Critical Reviews in Plant Sciences, 24:23–58, 2005 Copyright c Taylor & Francis Inc. ISSN: 0735-2689 print / 1549-7836 online DOI: 10.1080/07352680590910410 Drought and Salt Tolerance in Plants Dorothea Bartels and Ramanjulu Sunkar Institute of Molecular Physiology and Biotechnology of Plants, University of Bonn, Kirschallee 1, D-53115 Bonn, Germany Table of Contents I. INTRODUCTION ............................................................................................................................................. 25 II. OSMOTIC STRESS .......................................................................................................................................... 25 A. Osmotic Stress-Induced Growth Arrest .......................................................................................................... 25 B. Osmotic Stress Affects Cell Division and Elongation ...................................................................................... 30 III. SIGNAL PERCEPTION ................................................................................................................................... 30 IV. SIGNAL TRANSDUCTION .............................................................................................................................. 31 A. MAPKinase Pathways .................................................................................................................................. 31 B. SNF-1-Like Kinases Are Involved in Osmotic Stress Signalling ....................................................................... 32 C. Phosphatases ............................................................................................................................................... 32 D. Phospholipid Signalling ............................................................................................................................... 33 1. Inositol 1,4,5-Triphosphate (IP3) ......................................................................................................... 33 2. Phosphatidic Acid (PA) ...................................................................................................................... 34 E. Other Signalling Molecules .......................................................................................................................... 34 1. Salicylic Acid .................................................................................................................................... 34 2. Nitric Oxide (NO) ............................................................................................................................. 34 V. CALCIUM SIGNALLING DURING DEHYDRATION AND SALT STRESS .................................................... 35 A. Calcium-Dependent Protein Kinases (CDPKs) ............................................................................................... 35 B. Calcium-Binding Proteins ............................................................................................................................ 35 C. Ca 2+ -Mediated SOS Pathways Are Involved in Ion Homeostasis ..................................................................... 35 D. Calcineurin B (CBLS) and Osmotic Stress Responses ..................................................................................... 35 E. Ca 2+ ATPases ............................................................................................................................................. 36 VI. TRANSCRIPTIONAL REGULATION OF GENE EXPRESSION .................................................................... 36 A. ABA Response Elements (ABREs) ............................................................................................................... 36 B. The Dehydration Response Element (DRE) ................................................................................................... 37 C. Transcription Factors Modulated by Osmotic Stress ........................................................................................ 37 1. Basic Region Leucine Zipper (bZIP) Proteins ....................................................................................... 37 2. Homeodomain-Leucine Zipper Proteins (HD-ZIP) ................................................................................ 37 3. Zn-Finger Proteins ............................................................................................................................. 37 4. AP2/ERF-Type Transcription Factors .................................................................................................. 38 5. Myb-Like Proteins ............................................................................................................................. 38 6. Myc-Like Proteins ............................................................................................................................. 39 7. CDT-1 .............................................................................................................................................. 39 Address correspondence to Dorothea Bartels, Institute of Molecular Physiology and Biotechnology of Plants, University of Bonn, Kirschallee 1, D-53115 Bonn, Germany. E-mail: [email protected] 23

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Critical Reviews in Plant Sciences, 24:23–58, 2005Copyright c© Taylor & Francis Inc.ISSN: 0735-2689 print / 1549-7836 onlineDOI: 10.1080/07352680590910410
Drought and Salt Tolerance in Plants
Dorothea Bartels and Ramanjulu SunkarInstitute of Molecular Physiology and Biotechnology of Plants, University of Bonn, Kirschallee 1,D-53115 Bonn, Germany
Table of Contents
I. INTRODUCTION .............................................................................................................................................25
II. OSMOTIC STRESS ..........................................................................................................................................25A. Osmotic Stress-Induced Growth Arrest ..........................................................................................................25B. Osmotic Stress Affects Cell Division and Elongation ......................................................................................30
III. SIGNAL PERCEPTION ...................................................................................................................................30
IV. SIGNAL TRANSDUCTION ..............................................................................................................................31A. MAPKinase Pathways ..................................................................................................................................31B. SNF-1-Like Kinases Are Involved in Osmotic Stress Signalling .......................................................................32C. Phosphatases ...............................................................................................................................................32D. Phospholipid Signalling ...............................................................................................................................33
1. Inositol 1,4,5-Triphosphate (IP3) .........................................................................................................332. Phosphatidic Acid (PA) ......................................................................................................................34
E. Other Signalling Molecules ..........................................................................................................................341. Salicylic Acid ....................................................................................................................................342. Nitric Oxide (NO) .............................................................................................................................34
V. CALCIUM SIGNALLING DURING DEHYDRATION AND SALT STRESS ....................................................35A. Calcium-Dependent Protein Kinases (CDPKs) ...............................................................................................35B. Calcium-Binding Proteins ............................................................................................................................35C. Ca2+-Mediated SOS Pathways Are Involved in Ion Homeostasis .....................................................................35D. Calcineurin B (CBLS) and Osmotic Stress Responses .....................................................................................35E. Ca2+ ATPases .............................................................................................................................................36
VI. TRANSCRIPTIONAL REGULATION OF GENE EXPRESSION ....................................................................36A. ABA Response Elements (ABREs) ...............................................................................................................36B. The Dehydration Response Element (DRE) ...................................................................................................37C. Transcription Factors Modulated by Osmotic Stress ........................................................................................37
1. Basic Region Leucine Zipper (bZIP) Proteins .......................................................................................372. Homeodomain-Leucine Zipper Proteins (HD-ZIP) ................................................................................373. Zn-Finger Proteins .............................................................................................................................374. AP2/ERF-Type Transcription Factors ..................................................................................................385. Myb-Like Proteins .............................................................................................................................386. Myc-Like Proteins .............................................................................................................................397. CDT-1 ..............................................................................................................................................39
Address correspondence to Dorothea Bartels, Institute of Molecular Physiology and Biotechnology of Plants, University of Bonn, Kirschallee1, D-53115 Bonn, Germany. E-mail: [email protected]
23

24 D. BARTELS AND R. SUNKAR
VII. ACCUMULATION OF SUGARS AND COMPATIBLE SOLUTES ...................................................................39A. Sugars ........................................................................................................................................................39B. Cyclitols .....................................................................................................................................................40C. Proline ........................................................................................................................................................40D. Glycine Betaine ...........................................................................................................................................40
VIII. PROTECTIVE PROTEINS AND OTHER PATHWAYS INVOLVED IN STRESS ADAPTATION .....................40A. Late Embryogenesis-Abundant (LEA) Proteins ..............................................................................................40B. Aquaporins .................................................................................................................................................41C. Heat Shock Proteins (Hsps) ..........................................................................................................................41D. Proteases and Proteinase Inhibitors ...............................................................................................................42E. Polyamines .................................................................................................................................................42
IX. OXIDATIVE STRESS A CONSEQUENCE OF DEHYDRATION AND SALT STRESS ....................................42A. Formation of Reactive Molecules ..................................................................................................................42B. Enzymes That Detoxify Aldehydes ...............................................................................................................43C. Peroxiredoxins ............................................................................................................................................43D. Thioredoxins ...............................................................................................................................................43E. Protein Oxidation ........................................................................................................................................43
X. IONIC STRESS ................................................................................................................................................44A. Na+ Toxicity and Homeostasis .....................................................................................................................44B. Na+ Exclusion ............................................................................................................................................44C. Na+ Compartmentalization ...........................................................................................................................44D. Proton Transporters and Salt Tolerance ..........................................................................................................45E. SOS Pathway and Ion Homeostasis ...............................................................................................................45
XI. ABSCISIC ACID (ABA) ....................................................................................................................................46A. Regulation of ABA Levels ............................................................................................................................46B. The Role of ABA in Stomatal Closure ...........................................................................................................47C. ABA Signalling Components ........................................................................................................................47
XII. MONITORING GLOBAL GENE EXPRESSION USING MICROARRAY ANALYSIS .....................................48
XIII. CONCLUSIONS ...............................................................................................................................................48
ACKNOWLEDGMENTS ..................................................................................................................................49
REFERENCES .................................................................................................................................................49
Agricultural productivity worldwide is subject to increasing en-vironmental constraints, particularly to drought and salinity dueto their high magnitude of impact and wide distribution. Tradi-tional breeding programs trying to improve abiotic stress toler-ance have had some success, but are limited by the multigenicnature of the trait. Tolerant plants such as Craterostigma planta-genium, Mesembryanthemum crystallinum, Thellungiella halophilaand other hardy plants could be valuable tools to dissect the ex-treme tolerance nature. In the last decade, Arabidopsis thaliana, agenetic model plant, has been extensively used for unravelling themolecular basis of stress tolerance. Arabidopsis also proved to be
extremely important for assessing functions for individual stress-associated genes due to the availability of knock-out mutants andits amenability for genetic transformation. In this review, the re-sponses of plants to salt and water stress are described, the regula-tory circuits which allow plants to cope with stress are presented,and how the present knowledge can be applied to obtain tolerantplants is discussed.
Keywords dehydration, salinity, ABA, plant stress tolerance, trans-genic plants

DROUGHT AND SALT TOLERANCE IN PLANTS 25
I. INTRODUCTIONDrought and salinity are two major environmental factors
determining plant productivity and plant distribution. Droughtand salinity affect more than 10 percent of arable land, and de-sertification and salinization are rapidly increasing on a globalscale declining average yields for most major crop plants bymore than 50 percent (Bray et al., 2000). Understanding planttolerance to drought and salinity is therefore of fundamental im-portance and forms one of the major research topics. Plants canperceive abiotic stresses and elicit appropriate responses withaltered metabolism, growth and development. The regulatorycircuits include stress sensors, signalling pathways comprisinga network of protein-protein reactions, transcription factors andpromoters, and finally the output proteins or metabolites. Clas-sical breeding approaches revealed that stress tolerance traitsare mainly quantitative trait loci (QTLs), which make geneticselection of traits difficult. Nevertheless, very respectable stresstolerant crops have been obtained, mainly by introducing traitsfrom stress-adapted wild relatives.
As water and salt stresses occur frequently and can affect mosthabitats, plants have developed several strategies to cope withthese challenges: either adaptation mechanisms, which allowthem to survive the adverse conditions, or specific growth habitsto avoid stress conditions. Stress-tolerant plants have evolvedcertain adaptive mechanisms to display different degrees of tol-erance, which are largely determined by genetic plasticity. Dif-ferential stress tolerance could be attributed to differences inplant reactivity in terms of stress perception, signal transduc-tion and appropriate gene expression programs, or other novelmetabolic pathways that are restricted to tolerant plants. Thehypothesis that the genetic program for tolerance is at least tosome extent also present in nontolerant plants is supported by theobservation that gradual acclimation of sensitive plants leads toacquisition of tolerance to some degree. These plants may needgradual adaptation for proper expression of genes responsiblefor acquisition of tolerance (Zhu, 2001).
Our understanding of how plants respond to water and saltstress has advanced by analyzing stress-tolerant species like thedesiccation tolerant plant Cratesostigma plantagineum (Bartelsand Salamini, 2001) or the salt-tolerant plant Mesembryanthe-mum crystallinum (Bohnert and Cushman, 2000). Despite thefact that research with the desiccation-tolerant plant C. plan-tagineum revealed additional novel aspects such as specific car-bohydrate metabolism and the existence of CDT-1 gene thatwere unknown in other nontolerant plant species (Bartels andSalamini, 2001), the connection between these metabolites andtolerance is still correlative. Molecular genetic studies have beenperformed with Arabidopsis thaliana, which does not displayextreme stress tolerance, but shows many stress responses atthe molecular level and has therefore been successfully used fora genetic dissection of stress response pathways (Zhu, 2002;Shinozaki et al., 2003). More importantly, we learned that it isvery likely that the extreme tolerant model plants did not acquireunique genes since stress-relevant genes are ubiquitously present
in the plant kingdom. A particular gene expression pattern is of-ten associated with the tolerant phenotype and it is unknownto date how this is achieved. This may involve other molecu-lar aspects, like chromatin organization, which have not beenwell researched. Recently, the salt-tolerant plant Thellungiellahalophila was introduced as an attractive model plant to studymolecular genetics of salt tolerance, because it is a close rel-ative to Arabidopsis and amenable for transformation unlikethat of other tolerant model plants (Volkov et al., 2003; Bressanet al., 2001; Zhu, 2001). Thus it may be possible to combineprofound genetic knowledge with the expression of extremetolerance.
Exposure to drought or salt stress triggers many commonreactions in plants. Both stresses lead to cellular dehydration,which causes osmotic stress and removal of water from the cy-toplasm into the extracellular space resulting in a reduction ofthe cytosolic and vacuolar volumes. Another consequence is theproduction of reactive oxygen species which then in turn affectscellular structures and metabolism negatively. Early responsesto water and salt stress are largely identical except for the ioniccomponent. These similarities include metabolic processes suchas, for example, a decrease of photosynthesis or hormonal pro-cesses like rising levels of the plant hormone ABA. High in-tracellular concentrations of sodium and chloride ions are anadditional problem of salinity stress.
Adaptation to salinity and drought is undoubtedly one ofthe complex processes, involving numerous changes includ-ing attenuated growth, the activation/increased expression orinduction of genes, transient increases in ABA levels, accumu-lation of compatible solutes and protective proteins, increasedlevels of antioxidants and suppression of energy-consumingpathways. However, no consensus has been reached in defin-ing the key processes determining tolerance and the secondaryfollow-up processes. With the advancement of high throughputDNA technologies, several hundred stress-induced or upregu-lated genes have been identified. The search for stress-associatedgenes may have been saturated at least in Arabidopsis. How-ever, the function of only a limited number of gene products areknown (Ingram and Bartels, 1996; Bray, 1997; Shinozaki andYamaguchi-Shinozaki, 1997; Hasegawa et al., 2000; Ramanjuluand Bartels, 2002). Several stress-associated genes have beenevaluated or studies are in progress for their contribution todrought or salt tolerance in laboratory studies (Table 1). Morerigorous and perhaps field studies are required for possible uti-lization of these genes for improving stress tolerance in agricul-tural plants through biotechnological approaches.
II. OSMOTIC STRESS
A. Osmotic Stress-Induced Growth ArrestArrest of plant growth during stress conditions largely de-
pends on the severity of the stress. Mild osmotic stress leadsrapidly to growth inhibition of leaves and stems, whereas rootsmay continue to elongate (Westgate and Boyer, 1985; Sharp
26 D. BARTELS AND R. SUNKAR
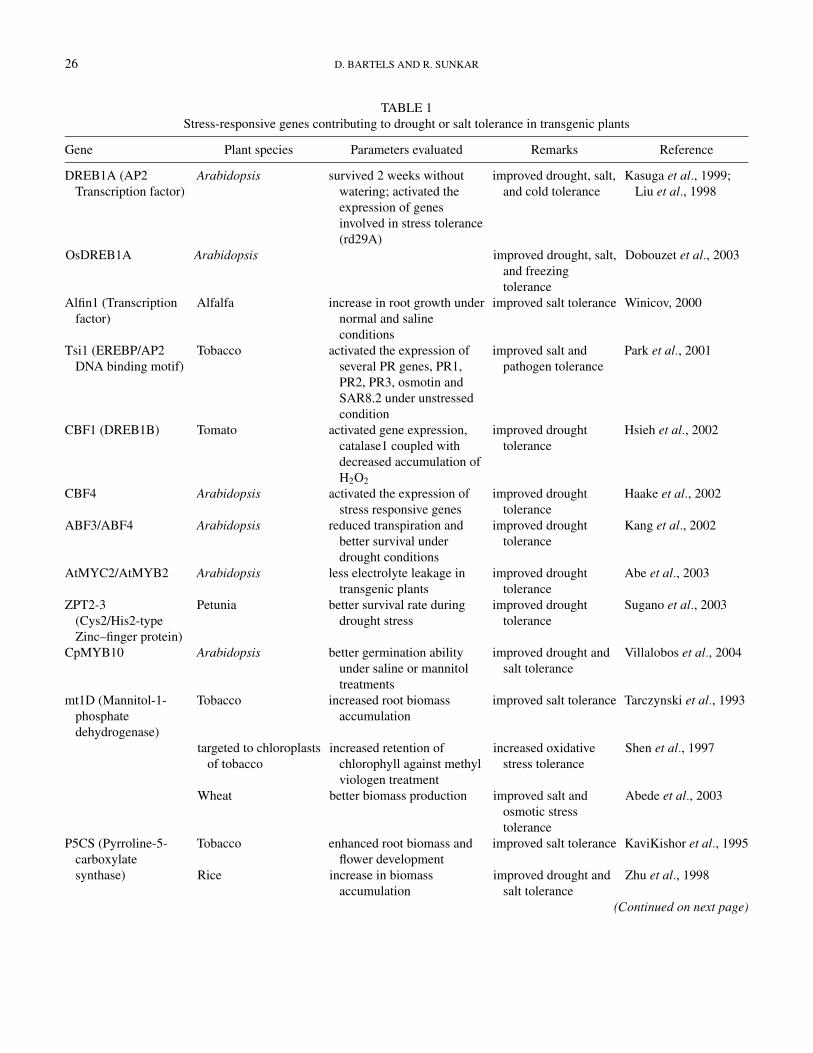
TABLE 1Stress-responsive genes contributing to drought or salt tolerance in transgenic plants
Gene Plant species Parameters evaluated Remarks Reference
DREB1A (AP2Transcription factor)
OsDREB1A
Arabidopsis
Arabidopsis
survived 2 weeks withoutwatering; activated theexpression of genesinvolved in stress tolerance(rd29A)
improved drought, salt,and cold tolerance
improved drought, salt,and freezingtolerance
Kasuga et al., 1999;Liu et al., 1998
Dobouzet et al., 2003
Alfin1 (Transcriptionfactor)
Alfalfa increase in root growth undernormal and salineconditions
improved salt tolerance Winicov, 2000
Tsi1 (EREBP/AP2DNA binding motif)
Tobacco activated the expression ofseveral PR genes, PR1,PR2, PR3, osmotin andSAR8.2 under unstressedcondition
improved salt andpathogen tolerance
Park et al., 2001
CBF1 (DREB1B) Tomato activated gene expression,catalase1 coupled withdecreased accumulation ofH2O2
improved droughttolerance
Hsieh et al., 2002
CBF4 Arabidopsis activated the expression ofstress responsive genes
improved droughttolerance
Haake et al., 2002
ABF3/ABF4 Arabidopsis reduced transpiration andbetter survival underdrought conditions
improved droughttolerance
Kang et al., 2002
AtMYC2/AtMYB2 Arabidopsis less electrolyte leakage intransgenic plants
improved droughttolerance
Abe et al., 2003
ZPT2-3(Cys2/His2-typeZinc–finger protein)
Petunia better survival rate duringdrought stress
improved droughttolerance
Sugano et al., 2003
CpMYB10 Arabidopsis better germination abilityunder saline or mannitoltreatments
improved drought andsalt tolerance
Villalobos et al., 2004
mt1D (Mannitol-1-phosphatedehydrogenase)
Tobacco
targeted to chloroplastsof tobacco
Wheat
increased root biomassaccumulation
increased retention ofchlorophyll against methylviologen treatment
better biomass production
improved salt tolerance
increased oxidativestress tolerance
improved salt andosmotic stresstolerance
Tarczynski et al., 1993
Shen et al., 1997
Abede et al., 2003
P5CS (Pyrroline-5-carboxylatesynthase)
Tobacco
Rice
enhanced root biomass andflower development
increase in biomassaccumulation
improved salt tolerance
improved drought andsalt tolerance
KaviKishor et al., 1995
Zhu et al., 1998
(Continued on next page)
DROUGHT AND SALT TOLERANCE IN PLANTS 27
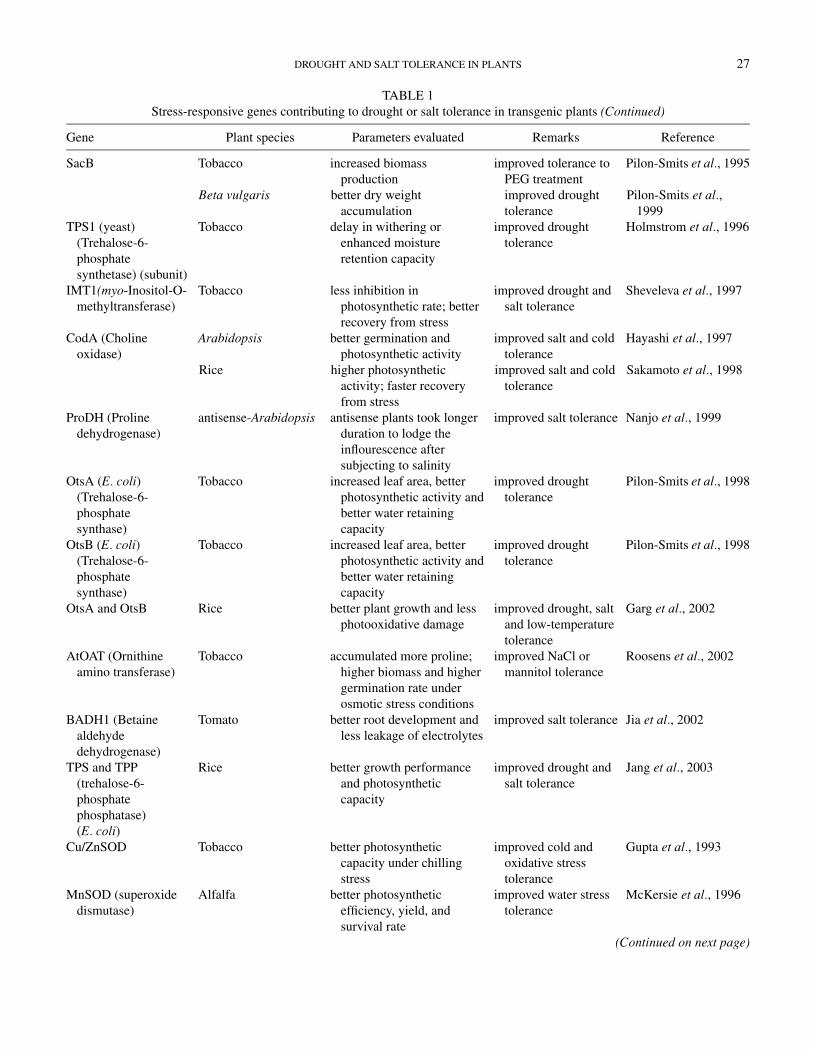
TABLE 1Stress-responsive genes contributing to drought or salt tolerance in transgenic plants (Continued)
Gene Plant species Parameters evaluated Remarks Reference
SacB Tobacco
Beta vulgaris
increased biomassproduction
better dry weightaccumulation
improved tolerance toPEG treatmentimproved droughttolerance
Pilon-Smits et al., 1995
Pilon-Smits et al.,1999
TPS1 (yeast)(Trehalose-6-phosphatesynthetase) (subunit)
Tobacco delay in withering orenhanced moistureretention capacity
improved droughttolerance
Holmstrom et al., 1996
IMT1(myo-Inositol-O-methyltransferase)
Tobacco less inhibition inphotosynthetic rate; betterrecovery from stress
improved drought andsalt tolerance
Sheveleva et al., 1997
CodA (Cholineoxidase)
Arabidopsis
Rice
better germination andphotosynthetic activity
higher photosyntheticactivity; faster recoveryfrom stress
improved salt and coldtolerance
improved salt and coldtolerance
Hayashi et al., 1997
Sakamoto et al., 1998
ProDH (Prolinedehydrogenase)
antisense-Arabidopsis antisense plants took longerduration to lodge theinflourescence aftersubjecting to salinity
improved salt tolerance Nanjo et al., 1999
OtsA (E. coli)(Trehalose-6-phosphatesynthase)
Tobacco increased leaf area, betterphotosynthetic activity andbetter water retainingcapacity
improved droughttolerance
Pilon-Smits et al., 1998
OtsB (E. coli)(Trehalose-6-phosphatesynthase)
Tobacco increased leaf area, betterphotosynthetic activity andbetter water retainingcapacity
improved droughttolerance
Pilon-Smits et al., 1998
OtsA and OtsB Rice better plant growth and lessphotooxidative damage
improved drought, saltand low-temperaturetolerance
Garg et al., 2002
AtOAT (Ornithineamino transferase)
Tobacco accumulated more proline;higher biomass and highergermination rate underosmotic stress conditions
improved NaCl ormannitol tolerance
Roosens et al., 2002
BADH1 (Betainealdehydedehydrogenase)
Tomato better root development andless leakage of electrolytes
improved salt tolerance Jia et al., 2002
TPS and TPP(trehalose-6-phosphatephosphatase)(E. coli)
Rice better growth performanceand photosyntheticcapacity
improved drought andsalt tolerance
Jang et al., 2003
Cu/ZnSOD Tobacco better photosyntheticcapacity under chillingstress
improved cold andoxidative stresstolerance
Gupta et al., 1993
MnSOD (superoxidedismutase)
Alfalfa better photosyntheticefficiency, yield, andsurvival rate
improved water stresstolerance
McKersie et al., 1996
(Continued on next page)
28 D. BARTELS AND R. SUNKAR
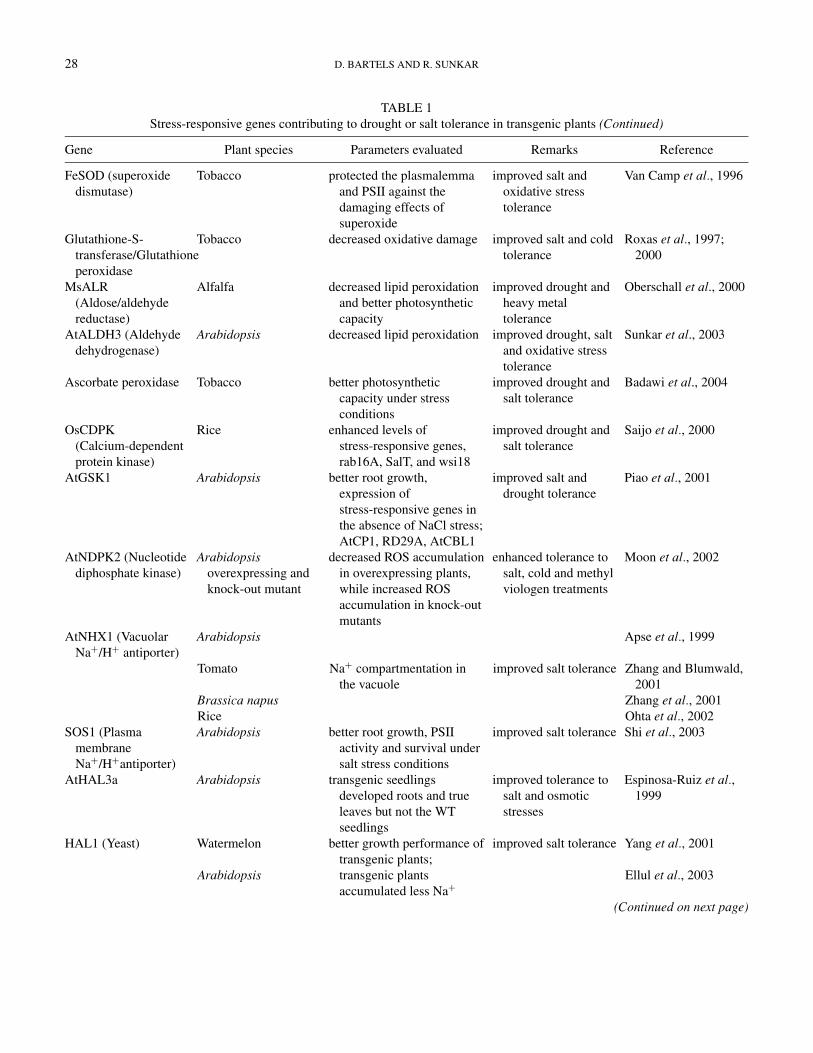
TABLE 1Stress-responsive genes contributing to drought or salt tolerance in transgenic plants (Continued)
Gene Plant species Parameters evaluated Remarks Reference
FeSOD (superoxidedismutase)
Tobacco protected the plasmalemmaand PSII against thedamaging effects ofsuperoxide
improved salt andoxidative stresstolerance
Van Camp et al., 1996
Glutathione-S-transferase/Glutathioneperoxidase
Tobacco decreased oxidative damage improved salt and coldtolerance
Roxas et al., 1997;2000
MsALR(Aldose/aldehydereductase)
Alfalfa decreased lipid peroxidationand better photosyntheticcapacity
improved drought andheavy metaltolerance
Oberschall et al., 2000
AtALDH3 (Aldehydedehydrogenase)
Arabidopsis decreased lipid peroxidation improved drought, saltand oxidative stresstolerance
Sunkar et al., 2003
Ascorbate peroxidase Tobacco better photosyntheticcapacity under stressconditions
improved drought andsalt tolerance
Badawi et al., 2004
OsCDPK(Calcium-dependentprotein kinase)
Rice enhanced levels ofstress-responsive genes,rab16A, SalT, and wsi18
improved drought andsalt tolerance
Saijo et al., 2000
AtGSK1 Arabidopsis better root growth,expression ofstress-responsive genes inthe absence of NaCl stress;AtCP1, RD29A, AtCBL1
improved salt anddrought tolerance
Piao et al., 2001
AtNDPK2 (Nucleotidediphosphate kinase)
Arabidopsisoverexpressing andknock-out mutant
decreased ROS accumulationin overexpressing plants,while increased ROSaccumulation in knock-outmutants
enhanced tolerance tosalt, cold and methylviologen treatments
Moon et al., 2002
AtNHX1 (VacuolarNa+/H+ antiporter)
Arabidopsis
Tomato
Brassica napusRice
Na+ compartmentation inthe vacuole
improved salt tolerance
Apse et al., 1999
Zhang and Blumwald,2001
Zhang et al., 2001Ohta et al., 2002
SOS1 (PlasmamembraneNa+/H+antiporter)
Arabidopsis better root growth, PSIIactivity and survival undersalt stress conditions
improved salt tolerance Shi et al., 2003
AtHAL3a Arabidopsis transgenic seedlingsdeveloped roots and trueleaves but not the WTseedlings
improved tolerance tosalt and osmoticstresses
Espinosa-Ruiz et al.,1999
HAL1 (Yeast) Watermelon
Arabidopsis
better growth performance oftransgenic plants;transgenic plantsaccumulated less Na+
improved salt tolerance Yang et al., 2001
Ellul et al., 2003
(Continued on next page)
DROUGHT AND SALT TOLERANCE IN PLANTS 29
TABLE 1Stress-responsive genes contributing to drought or salt tolerance in transgenic plants (Continued)
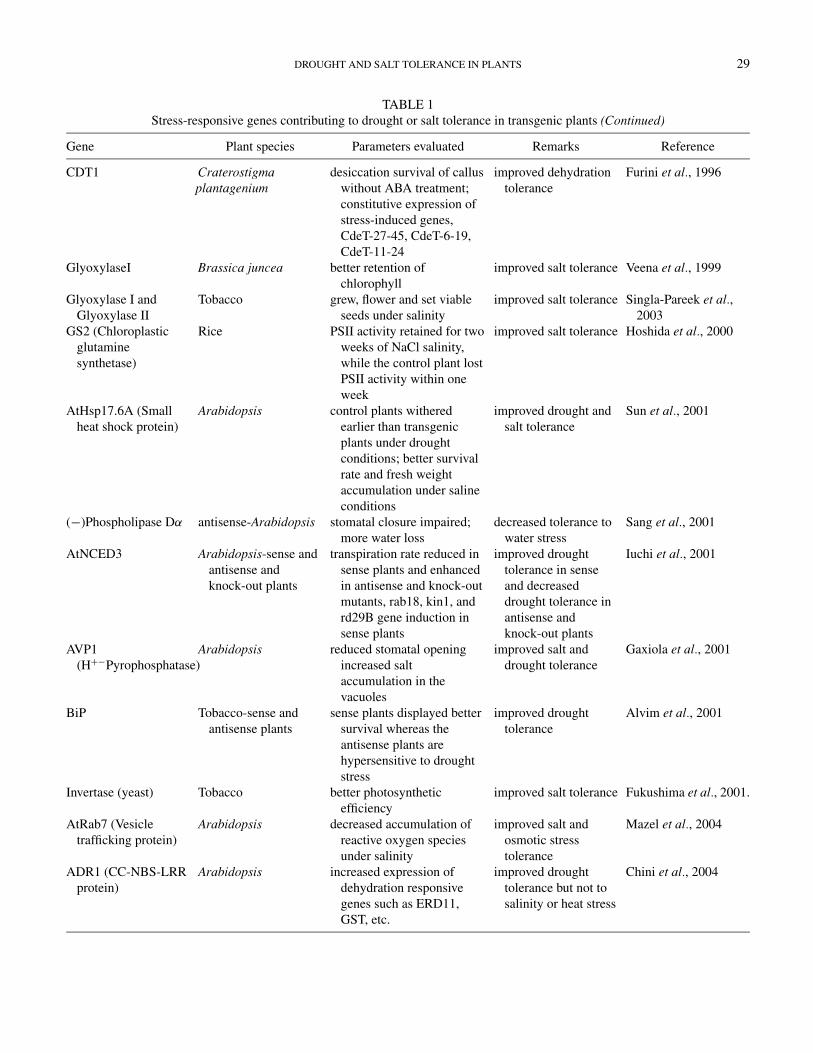
Gene Plant species Parameters evaluated Remarks Reference
CDT1 Craterostigmaplantagenium
desiccation survival of calluswithout ABA treatment;constitutive expression ofstress-induced genes,CdeT-27-45, CdeT-6-19,CdeT-11-24
improved dehydrationtolerance
Furini et al., 1996
GlyoxylaseI Brassica juncea better retention ofchlorophyll
improved salt tolerance Veena et al., 1999
Glyoxylase I andGlyoxylase II
Tobacco grew, flower and set viableseeds under salinity
improved salt tolerance Singla-Pareek et al.,2003
GS2 (Chloroplasticglutaminesynthetase)
Rice PSII activity retained for twoweeks of NaCl salinity,while the control plant lostPSII activity within oneweek
improved salt tolerance Hoshida et al., 2000
AtHsp17.6A (Smallheat shock protein)
Arabidopsis control plants witheredearlier than transgenicplants under droughtconditions; better survivalrate and fresh weightaccumulation under salineconditions
improved drought andsalt tolerance
Sun et al., 2001
(−)Phospholipase Dα antisense-Arabidopsis stomatal closure impaired;more water loss
decreased tolerance towater stress
Sang et al., 2001
AtNCED3 Arabidopsis-sense andantisense andknock-out plants
transpiration rate reduced insense plants and enhancedin antisense and knock-outmutants, rab18, kin1, andrd29B gene induction insense plants
improved droughttolerance in senseand decreaseddrought tolerance inantisense andknock-out plants
Iuchi et al., 2001
AVP1(H+−Pyrophosphatase)
Arabidopsis reduced stomatal openingincreased saltaccumulation in thevacuoles
improved salt anddrought tolerance
Gaxiola et al., 2001
BiP Tobacco-sense andantisense plants
sense plants displayed bettersurvival whereas theantisense plants arehypersensitive to droughtstress
improved droughttolerance
Alvim et al., 2001
Invertase (yeast) Tobacco better photosyntheticefficiency
improved salt tolerance Fukushima et al., 2001.
AtRab7 (Vesicletrafficking protein)
Arabidopsis decreased accumulation ofreactive oxygen speciesunder salinity
improved salt andosmotic stresstolerance
Mazel et al., 2004
ADR1 (CC-NBS-LRRprotein)
Arabidopsis increased expression ofdehydration responsivegenes such as ERD11,GST, etc.
improved droughttolerance but not tosalinity or heat stress
Chini et al., 2004

30 D. BARTELS AND R. SUNKAR
et al., 1988; Nonami and Boyer, 1990; Spollen et al., 1993). Thedegree of growth inhibition due to osmotic stress depends on thetime scale of the response, the particular tissue and species inquestion, and how the stress treatment was given (rapid or grad-ual). Growth arrest can be considered as a possibility to preservecarbohydrates for sustained metabolism, prolonged energy sup-ply, and for better recovery after stress relief. The inhibitionof shoot growth during water deficit is thought to contribute tosolute accumulation and thus eventually to osmotic adjustment(Osorio et al., 1998). For instance, hexose accumulation ac-counts for a large proportion of the osmotic potential in the cellelongation zone in cells of the maize root tip (Sharp et al., 1990).On the other hand, continuation of root growth under droughtstress is an adaptive mechanism that facilitates water uptakefrom deeper soil layers. Similarly, continued root growth undersalt stress may provide additional surfaces for sequestration oftoxic ions, leading to lower salt concentration. For example, salttolerance of barley was correlated with the better root growthrates coupled with fast development and early flowering (Munnset al., 2000).
B. Osmotic Stress Affects Cell Division and ElongationPlants have the unique attribute of modulating their devel-
opment with the prevailing environmental conditions involv-ing plant hormones, which in turn influence gene expressionprograms. Cell division is the principal determinant of meris-tem activity and determines the overall plant growth rate. It hasbeen proposed that environmental and developmental controlsof growth rate act by regulating cyclin-dependent kinase (CDK)activity and cell division (Cockcroft et al., 2000; West et al.,2004). CDKs are a family of protein kinases, each with a posi-tive regulatory subunit termed a cyclin and the catalytic subunitCDK (den Boer and Murray, 2000). CDKs are emerging as keyplayers in regulation of cell division and are likely to be regulatedat both transcriptional and post-translational levels in responseto stress. For example, maize ZmCdc2 (a member of the CDKfamily) was shown to be downregulated by water stress lead-ing to a decrease in mitotic cell cycling (Setter and Flanningan,2001). The decrease in cell division in response to water stressis characterized by lower CDK activity, which is correlated withtyrosine phosphorylation (Schuppler et al., 1998). The recentfinding that ABA induces expression of an inhibitor of CDK(ICK1) links cell division and ABA (Wang et al., 1998). This isfurther supported by studies of Kang et al. (2002) who reportedthat expression of the ABA inducible cell cycle regulator ICK1was increased in transgenic plants overexpressing ABF3 andABF4 resulting in dwarf phenotypes. These mechanisms couldbe responsible for ABA-dependent cell cycle arrest during os-motic stress in plants.
Cell expansion is a coordinately regulated process at thewhole plant level and is influenced by external stimuli includingwater availability. The rate of cell expansion is mainly deter-mined by two parameters, cell wall extensibility and cellular
osmotic potential. The enlargement of plant cells involves con-trol of wall synthesis and expansion, solute and water transport,membrane synthesis, Golgi secretion, ion transport and otherprocesses (Cosgrove, 1997). Expansins are a family of plant pro-teins essential for acid-induced cell wall loosening (Cosgrove,1997).
Expression of three expansin genes Exp1, Exp5, and ExpB8was upregulated in the apical region of roots after growth atlow water potential leading to higher amounts of expansin pro-tein, which is closely correlated with the root elongation (Wuet al., 2001). These results are consistent with the hypothesisthat the adaptive wall loosening and growth maintenance in theapical region of maize roots are partly due to altered expansingene expression in the root tip at low water potentials (Wu et al.,2001). Expansin genes in Craterostigma plantagineum, CpExp1and CpExp3, are upregulated to different degrees in response todehydration. In addition CpExp1 but not CpExp3, is also upreg-ulated in response to rehydration (Jones and McQueen-Mason,2004). These results implicate a role for expansins during de-hydration and rehydration, particularly in increasing wall flexi-bility. However, the effect of osmotic stress on cell enlargementis still not clear and may also involve other hormones such asauxin, cytokinin, or gibberellins.
III. SIGNAL PERCEPTIONIt is still an open question how plants sense osmotic stress.
Because no plant molecule has truely been identified as os-mosensor, scientists oriented themselves to study how yeast andmicroorganisms sense osmotic stress. In yeast, hyperosmoticstress is sensed by two types of osmosensors, SLN1 and SHO1,that feed finally into HOG (high-osmolarity glycerol) MAPKpathway. High osmolarity induces loss of turgor that leads con-comitantly to shrinkage of cell volume and an increase in thedistance between plasma membrane and cell wall. SLN1 is likelyto sense the change in turgor pressure (Reiser et al., 2003).SLN1 is a two-component regulatory system, which is also awell characterized signal transduction element in prokaryotes.The two-component regulatory system consists of a phospho-relay between three proteins. The yeast osmosensor SLN1 is afused two-component system that autophosphorylates a histi-dine residue in the N-terminal sensor domain and then transfersthe phosphate group to an aspartate residue in the C-terminal-located response-regulator domain. The phosphate is transferredto YPD1, which functions as a second histidine phosphorelayintermediate between SLN1 and the response regulator SSK1.SSK1 finally feeds into the HOG pathway, which responds toincreased extracellular osmolarity and is responsible for os-molyte (glycerol) accumulation (Posas et al., 1996; Wurgler-Murphy and Saito, 1997). Another yeast membrane protein in-volved in osmosensing is SHO1, which also converges withthe MAPK pathway. Which pathway is activated depends onthe osmotic stress level. In Arabidopsis an SLN1 homologue,AtHK1, was identified; it can function as an osmosensor in yeast

DROUGHT AND SALT TOLERANCE IN PLANTS 31
and complements SLN1-deficient yeast mutants (Urao et al.,1999).
Recently, NtC7, which is originally identified as a gene thatis responsive to wounding, has been suggested as another proba-ble candidate for sensing osmotic stress in plants (Tamura et al.,2003). NtC7 transcript accumulates rapidly and transiently notonly in response to wounding but also to salt and osmotic stress.The NtC7 gene encodes a receptor-like membrane protein andoverexpression improved osmotic stress tolerance induced bymannitol but not by NaCl (Tamura et al., 2003). These resultssuggest that NtC7 may play a role in sensing specifically os-motic stress. Cre1 (cytokinin response 1) is a cytokinin receptorin Arabidopsis. Recent experiments implicate Cre1 as anotherlikely candidate for sensing osmotic stress in plants (Reiseret al., 2003). Cre1 and Sln1 have similar organizations of thecytoplasmic histidine kinase and receiver domains. These ob-servations suggest that several candidates for osmosensors havebeen proposed, although their role as osmosensors has yet to bedemonstrated.
IV. SIGNAL TRANSDUCTIONPlants react to external stimuli by initiating signalling cas-
cade which activate the expression of appropriate responses. Incontrast to signal perception various components of the signaltransduction have been identified, although it is largely unknownhow the different molecules interact with each other and wherethey are positioned in the complex signalling network. Thesesignalling pathways comprise a network of protein-protein re-actions and signalling molecules (for example, ROS, Ca2+ etc.).Reversible phosphorylation of proteins is an important mecha-nism, by which organisms regulate cellular processes in responseto environmental cues. In this review, we will consider severalclasses of protein kinases and phosphatases as signal transduc-ers that were shown to be involved in osmotic stress signalling.This will be followed by a description of the role of calcium assecond messenger molecules.
A. MAPKinase PathwaysProtein phosphorylation is one of the major mechanisms for
controlling cellular functions in response to external signals. Themitogen-activated protein kinase (MAPK) cascades are commonsignalling modules in eukaryotic cells including plants. A gen-eral feature of MAPK cascades is their composition of threefunctionally linked protein kinases. MAP kinase activation re-quires the phosphorylation of conserved threonine and tyrosineresidues in the so-called TEY (Thr, Glu, Tyr) activation loop bya specific MAPK kinase (MAPKK). A MAPKK kinase (MAP-KKK) activates through phosphorylation of conserved threonineand/or serine residues. At the downstream end of the cascade, ac-tivation of the cytoplasmic MAPK module often induces translo-cation of the MAPK into the nucleus where the kinase is ableto activate genes through phosphorylation of transcription fac-tors (Triesmann, 1996). In some other cases, a given MAPK
may translocate to other sites in the cytoplasm to phosphory-late specific enzymes or cytoskeletol components (Robinson andCobb, 1997). By tight regulation of the MAPK localization andthrough expression of signalling components and substrates intarget cells, tissues or organs, MAPK pathways can mediatesignalling of an extracellular stimulus and bring about specificresponses.
MAPK pathways may integrate a variety of upstream signalsthrough interaction with other kinases or G proteins (Robinsonand Cobb, 1997). The G proteins often directly serve as couplingagent between a plasma membrane–located receptor protein thatsenses an extracellular stimulus and a cytoplasmic module. Gproteins and kinases in yeast and mammals have been shown toregulate MAPKKKs (Kyriakis and Avruch, 2001). The actualmechanisms of MAPKKK activation by osmotic stress in mam-malian cells remain largely uncharacterized. Mammalian cellsactivate three different MAPKs in response to osmotic stress:P38, JNK, and ERK5 (extracellular signal regulated proteinkinases) (de Nadal et al., 2002). When compared with mam-malian MAPKs, all plant MAPKs have highest homology to theERK subfamily. Control of gene expression is a major outcomeof stress-activated MAPK pathways. In mammalian cells, P38(MAPK) controls the expression of >100 genes, while in yeast,genome-wide transcription studies revealed that a large numberof genes (∼7%) show transient changes in their expression levelsafter a mild osmotic shock and that the HOG1 MAPK pathwayplays a key role (de Nadal et al., 2002).
At least 20 MAPK, 10 MAPKK and 60 MAPKKK geneshave been identified in Arabidopsis on the basis of sequence sim-ilarities (Riechmann et al., 2000; Ichimura et al., 2002). Giventhe imbalance in numbers it is likely that the pathways are notbeing linear and convergence of pathways is expected. We arealready aware that some MAPKinase-activated pathways appar-ently overlap, i.e., several genes are induced by more than onestressor (Knight and Knight, 2001). For instance, AtMPK6 andAtMPK3 are activated by osmotic stress in Arabidopsis, and thetobacco orthologs SIPK (salicylic acid–inducible protein kinase)and WIPK (wound-inducible protein kinase) are also activatedby biotic stresses indicating the point of convergence of differ-ent signalling cascades and yet leading to appropriate responses(Singh et al., 2002). Recent studies suggest that different stimuliactivate MAPKs to different levels and with different kinetics,which may encode signal specificity. Thus, the same MAPKsmay participate in different signalling events (reviewed in Tenaet al., 2001).
Moderate and extreme hyperosmotic stress activated two dis-tinct kinases in alfalfa cells, a 46 kDa MAP kinase identified tobe SIMK under moderate osmotic stress conditions, whereas a38 kDa protein kinase becomes activated under extreme hyper-osmotic stress conditions (Munnik et al., 1999). This situationresembles the operation of the osmosensing SLN1and SHO1pathways in yeast, and suggests the possible existence of dis-tinct sensors for moderate and extreme hyperosmotic stress inplants. To determine the upstream activator of SIMK, Kiegerl

32 D. BARTELS AND R. SUNKAR
et al. (2000) used SIMK as bait in yeast two-hybrid screeningand isolated MAPKK (SIMK kinase). SIMKK encodes a func-tional protein kinase that specifically activates SIMK in vitro andin vivo. SIMKK phosphorylates SIMK on the threonine and ty-rosine residues of the activation loop, establishing that SIMKKis a specific dual-specificity protein kinase of SIMK (Kiegerlet al., 2000). The interaction of SIMK with SIMKK was furtherconfirmed in an in vivo system. SIMK is only partially activatedby NaCl in transfected parsley protoplasts, whereas it can befully activated by co-transfection of SIMKK.
In Arabidopsis, AtMEKK1 (a MAPKinase-kinase-kinase)and AtMPK3 (a MAPKinase) are activated by dehydration, touchand cold (Mizoguchi et al., 1996). In accordance with a role forAtMPK3 during dehydration, the closely related alfalfa homo-logue SAMK is also transcriptionally upregulated upon droughtstress (Jonak et al., 1996). In Arabidopsis, AtMPK4 and AtMPK6are post-translationally activated by cold, osmotic stress, andwounding (Ichimura et al., 2000). Recently, OXI1, a serine/threonine kinase was identified as a downstream componentof ROS signalling, possibly acting upstream to the AtMPK3and AtMPK6. OXI1 is transcriptionally upregulated by H2O2,wounding pathogen attack and osmotic stress (Rentel et al.,2004). Downstream targets of OXI1 could be AtMPK3 andAtMPK6, since the OXI1 is required for the activation of MPK3and MPK6.
ADR1, a CC-NBS-LRR gene, that shows homology withthe serine/threonine protein kinases and that has been originallyimplicated as a component of disease resistance turns out to beimportant also for abiotic stress responses (Chini et al., 2004).The ADR1 transgenic plants display dual but opposite responsesto dehydration and other abiotic stresses. These plants exhibitimproved dehydration tolerance but are hypersensitive to salinityand high temperature stresses. The improved tolerance appearsto be correlated with the dehydration-responsive gene expression(Chini et al., 2004). These observations implicate overlappingbiotic and abiotic stress signalling pathways using kinases asconverging points of stress signalling pathway and yet capableof eliciting stress-specific responses.
B. SNF-1-Like Kinases Are Involved in OsmoticStress Signalling
Another family of protein kinases are the SNF1/AMP-activated protein kinases, which were first analysed in yeast fromwhere the name originated (SNF = sucrose-nonfermenting).These kinases may sense the ATP/AMP ratio and thus controlfluxes between anabolism and catabolism via transcription ofgenes encoding enzymes related to carbohydrate metabolism.In plants some members of this group of kinases are expressedin response to dehydration or ABA. The kinases range between∼40 and 50 kDa, and are activated by phosphorylation of serineor threonine. SNF-1-related protein kinases (SnRKs) have beenclassified into three families, SnRK1, SnRK2, and SnRK3 withunknown function (Halford and Hardie, 1998). PKABA1 from
wheat (Anderberg and Walker-Simmons, 1992), ARSK1 (Ara-bidopsis root specific kinase 1) (Hwang and Goodman, 1995),two kinases in Dunaliella (Yuasa and Muto, 1996), a maize45 kDa kinase (Conley et al., 1997), SPK3 and SPK4-kinasesfrom soybean (Yoon et al., 1997), a 42 kDa kinase from to-bacco (Mikolayczyk et al., 2000), a 38 kDa kinase in alfalfa(Munnik et al., 1999; Munnik and Meijer, 2001), a guard cell-localized ABA-activated protein kinase, AAPK from Vicia faba(Li et al., 2000), Arabidopsis OST1 protein kinase which is re-lated to AAPK of Vicia faba (Mustilli et al., 2002) are predictedto belong to this group of kinases and are activated in responseto osmotic stress. The rice genome appears to encode 10 pro-tein kinases belonging to this class and surprisingly all are acti-vated by osmotic stress, and three of them (SAPK8, SAPK9,and SAPK10) are also activated by ABA (Kobayashi et al.,2004). Both SAPK1 and SAPK2 from rice are highly homol-ogous to PKABA1. Additionally, these kinases seem to be tran-scriptionally either upregulated or downregulated by osmoticstress and ABA. The biological significance of the osmotic- orABA-activation of these kinases is largely unknown. However,their activation in response to different abiotic stresses to differ-ent levels implicates a major role for these kinases.
C. PhosphatasesThe action of the protein kinases is counteracted by phos-
phatases providing modulation and reversibility of the phos-phoregulatory mechanism. Phosphatases are classified accord-ing to their substrate specificity. There are two major groupsof phosphatases: phosphoprotein (serine/threonine) phophatases(or PPases) and phosphotyrosine (protein tyrosine phosphatasesor PTPases). PPases are classified into four groups (PP1, PP2A,PP2B, and PP2C) based on their biochemical and pharmaco-logical properties (Cohen, 1989). The PTPases form three sub-groups: receptor-like PTPases, intracellular PTPases, and dual-specific PTPases.
Two major families of phosphatases interact with and inac-tivate HOG1 in yeast: the serine/threonine protein phosphatasetype 2C (PP2C) and the protein tyrosine phosphatases (PTPases)(reviewed in de Nadal et al., 2002). Tyrosine specific phos-phatases (PTPases) play a major role in the regulation of MAPKpathway in yeast (Shinozaki and Russel, 1995). PTP2 and PTP3are major tyrosine specific PTPases responsible for dephos-phorylation and inactivation of HOG1 (Wurgler-Murphy et al.,1997). Expression of genes encoding PTPases is often upregu-lated by the MAPK pathway, forming a negative feedback loopfor MAPK regulation (Jacoby et al., 1997; Wurgler-Murphyet al., 1997). Extrapolating from mammals, transient and low-level MAPK activation may contribute to stress tolerance inplants, whereas prolonged and high level activation may bedetrimental to the organism. Salt stress upregulated transientlythe AtPTP1 gene in Arabidopsis (Xu et al., 1998), which isdownregulated by cold stress. This implicates a unique mecha-nism for AtPTP1 in response to environmental stresses. AtPTP1

DROUGHT AND SALT TOLERANCE IN PLANTS 33
dephosphorylates AtMPK4 resulting in a complete loss of en-zyme activity (Huang et al., 2000). This is important, becauseboth AtPTP1 and ATMKP4 respond to salt stress (Xu et al.,1998).
The Arabidopsis mutant Atmkp1 is hypersensitive to geno-toxic stress conditions such as UV-C and MMS (Ulm et al.,2001). AtMKP1 codes for a dual-specificity kinase phosphatase.Disruption of Atmkp1 has no effect on the plant phenotype undernormal conditions, however, the mutant exhibits hypersensitiv-ity to stress caused by xenotoxic stresses but not to other abioticstresses. In wild-type plants, AtMKP1 is specifically activatedin response to UV and MMS. This is the only in vivo analysisof a phosphatase available up to now that has a role in stressconditions.
Protein phosphatases 2Cs are serine/threonine phosphatasesand their involvement in stress is well studied in fungal mod-els. In yeast PP2C interacts with a MAP kinase cascade thatcontrols osmolyte biosynthesis (HOG pathway). Several PP2Cs(Mpcs) from M. crystallinum have been isolated and studied fortheir tissue, developmental stage- and stress-specific responses.Out of ten PP2Cs, four transcripts, MPC2, MPC3, MPC5, andMPC8, are transiently increased by salt and dehydration, sug-gesting a role for these PP2Cs during stress (Miyazaki et al.,1999). Arabidopsis PP2Cs appear to be the largest protein phos-phatase family with 76 genes (reviewed in Schweighofer et al.,2004). Most of the research is focused on PP2C action in rela-tion to ABA signalling. PP2C has been described as abi1 andabi2 Arabidopsis mutants defective in a PP2C isoform (Leunget al., 1994), which seems to function as a negative regulatorin a pathway that mediates responses to environmental stressesinvolving ABA. However, the substrates of these PP2C in Ara-bidopsis are still unknown. Additional studies have shown thatguard cells of abi1-1 and abi2-1 plants are disrupted in ABA acti-vation of hyperpolarization-activated Ca2+(ICa) channels (Allenet al., 1999; Murata et al., 2001). Further, experiments on abi1-1 and abi2-1 mutants revealed both PP2Cs are acting at differ-ent levels of the same pathway, i.e., abi1-1 acts upstream andabi2-1 downstream of ABA-induced ROS production in guardcells (Murata et al., 2001). It was found that ABI1 can interactwith the ABA-inducible transcription factor ATHB6. ATHB6promoter-reporter expression was abrogated in abi1-1 mutant,suggesting that ABI1 acts upstream of the transcription factorATHB6 (Himmelbach et al., 2002).
Recently, the involvement of protein phosphatases in stom-atal regulation has been demonstrated using pharmacologicalapproaches. Application of phenylarsine oxide, a specific in-hibitor of protein tyrosine phosphatase prevented stomatal clo-sure in response to four stomatal closing signals such as ABA,H2O2, Ca2+ and dark (MacRobbie, 2002). This suggests thatprotein tyrosine dephosphorylation is involved mostly down-stream of the Ca2+ signalling which is responsible for stomatalclosure. Stomatal aperture regulation is dependent on ion ef-flux from guard cells. Identification of the target protein whosedephosphorylation results in activation of ion release from the
vacuole will allow us to connect the signalling events in responseto osmotic stress and ABA.
D. Phospholipid SignallingThe plasma membrane must play an important role in per-
ceiving and transmitting environmental signals. Osmotic stressoften leads to altered membrane fluidity and changes in phos-pholipids have recently been recognized as important eventsmediating osmotic stress signals in plants (Munnik and Meijer,2001). The current hypothesis is that phopspholipids are cleavedby phospholipases, which produce phospholipid-derived secondmessengers. In plants, like in other organisms four major classesof phospholipases are distinguished based on their cleavage site:phospholipase C (PLC), phospholipase D (PLD), and phospho-lipase A1 and A2 (PL A1 and PL A2) (Wang, 2002). Phospho-lipid signalling may be regulated through G-proteins and maybe tightly linked with calcium. The major phospholipid-derivedsignalling molecules which will be considered in the context ofosmotic stress are inositol 1,4,5-triphosphate (IP3 ), diacylglyc-erol (DAG) and phosphatidic acid (PA).
1. Inositol 1,4,5-Triphosphate (IP3)PLC cleaves the phospholipid phosphatidylinositol 4,5-
bisphosphate (PIP2) into the soluble IP3 and the membranebound DAG. Osmotic stress rapidly increases PIP2 synthesisin Arabidopsis (Pical et al., 1999; De Wald et al., 2001) and con-comitantly the transcript levels increase of PIP5K, aphosphatidylinositol-kinase, that synthesizes PIP2 (Mikamiet al., 1998). PIP2 is a signal molecule which in animal cellsleads to K+- desensitization (Kobrinsky et al., 2000). If thiscould also be confirmed for plant cells, then ion flux and osmoticstress can be linked via PIP2. There are reports from several plantspecies that osmotic stress leads to increased PLC transcript lev-els. One of the Arabidopsis phospholipase C genes (AtPLC1) isinduced by dehydration, salinity and low temperature (Hirayamaet al., 1995). Takahashi et al. (2001) have shown that the hyper-osmotic stress induces a rapid and transient elevation in IP3levels due to activation of PI-PLC in Arabidopsis cell cultures.Two genes, VuPLC1 and VuPLC2, were isolated from drought-tolerant and -sensitive varieties of cowpea. Whereas VuPLC1is constitutively expressed and decreases under drought stress,VuPLC2 transcript levels accumulate under water stress in thetolerant plant and progressively declines in the sensitive plant(El-Maarouf et al., 2001).
Involvement of PLC-genes in dehydration has also been ob-served in potato (Kopka et al., 1998). PLC activation leads tothe synthesis of IP3 and DAG. IP3 then releases Ca2+ from in-ternal stores (Sanders et al., 1999; Schroeder et al., 2001), whileDAG may be converted to PA or activates a protein kinase C,which, however, has not been isolated yet in plants. Hyperos-motic stress increased IP3 levels were reported in several stud-ies on osmotic stress (Takahashi et al., 2001; De Wald et al.,2001; Drøbak and Watkins, 2000). PI-PLC was also shown to

34 D. BARTELS AND R. SUNKAR
be involved in the regulation of stomatal movements. Drought-induced activation of PI-PLC led probably via IP3 to an increasein cytosolic Ca2+ in guard cells, which triggers stomatal closureand thus presents a drought avoidance mechanism (Staxen et al.,1999).
2. Phosphatidic Acid (PA)Phosphatidic acid (PA) is another second messenger in animal
cells that can activate PLC and protein kinase C (Munnik andMeijer, 2001). PA is synthesized through cleavage of membranephospholipids by PLD. The predicted structure of plant PLDsdiffer from that of yeast and animals in that most plant PLDscontain not only the conserved catalytic motifs, but also a Ca2+-binding domain (a C2 domain) which is not found in the PLDsof other organisms (Wang, 2001). This feature indicates a directregulation of PLD by calcium, in addition to changes in PLDgene expression.
Elevated PLD activity was found to be correlated with drought-stress in a drought-sensitive cowpea strain, when it was com-pared with a drought-tolerant strain (El-Maarouf et al., 1999).Further evidence was provided by Frank et al. (2000) by isolatingtwo phospholipase D cDNAs from C. plantagineum: CpPLD1is constitutively expressed and is likely to be involved in earlyresponses to dehydration, producing the second messenger phos-photidic acid to amplify the signal after its perception, while thedehydration-responsive CpPLD2 may be involved in phospho-lipid metabolism and membrane rearrangements at later stagesof dehydration. Four PLD genes were analyzed for their re-sponse to osmotic stress conditions in Arabidopsis (Katagiriet al., 2001).
Only the transcript levels of PLDδ were transiently elevatedin response to dehydration, whereas the expression was rapidlyinduced by high salt stress. Plants transformed with a PLDδ
promoter fused to a reporter gene showed strong induction un-der dehydration, mainly in vascular tissues of leaves, cotyle-dons and roots. Increased PLD activity coincides with the el-evation in phosphatidic acid levels in response to dehydration.Evidence to support that the elevated phosphatidic acid levelswere linked with stress-increased PLDδ activity was providedby antisense PLD transgenic plants, in which the dehydration-induced production of phosphatidic acid accumulation was sub-stantially reduced (Katagiri et al., 2001). In castor bean leavesPLDα mRNA and enzymatic activity increased by ABA treat-ment. Further, an increase was observed in the proportion ofenzyme activity associated with microsomal membranes (Ryuand Wang, 1995) implying both enzyme level and its locationare important factors in the ABA response. Like PLC, PLD hasa role in the regulation of the stomatal aperture during osmoticstress. ABA-promoted stomatal closure was shown to be me-diated by guard cell PLD activity in response to water stress(Jacob et al., 1999). Antisense suppression of PLDα impairsstomatal closure mediated by ABA or water stress and increaseswater loss in Arabidopsis, whereas the overexpression has theopposite effect and leads to a decreased water loss by enhancing
the sensitivity to ABA (Sang et al., 2001). These experimentsprovided direct evidence for PLDα in regulating water loss. Theinvolvement of PLD in ABA responses obtained further supportfrom experiments using rice protoplasts. The expression of sev-eral ABA-related genes was repressed when PLD-derived PAwas blocked, which indicates that PLD activity is important forABA-inducible gene expression (Gampala et al., 2001; 2002).Osmotic stress activated PLDs implies a role for PA that is pro-duced by PLD. The targets of PA in plants are unknown, however,PIP kinase, PDK (phosphoinositide dependent kinase), MAPKpathway, MGDG synthetase, K+ channel are possible targets(Munnik, 2001).
E. Other Signalling Molecules1. Salicylic Acid
It has been established for quite some time that salicylic acid(SA) plays an important role in the defense response in manyplant species to pathogen attack. SA mediates the oxidative burstthat leads to cell death in the hypersensitive response, and actsas a signal for the development of the systemic acquired resis-tance (Shirasu et al., 1997). Recently the involvement of SA inosmotic stress was demonstrated by using an SA-deficient trans-genic line expressing a salicylate hydroxylase (NahG). Wild typeseeds germinated in the presence of NaCl or mannitol showedextensive necrosis in the shoot, but not in the NahG mutant. Wildtype and NahG behaved similarly during germination at variousLi+ concentrations, excluding the possible involvement of ioniccomponents. Greater oxidative damage occurred in wild-typeseedlings compared with NahG seedlings under NaCl stress.Methyl viologen treatment resulted in necrotic phenotypes onlyin wild-type plants. These different observations lend support tothe hypothesis that SA potentiates the effects of salt and osmoticstress by enhancing the generation of ROS during photosynthe-sis (Borsani et al., 2001).
2. Nitric Oxide (NO)Recent studies revealed that NO is an important signalling
molecule involved in several physiological functions rangingfrom plant development to defense responses (Wendehenne et al.,2001). Nitric oxide is a labile free radical that is produced fromL-arginine by NO synthase in various mammalian cells, whereit has been shown to be protective against damages caused byoxidative stress conditions. Exogenous application of NO im-proved water stress tolerance both in wheat seedlings and indetached leaves (Mata and Lamattina, 2001). Detached leavespretreated with the NO releasing agent SNP withstand the im-posed stress by retaining a higher water content, lower ion leak-age, and greater accumulation of LEA 3 transcripts compared tocontrol leaves. The drought tolerance of NO could be attributedto its ability to maintain higher RWC, decreasing the rate oftranspiration by closing stomata and the ability to induce geneexpression involved in stress tolerance.

DROUGHT AND SALT TOLERANCE IN PLANTS 35
V. CALCIUM SIGNALLING DURING DEHYDRATIONAND SALT STRESS
In plant cells, calcium functions as a second messenger cou-pling a wide range of extracellular stimuli to intracellularresponses (Snedden and Fromm, 1998, 2001). Different ex-tracellular stimuli elicit specific calcium signatures: kinetics,amplitude and duration of Ca2+ transients specify the natureand the intensity of stimulus. To date, three major classes ofCa2+ sensors have been characterized in plants. These classesare calmodulin, CDPKs (calcium-dependent protein kinase) andCBLs (calcineurin B-like proteins) (Yang and Poovaiah, 2003).Several lines of evidence suggest that all these three classes ofCa2+ sensors are involved in stress signal transduction (Sneddenand Fromm, 2001; Luan et al., 2002; Zhu, 2000).
The involvement of Ca2+ signalling in response to osmoticand ionic stress is well documented. NaCl causes a rapid andtransient increase in cytosolic calcium, that in turn triggers manysignal transduction pathways, including the regulation of enzy-matic activity, ion channel activity, and gene expression whichresults in diverse cellular responses (Snedden and Fromm, 1998,2001) and mediates salt adaptation (Bressan et al., 1998; Liuand Zhu, 1998; Serrano et al., 1999). Recent progress providedinsights into how the response to different osmotic stresses is en-coded in the spatial and temporal dynamics of the Ca2+ signal(Knight and Knight, 2001). The role of calcium and its dynam-ics were investigated using either pharmacological approachesor using transgenic plants which express the calcium reporterprotein aequorin in different cellular compartments or under thecontrol of promoters with different responsiveness to environ-mental stimuli. This has allowed determining calcium fluxeswithin one cell and in different tissues. For instance, mannnitoland sodium chloride-induced increased cytosolic [Ca2+] is dueto release of calcium from the vacuole (Knight et al., 1997).Cell-type specific changes in cytosolic calcium levels were ob-served in Arabidopsis root cells in response to drought, salinity,and low temperature (Kiegle et al., 2000).
A. Calcium-Dependent Protein Kinases (CDPKs)Osmotic stress-induced CDPKs have been reported from sev-
eral plants (Kawasaki et al., 2001; Seki et al., 2002; Ozturk et al.,2002). Recent experiments on salt-tolerant and salt-sensitive ricevarieties have strengthened the importance of CDPKs in osmoticstress responses. A specific CDPK is induced earlier and its ex-pression is sustained for longer duration in the tolerant varietycompared to the sensitive variety (Kawasaki et al., 2001). Furtherevidence for the involvement of CDPKs in stress was obtainedfrom studying Arabidopsis CDPKs (Sheen, 1996). Out of sev-eral CDPKs tested, only AtCDPK1 and AtCDPK1a were ableto transcriptionally activate selected reporter genes indicatingthat specific CDPK isoforms mediate the effects of stress. Inter-estingly, in the salt-tolerant Mesembryanthemum crystallinumosmotic stress-induced CDPK (McCDPK1) was shown to inter-act with CSP1 (calcium-dependent protein kinase Substarte Pro-
tein1), which is a transcription factor belonging to a class of two-component pseudoresponse regulators (Patharkar and Cushman,2000). These results establish a role for specific CDPKs in stress-induced gene expression.
B. Calcium-Binding ProteinsModulation of intracellular calcium levels is partly regulated
by calcium-binding proteins such as calmodulin, which is ac-tivated by increased calcium concentrations and then inducesspecific kinases. The importance of the calcium-binding pro-teins has first been derived from yeast mutant analysis. Muta-tions in calmodulin genes of yeast render the yeast cells sensi-tive to high NaCl concentrations and NaCl-induced genes areno longer induced, suggesting that calmodulin is involved inthe NaCl-stress signal transduction pathway (Cunningham andFink, 1996). The regulation of calcium-binding proteins can oc-cur in a cell- or tissue-specific manner as the following examplesillustrate. Ca2+/calmodulin dependent kinases have been shownto be regulated by salt stress, e.g., PsCCaMK, a Ca2+/calmodulindependent protein kinase in pea was specifically upregulated byNaCl in roots, while the shoot kinase was not affected (Pandeyet al., 2002). A family of calmodulin binding transcription acti-vators were first discovered in drought-stressed Brassica napusand were subsequently shown to be only present in multicellularorganisms (Bouche et al., 2002). Further studies are needed todetermine the importance of this family during osmotic stress.
Other examples for osmotic stress-activated calcium-bindingproteins are the Arabidopsis protein AtCP1, the membrane-associated rice protein OsEFA27, and the Arabidopsis counter-part RD20 (Frandsen et al., 1996; Jang et al., 1998; Takahashiet al., 2001). A recently identified calmodulin binding proteinin Arabidopsis, AtCaMBP25, has been implicated as a negativeregulator of osmotic stress. AtCaMBP25 is upregulated by os-motic stress and its overexpression leads to sensitivity to osmoticor salt stresses, whereas the suppression by antisense technologyresulted in improved tolerance (Perruc et al., 2004) suggestingthat AtCaMBP25 is a negative regulator of osmotic and saltstresses. Further, microarray analysis in these plants will revealif there is a correlation between the observed phenotypes andaltered gene expression.
C. Ca2+-Mediated SOS Pathways Are Involvedin Ion Homeostasis
Insight into the underlying mechanisms of external calciumon cellular responses to salt has been provided by the isolation ofsalt hypersensitive mutants sos1, sos2 and sos3 from Arabidopsisthaliana (Zhu, 2003). Cloning and characterization of SOS geneshas led to the discovery of a novel Ca-dependent pathway forthe regulation of ion homeostasis and plant salt tolerance (seelater in Na+ extrusion).
D. Calcineurin B (CBLS) and Osmotic Stress ResponsesDespite the genetic analysis of the interaction of the SOS 1
to 3 genes, the complexity of the calcium signals requires a lot

36 D. BARTELS AND R. SUNKAR
more mutant analysis which becomes apparent by the fact thatArabidopsis thaliana has at least 10 calcineuron B-like (CBL)genes encoding highly similar but functionally distinct Ca2+-binding proteins (Kudla et al., 1999). Drought, high salt, coldand wound signals induce AtCBL1 gene transcripts (Kudla et al.,1999). Both CBL1 and CBL2 respond to light, but CBL2 lacksthe other responses of CBL1 (Nozawa et al., 2001). Knockoutmutant plants of SCaBP5/CBL1 displayed hypersensitivity todrought and salinity, in contrast to CBL1 overexpressing plantsthat showed drought tolerance (Albrecht et al., 2003). Such ex-pression patterns suggest that CBL1 and CBL2 have both over-lapping and specific functions.
E. Ca2+ ATPasesThe major physiological role of Ca2+-ATPases is to restore
and maintain homeostasis by pumping Ca2+ out of the cytosolto terminate a signalling event, which is critical in all eukaryoticcells (Sze et al., 2000). Plant and animal cells utilize two distincttypes of Ca2+ pumps, identified as type IIA and type IIB, basedon their protein sequences. NaCl stress has been reported to in-duce the expression of genes encoding the type IIA Ca2+ pumpin tobacco, tomato, and soybean (Wimmers et al., 1992; Chunget al., 2000). The consequence of upregulating the Ca2+ pumpin response to NaCl is not known, but it was speculated that itis likely to provide an adaptive response such that a stimulatedcell would acquire an enhanced efflux capacity capable of sub-sequently decreasing the magnitude or duration of a calcium re-lease to further exposure to a given stimulus (Chung et al., 2000).The soybean Ca2+-ATPase1 was induced by NaCl but not byKCl and mannitol, indicating that specific calcium signals trig-ger an increase in transcription (Chung et al., 2000). Geisler et al.(2000a) have cloned and characterized Ca2+-ATPase isoform 4(ACA4), a calmodulin-regulated Ca2+-ATPase from Arabidop-sis. Two lines of evidence suggest that ACA4 might be part ofthe Ca2+-dependent signal transduction pathway linked to saltstress: (1) Arabidopsis seedlings treated with different concen-trations of NaCl for 24 h showed a dose-dependent increase inACA4 transcript levels, (2) when N-terminal truncated ACA4was expressed in yeast, it conferred increased NaCl tolerance toits host (Geisler et al., 2000b).
VI. TRANSCRIPTIONAL REGULATIONOF GENE EXPRESSION
Stress responses primarily include transcriptional regulationof gene expression and this depends on the interaction of tran-scription factors with cis-regulatory sequences. In several in-stances, the quantity and availability of regulatory proteins maydepend on their own expression patterns. Such autocatalytic con-trols may be exerted on the transcriptional, post-transcriptionalor translational level. Phosphorylation of regulatory proteins isa major event in controlling the gene expression in eukary-otes. Therefore, multiple protein-protein and/or protein-DNAinteractions frequently determine the rate of transcription by
activation/repression of a promoter under given environmentalconditions.
Although the cis-elements of dehydration and ABA-responsivegenes have been intensively studied, our understanding is lim-ited. More efforts are still needed to identify additional novelcis- and trans-acting elements that function in ABA-dependentas well as ABA-independent gene expression. Presently, twoclasses of drought-responsive DNA elements have been wellcharacterized: the ABA-responsive element (ABRE) and thedehydration responsive element (DRE, also referred to as C-repeat; Baker et al., 1994). It is highly likely that yet unidentifiedosmotic-stress responsive cis-elements exist in plants.
A. ABA Response Elements (ABREs)Most, but not all of the dehydration-induced genes are also in-
duced by the application of the phytohormone ABA(Chandler and Robertson, 1994; Leung and Giraudat, 1998) (fordetails see section XI). Until now, two ABA-dependent pathwaysare known to mediate gene expression in plants during osmoticstress. The distinction is largely based on cis-elements that existin the promoters of ABA-inducible genes. The ABA-dependentpathways are thought to mediate the gene expression throughan ABRE-element and b-ZIP transcription factors (Busk andPages, 1998), while the other pathway is through a MYC andMYB elements and transcription factors (Yamaguchi-Shinozakiand Shinozaki, 1993).
Many cis-regulatory elements known as ABA-responsive ele-ments (ABREs) have been identified. Among them, (C/T)ACGTG(G/T)C motifs have been reported to function as ABREsin many genes (Guiltinan et al., 1990; Mundy et al., 1990; In-gram and Bartels, 1996; Busk and Pages, 1998). The core el-ement of these ABREs is the CACGTG motif also known asG-box motif, which functions in the regulation of plant genesstimulated by a variety of environmental signals. SystematicDNA-binding studies have shown that nucleotides flanking theACGT core specify the DNA-binding interactions and subse-quent gene activation (Williams et al., 1992; Izawa et al., 1993).Nevertheless, in promoters such as CDeT27-45 and CDeT6-19,isolated from C. plantagineum, G-box related ABREs do not ap-pear to be major determinants of the ABA or drought response(Michel et al., 1993, 1994).
Ho and coworkers identified cis-elements called couplingelements which are active in combination with an ABRE butnot alone (Shen et al., 1996). In the promoters of the barleygenes HVA22 and HVA1 the coupling elements CE1 and CE3(ACGCGTGTCCTC) are necessary for activation by ABA. Dis-section of these promoters defined ABA response complexes(ABRC) consisting of a coupling element and an ABRE capa-ble of conferring ABA-inducible transcription. Other ABREsthat do not belong to the above groups have also been reported:a Sph element-containing sequence (CGTGTCGTCCATGCAT)of the maize C1gene (Kao et al., 1996), the MYB and the MYCbinding sites of the Arabidopsis rd22 gene (Abe et al., 1997),

DROUGHT AND SALT TOLERANCE IN PLANTS 37
and a novel element present in the CDeT27-45 gene of C. plan-tagineum (Michel et al., 1993).
B. The Dehydration Response Element (DRE)The investigation of dehydration-induced genes in Arabidop-
sis has also revealed ABA-independent signal transduction path-ways (Shinozaki and Yamaguchi-Shinozaki, 2000). In aba (ABAdeficient) and abi (ABA insensitive) mutants, several genes areinduced by dehydration indicating that these genes do not re-quire ABA for their expression under drought conditions. TheA. thaliana genes rd29A (also described as COR 78 or LTI78) andrd29B are differentially induced under conditions of dehydra-tion, salt, cold stress, or ABA treatment. This multiple expressionpattern requires at least two cis-acting elements for the rd29Agene. The 9-base pair direct repeat (TACCGACAT), termed thedehydration responsive element (DRE), functions in the initialrapid response of rd29A to dehydration, salt, or low temper-ature. The slower ABA response is mediated by another pro-moter fragment that contains an ABRE (Yamaguchi-Shinozakiand Shinozaki, 1993, 1994). The DRE is an essential cis-actingelement for the regulation of rd29A induction in the ABA-independent response to dehydration in Arabidopsis. DRE-related motifs have been reported in promoters of several genesregulated by osmotic and low temperature stress, including kin1,cor6.6/kin2, and rd17/cor47 in Arabidopsis (Wang et al., 1995;Iwasaki et al., 1997). A similar motif was also reported in the pro-moter region of cold and dehydration-inducible cor15A (Bakeret al., 1994). In a recent study 16 genes containing DRE (TAC-CGACAT) or DRE-related core motif (CCGAC) were identifiedin the promoters of drought stress inducible Arabidopsis genes,which are likely targets of DREB1 or DREB2 transcription fac-tors (see below) (Seki et al., 2002). In another microarray ex-periment which analysed DREB1A overexpressing plants Sekiet al. (2001) have identified 12 target genes of DREB. Amongthem, 11 promoters contain the CCGAC sequence motif, and6 promoters contain ABRE elements (Seki et al., 2001), whichimplies additional unidentified stress responsive cis-elements.
C. Transcription Factors Modulated by Osmotic StressPlants need a large number of transcription factors governing
proper and strict transcriptional regulation in response to devel-opmental and environmental cues. Indeed, over 5 percent of theArabidopsis genome is devoted to encoding more than 1,500transcription factors (Riechmann et al., 2000). Despite the factthat not many stress-specific consensus sequences were identi-fied in promoters of stress specific genes transcription factorsbelonging to different families have been shown to be modi-fied by stress. To activate or repress transcription, transcriptionfactors must be located in the nucleus, bind DNA, and interactwith the basal transcription apparatus. Therefore, environmentalsignals that regulate transcription factor activity may affect anyone or a combination of these processes. Regulation of a tran-scription factor is achieved by reversible phosphorylation or by
de novo synthesis of transcription factors. The current analysisof identified transcription factors suggests that different stresssignalling pathways may overlap or converge at specific points.
1. Basic Region Leucine Zipper (bZIP) ProteinsBasic region leucine zipper (bZIP) proteins contain a DNA-
binding domain rich in basic residues and adjacent to a leucinezipper dimerization domain. bZIPs are a large family of tran-scription factors in plants and are represented by 75 membersin Arabidopsis (Jacoby et al., 2002). All plant bZIP proteinsbind to an ACGT core sequence, but the sequences flanking thecore sequence affect the precise DNA binding. Several bZIPfactors that bind ABREs have been cloned as candidates forABA-responsive transcription factors that induce gene expres-sion by osmotic stress and/or ABA. One bZIP subfamily hasbeen linked genetically to an ABA response: ABI5 and its ho-mologs, the ABRE binding factors (ABFs/AREBs). ABRE bind-ing factors (ABFs)/ABA-responsive element binding (AREBs)proteins respond at the transcriptional and post-transcriptionallevel to dehydration and salt stress (Choi et al., 2000; Uno et al.,2000). ABF3 and ABF4 overexpression in transgenic Arabidop-sis plants resulted in enhanced drought tolerance accompaniedby decreased transpiration, suggesting that ABF3 and ABF4 areinvolved in stomatal closure mediated by ABA (Kang et al.,2002). Promoters of both ABF3 and ABF4 were found to bemost active in roots and guard cells, consistent with their rolesin stomatal regulation and water stress response.
2. Homeodomain-Leucine Zipper Proteins (HD-ZIP)HD-ZIP genes encode proteins that have only been identi-
fied in plants so far and are thought to regulate developmentalprocesses and responses to environmental cues. HD-ZIP pro-teins are characterized by the presence of a DNA-binding home-odomain with a closely linked leucine zipper motif. The activityof HD-ZIP resides primarily in the specific DNA-binding prop-erty of the homeodomain and the ability of the leucine zipperto mediate protein-protein interaction with other HD-ZIPs. HD-ZIPs have been described for several plant species such as Ara-bidopsis, carrot, tomato, rice, sunflower, and C. plantagineum.Seven HD-ZIP are affected by dehydration in C. plantagineum,which are placed in different branches of the signalling network(Deng et al., 2002).
In Arabidopsis three HD-ZIP genes (ATHB-6, ATHB-7, andATHB-12) are shown to be responsive to dehydration (Sodermanet al., 1996, 1999; Lee and Chun, 1998). Hahb-4 is upregu-lated by drought and ABA in sunflower roots, stems, and leaves(Gago et al., 2002). HD-ZIP proteins can form both homo- andheterodimers, which gives many degrees of freedom in regula-tion processes. Direct targets of HD-ZIP genes have still to beidentified.
3. Zn-Finger ProteinsA zinc-finger motif represents the sequence in which cys-
teines and/or histidines coordinate a zinc atom(s) to form local

38 D. BARTELS AND R. SUNKAR
peptide structures that are required for their specific functions.The zinc-finger motifs are classified based on the arrangementof the zinc-binding amino acids. They play critical roles in in-teractions with other molecules. The cDNA clone for Alfin1 wasisolated by differential screening of salt-tolerant alfalfa cellsgrown on NaCl (Winicov, 1993). Alfin1 cDNA encodes a novelmember of putative Zn-finger proteins, which binds to promotersequences of MsPRP2 in vitro (Bastola et al., 1998). Overex-pression of the Alfin1 gene in transgenic plants increased theMsPRP2 transcript levels in root and enhanced NaCl tolerance,indicating that the Alfin1 gene product regulates MsPRP2 ex-pression in vivo (Winicov and Bastola, 1999). The function ofMSPRP2 is yet unknown, however, it was predicted that it islikely to be a cell wall protein with potential as an anchor to themembrane.
4. AP2/ERF-Type Transcription FactorsThe group of AP2/ERF DNA binding proteins were iden-
tified in different scientific contexts and the first proteins de-scribed gave this group its name. AP2/ERF domain proteinsinclude the DREB or CBF proteins binding to dehydration re-sponse elements (DRE) or C-repeats. The Arabidopsis genomeencodes 145 DREB/ERF-related proteins (Sakuma et al., 2002).The Arabidopsis AP2 proteins have been classified into fiveclasses based on similarities in their DNA-binding domains:AP2 subfamily (14 genes), RAV (related to ABI3/VP1) sub-family (6 genes), DREB subfamily (55 genes; Group A), ERFsubfamily (65 genes, Group B), and others (4 genes) (Sakumaet al., 2002). A major transcriptional regulatory system is repre-sented by DRE/C-repeat promoter sequences in stress-activatedgenes and DREBs/CBF factors that control stress gene expres-sion (Stockinger et al., 1997; Liu et al., 1998). A C-repeatbinding factor, CBF1, was isolated by screening an ArabidopsiscDNA expression library using the C-repeat of the cold stressCOR15a gene as target (Stockinger et al., 1997). The samefactor and other CBF family members, referred to as DREBs(Dehydration Responsive Element Binding proteins) have beenreported from studies of dehydration stress genes (Liu et al.,1998). A detailed expression analysis showed that these factorsmay be associated with different physiological conditions. Forexample, expression of DREB1A, B, C/CBF1, 2, 3 is inducedby low temperature, DREB1A, DREB1D/CBF4, DREB2A andDREB2B are induced by salt and dehydration, DREB1F is in-duced by salt, whereas, DREB1E is induced by ABA (Liu et al.,1998; Shinwari et al., 1998; Nakashima et al., 2000; Haakeet al., 2002; Sakuma et al., 2002). Several stress-inducible genessuch as rd29A, Cor6.6, Cor15a and Kin1 are target genes ofDREBs/CBFs in Arabidopsis and contain DRE/C-repeat se-quences in their promoters. Seki et al. (2001) have identified sixnew genes containing both DRE/C-repeat and ABRE motifs intheir promoters, implying complex regulation of stress-inducedgenes by ABA-dependent and ABA-independent pathways. Theemerging picture from functional studies of CBFs/DREBs indi-cates that CBF genes coordinate both, activation or repression
of stress responsive genes. Ectopic overexpression of CBF1/DREB1B and CBF3/DREB1A in Arabidopsis results in the con-stitutive expression of downstream stress-inducible genes andenhances freezing tolerance and dehydration/salt tolerances, re-spectively (Kasuga et al., 1999; Gilmour et al., 2000). On theother hand, the functional analysis of cbf2/dreb1c knock-out mu-tant plants revealed somewhat surprising results. Cbf2 mutantplants displayed tolerant phenotype to dehydration and salin-ity (Novillo et al., 2004). Another interesting feature of thesemutant plants is enhanced expression of CBF1/DREB1B andCBF3/DREB1A and the downstream genes such as LTI78, KIN1,COR47, and COR15A which are known to impart stress toler-ance. These results imply that CBF2 is a negative regulator ofCBF1 and CBF3 transcription factors and of the correspondingdownstream genes.
Rice AP2 transcription activators, OsDREB1A, OsDREB1B,OsDREB1C, OsDREB1D and OsDREB2A have been isolated.OsDREB2A was induced by dehydration and high salinity stress.Overexpression of OsDREB1A in transgenic Arabidopsis im-proved stress tolerance (Dubouzet et al., 2003). Two novel DREbinding factors (DBF1 and DBF2) were isolated from maize(Kizis and Pages, 2002). DBF1 and DBF2 are capable of bind-ing DRE motifs and are not homologues of DREBs from Ara-bidopsis. DBF1 is induced by dehydration, salinity, and ABAwhereas DBF2 is constitutively expressed at low levels and notresponsive to stress.
The Tsi1 (tobacco stress-inducible gene 1) from tobacco be-longs to the ERF subfamily (Sakuma et al., 2002) and is inducedby NaCl, salicyclic acid, and ethylene. Tsi1 was able to bind GCCmotifs in both ethylene response elements and DRE/CRT se-quences. Tobacco plants overexpressing Tsi1 showed enhancedresistance to osmotic stress and pathogen attack (Tsugane et al.,1999). This demonstrates that overexpression of a single tran-scription factor effects both biotic and abiotic stress responsessuggesting convergence of both signalling cascades.
5. Myb-Like ProteinsThe Myb-motif comprises three imperfect repeats forming a
helix-turn-helix–related motif. Each repeat contains three con-served tryptophan residues every 18 to 19 amino acids, whichpromotes a secondary structure with a functional Myb-domain.In plants, the first tryptophan of R3 is substituted by phenylala-nine or isoleucine. This latter amino acid is present in Atmyb2 inArabidopsis and myb-related genes from C. plantagineum cp-MYB7 and cpMYB10 whose expression is upregulated by dehy-dration and ABA (Urao et al., 1993; Iturriga et al., 1996). Ectopicexpression of CpMYB10 in transgenic Arabidopsis plants re-sulted in drought and salinity tolerance (Villalobos et al., 2004).The Myb gene family is represented by 190 genes in Arabidop-sis (Riechmann et al., 2000). Atmyb-2 has been identified as apositive regulator of stress-induced rd22 gene expression (Abeet al., 1997). Consistent with the role, AtMYB-2 overexpress-ing transgenic plants exhibited hypersensitivity whereas knock-out mutants showed insensitivity to ABA (Abe et al., 2003). In

DROUGHT AND SALT TOLERANCE IN PLANTS 39
addition, three additional Arabidopsis Myb-like transcriptionfactors were identified as upregulated by dehydration, high salin-ity, or cold stress in a microarray study (Seki et al., 2002).
6. Myc-Like ProteinsMyc-like proteins contain the basic helix-loop-helix (bHLH)
domain, which is composed of two subdomains: the basic re-gion (as found in bZIPs) responsible for DNA binding and theHLH (helix-loop-helix) region for dimerization with interactingproteins. The Arabidopsis rd22BP1 gene encodes a Myc-liketranscription factor and is induced by dehydration, high saltconditions, and ABA. It activates the downstream rd22 geneby binding to the myc promoter motif (Abe et al., 1997). Theexpression of rd22BP1 (AtMYC-2) precedes the rd22 gene ex-pression, which underlines the hierarchic relationship. Overex-pression and knock-out mutants displayed contrasting pheno-types with respect to ABA sensitivity, similar to AtMYb-2 (Abeet al., 2003). Hypersensitivity to ABA was enhanced in trans-genics when both genes are overexpressed together (Abe et al.,2003), confirming the cooperation of AtMYB-2 and AtMYC-2interaction in vivo in stress-activated gene expression.
7. CDT-1A novel gene CDT-1 was isolated from C. plantagineum us-
ing a T-DNA activation tagging approach. A gain-of-functionphenotype in C. plantagineum calli was due to the activationof the CDT-1 gene that confers desiccation tolerance to calluscells and activates stress genes independent of exogenous ABA(Furini et al., 1997). However, it is not known whether CDT-1functions as RNA or as a peptide; recent experiments favor RNAas functional molecule (Furini and Bartels, unpublished). Thereis no direct sequence homologue to CDT-1 in Arabidopsis.
VII. ACCUMULATION OF SUGARSAND COMPATIBLE SOLUTES
Almost all organisms, ranging from microbes to animalsand plants, synthesize compatible solutes in response to os-motic stress (Burg et al., 1996). Compatible solutes are non-toxic molecules such as amino acids, glycine betaine, sugars, orsugar alcohols. They do not interfere with normal metabolismand accumulate predominantly in the cytoplasm at high con-centrations under osmotic stress (reviewed in Chen and Murata,2002). These molecules may have a primary role of turgor main-tenance but they may also be involved in stabilizing proteinsand cell structures (Yancey et al., 1982). Initially it was thoughtthat compatible solutes have their main role in osmotic adjust-ment, but there is increasing discussion of other roles (Serraj andSinclair, 2002). The accumulation of these solutes per se maynot be important for osmotic stress tolerance but the metabolicpathways may have adaptive value (Hasegawa et al., 2000). Afurther hypothesis is that compatible solutes are also involved inscavenging reactive oxygen species (Shen et al., 1997a, b; Honget al., 2000; Akashi et al., 2001; Chen and Murata, 2002). Engi-
neering the synthesis of compatible solutes has been a relativelysuccessful approach to obtain stress tolerant plants (see Table 1).
A. SugarsSeveral physiological studies suggested that under stress con-
ditions nonstructural carbohydrates (sucrose, hexoses, and sugaralcohols) accumulate although to varying degree in differentplant species. A strong correlation between sugar accumulationand osmotic stress tolerance has been widely reported, includingtransgenic experiments (Ramanjulu et al., 1994a; Abd-El Bakiet al., 2000; Gilmour et al., 2000; Streeter et al., 2001; Taji et al.,2002).
The increase in sugars mostly results in increased starch hy-drolysis, which requires activities of hydrolytic enzymes. Resur-rection plants and seeds of many higher plants are good examplesfor a link of carbohydrate (in particular sucrose) accumulationand the acquisition of stress tolerance (Hoekstra et al., 2001;Phillips et al., 2002). This is illustrated with the example ofthe resurrection plant C. plantagineum, which contains a highamount of the unusual sugar octulose (an 8-carbon sugar), whichis rapidly converted into sucrose during dehydration (Bianchiet al., 1991). This sugar conversion is coupled with the in-creased expression of sucrose synthase and sucrose phosphatesynthase genes (Ingram et al., 1997). The currant hypothesis isthat sugars either act as osmotica and/or protect specific macro-molecules and contribute to the stabilization of membrane struc-tures. Sugars may protect cells during desiccation by formingglasses (Black and Pritchard, 2002). Sugars are also thought tointeract with polar headgroups of phospholipids in membranesso that membrane fusion is prevented. It is unknown whethersugars fulfil this function on their own or in conjunction withother molecules such as LEA proteins. Many seeds accumulateconsiderable amounts of raffinoase-type oligosaccarides (RFOs)such as raffinose and stachyose which are thought to play a rolein the acquisition of desiccation tolerance. Despite many studiesthe link between the presence of these carbohydrates and des-iccation tolerance has not always been confirmed. Galactinolsynthase catalyzes the first committed step in the biosynthesisof RFOs. Overexpression of galactinol synthese resulted in accu-mulation of galactinol and raffinose under controlled conditionsand improved drought tolerance (Taji et al., 2002), confirmingthe importance of these sugars under stress conditions.
A sugar which has been shown to contribute to desiccationtolerance in yeast and some nematodes is trehalose. In higherplants substantial amounts of trehalose were identified in tworesurrection plants Myrothamnus flabellifolia and Sporobolusstapfianus (Phillips et al., 2002). It has been reported that manyhigher plants possess trehalase activity, which is perhaps re-sponsible for rapid degradation of any trehalose synthesized.Arabidopsis thaliana has at least one gene which encodestrehalose-6-phosphate phosphatase which is required for tre-halose synthesis, but the physiological role of this enzyme is notclear (Vogel et al., 1998).

40 D. BARTELS AND R. SUNKAR
B. CyclitolsAccumulation of cyclic polyols such as D-pinitol (1D-3-
O-methyl-chiro-inositol) or D-ononitol (1D-4-O-methyl-myo-inositol) has frequently been reported in response to droughtand salinity (Vernon and Bohnert, 1992; Streeter et al., 2001).Pinitol may accumulate in chloroplasts, which is consistent withthe positive correlation between cyclitol accumulation and CO2
assimilation under drought (Sheveleva et al., 1997). Direct evi-dence for a role of cyclitols has been provided by transgenic to-bacco plants that accumulated ononitol which resulted in droughtand salt stress tolerance (Sheveleva et al., 1997).
C. ProlineProline is probably the most widely distributed osmolyte,
and it occurs not only in plants but also in many other organisms(McCue and Hanson, 1990; Delauney, 1993). Besides osmoticadjustment other roles have been proposed for proline in os-motically stressed plant tissues: protection of plasma membraneintegrity (Mansour et al., 1998), a sink of energy or reducingpower (Verbruggen et al., 1996), a source for carbon and nitrogen(Ahmad and Hellebust, 1988; Peng et al., 1996), or hydroxyl rad-ical scavenger (Smirnoff and Cumbes, 1989; Hong et al., 2000).
Proline accumulation can occur via two biosynthetic path-ways in plants: the ornithine-dependent pathway and the glutamate-dependent pathway. Proline biosynthesis from glutamate ap-pears to be the predominant pathway, especially under stressconditions (Delauney and Verma, 1993; Delauney et al., 1993).L-proline is synthesized from L-glutamic acid via �1-pyrroline-5-carboxylate (P5C). This reaction is catalyzed by two enzymes,P5C synthetase (P5CS) and P5C reductase (P5CR). Genes en-coding the enzymes involved in proline metabolism have beenisolated from several plant species (Yoshiba et al., 1997). Thesecond proline biosynthesis pathway involves transaminationof ornithine and is catalyzed by ornithine-δ-aminotransferase(OAT) yielding two possible intermediates P5C (�-pyrroline-2-carboxylate) and P2C (�-pyrroline-2-carboxylate), which canboth be reduced to proline (Mestichelli et al., 1979). There areindications from Arabidopsis that the ornithine pathway oper-ates mainly in young seedlings (Roosens et al., 1998). The otherimportant process that controls proline levels is oxidation ofL-proline by proline dehydrogenase (ProDH) to P5C, which isconverted to L-glutamic acid by P5C dehydrogenase.
Direct evidence for the role of proline during osmotic stresshas been provided by transgenic approaches. Different strategieswere used to manipulate proline biosynthesis including over-expression of P5CS in tobacco, rice and Arabidopsis plants,overexpression of OAT, expression of a feedback inhibition-insensitive form of P5CS, and antisense suppression of prolineoxidation by ProDH (Kavi Kishor et al., 1995; Zhu et al., 1998;Nanjo et al., 1999; Hong et al., 2000; Roosens et al., 2002).All approaches resulted in elevated proline pools and improvedosmotic stress tolerance. In contrast to these observations is areport about antisense ProDH transgenic Arabidopsis plants,
which accumulated proline but showed no change in osmoticstress tolerance (Mani et al., 2002).
D. Glycine BetaineGlycine betaine is another extensively studied compatible so-
lute. Glycine betaine is thought to protect the plant by maintaningthe water balance between the plant cell and the environment andby stabilizing macromolecules (reviewed in Chen and Murata,2002; Rontein et al., 2002). Plants synthesize glycine betaine viaa two-step oxidation of choline: Choline → betaine aldehyde →glycine betaine (Rhodes and Hanson, 1993). The first reactionis catalyzed by a ferredoxin-dependent choline monooxygenase(CMO) and the second step by a NAD+-dependent betaine alde-hyde dehydrogenase (BADH) (Chen and Murata, 2002; Ronteinet al., 2002). Glycine betaine accumulation is associated withupregulated CMO and BADH gene expression concomitantlyleading to elevated enzymatic activity.
Glycine betaine accumulation marginally improves osmoticstress tolerance in transgenic plants (Hayashi et al., 1997). Thelevels of glycine betaine thus far obtained by engineering arelow, and the increments in stress tolerance are small (Nuccioet al., 1999). The major factors that limit the accumulation ofglycine betaine are the available choline as the substrate forthe reaction and its transport from the chloroplast (where it issynthesized) to the cytosol (Nuccio et al., 1998, 2000; McNeilet al., 2000; Huang et al., 2000; Chen and Murata, 2002; Ronteinet al., 2002).
VIII. PROTECTIVE PROTEINS AND OTHER PATHWAYSINVOLVED IN STRESS ADAPTATION
A. Late Embryogenesis-Abundant (LEA) ProteinsLea genes encode a diverse group of stress-protection pro-
teins expressed during embryo maturation in all angiospermsincluding orthodox and recalcitrant seeds. LEA proteins werefirst identified and characterized in cotton and represents thedominant protein and mRNA species in mature embryos (Dureet al., 1981; Galau et al., 1986). Accumulation of LEA pro-teins during embryogenesis correlates with increased levels ofABA and with acquisition of desiccation tolerance (Galau et al.,1986). ABA treatment of embryos isolated from an early de-velopmental stage results in a precocious accumulation of LEAproteins and the acquisition of desiccation tolerance indicatinga connection between these parameters (Bartels et al., 1988).LEA proteins are not normally expressed in vegetative tissues,but they are induced by osmotic stress or exogenous applicationof ABA (Ingram and Bartels, 1996). LEA proteins comprise thevast majority of stress-responsive proteins. Evidence derivedfrom expression profiles strongly supports a role for LEA pro-teins as protective molecules, which enable the cells to surviveprotoplasmic water depletion (Ingram and Bartels, 1996). Thisis corroborated by the finding that a lea transcript is expressedin response to dehydration in the nematode Aphelenchus avenae

DROUGHT AND SALT TOLERANCE IN PLANTS 41
(Browne et al., 2002). This nematode belongs to the group ofanimals that enter a state of suspended animation known as anhy-drobiosis (Crowe et al., 1992), surviving for indefinite periodsuntil rehydration allows them to resume normal metabolism.The genomes of some microorganisms also contain sequencesthat encode LEA-like proteins (Dure, 2001). This suggests thatplants, animals, and microorganisms may use common protec-tive strategies against dehydration, although the biochemicalfunction of the LEA proteins has not been proven. Some positiveevidence for LEA proteins as protective molecules has been de-rived from overexpression studies in plants and yeast (Table 1).
LEA proteins have been divided into different groups basedon conserved structural features (Dure et al., 1989; Dure, 1993),although there is some debate about the classification and bettersystems may evolve. Group 1 LEA proteins are characterizedby high glycine contents (ca. 20%), amino acids with chargedR-groups (ca. 40%) and the presence of a stretch of 20 hy-drophilic amino acids. The high hydrophilicity of these proteinsrenders them soluble after boiling suggesting that these pro-teins are highly hydrated and do not assume a globular tertiarystructure (Dure, 1993). The group 1 proteins may be involvedin binding or replacement of water. The group 2 LEA proteins(dehydrins) are characterized by a tract of serine residues, a con-served motif containing the consensus sequence DEYGNP nearthe N-terminus and a lysine-rich 15 amino acid sequence motiftermed K-segment (EKKGIMDKIKEKLGP), which is presentin most cases at or near the carboxy terminus and is predictedto form an amphipathic α-helix. Group 2 and Group 4 proteinsmay contribute to maintenance of protein and membrane struc-tures (Dure, 1993; Ingram and Bartels, 1996). The main char-acteristic of group 3 LEA proteins is the presence of severalcopies of an 11-amino acid peptide predicted to form an am-phipathic α-helix with possibilities for intra-and intermolecularinteractions (Dure, 1993). Group 4 is characterized by a con-served N-terminus predicted to form an α-helix and a diverseC-terminal part with a random coil structure. Group 5 LEA pro-teins contain a higher proportion of hydrophobic residues thanthe other four groups and probably adopt a globular conforma-tion. Proteins of group 3 and 5 have been suggested to formdimers with a coiled-coil structure capable of sequestering ions,which accumulate due to water depletion (Dure et al., 1989).
B. AquaporinsDehydration and salt stress require changes in water flow
to allow cells and tissue to adapt to the stress situation. Therate of water flux into or out of cells is determined by the wa-ter potential gradient that acts as the driving force for transportand by the water permeability of the membrane. Evidence hasbeen accumulating that aquaporins are central components inplant water relations; this subject has recently been reviewed indetail by Tyerman et al. (2002). Aquaporin proteins facilitateosmosis by forming water-specific pores as an alternative to wa-ter diffusion through the lipid bilayer, thus increasing the water
permeability of the membrane (Schaffner, 1998; Kjellbom et al.,1999). Aquaporins are members of a large super-family of mem-brane spanning proteins, the major intrinsic proteins (MIPs). Inplants, aquaporins localized in the tonoplast are called tonoplastintrinsic proteins (TIPs), while those in the plasma membraneare PIPs. Across different plant species aquaporins have veryconserved structures and hence are encoded by similar DNA se-quences. In A. thaliana, the MIPs are forming a family of at least30 genes that display differential patterns of expression. In gen-eral, MIPs are most abundantly expressed in rapidly growing tis-sues and in cells involved in high-volume water flux (Weig et al.,1997; Kjellbom et al., 1999). There are several reports that aqua-porin genes are induced by dehydration and salt stress; this maytrigger greater osmotic water permeability and facilitate waterflux (Guerrero et al., 1990; Yamaguchi-Shinozaki et al., 1992;Fray et al., 1994; Yamada et al., 1997; Sarda et al., 1999). On theother hand, there are also examples of downregulated aquaporinsunder dehydration or salt stress, which should result in decreasedmembrane permeability and may allow cellular water conserva-tion (Yamada et al., 1997; Johansson et al., 1998). Guard cellMIP genes (TIPs) NgMIP2, NgMIP3 and (PIP) NgMIP4 aredownregulated by drought stress (Smart et al., 2001). A role foraquaporins in fine tuning of water availability is supported by arecent example from the resurrection plant C. plantagineum. It isshown that a specific aquaporin gene is upregulated by dehydra-tion but downregulated by salt stress, which coincides with thetolerance behavior of this plant (Smith-Espinoza et al., 2003).Direct evidence for a role of aquaporins in maintaining the wa-ter status of plants has been derived from aquaporin antisenseexperiments in Arabidopsis or tobacco (Kaldenhoff et al., 1998;Siefritz et al., 2002). Kaldenhoff et al. (1998) showed that Ara-bidopsis plants, in which the expression of the plasma membraneaquaporin PIP1b is suppressed, performed better in a hypotonicsolution than wild-type plants, and these plants compensatedfor the reduced water flow by increasing the root system. Ara-bidopsis plants with reduced aquaporin1 gene expression hadreduced hydraulic conductivity and decreased drought tolerance(Siefritz et al., 2002).
C. Heat Shock Proteins (Hsps)Synthesis of correctly structured proteins and their mainte-
nance is important for efficient cellular functions. The heat shockprotein family encompasses many chaperones, which have animportant role in the folding and assembly of proteins duringsynthesis, and in the removal and disposal of nonfunctional anddegraded proteins. Heat shock proteins (Hsps) are usually unde-tectable in vegetative tissues under normal growth conditions,but can be induced by environmental stress and developmen-tal stimuli. Accumulation of Hsps coincides with acquisitionof stress tolerance. In desiccation sensitive Arabidopsis mutantseeds Hsp expression is reduced (Wehmeyer and Vierling, 2000).Hsps are induced by water stress in several plants (Alamillo et al.,1995; Coca et al., 1996; Campalans et al., 2001). Transgenic

42 D. BARTELS AND R. SUNKAR
Arabidopsis plants overexpressing AtHSP17.7 accumulate highlevels of AtHSP17.7 protein and show enhanced tolerance todrought and salinity (Sun et al., 2001). This provides in vivo ev-idence for a protective role of Hsps in plants. Experiments in vitrosuggest that cytosolic Hsps function as molecular chaperonesby preventing the thermal aggregation of substrate proteins andfacilitating the subsequent reactivation (Lee et al., 1995). Theconcept that low RWC impairs protein structure explains the ne-cessity of molecular chaperones to accumulate under a range ofstresses. The abundance of small heat shock proteins (sHsps) inplants and their functional characteristics of binding and stabi-lizing denatured proteins suggest that sHsps play an importantrole in plant stress tolerance (reviewed in Wang et al., 2004).
Targeting is controlled by molecular chaperones of which oneof the major players is the chaperone binding protein (BiP), amember of the HSP70 protein family. Synthesis of BiP is inducedby physiological stress conditions that promote accumulationof mis-folded proteins. Plant BiPs were shown to be inducedby a variety of environmental stresses, including water stress(Cascardo et al., 2000). Overexpression of BiP-enhanced wa-ter stress tolerance in transgenic tobacco plants, whereas theantisense transgenic plants showed hypersensitivity to waterstress. The water-stress tolerant phenotype of BiP overexpress-ing plants is likely to be due to better osmotic adjustment whichprevents turgor loss (Alvim et al., 2001).
D. Proteases and Proteinase InhibitorsProteolysis is an important cellular activity to maintain pro-
tein homeostasis. Increased proteolysis in response to stress isfrequently observed and can be interpreted as a way to get ridof damaged proteins or to mobilize nitrogen (Vierstra, 1996).Increased proteolysis during drought/salt stress conditions hasbeen reported (Guerrero et al., 1990; Ramanjulu et al., 1994b;Ramanjulu and Sudhakar, 1997; de Carvalho et al., 2001). Cys-teine proteases were shown to be induced during drought andsalinity (Koizumi et al., 1993; Forsthoefel et al., 1998; Khanna-Chopra et al., 1999; Campalans et al., 2001; Seki et al., 2002).The water and salinity stress-induced Arabidopsis ERD1 geneencodes a chloroplast-localized protein with sequence homol-ogy to the regulatory ATPase subunit ClpA of the ATP-dependentClp protease from E. coli (Nakashima et al., 1997). The ERD1gene product may interact with other subunits such as ClpP (achloroplast gene) of the ATP-dependent protease and may func-tion in the degradation of chloroplast proteins. An ArabidopsisDegP2 gene encoding a novel chloroplast homologue of theprokaryotic trypsin was isolated and was shown to increase tohigh concentrations of NaCl, desiccation and high light (Hausuhlet al., 2001). AtDegP2 is likely to have a role in primary cleavageof the photo-damaged D1 protein of PSII prior to its removal bysecondary proteolysis.
E. PolyaminesThe polyamines spermidine, spermine, and putrescine are
polycationic small aliphatic amines that have been implicated
in a variety of physiological processes such as growth and de-velopment in plants. A role for polyamines has been proposedin stress responses. Salt-tolerant Pokkali rice plants accumu-late higher levels of polyamines compared to the salt-sensitiverice plants in response to salinity stress (Chattopadhyay et al.,2002). Exogenously supplied putrescine prevented stress dam-age and increased stress tolerance in Conyza bonariensis andmaize (Ye et al., 1998). Biosynthesis of polyamines in plantsis controlled by the enzymes ornithine decarboxylase and argi-nine decarboxylase which are responsible for the productionof putrescine, and S-adenosyl-L-methionine (SAM) decarboxy-lase that is necessary for the formation of spermidine and sper-mine. Plants subjected to osmotic stress show a rapid increasein putrescine levels due to transcription and activation of argi-nine decarboxylase (Borrell et al., 1996; Flores and Galston,1982). Arabidopsis mutants spe1-1 and spe2-1 with reduced ac-tivity of arginine decarboxylase are more sensitive to salt stressthan wild-type plants (Kasinathan and Wingler, 2002). Further-more, AtADC2 (arginine decarboxylase) expression correlatedwith free putrescine accumulation under salinity and dehydra-tion (Urano et al., 2003). Ds insertion mutant adc2-1 displayeda salt-sensitive phenotype coupled with reduced accumulationof putrescine (Urano et al., 2004). Transgenic rice plants withenhanced level of ADC gene expression increased biomass com-pared to control plants under saline conditions (Roy and Wu,2001). These observations lend support for the protective func-tion of polyamines. Polyamines may possibly exert their protec-tive function by scavanging ROS, which may occur as a conse-quence of stress (Tiburcio et al., 1994). However, accumulationof polyamines seems to be toxic to the plants under normal con-ditions and therefore constitutive overexpression may not be theappropriate way to obtain stress tolerance.
IX. OXIDATIVE STRESS A CONSEQUENCEOF DEHYDRATION AND SALT STRESS
A. Formation of Reactive MoleculesA secondary effect of dehydration and salt stress is the in-
crease of reactive oxygen species (ROS), which include singletoxygen, superoxide anion radicals, hydroxyl radicals, and hy-drogen peroxide (Smirnoff, 1998; Bartels, 2001; Apel and Hirt,2004). ROS are predominantly generated in the chloroplast bydirect transfer of excitation energy from chlorophyll to producesinglet oxygen, or by univalent oxygen reduction at photosys-tem I, in the Mehler reaction (Foyer et al., 1994; Allen, 1995)and to some extent in mitochondria. Chloroplasts are the firsttargets in plant cells since this is the major site of ROS produc-tion. The increased concentration of ROS inhibits the ability torepair damage to photosystem II and inhibits the synthesis ofthe D1 protein. Stress-enhanced photorespiration and NADPHactivity also contributes to the increased H2O2 accumulation,which may inactivate enzymes by oxidizing their thiol groups.The toxicity of H2O2 is not due to its reactivity per se, butrequires the presence of a metal reductant to form the highly

DROUGHT AND SALT TOLERANCE IN PLANTS 43
reactive hydroxyl radical, which potentially reacts with all bio-logical molecules. Transition metals such as cuprous and ferrousions may be released from enzymes and electron carriers dur-ing stress and promote the Fenton reaction to produce highlyreactive hydroxyl radicals, which extensively oxidizes proteins,lipids and nucleic acids (Halliwell and Gutteridge, 1999).
The alleviation of oxidative damage and increased resistanceto environmental stresses is often correlated with an efficient an-tioxidative system (Smirnoff, 1998; Shalata et al., 2001; Kranneret al., 2002). Overproduction of SOD, APX and catalase havebeen shown to improve oxidative stress tolerance in transgenicplants (Allen, 1995; Roxas et al., 1997). The importance of an-tioxidant systems has been demonstrated in the resurrectionplant Myrothamnus flabellifolia (Kranner et al., 2002). Thisplant was able to recover after four months of desiccation, butnot after eight months when the antioxidant defense mecha-nisms were broken down. The ability to recover after desicca-tion has been correlated with the ability to maintain/resynthesizeantioxidants such as α-tocopherol, ascorbate, and glutathione.Strategies have been developed to keep the concentrations ofROS under tight control by detoxification. ROS detoxificationmechanisms can be broadly divided into nonenzymatic and en-zymatic mechanisms. Major nonenzymatic antioxidants includeascorbate (vitamin C) and glutathione in plants, although toco-pherol (vitamin E), flavonoids, alkaloids, and carotenoids canalso act as antioxidants. Enzymatic mechanisms include super-oxide dismutase, peroxidases, and catalase. These aspects havebeen discused extensively in recent reviews (Mittler, 2002; Apeland Hirt, 2004). Here we would like to highlight additional com-ponents that aid in detoxification of ROS and secondary productsthat are derived from ROS interaction with biomolecules.
B. Enzymes That Detoxify AldehydesFree radical-mediated lipid peroxidation in biological sys-
tems is always accompanied by the generation of complex pat-terns of aldehydes. Aldehydes are highly reactive moleculesand toxic at low concentrations. Aldehyde dehydrogenases havebeen emerging as crucial enzymes in detoxification processes ina wide range of organisms. These enzymes are critical in the con-version of toxic aldehydes to the less reactive carboxylic acidforms.
In plants, not much is known about aldehyde and detoxify-ing aldehydes. An aldehyde dehydrogenase from C. plantage-nium (Cp-ALDH) and its Arabidopsis homologue Ath-ALDH3were induced by diverse abiotic stressors that induce oxidativestress (Kirch et al., 2001; Sunkar et al., 2003). Osmotic stress-induced aldehyde dehydrogenases have also been described inother plants (Seki et al., 2002; Ozturk et al., 2002; Chen et al.,2002). Another group of enzymes are aldose/aldehyde reduc-tases which reduce a wide range of aldehydes and ketones toalcohols. Osmotic stress-inducible aldose/aldehyde reductaseswere isolated from plants (Oberschall et al., 2000; Mundreeet al., 2000). Functional analyses of aldose/aldehyde reductase
and aldehyde dehydrogenases point to a role of detoxification ofaldehydes in vitro. The function is also supported by transgenicplants overexpressing these enzymes (Oberschall et al., 2000;Sunkar et al., 2003).
C. PeroxiredoxinsPeroxiredoxins are a group of enzymes with a catalytic func-
tion in the detoxification of cellular-toxic peroxides (Dietz et al.,2002). Peroxiredoxins reduce peroxide to the corresponding al-cohol. Peroxiredoxins are able to protect DNA, membranes andcertain enzymes in vitro against damage by ROS and to removeH2O2, alkyl hydroperoxides and hydroxyl radicals (Lim et al.,1993). Stress-inducible peroxiredoxins have been identified inplants (Seki et al., 2001; Mowla et al., 2002). The peroxire-doxin transcript is constitutively expressed at high levels in35S:DREB1A overexpressing plants (Kasuga et al., 1999; seeunder section VI C AP2/ERF-type-transcription factors) sug-gesting that it is one of the targets of DREB1A.
D. ThioredoxinsThioredoxins are small proteins functioning as hydrogen
donor and thereby reducing disulfide bridges in proteins. Theirinvolvement in the response to oxidative stress is well docu-mented in bacteria, yeast, and animal cells (Arner and Holmgren,2000). The role of thioredoxins in oxidative stress responsesin plants is largely unexplored. Drought- and high light stress-inducible chloroplast localized protein CDSP32 has been iden-tified in plants (Pruvot et al., 1996; Rey et al., 1998; Broinet al., 2000). The mature CDSP32 protein exhibits structuralfeatures similar to those described for bacterial thioredoxins. Adecreased photochemical efficiency by suppression of CDSP32in antisense plants suggested its role in oxidative stress. How-ever, overexpression of CDSP32 in transgenic plants leads tosusceptibility to oxidative stress and it was hypothesized thatoverproduction of thioredoxin could disturb the plastidic redoxstate (Broin et al., 2002).
E. Protein OxidationROS can lead to oxidation of amino acid residue side chains,
formation of protein-protein cross-linkages and oxidation of theprotein backbone, resulting in protein fragmentation (Berlett andStadtman, 1997). Such modified proteins are more susceptible todegradation by intracellular proteases. The sulphur-containingamino acids methionine (Met) and cysteine (Cys) in proteins aremore susceptible than others. Oxidation of Met leads to the for-mation of the methionine sulfoxide [Met(O)], a process whichleads to the loss of function (Rodrigo et al., 2002). However, en-zymes such as peptide-methionine sulfoxide reductases (MsrA)found in several organisms can compensate the damage by cat-alyzing the reduction of free and protein-bound (Met(O) residuesto Met (Moskovitz et al., 1999). The Arabidopsis gene AtSXL3is induced by dehydration and oxidative stress. Recently Ro-drigo et al. (2002) have analyzed SXL genes in knock-out yeast

44 D. BARTELS AND R. SUNKAR
and Arabidopsis mutants. Knock-out mutants have significantlygreater quantities of Met(O) compared to wild type and rever-tants under oxidative stress.
X. IONIC STRESSSimilar to osmotic stress, high concentrations of Na+ in the
soil/increased Na+ accumulation in the plant system may be rec-ognized by extracellular and intracellular sensors such that theeffective counteracting mechanisms will be initiated. As yet, themolecular identity of Na+ sensor(s) is unknown, but the long C-terminal cytoplasmic domain of SOS1 (see below) is predictedas a possible cytosolic Na+ sensor (Zhu, 2003). The identifica-tion of Na+ sensor is as important as that of osmosensor.
A. Na+ Toxicity and HomeostasisThe complexity of the plant response to salt stress can be
partially explained by the fact that salinity imposes salt toxi-city in addition to osmotic stress (Niu et al., 1995; Hasegawaet al., 2000). Sodium is toxic to many organisms, except forhalotolerant organisms like halobacteria and halophytes, whichpossess specific mechanisms that keep intracellular sodium con-centrations low. Accumulation of sodium in the cytoplasm isprevented by restricting its uptake across the plasma membraneand by promoting its extrusion or sequestration in halophytes(Hasegawa et al., 2000). High salt concentrations (>400 mM)inhibit the activities of most enzymes because of perturbation ofthe hydrophobic-electrostatic balance between the forces main-taining protein structure. However, toxic effects on cells occurat much lower salt concentrations (about 100 mM), pointing tospecific salt toxicity targets (Serrano, 1996). Surprisingly, thecytosolic enzymes from glycophytes are as sensitive as those ofhalophytes to Na+ (Blumwald et al., 2000). On the other hand,extracellular (apoplastic) enzymes from glycophytes and halo-phytes have been shown in vitro to be remarkably salt-insensitivetolerating concentrations up to 500 mM NaCl (Thiyagarajahet al., 1996).
Many physiological studies have demonstrated that Na+ tox-icity is not only due to toxic effects of Na+ in the cytosol, butalso because K+ homeostasis is disrupted possibly due to theability of Na+ competing for K+ binding sites.
Considerable progress has been made in understanding ionhomeostasis (Hasegawa et al., 2000; Blumwald et al., 2000;Apse and Blumwald, 2002; Zhu, 2003). Ion transporters areconsidered to play an important role in salt tolerance. In princi-ple, three mechanisms exist to prevent excess Na+ accumulationin the symplast of plant cells:
1. Restricting the Na+ permeation and entry into plants by Na+
transporters, whose molecular identity is unknown.2. Compartmentalizing the Na+ in the vacuole.3. Extruding Na+: cytosolic Na+ can be transported back to
the external medium or the apoplast via plasma membraneNa+/H+ antiporter activity.
B. Na+ ExclusionNa+ levels should be controlled at the entry point (Munns,
2002). The Na+ uptake across the plasma membrane has beenattributed to low Na+ permeability properties of systems thattransport essential K+ (Blumwald et al., 2000; Hasegawa et al.,2000). The Arabidopsis transporter AtHKT1 has been identifiedas one of the pathways of Na+ entry into plants. Mutation inAtHKT1 suppresses the hypersensitivity of sos3 mutants (Ruiset al., 2001) suggesting that the wild-type SOS3 may inhibitthe activity of AtHKT1 as a Na+ influx transporter. In addition,nonselective cation channels present in the root plasma mem-brane may also contribute to Na+ entry into plants (Amtmannand Sanders, 1999), although the molecular identity of suchtransporters is still unknown.
C. Na+ CompartmentalizationIn plants, the central vacuole plays a vital role in regulation of
cytoplasmic ion homeostasis. Exclusion of excess Na+ from thecytoplasm and the accumulation in the vacuole represents oneof the adaptive mechanisms during salt stress. Halophytes havethe ability to use Na+ as an osmoticum by compartmentalizingit into vacuoles. The vacuolar sodium sequestration is medi-ated by a Na+/H+ antiport at the tonoplast (Apse et al., 1999).Sequestration or compartmentalization of sodium into the vac-uole through vacuolar Na+/H+ antiporters uses the proton mo-tive force generated by the vacuolar H+-translocating enzymes,H+-adenosine triphosphatase (ATPase), and H+-inorganic py-rophosphatase (PPiase), to couple the downhill movement ofH+ with the uphill movement of Na+ against the electrochemi-cal potential (Blumwald and Gelli, 1997).
The presence of Na+/H+ antiporter activities has been phys-iologically characterized in tonoplast vesicles and it is molecu-larly represented by six Arabidopsis genes AtNHX1-6 (Blumwaldet al., 2000; Yokoi et al., 2002). AtNHX1 steady-state transcriptlevels were increased in response to NaCl, KCl, sorbitol, andABA suggesting that the AtNHX1 transcript upregulation is notspecific to ionic stress but common to osmotic stress (Gaxiolaet al., 1999; Quintero et al., 2002; Shi and Zhu, 2002; Yokoiet al., 2002). AtNHX1 was able to suppress the salt-sensitive phe-notype of the nhx1 yeast mutant (Gaxiola et al., 1999; Quinteroet al., 2002). Evidence for the in vivo function was derivedfrom ectopically expressing AtNHX1 in Arabidopsis, tomato andcanola (Apse et al., 1999; Zhang and Blumwald, 2001; Zhanget al., 2001). This was further supported by the overexpressionof the rice homologue in transgenic rice plants (Fukuda et al.,2004). Tonoplast vesicles isolated from transgenic plants dis-played significantly higher Na+/H+exchange rates than those ofwild-type plants.
Expression of both AtNHX2 and 5 also suppressed the al-kali cation-sensitive phenotype of an nhx1 mutant yeast, likeAtNHX1 (Yokoi et al., 2002). AtNHX2 mediated relatively highertolerance than AtNHX1 and 5 in yeast, which may suggest thatNHX proteins perform similar functions but with varying

DROUGHT AND SALT TOLERANCE IN PLANTS 45
efficiency. AtNHX1 and 2 are responsive to NaCl, sorbitol andABA suggesting that AtNHX1 and 2 are responsive to hyper-osmotic stress rather than to Na+ toxicity, while AtNHX5 wasspecifically increased in response to NaCl but not to sorbitol,an indication for its involvement in Na+ exchange (Yokoi et al.,2002).
AtNHX1 is constitutively expressed and thus it is likely tohave physiological roles under nonstress conditions. One suchfunction seems to be pH regulation (Fukuda-Tanaka et al., 2000).Venema et al. (2002) showed that AtNHX1 was able to mediatelow affinity Na+ as well as K+ transport in artificial, reconsti-tuted liposomes and therefore AtNHX1 may also have a role ofK+ accumulation in vacuoles. These observations implicate thatNHX proteins may have additional roles such as K+ exchangeactivity and pH regulation.
In yeast the protein phosphatase PPZ1 and its regulatory sub-unit HAL3 act as important determinants of salt tolerance byregulating the expression of the ENA1 gene encoding the ma-jor sodium extrusion pump (de Nadal et al., 1998). TransgenicArabidopsis plants constitutively overexpressing AtHAL3a, theHAL3 Arabidopsis homologue, showed improved growth andosmotic stress tolerance (Espinosa-Ruiz et al., 1999). This pos-sibly suggests a similar role for AtHAL3 in plants as for its yeastcounterpart HAL3.
D. Proton Transporters and Salt ToleranceRegulation of ion transport is one of the important factors for
salt tolerance of plants. Protons are used as coupling ions for iontransport systems, and the proton gradient, generated by protonpumps in the membrane systems, is the driving force for Na+
transport across membranes. The plasma membrane H+-ATPase(P-H+-ATPases) and vacuolar ATPases (V-ATPases) coupledwith vacuolar H+-PPase (pyrophosphatase) (V-H+-PPases) actat the plasma membrane and tonoplast, respectively. The tono-plast H+-ATPase and H+-PPase play important roles in trans-porting H+ into the vacuole and generation of proton motiveforce across the tonoplast, which operates Na+/H+antiporters(Hasegawa et al., 2000; Blumwald et al., 2000).
The P-H+-ATPases (proton pumps) are encoded by at least12 genes in Arabidopsis. P-H+-ATPase from Arabidopsis wasidentified as one of the important components of salt tolerance inplants (Vitart et al., 2001). A mutation in the ATPase gene AHA4causes a reduction in root and shoot growth under NaCl stress.Salt stress resulted in more Na+ and less K+ accumulation inthe leaves of aha4-1 mutant plants compared to wild type. It ishypothesized that AHA4 functions in the control of Na+ fluxacross the root endodermis, possibly by partially disrupting theactivity of other pumps (Vitart et al., 2001).
The V-type ATPase is a multimeric enzyme localized in en-domembranes of eukaryotic cells that establishes an electro-chemical H+-gradient (Luttge and Ratajczak, 1997). TheV-ATPases are of prime importance in energizing sodium se-questration into the central vacuole. Transcripts encoding
subunits of V-ATPases accumulate in response to salt stressleading to increased enzyme activity (Ratajczak et al., 1994).Salt induced the transcription of several V-ATPase subunits (A,B, C, and E) in the common ice plant (Dietz and Arbinger,1996), subunit A in salt-adapted tobacco cell suspension culture(Narasimhan et al., 1991) and subunits A and C in sugar beet(Kirsch et al., 1996; Lehr et al., 1999). A close correlation be-tween the expression of subunit E of the V-ATPase and salttolerance was reported for the ice plant, i.e., juvenile non-salt-tolerant plants did not alter subunit E; whereas the transcript lev-els increased in salt-tolerant mature leaves (Golldack and Dietz,2001). Coupled with ATPase activity is the vacuolar H+-PPase(pyrophosphatase) activity, which is encoded by Avp1in Ara-bidopsis. The activity of Avp1 is correlated with increased ioncontent of the plants. Overexpression of Avp1 leads to enhancedsalt and drought tolerance in transgenic plants (Gaxiola et al.,2001). This suggests that pumping of H+ across the vacuolarmembrane is an additional driving force for vacuolar sodiumaccumulation. Therefore saltadaptation is correlated with V-ATPase in many plants. In summary, salt adaptation is correlatedwith the tonoplast Na+/H+ antiporter and with increased activ-ity of V-H+-ATPase submits. The coordination of both systemsis crucial for sequestering Na+ in cellular compartments.
E. SOS Pathway and Ion HomeostasisGenetic approaches and yeast complementation assays have
identified individual components of sodium homeostasis, butonly the systematic genetic approach in Arabidopsis by Zhuand colleagues have led to the discovery of a linear salt-stress-induced Ca2+-regulated pathway involved in Na+ homeostasis.The molecular basis for this was the characterization of the SOS(Salt Overly Sensitive) mutants, which render the SOS plantshypersensitive to NaCl. The identification of SOS genes has un-covered a novel Ca2+-dependent pathway determining ion (Na+
and K+) homeostasis and salt tolerance. Molecular interactionand complementation analysis indicate that SOS3 is requiredfor activation of SOS2 that regulates SOS1 transcription. Doublemutant analysis showed that three SOS genes function in a linearpathway (Zhu, 2001a). The SOS3 gene encodes a Ca2+ sensorprotein similar to the calcineurin B subunit from yeast, a Ca2+
dependent protein phosphatase, and neuronal calcium sensorsfrom animals (Liu and Zhu, 1998). Calcineurin B and neuronalcalcium sensors are both activated by Ca2+, which raises thepossibility that the SOS3 gene product might control K+/Na+
transport via a Ca2+-regulated pathway (Liu and Zhu, 1998).Calcineurin is a protein phosphatase type 2B that plays a vitalrole in signalling pathways in regulating ion homeostasis andsalt tolerance of yeast (Mendoza et al., 1994) and plants (Pardoet al., 1998; Liu and Zhu, 1998). Evidence suggests that SOS3is involved in regulating the SOS2 protein kinase (Halfter et al.,2000). SOS2 encodes a serine/threonine protein kinase with anN-terminal kinase domain and a unique C-terminal domain withFISL motif that is sufficient for interaction with SOS3 (Liu et al.,

46 D. BARTELS AND R. SUNKAR
2000). Removal of the FISL motif renders the SOS2 kinase con-stitutively active (Guo et al., 2001). SOS1 mutant plants are hy-persensitive to salinity and accumulate high amounts of Na+
compared to wild-type plants (Liu and Zhu, 1997). The SOS1gene encodes a plasma membrane Na+/H+ antiporter, with along cytoplasmic C-terminal tail (Shi et al., 2000). The upregu-lation of SOS1 in response to NaCl is reduced in sos2 and sos3mutant plants, suggesting that SOS1 expression is controlled bySOS2/SOS3, which is further evidence for the interaction of thethree SOS genes. SOS3 forms a complex with SOS2, and thiscomplex is necessary for the phosphorylation and subsequentactivation of SOS1, the Na+/H+ antiporter (Qiu et al., 2002).Overexpression of SOS1 in Arabidopsis plants improved salttolerance with reduced Na+ accumulation in shoots. SOS1 prob-ably retrieves Na+ from the xylem (Shi et al., 2003). It seemsthat the SOS pathway also regulates the vacuolar Na+/H+ ex-change activity and contributes to Na+ compartmentalization(Qiu et al., 2004). The emerging picture from these studies isthat the SOS pathway coordinately regulates plasma membraneand tonoplast Na+/H+ antiporter activity which leads to Na+
homeostasis and thus salt tolerance.
XI. ABSCISIC ACID (ABA)The plant hormone abscisic acid (ABA) plays a central role
in many aspects of stress responses as well as seed develop-ment, dormancy, and germination. Different aspects have re-cently been covered in excellent reviews (Leung andGiraudat, 1998; Rock, 2000; Finkelstein et al., 2002). Here wewill focus on the role of ABA in osmotic stress. During vege-tative growth, ABA-mediated adaptive responses are critical toplant survival during drought, salt, and cold stress. These stres-sors serve as a trigger for the accumulation of ABA, which inturn activates various stress-associated genes that are thoughtto function in the accumulation of osmoprotectants, (LEA) pro-teins, signalling, transcriptional regulation etc. Exposure to ex-ogenous ABA mimics the induction of genes similar to stresstreatment. This gene induction can be correlated with acquisi-tion of desiccation tolerance similar to what has been shownfor callus derived from the resurrection plant C. plantagineum(Bartels and Salamini, 2001). ABA-insensitive and biosyntheticmutants have confirmed the role of ABA as an intermediatemolecule between stress perception and cellular stressresponse.
A. Regulation of ABA LevelsMutant plants with altered biosynthesis, perception, or re-
sponse have been crucial in identification of various compo-nents involved in ABA biosynthesis and signalling. The geneticscreens and selections that have been used include productionof nondormant seeds, loss or gain of sensitivity to ABA dur-ing germination, seedling or root growth, and altered expres-sion of reporter genes. These approaches have yielded threeclasses of ABA mutants: ABA-deficient, –hypersensitive, and
–insensitive. ABA-deficient mutants (impaired ABA biosyn-thesis) exhibited a wilty phenotype due to impaired stomatalclosure.
Significant progress has been made in recent years with cloninggenes encoding enzymes involved in ABA biosynthetis. Thisrevealed the critical steps in the regulation of ABA levels. Themain ABA biosynthetic pathway starts from carotenoids C-40(Zeevart and Creelman, 1988). Biochemical studies have in-dicated that 9-cis-epoxycarotenoid dioxygenase (NCED) is akey enzyme in ABA biosynthesis (Kende and Zeevaart, 1997).NCED catalyzes the cleavage reaction of epoxycarotenoids whichproduces xanthoxin (the first C15 intermediate). The ABA ac-cumulation is regulated at the transcriptional level in response toosmotic stress (Guerro and Mullett, 1986; Stewart et al., 1986).Accordingly, the induction of the NCED gene by drought stresshas been reported in maize, tomato, Arabidopsis, bean, and cowpea (Burbridge et al., 1997; Tan et al., 1997; Neill et al., 1998;Qin and Zeevaart et al., 1999; Iuchi et al., 2000; Thompsonet al., 2000a). The timing of the induction of the VuNCED1 genewas shown to be slightly earlier than that of ABA accumulationunder drought stress suggesting that the transcriptional regula-tion of the genes involved in the ABA biosynthesis pathwayare responsible for drought-induced ABA accumulation (Iuchiet al., 2000). The important role of NCED was confirmed bytransgenic and mutant studies which confirmed for Arabidop-sis and tomato that the expression of the NCED gene causedan increase in ABA levels (Iuchi et al., 2001; Thompson et al.,2000b). Plants overexpressing AtNCED3 showed reduced tran-spiration rates and improved drought tolerance, whereas plantswith nonfunctional NCED genes had a drought-sensitive pheno-type (Iuchi et al., 2001). These experiments provide convincingevidence that modulating of endogenous ABA levels is possiblevia engineering NCED expression, which subsequently changesdrought tolerance and most likely other responses to osmoticstress. The Arabidopsis genome contains at least seven homol-ogous NCED genes, but there is hardly any knowledge of theirindividual functions.
Recent findings confirmed that ZEP and NCED are locatedin plastids, and that the product of the ABA levels are modu-lated not only by the rate of synthesis, but also by the activityof degradation enzymes. The hydroxylation at the 8′-positionof ABA is thought to be the key step of ABA catabolism, andthis reaction is catalyzed by ABA 8′-hydroxylase, a cytochromeP450. The gene responsible for this process in Arabidopsis isCYP707A3. ABA treatment resulted in increased transcript ac-cumulation of CYP707A3 (Saito et al., 2004). Further, plantsthat accumulated ABA due to overexpression of NCED alsohyperaccumulated phaseic acid (Qin and Zeevaart, 2002), sug-gesting that the ABA catabolism has been increased in theseplants. These observations support the notion that ABA mightrestrict its own accumulation by activating its degradation. Thefindings suggest that both ABA biosynthesis and catabolism areactivated by stress/ABA in fine tuning the levels of ABA. There-fore, it is possible that ABA negatively regulates the biosynthetic

DROUGHT AND SALT TOLERANCE IN PLANTS 47
genes and positively regulates the catabolic genes in feed-backand feed-forward mechanisms.
B. The Role of ABA in Stomatal ClosureTranspiration through stomatal pores is a crucial response of
the plant under osmotic stress regulated by ABA. The closureof stomatal pores in aerial tissues is an important mechanism bywhich higher plants regulate their water balance. Guard cells,which flank stomatal pores integrate and respond appropriatelyto changes in water levels with ABA as a major signal. ABA-deficient mutants are prone to wilting and cannot withstand wa-ter deficit conditions due to unregulated stomatal control. Therehave been major advances in our understanding of the cellularevents that underlie the regulation of stomatal aperture in re-sponse to osmotic stress (reviewed in Schroeder et al., 2001).Different signalling components in this process such as proteinkinases and phosphatses, Ca2+, ROS, IP3, IP6, NO, syntaxin,sphingosine-1-phosphate, phosphatidic acid, G proteins, G pro-tein coupled receptor (GCR1) and cADPR have been identi-fied recently (Leung et al., 1994; Leymzann et al., 1999; Peiet al. 2000; Li et al., 2000; Ng et al., 2001; Wang, 2001; Mataand Lamattina, 2001; Zhu et al., 2002; Coursol et al., 2003;Pandey and Assmann, 2004). ABA-induced stomatal closureis mediated by a reduction in the turgor pressure of guard cells,which requires an efflux of K+ and Cl−, sucrose removal and theconversion of malate to osmotically inactive starch (Schroederet al., 2001). ABA triggers an increase in cytosolic calcium lev-els in guard cells which regulate ion channels hat control ionefflux and stomatal closure. These aspects have been coveredin some recent excellent reviews by Assmann and Wang (2001)and Schroeder et al. (2001).
ABA-induced stomatal closure requires a reorganization ofthe actin cytoskeleton of guard cells (Eun and Lee, 1997), sug-gesting that the ABA-triggered cytoskeleton reorganization isimportant for the process of stomatal closure. A link for suchan action has been demonstrated between ABA and actin throughAtRac1. AtRac1 (Arabidopsis Rho-related small guanosine triphos-phatase, GTPase) has been identified as a central component inthe ABA-mediated stomatal closure. In animals and yeast RhoGTPases are key regulators of the actin cytoskeleton. GTPasesare inactivated by ABA treatment leading to the disruption of theguard cell actin in wild-type but not in abi1-1 mutant. Furtherexpression of a dominant-negative mutant resulted in stomatalclosure, while the dominant-positive mutant inhibited the induc-tion of stomatal closure by ABA (Lemichez et al., 2001). A linkbetween ABA-induced increases in [Ca2+] and the cytoskeletonreorganization has been also established (Hwang and Lee, 2001)suggesting that the ABA-induced increases in [Ca2+] is a com-ponent in the pathways leading to both alteration in the activityof ion channels and the control of cytoskeletal reorganization.
C. ABA Signalling ComponentsCharacterization of ABA hypersensitive mutants led to the
identification of several components of ABA signalling. ABA
hypersensitive mutants display enhanced sensitivity to ABA,resulting in diminished germination rates at low ABA concen-trations and reduced water loss due to enhanced ABA-inducedstomatal closure (Arabidopsis era1, sad1, abh1, hyl1, rop10 andfiery 1). The ERA1 gene encodes a protein farnesyltransferase,HYL1, SAD1, and ABH1 genes encode different types of RNAbinding proteins. Farnesyltransferases influence protein struc-ture or localization through mechanisms other than phosphory-lation. Protein farnesylation mediates the COOH-terminal lipi-dation of specific cellular proteins such as Ras and G-proteins.Pei et al. (1998) reported that deletion of the Arabidopsis farne-syltransferase gene ERA1 or application of farnesyltransferaseinhibitors resulted in ABA hypersensitivity of guard cell anion-channel activation and of stomatal closure implicating a rolefor farnesyltransferases in stomatal regulation. The era1 mutantplants exhibited a reduction of transpirational water loss duringdehydration suggesting ERA1 is a negative regulator in this pro-cess (Pei et al., 1998). FRY1 encodes an inositolpolyphosphate-1-phosphatase. Fry1 mutation results in elevated levels of inositol-1, 4, 5-triphosphate, super-induction of ABA and stress-induciblegenes in response to ABA, salt, and dehydration. Therefore,FRY1 is a negative regulator of ABA and stress signalling andestablishes a connection between phosphoinositols and ABAand stress signalling in plants (Xiong et al., 2001c).
Recent identification of RNA-binding proteins ABH1 (ABAhypersensitive), SAD1 (supersensitive to ABA and drought 1)and HYL1 (hyponastic leaves 1), whose mutation confers anABA-hypersensitive phenotype (Lu and Federoff, 2000;Hugouvieux et al., 2001; Xiong et al., 2001b) and AKIP1 (an hn-RNP binding protein) (Li et al., 2002) implicates that ABA mayalso play a role in post-transcriptional RNA processing. HYL1(hyponastic leaves 1) is a double stranded–RNA binding proteinand it was shown recently that HYL1 is involved in microRNAproduction. Another modulator of ABA signalling Abh1 encod-ing a mRNA cap binding protein was identified (Hugouvieuxet al., 2001). Abh1 plants exhibited drought tolerance comparedto wild-type due to increased stomatal closure coupled with el-evations in cytosolic calcium levels.
DNA chip analysis indicated only a few of the genes im-plicated in ABA signalling are downregulated in the mutant.One important gene is type 2C phosphoprotein phosphatase thathas been proposed to function as a negative regulator of ABAsignal transduction (Sheen, 1998). It seems that ABH1 influ-ences ABA signalling pathways upstream of [Ca2+], since abh1mutant guard cells showed hypersensitive ABA-induced Ca2+
elevations when compared with wild-type. Sad1 (supersensitiv-ity to ABA and drought 1) mutant is impaired in the last stepof drought-induced ABA biosynthesis, i.e., conversion of ABAaldehyde to ABA (Xiong et al., 2001). SAD1 encodes a Sm-like small ribonucleoprotein that is predicted to be involved inmRNA splicing, export, and degradation. Further studies are re-quired to establish the link between SAD1 expression and ABAbiosynthesis (Xiong and Zhu, 2003). Another RNA binding pro-tein, AKIP1 was specifically phosphorylated by ABA-activated

48 D. BARTELS AND R. SUNKAR
AAPK that increases the affinity of AKIP1 for the dehydrin tran-script (Li et al., 2002). How these proteins execute their role inthe ABA signalling pathway is unknown but the findings impli-cate a connection between RNA processing and ABA signalling.
XII. MONITORING GLOBAL GENE EXPRESSIONUSING MICROARRAY ANALYSIS
The field of plant stress tolerance has recently devoted consid-erable research effort towards identifying stress response genes.This approach is largely boosted by the availability of Ara-bidopsis and rice genome sequences. Genomics technologiesnow provide high-throughput integrated approaches for tran-script and protein analysis, which should complete our view ofstress-activated gene expression and may also lead to novel genediscovery. Although it is believed that high-throughput experi-mentation will be highly useful in finding the whole set of genesthat are involved in the process, single-gene studies remain es-sential to verify and evaluate high-throughput data sets. The firstavailable results on microarray data in relation to abiotic stressesin Arabidopsis and rice will be summarized here. These studiesprovide important information, given the complexity of plantresponse to abiotic stresses. Seki et al. (2001) have constructedArabidopsis full-length cDNA microarrays using about 1300full-length cDNAs. Forty-four genes were identified as droughtinducible, of which 30 genes are novel drought-inducible genes.In a subsequent experiment, Seki et al. (2002) have analyzedthe expression of 7000 Arabidopsis full-length cDNAs in re-sponse to dehydration, high-salinity, and cold stress treatment.Dehydration and high-salinity treatments resulted in increasedexpression of 277 and 194 genes, respectively. Nearly 70 per-cent (141 genes) of the high-salinity–inducible genes are alsoinduced by dehydration, emphasizing an extensive overlap of de-hydration and salt stress. The same array experiment identified40 transcription factor genes as dehydration, high-salinity, orcold-inducible genes. These include six members of the DREBfamily, two members of the ERF family, ten members of the Zn-finger family, four members of the WRKY family, three mem-bers of the Myb family, two members of the bHLH family, fourmembers of the bZIP family, five members of the NAC familyand three members of the HDZIP.
In another study, a comparative analysis was made betweensalt-tolerant and salt-sensitive rice varieties in terms of saltshock–induced gene expression profiles (Kawasaki et al., 2001).Salt-tolerant Pokkali rice plants survived a 150 mM salt shocktreatment, whereas salt-sensitive IR29 plants collapsed within24 h of the treatment. Thirtythree percent of the Pokkali tran-scripts were upregulated after 1 h treatment with 150 mM NaClshock, whereas only 7 percent of the salt-sensitive IR29 tran-scripts were upregulated under similar conditions. Mostly, tran-scripts homologous with ribosomal proteins were upregulatedin Pokkali but not in IR29. After 3 h 38 percent of the transcriptswere altered in both varieties not showing significant differencesin the two lines. After 6 h of treatment, 38 percent of the tran-
scripts were downregulated in IR29, but only 13 percent weredownregulated in Pokkali. The differences in expression behav-ior between the two lines include a delay in the initial response byIR29. For instance, one of the early responsive genes, CDPK-likesequence was threefold upregulated in Pokkali but not in IR29after 1 h of salt shock treatment, suggesting a difference in signaltransduction at the early stages of stress (Kawasaki et al., 2001).Global expression profiles also provide an overview of thosegenes, which are downregulated by dehydration or salt stress.A significant percent of genes are downregulated by dehydra-tion or salinity (Bockel et al., 1998; Kawasaki et al., 2001; Sekiet al., 2002; Oztur et al., 2002). For example, Seki et al. (2002)have identified 79 and 89 Arabidopsis genes downregulated bydrought and high-salinity, respectively. Mostly, photosynthesis-related genes and the components of PSI and PSII belong tothis group of genes (Seki et al., 2002; Bockel et al., 1998). Alogical hypothesis for such a response is that photosynthesis isdecreased as a consequence of stress imposition, which in turndownregulates the components involved in this process. Thisrepresents an adaptive mechanism to decrease the formation ofROS. On the other hand, some of the downregulated genes maybe of direct adaptive value. For example, downregulation of pro-line dehydrogenase is important for proline accumulation underosmotic stress conditions. In addition, some of the downregu-lated genes may include negative regulators of stress responses.
Microarray studies show that at least several hundred genesare involved in response to drought or salt stress in plants. A care-ful analysis of the transcription profiles should reveal not only in-dividual stress-activated genes but pathways. With the advancesin proteomics we should be able to simultaneously quantify thelevels of individual proteins or follow post-translational modifi-cations that occur in response to abiotic stresses. Unfortunately,the global expression analysis will at present be restricted toa few species, in particular Arabidopsis, rice, and maize, fromwhich large EST collections are available. Interesting stress-tolerant species are not amenable to this analysis because oflack of tools. It remains to be seen to what extent the data canbe extrapolated from those species, which can undergo globalexpression analysis.
XIII. CONCLUSIONSDespite the fact that research efforts have produced an enor-
mous amount of information, we are far from understanding thecomplete circuits of stress reactions. Only a few componentsof many pathways have been the subject of investigations. Fu-ture priorities should be aimed at the identification of moleculesconnecting pathways and of key components in each pathway.It is proposed that our understanding of plant stress tolerancecan be greatly refined by thorough characterization of individ-ual genes and assessing their contribution to stress tolerance.Such experiments indicated that many individual genes appearto have some positive impact on stress tolerance; mainly mas-ter switches such as transcription factor or upstream signalling

DROUGHT AND SALT TOLERANCE IN PLANTS 49
molecules are promising candidate genes for biotechnologicalapproaches. However, a combination of such master genes thatact in different pathways, e.g., ROS scavenging and osmoticadaptation, may prove even more beneficial for improving stresstolerance.
ACKNOWLEDGMENTSThe work in the laboratory of D. Bartels was supported by
grants from the European Commission and the German researchCouncil (DFG). S. Ramanjulu was supported by a fellowshipfrom the Alexander von Humboldt Foundation. We appologizeto all colleagues whose work has not been cited due to restric-tions in length.
REFERENCESAbd-El Baki, G. K., Siefritz, F., Man, H. M., Weiner, H., Kaldenhoff, R., and
Kaiser, W. 2000. Nitrate reductase in zea mays L. under salinity. Plant CellEnviron. 23: 515–521.
Abe, H., Urao, T., Ito, T., Seki, M., Shinozaki, K., and Yamaguchi-Shinozaki,K. 2003. Arabidopsis AtMYC2 (b-HLH) and AtMYB2 (MYB) function astranscriptional activators in abscisic acid signalling. Plant Cell 15: 63–78.
Abe, H., Yamaguchi-Shinozaki, K., Urao, T., Iwasaki, T., Hosakawa, D., andShinozaki, K. 1997. Role of Arabidopsis MYC and MYB homologs indrought- and abscisic acid-regulated gene expression. Plant Cell 9: 1859–1868.
Abede, T., Guenzi, A. C., Martin, B., and Cushman, J. C. 2003. Tolerance ofmannitol-accumulating transgenic wheat to water stress and salinity. PlantPhysiol. 131: 1748–1755.
Ahmad, I. and Hellebust, J. A. 1988. The relationship between inorganic nitrogenmetabolism and proline accumulation in osmoregulatory responses of twoeuryhaline microalgae. Plant Physiol. 88: 348–354.
Akashi, K., Miyake, C., and Yokota, A. 2001. Citrulline, a novel compatiblesolute in drought-tolerant wild watermelon leaves, is an efficient hydroxylradical scavenger. FEBS Lett. 508: 438–442.
Alamillo, J., Almogura, C., Bartels, D., and Jordano, J. 1995. Constitutive ex-pression of small heat shock proteins in vegetative tissues of the resurrectionplant Craterostigma planatgenium. Plant Mol. Biol. 29: 1093–1099.
Albrecht, V., Wein, S., Blazevic, D., D’Angelo, C., Batistic, O., Kolukisaoglu,U., Bock, R., Schulz, B., Harter, K., and Kudla, J. 2003. The calcium sensorCBL1 integrates plant responses to abiotic stresses. Plant J. 36: 457–470.
Allen, G. J., Kuchitsu, K., Chu, S. P., Murata, Y., and Schroeder, J. I. 1999.Arabidopsis abi1-1 and abi2-1 phosphatase mutations reduce abscisic acid-induced cytosolic calcium rises in guard cells. Plant Cell 11: 1785–1798.
Allen, R. 1995. Dissection of oxidative stress tolerance using transgenic plants.Plant Physiol. 107: 1049–1054.
Alvim, F. C., Carolino, S. M. B., Cascardo, J. C. M., Nunes, C. C., Martinez,C. A., Otoni, W. C., and Fontes, E. P. B. 2001. Enhanced accumulation ofBiP in transgenic plants confers tolerance to water stress. Plant Physiol. 126:1042–1054.
Amtmann, A. and Sanders, D. 1999. Mechanisms of Na+ uptake by plant cells.Advances in Botanical Research 29: 75–112.
Apel, K. and Hirt, H. 2004. Reactive oxygen species: Metabolism, oxidativestress, and signal transduction. Annu. Rev. Plant. Biol. 55: 373–399.
Apse, M. P., Aharon, G. S., Snedden, W. A., and Blumwald, E. 1999. Salttolerance conferred by overexpression of a vacuolar Na+/H+ antiport inArabidopsis. Science 285: 1256–1258.
Apse, M. P. and Blumwald, E. 2002. Engineering salt tolerance in plants. Curr.Opin. Biotechnol. 13: 146–150.
Arner, E. S. J. and Holmgren, A. 2000. Physiological functions of thioredoxinand thioredoxin reductase. Eur. J. Biochem. 267: 6102–6109.
Assmann, S. M. and Wang, X. O. 2001. From milliseconds to millions of years:Guard cells and environmental responses. Curr. Opin. Plant Biol. 4: 421–428.
Badawi, G. H., Kawano, N., Yamauchi, Y., Shimada, E., Sasaki, R., Kubo, A.,and Tanaka, K. 2004. Over-expression of ascorbate peroxidase in tobaccochloroplasts enhances the tolerance to salt stress and water deficit. Physiol.Plant 121: 231–237.
Baker, S. S., Wilhelm, K. S., and Thomashow, M. F. 1994. The 5′-region of Ara-bidopsis thaliana cor15a has cis-acting elements that confer cold-, drought-and ABA-regulated gene expression. Plant Mol. Biol. 24: 701–713.
Bartels, D. 2001. Targeting detoxification pathways: An efficient approach toobtain plants with multiple stress tolerance? Trends Plant Sci. 6: 284–286.
Bartels, D. and Salamini, F. 2001. Desiccation tolerance in the resurrection plantCraterostigma plantagineum. A contribution to the study of drought toleranceat the molecular level. Plant Physiol. 127: 1346–1353.
Bartels, D., Singh, M., and Salamini, F. 1988. Onset of desiccation toleranceduring development of the barley embryo. Planta 175: 485–492.
Bastola, D. R., Pethe, V. V., and Winicov, I. 1998. Alfin1, a novel zinc fingerprotein in alfalfa roots that binds to promoter in the salt-inducible MSPRP2gene. Plant Mol. Biol. 38: 1123–1135.
Berlett, B. S. and Stadtman, E. R. 1997. Protein oxidation in aging, disease andoxidative stress. J. Biol. Chem. 272: 20313–20316.
Bianchi, G., Gamba, A., Murelli, C., Salamini, F., and Bartels, D. 1991.Novel carbohydrate metabolism in the resurrection plant Craterostigma plan-tagineum. Plant J. 1: 355–359.
Blumwald, E., Aharon, G. S., and Apse, M. P. 2000. Sodium transport in plantcells. Biochimica et Biophysica Acta 1465: 140–151.
Blumwald, E. and Gelli A. 1997. Secondary inorganic ion transport in plantvacuoles. Adv. Bot. Res. 25: 401–407.
Bockel, C., Salamini, F., and Bartels, D. 1998. Isolation and characterizationof genes expressed during early events of the dehydration process in theresurrection plant Craterostigma plantagineum. J. Plant Physiol. 25: 158–166.
Bohnert, H. J. and Cushman, J. C. 2000. The ice plant cometh: Lessons in abioticstress tolerance. J. Plant Growth Regul. 19: 334–346.
Borrell, A., Besford, R. T., Altabella, T., Masgrau, C., and Tiburcio, A. F. 1996.Regulation of arginine decarboxylase by spermine in osmotically-stressed oatleaves. Physiol. Plant 98: 105–110.
Borsani, O., Valupesta, V., and Botella, M. A. 2001. Evidence for a role ofsalicylic acid in the oxidative damage generated by NaCl and osmotic stressin Arabidopsis seedlings. Plant Physiol. 126: 1024–1030.
Bouche, N., Scharlat, A., Snedden, W., Bouchez, D., and Fromm, H. 2002. Anovel family of calmodulin-binding transcription activators in multicellularorganisms. J. Biol. Chem. 277: 21851–21861.
Bray, E. A. 1997. Plant responses to water deficit. Trends Plant Sci. 2: 48–54.Bray, E. A., Bailey-Serres J., and Weretilnyk, E. 2000. Responses to abiotic
stresses. In: Biochemistry and Molecular Biology of Plants. Gruissem, W.,Buchnnan, B., and Jones, R. eds. American Society of Plant Physiologists,Rockville, MD, 1158–1249.
Bressan, R. A., Hasegawa, P. M., and Pardo, J. M. 1998. Plants use calcium toresolve salt stress. Trends Plant Sci. 3: 411–412.
Bressan, R. A., Zhang, C. Q., Zhang, H., Hasegawa, P. M., Bohnert, H. J., andZhu, J. K. 2001. Learning from the Arabidopsis experience. The next genesearch paradigm. Plant Physiol. 127: 1354–1360.
Broin, M., Cuine, S., Eymery, F., and Rey, P. 2002. The plastidic 2-cysteinperoxiredoxin is a target for a thioredoxin involved in the protection of thephotosynthetic apparatus against oxidative damage. Plant Cell 14: 1417–1432.
Broin, M., Cuine, S., Peltier, G., and Rey, P. 2000. Involvement of CDSP32a drought-induced thioredoxin in the response to oxidative stress in potatoplants. FEBS Lett. 467: 245–248.
Browne, J., Tunnacliffe, A., and Burnell, A. 2002. Plant desiccation gene foundin a nematode. Nature 416: 38.
Burbridge, A., Grieve, T. M., Jackson, A., Thompson, A., and Taylor, I. B. 1997.Structure and expression of a cDNA encoding a putative neoxanthin cleavage

50 D. BARTELS AND R. SUNKAR
enzyme (NCE) isolated from a wilt-related tomato (Lycopersicon esculentumMill) library. J. Exp. Bot. 47: 2111–2112.
Burg, M. B., Kwon, E. D., and Kultz, D. 1996. Osmotic regulation of geneexpression. FASEB J. 10: 1598–1606.
Busk, P. K. and Pages, M. 1998. Regulation of abscisic acid-induced transcrip-tion. Plant Mol. Biol. 37: 425–435.
Campalans, A., Pages, M., and Messeguer, R. 2001. Identification of differen-tially expressed genes by the cDNA-AFLP technique during dehydration ofalmond (Prunus amygdalus). Tree Physiol. 21: 633–643.
Cascardo, J. C. M., Almeida, R. S., Buzeli, R. A. A., Carolino, S. M. B., Otoni,W. C., and Fontes, E. P. B. 2000. The phosphorylation state and expression ofsoybean BiP isoforms are differentially regulated following abiotic stresses.J. Biol. Chem. 275: 14494–14500.
Chandler, P. and Robertson, M. 1994. Gene expression regulated by abscisicacid and its relation to stress tolerance. Annu. Rev. Plant Physiol. Plant Mol.Biol. 25: 113–141.
Chattopadhyay, M. K., Tiwari, B. S., Chattopadhyay, G., Bose, A., Sengupta, D.N., and Ghosh, B. 2002. Protective role of exogenous polyamines on salinity-stressed rice (Oryza sativa) plants. Physiol. Plant 116: 192–199.
Chen, T. H. H. and Murata, N. 2002. Enhancement of tolerance of abiotic stressby metabolic engineering of betaines and other compatible solutes. Curr.Opin. Plant Biol. 5: 250–257.
Chini, A., Grant, J. J., Seki, M., Shinozaki, K., and Loake, G. J. 2004. Drought tol-erance established by enhanced expression of the CC-NBS-LRR gene, ADR1,requires salicylic acid, EDS1 and ABI1. Plant J. 38: 810–822.
Choi, H. I., Hong, J. H., Ha, J. O., Kang, J. Y., and Kim, S. Y. 2000. ABFs,a family of ABA-responsive element binding factors. J. Biol. Chem. 275:1723–1730.
Chung, W. S., Lee, S. H., Kim, J. C., Heo, W. D., Kim, M. C., Park, C. Y., Park,H. C., Lim, C. O., Kim, W. B., Harper, J. F,. and Cho, M. J. 2000. Identificationof a calmodulin-regulated Ca2+-ATPase (SCA1) that is located in the plasmamembrane. Plant Cell 12: 1393–1407.
Coca, M., Almoguera, C., Thomas, T., and Jordano, J. 1996. Differentail regula-tion of small heat-shock genes in plants: Analysis of a water stress-inducibleand developmentally activated sunflower promoter. Plant Mol. Biol. 31: 863–876.
Cockcroft, C. E., den Boer, B. G. W., Healyy, J. M. S., and Murray, J. A. H.2000. Cyclin D control of plant growth rate in plants. Nature 405: 575–579.
Cohen, P. 1989. The structure and regulation of protein phosphatases. Annu. Rev.Biochem. 58: 453–508.
Conley, T. R., Sharp, R. E., and Walker, J. C. 1997. Water deficit rapidly stim-ulates the activity of a protein kinase in the elongation zone of the maizeprimary root. Plant Physiol. 113: 219–226.
Cosgrove, D. J. 1997. Relaxation in a high stress environment: The molecuklarbasis of extensible cell walls and cell enlargement. Plant Cell 9: 1031–1041.
Coursol, S., Fan, L. M., Le Stunff, H., Spiegel, S., Gilroy, S., and Assmann,S. 2003. Sphingolipid signalling in Arabidopsis guard cells involves het-erotrimeric G proteins. Nature 423: 651–654.
Crowe, J. H., Hoekstra, F. A., and Crowe, L. M. 1992. Anhydrobiosis. Annu RevPlant Physiol. 54: 579–599.
Cunningham, K. W. and Fink, G. R. 1996. Calcineurin inhibits VCX1-dependentH+/Ca2+ exchange and induced Ca2+ ATPases in Saccharomyces cerevisiae.Mol. Cell Biol. 16: 2226–2237.
de Carvalho, M. H., dArcy-Lameta, A., Roy-Macauley, H., Garcil, M., ElMaarouf, H., Pham-Thi, A.-T., and Zuily-Fodil, Y. 2001. Aspartic protease inleaves of common bean (Phaseolus vulgaris L.) and cowpea (Vigna unguic-ulata L. Wasp): Enzymatic activity, gene expression and relation to droughtsusceptibility. FEBS Lett. 492: 242–246.
Delauney, A. J., Hu, C.-A. A., Kavi Kishor, P. B., and Verma, D. P. S. 1993.Cloning of ornithine δ-aminotransferase cDNA from Vigna aconitifolia bytrans-complementation in Escherichia coli and regulation of proline biosyn-thesis. J. Biol. Chem. 268: 18673–18678.
Delauney, A. and Verma, D. P. S. 1993. Proline biosynthesis and osmoregulationin plants. Plant J. 4: 215–223.
de Nadal, E., Alepuz, P. A., and Posas, F. 2002. Dealing with osmostress throughMAP kinase activation. EMBO Reports 3: 735–740.
de Nadal, E., Clotet, J., Posas, F., Serrano, R., Gomez, N., and Arino, J. 1998.The yeast halotolerance determinant Hal3p is an inhibitory subunit of thePpz1p Ser/Thr protein phosphatase. Proc. Natl. Acad. Sci. U S A 95: 7357–7362.
den Boer, B. G. W. and Murray, J. A. H. 2000. Triggering the cell cycle in plants.Trends Cell Biol. 10: 245–250.
Deng, X., Philips, J., Meijer, A., Salamini, F., and Bartels, D. 2002. Charac-terization of five novel dehydration-responsive homeodomain leucine zippergenes from the resurrection plant Craterostigma plantagenium. Plant Mol.Biol. 49: 601–610.
De Wald, D. B., Torabinejad, J., Jones, C. A., Shope, J. C., Cangelosi, A. R.,Thompson, J. E., Prestwich, G. D., and Hama, H. 2001. Rapid accumulationof phosphatidylinositol 4,5-bisphosphate and inositol 1,4,5-trisphosphate cor-relates with calcium mobilization in salt-stressed Arabidopsis. Plant Physiol.126: 759–769.
Dietz, K.-J. and Arbinger, B. 1996. cDNA sequence and expression of subunitE of the vacuolar H+-ATPase in the inducible crassulacean acid metabolismplant Mesembryanthemum crystallinum. Biochim. Biophys. Acta 1281: 134–138.
Dietz, K.-J., Horhing, F., Konig, J., and Baier, M. 2002. The function of chloro-plast 2-cysteine peroxiredoxin in peroxide detoxification and its regulation.J. Exp. Bot. 53: 1321–1329.
Drøbak, B. K. and Watkins, P. A. C. 2000. Inositol(1,4,5)trisphosphate produc-tion in plant cells: an early response to salinity and hyperosmotic stress. FEBSLett. 481: 240–244.Dubouzet, J. G., Sakuma, Y., Ito, Y., Kasuga, M., Dubouzet, E., Miura, S., Seki,M., Shinozaki, K., and Yamaguchi-Shinozaki. K. (2003). OsDREB genes inrice, Oryza sativa L., encode transcription activators that function in drought-,high-salt- and cold-responsive gene expression. Plant J. 33: 751–763.
Dure, III L. 1993. Structural motifs in LEA proteins. In: Close T. J. Ed. PlantResponses to cellular dehydration during environmental stress: pp 91–103.American Society of Plant Physiologists, Rockville, MN.
Dure, L. 2001. Occurrence of a repeating Il-mer amino acid sequence motif indiverse organisms. Protein Pept. Lett. 8: 115–122.
Dure, III L., Crouch, M., Harada, J., Ho, T. H. D., Mundy, J., Quatrano, R. S.,Thomas, T., and Sung, Z. R. 1989. Common amino acid sequence domainsamong the LEA proteins of higher plants. Plant Mol. Biol. 12: 475–486.
Dure, III L., Greenway, S., and Galau, G. A. 1981. Developmental biochem-istry of cotton seed embryogenesis and germination XIV. Changing mRNApopulations as shown in vitro and in vivo protein synthesis. Biochemistry 20:4162–4168.
Ellul, P., Rios, G., Atares, A., Roig, L. A., Serrano, R., and Moreno, V. 2003.The expression of the Saccharomyces cerevisiae HAL1 gene increases salttolerance in transgenic watermelon [Citrullus lanatus (Thunb.) Matsun. &Nakai. ]. Theor. Appl. Genet. 107: 462–469.
El-Maarouf, H., Arcy-Lameta, A., Gareil, M., Zuily-Fodil, Y., and Pham-Thi,A.-T. 2001. Cloning and expression under drought of cDNAs coding for twoPI-PLCs in cowpea leaves. Plant Physiol. Biochem. 39: 167–172.
El-Maarouf, H., Zuily-Fodil, Y., Gareil, M., Arcy-Lameta, A., and Pham-Thi,A.-T. 1999. Enzymatic activity and gene expression under water stress ofphospholipase D in two cultivars of Vigna unguiculata L. Walp. differing indrought tolerance. Plant Mol. Biol. 39: 1257–1265.
Espinosa-Ruiz, A., Belles, J. M., Serrano, R., and Culianez-Macia, F. A. 1999.Arabidopsis thaliana AtHAL3: A flavoprotein related to salt and osmotictolerance and plant growth. Plant J. 20: 529–539.
Eun, S. O. and Lee, Y. 1997. Actin filaments of guard cells are reorganized inresponse to light and abscisic acid. Plant Physiol. 115: 1491–1498.
Finkelstein, R. R., Gampala, S. S., and Rock, C. D. 2002. Abscisic acid signalingin seeds and seedlings. Plant Cell 14 Suppl: S15–S45.
Flores, H. E. and Galston, A. W. 1982. Osmotic stress-induced polyamine accu-mulation in cereal leaves. I. Physiological parameters of the response. PlantPhysiol. 75: 102–109.

DROUGHT AND SALT TOLERANCE IN PLANTS 51
Forsthoefel, N. R., Cushman, M. A. F., Ostrem, J. A., and Cushman, J. C.1998. Induction of a cysteine protease cDNA from Mesembryanthemum crys-tallinum leaves by environmental stress and plant growth regulators. Plant Sci.136: 195–206.
Foyer, C. H., Descourvieres, P., and Kunert, K. J. 1994. Protection against oxygenradicals, an important defense mechanism studied in transgenic plants. PlantCell Environ. 17: 507–523.
Frandsen, G., Muller-Uri, F., Nielsen, M., Mundy, J., and Skriver, K. 1996.Novel plant Ca2+-binding protein expressed in response to abscisic acid andosmotic stress. J. Biol. Chem. 271: 343–348.
Frank, W., Munnik, T., Kerkmann, K., Salamini, F., and Bartels, D. 2000.Water deficit triggers phospholipase D activity in the resurrection plantCraterostigma plantagineum. Plant Cell 12: 111–123.
Fray, R. G., Wallace, A., Grierson, D., and Lycett, G. W. 1994. Nucleotidesequence and expression of a ripening and water stress-related cDNA fromtomato with homology to the MIP class of membrane channel proteins. PlantMol. Biol. 24: 539–543.
Fukuda, A., Nakamura, A., Tagiri, A., Tanaka, H., Miyao, A., Hirochika, H., andTanaka, Y. 2004. Function, intracellular localization and the importance in salttolerance of a vacuolar Na+/H+ antiporter from rice. Plant Cell Physiol. 45:146–159.
Fukada-Tanaka, S., Inagaki, Y., Yamaguchi, T., Saito, N., and Iida, S. 2000.Colour-enhancing protein in blue petals. Nature 407: 581.
Fukushima, E., Arata, Y., Endo, T., Sonnewald, U., and Sato, F. 2001. Improvedsalt tolerance of transgenic tobacco expressing apoplastic yeast-derived in-vertase. Plant Cell Physiol. 42: 245–249.
Furini, A., Koncz, C., Salamini, F., and Bartels, D. 1997. High level transcriptionof a member of a repeated gene family confers dehydration tolerance to callustissue of Craterostigma plantagineum. EMBO J. 16: 3599–3608.
Gago, G. M., Almoguera, C., Jordano, J., Gonzalez, D. H., and Chan, R. L. 2002.Hahb-4, a homeobox-leucine zipper gene potentially involved in abscisicacid-dependent responses to water stress in sunflower. Plant Cell Environ.25: 633–640.
Galau, G. A, Hughes, D. W., and Dure, III L. 1986. Abscisic acid induction ofcloned cotton late embryogenesis-abundant (Lea) mRNAs. Plant Mol. Biol.7: 155–170.
Gampala, S. S., Finkelstein, R. R., Sun. S. S., and Rock, C. D. 2002. ABI5interacts with abscisic acid signalling effectors in rice protoplasts. J. Biol.Chem. 277: 1689–1694.
Gampala, S. S., Hagenbeek, D., and Rock, C. D. 2001. Functional interactionsof lanthanium and phospholipase D with the abscisic acid signalling effectorsVP1 and ABI1-1 in rice protoplasts. J. Biol. Chem. 276: 9855–9860.
Garg, A. K., Kim, J-K., Owens, T. G., Ranwala, A. P., Choi, Y. D., Kochian, L.V., and Wu, R. J. 2002. Trehalose accumulation in rice plants confers hightolerance levels to different abiotic stresses. Proc. Nat. Acad. Sci USA 99:15898–15903.
Gaxiola, R. A., Rao, R., Sherman, A., Grisafi, P., Alper, S. L., and Fink, G. R.1999. The Arabidopsis thaliana proton transporters, Atnhx1 and Avp1, canfunction in cation detoxification in yeast. Proc. Natl. Acad. Sci. USA 96:1480–1485.
Gaxiola, R., Li, J., Undurraga, S., Dang, L. M., Allen, G. J., Alper, S. L., andFink, G. R. 2001. Drought- and salt-tolerant plants result from overexpressionof the AVP1 H+ pump. Proc. Nat. Acad. Sci. USA 98: 11444–11449.
Geisler, M., Axelsen, K. B., Harper, J. F., and Palmgren, M. G. (2000a). Molec-ular aspects of higher plant P-type Ca(2+)-ATPases. Biochim. Biophys. Acta1465: 52–78.
Geisler, M., Frangne, N., Gomes, E., Matinoia, E., and Palmgreen, M. G.(2000b). The ACA4 gene of Arabidopsis encodes a vacuolar membrane cal-cium pump that improves salt tolerance in yeast. Plant Physiol. 124: 1814–1827.
Gilmour, S. J., Seblot, A. M., Salazar, M. P., Everard, J. D., and Thomashow,M. F. 2000. Overexpression of the Arabidopsis CBF3 transcriptional activatormimics multiple biochemical changes associated with cold acclimation. PlantPhysiol. 124: 1854–1865.
Golldack, D. and Dietz, K-J. 2001. Salt-induced expression of the vacuolar H+-ATPase in the common ice plant is developmentally controlled and tissuespecific. Plant Physiol. 125: 1643–1654
Guerroro, F. and Mullet, J. E. 1986. Increased abscisic acid biosynthesis duringplant dehydration requires transcription. Plant Physiol. 80: 588–591.
Guerrero, F. D., Jones, J. T., and Mullet, J. E. 1990. Turgor responsive genetranscription and RNA levels increase rapidly when pea shoots are wilted:sequence and expression of three inducible genes. Plant Mol. Biol. 15: 11–26.
Guiltinan, M. J., Marcotte, W. R., and Quatrano, R. S. 1990. A plant leucinezipper protein recognizes an abscisic acid responsive element. Science 25:267–271.
Guo, Y., Halfter, U., Ishitani, M., and Zhu, J.-K. 2001. Molecular characterizationof functional domains in the protein kinase SOS2 that is required for plantsalt tolerance. Plant Cell 13: 1383–1400.
Gupta, A. S., Heinen, J. L., Holaday, A. S., Burke, J. J., and Allen, R. D. 1993.Increased resistance to oxidative stress in transgenic plants that overexpresschloroplastic Cu/Zn superoxide dismutase. Proc. Natl. Acad. Sci. USA 90:1629–1633.
Haake, V., Cook, D. Riechmann, J. L., Pineda, O., Thomashow, M. F., and Zhang,J. Z. 2002. Transcription factor CBF4 is a regulator of drought adaptation inArabidopsis. Plant Physiol. 130: 639–648.
Halford, N. G. and Hardie, D. G. 1998. SNF-1 related protein kinases: Globalregulators of carbon metabolism in plants? Plant Mol. Biol. 37: 735–748.
Halfter, U., Ishitani, M., and Zhu, J. -K. 2000. The Arabidopsis SOS2 proteinkinase physically interacts with and is activated by the calcium-binding proteinSOS3. Proc. Natl. Acad. Sci. USA 97: 3735–3740.
Halliwell, B. and Gutteridge, J. M. C. 1999. Free Radicals in Biology andMedicine: 3rd ed., Oxford University Press, New York.
Hasegawa, P. M., Bressan, R., Zhu, J.-K., and Bohnert, H.-J. 2000. Plant cellularand molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol.Biol. 51: 463–499.
Haushul, K., Andersson, B., and Adamska, I. 2001. A chloroplast DegP2 pro-tease performs the primary cleavage of the photodamaged D1 protein in plantphotosystem II. EMBO J. 20: 713–722.
Hayashi, H., Alia, Mustardy, L., Deshnium, P., Ida, M., and Murata, N. 1997.Transformation of Arabidopsis thaliana with the codA gene for choline oxi-dase; accumulation of glycinebetaine and enhanced tolerance to salt and coldstress. Plant J. 12: 133–142.
Himmelbach, A., Hoffmann, T., Leube, M., Hohener, B., and Grill, E. 2002.Homeodomain protein ATHB6 is a target of protein phosphatase ABI1and regulates hormone responses in Arabidopsis. EMBO J. 21: 3029–3038.
Hirayama, T., Ohto, C., Mizoguchi, T., and Shinozaki, K. 1995. A gene encodinga phosphotidylinositol-specific phospholipase C is induced by dehydrationand salt stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 92: 3903–3907.
Hoekstra, F. A., Golovina, E. A., and Buitink, J. 2001. Mechanisms of plantdesiccation tolerance. Trends Plant Sci. 6: 431–438.
Hong, Z., Lakkineni, K., Zhang, Z., and Verma, D. P. S. 2000. Removal offeedback inhibition of �1-pyrroline-5-carboxylate synthetase results in in-creased proline accumulation and protection of plants from osmotic stress.Plant Physiol. 122: 1129–1136.
Hoshida, H., Tanaka, Y., Hibino, T., Hayashi, Y., Tanaka, A., Takabe, T., andTakabe, T. 2000. Enhanced tolerance to salt stress in transgenic rice thatoverexpresses chloroplast glutamine synthetase. Plant Mol. Biol. 43: 103–111.
Hsieh, T.-H., Lee, J.-T., Charng, Y.-Y., and Chan, M.-T. 2002. Tomato plantsectopically expressing Arabidopsis CBF1 show enhanced resistance to waterdeficit stress. Plant Physiol. 130: 618–626.
Huang, J., Hirji, R., Adam, L., Rozwadowski, K. L., Hammerlindl, J. K., Keller,W. A., and Selvaraj, G. 2000. Genetic engineering of glycinebetaine produc-tion toward enhancing stress tolerance in plants: metabolic limitations. PlantPhysiol. 122: 747–756.

52 D. BARTELS AND R. SUNKAR
Huang, Y., Li, H., Gupta, R., Morris, P. C., Luan, S., and Kieber, J. J. 2000.ATMPK4, an Arabidopsis homolog of mitogen-activated protein kinase, is ac-tivated in vitro by AtMEK1 through threonine phosphorylation. Plant Physiol.122: 1301–1310.
Hugovieux, V., Kwak, J. M., and Schroeder, J. I. 2001. An mRNA cap bindingprotein, ABH1, modulates early abscisic acid signal transduction. Cell 106:477–487.
Hwang, I. and Goodman, H. M. 1995. An Arabidopsis thaliana root-specifickinase homolog is induced by dehydration, ABA, and NaCl. Plant J. 8: 37–43.
Hwang, J. and Lee, Y. 2001. Abscisic acid-induced actin reorganization in guardcells of dayflower is mediated by cytosolic calcium levels and by proteinkinase and protein phosphatase activities. Plant Physiol. 125: 2120–2128.
Ichimura, K., Mizoguchi, T., Yoshida, R., Yuasa, T., and Shinozaki, K. 2000.Various abiotic stresses rapidly activate Arabidopsis MAP kinases AtMAPK4and AtMPK6. Plant J. 24: 655–665.
Ichimura, K., Shinozaki, K., Tina, G., Sheen, J., Henry, Y. et al. 2002. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends PlantSci. 7: 301–308.
Ingram, J. and Bartels, D. 1996. The molecular basis of dehydration tolerancein plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47: 377–403.
Ingram, J., Chandler, J. W., Gallagher, L., Salamini, F., and Bartels, D. 1997.Analysis of cDNA clones encoding sucrose-phosphate synthase in relation tosugar interconversions associated with dehydration in the resurrection plantCraterostigma plantagineum Hochst. Plant Physiol. 115: 113–121.
Iturriga, G., Leyns, L., Villegas, A., Gharabeih, R., Salamini, F., and Bartels,D. 1996. A family of novel myb-related genes from the resurrection plantCraterostigma plantagineum are specifically expressed in callus and roots inresponse to ABA or desiccation. Plant Mol. Biol. 25: 707–716.
Iuchi, S., Kobayashi, M., Taji, T., Naramoto, M., Seki, M., Kato, T., Tabata, S.,Kakubari, Y., Yamaguchi-Shinozaki, K., and Shinozaki, K. 2001. Regulationof drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxy-genase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J.27: 325–333.
Iuchi, S., Kobayashi, M., Yamaguchi-Shinozaki, K., and Shinozaki, K. 2000.A stress-inducible gene for 9-cis-epoxycarotenoid dioxygenase involved inabscisic acid biosynthesis under water stress in drought tolerant cow pea.Plant Physiol. 123: 553–562.
Iwasaki, T., Kiyosue, T., Yamaguchi-Shinozaki, K., and Shinozaki, K. 1997.The dehydration-inducible rd17 (cor47) gene and its promoter region in Ara-bidopsis thaliana. Plant Physiol. 115: 1287–1294.
Izawa, T., Foster, R., and Chua, N.-H. 1993. Plant bZIP Protein DNA BindingSpecificity J. Mol. Biol. 230: 1131–1144.
Jacob, T., Ritchie, S., Assmann, S. M., and Gilroy, S. 1999. Abscisic acid signaltransduction in guard cells is mediated by phospholipase D activity. Proc.Natl. Acad. Sci. USA 95: 2697–2702.
Jacoby, M., Weisshaar, B., Droge-Laser, W., Vicente-Carbajosa, J., Tiedemann,J., Kroj, T., and Parcy, F. 2002. bZIP transcription factors in Arabidopsis.Trends Plant Sci. 7: 106–111.
Jacoby, T., Flanagan, H., Faykin, A., Seto, A. G., Mattison, C., and Ota, I.1997. Two protein-tyrosine phosphatases inactivate the osmotic stress re-sponse pathway in yeast by targeting the mitogen activated protein kinase,HOG1. J. Biol. Chem. 272: 17749–17755.
Jang, H. J., Pih, K. T., Kang, S. G., Lim, J. H., Jin, J. B., Piao, H. L., and Hwang,I. 1998. Molecular cloning of a novel Ca2+− binding protein that is inducedby NaCl stress. Plant Mol. Biol. 37: 839–847.
Jang, I.-C., Oh, S.-J., Seo, J.-S., Choi, W.-B., Song, S. I., Kim, C. H., Kim, Y.S., Seo, H.-S., Choi, Y. D., Nahm, B. H., and Kim, J.-K. 2003. Expression ofa bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphatesynthase and trehalose-6-phosphate phosphatase in transgenic rice plants in-creases trehalose accumulation and abiotic stress tolerance without stuntinggrowth. Plant Physiol. 131: 516–524.
Johansson, I., Karlsson, M., Shukla, V. K., Chrispeels, M. J., Larsson, C.,and Kjellbom, P. 1998. Water transport activity of the plasma membrane
aquaporin PM28A is regulated by phosphorylation. Plant Cell 25: 451–460.
Jonak, C., Kiegerl, S., Ligterink, W., Barker, P. J., Huskisson, N. S., and Hirt, H.1996. Stress signaling in plants: A mitogen-activated protein kinase pathwayis activated by cold and drought. Proc. Natl. Acad. Sci. USA 93: 11274–11279.
Jones, L. and McQueen-Mason, S. 2004. A role for expansins in dehydrationand rehydration of the resurrection plant Craterostigma plantagineum. FEBSLett. 559: 61–65.
Kaldenhoff, R., Grote, K., Zhu, J. J., and Zimmermann, U. 1998. Significance ofplasmalemma aquaporins for water-transport in Arabidopsis thaliana. PlantJ. 14: 121–128.
Kang, J., Choi, H., Im, M., and Kim, S. Y. 2002. Arabidopsis basic leucine zipperproteins that mediate stress-responsive abscisic acid signaling. Plant Cell 14:343–357.
Kao, C. Y., Cocciolone, S. M., Vasil, I. K., and McCarty, D. R. 1996. Localizationand interaction of the cis-acting elements for abscisic acid, VIVIPAROUS1,and light activation of the C1 gene of maize. Plant Cell 8: 1171–1179.
Kasinathan, V. and Wingler, A. 2002. Effect of reduced arginine decarboxylaseactivity on salt tolerance and on polyamine formation during salt stress inArabidopsis thaliana. Physiol. Plant 121: 101–107.
Kasuga, M., Liu, Q., Miura, S., Yamaguchi–Shinozaki, K., and Shinozaki, K.1999. Improving plant drought, salt, and freezing tolerance by gene transferof a single stress-inducible transcription factor. Nat. Biotechnol. 17: 287–291.
Katagiri, T., Takahashi, S., and Shinozaki, K. 2001. Involvement of a novelArabidopsis phospholipase D, AtPLD in dehydration-inducible accumulationof phosphatidic acid in stress signalling. Plant J. 26: 595–605.
KaviKishor, P. B., Hong, Z., Miao, G.-H., Hu, C.-A. A., and Verma, D. P. S. 1995.Over-expression of �-pyrroline-5-carboxylate synthetase increases prolineproduction and confers osmotolerance in transgenic plants. Plant Physiol.108: 1387–1394.
Kawasaki, S., Borchert, C., Deyholos, M., Wang, H., Brazille, S., Kawai, K.,Galbraith, D., and Bohnert, H. 2001. Gene expression profiles during theinitial phase of salt stress in rice. Plant Cell 13: 889–905.
Kende, H. and Zeevaart, J. 1997. The five “classical” plant hormones. Plant Cell9: 1197–1210.
Khanna-Chopra, R., Srivalli, B., and Ahlawat, Y. 1999. Drought induces manyforms of cysteine proteases not observed during natural senescence. Biochem.Biophys. Res. Commun. 255: 324–327.
Kiegerl, S., Cardinale, F., Siligan, C., Gross, A., Baudouin, E., Liwosz, A.,Ekloff, S., Till, S., Bogre, L., Hirt, H., and Meskiene, I. 2000. SIMKK, amitogen-activated protein kinase (MAPK) kinase is a specific activator of thesalt stress-induced MAPK, SIMK. Plant Cell 12: 2247–2258.
Kiegle, E., Moore, C. A., Haseloff, J., Tester, M. A., and Knight, M. R. 2000. Cell-type-specific calcium responses to drought, salt and cold in the Arabidopsisroot. Plant J. 23: 267–278.
Kirch, H.-H., Nair, A., and Bartels, D. 2001. Novel ABA- and dehydration-inducible aldehyde dehydrogenase genes isolated from the resurrection plantCraterostigma plantagineum and Arabidopsis thaliana. Plant J. 28: 555–567.
Kirsch, M., An, Z., Viereck, R., Low, R., and Rausch, T. 1996. Salt stress inducesan increased expression of V-type H(+)-ATPase in mature sugar beet leaves.Plant Mol. Biol. 32: 543–547.
Kizis, D. and Pages, M. 2002. Maize DRE-binding proteins DBF1 and DBF2are involved in rab17 regulation through the drought-responsive element inan ABA-dependent pathway. Plant J. 30: 679–689.
Kjellbom, P., Larsson, C., Johansson, I., Karlsson, M., and Johanson, U. 1999.Aquaporins and water homeostasis in plants. Trends Plant Sci. 25: 308–314.
Knight, H. and Knight, M. R. 2001. Abiotic stress signalling pathways: Speci-ficity and cross-talk. Trends Plant Sci. 6: 262–267.
Knight, H., Trewavas, A. J., and Knight, M. R. 1997. Calcium signalling inArabidopsis thaliana responding to drought and salinity. Plant J. 12: 1067–1078.
Kobayashi, Y., Yamamoto, S., Minami, H., Kagaya, Y., and Hattori, T. 2004. Dif-ferential activation of the rice sucrose nonfermenting1-related protein kinase2family by hyperosmotic stress and abscisic acid. Plant Cell 16: 1163–1177.

DROUGHT AND SALT TOLERANCE IN PLANTS 53
Kobrinsky, E., Mirshahi, T., Zhang, H, Jin, T., and Logothetis, D. E. 2000.Receptor-mediated hydrolysis of plasma membrane messenger PIP2 leads toK+-current desensitization. Nat. Cell. Bio. 2: 507–514.
Koizumi, M., Yamaguchi-Shinozaki, K., Tsuji, H., and Shinozaki, K. 1993.Structure and expression of two genes that encode distinct drought-induciblecysteine proteinases in Arabidopsis thaliana. Gene 129: 175–182.
Kopka, J., Pical, C., Gray, J. E., and Muller-Rober, B. 1998. Molecular andenzymatic characterization of three phosphoinositide-specific phospholipaseC isoforms from potato. Plant Physiol. 116: 239–250.
Kranner, I., Beckett, R. P., Wornik, S., Zorn, M., and Pfeifhofer, H. W. 2002.Revival of resurrection plant correlates with its antioxidant status. Plant J.31: 13–24.
Kudla, J., Xu, Q., Harter, K., Gruissem, W., and Luan, S. 1999. Genes forcalcineurin B-like proteins in Arabidopsis are differentially regulated by stresssignals. Proc. Natl. Acad. Sci. USA 96: 4718–4723.
Kyriakis, J. M. and Avruch, J. 2001. Mammalian mitogen-activated proteinkinase signal transduction pathways activated by stress and inflammation.Physiol. Rev. 81: 807–869.
Lee, G., Pokala, N., and Vierling, E. 1995. Structure and in vitro molecularchaperone activity of cytosolic small heat shock proteins from pea. J. Biol.Chem. 270: 10432–10438.
Lee, Y. H. and Chun, J. Y. 1998. A new homeodomain-leucin zipper gene fromArabidopsis thaliana induced by water stress and abscisic acid. Plant Mol.Biol. 37: 377–384.
Lehr, A., Kirsch, M., Viereck, R., Schiemann, J., and Rausch, T. 1999. cDNA andgenomic cloning of sugar beet V-type H+-ATPase subunit A and c isoforms:Evidence for coordinate expression during plant development and coordinateinduction in response to high salinity. Plant Mol. Biol. 39: 463–75.
Lemichez, E., Wu, Y., Sanchez, J.-P., Mettouchi, A., Mathur, J., and Chua, N.-H.2001. Inactivation of AtRac1 by abscisic acid is essential for stomatal closure.Genes Dev. 15: 1808–1816.
Leung, J., Bouvier-Durand, M., Morris, P. C., Guerrier, D., Chedfor, F., andGiraudat, J. 1994. Arabidopsis ABA-response gene ABI1: Features of acalcium-modulated protein phosphatase. Science 264: 1448–1452.
Leung, J. and Giraudat, J. 1998. Abscisic acid signal transduction. Annu. Rev.Plant Physiol. Plant Mol. Biol. 25: 199–221.
Leymann, B., Geelen, D., Quintero, F. J., and Blatt, M. R. 1999. A tobaccosyntaxin with a role in hormonal control of guard cell ion channels. Science283: 537–540.
Li, J., Kinoshita, T., Pandey, S., Ng, C. K-Y., Gygl, S. P., Shimazaki, K-I., andAssmann, S. M. 2002. Modulation of an RNA-binding protein by abscisic-acid-activated protein kinase. Nature 418: 793–797.
Li, J., Wang, X. -Q., Watson, M. B., and Assmann, S. M. 2000. Regulationof abscisic acid-induced stomatal closure and anion channels by guard cellAAPK kinase. Science 287: 300–303.
Lim, Y. S., Cha, M-K., Kim, H. K., Uhm, T. B., Park, J. W., Kim, K., and Kim, I.H. 1993. Removal of hydrogen peroxide and hydroxyl radical by thiol-specificantioxidant protein as a possible role in vivo. Biochem. Biophy. Res. Comm.192: 273–280.
Liu, J., Ishitani, M., Halfter, U., Kim, C.-S., and Zhu, J.-K. 2000. The Arabidopsisthaliana SOS2 gene encodes a protein kinase that is required for salt tolerance.Proc. Natl. Acad. Sci. USA 97: 3730–3734.
Liu, J. and Zhu, J.-K. 1997. An Arabidopsis mutant that requires increasedcalcium for potassium nutrition and salt tolerance. Proc. Natl. Acad. Sci. USA94: 14960–14964.
Liu, J. and Zhu J.-K. 1998. A calcium sensor homolog required for plant salttolerance. Science 280: 1943–1945.
Liu, Q., Kasuga, M., Sakuma, Y., Abe, H., Miura, S., Yamaguchi-Shinozaki, K.,and Shinozaki, K. 1998. Two transcription factors, DREB1 and DREB2, withan EREBP/AP2 DNA binding domain separate two cellular signal transduc-tion pathways in drought- and low-temperature-responsive gene expression,respectively, in Arabidopsis. Plant Cell 10: 1391–1406.
Lu, C. and Fedoroff, N. 2000. A mutation in the Arabidopsis HYL1 gene en-coding a dsRNA binding protein affects responses to abscisic acid, auxin, andcytokinin. Plant Cell 12: 2351–2366.
Luan, S., Kudla, J., Rodriguez-Concepcion, M., Yalovsky, S., and Gruissem,W. 2002. Calmodulins and calcineurin-B like proteins: Calcium sensors forspecific signal response coupling in plants. Plant Cell 14: S389–S400.
Luttge, U. and Ratajczak, R. 1997. The physiology, biochemistry and molecularbiology of the plant vacuolar ATPase. Adv. Bot. Res. 25: 253–296.
MacRobbie, E. A. C. 2002. Evidence for a role for protein tyrosine phosphatasein the cytosol of ion release from the guard cell vacuole in stomatal closure.Proc. Natl. Acad. Sci. USA 99: 11963–11968.
Mani, S., van de Cotte, B., Van Montagu, M., and Verbruggen, N. 2002. Al-tered levels of proline dehydrogenase cause hypersensitivity to proline andits analogs in Arabidopsis. Plant Physiol. 128: 73–83.
Mansour, M. M. F. 1998. Protection of plasma membrane of onion epidermalcells by glycinebetaine and proline against NaCl stress. Plant Physio. Biochem36: 767–772.
Mata, C. G. and Lamattina, L. 2001. Nitric oxide induces stomatal closure andenhances the adaptive plant responses against drought stress. Plant Physiol.126: 1196–1204.
Mazel, A., Leshem, Y., Tiwari, B. S., and Levin, A. 2004. Induction of saltand osmotic stress tolerance by overexpression of an intracellular vesicletrafficking protein AtRab7 (AtRabG3e). Plant Physiol. 134: 118–128.
McCue, K. F., and Hanson, A. D. 1990. Drought and salt tolerance: towardsunderstanding and application. Trends Biotech. 8: 358–362.
McKersie, B. D., Bowley, S. R., Harjanto, E., and Leprince, O. 1996. Water-deficit tolerance and field performance of transgenic alfalfa overexpressingsuperoxide dismutase. Plant Physiol. 111: 1177–1181.
McNeil, S. D., Rhodes, D., Russell, B. L., Nuccio, M. L., Shachar-Hill, Y.,and Hanson A. D. 2000. Metabolic modeling identifies key constraints on anengineered glycine betaine synthesis pathway in tobacco. Plant Physiol. 124:153–162.
Mendoza, I., Rubio, F., Rodriguez-Navarro, A., and Pardo, J. M. 1994. The pro-tein phosphatase calcineurin is essential for NaCl tolerance of Saccharomycescerevisiae. J. Biol. Chem. 269: 8792–8796.
Mestichelli, L. J., Gupta, R. N., and Spenser, I. D. 1979. The biosynthetic routefrom ornithine to proline. J. Biol. Chem. 254: 640–647.
Michel, E., Salamini, R., Barels, E., Dale, P., Baga, M., and Szalay, A. 1993.Analysis of a desiccation and ABA-responsive promoter isolated from theresurrection plant Craterostigma plantagineum. Plant J. 4: 29–40.
Michel, D. Furini, A., Salamini, F., and Bartels, D. 1994. Structure and regula-tion of an ABA- and desiccation-responsive gene from the resurrection plantCraterostigma plantagineum. Plant Mol. Biol. 24: 549–560.
Mikami, K., Katagiri, T., Luchi, S., Yamaguchi-Shinozaki, K., and Shinozaki, K.1998. A gene encoding phosphatidylinositol 4-phosphate 5-kinase is inducedby water stress and abscisic acid in Arabidopsis thaliana. Plant J. 15: 563–568.
Mikolajczyk, M., Awotunde, O. S., Muszynska, G., Klessig, D. F., andDobrowolska, G. 2000. Osmotic stress induced rapid activation of a sali-cylic acid-induced protein kinase and a homolog of protein kinase ASK1 intobacco cells. Plant Cell 12: 165–178.
Mittler, R. 2002. Oxidative stress, antioxidants and stress tolerance. Trends PlantSci. 7: 405–410.
Miyazaki, S., Koga, R., Bohnert, H. J., and Fukuhara, T. 1999. Tissue- andenvironmental response-specific expression of 10PP2C transcripts in Mesem-bryanthemum crystallinum. Mol. Gen. Genet. 261: 307–316.
Mizoguchi, T., Irie, K., Hirayama, T., Hayashida, N., Yamaguchi-Shinozaki,K., Matsumoto, K., and Shinozaki, K. 1996. A gene encoding a mitogen-activated protein kinase kinase kinase is induced simultaneously with genesfor a mitogen-activated protein kinase and an S6 ribosomal protein kinase bytouch, cold, and water stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci.USA 93: 765–769.
Moskovitz, J., Berlett, B. S., Poston, J. M., and Stadtman, E. R. 1999. Methion-ine sulfoxide reductase in antioxidant defense. Methods Enzymol. 300: 239–244.
Mowla, S. B., Thomson, J., Farrant, J. M., and Mundree, S. G. 2002. A novelstress-inducible antioxidant enzyme identified from the resurrection plantXerophyta viscosa baker. Planta 215: 716–726.

54 D. BARTELS AND R. SUNKAR
Mundree, S. G., Whittaker, A., Thomson, J., and Farrant, J. M. 2000. An al-dose reductase homolog from the resurrection plant Xerophyta viscosa Baker.Planta 211: 693–700.
Mundy, J., Yamaguchi-Shinozaki, K., and Chua, N. H. 1990. Nuclear proteinsbind conserved elements in the abscisic acid-responsive promoter of a ricerab gene. Proc. Natl. Acad. Sci. USA 87: 1406–1410.
Munnik, T. 2001. Phosphatidic acid: an emerging plant lipid second messenger.Trends Plant Sci. 6: 227–233.
Munnik, T., Ligternick, W., Meskiene, I., Calderini, O., Beyerly, J., Musgrave,A., and Hirt, H. 1999. Distinct osmo-sensing protein kinase pathways areinvolved in signalling moderate and severe hyper-osmotic stress. Plant J. 20:381–388.
Munnik, T. and Meijer, H. J. G. 2001. Osmotic stress activates distinct lipid andMAPK signalling pathways in plants. FEBS Lett. 498: 172–178.
Munns, R. 2002. Comparative physiology of salt and water stress. Plant CellEnviron. 25: 239–250.
Munns, R., Passioura, J. B., Guo, J., Ehazen, O., and Cramer, G. R. 2000. Waterrelations and leaf expansion: Importance of timing. J. Exp. Bot. 51: 1495–1504.
Murata, Y., Pei, Z. M., Mori, I. C., and Schroeder, J. I. 2001. Abscisic acid ac-tivation of plasma membrane Ca2+ channels in guard cells require cytosolicNAD(P)H and is differentially disrupted upstream and downstream of reac-tive oxygen species production in abi1-1 and abi2-1 protein phosphatase 2Cmutants. Plant Cell 13: 2513–2523.
Mustilli, A.-C., Merlot, S., Vavasseur, A., Fenzi, F., and Giraudat, J. 2002. Ara-bidopsis OST1 protein kinase mediates the regulation of stomatal aperture byabscisic acid and acts upstream of reactive oxygen species production. PlantCell 14: 3089–3099.
Nakashima, K., Kiyosue, T., Yamaguchi-Shinozaki, K., and Shinozaki, K. 1997.A nuclear gene, erd1, encoding a chloroplast-targeted Clp protease regulatorysubunit homolog is not only induced by water stress but also developmentallyup-regulated during senescence in Arabidopsis thaliana. Plant J. 12: 851–861.
Nakashima, K., Shinwari, Z. K., Sakuma, Y., Seki, M., Miura, S., Shinozaki,K., and Yamaguchi-Shinozaki, K. 2000. Organization and expression oftwo Arabidopsis DREB2 genes encoding DRE-binding proteins involved indehydration- and high-salinity-responsive gene expression. Plant Mol. Biol.42: 657–665.
Nanjo, T., Kobayashi, M., Yoshiba, Y., Kakubari, Y., Yamaguchi-Shinozaki,K., and Shinozaki, K. 1999. Antisense suppression of proline degradationimproves tolerance to freezing and salinity in Arabidopsis thaliana. FEBSLett. 461: 205–210.
Narasimhan, M. L., Binzel, M. L., Perez-Prat, E., Chen, Z., Nelson, D. E., Singh,N. K., Bressan, R. A., and Hasegawa, P. M. 1991. NaCl regulation of tonoplastATPase 70-kilodalton subunit mRNA in tobacco cells. Plant Physiol. 97: 562–568.
Neill, S. J., Burnett, E. C., Desikan, R., and Hancock, J. T. 1998. Cloning ofa wilt-responsive cDNA from an Arabidopsis thaliana suspension culturecDNA library that encodes a putative 9-cis-epoxycarotenoid dioxygenase. J.Exp. Bot. 49: 1893–1894.
Ng, C. K.-Y., Carr, K., McAinsh, M. R., Powell, B., and Hetherington, A. M.2001. Drought-induced guard cell signal transduction involves sphingosine-1-phosphate. Nature 410: 596–599.
Niu, X., Bressan, R. A., Hasegawa, P. M., and Pardo, J. M. 1995. Ion homeostasisin NaCl stress environments. Plant Physiol. 109: 735–742.
Nonami, H. and Boyer, J. S. 1990. Primary events regulating stem growth at lowwater potentials. Plant Physiol. 94: 1601–1609.
Novillo, F., Alonso, J. M., Ecker, J. R., and Salinas, J. 2004. CBF2/DREB1C isa negative regulator of CBF1/DREB1B and CBF3/DREB1A expression andplays a central role in stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci.USA 101: 3985–3990.
Nozawa, A., Koizumi, N., and Sano, H. 2001. An Arabidopsis SNF-1relatedprotein kinase, atsr1, interacts with a calcium-binding protein, AtCBL2, ofwhich transcripts respond to light. Plant Cell Physiol. 42: 976–981.
Nuccio, M. L., McNeil, S. D., Ziemak, M. J., Hanson, A. D., Jain, R. K., andSelvaraj, G. 2000. Choline import into chloroplasts limits glycine betaine
synthesis in tobacco: analysis of plants engineered with a chloroplastic or acytosolic pathway. Metab Eng. 2: 300–311.
Nuccio, M. L., Rhodes, D., McNeil, S. D., and Hanson, A. D. 1999. Metabolicengineering of plants for osmotic stress resistance. Curr. Opin. Plant Biol. 2:128–134.
Nuccio, M. L., Russell, B. L., Nolte, K. D., Rathinasabapathi, B., Gage, D. A.,and Hanson, A. D. 1998. The endogenous choline supply limits glycine be-taine synthesis in transgenic tobacco expressing choline monooxygenase.Plant J. 16: 101–110.
Oberschall, A., Deak, M., Torok, K., Saa, L., Vass, I., Kovacs, I., Feher, A.,Dudits, D., and Horvath, G. V. 2000. A novel aldose/aldehyde reductase pro-tects transgenic plants against lipid peroxidation under chemical and droughtstresses. Plant J. 24: 437–446.
Ohta, M., Hayashi, Y., Nakashima, A., Hamada, A., Tanaka, A., Nakamura,T., and Hayakawa, T. 2002. Introduction of a Na+/H+ antiporter gene fromAtriplex gmelini confers salt tolerance to rice. FEBS Lett. 532: 279–282.
Osorio, J., Osorio, M. L., Chaves, M. M., and Pereira, J. S. 1998. Water deficitsare more important in delaying growth than in changing patterns of carbonallocation in Eucalyptus globulus. Tree Physiol. 18: 363–373.
Oztur, Z. N., Talame, V., Deyholds, M., Michalowski, C. B., Galbraith, D. W.,Gozukirmizi, N., Tubrosa, R., and Bohnert, H. J. 2002. Monitoring large-scale changes in transcript abundance in drought- and salt-stressed barley.Plant Mol. Biol. 48: 551–573.
Pandey, S. and Assmann, S. 2004. The Arabidopsis putative G protein-coupledreceptor GCR1 interacts with the G protein α subunit GPA1 and regulatesabscisic acid signalling. Plant Cell 16: 1616–1632.
Pandey, S., Tiwari, S. B., Tyagi, W., Reddy, M. K., Upadhyaya, K. C., and Sopory,S. K. 2002. A Ca2+/CaM-dependent kinase from pea is stress regulated andin vitro phosphorylates a protein that binds to AtCaM5 promoter. Eur. J.Biochem. 269: 3193–3204.
Pardo, J. M., Reddy, M. P., Yang, S., Maggio, A., Huh, G. H., Matsumoto, T.,Coca, M. A., Paino-D’Urzo, M., Koiwa, H., Yun, D. J., Watad, A. A., Bressan,R. A., and Hasegawa, P. M. 1998. Stress signaling through Ca2+/calmodulin-dependent protein phosphatase calcineurin mediates salt adaptation in plants.Proc. Natl. Acad. Sci. USA 95: 9681–9686.
Park, J. M., Park, C.-J., Lee, S.-B., Ham, B.-K., Shin, R., and Paek, K.-H.2001. Overexpression of the tobacco Tsi1 gene encoding an EREBP/AP2-typetranscription factor enhances resistance against pathogen attack and osmoticstress in tobacco. Plant Cell 13: 1035–1046.
Patharkar, O. R. and Cushman, J. C. 2000. A stress-induced calcium-dependentprotein kinase from Mesembryanthemum crystallium phosphorylates a two-component pseudo-response regulator. Plant J. 24: 679–691.
Pei, Z. M., Ghassemian, Kwak, C. M., McCourt, P., and Schroeder, J. I. 1998.Role of farnesyltransferase in ABA regulation of guard cell anion channelsand plant water loss. Science 282: 287–290.
Pei, Z. M., Murata, Y., Benning, G., Thomine, S., Klusener, B., Allen, G. J.,Grill, E., and Schroeder, J. I. 2000. Calcium channels activated by hydrogenperoxide mediate abscisic acid signaling in guard cells. Nature 406: 731–734.
Peng, Z., Lu, Q., and Verma, D. P. 1996. Reciprocal regulation of �1-pyrroline-5-carboxylate synthetase and proline dehydrogenase genes controls prolinelevels during and after osmotic stress in plants. Mol. Gen. Genet. 253: 334–341.
Perruc, E., Charpenteau, M., Ramirez, B. C., Jauneau, A., Galaug, J.-P., Ranjeva,R., and Ranty, B. 2004. A novel calmodulin-binding protein functions as a neg-ative regulator of osmotic stress tolerance in Arabidopsis thaliana seedlings.Plant J. 38: 410–420.
Phillips, J. R., Oliver, M. J., and Bartels, D. 2002. Molecular genetics of desicca-tion and tolerant systems. In CAB International Desiccation and Survival inPlants: Drying without dying, M. Black and H. Pritchard. (Eds.), pp. 319–341.
Piao, H. L., Lim, J. H., Kim, S. J., Cheong, G. W., and Hwang, I. 2001. Constitu-tive over-expression of AtGSK1 induces NaCl stress responses in the absenceof NaCl stress and results in enhanced NaCl tolerance in Arabidopsis. PlantJ. 27: 305–314.
Pical, C., Westergren, T., Dove, S. K., Larsson, C., and Sommarin,M. 1999. Salinity and hyperosmotic stress induce rapid increases in

DROUGHT AND SALT TOLERANCE IN PLANTS 55
phosphotidylinositol-4,5-bisphosphate, diacylglycerol pyrophosphate, andphosphotidylcholine in Arabidopsis thaliana cells. J. Biol. Chem. 274: 38232–38240.
Pilon-Smits, E. A. H., Ebskamp, M. J. M., Paul, M. J., Jeuken, M. J. W.,Weisbeek, P. J., and Smeekens, S. C. M. 1995. Improved performance oftransgenic fructan-accumulating tobacco under drought stress. Plant Physiol.25: 125–130.
Pilon-Smits, E. A. H., Terry, N., Sears Tobin, K. H., and Van Dun, K. 1999.Enhanced drought resistance in fructan-producing sugar beet. Plant Physiol.Biochem 25: 313–317.
Pilon-Smits, E. A. H., Terry, N., Sears Tobin, K. H. et al. 1998. Trehalose-producing transgenic tobacco plants show improved growth performance un-der drought stress. J Plant Physiol. 25: 525–532.
Posas, F., Wurgler-Murphy, S. M., Maeda, T., Witten, E. A., Thai, T. C., andSaito, H. 1996. Yeast HOG1 MAPkinase cascade is regulated by a multi-step phosphorelay mechanism in the SLN1-YPD1-SSK1 “two-component”osmosensor. Cell 86: 865–875.
Pruvot, G., Cuine, S., Peltier, G., and Rey, P. 1996. Characterization of a noveldrought-induced 34-kDa protein located in the thylakoids of Solanum tubero-sum L. plants. Planta 25: 471–479.
Qin, X. and Zeevaart, J. A. D. 1999. The 9-cis-epoxycarotenoid cleavage reactionis the key regulatory step of abscisic acid biosynthesis in water-stressed bean.Proc. Natl. Acad. Sci. USA 96: 15354–15361.
Qin, X. and Zeevaart, J. A. D. 2002. Overexpression of a 9-cis-epoxycarotenoiddioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid andphaseic acid levels and enhances drought tolerance. Plant Physiol. 128: 544–551.
Qiu, Q. S., Guo, Y., Dietrich, M., Schumaker, K. S., and Zhu, J.-K. 2002. Regula-tion of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thalianaby SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 99: 8436–8441.
Qiu, Q.-S., Guo, Y., Quintero, F. J., Pardo, J. M., Schumaker, K. S., and Zhu,J.-K. 2004. Regulation of vacuolar Na+/H+ exchange in Arabidopsis thalianaby the salt-overly-sensitive (SOS) pathway. J. Biol. Chem. 279: 207–215.
Quintero, F. J., Ohta, M., Shi, H., Zhu, J.-K., and Pardo, J. M. 2002. Reconstitu-tion in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis.Proc. Natl. Acad. Sci. USA 99: 9061–9066.
Ramanjulu, S. and Bartels, D. 2002. Drought- and desiccation-induced modu-lation of gene expression in plants. Plant Cell Environ. 25: 141–151.
Ramanjulu, S. and Sudhakar, C. 1997. Drought tolerance is partly related toamino acid accumulation and ammonia assimilation: A comparative study intwo mulberry genotypes differing in drought sensitivity. J Plant Physiol. 150:345–350.
Ramanjulu, S., Veeranjaneyulu, K., and Sudhakar, C. 1994a. Relative toleranceof certain mulberry (Morus alba L.) varieties to NaCl salinity. Serecologia34: 695–702.
Ramanjulu, S., Veeranjaneyulu, K., and Sudhakar, C. 1994b. Short-term shiftsin nitrogen metabolism in mulberry, Morus alba under salt shock. Phytochem.37: 991–995.
Ratajczak, R., Richter, J., and Luttge, U. 1994. Adaptation of the tonoplast V-type H+-ATPase of Mesembryanthemum crystallinum to salt stress, C3-CAMtransition and plant age. Plant Cell Environ. 17: 1101–1112.
Reiser, V., Raitt, D. C., and Saito, H. 2003. Yeast osmosensor Sln1 and plantcytokinin receptor Cre1 respond to changes in turgor pressure. J. Cell. Biol.161: 1035–1040.
Rentel, M. C., Lecourieux, D., Ouaked, F., Usher, S. L., Petersen, L., Okamoto,H., Knight, H., Peck, S. C., Grierson, C. S., Hirt, H., and Knight, M. R.2004. OXI1 kinase is necessary for oxidative burst-mediated signalling inArabidopsis. Nature 427: 858–861.
Rey, P., Pruvot, G., Becuwe, N., Eymery, F., Rumeau, D., and Peltier, G. A.1998. A novel thioredoxin-like protein located in the chloroplast is inducedby water deficit in Solanum tuberosum L. plants. Plant J. 25: 97–107.
Rhodes, D. and Hanson, A. D. 1993. Quaternary ammonium and tertiary sul-fonium compounds in higher plants. Annu Rev Plant Physiol Plant Mol Biol44: 357–384.
Riechmann, J., Heard, G., Martin, L., Reuber, C.-Z., Jiang, J., Keddie, L., Adam,O., Pineda, O. J., Ratcliffe, R. R., Samaha, R., Creelman, M., Pilgrim, P.,Broun, J. Z., Zhang, D., Ghandehari, B. K., Sherman, G., and Yu L. 2000.Arabidopsis transcription factors: Genome-wide comparative analysis amongeukaryotes. Science 290: 2105–2110.
Robinson, M. J. and Cobb, M. H. 1997. Mitogen-activated protein kinase path-ways. Curr. Opin. Cell Biol. 9: 180–186.
Rock, C. 2000. Pathways to abscisic acid-regulated gene expression. New Phytol.148: 357–396.
Rodrigo, M.-J., Moskovitz, J., Salamini, F., and Bartels, D. 2002. Reverse geneticapproaches in plants and yeast suggest a role for novel, evolutionarily con-served, selenoprotein-related genes in oxidative stress defense. Mol. Genet.Genomics 267: 613–621.
Rontein, D., Basset, G., and Hanson, A. D. 2002. Metabolic engineering ofosmoprotectant accumulation in plants. Metab. Eng. 4: 49–56.
Roosens, N., Hal Bitar, F., Loenders, K., Angenon, G., and Jacobs, M. 2002.Overexpression of ornithine-δ-aminotransferase increases proline biosynthe-sis and confers osmotolerance in transgenic plants. Mol. Breed. 9: 73–80.
Roosens, N. H. J., Thu, T. T., Iskandar, H. M., and Jacobs, M. 1998. Isolation ofornithine-δ-aminotransferase cDNA and effect of salt stress on its expressionin Arabidopsis. Plant Physiol. 117: 263–271.
Roxas, V. P., Smith, Jr, R. K., Allen, E. R., and Allen, R. D. 1997. Overexpres-sion of glutathione-S-transferase/glutathione peroxidase enhances the growthof transgenic tobacco seedlings during stress. Nat. Biotechnol. 15: 988–991.
Roy, M. and Wu, R. 2001. Arginine decarboxylase transgene expression andanalysis of environmental stress tolerance in transgenic rice. Plant Sci. 160:869–875.
Rus, A., Yokoi, S., Sharkhuu, A., Reddy, M., Lee, B. H., Matsumoto, T. K.,Koiwa, H., Zhu, J. K., Bressan, R. A., and Hasegawa, P. M. 2001. AtHKT1 isa salt tolerance determinant that controls Na(+) entry into plant roots. ProcNatl Acad Sci USA. 98: 14150–1455.
Ryu, S. B. and Wang, X. 1995. Expression of phospholipase D during castorbean senescence. Plant Physiol. 108: 713–719.
Saijo, Y., Hata, S., Kyozuka, J., Shimamoto, K., and Izui, K. 2000. Over-expression of a single Ca2+-dependent protein kinase confers both cold andsalt/drought tolerance on rice plants. Plant J. 23: 319–327.
Saito, S., Hirai, N., Matsumoto, C., Ohigashi, H., Ohta, D., Sakata, K., andMizutani, M. 2004. Arabidopsis CYP707As encode (+)-abscisic acid 8′-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid. PlantPhysiol. 134: 1439–1449.
Sakuma, Y., Liu, Q., Dubouzet, J. G., Abe, H., Shinozaki, K., and Yamaguchi-Shinozaki, K. 2002. DNA-binding specificity of the ERF/AP2 domain ofArabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 290: 998–1009.
Sanders, D., Brownlee, C., and Harper, J. F. 1999. Communicating with calcium.Plant Cell 11: 691–706.
Sang, Y., Zhang, S., Li, W., Huang, and Wang, X. 2001. Regulation of plantwater loss by manipulating the expression of phospholipase Dα. Plant J. 28:135–144.
Sarda, X., Tousch, D., Ferrare, K., Cellier, F., Alcon, C., Dupuis, J. M., Casse, F.,and Lamaze, T. 1999. Characterization of closely related delta-TIP genes en-coding aquaporins which are differentially expressed in sunflower roots uponwater deprivation through exposure to air. Plant Mol. Biol. 25: 179–191.
Schaffner, A. R. 1998. Aquaporin function, structure, and expression: Are theremore surprises to surface in water relations? Planta 25: 131–139.
Schroeder, J. I., Kwak, J. M., and Allen, G. J. 2001. Guard cell abscisic acidsignalling and engineering drought hardiness in plants. Nature 410: 327–330.
Schuppler, U., He, P.-H., John, P. C. L., and Munns, R. 1998. Effect of waterstress on cell division and cell-division-cycle 2-like cell-cycle kinase activityin wheat leaves. Plant Physiol. 117: 667–678.
Schweighofer, A., Hirt, H., and Meskiene, I. 2004. Plant PP2C phosphatases:Emerging functions in stress signalling. Trends Plant Sci. 9: 236–243.

56 D. BARTELS AND R. SUNKAR
Seki, M., Narusaka, M., Abe, H., Kasuga, M., Yamaguchi-Shinozaki, K.,Carninic, P., Hayashizaki, Y., and Shinozaki, K. 2001. Monitoring theexpression pattern of 1300 Arabidopsis genes under drought and cold stressesby using a full-length cDNA microarray. Plant Cell 13: 61–72.
Seki, M., Narusaka, M., Ishida, J., Nanjo, T., Fujita, M., Oono, Y., Kamiya,A., Nakajima, M., Enju, A., Sakurai, T., Satou, M., Akiyama, K., Taji, T.,Yamaguchi-Shinozaki, K., Carninci, P., Kawai, J., Hayashizaki, Y., and Shi-nozaki, K. 2002. Monitoring the expression profiles of ca. 7000 Arabidopsisgenes under drought, cold, and high-salinity stresses using a full-length cDNAmicroarray. Plant J. 31: 279–292.
Serraj, R. and Sinclair, T. R. 2002. Osmolyte accumulation: Can it really helpincrease in crop yield under drought conditions? Plant Cell Environ. 25: 333–341.
Serrano, R. 1996. Salt tolerance in plants and microorganisms: Toxicity targetsand defense responses. Int. Rev. Cytol. 165: 1–52.
Serrano, R., Mulet, J. M., Rios. G., Marquez. J. A., de Larrinoa, I., Leube, M.P., Mendizabal, I., Pascual-Ahuir, A., Proft, M., Ros, R., and Montesinos, C.1999. A glimpse of the mechanisms of ion homeostasis during salt stress. J.Exp. Bot. 50: 1023–1036.
Setter, T. L. and Flannigan, B. A. 2001. Water deficit inhibits cell division andexpression of transcripts involved in cell proliferation and endoreduplicationin maize endosperm. J. Exp. Bot. 52: 1401–1408.
Shalata, A., Mittova, V., Volokita, M., Guy, M., and Tal, M. 2001. Response ofthe cultivated tomato and its wild salt-tolerant relative Lycopersicon pennelliito salt-dependent oxidative stress: The root antioxidative system. Physiol.Plant 112: 487–494.
Sharp, R. E., Hsiao, T. C., and Silk, W. K. 1988. Growth of the maize primaryroot at low water potentials. I. Spatial distribution of expansive growth. PlantPhysiol. 87: 50–57.
Sharp, R. E., Hsiao, T. C., and Silk, W. K. 1990. Growth of the maize primaryroot at low water potentials. II. Role of growth and deposition of hexose andpotassium in osmotic adjustment. Plant Physiol. 93: 1337–1346.
Sheen, J. 1996. Ca2+-dependent protein kinases and stress signal transductionin plants. Science 274: 1900–1902.
Sheen, J. 1998. Mutational analysis of protein phosphatase 2C involved in ab-scisic acid signal transduction in higher plants. Proc. Natl. Acad. Sci. USA95: 975–80.
Shen, B., Jensen, R. G., and Bohnert, H. J. 1997a. Mannitol protects againstoxidation by hyderoxyl radicals. Plant Physiol. 115: 527–532.
Shen, B., Jensen, R. G., and Bohnert, H. J. 1997b. Increased resistance to oxida-tive stress in transgenic plants by targeting mannitol biosynthesis to chloro-plasts. Plant Physiol. 113: 1177–1183.
Shen, Q., Zhang, P., and Ho, T. -H. D. 1996. Modular nature of abscisic acid(ABA) response complexes: Composite promoter units that are necessaryand sufficient for ABA induction of gene expression in barley. Plant Cell 8:1107–1119.
Sheveleva, E., Chmara, W., Bohnert, H. J., and Jensen, R. G. 1997. Increasedsalt and drought tolerance by D-ononitol production in transgenic Nicotianatabacum L. Plant Physiol. 115: 1211–1219.
Shi, H., Ishitani, M., Kim, C., and Zhu, J.-K. 2000. The Arabidopsis thalianasalt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl.Acad. Sci. USA 97: 6896–6901.
Shi, H., Lee, B.-H., Wu, S.-J., and Zhu, J.-K. 2003. Overexpression of a plasmamembrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsisthaliana. Nature Biotechnol. 21: 81–85.
Shi, H. and Zhu, J.-K. 2002. Regulation of expression of the vacuolar Na+/H+antiporter gene AtNHX1 by salt stress and ABA. Plant Mol. Biol. 50: 543–550.
Shinozaki, K. and Russel, P. 1995. Cell-cycle control linked to extracellularenvironment by MAP kinase pathway in fission yeast. Nature 378: 739–743.
Shinozaki, K. and Yamaguchi-Shinozaki, K. 1997. Gene expression and signaltransduction in water-stress response. Plant Physiol. 25: 327–334.
Shinozaki, K. and Yamaguchi-Shinozaki, K. 2000. Molecular response to dehy-dration and low temperature: Differences and cross-talk between two stresssignaling pathways. Curr. Opin. Plant Biol. 3: 217–223.
Shinozaki, K., Yamaguchi-Shinozaki, K., and Seki, M. 2003. Regulatory net-work of gene expression in the drought and cold stress responses. Curr. Opin.Plant Biol. 6: 410–417.
Shinwari, Z. K., Nakashima, K., Miura, S., Kasuga, M., Seki, M., Yamaguchi-Shinozaki K., and Shinozaki, K. 1998. An Arabidopsis gene family encodingDRE/CRT binding proteins involved in low-temperature-responsive gene ex-pression. Biochem. Biophys. Res. Commun. 250: 161–170.
Shirasu, K., Nakajima, H., Rajasekhar, K., Dixon, R. A., and Lamb, C. 1997.Salicylic acid potentiates an agonist-dependent gain control that amplifiespathogen signal in the activation of defense mechanisms. Plant Cell 9: 261–270.
Siefritz, F. Tyree, M. T., Lovisolo, C., Schubert, A., and Kaldenhoff, R. 2002.PIP1 plasma membrane aquaporins in tobacco: From cellular effects to func-tion in plants. Plant Cell 14: 869–876.
Singh, K. B., Foley, R. C., and Onate-Sanchez, L. 2002. Transcription fac-tors in plant defense and stress response. Curr. Opin. Plant Biol. 5: 430–436.
Singla-Pareek, S. L., Reddy, M. K., and Sopory, S. K. 2003. Genetic engineeringof the glyoxalase pathway in tobacco leads to enhanced salinity tolerance.Proc. Natl. Acad. Sci. USA 100: 14672–14677.
Smart, L. B., Moskal, W. A., Cameron, K. D., and Bennett, A. 2001. MIPgenes are down-regulated under drought stress in Nicotiana glauca. PlantCell Physiol. 42: 686–693.
Smirnoff, N. 1998. Plant resistance to environmental stress. Curr. Opin. PlantBiol. 9: 214–219.
Smirnoff, N., and Cumbes, Q. J. 1989. Hydroxyl radical scavenging activity ofcompatible solutes. Phytochem. 28: 1057–1060.
Smith-Espinoza, C. J., Richter, A., Salamini, F., and Bartels, D. 2003. Dis-secting the response to dehydration and salt (NaCl) in the resurrection plantCraterostigma Plantagenium. Plant Cell Environ. 26: 1307–1315.
Snedden, W. A. and Fromm, H. 1998. Calmodulin, calmodulin-related proteinsand plant responses to the environment. Trends Plant Sci. 3: 299–304.
Snedden, W. A. and Fromm, H. 2001. Calmodulin as a versatile calcium signaltransducer in plants. New Phytol. 151: 35–66.
Soderman, E., Hjellstrom, M., Fahleson, J., and Engstrom, P. 1999. The HD-ZIP gene ATHB6 in Arabidopsis is expressed in developing leaves, rootsand carpels and up-regulated by water-deficit conditions. Plant Mol. Biol. 40:1073–1083.
Soderman, E., Mattsson, J., and Engstrom, P. 1996. The Arabidopsis homeoboxgene ATHB-7 is induced by water deficit and by abscisic acid. Plant J. 10:375–381.
Spollen, W. G., Sharp, R. E., Saab, I. N., and Wu, Y. 1993. Regulation of cellexpansion in roots and shoots at low water potentials. In: Water Deficits: PlantResponses from Cell to Community. Smith J. A. C., Griffiths H., Eds. BIOSScientific Publishers, Oxford, pp. 37–52.
Staxen, I., Pical, C., Montgomery, L. T., Gray, J. E., Hetherington, A. M., andMcAinsh, M. R. 1999. Abscisic acid induces oscillations in guard-cell cy-tosolic free calcium that involve phosphoinositide-specific phospholipase C.Proc. Natl. Acad. Sci. USA 96: 1779–1784.
Stewart, C. R., Voetberg, G., and Rayapati, P. J. 1986. The effects of benzylade-nine, cycloheximide and cordycepin on wilting-induced abscisic acid andpraline accumulations and abscisic acid- and salt-induced praline accumula-tion in barley leaves. Plant Physiol. 82: 703–707.
Stockinger, E. J., Gilmour, S. J., and Thomashow, M. F. 1997. Arabidopsisthaliana CBF1 encodes an AP2 domain-containing transcriptional activatorthat binds to the C-repeat/DRE, a cis-acting DNA regulatory element thatstimulates transcription in response to low temperature and water deficit.Proc. Natl. Acad. Sci. USA 94: 1035–1040.
Streeter, J. G., Lohnes, D. G., and Fioritto, R. J. 2001. Pattern of pinitol accu-mulation in soybean plants and relationships to drought tolerance. Plant CellEnviron. 24: 429–438.
Sugano, S. Kaminaka, H. Rybka, Z. Catala, R. Salinas, J., Matsui, K. Ohme-Tkagi, M., and Takatsuji, H. 2003. Stress-responsive zinc finger geneZPT2-3 plays a role in drought tolerance in petunia. Plant J. 36: 830–841.

DROUGHT AND SALT TOLERANCE IN PLANTS 57
Sun, W., Bernard, C., van de Cotte, Van Montagu, M., and Verbruggen, N.2001. At-HSP17. 6A, encoding a small heat-shock protein in Arabidopsis,can enhance osmotolerance upon overexpression. Plant J. 27: 407–415.
Sunkar, R., Bartels, D., and Kirch, H.-H. 2003. Overexpression of a stress-inducible aldehyde dehydrogenase gene from Arabidopsis thaliana in trans-genic plants improves stress tolerance. Plant J. 35: 452–464.
Sze, H., Liang, F., Hwang, I., Curran, A. C., and Harper, J. F. 2000. Diversityand regulation of plant Ca2+ pumps: Insights from expression in yeast. Annu.Rev. Plant Physiol. Plant Mol. Biol. 51: 433–462.
Taji, T., Ohsumi, C., Iuchi, S., Seki, M., Kasuga, M., Kobayashi, M., Yamaguchi-Shinozaki, K., and Shinozaki, K. 2002. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsisthaliana. Plant J. 29: 417–426.
Takahashi, S., Katagiri, T., Hirayama, T., Yamaguchi-Shinozaki, K., andShinozaki, K. 2001. Hyperosmotic stress induces a rapid and transient increasein Inositol1,4,5-triphosphate independent of abscisic acid in Arabidopsis cellcultures. Plant Cell Physiol. 42: 214–222.
Tamura, T., Hra, K., Yamaguchi, Y., Koizumi, N., and Sano, H. 2003. os-motic stress tolerance of transgenic tobacco expressing a gene encoding amembrane-located receptor-like protein from tobacco plants. Plant Physiol.131: 454–462.
Tan, B. C., Schwartz, S. H., Zeevart, J. A., and McCarty, D. R. 1997. Geneticcontrol of abscisic acid biosynthesis in maize. Proc. Natl. Acad. Sci. USA 94:12235–12240.
Tena, G., Asai, T., Chiu, W.-L., and Sheen, J. 2001. Plant mitogen-activatedprotein kinase signaling cascades. Curr. Opin. Plant Biol. 4: 392–400.
Thiyagarajah, M., Fry, S., and Yeo, A. 1996. In vitro salt tolerance of cell wallenzymes from halophytes and glycophytes. J. Exp. Bot. 47: 1717–1724.
Thompson, A. J., Jackson, A. C., Parker, R. A., Morpeth, D. R., Burbidge, A.,and Taylor, I. B. 2000a. Abscisic acid biosynthesis in tomato: Regulationof zeaxanthin epoxidase and 9-cis-epoxycarotenoid dioxygenase mRNAs bylight/dark cycles, water stress and abscisic acid. Plant Mol. Biol. 42: 833–845.
Thompson, A. J., Jackson, A. C., Symonds, R. C., Mulholland, B. J., Dadswell,A. R., Blake, P. S., Burbidge, A., and Taylor, I. B. 2000b. Ectopic expressionof a tomato 9-cis-epoxycarotenoid dioxygenase gene causes over-productionof abscisic acid. Plant J. 23: 363–374.
Tiburcio, A. F., Besford, R. T., Capell, T., Borrell, A., Testillano, P. S., andRisueno, M. C. 1994. Mechanisms of polyamine action during senescenceresponses induced by osmotic stress. J. Exp. Bot. 45: 1789–1800.
Triesmann, R. 1996. Regulation of transcription by MAP kinase cascades. Curr.Opin. Cell Biol. 8: 205–215.
Tsugane, K., Kobayashi, K., Niwa, Y., Ohba, Y., Wada, K., and Kobayashi, H.1999. A recessive Arabidopsis mutant that grows photoautotrophically undersalt stress shows enhanced active oxygen detoxification. Plant Cell 11: 1195–1206.
Tyerman, S. D., Niemietz, C. M., and Bramley, H. 2002. Plant aquaporins:Multifunctional water and solute channels with expanding roles. Plant CellEnviron. 25: 173–194.
Ulm, R., Rekenkova, E., Sansebastiano, G. P., Bechtold, N., and Paszkowski, J.2001. Mitogen-activated protein kinase phosphatse is required for genotoxicstress releif in Arabidopsis. Genes Dev. 15: 699–709.
Uno, Y., Furihata, T., Abe, H., Yoshida, R., Shinozaki, K., and Yamaguchi-Shinozaki, K. 2000. Arabidopsis basic leucine zipper transcription factorsinvolved in an abscisic acid-dependent signal transduction pathway underdrought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 97: 11632–11637.
Urano, K., Yoshiba, Y., Nanjo, T., Igarashi, Y., Seki, M., Sekiguchi, K.,Yamaguchi-Shinozaki, K., and Shinozaki, K. 2003. Characterization of Ara-bidopsis genes involved in biosynthesis of polyamines in abiotic stress re-sponses and developmental stages. Plant Cell Environ. 26: 1917–1926.
Urano, K., Yoshiba, Y., Nanjo, T., Ito, T., Yamaguchi-Shinozaki, K., andShinozaki, K. 2004. Arabidopsis stress-inducible gene for arginine decar-boxylase AtADC2 is required for accumulation of putrescine in salt tolerance.Biochem. Biophys. Res. Commun. 313: 369–375.
Urao, T., Yakubov, B., Satoh, R., Yamaguchi-Shinozaki, K., Seki, B., Hirayama,T., and Shinozaki, K. 1999. A transmembrane hybrid-type histidine kinase inArabidopsis functions as an osmosensor. Plant Cell 11: 1743–1754.
Urao, T., Yamaguchi-Shinozaki, K., Urao, S., and Shinozaki, K. 1993. An Ara-bidopsis myb homolog is induced by dehydration stress and its gene prod-uct binds to the conserved MYB recognition sequence. Plant Cell 5: 1529–1539.
Van Camp, W., Capiau, K., Van Montagu, M., Inze, D., and Slooten, L. 1996.Enhancement of oxidative stress tolerance in transgenic tobacco plants over-producing Fe-superoxide dismutase in chloroplasts. Plant Physiol. 112: 1703–1714.
Veena, Reddy, V. S., and Sopory, S. K. 1999. Glyoxylase I from Brassica juncea:Molecular cloning, regulation and its overexpression confer tolerance in trans-genic tobacco under stress. Plant J. 17: 385–395.
Venema, K., Quintero, F. J., Pardo, J. M., and Donaire, J. P. 2002. The Arabidop-sis Na+/H+ exchanger AtNHX1 catalyses low affinity Na+ and K+ transportin reconstituted liposomes. J. Biol. Chem. 277: 2413–2418.
Verbruggen, N., Hua, X. J., May, M., and VanMontagu, M. 1996. Environmen-tal and developmental signals modulate proline homeostasis: evidence fora negative transcriptional regulator. Proc. Natl. Acad. Sci. USA, 93: 8787–8791.
Vernon, D. M., and Bohnert, H. J. 1992. A novel methyl transferase induced byosmotic stress in the facultative halophyte, Mesembryanthemum crystallinum.EMBO J. 11: 2077–2085.
Vierstra, R. D. 1996. Proteolysis in plant: mechanisms and functions. Plant Mol.Biol. 32: 275–302.
Villalobos, M. A., Bartels, D., and Iturriga, G. 2004. Stress tolerance and glucoseinsensitive phenotypes in Arabidopsis overexpressing the CpMYB10 tran-scription factor gene. Plant Physiol. 135: 309–324.
Vitart, V., Baxter, I., Doerner, P., and Harper, J. F. 2001. Evidence for a rolein growth and salt resistance of a plasma membrane H+-ATPase in the rootendodermis. Plant J. 27: 191–201.
Vogel, G., Aeschbacher, R. A., Muller, J., Boller, T., and Wiemken, A. 1998.Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: identifica-tion by functional complementation of the yeast tps2 mutant. Plant J. 13:673–683.
Volkov, V., Wang, B., Dominy, P. J., Fricke, W., and Amtmann, A. 2003. Thel-lungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesseseffective mechanisms to discriminate between potassium and sodium. Plant,Cell Environ. 27: 1–14.
Wang, H., Datla, R., Georges, F., Loewen, M., and Cuter, A. J. 1995. Promot-ers from kin1 and cor6. 6, two homologues of Arabidopsis thaliana genes:Transcriptional regulation and gene expression induced by low temperature,ABA, osmoticum and dehydration. Plant Mol. Biol. 28: 605–617.
Wang, H., Qi, Q., Schorr, P., Cutler, A. J., Crosby, W. L., and Fowke, L. C.1998. ICK1, a cyclin-dependent protein kinase inhibitor from Arabidopsisthaliana interacts with both Cdc2a and CycD3, and its expression is inducedby abscisic acid. Plant J. 15: 501–510.
Wang, W., Vinocur, B., Shoseyov, O., and Altman, A. 2004. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. TrendsPlant Sci. 9: 244–252.
Wang, X. 2001. Plant phospholipases. Annu. Rev. Plant Physiol. Plant Mol. Biol.52: 211–231.
Wang, X. 2002. Phospholipase D in hormonal and stress signalling. Curr. Opin.Plant Biol. 5: 408–414.
Wehmeyer, N. and Vierling, E. 2000. The expression of small heat shock proteinsin seeds responds to discrete developmental signals and suggests a generalprotective role in desiccation tolerance. Plant Physiol. 122: 1099–1108.
Weig, A., Deswarte, C., and Chrispeels, M. J. 1997. The major intrinsic proteinfamily of Arabidopsis has 23 members that form three distinct groups withfunctional aquaporins in each group. Plant Physiol. 25: 1347–1357.
Wendehenne, D., Pugin, A., Klessig, D. F., and Durner, J. 2001. Nitric oxide:Comparative synthesis and signalling in animal and plant cells. Trends PlantSci. 6: 177–183.

58 D. BARTELS AND R. SUNKAR
West, G., Inze, D., and Beemster, G. T. S. 2004. Cell cycle modulation in theresponse of the primary root of Arabidopsis to salt stress. Plant Physiol. 135:1050–1058.
Westgate, M. E. and Boyer, J. S. 1985. Osmotic adjustment and the inhibitionof leaf, root, stem and silk growth at low water potentials in maize. Planta164: 540–549.
Williams, M. E., Foster, R., and Chua, N.-H. 1992. Sequences flanking thehexameric G-box core CACGTG affect the specificity of protein binding.Plant Cell 4: 485–496.
Wimmers, L. E., Ewing, N. N., and Bennett, A. B. 1992. Higher plant Ca2+-ATPase: Primary structure and regulation of mRNA abundance by salt. Proc.Natl. Acad. Sci. USA 89: 9205–9209
Winicov, I. 1993. cDNA encoding zinc finger motifs from salt-tolerant alfalfa(Medicago sativa L.) cells. Plant Physiol. 102: 681–682.
Winicov, I. 2000. Alfin1 transcription factor overexpression enhances plant rootgrowth under normal and saline conditions and improves salt tolerance inalfalfa. Planta 210: 416–422.
Winicov, I. and Bastola, D. R. 1999. Transgenic overexpression of the tran-scription factor Alfin1 enhances expression of the endogenous MSPRP2 genein alfalfa and improves salinity tolerance of the plants. Plant Physiol. 120:473–480.
Wu, Y., Thorne, E. T., Sharp, R. E., and Cosgrove, D. J. 2001. Modification ofexpansin transcript levels in the maize primary root at low water potentials.Plant Physiol. 126: 1471–1479.
Wurgler-Murphy, S. M., Maeda, T., Witten, E. A., and Saito, H. 1997. Regulationof the Saccharomyces cerevisiae HOG1 mitogen-activated protein kinase bythe PTP2 and PTP3 protein tyrosine phosphatases. Mol. Cell Biol. 17: 1289–1297.
Wurgler-Murphy, S. M., and Saito, S. 1997. Two-component signal transducersand MAPK cascades. Trends Biochem. Sci. 25: 172–176.
Xiong, L., Gong, Z., Rock, C. D., Subramanian, S., Guo, Y., Xu, W., Galbraith,D., and Zhu, J. K. 2001. Modulation of abscisic acid signal transduction andbiosynthesis by an Sm-like protein in Arabidopsis. Dev. Cell 1: 771–778.
Xiong, L., Ishitani, M., Lee, H., and Zhu, J.-K. 2001. The ArabidopsisLOS5/ABA3 locus encodes a molybdenum cofactor sulfurylase and modu-lates cold stress and osmotic stress-responsive gene expression. Plant Cell13: 2063–2083.
Xiong, L., Lee, B. H., Ishitani, M., Lee, H., Zhang, C., and Zhu, J. K. 2001c.FIERY1 encoding an inositol polyphosphate 1-phosphatase is a negative reg-ulator of abscisic acid and stress signalling in Arabidopsis. Genes Dev. 15:1971–1984.
Xiong, L. and Zhu, J. K. 2003. Regulation of abscisic acid biosynthesis. PlantPhysiol. 133: 29–36.
Xu, Q., Fu, H.-H., Gupta, R., and Luan, S. 1998. Molecular characterization ofa tyrosine-specific protein phosphatase encoded by a stress-responsive genein Arabidopsis. Plant Cell 10: 849–857.
Yamada, S., Komori, T., Myers, P. N., Kuwata, S., Kubo, T., and Imaseki, H.1997. Expression of plama membrane water channel genes under water stressin Nicotiana excelsior. Plant Cell Physiol. 25: 1226–1231.
Yamaguchi-Shinozaki, K., Koizumi, M., Urao, S., and Shinozaki, K. 1992.Molecular cloning and characterization of 9 cDNAs for genes that are re-sponsive to desiccation in Arabidopsis thaliana: Sequence analysis of onecDNA that encodes a putative transmembrane channel protein. Plant CellPhysiol. 25: 217–224.
Yamaguchi-Shinozaki, K. and Shinozaki, K. 1993. The plant hormone abscisicacid mediates the drought-induced expression but not the seed-specific expres-
sion of rd22, a gene responsive to dehydration stress in Arabidopsis thaliana.Mol. Gen. Genet. 238: 17–25.
Yamaguchi-Shinozaki, K. and Shinozaki, K. 1994. A novel cis-acting ele-ment in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 6: 251–264.
Yancey, P. H., Clark, M. E., Hand, S. C., Bowlus, R. D., and Somero, G. N.1982. Living with water stress: Evolution of osmolyte systems. Science 217:1214–1222.
Yang, S. X., Zhao, Y. X., Zhang, Q., He, Y. K., Zhang, H., and Luo. 2001. HAL1mediate salt adaptation in Arabidopsis thaliana. Cell Res. 11: 142–148.
Yang, T. and Poovaiah, B. W. 2003. Calcium/Calmodulin-mediated signal net-work in plants. Trends Plant Sci. 8: 505–512.
Ye, B., Muller, H. H., Zhang, J., and Gressel, J. 1998. Constitutively elevatedlevels of putrescine and putrescine generating enzymes correlated with oxi-dant stress resistance in Conyza bonariensis and wheat. Plant Physiol. 115:1443–1451.
Yokoi, S., Quintero, F. J., Cubero, B., Ruiz, M. T., Bressan, R. A., Hasegawa, P.M., and Pardo, J. M. 2002. Differential expression and function of Arabidopsisthaliana NHX Na+/H+ antiporters in the salt stress response. Plant J. 30:529–539.
Yoon, H. W., Kim, M. C., Shin, P. G., Kim, J-S., Lee, S. Y., Hwang, I., Bahk, J.D., Hong, J. C., Han, C., and Cho, M. J. 1997. Differential expression of twofunctional serine/threonine protein kinases from soybean that have an unusualacidic domains at the carboxy terminus. Mol. Gen. Genet. 255: 359–371.
Yoshiba, Y., Kiyosue, T., Nakashima, K., Yamaguchi-Shinozaki, K., andShinozaki, K. 1997. Regulation of levels of proline as an osmolyte in plantsunder water stress. Plant Cell Physiol. 38: 1095–1102.
Yuasa, T., Ichimura, K., Mizoguchi, T., and Shinozaki, K. 2001. Oxidative stressactivates ATMPK6, an Arabidopsis homologue of MAP kinase. Plant CellPhysiol. 42: 1012–1016.
Yuasa, T. and Muto, S. 1996. Activation of 40-kDa protein kinases in responseto hypo- and hyperosmotic shocks in a halotolerant green alga Dunaliellatertiolecta. Plant Cell Physiol. 37: 35–42.
Zeevaart, J. A. D. and Creelman, R. A. 1988. Metabolism and physiology ofabscisic acid. Annu. Rev. Plant Physiol. Plant Mol. Biol. 39: 439–473.
Zhang, H.-X. and Blumwald, E. 2001. Transgenic salt-tolerant tomato plantsaccumulate salt in foliage but not in fruit. Nature Biotechnol. 19: 765–768.
Zhang, H.-X., Hodson, J. N., Williams, J. P., and Blumwald, E. 2001. Engi-neering salt-tolerant Brassica plants: Characterization of yield and seed oilquality in transgenic plants with increased vacuolar sodium accumulation.Proc. Natl. Acad. Sci. USA 98: 12832–12836
Zhu, B. C., Su, J., Chan, M. C., Verma, D. P. S., Fan, Y. L., and Wu, R. 1998.Over-expression of a �-pyrroline-5-carboxylate synthetase gene and analysisof tolerance to water-stress and salt-stress in transgenic rice. Plant Sci. 139:41–48.
Zhu, J., Gong, Z., Zhang, C., Song, C.-P., Damsz, B., Inan, G., Koiwa, H., Zhu,J.-K., Hasegawa, P. M., and Bressan, R. A. 2002. OSM1/SYP61: A syntaxinprotein in Arabidopsis controls abscisic acid-mediated and non-abscisic acid-mediated responses to abiotic stress. Plant Cell 14: 3009–3028.
Zhu, J.-K. 2000. Genetic analysis of plant salt tolerance using Arabidopsis. PlantPhysiol. 124: 941–948.
Zhu, J.-K. 2001. Plant salt tolerance. Trends Plant Sci. 6: 66–71.Zhu, J.-K. 2002. Salt and drought stress signal transduction in plants. Annu. Rev.
Plant Biol. 53: 247–273.Zhu, J. K. 2003. Regulation of ion homeostasis under salt stress. Curr. Opin.
Plant Biol. 6: 441–445.
Related Documents











