Original Contribution Doxorubicin as an antioxidant: Maintenance of myocardial levels of lycopene under doxorubicin treatment Ana Lucia Anjos Ferreira a, ⁎ , Kyung-Jin Yeum b , Luiz Shiguero Matsubara a , Beatriz Bojikian Matsubara a , Camila R. Correa a , Elenize Jamas Pereira a , Robert Mitchell Russell b , Norman I. Krinsky b,c , Guangwen Tang b a Department of Internal Medicine, Botucatu Faculty of Medicine, UNESP, São Paulo State University, CEP: 18618-970, Botucatu, SP, Brazil b Jean Mayer United States Department of Agriculture, Human Nutrition Research Centeron Aging at Tufts University, Boston, MA 02111, USA c Department of Biochemistry, School of Medicine, Tufts University, Boston, MA, USA Received 21 December 2006; revised 25 April 2007; accepted 2 May 2007 Available online 10 May 2007 Abstract The mechanism of doxorubicin-induced cardiotoxicity remains controversial. Wistar rats (n = 96) were randomly assigned to a control (C), lycopene (L), doxorubicin (D), or doxorubicin + lycopene (DL) group. The L and DL groups received lycopene (5 mg/kg body wt/day by gavage) for 7 weeks. The D and DL groups received doxorubicin (4 mg/kg body wt intraperitoneally) at 3, 4, 5, and 6 weeks and were killed at 7 weeks for analyses. Myocardial tissue lycopene levels and total antioxidant performance (TAP) were analyzed by HPLC and fluorometry, respectively. Lycopene metabolism was determined by incubating 2 H 10 -lycopene with intestinal mucosa postmitochondrial fraction and lipoxygenase and analyzed with HPLC and APCI mass spectroscopy. Myocardial tissue lycopene levels in DL and L were similar. TAP adjusted for tissue protein were higher in myocardium of D than those of C (P = 0.002). Lycopene metabolism study identified a lower oxidative cleavage of lycopene in D as compared to those of C. Our results showed that lycopene was not depleted in myocardium of lycopene-supplemented rats treated with doxorubicin and that higher antioxidant capacity in myocardium and less oxidative cleavage of lycopene in intestinal mucosa of doxorubicin- treated rats suggest an antioxidant role of doxorubicin rather than acting as a prooxidant. © 2007 Elsevier Inc. All rights reserved. Keywords: Lycopene; Doxorubicin; Heart; Rat; Enzymatic cleavage; Oxidative cleavage; Antioxidant capacity Introduction The anthracycline, doxorubicin (DOX), is an excellent drug for the treatment of a wide variety of human tumors. However, the development of irreversible cardiotoxicity has limited its use [1]. Due to the successful action of DOX as a chemotherapic agent, several strategies have been tried to prevent/attenuate the side effects of DOX. Although DOX-induced injury appears to be multifactorial, a common denominator to most of the proposed mechanisms is cellular damage mediated by reactive oxygen species (ROS) [2,3]. However, it remains controversial as to whether antioxidants can prevent such side effects [4–8]. Among these molecules, metal ion chelators, vitamin A [9], β- carotene [6,7], and others have been widely investigated in relation to DOX toxicity. Free Radical Biology & Medicine 43 (2007) 740 – 751 www.elsevier.com/locate/freeradbiomed Abbreviations: AAPH, 2,2′-azobis-(2-amidinopropane) dihydrochloride; AMVN, 2,2′-azobis 2,4-dimethylvaleronitrile; BODIPY 581/591, 4,4-di- fluoro-5-(4-phenyl-1,3-butadienyl)-4-bora-3a,4a-diaza-s-indacene-3-undecanoic acid; DCFH, 2′,7′-dichlorodihydrofluorescein; DCFH-DA, DCFH diacetate; DOX, doxorubicin; DTT, dithiothreitol; EDTA, ethylenediamine-tetraacetic acid; Hepes, N-(2-hydroxyethyl)piperazine-N′–(2-ethanesulfonic acid); HPLC, high- performance liquid chromatography; KCl, potassium chloride; LC/APCI-MS, liquid chromatography/atmospheric pressure chemical ionization-mass spectro- metry; LOX, lipoxygenase (soybean); MeO-AMVN, 2,2′-azobis(4-methoxy-2,4- dimethylvaleronitrile); NAD + , nicotinamide adenine dinucleotide; ROS, reactive oxygen species; RT, retention time; TAP, total antioxidative performance; THF, tetrahydrofuran. ⁎ Corresponding author. Departamento de Clinica Médica, Faculdade de Medicina de Botucatu, UNESP, CEP 18618–970, Botucatu, SP, Brazil. Fax: +55 14 3882 2969/3882 2238. E-mail address: [email protected] (A.L.A. Ferreira). 0891-5849/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.freeradbiomed.2007.05.002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Free Radical Biology & Medicine 43 (2007) 740–751www.elsevier.com/locate/freeradbiomed
Original Contribution
Doxorubicin as an antioxidant: Maintenance of myocardial levels oflycopene under doxorubicin treatment
Ana Lucia Anjos Ferreira a,⁎, Kyung-Jin Yeum b, Luiz Shiguero Matsubara a,Beatriz Bojikian Matsubara a, Camila R. Correa a, Elenize Jamas Pereira a,
Robert Mitchell Russell b, Norman I. Krinsky b,c, Guangwen Tang b
a Department of Internal Medicine, Botucatu Faculty of Medicine, UNESP, São Paulo State University, CEP: 18618-970, Botucatu, SP, Brazilb Jean Mayer United States Department of Agriculture, Human Nutrition Research Center on Aging at Tufts University, Boston, MA 02111, USA
c Department of Biochemistry, School of Medicine, Tufts University, Boston, MA, USA
Received 21 December 2006; revised 25 April 2007; accepted 2 May 2007Available online 10 May 2007
Abstract
The mechanism of doxorubicin-induced cardiotoxicity remains controversial. Wistar rats (n=96) were randomly assigned to a control (C),lycopene (L), doxorubicin (D), or doxorubicin+ lycopene (DL) group. The L and DL groups received lycopene (5 mg/kg body wt/day by gavage)for 7 weeks. The D and DL groups received doxorubicin (4 mg/kg body wt intraperitoneally) at 3, 4, 5, and 6 weeks and were killed at 7 weeks foranalyses. Myocardial tissue lycopene levels and total antioxidant performance (TAP) were analyzed by HPLC and fluorometry, respectively.Lycopene metabolism was determined by incubating 2H10-lycopene with intestinal mucosa postmitochondrial fraction and lipoxygenase andanalyzed with HPLC and APCI mass spectroscopy. Myocardial tissue lycopene levels in DL and L were similar. TAP adjusted for tissue proteinwere higher in myocardium of D than those of C (P=0.002). Lycopene metabolism study identified a lower oxidative cleavage of lycopene in D ascompared to those of C. Our results showed that lycopene was not depleted in myocardium of lycopene-supplemented rats treated withdoxorubicin and that higher antioxidant capacity in myocardium and less oxidative cleavage of lycopene in intestinal mucosa of doxorubicin-treated rats suggest an antioxidant role of doxorubicin rather than acting as a prooxidant.© 2007 Elsevier Inc. All rights reserved.
Keywords: Lycopene; Doxorubicin; Heart; Rat; Enzymatic cleavage; Oxidative cleavage; Antioxidant capacity
Abbreviations: AAPH, 2,2′-azobis-(2-amidinopropane) dihydrochloride;AMVN, 2,2′-azobis 2,4-dimethylvaleronitrile; BODIPY 581/591, 4,4-di-fluoro-5-(4-phenyl-1,3-butadienyl)-4-bora-3a,4a-diaza-s-indacene-3-undecanoicacid; DCFH, 2′,7′-dichlorodihydrofluorescein; DCFH-DA, DCFH diacetate;DOX, doxorubicin; DTT, dithiothreitol; EDTA, ethylenediamine-tetraacetic acid;Hepes, N-(2-hydroxyethyl)piperazine-N′–(2-ethanesulfonic acid); HPLC, high-performance liquid chromatography; KCl, potassium chloride; LC/APCI-MS,liquid chromatography/atmospheric pressure chemical ionization-mass spectro-metry; LOX, lipoxygenase (soybean); MeO-AMVN, 2,2′-azobis(4-methoxy-2,4-dimethylvaleronitrile); NAD+, nicotinamide adenine dinucleotide; ROS, reactiveoxygen species; RT, retention time; TAP, total antioxidative performance; THF,tetrahydrofuran.⁎ Corresponding author. Departamento de Clinica Médica, Faculdade de
Medicina de Botucatu, UNESP, CEP 18618–970, Botucatu, SP, Brazil. Fax: +5514 3882 2969/3882 2238.
E-mail address: [email protected] (A.L.A. Ferreira).
0891-5849/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.freeradbiomed.2007.05.002
Introduction
The anthracycline, doxorubicin (DOX), is an excellent drugfor the treatment of a wide variety of human tumors. However,the development of irreversible cardiotoxicity has limited its use[1]. Due to the successful action of DOX as a chemotherapicagent, several strategies have been tried to prevent/attenuate theside effects of DOX. Although DOX-induced injury appears tobe multifactorial, a common denominator to most of theproposed mechanisms is cellular damage mediated by reactiveoxygen species (ROS) [2,3]. However, it remains controversialas to whether antioxidants can prevent such side effects [4–8].Among these molecules, metal ion chelators, vitamin A [9], β-carotene [6,7], and others have been widely investigated inrelation to DOX toxicity.

741A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
Lycopene is one of the most potent antioxidants among thedietary carotenoids due mainly to its many conjugated doublebounds [10,11]. The antioxidant activity of lycopene is mainlydependent on its scavenging properties of singlet oxygen [12]and peroxyl radicals [13]. Antioxidant properties of lycopeneagainst DOX-induced cardiomyopathy were first described byKarimi et al. [14]. Although lycopene has been shown to offersome protection against adriamycin-induced cardiomyopathy[14,15], its metabolism during this pathological condition is notknown.
Our laboratory has carried out extensive studies on themetabolism of carotenoids. We found that the cleavage of β-carotene to retinoids can occur by either an excentric or acentral cleavage pathway depending on the absence or pre-sence of antioxidants [16], and have identified various inter-mediates of the excentric cleavage of β-carotene [17–19].Interestingly, it was found that β-carotene may react witheither fatty acid hydroperoxides or their derivatives andquench alkoxyl and/or peroxyl radicals [20]. Furthermore,we have demonstrated that both central and excentric clea-vage of lycopene occur in the intestinal mucosa postmitochon-drial fraction in the presence of soy lipoxygenase (LOX) andhave partially identified the corresponding cleavage products[21].
The present study investigated the effect of long-term DOXtreatment on the myocardial levels and metabolism of lycopene.First, we determined lycopene levels in the myocardium oflycopene-supplemented rats with and without DOX treatment.Secondly, the effect of DOX therapy on lycopene metabolismwas evaluated in the rat. Thirdly, the effect of DOX therapy onthe antioxidant capacity in the myocardium was determined inthe rat. Finally, the antioxidant activity of DOX was evaluatedin vitro.
Fig. 1. Doxorubicin and
Materials and methods
Chemical products
Doxorubicin hydrocloride was purchased from Pharmacia(Milan, Italy) and from Sigma Chemical (St. Louis, MO) (Fig.1A). Idarubicin hydrochloride was obtained from SigmaChemical. Pentobarbital sodic (Cristalia, Paulinia, SP, Brazil)was used for anesthesia. All-trans-β-carotene (type IV),lycopene, α-tocopherol, ascorbic acid, albumin, and ammoniumacetate were purchased from Sigma Chemical. Lutein waspurchased fromKemin Industries (DesMoines, IA). Zeaxanthin,cryptoxanthin, 13-cis-β-carotene, 9-cis-βcarotene, and echine-none were kindly provided by Hoffmann-La Roche (Nutley, NJ).Deuterated all-trans-lycopene (12, 12′, 14, 14′, 19, 19, 19, 19′,19′, 19′-2H10-lycopene was provided by BASF AG (Ludwig-shafen, Germany) (Fig. 1B). Deuterated lycopene solutions wereprepared under red light immediately before use. The standardsof lycopene 1,2-epoxide, 2,6-cyclolycopene-1,5-epoxide I, 2,6-cyclolycopene-1,5-epoxide II, and 2,6-cyclolycopene-1,5-diolwere kindly provided by Dr. Frederick Khachik (University ofMaryland). Lycopene from tomato oleoresin, Lyc-O-Mato, was agift from LycoRed Natural Products Industries (Beer-Sheva,Israel). Solutions of carotenoids and retinoids were preparedunder red light immediately before use. HPLC-grade methanolandwater were obtained from J. T. Baker Chemical (Philipsburg,NJ). Methyl-tert-butyl ether was purchased from AldrichChemical (Milwaukee, WI). Hepes, soy lipoxygenase, NAD+,DTT, Tween 40, and formaldehyde were purchased fromSigma Chemical Co. All HPLC solvents were passed through a0.45-μm membrane filter before use. All carotenoid standardswere stored at −70°C until used. The fatty acid analogue 4,4-difluoro-5-(4-phenyl-1,3-butadienyl)-4-bora-3a,4a-diaza-s-
lycopene structures.

742 A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
indacene-3-undecanoic acid (BODIPY 581/591) was pur-chased from Molecular Probes (Eugene, OR). The radical ini-tiator, MeO-AMVN, was purchased from Wako Chemicals(Richmond, VA).
Animals
Male rats of the Wistar strain (400–500 g) were purchasedfrom CEMIB (Centro Multidisciplinar de Investigação Bioló-gica, Unicamp, Campinas, SP, Brazil). Animals were housed for7 weeks in the animal facility at Laboratorio Experimental atFaculdade de Medicina de Botucatu, UNESP and consumedwater and commercial diet [22] ad libitum. Rats were randomlyassigned to four groups: C (control, n=24), D (doxorubicin,n=40), L (lycopene, n=8), and DL (lycopene+doxorubicin,n=40). Intraperitoneal (ip) treatments of DOXwere adminis-tered at 3, 4, 5, and 6 weeks: Rats from C and L groups receivedan appropriate volume of 0.9% NaCl (ip), while D and DLreceived doxorubicin diluted in 0.9% NaCl divided in four equalinjections [each containing 4 mg/(kg body wt)] reaching acumulative dose of 16 mg/kg body wt. Lycopene oleoresinsupplement mixed with corn oil [5 mg lycopene/(kg body wt ·day)] was given daily by gavage to animals in the L and DLgroups, while corn oil alone was given to C and D groups. Toavoid differences in the energy provided, all groups received thesame corn oil volume [(∼0.5 mL/(kg body wt · day)]. After thefinal ip injections, all rats were observed for 1 week and thenkilled under penthobarbital sodic ip (50 mg/kg) anesthesia.Hearts were collected and stored at −80°C until carotenoidanalyses and TAPmeasurements. The upper half of the intestinesof the rats from the C and D groups was collected fordetermining lycopene metabolism using incubation procedures.
A high number of animals was used in the DOX-treatedgroup (n=40) and DOX and lycopene-treated (n=40) groups.These numbers were chosen because of the elevated mortality(N50%) in DOX groups that was observed in the pilot study.Twenty animals from D and 23 from DL groups died after aDOX injection prior to protocol completion. This mortality rateis consistent with other reports [9,23,24].
The protocol used was in accordance with Ethical Principlesfor Animal Research adopted by the Brazilian College of AnimalExperimentation (COBEA) and was approved by the localEthical Committee for Animal Research (Protocol No. 267).
The strategy for using different tissues (myocardium andintestinal mucosa) in the current study is as follows. Myocardialhomogenate was used to check lycopene absorption in lycopene-supplemented rats in the target organ (L and DL groups) and toevaluate the effect of DOX on cardiac lycopene levels (DLgroup). The cardiac lycopene level findings (DL group)stimulated us to evaluate the effect of DOX on lycopenemetabolism. Due to this, we incubated labeled lycopene (D10
lycopene) with cofactors and postmitochondrial from intestinalmucosa (C and D groups) as the most active tissue to studycarotenoid metabolism [16,21,25,26]. Afterward, we usedmyocardial homogenate once again to check for DOX inter-ference with oxidative stress by using the total antioxidant po-tential assay (C and D groups).
Lycopene preparation given by gavage
Tomato oleoresin, Lyc-O-Mato 6% dewaxed, was mixedwith corn oil and stored at 4°C in the dark until used. Itcontained 13-cis-β-carotene (1%), all-trans-β-carotene (5%),and total lycopene (95%). Of the total lycopene, 93% was in theall-trans form and 7% was as cis-lycopene isomers. The tomatooleoresin-corn oil mixture was stirred for 20 min in a water bathat 54°C before being fed to the animals. Each milliliter ofsolution contained 5 mg total lycopene. The stability oflycopene in corn oil was monitored by HPLC.
Carotenoid analyses in diet and myocardium
Diet and myocardium were analyzed as previously described[27]. The diet samples were extracted with hexane/acetone/ethanol/toluene (50/35/30/35, v/v/v/v), while the saponifiedcardiac tissue (left ventricle) was extracted using CHCl3/CH3OH (2/1, v/v) and hexane. All sample analyses were donein duplicate. All sample processing was carried out under redlight. The recovery of the added internal standard wasconsistently N90%.
HPLC analyses
Carotenoids in the diet and myocardium were analyzed usingthe HPLC system described below. The HPLC system was aWaters Alliance 2695 (Waters, Wilmington, MA) consisting of apump with chromatography bound to a 2996 programmablephotodiode array detector and a 2475 fluorescence detector, aC30 carotenoid column (3 μm, 150×4.6 mm, YMC, Wilming-ton, MA), and Empower software. The Waters 2996 program-mable photodiode array detector was set at 340 nm for retinoidsand 450 nm for carotenoids. The HPLC mobile phase wasmethanol/methyl-tert-butyl ether/water (83/15/2, v/v/v, 15 g/Lammonium acetate in the water, solvent A) and methanol/methyl-tert-butyl ether/water (8/90/2, v/v/v, 10 g/L ammoniumacetate in the water, solvent B). The gradient procedure, at a flowrate of 1 mL/min (16°C), was as follows: (1) 100% solvent Awasused for 2min followed by a 6-min linear gradient to 70% solventA; (2) a 3-min hold followed by a 10-min linear gradient to 45%solvent A; (3) a 2-min hold, and then a 10-min linear gradient to5% solvent A; (4) a 4-min hold, and then a 2-min linear gradientback to 100% solvent A. Using this method, lutein, zeaxanthin,cryptoxanthin, 13-cis β-carotene, all-trans-β-carotene, 9-cis-β-carotene, 9-cis-β-carotene, 13-cis-, 15-cis-, and all-trans-lyco-pene were adequately separated. Carotenoids and retinoids werequantified by determining peak areas in the HPLC chromato-grams calibrated against known amounts of standards. Theamounts were corrected for extraction and handling losses bymonitoring the recovery of the internal standards.
Measurement of total antioxidant performance (TAP)
Liposome preparationLiposomes were prepared as follows. The soybean phos-
phatidylcholine was dissolved in chloroform and evaporated

743A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
very slowly under a gentle stream of nitrogen, rolling the flaskto deposit a thin lipid film. The flask was then maintained undera nitrogen stream for an additional 20 min to eliminate residualsolvent. Then the lipid film was rehydrated with phosphate-buffered saline (PBS, 40 mM, pH 7.4) to a concentration of2.5 mg of phospholipids/mL and frozen and thawed 3 timesfollowed by sonication (Branson Sonifer 450, Danbrrry, CT)under ice for 30 s and repeated 3 times.
Effect of DOX on TAPThe TAP of rat myocardium was quantified by comparing
the area under the curve (AUC) relative to the oxidation kineticsof BODIPY with respect to a phosphatidylcholine (PC)liposome suspension used as the reference lipid matrix [28].The TAP method was applied for the measurement of theantioxidant status of myocardium from animals treated with (D)and without (C) DOX.
Myocardium (left ventricle) was weighed (0.3 g), minced,and homogenized for 20 s on ice with 3 mL of phosphate buffer(100 mM, pH 7.4) using a Brinkmann polytron homogenizer(Westbury, NY). Aliquots of supernatant were collected aftercentrifugation of the cardiac tissue homogenate at 800 g for15 min in a Sorvall RT 6000 refrigerated centrifuge (Du PontCo., Newtown, CT). The aliquots from cardiac tissue sampleswere used for TAP procedures. The BCA Protein Assay wasused to determine protein concentration.
As reported previously [29], 200 μL of cardiac tissuesamples was used for TAP assay. After adding 200 μL of PBS,BODIPY 581/591 (100 μL) was incorporated into the lipidcompartment of tissue at a final concentration of 2 μmol/L byvortexing gently for 10 s at low speed followed by 10 minincubation at 37°C. After incubation, cold phosphate buffer(485 μL, pH 7.4) was added and vortexed gently for 10 s at lowspeed. The radical initiator, MeO-AMVN (15 μL, finalconcentration 2 mM), was added into the reaction mixturewith gentle mixing with a magnetic stirring bar. Externalstandard curves were obtained by the oxidation kinetics of acontrol solution in the presence of butylated hydroxytoluene(BHT) at various concentrations (2, 4, and 8 μmol/L). Theoxidation of tissue samples was monitored by the greenfluorescent oxidation product of BODIPY 581/591 (finalconcentration λex=500, λem=520 nm, slit 10 nm) using amicroplate reader (Wallac Vitor 2, Perkin-Elmer, Boston, MA).
PC liposomes were used for all in vitro studies in order todetermine total antioxidant performance of DOX. Doxorubicinhydrochloride was added to the incubation solution prior to theincubation at final concentrations of 1, 2, 4, and 8 μM. Anotheranthracycline analog widely used in chemotherapy, idarubicinhydrochloride, was also studied at the same concentrations ofdoxorubicin hydrochloride.
All sample analyses were done in triplicate.
Incubation procedure
Lycopene purificationD10 lycopene was purified by neutral aluminum oxide
column chromatography [20].
Sample preparation and incubationThe preparation of the postmitochondrial fraction and its
incubation with lycopene were essentially the same as describedpreviously [16,21,30] with minor alterations. Briefly, the freshupper half of the intestinal mucosa was homogenized with50 mM Hepes buffer (pH 7.4). The postnuclear fraction waspurified by applying to an Econo-Pac 10 DG disposable column(30×10 mL, Bio-Rad Laboratories, Hercules, CA). Thepostmitochondrial fractions were used 24 h to 2 weeks aftercollection. The protein concentration was determined using theBCA Protein Assay (Pierce Co., Rockford, IL). The standardincubation mixture contained 2.0 mg of protein fraction, 20 mMHepes buffer, pH 8.0, 800 units LOX, 0.2 mM NAD+, 2 mMDTT, 150 mM KCl, in a total volume of 1000 μL. Afterpreincubation at 37°C in a shaking water bath for 5 min, theenzyme reaction was started by adding 40 μL of lycopenesolubilized in aqueous Tween 40 to 920 μL of the incubationmixture with a final concentration of 2.5 μM lycopene. Thevials were uncovered and the incubation mixture was exposedto room air as the gas phase. After incubation for up to 90 min at37°C, the incubation was terminated by adding 100 μL offormaldehyde (37%, w/w), and postincubated for 10 min at37°C. Three control vials (Blank) were run lacking lycopeneand/or the protein fraction. Based on the pilot and the previousstudy we carried out the incubation procedure with bothintestinal mucosa postmitochondrial fraction and LOX to obtainmaximum metabolite production [21].
Crystalline D10 lycopene was solubilized in Tween 40/acetone (1/19, v/v) in a 1:10,000 ratio by weight (0.3 mg of D10
lycopene dissolved in 3.3 g of Tween 40/acetone) in a quartztube since lycopene tends to stick to the surface of regular glasstest tubes. To yield a clear mixture, the lycopene solution wasvortexed and sonicated 5×3 min under ice, and the acetone wasevaporated under N2. Water (9 mL) was then added, and thesolution was again vortexed and sonicated 5×3 min at 4°C. Thelycopene solution was added to the incubation mixture, as soonas it was prepared. All experimental procedures were repeatedthree to eight times and carried out under red light.
The intestinal tissue, a major tissue at where carotenoidmetabolism takes place, was chosen for elucidating lycopenemetabolism. The postmitochondrial fraction was prepared as theenzymes involved in carotenoid metabolism are reported to berich in this fraction [16,21,25,26]. Lipoxygenase, which isshown to stimulate lycopene cleavage [21], was added to theincubation system in order to increase production of lycopenemetabolites.
HPLC analyses
Extraction of the postmitochondrial fraction from intestinalmucosa was done using 1.5 mL of CHCL3/CH3OH (2/1, v/v)plus 0.5 mL of hexane, followed by 1.5 mL of hexane. Themixture was centrifuged for 20 min at 800g at 4°C. Aftercentrifugation, the chloroform and hexane layers were com-bined and evaporated to dryness under N2. The residue wasredissolved in 150 μL ethanol, sonicated twice for 30 s eachtime, and vortexed for 30 s. A 50 μL aliquot of the final extract

Fig. 2. Concentrations of total lycopene in myocardium from rats after tomatooleoresin supplementation (5 mg lycopene/(kg body wt · day/7week) and DOXtherapy (4 mg DOX/kg body wt · week/4week). Groups: C, control (n=24); D,doxorubicin (n=20); L, lycopene (n=8); DL, doxorubicin+ lycopene (n=17).Values are means±SE, n, sample number; ND, not detectable; Student's t testwas used to compare L and DL groups, PN0.05.
744 A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
was injected into the HPLC system after centrifugation for2 min at 800g at 4°C.
The HPLC system consisted of Waters 717 plus autosampler(Waters Chromatography Division of Millipore Corp., Milford,MA), Perkin-Elmer Series 200 Diode Array Detector II, LCpump, and a Pecosphere-3 C18 0.46×8.3 cm cartridge columnand a Total Chrom Workstation (Perkin-Elmer Instruments,Shelton, CT). The HPLC mobile phase was CH3CN/THF/H2O(50/20/30, v/v/v, 1% ammonium acetate in H2O, solvent A) andCH3CN/THF/H2O (50/44/6, v/v/v, 1% ammonium acetate inH2O, solvent B). The gradient procedure at a flow rate of1 mL/min was as follows: 90% solvent A and 10% solvent Bwere used for 2 min followed by a 12 min linear gradient to100% solvent B, a 15 min hold at 100% solvent B, and then a2 min gradient back to 90% solvent A and 10% of solvent B.The Waters 490E multiwavelength spectrophotometer detectorwas set at 340, 375, and 450 nm, respectively. Unknown peakswere quantified by determining peak areas in the HPLC chro-matograms. Furthermore, absorption spectra were recorded bythe diode array detector for the unknown peaks and comparedwith standards.
LC/APCI-MS analysis of metabolites
The final extracts from the incubations were redissolved in100 μL ethanol and evaporated to dryness under N2. Theresidue was resuspended in 600 μL CHCl3 and applied to Sep-Pak Vac NH2 cartridges (Waters, Milford, MA) rinsed withhexane. The Sep-Pak column was then eluted with 6 mL hexaneand fraction 1 was collected. The Sep-Pak column was theneluted with 6 mL CHCl3/3-propanol (2/1) and fraction 2 wascollected. Both fractions were evaporated under N2, and theresidue was resuspended in 120 μL ethanol for LC/MS analysisusing atmospheric pressure chemical ionization (APCI). Thesame mobile phase described for HPLC (except the absence ofammonium acetate) was pumped through a 0.005 in. ID PEEKby an Agilent 1100 LC pump at a flow rate of 1.0 mL/min. TheAgilent 1100 HPLC was equipped with an autosampler, UV/VIS diode array detector, and a quaternary pump. An EsquireLC (Wilmington, DE) mass spectrometer (Bruker Daltonic,Inc., Bremen, Germany) with APCI source and ion trap wasused for analyses. APCI was performed in the positive mode[21]. The vaporizer temperature, drying gas temperature, andcorona current were all optimized with regard to maximumsignal intensity of molecular ions. The following optimalconditions were used: vaporizer temperature, 300°C; nebulizerpressure, 50 psi of N2; drying gas temperature 350°C. Thecountercurrent drying gas was supplied from a liquid N2
cylinder, and was optimized at a flow rate of 4.0 L/min. TheAPCI source was a corona discharge from a needle generated bythe following potentials: corona needle=+2200 V, endplateoffset=−500 V, and capillary=−4000 V. The mass scan rangewas at 200 to 700 Da in 0.1-Da steps with an accumulation timeat 50 s and a half-maximum peak width of 0.6 Da. From the LC/APCI-MS analysis, the mass spectra for unknown metaboliteswith a characteristic isotopic profile similar to D10 lycopenewere identified to determine their mass.
Statistical analysis
Results are expressed as mean±SE and the significance ofdifferences was calculated by Student's t test and ANOVAusing SigmaStat version 2.0 for Windows 95, NT and 3.1(Jandel Scientific Software, San Rafael, CA). Differences wereconsidered significant when Pb0.05.
Results
Lycopene concentrations in diet, tomato oleoresin, andmyocardium
No lycopene was detected in the diets fed to the animal.From the tomato oleoresin corn oil mixture, the animalsreceived 5.0, 0.77 and 2.0 mg/(kg body wt · day) of lycopene,13-cis-β-carotene, and all-trans-β-carotene, respectively. Lyco-pene was determined as both cis and trans isomers in myo-cardium. After 7 weeks of tomato oleoresin supplementation,myocardial lycopene levels reached the mean of 49 nmol/kg inDL and 45 nmol/kg in L groups. There was no detectable lyco-pene in the myocardium of C or D groups (Fig. 2).
Effect of DOX on total antioxidant performance
Table 1 shows the results of the the total antioxidantperformance assay in the myocardium of animals treated withDOX (D, n=10) and without DOX (C, n=10). Mean TAPvalues whether adjusted for cardiac tissue protein concentra-tions, adjusted for tissue weight, or not adjusted were higher inthe myocardium from the D group than that of C group.
Fig. 3 shows the oxidation kinetics of PC liposomes in thepresence of either doxorubicin hydrochloride or idarubicinhydrochloride at 1, 2, 4, and 8 μM. Both doxorubicin hydro-

Table 1Total antioxidant performance of myocardium from control and DOX animals
C DOX Pn=10 n=10
TAP (BHTeq.) 19.48±9.29 50.62±32.21 0.009850TAP (BHTeq.)/protein 21.43±9.52 73.47±42.45 0.002054TAP (BHTeq.)/g 62.37±29.73 163.56±104.78 0.010496Protein a (g) 0.92±0.17 0.67±0.80 0.00418Tissue wt (g) 0.31±0.003 0.31±0.006 N0.05
Values are means±SE, n, sample number; C, control group; DOX, doxorubicingroup; wt, wet weight; Student's t test was used to compare C and DOX groups;TAP, total antioxidant performance; BHTeq, BHT equivalent; radicals initiatedby MeO-AMVN, and lipid peroxidation was determined by oxidation ofBODIPY 581/591.a Protein concentrations determined using BCA.
Fig. 3. Oxidation kinetics of the total antioxidant performance assay using 0, 1,2, 4, and 8 μM of either doxorubicin (A) or idarubicin (B). Each cyclecorresponds to 5 min.
745A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
chloride and idarubicin hydrochloride prevented the oxidationof control solution (PC liposomes) in a dose-dependent manner.
Cleavage products of lycopene
Peak identification by HPLCAfter incubation of lycopene alone, in the absence of
intestinal mucosa postmitochondrial fraction and LOX, therewere only minor peaks observed at 340 nm (Fig. 4A) and somedistinct peaks at 450 nm (Fig. 4B), suggesting that there wassome autoxidation of lycopene during the 90 min incubation at37°C. After incubation of lycopene with intestinal mucosapostmitochondrial fraction and LOX, new peaks were observedin the HPLC analysis at both 340 (Figs. 4C and 4E) and 450 nm(Figs. 4D and 4F). We were able to identify seven major peaks(peaks: 1, 2, 2′, 3, 4, 5, 5′) based on their UV-visible spectra.These peaks were analyzed further by LC/APCI-MS todetermine the possible mass of each peak. As shown in Fig.4, peak 1 had a retention time of 3.5 min and was seen at awavelength setting at 340 nm (Figs. 4C and 4E). Peaksmonitored at 450 nm were at retention times 9.7, 10.9, 14.4,18.1, 21.0, and 21.7 min, and were labeled as peaks 2, 2′, 3, 4, 5,and 5′, respectively (Figs. 4D and 4F). Relatively minor peaksin the HPLC profile were not studied for identification becausethese peaks consisted of a mixture of products without well-defined spectral properties, or were the same as the peaksappearing in the absence of intestinal mucosa and LOXincubations.
When LOX and intestinal mucosa postmitochondrial frac-tion either from control or DOX-treated rats were added to theincubation mixture, the peak areas for the all peaks weresignificantly increased (except peak 4, PN0.05). This augmen-tation was 1- to 5-fold over baseline (blank) (Table 2).
When the incubation mixture using LOX+intestinal mucosapostmitochondrial fraction from control rats was compared toLOX+intestinal mucosa postmitochondrial fraction from DOXrats, we identified different quantitative production for allpeaks at 450 nm. Except for peak 4 (PN0.05), all peaksidentified at 450 nm were significantly lower than those ofcontrol rats. However, peak 1 identified at 340 nm from DOXrats was significantly higher than those of control rats (Table 2,Fig. 4).
Peak identification using UV/Vis and LC/APCI-MSThe in vitro metabolites which were collected from the
HPLC system were applied to LC-APCI-MS (positive-ionmode) to determine the molecular masses. Metabolites wereidentified by the presence of a semisymmetrical pattern ofmolecules, each differing by a single mass unit attributable tothe deuterium from the parent D10 lycopene [21]. Theincubation of deuterated lycopene with both postmitochon-drial fractions of rat intestinal mucosa and lipoxygenaseproduced two groups of enzymatic products: cleavage (peak1) and oxidative (peaks 2–5′) products. All the proposedstructures of the metabolites from the incubation are presentedin Fig. 5.
Enzymatic cleavage products of lycopene
Peak 1 (retention time=3.5 min) had a single absorptionpeak at 365 nm and m/z=277 for the protonated molecularmass. When corrected for H+ and for 4 deuterium atoms, themolecular mass was 272 and the proposed composition isC18H24O2. The absorption maximum (365 nm) of this peak issimilar to a compound with a carbonyl conjugated to 4 doublebonds (365 nm) and was reported as a product of autoxidationof lycopene [31] and also from our previous study [21]. Sincepeak 1 has an additional atom of oxygen, we are proposing thestructure for peak 1 as 3-keto-apo-13-lycopenone (Fig. 5A).
Enzymatic oxidative cleavage products of lycopene
Peak 2 (retention time=9.7 min) has a λmax=409, 434, and460 nm and m/z=521 for a protonated molecular ion. Cor-recting for H+ and 10 deuterium atoms would give a m/z=510,

Fig. 4. The oxidative and enzyme-catalyzed cleavage products of lycopene. Purified D10 lycopene 2.5 μM (solubilized in aqueous Tween 40) plus LOX-800 units, andcofactors (NAD+, DTT) were incubated in the absence (A, B) or presence (C, D, E, F) of purified postmitochondrial fraction of rat intestine (2 mg protein) from controlgroup (C, D) and doxorubicin group (E, F) for 90 min at 37°C in a total volume of 1 mL. The HPLC profiles of the extracts are displayed when monitored at both340 nm (A, C, E) and 450 nm (B, D, F). The peak at 24 min is lycopene.
746 A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
corresponding to a C37H50O composition. The spectrumindicates that there are no conjugated carbonyls, and is verysimilar to ε,ε-carotene with 9 conjugated double bonds [32], andit was identified in our previous study [21]. The structureproposed is 2-ene-5,8-lycopene-furanoxide (Fig. 5B).
Peak 2′ (retention time=10.9 min) has a λmax=328, 409,434, and 460 nm and m/z=521 for a protonated molecular ion.Correcting for 10 deuterium atoms would give a m/z=510,corresponding to a C37H50O composition and is similar to peak2. Since peak 2′ has an additional absorption peak at 328 nm,we are proposing its structure as cis-2-ene-5,8-lycopene-furanoxide (Fig. 5B).
Peak 3 (retention time=14.4 min) showed λmax=435, 457,and 490 nm and m/z=579 for (M+H)+. Correcting for 10deuterium atoms yields a mass of 568, corresponding to aC40H56O2 composition. Since a cis peak at 346 nm wasobserved, it was assigned as cis-lycopene 1,2,5′,6′-diepoxide(Fig. 5B). This structure had not been identified in our previousexperiment. However, this assignment matches the previousreport on the diepoxide formed from oxidation of lycopene withm-chloroperoxybenzoic acid [33].
Peak 4 (retention time=18.1 min) has a λmax=415, 440,and 472 nm and m/z=579 for (M+H)+. Correcting for 10deuterium atoms yields a mass of 568, corresponding to aC40H56O2 composition. It was assigned as lycopene 5,6,5′,6′-diepoxide (Fig. 5B). This assignment is in agreement with thestructure with an identical mass and absorption maximumdescribed earlier [21,34].
Peak 5 (retention time=21.0 min) had λmax=348, 362, 440,464, and 495 nm and m/z=563 for (M+H)+. Correcting for 10deuterium atoms yields a mass of 552, corresponding to aC40H56O composition. Since the most intensive absorbance is at464 nm, peak 5 closely matches the UVabsorption of lycopene1,6,2,5-diepoxide identified earlier with UV absorbance at 351,439, 465, and 497 [33]. Given that the mass of peak 5 showedonly one oxygen atom in the molecule and UV absorbance at348 and 362 of a typical cis peak absorbance, it was assigned ascis-lycopene 1,6-epoxide (Fig. 5B). This structure has not beenidentified before.
Peak 5′ (retention time=21.7 min) had λmax=441, 466, and497 nm and m/z=563 for (M+H)+. Correcting for 10 deuteriumatoms yields a mass of 552, corresponding to a C40H56O
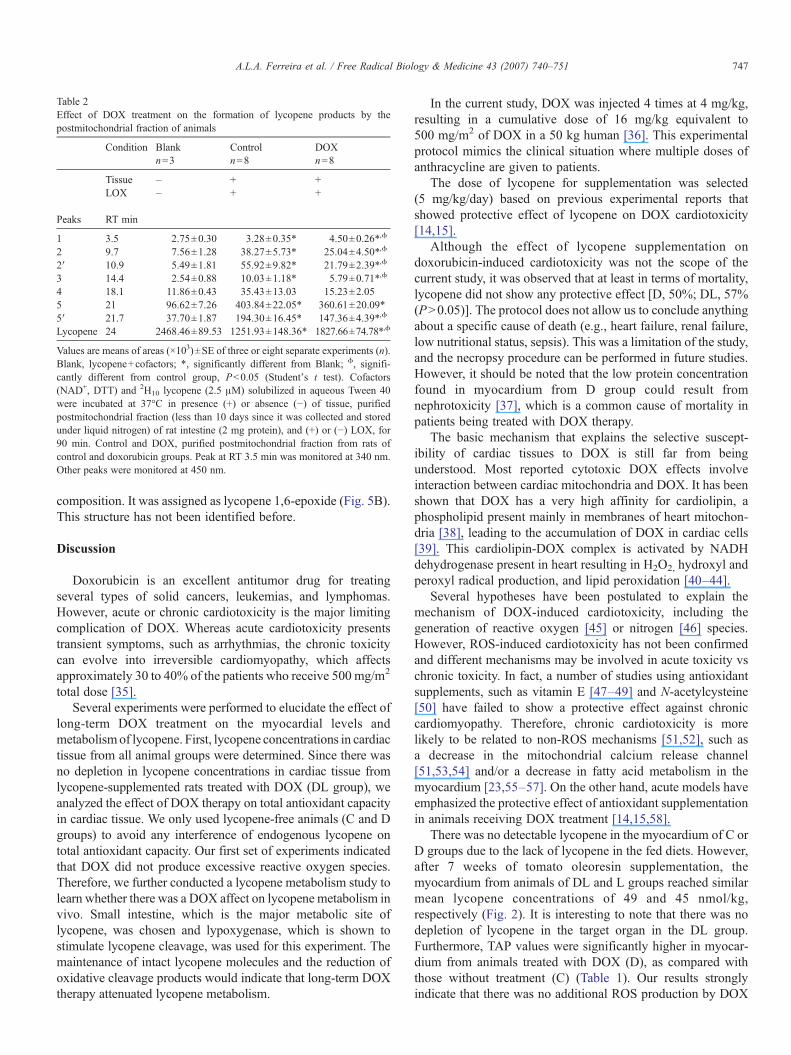
Table 2Effect of DOX treatment on the formation of lycopene products by thepostmitochondrial fraction of animals
Condition Blank Control DOXn=3 n=8 n=8
Tissue – + +LOX – + +
Peaks RT min
1 3.5 2.75±0.30 3.28±0.35* 4.50±0.26*,ϕ
2 9.7 7.56±1.28 38.27±5.73* 25.04±4.50*,ϕ
2′ 10.9 5.49±1.81 55.92±9.82* 21.79±2.39*,ϕ
3 14.4 2.54±0.88 10.03±1.18* 5.79±0.71*,ϕ
4 18.1 11.86±0.43 35.43±13.03 15.23±2.055 21 96.62±7.26 403.84±22.05* 360.61±20.09*5′ 21.7 37.70±1.87 194.30±16.45* 147.36±4.39*,ϕ
Lycopene 24 2468.46±89.53 1251.93±148.36* 1827.66±74.78*,ϕ
Values are means of areas (×103)±SE of three or eight separate experiments (n).Blank, lycopene+cofactors; *, significantly different from Blank; ϕ, signifi-cantly different from control group, Pb0.05 (Student's t test). Cofactors(NAD+, DTT) and 2H10 lycopene (2.5 μM) solubilized in aqueous Tween 40were incubated at 37°C in presence (+) or absence (−) of tissue, purifiedpostmitochondrial fraction (less than 10 days since it was collected and storedunder liquid nitrogen) of rat intestine (2 mg protein), and (+) or (−) LOX, for90 min. Control and DOX, purified postmitochondrial fraction from rats ofcontrol and doxorubicin groups. Peak at RT 3.5 min was monitored at 340 nm.Other peaks were monitored at 450 nm.
747A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
composition. It was assigned as lycopene 1,6-epoxide (Fig. 5B).This structure has not been identified before.
Discussion
Doxorubicin is an excellent antitumor drug for treatingseveral types of solid cancers, leukemias, and lymphomas.However, acute or chronic cardiotoxicity is the major limitingcomplication of DOX. Whereas acute cardiotoxicity presentstransient symptoms, such as arrhythmias, the chronic toxicitycan evolve into irreversible cardiomyopathy, which affectsapproximately 30 to 40% of the patients who receive 500 mg/m2
total dose [35].Several experiments were performed to elucidate the effect of
long-term DOX treatment on the myocardial levels andmetabolismof lycopene. First, lycopene concentrations in cardiactissue from all animal groups were determined. Since there wasno depletion in lycopene concentrations in cardiac tissue fromlycopene-supplemented rats treated with DOX (DL group), weanalyzed the effect of DOX therapy on total antioxidant capacityin cardiac tissue. We only used lycopene-free animals (C and Dgroups) to avoid any interference of endogenous lycopene ontotal antioxidant capacity. Our first set of experiments indicatedthat DOX did not produce excessive reactive oxygen species.Therefore, we further conducted a lycopene metabolism study tolearn whether there was a DOX affect on lycopene metabolism invivo. Small intestine, which is the major metabolic site oflycopene, was chosen and lypoxygenase, which is shown tostimulate lycopene cleavage, was used for this experiment. Themaintenance of intact lycopene molecules and the reduction ofoxidative cleavage products would indicate that long-term DOXtherapy attenuated lycopene metabolism.
In the current study, DOX was injected 4 times at 4 mg/kg,resulting in a cumulative dose of 16 mg/kg equivalent to500 mg/m2 of DOX in a 50 kg human [36]. This experimentalprotocol mimics the clinical situation where multiple doses ofanthracycline are given to patients.
The dose of lycopene for supplementation was selected(5 mg/kg/day) based on previous experimental reports thatshowed protective effect of lycopene on DOX cardiotoxicity[14,15].
Although the effect of lycopene supplementation ondoxorubicin-induced cardiotoxicity was not the scope of thecurrent study, it was observed that at least in terms of mortality,lycopene did not show any protective effect [D, 50%; DL, 57%(PN0.05)]. The protocol does not allow us to conclude anythingabout a specific cause of death (e.g., heart failure, renal failure,low nutritional status, sepsis). This was a limitation of the study,and the necropsy procedure can be performed in future studies.However, it should be noted that the low protein concentrationfound in myocardium from D group could result fromnephrotoxicity [37], which is a common cause of mortality inpatients being treated with DOX therapy.
The basic mechanism that explains the selective suscept-ibility of cardiac tissues to DOX is still far from beingunderstood. Most reported cytotoxic DOX effects involveinteraction between cardiac mitochondria and DOX. It has beenshown that DOX has a very high affinity for cardiolipin, aphospholipid present mainly in membranes of heart mitochon-dria [38], leading to the accumulation of DOX in cardiac cells[39]. This cardiolipin-DOX complex is activated by NADHdehydrogenase present in heart resulting in H2O2, hydroxyl andperoxyl radical production, and lipid peroxidation [40–44].
Several hypotheses have been postulated to explain themechanism of DOX-induced cardiotoxicity, including thegeneration of reactive oxygen [45] or nitrogen [46] species.However, ROS-induced cardiotoxicity has not been confirmedand different mechanisms may be involved in acute toxicity vschronic toxicity. In fact, a number of studies using antioxidantsupplements, such as vitamin E [47–49] and N-acetylcysteine[50] have failed to show a protective effect against chroniccardiomyopathy. Therefore, chronic cardiotoxicity is morelikely to be related to non-ROS mechanisms [51,52], such asa decrease in the mitochondrial calcium release channel[51,53,54] and/or a decrease in fatty acid metabolism in themyocardium [23,55–57]. On the other hand, acute models haveemphasized the protective effect of antioxidant supplementationin animals receiving DOX treatment [14,15,58].
There was no detectable lycopene in the myocardium of C orD groups due to the lack of lycopene in the fed diets. However,after 7 weeks of tomato oleoresin supplementation, themyocardium from animals of DL and L groups reached similarmean lycopene concentrations of 49 and 45 nmol/kg,respectively (Fig. 2). It is interesting to note that there was nodepletion of lycopene in the target organ in the DL group.Furthermore, TAP values were significantly higher in myocar-dium from animals treated with DOX (D), as compared withthose without treatment (C) (Table 1). Our results stronglyindicate that there was no additional ROS production by DOX

Fig. 5. Proposed structures of the metabolites detected through the incubation of D10-lycopene with rat intestinal mucosa postmitochondrial fraction and soylipoxygenase. The purified postmitochondrial fraction of rat intestine (2 mg protein) was incubated at 37°C for 90 min with LOX (800 units), cofactors (NAD+, DTT),and 2.5 μM lycopene solubilized in aqueous Tween 40.
748 A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
treatment in our animal model. The current results are inagreement with other studies where DOX did not induce thereduction of lipophilic [59–61] antioxidant components incardiac tissues. In fact, the maintenance of some antioxidantdefense components after DOX infusion in animal hearts hasbeen reported. Retinol and α-tocopherol concentrations [9],glutathione peroxidase, superoxide dismutase, catalase activ-ities, and GSH levels [59–61] were all found to be unchanged inhearts from animals subjected to DOX therapy. Similar resultswere seen in humans such as reduction of lipid peroxidationproducts [62], unchanged plasma α-tocopherol [63], andunchanged plasma β-carotene levels [64]. Furthermore, our invitro experiment, showing antioxidant activity of DOX in adose-dependent-manner (Fig. 3), strongly indicates antioxidantactivity of DOX. As far as we know, this is the first time that theantioxidant capacity of an anthracycline has been assessed byTAP. Thus, taken all together, these results suggest that DOXtherapy given under the current conditions may actually reducecardiac lipid peroxidation.
The probable mechanism that explains the lack of oxidativestress found in myocardium from animals treated with DOX canbe in part explained by alterations in lipoxygenase activity andcalcium concentration identified with long-term DOX therapy.Mitochondrial cardiac lipid peroxidation (cardiolipin) alsodepends on LOX activity, which is affected by calciumconcentration [65,66]. This event sequence was confirmed inchronic cardiomyopathy induced by DOXwhere the cumulativedose (12–18 mg/kg) given fractionally resulted in an irrever-sible decrease in the amount of calcium release of thesarcosplasmatic reticulum [53] and of cardiac mitochondria[54]. Even though we did not evaluate calcium loading capacitybecause this was not the aim of our study, several studiessupport this hypothesis. The low cardiac inflammation levelfound in rats [67] or patients [68] under long-term DOX therapyis also another interesting event that can be related to the lowoxidative stress status.
In order to avoid interference of supplemental lycopene, Cand D groups were selected to analyze total antioxidant per-

749A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
formance in myocardium and lycopene metabolism in theintestinal mucosa postmitochondrial fraction.
Although lycopene has been shown to offer some protectionagainst DOX-induced cardiomyopathy [14,15], the metabolismof lycopene in this pathological condition is not known. Tostudy the metabolism of lycopene, intestinal mucosa post-mitochondrial fraction, several cofactors and LOX were used.As previously demonstrated, we added LOX to obtainmaximum production of the metabolic products [21]. Deuter-ated lycopene (2H10) was used with a characteristic enrichmentprofile that helped us to identify the lycopene cleavageproducts.
The current results showed that both cleavage and oxidationof lycopene occur in rat intestinal mucosa postmitochondrialfraction in the presence of soy LOX, as demonstrated in ourrecent study [21]. Lycopene metabolites consisted of bothenzyme-catalyzed cleavage products (peak 1) as well asoxidative products (peak 2–5′). When compared with ourprevious study, new oxidative products were found, such aspeak 3, peak 5, and peak 5′. This difference may be due to thedifferent rat strain used. Only one of those peaks (peak 3) hasbeen shown earlier as an epoxide product [33]. Peaks 5 and 5′have not been reported earlier with a single oxygen atomincorporated into the lycopene molecule at the 1 and 6 positions(Fig. 5).
The quantification of these products revealed that, whencompared to C group, intestinal mucosa postmitochondrialfraction from D group had significantly higher amounts of intactlycopene and lower oxidative cleavage products, suggestingthat DOX may have contributed to preventing the lycopenebreakdown process and therefore preserving lycopene in itsintact form (Table 2, Fig. 4). This is an important issue becauseit can in part explain why DOX therapy did not depletelycopene levels in cardiac tissues from animals supplementedwith lycopene. From these data, we cannot suggest that cancerpatients under DOX therapy should or not eat tomato products.Our results indicated that DOX seems to retard lycopenemetabolism (i.e., preserves D10 lycopene in its intact form (all-trans-) and lowers oxidative cleavage products of D10
lycopene). Considering that the intact lycopene has higherantioxidant activity as compared to those of its metabolites, andthat there is no known toxicity of lycopene, it may be beneficialfor cancer patients with DOX therapy (especially in the acutetreatment) to consume lycopene-rich foods such as tomatoes ortomatoes products.
Our results show that DOX: (1) maintains lycopene levels inmyocardium of Wistar rats supplemented with lycopene; (2)increases total antioxidant performance in myocardium ofWistar rats; (3) decreases oxidative cleavage of lycopene ofWistar rats; (4) prevents oxidation of phosphatidylcholineliposomes in a dose-dependent manner, in vitro. Therefore, it ishighly probable that the mechanism of chronic DOX-inducedcardiotoxicity is related to mechanisms other than ROSinterference. Although the experimental design using multipledoses of DOX did mimic the clinical situation, it would beimprudent to extrapolate the current results to humans.However, the present study represents a contribution to our
understanding of the metabolism of lycopene and myocardiumoxidative stress under chronic DOX treatment.
Acknowledgments
Supported in part by Fundação de Amparo à Pesquisa doEstado de São Paulo, FAPESP, São Paulo, SP, Brazil under acontract number 03/13979-3 and by a grant from the U.S.Department of Agriculture, Agricultural Research Service underCooperative Agreement number 1950-51000-065. Any opi-nions, findings, conclusion, or recommendations expressed inthis publication are those of the author(s) and do not necessarilyreflect the view of the UNESP or U.S. Dept of Agriculture. Wethank Alexandre L. Loureiro, Corina Correa, Elizabeth Johnson,Jose A. Souza, Jose C. Georgete, Jian Qin, Mario A. Dallaqua,Mario B. Bruno, Rogerio A. Monteiro, Sandra A. Fábio, SueliClara, and Vitor M. Souza for their help in this study.
References
[1] Buja, L. M.; Ferrans, V. J.; Mayer, R. J.; Roberts, W. C.; Henderson, E. S.Cardiac ultraestructural changes induced by daunorubicin therapy. Cancer32:771–788; 1973.
[2] Li, T.; Danelisen, I.; Singal, P. K. Early changes in myocardial antioxidantenzymes in rats treated with adriamycin. Mol. Cell. Biochem. 232:19–26;2002.
[3] Lee, V.; Randhawa, A. K.; Singal, P. K. Adriamycin-induced myocardialdysfunction in vitro is mediated by free radicals. Am. J. Physiol. 261;1991.
[4] Wahab, M. H. A.; Akoul, E. E. M. S.; Abdelaziz, A. H. Modulatory effectsof melatonin and vitamin-E on doxorubicin-induced cardiotoxicity inEhrlich ascites carcinoma bearing mice. Tumori 86:157–162; 2000.
[5] Antunes, L. M.; Takahashi, C. S. Effects of high doses of vitamins C and Eagainst doxorubicin-induced chromosomal damage in Wistar rat bonemarrow cells. Mutat. Res. 419:137–143; 1998.
[6] Vile, G. F.; Winterbourn, C. C. Inhibition of adriamycin-promotedmicrosomal lipid peroxidation by beta-carotene, alpha-tocopherol andretinol at high and low oxygen partial pressures. FEBS Lett. 238:353–356;1988.
[7] Teicher, B. A.; Schwartz, J. L.; Holden, S. A.; Ara, G.; Northey, D. In vivomodulation of several anticancer agents by beta-carotene. CancerChemother. Pharmacol. 34:235–241; 1994.
[8] van Acker, F. A.; Hulshof, J. W.; Haenen, G. R.; Menge, W. M.; van derVijgh, W. J.; Bast, A. New synthetic flavonoids as potent protectors againstdoxorubicin-induced cardiotoxicity. Free Radic. Biol. Med. 31:31–37;2001.
[9] Danelisen, I.; Palace, V.; Lou, H.; Singal, P. K. Maintenance of myocardiallevels of vitamin A in heart failure due to adriamycin. J. Mol. Cell.Cardiol. 34:789–795; 2002.
[10] Arab, L.; Steck, S. Lycopene and cardiovascular disease. Am. J. Clin. Nutr.71:1691S–1695S; 2000.
[11] Stahl, W.; Sies, H. Physical quenching of singlet oxygen and cis-transisomerization of carotenoids. Ann. N. Y. Acad. Sci. 691:10–19; 1993.
[12] Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biologicalcarotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 274:532–538; 1989.
[13] Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Aspects Med.24:345–351; 2003.
[14] Karimi, G.; Ramezani, M.; Abdi, A. Protective effects of lycopene andtomato extract against doxorubicin-induced cardiotoxicity. Phytother. Res.19:912–914; 2005.
[15] Yilmaz, S.; Atessahin, A.; Sahna, E.; Karahan, I.; Ozer, S. Protective effectof lycopene on adriamycin-induced cardiotoxicity and nephrotoxicity.Toxicology 218:164–171; 2006.

750 A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
[16] Yeum, K.-J.; Ferreira, A. L. A.; Smith, D.; Krinsky, N. I.; Russell, R. M.The effect of a-tocopherol on the oxidative cleavage of β-carotene. FreeRadic. Biol. Med. 29:105–114; 2000.
[17] Tang, G. W.; Wang, X. D.; Russell, R. M.; Krinsky, N. I. Characterizationof beta-apo-13-carotenone and beta-apo-14′-carotenal as enzymaticproducts of the excentric cleavage of beta-carotene. Biochemistry 30(41):9829–9834; 1991.
[18] Wang, X.-D.; Krinsky, N. I.; Tang, G.; Russell, R. M. Retinoic acid can beproduced from excentric cleavage of β-carotene in human intestinalmucosa. Arch. Biochem. Biophys. 293:293–304; 1992.
[19] Krinsky, N. I.; Wang, X.-D.; Tang, G.; Russell, R. M. Conversion ofcarotenoids to retinoids. New York: Dekker; 1993.
[20] Yeum, K.-J.; Lee-Kim, Y. C.; Yoon, S.; Lee, K. Y.; Park, I. S.; Lee, K. S.;Kim, B. S.; Tang, G.; Russell, R. M.; Krinsk, N. I. Similar metabolitesformed from β-carotene by either human gastric mucosal homogenates,lipoxygenase or linoleic acid hydroperoxide. Arch. Biochem. Biophys.321:167–174; 1995.
[21] Ferreira, A. L. A.; Yeum, K.-J.; Russell, R. M.; Tang, G.; Krinsky, N. I.Enzymatic oxidative metabolites of lycopene. J. Nutr. Biochem.14:531–540; 2003.
[22] Paiva, S. A.; Novo, R.; Matsubara, B. B.; Matsubara, L. S.; Azevedo, P. S.;Minicucci, M. F.; Campana, A. O.; Zornoff, L. A. Beta-carotene attenuatesthe paradoxical effect of tobacco smoke on the mortality of rats afterexperimental myocardial infarction. J. Nutr. 135:2109–2113; 2005.
[23] Abdel-aleem, S.; el-Merzabani, M. M.; Sayed-Ahmed, M.; Taylor, D. A.;Lowe, J. E. Acute and chronic effects of adriamycin on fatty acid oxidationin isolated cardiac myocytes. J. Mol. Cell. Cardiol. 29:789–797; 1997.
[24] Matsui, H.; Morishima, I.; Numaguchi, Y.; Toki, Y.; Okumura, K.;Hayakawa, T. Protective effects of carvedilol against doxorubicin-inducedcardiomyopathy in rats. Life Sci. 65:1265–1274; 1999.
[25] Nagao, A.; During, A.; Hoshino, C.; Terao, J.; Olson, J. A. Stoichiometricconversion of all trans-b-carotene to retinal by pig intestinal extract. Arch.Biochem. Biophys. 328:57–63; 1996.
[26] During, A.; Nagao, A.; Hoshino, C.; Terao, J. Assay of β-carotene 15,159-dioxygenase activity by reverse-phase high-pressure liquid chromatogra-phy. Anal. Biochem. 241:199–205; 1996.
[27] Ferreira, A. L. A.; Yeum, K.-J.; Liu, C.; Smith, D.; Krinsky, N. I.; Wang,X.-D.; Russell, R. M. Tissue distribution of lycopene in ferrets and ratsafter lycopene supplementation. J. Nutr. 130:1256–1260; 2000.
[28] Beretta, G.; Aldini, G.; Facino, R. M.; Russell, R. M.; Krinsky, N. I.;Yeum, K. J. Total antioxidant performance: a validated fluorescenceassay for the measurement of plasma oxidizability. Anal. Biochem. 354:290–298; 2006.
[29] Aldini, G.; Yeum, K.; Russell, R. M.; Krinsky, N. I. A method to measurethe oxidizability of both the aqueous and lipid compartments of plasma.Free Radic. Biol. Med. 31:1043–1050; 2001.
[30] Wang, X. D.; Tang, G. W.; Fox, J. G.; Krinsky, N. I.; Russell, R. M.Enzymatic conversion of beta-carotene into beta-apo-carotenals andretinoids by human, monkey, ferret, and rat tissues. Arch. Biochem.Biophys. 285:8–16; 1991.
[31] Kim, S. J.; Nara, E.; Kobayashi, H.; Terao, J.; Naga, A. Formation ofcleavage products by autoxidation of lycopene. Lipids 36:191–199; 2001.
[32] Manchand, P. S.; Ruegg, R.; Schwieter, U.; Siddons, P. T.; Weedon,B. C. L. Carotenoids and related compounds. XI. Syntheses of delta-carotene and epson-carotene. J. Chem. Soc. 46:2019–2026; 1965.
[33] Khachik, F.; Steck, A.; Niggli, U. A.; Pfander, H. Partial synthesis andstructural elucidation of the oxidative metabolites of lycopene identified intomato paste, tomato juice, and human serum. J. Agric. Food Chem.46:4874–4884; 1998.
[34] Paetau, I.; Khachik, F.; Brown, E. D.; Beecher, G. R.; Kramer, T. R.;Chittams, J.; Clevidence, B. A. Chronic ingestion of lycopene-rich tomatojuice or lycopene supplements significantly increases plasma concentra-tions of lycopene and related tomato carotenoids in humans. Am. J. Clin.Nutr. 68:1187–1195; 1998.
[35] Lefrak, E. A.; Pitha, J.; Rosenheim, S.; Gottieb, J. A. A clinicopathologicanalysis of adriamycin cardiotoxicity. Cancer 32:302–314; 1973.
[36] Hiroe, M.; Ohta, Y.; Fujita, N.; Nagata, M.; Toyozaki, T.; Kusakabe, K.;Sekiguchi, M.; Marumo, F. Myocardial uptake of 111In monoclonal
antimyosin Fab in detecting doxorubicin cardiotoxicity in rats. Morpho-logical and hemodynamic findings. Circulation 86:1965–1972; 1992.
[37] Deepa, P. R.; Varalakshmi, P. The cytoprotective role of a low-molecular-weight heparin fragment studied in an experimental model of glomer-ulotoxicity. Eur. J. Pharmacol. 478:199–205; 2003.
[38] Chen, D.; Zhang, X. Y.; Shi, Y. Identification and functional characteriza-tion of hCLS1, a human cardiolipin synthase localized in mitochondria.Biochem. J. 398:169–176; 2006.
[39] Goormaghtigh, E.; Ruysschaert, J. M. Anthracycline glycoside-/membraneinteractions. Biochim. Biophys. Acta 779:271–288; 1984.
[40] Davies, K. J. A.; Doroshow, J. H. Redox cycling of anthracyclines bycardiac mitochondria. I. Anthracycline radical formation by NADHdehydrogenase. J. Biol. Chem. 261:3060–3067; 1986.
[41] Nohl, H. Identification of the site of adriamycin-activation in the heart cell.Biochem. Pharmacol. 37:2633–2637; 1988.
[42] Nohl, H. Demonstration of the existence of an organo-specific NADHdehydrogenase in heart mitochondria. Eur. J. Biochem. 169:585–591;1987.
[43] Schönheit, K.; Nohl, H. Oxidation of cytosolic NADH via complex I ofheart mitochondria. Arch. Biochem. Biophys. 327:319–323; 1996.
[44] Gille, L.; Nohl, H. Analyses of the molecular mechanism of adriamycin-induced cardiotoxicity. Free Radic. Biol. Med. 23:775–782; 1997.
[45] Singal, P. K.; Siveski-Iliskovic, N.; Hill, M.; Thomas, T. P.; Li, T.Combination therapy with probucol prevents adriamycin-induced cardio-myopathy. J. Mol. Cell. Cardiol. 27:1055–1063; 1995.
[46] Kotamraju, S.; Konorev, E. A.; Joseph, J.; Kalyanaraman, B. Doxorubicin-induced apoptosis in endothelial cells and cardiomyocytes is amelioratedby nitrone spin traps and ebselen. Role of reactive oxygen and nitrogenspecies. J. Biol. Chem. 275:33585–33592; 2000.
[47] Van Vleet, J. F.; Ferrans, V. J.; Weirich, W. E. Cardiac disease induced bychronic adriamycin administration in dogs and an evaluation of vitamin Eand selenium as cardioprotectants. Am. J. Pathol. 99:13–42; 1980.
[48] Breed, J. G.; Zimmerman, A. N.; Dormans, J. A.; Pinedo, H. M. Failure ofthe antioxidant vitamin E to protect against adriamycin-induced cardio-toxicity in the rabbit. Cancer Res. 40:2033–2038; 1980.
[49] Legha, S. S.; Wang, Y. M.; Mackay, B.; Ewer, M.; Hortobagyi, G. N.;Benjamin, R. S.; Ali, M. K. Clinical and pharmacologic investigation ofthe effects of alpha-tocopherol on adriamycin cardiotoxicity. Ann. N. Y.Acad. Sci. 393:411–418; 1982.
[50] Myers, C.; Bonow, R.; Palmeri, S.; Jenkins, J.; Corden, B.; Locker, G.;Doroshow, J.; Epstein, S. A randomized controlled trial assessing theprevention of doxorubicin cardiomyopathy by N-acetylcysteine. Semin.Oncol. 10:53–55; 1983.
[51] Olson, R. D.; Mushlin, P. S.; Brenner, D. E.; Fleischer, S.; Cusack, B. J.;Chang, B. K.; Boucek, R. J. Doxorubicin cardiotoxicity may be caused byits metabolite, doxorubicinol. Proc. Natl. Acad. Sci. USA 85:3585–3589;1988.
[52] Minotti, G.; Cavaliere, A. F.; Mordente, A.; Rossi, M.; Schiavello, R.;Zamparelli, R.; Possati, G. Secondary alcohol metabolites mediate irondelocalization in cytosolic fractions of myocardial biopsies exposed toanticancer anthracyclines: novel linkage between anthracycline metabo-lism and iron-induced cardiotoxicity. J. Clin. Invest. 95:1595–1605; 1995.
[53] Dodd, D. A.; Atkinson, J. B.; Olson, R. D.; Buck, S.; Cusack, B. J.;Fleischer, S.; Boucek, R. J. J. Doxorubicin cardiomyopathy is associatedwith a decrease in calcium release channel of the sarcoplasmic reticulum ina chronic rabbit model. J. Clin. Invest. 91:1697–1705; 1993.
[54] Zhou, S.; Starkov, A.; Froberg, M. K.; Leino, R. L.; Wallace, K. B.Cumulative and irreversible cardiac mitochondrial dysfunction induced bydoxorubicin. Cancer Res. 61:771–777; 2001.
[55] Siveski-Iliskovic, N.; Singal, P. K. Lipid lowering: an important factorin preventing adriamycin- induced heart failure. Am. J. Pathol. 150:727–734; 1997.
[56] Hong, Y. M.; Kim, H. S.; Yoon, H.-R. Serum lipid and fatty acid profiles inadriamycin-treated rats after administration of L-carnitine. Pediatr. Res.51:249–255; 2002.
[57] Kitagawa, K.; Takeda, K.; Saito, K.; Okamoto, S.; Makino, K.; Maeda, H.;Ichihara, T. Differences in fatty acid metabolic disorder between ischemicmyocardium and doxorubicin-induced myocardial damage: assessment

751A.L.A. Ferreira et al. / Free Radical Biology & Medicine 43 (2007) 740–751
using BMIPP dynamic SPECT with analysis by the Rutland method.J. Nucl. Med. 43:1286–1294; 2002.
[58] Lü, H. Z.; Geng, B. Q.; Zhu, Y. L.; Yong, D. G. Effects of beta-carotene ondoxorubicin-induced cardiotoxicity in rats. Acta Pharmacol. Sin.17:317–320; 1996.
[59] Ciaccio, M.; Valenza, M.; Tesoriere, L.; Bongiorno, A.; Albiero, R.;Livrea, M. A. Vitamin-A inhibits doxorubicin-induced membrane lipid-peroxidation in rat-tissues in vivo. Arch. Biochem. Biophys. 302:103–108;1993.
[60] Yin, X.; Wu, H.; Chen, Y.; Kang, Y. J. Induction of antioxidants byadriamycin in mouse heart. Biochem. Pharmacol. 56:87–93; 1998.
[61] Dziegiel, P.; Murawska-Cialowicz, E.; Jethon, Z.; Januszewska, L.;Podhorska-Okolow, M.; Surowiak, P.; Zawadzki, M.; Rabczynski, J.;Zabel, M. Melatonin stimulates the activity of protective antioxidativeenzymes in myocardial cells of rats in the course of doxorubicinintoxication. J. Pineal Res. 35:183–187; 2003.
[62] Minotti, G.; Mancuso, C.; Frustaci, A.; Mordente, A.; Santini, S. A.;Calafiore, A. M.; Liberi, G.; Gentiloni, N. Paradoxical inhibition of cardiaclipid peroxidation in cancer patients treated with doxorubicin. Pharmaco-logic and molecular reappraisal of anthracycline cardiotoxicity. J. Clin.Invest. 98:650–661; 1996.
[63] Erhola, M.; Kellokumpu-Lehtinen, P.; Metsa-Ketela, T.; Alanko, K.;Nieminen, M. M. Effects of anthracyclin-based chemotherapy on total
plasma antioxidant capacity in small cell lung cancer patients. Free Radic.Biol. Med. 21:383–390; 1996.
[64] Faure, H.; Coudray, C.; Mousseau, M.; Ducros, V.; Douki, T.; Bianchini,F.; Cadet, J.; Favier, A. 5-Hydroxymethyluracil excretion, plasma TBARSand plasma antioxidant vitamins in adriamycin-treated patients. FreeRadic. Biol. Med. 20:979–983; 1996.
[65] Iwase, H.; Sakurada, K.; Takatori, T.; Nagao, M.; Niijima, H.; Matsuda, Y.;Kobayashi, M. Calcium ions potentiate lipoxygenase activity ofcytochrome c at the physiological pH. Biochem. Biophys. Res. Commun.243:485–491; 1998.
[66] Hammarberg, T.; Provost, P.; Persson, B.; Radmark, O. The N-terminaldomain of 5-lipoxygenase binds calcium and mediates calciumstimulation of enzyme activity. J. Biol. Chem. 275:38787–38793;2000.
[67] Ferreira, A. L. A.; Russell, R. M.; Rocha, N. S.; Ladeira, M. S. P.;Salvadori, D. M. F.; Nascimento, M. C. M. O.; Matsui, M.; Carvalho, F. A.;Tang, G.; Matsubara, L. S.; Matsubara, B. B. Effect of Lycopene ondoxorubicin-induced cardiotoxicity: an echocardiographic, histological,and morphometrical assessment. Basic Clin. Pharmacol. Toxicol.101:16–24; 2007.
[68] Billingham, M. E.; Mason, J. W.; Bristow, M. R.; Daniels, J. R. Anthra-cycline cardiomyopathy monitored by morphologic changes. CancerTreat. Rep. 62:865–872; 1978.
Related Documents











