethylase in Candida albicans.Antimicrob Agents Chemother 1997; 41 (7): 1488-1494.
78. Sanglard D, Ischer F, Koymans L, Bille J.Amino acid substitutions in the cytochrome P-450 lanosterol 14alpha- demethylase (CYP51A I ) from azole-resistant Candida albicans clinical isolates contribute to resistance to azole antifungal agents [In Process Citation].Antimicrob Agents Chemother 1998; 42(2): 241-253.
79. Vanden Bossche H, Marichal P, Odds FC. Molecular mechanisms of drug resistance in fungi.Trends Microbiol 1994; 2( 10): 393-400.
80. Ramos H, Saint-Pierre-Chazalet M, Bolard J, Cohen BE. Effect of ketoconazole on lethal action of amphotericin B on Leishmania mexicana promastigotes.Antimicrob Agents Chemother 1994; 38(5): 1079-10784.
81. Marr JJ. Purine analogs as chemotherapeutic agents in leishmaniasis and American trypanosomiasis.J Lab Clin Meal 1991; 118(2):11 I - I 19.
82. Velez I,Agudelo S, Hendrickx E et al. Inefficacy of allopurinol as monotherapy for Colombian cutaneous leishmaniasis.A randomized, controlled trial.Ann Intern Meal 1997; 126(3): 232-236.
83. Hwang HY, UIIman B. Genetic analysis of purine metabolism in Leishmania donovani. J Biol Chem 1997; 272(31 ): 19488-194896.
© 1998 Harcourt Brace & Co. Ltd Drug Resistance Updates (I 998) I, 266-278

84. Pfefferkorn ER, Borotz SE. Toxoplasma gondii: characterization of a mutant resistant to 6-thioxanthine. Exp Parasitol 1994; 79(3): 374-382.
85. Iovannisci DM, Kaur K,Young L, UIIman B. Genetic analysis of nucleoside transport in Leishmania donovani. Mol Cell Biol 1984; 4(6): 1013-1019.
86. Rainey P, Santi DV. Formycin B resistance in Leishmania. Biochem Pharmacol 1984; 33(8): 1374-1374.
87. Aronow B, Kaur K, McCartan K, Ullman B.Two high affinity nucleoside transporters in Leishmania donovani. Mol Biochem Parasitol 1987; 22(I): 29-37.
88. Finley RW, Cooney DA, Dvorak JA. Nucleoside uptake in Trypanosoma cruzi: analysis of a mutant resistant to tubercidin. Mol Biochem Parasitol 1988; 31(2): 133-140.
89. NozakiT, Dvorak JA. Molecular biology studies of tubercidin resistance in Trypanosoma cruzi. Parasitol Res 1993; 79(6): 451-455.
90. Hedstrom L,Wang CC. Purine base transport in wild-type and mycophenolic acid-resistant Tritrichomonas foetus. Mol Biochem Parasitol 1989; 35(3): 219-227.
91. Ross CA, Barns AM.Alteration to one of three adenosine transporters is associated with resistance to cymelarsan in Trypanosoma evansi. Parasitol Res 1996; 82(2): 183-188.
92. Barrett MR Zhang ZQ, Denise He t aI.A diamidine-resistant Trypanosoma equiperdum clone contains a P2 purine transporter with reduced substrate affinity. Mol Biochem Parasitol 1995; 73(I-2): 223-229.
93. Kerby BR, Detke S. Reduced purine accumulation is encoded on an amplified DNA in Leishmania mexicana amazonensis resistant to toxic nucleosides. Mol Biochem Parasitol 1993; 60(2): 17 I - I 85.
94. Detke S. Identification of a transcription factor like protein at the TOR locus in Leishmania mexicana amazonensis. Mol Biochem Parasitol 1997; 90(2): 505-51 I.
95. Wilson K, Beverley SM, UIIman B. Stable amplification of a linear extrachromosomal DNA in mycophenolic acid-resistant Leishmania donovani. Mol Biochem Parasito[ 1992; 55(I-2): 197-206.
96. Wilson K, Berens RL, Sifri CD, Ullman B.Amplification of the inosinate dehydrogenase gene in Trypanosoma brucei gambiense due to an increase in chromosome copy number.J Biol Chem 1994; 269(46): 28979-28987.
97. Ferone R, Roland S. Dihydrofolate reductase: thymidylate synthase, a bifunctional polypeptide from Crithidia fasciculata. Proc Natl Acad Sci U S A 1980; 77( I 0): 5802-5806.
98. Ouellette M, Leblanc I~, KCindig C, Papadopoulou B.Antifolate resistance mechanisms from bacteria to cancer cells with emphasis on parasites. In: Rosen BR Mobashery S, eds. Resolving the antibiotic paradox. NewYork: Plenum Publishing Corporation, 1998: in press.
99. Campbell CC, Collins WE, Milhous WK et aI.Adaptation of the Indochina I/CDC strain of Plasmodium falcipamm to the squirrel monkey (Saimiri sciureus).Am J Trop Med Hyg 1986; 35(3): 472-475.
100. Nzila-MoundaA, Mberu EK, Sibley CH et al. Kenyan Plasmodium falciparum field isolates: correlation between pyrimethamine and chlorcycloguanil activity in vitro and point mutations in the dihydrofolate reductase domain.Antimicrob Agents Chemother 1998;42(l): 164-169.
101. Nare B, Hardy LW, Beverley SM.The roles of pteridine reductase I and dihydrofolate reductase- thymidylate synthase in pteridine
metabolism in the protozoan parasite Leishmania major.J Biol Chem 1997; 272(2 I): 13883-1389 I.
102. Coderre JA, Beverley SM, Schimke RT, Santi DV. Overproduction of a bifunctional thymidylate synthetase-dihydrofolate reductase and DNA amplification in methotrexate-resistant Leishmania tropica. Proc Natl Acad Sci U S A 1983; 80(8): 2132-2136.
103. Beverley SM, Coderre JA, Santi DV, Schimke RT. Unstable DNA amplifications in methotrexate-resistant Leishmania consist of extrachromosomal circles which relocalize during stabilization. Cell 1984; 38(2): 431-439.
104. Arrebola R, Olmo A, Reche Pet al. Isolation and characherization of a mutant dihydrofolate reductase- thymidylate synthase from methotrexate-resistant Leishmania cells.J Biol Chem 1994; 269(I 4): 10590-10596.
105. Huovinen P, Sundstrom L, Swedberg G, Skold O.Trimethoprim and sulfonamide resistance.Antimicrob Agents Chemother 1995; 39(2): 279-289.
106. Bertino JR. Karnofsky memorial lecture. Ode to methotrexate. J Clin Oncol 1993; I l(I):5-14.
107. Huennekens FM. In search of dihydrofolate reductase. Protein Sci 1996; 5 (6): 120 I - 1208.
108. Cowman AF, Morry MJ, Biggs BA, Cross GA, Foote SJ.Amino acid changes linked to pyrimethamine resistance in the dihydrofolate reductase-thymidylate synthase gene of Plasmodium falciparum. Proc Natl Acad Sci U S A 1988; 85(23): 9109- I 13.
109. Wang J, Leblanc E, Chang CF et al. Pterin and folate reduction by the Leishmania tarentolae H locus short- chain dehydrogenase/ reductase PTRI.Arch Biochem Biophys 1997; 342(2): 197-202.
110. EllenbergerTE, Beverley SM. Reductions in methotrexate and folate influx in methotrexate-resistant lines of Leishmania major are independent of R or H region amplification.J Biol Chem 1987; 262(28): 1350 I - I 3506.
I. Papadopoulou B, Roy G, Ouellette M. Frequent amplification of a short chain dehydrogenase gene as part of circular and linear amplicons in methotrexate resistant Leishmania. Nucleic Acids Res 1993; 21 (I 8): 4305-4312.
I 12. Olmo A, Arrebola R, BernierV et al. Co-existence of circular and multiple linear amplicons in methotrexate- resistant Leishmania. Nucleic Acids Res 1995; 23( 15): 2856-2864.
I 13. Papadopoulou B, Roy G, Ouellette M.A novel antifolate resistance gene on the amplified H circle ofLeishmania. EmboJ 1992; I I(10): 3601-3608.
114. Callahan HL, Beverley SM.A member of the aldoketo reductase family confers methotrexate resistance in Leishmania.J Biol Chem 1992; 267(34): 24165-24168.
I 15. Papadopoulou B, Roy G, Mourad W e t al. Changes in folate and pterin metabolism after disruption of the Leishmania H locus short chain dehydrogenase gene. J Biol Chem 1994; 269(I 0): 7310-7315.
I 16. Bello AR, Nare B, Freedman D et al. PTRI: a reductase mediating salvage of oxidized pteridines and methotrexate resistance in the protozoan parasite Leishmania major. Proc Natl Acad Sci U S A 1994; 91 (24): I 1442- I 1446.
I 17. Leblanc E, Papadopoulou B, Bernatchez C, Ouellette M. Residues involved in co-factor and substrate binding of the short-chain dehydrogenase/reductase PTRI producing methotrexate resistance in Leishmania. Eur J Biochem 1998; 251 (3): 768-774.
118. Hardy LW, Matthews W, Nare B, Beverley SM. Biochemical and Genetic Tests for Inhibitors of Leishmania Pteridine Pathways. Exp Parasitol 1997; 87(3): 158-170.
Drug Resistance Updates (I 998) I, 266-278 © 1998 Harcourt Brace & Co. Ltd

I 19. Robello C, Navarro R Castanys S, Gamarro F.A pteridine reductase gene ptrl contiguous to a P-glycoprotein confers resistance to antifolates in Trypanosama cruzi. Mol Biochem Parasitol 1997; 90(2): 525-535.
120. Dewes H, Ostergaard HL, Simpson L. Impaired drug uptake in methotrexate resistant Crithidia fasciculata without changes in dihydrofolate reductase activity or gene amplification. Mol Biochem Parasitol 1986; 19(2): 149-16 I.
12 I. Kaur K, Coons T, Emmett K, UIIman B. Methotrexate-resistant Leishmania donovani genetically deficient in the folate- methotrexate transporter. J Biol Chem 1988; 263(15): 7020-7028.
122. Papadopoulou B, Ouellette M. Frequent amplification of a short chain dehydrogenase gene in methotrexate resistant Leishmania. Adv Exp Meal Biol 1993; 338: 559-562•
123. Gamarro F, Chiquero MJ,Amador MV et al. P-glycoprotein overexpression in methotrexate-resistant Leishmania tropica. Biochem Pharmacol 1994;47(I I): 1939-1947.
124. Ouellette M, Papadopoulou B, HaimeurA et aI.Transport of antifolates and antimonials in drug-resistant Leishmania. In: Georgopapadakou N, ed. Drug Transport in Antimicrobial and anticancer chemotherapy. NewYork: Marcel Dekker, Inc, 1995: 377-402.
125. Beck JT, UIIman B. Biopterin conversion to reduced folates by Leishmania donovani promastigotes. Mol Biochem Parasitol 199 I; 49(I): 21-28.
126. Nare B, Luba J, Hardy LW, Beverley S. New approaches to Leishmania chemotherapy: pteridine reductase I (PTRI) as a target and modulator of antifolate sensitivity. Parasitology 1997; 114 Suppl: SI01-110.
127. Myler PJ, Lodes MJ, Merlin Ge t aI.An amplified DNA element in Leishmania encodes potential integral membrane and nucleotide- binding proteins. Mol Biochem Parasitol 1994; 66(I):11-20.
128. Segovia M, Ortiz G. LDI amplifications in Leishmania. Parasitol Today 1997; 13: 342-8.
129. Gorlick R, Goker E,TrippettT et al. Defective transport is a common mechanism of acquired methotrexate resistance in acute lymphocytic leukemia and is associated with decreased reduced folate carrier expression. Blood 1997; 89(3): I 013- I 018.
130. Poll A, Sozzi S, Guidi G et al. Comparison of aminosidine (paromomycin) and sodium stibogluconate for treatment of canine leishmaniasis.Vet Parasitol 1997; 71 (4): 263-27 I.
13 I. Fang D, Chan MM, Rodriguez R et al. Paromomycin resistance in Leishmania tropica: lack of correlation with mutation in the small subunit ribosomal RNA gene.Am J Trap Med Hyg 1994; 51 (6): 758-766.
132. el-On J, Sulitzeanu A, Schnur LF. Leishmania major: resistance of promastigotes to paromomycin, and susceptibility of amastigotes to paromomycin-methylbenzethonium chloride ointment.Ann Trap Med Parasitol 199 I; 85(3): 323-328.
133. Gueiros-Filho FJ, Beverley SM. On the introduction of genetically modified Leishmania outside the laboratory. Exp Parasitol 1994; 78(4): 425-428.
134. Henderson DM, Sifri CD, Rodgers Met al. Multidrug resistance in Leishmania donovani is conferred by amplification of a gene homologous to the mammalian mdrl gene. Mol Cell Biol 1992; 12(6): 2855-2865.
135. Chow LM,WongAK, UIIman B,Wirth DF. Cloning and functional analysis of an extrachromosomally amplified multidrug resistance- like gene in Leishmania enriettii. Mol Biochem Parasitol 1993; 60(2): 195-208.
136. Gueiros-Filho FJ,Viola JR Games FC et al. Leishmania amazonensis: Multidrug resistance in vinblastine-resistant promastigotes is associated with rhodamine 123 efflux, DNA amplification, and RNA overexpression of a Leishmania mdrl gene. Exp Parasitol 1995; 81 (4): 480--490.
137. Chiquero MJ, Perez-Victoria JM, O'Valle F et aI.Altered drug membrane permeability in a multidrug-resistant Leishmania tropica line. Biochem Pharmacol 1998; 55(2): 13 I - I 39.
138. Ullman B. Multidrug resistance and P-glycoproteins in parasitic protozoa. J Bioenerg Biomembr 1995; 27(I): 77-84.
139. Ouellette M, HaimeurA, Grondin K et aI.Amplification of the ABC transporter gene pgpA and of other metal resistance genes in Leishmania tarentolae and their study by gene transfection and gene disruption. In:Ambudkar SV, Gottesman MM, eds. Methods Enzymol. San Diego:Academic Press, 1998: in press, vol ABC Transporters: Biochemical, cellular, and molecular aspects).
140. Liu X, Chang KRThe 63-kilobase circular amplicon of tunicamycin-resistant Leishmania amazonensis contains a functional N-acetylglucosamine-I-phosphate transferase gene that can be used as a dominant selectable marker in transfection. Mol Cell Biol 1992; 12(9): 4112-4 122.
141. Beverley SM. Gene amplification in Leishmania.Annu Rev Microbial 1991;45:417-444.
142. Ouellette M, Hettema E,Wust D et al. Direct and inverted DNA repeats associated with P-glycoprotein gene amplification in drug resistant Leishmania. Embo J 199 I; 10(4): 1009-I 016.
143. Grondin K, Roy G, Ouellette M. Formation of extrachromosomal circular amplicons with direct or inverted duplications in drug- resistant Leishmania tarentolae. Mol Cell Biol 1996; 16(7): 3587-3595.
144. Papadopoulou B, Roy G, Ouellette M.Autonomous replication of bacterial DNA plasmid oligomers in Leishmania. Mol Biochem Parasitol 1994; 65( I): 39-49.
145. Pamaik PK, BellofattoV, Hartree D, Cross GA.An episome of Trypanosoma brucei can exist as an extrachromosomal element in a broad range of trypanosomatids but shows different requirements for stable replication. Mol Biochem Parasitol 1994; 66(I): 153-156.
146. Cowman AF, Lew AM.Antifolate drug selection results in duplication and rearrangement of chromosome 7 in Plasmodium chabaudi. Mol Cell Biol 1989; 9( I I ): 5182-5188.
147. Wahl GM.The importance of circular DNA in mammalian gene amplification. Cancer Res 1989; 49(6): 1333-1340.
148. Quijada L, Soto M,Alonso C, Requena JM.Analysis of post- transcriptional regulation operating on transcription products of the tandemly linked Leishmania infantum hsp 70 genes.J Biol Chem 1997; 272(7): 4493-4499.
149. Aly R,Argaman M, Halman S, Shapira M.A regulatory role for the 5' and 3' untranslated regions in differentia[ expression of hsp83 in Leishmania. Nucleic Acids Res 1994; 22( 15): 2922-2929.
150. Svensson F, Ceriani C,Wallstrom EL et al. Cloning of a trypanosomatid gene coding for an ornithine decarboxylase that is metabolically unstable even though it lacks the C-terminal degradation domain. Proc Natl Acad Sci U S A 1997; 94(2): 397-402.
15 I. Bates PA. Complete developmental cycle of Leishmania mexicana in axenic culture. Parasitology 1994; 108(Pt I ): I-9.
152. Roberts WL, Berman JD, Rainey PM. In vitro antileishmanial properties of tri- and pentavalent antimonial preparations. Antimicrob Agents Chemother 1995; 39(6): 1234-1239.
© 1998 Harcourt Brace & Co. Ltd Drug Resistance Updates (1998) I, 266-278

153. Callahan HL, Portal AC, Devereaux R, Grogl M.An axenic amastigote system for drug screening.Antimicrob Agents Chemother 1997; 41 (4): 818-822.
154. Ephros M,Waldman E, Zilberstein D. Pentostam induces resistance to antimony and the preservative chlorocresol in Leishmania
donovani promastigotes and axenically grown amastigotes. Antimicrob Agents Chemother 1997; 41 (5): 1064-1068.
155. Sereno D, Lemesre JL. In vitro life cycle of pentamidine-resistant amastigotes: stability of the chemoresistant phenotypes is dependent on the level of resistance induced.Antimicrob Agents Chemother 1997; 41 (9): 1898-1903.
156. Ibrahim ME, Hag-Ali M, eI-Hassan AM et al. Leishmania resistant to sodium stibogluconate: drug-associated macrophage-dependent killing. Parasitol Res 1994; 80(7): 569-574.
Drug Resistance Updates (I 998) I, 266-278 © 1998 Harcourt Brace & Co. Ltd
Related Documents











