This article was downloaded by: [Professor Gianluca Polese] On: 06 November 2014, At: 13:34 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Aquatic Animal Health Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/uahh20 Dose-Dependent Effects of the Clinical Anesthetic Isoflurane on Octopus vulgaris: A Contribution to Cephalopod Welfare Gianluca Polese a , William Winlow b & Anna Di Cosmo a a Department of Biology, University of Napoli Federico II, Via Cinthia, Building 7, Campus MSA, 80126 Napoli, Italy b University of Liverpool, Liverpool, Merseyside L69 3BX, UK; and University of Central Lancashire, Fylde Road, Preston, Lancashire PR1 2HE, UK Published online: 04 Nov 2014. To cite this article: Gianluca Polese, William Winlow & Anna Di Cosmo (2014) Dose-Dependent Effects of the Clinical Anesthetic Isoflurane on Octopus vulgaris: A Contribution to Cephalopod Welfare, Journal of Aquatic Animal Health, 26:4, 285-294, DOI: 10.1080/08997659.2014.945047 To link to this article: http://dx.doi.org/10.1080/08997659.2014.945047 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [Professor Gianluca Polese]On: 06 November 2014, At: 13:34Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Aquatic Animal HealthPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/uahh20
Dose-Dependent Effects of the Clinical AnestheticIsoflurane on Octopus vulgaris: A Contribution toCephalopod WelfareGianluca Polesea, William Winlowb & Anna Di Cosmoa
a Department of Biology, University of Napoli Federico II, Via Cinthia, Building 7, CampusMSA, 80126 Napoli, Italyb University of Liverpool, Liverpool, Merseyside L69 3BX, UK; and University of CentralLancashire, Fylde Road, Preston, Lancashire PR1 2HE, UKPublished online: 04 Nov 2014.
To cite this article: Gianluca Polese, William Winlow & Anna Di Cosmo (2014) Dose-Dependent Effects of the ClinicalAnesthetic Isoflurane on Octopus vulgaris: A Contribution to Cephalopod Welfare, Journal of Aquatic Animal Health, 26:4,285-294, DOI: 10.1080/08997659.2014.945047
To link to this article: http://dx.doi.org/10.1080/08997659.2014.945047
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Aquatic Animal Health 26:285–294, 2014C© American Fisheries Society 2014ISSN: 0899-7659 print / 1548-8667 onlineDOI: 10.1080/08997659.2014.945047
ARTICLE
Dose-Dependent Effects of the Clinical AnestheticIsoflurane on Octopus vulgaris: A Contribution toCephalopod Welfare
Gianluca PoleseDepartment of Biology, University of Napoli Federico II, Via Cinthia, Building 7, Campus MSA,80126 Napoli, Italy
William WinlowUniversity of Liverpool, Liverpool, Merseyside L69 3BX, UK; and University of Central Lancashire,Fylde Road, Preston, Lancashire PR1 2HE, UK
Anna Di Cosmo*Department of Biology, University of Napoli Federico II, Via Cinthia, Building 7, Campus MSA,80126 Napoli, Italy
AbstractRecent progress in animal welfare legislation relating to invertebrates has provoked interest in methods for the
anesthesia of cephalopods, for which different approaches to anesthesia have been tried but in most cases withouttruly anesthetizing the animals. For example, several workers have used muscle relaxants or hypothermia as formsof “anesthesia.” Several inhalational anesthetics are known to act in a dose-dependent manner on the great pondsnail Lymnaea stagnalis, a pulmonate mollusk. Here we report, for the first time, on the effects of clinical doses of thewell-known inhalational clinical anesthetic isoflurane on the behavioral responses of the common octopus Octopusvulgaris. In each experiment, isoflurane was equilibrated into a well-aerated seawater bath containing a single adultO. vulgaris. Using a web camera, we recorded each animal’s response to touch stimuli eliciting withdrawal of thearms and siphon and observed changes in the respiratory rate and the chromatophore pattern over time (before,during, and after application of the anesthetic). We found that different animals of the same size responded withsimilar behavioral changes as the isoflurane concentration was gradually increased. After gradual application of 2%isoflurane for a maximum of 5 min (at which time all the responses indicated deep anesthesia), the animals recoveredwithin 45–60 min in fresh aerated seawater. Based on previous findings in gastropods, we believe that the processof anesthesia induced by isoflurane is similar to that previously observed in Lymnaea. In this study we showed thatisoflurane is a good, reversible anesthetic for O. vulgaris, and we developed a method for its use.
Anesthetics have long been used to diminish pain and suffer-ing in human beings, and the assumption is made that they havesimilar actions on other vertebrate animals. Although these ani-mals are not able to communicate directly with the investigators,assumptions about their level of anesthesia are based on theirbehavioral responses. In vertebrates, anesthetics have been usedto abolish the perception of pain, either to allow humane han-
*Corresponding author: [email protected] February 14, 2014; accepted July 7, 2014
dling of an animal or to allow surgical manipulations. However,pain in invertebrates is as yet a largely unexplored field to whichexperimental biologists have paid little attention during exper-imental procedures. Therefore, the procedures for invertebrateanesthesia, analgesia, and euthanasia that are available at themoment are more focused on the scientists’ experimental needsthan on the effective reduction of pain and suffering and show
285
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14

286 POLESE ET AL.
little regard for the animal’s well-being (Cooper 2011; Mather2011). However, the recent growth of interest in invertebratewelfare (Moltschaniwskyj et al. 2007; Elwood 2011; De Lisaet al. 2012; Andrews et al. 2013; Horvath et al. 2013; Magee andElwood 2013) necessitates the use of agents such as isofluranewith known anesthetic properties (Eger 1981) during invasiveprocedures, along with the establishment protocols for their usethat are suitable for individual species.
Among marine invertebrates the most developed and intel-ligent animals are probably cephalopod mollusks and decapodcrustaceans (Advocates for Animals 2005). Cephalopods dis-play advanced cephalization of the ganglionic masses, whichare associated with hierarchical organization of function andthe development of advanced cognitive capabilities. Based onthese capabilities and structures as well as their complexity,individuality (Mather and Anderson 1993), adaptability, andability to learn, it should be assumed that cephalopods feel pain.Although they respond to noxious stimuli (Crook and Walters2011), their central neural circuits for pain perception are un-known at present apart from Gray’s suggestion (Gray 1970) thattheir pain center is located in the subvertical lobe.
Recently introduced legislation stresses the necessity of treat-ing cephalopods humanely. This has resulted from changes tolaws for the protection of animals in the United Kingdom in1993 (when the Animals [Scientific Procedures] Act of 1986was amended to include Octopus vulgaris; UK Statutory Instru-ments 1993), and in Australia (Sharman 2004) and the EuropeanUnion (European Parliament 2010) to cover all live cephalopodmollusks. However, there is no protection of cephalopods underU.S. law (Latham 2012).
Previous attempts to anesthetize cephalopods have used avariety of substances, including muscle relaxants such as mag-nesium chloride (Messenger et al. 1985; Mooney et al. 2010;Goncalves et al. 2012) and low temperatures (Andrews andTansey 1981), resulting in paralysis of the animals rather thananesthesia. A recent study by Goncalves et al. (2012) comparesseveral different agents commonly used to immobilize commoncuttlefish Sepia officinalis for short-term handling and indicatesthat hypothermia causes severe stress reactions during the re-covery phase; the authors recommend MgCl2 as an appropriate“anesthetic” agent because it causes little stress to the animalunder these conditions. However, MgCl2 is not an anestheticagent, and neither of these treatments is acceptable as a formof anesthesia because the loss of consciousness cannot be as-sumed in the absence of movement without detailed analysisof the behavioral signs of anesthesia. Magnesium salts such asMgSO4 may be used as adjuncts to anesthetics and have beenfound to decrease the time to onset of neuromuscular blockageinduced by clinical blocking agents such as rocuronium (Ro-tava et al. 2013). Additionally, cephalopods are relatively largeanimals and it is unlikely that immersion in magnesium chlo-ride would have rapid effects on the central nervous system asasserted by Messenger et al. (1985). Furthermore, the octopisubjected to MgCl2 treatment only survived for 2 d posttreat-
FIGURE 1. Anesthetic delivery system consisting of an electric pump (A), aflowmeter (B), and a specific (isoflurane) vaporizer (C) terminating in an airstone (D) in a tank (E) filled with 1.6 L of seawater. The ambient temperatureof the water was 18◦C. A recording camera (F) was placed just over the tank torecord the movements and color changes of the animal. The whole system wasplaced in an enclosure (H) and air was expelled to the outside using an extractorfan (G).
ment. Other muscle relaxants used with cephalopods includegallamine (Flaxedil) and tricaine methanesulfonate (or mesy-late [MS-222]) (Mooney et al. 2010). It should be noted thatalthough MS-222 is used as an anesthetic and sedative for fish(Palmer and Mensinger 2004), it has only been shown to actlike a local anesthetic (Frazier and Narahashi 1975) or a mus-cle relaxant in cephalopods. While muscle relaxants may havea role in the induction of anesthesia, none of these substanceshave anesthetic actions in themselves. The application of gal-lamine to the longfin inshore squid Doryteuthis peleii resultedin the death of the animals, as did that of the local anestheticbenzocaine and the analgesic clove oil (Mooney et al. 2010).Other substances have also proved problematical; solutions ofthe intravenous anesthetic urethane (ethyl carbamate) were usedto anesthetize cephalopods in the past (Messenger 1968; Young1971). However, urethane fell into disfavor when it was foundto be carcinogenic and endangered laboratory personnel, thoughit could still be used with appropriate precautions as an anes-thetic agent for laboratory animals when procedures that did notentail recovery were to be carried out (Field and Lang 1988).Ethanol has also been tried as a cephalopod anesthetic, but ittends to cause the escape behaviors of jetting and inking oninitial immersion of the animal (Andrews and Tansey 1981)—aphenomenon that is not always reported (Froesch and Marthy1975; Harms et al. 2006). Mooney et al. (2010) describedsimilar phenomena when D. peleii were placed in ethanol,along with dramatic changes in color not normally seen in theanimal.
Ideally, an anesthetic would be a good muscle relaxant, haveboth analgesic and anesthetic properties, induce unconscious-ness, and have no side effects. This ideal is difficult to achieve,which is why muscle relaxants are used in the induction phaseof clinical or veterinary anesthesia to minimize the voluntaryand involuntary excitation and struggling that would otherwise
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
EFFECTS OF ISOFLURANE ON OCTOPUS VULGARIS 287
be observed. These behaviors result in an increase in both res-piratory rate and heart rate during induction.
Inhalational anesthetics are widely used clinically, along witha wide variety of adjunctive agents, because their physiologicaleffects are well known and relatively easy to monitor and reverse(Ebert 2006). Inhalational anesthetics are delivered in a carrierstream of air or oxygen using an anesthetic vaporizer to ensurethat the subject remains adequately oxygenated throughout theprocedure. Among mollusks, the gastropod Lymnaea stagnalishas proved to be a useful model for the study of anesthetics(Girdlestone et al. 1989; Winlow et al. 1995, 1998). The clini-cal inhalational anesthetics halothane, enflurane, and isofluranehave all been used to induce anesthesia in Lymnaea in a dose-dependent manner, reducing L-type calcium (Yar and Winlow1993) currents and potassium currents (Moghadam and Winlow1995). There is evidence from cell culture that such anesthet-ics block excitatory chemical synapses more effectively thaninhibitory synapses (Spencer et al. 1996). In addition, duringthe anesthetization of Lymnaea an early excitatory phase is fol-lowed by suppression (McCrohan et al. 1987), similar to thephases of anesthesia observed in humans under anesthesia by
ether (Guedel 1937) but not normally observable under cur-rent practices due to the use of muscle relaxants. We chose theinhalational anesthetic isoflurane to develop an appropriate pro-tocol for anesthetization of O. vulgaris because it is very stable(Ebert 2006), reliable, easily available, relatively inexpensive,and “can produce adequate muscle relaxation for any surgicalprocedure” (Eger 1981). Here we describe for the first time amethod for the successful use of isoflurane as an anesthetic forcephalopods.
METHODSAnimals.—Sixteen specimens of Octopus vulgaris (body
weight, 0.38–0.43 kg), of which eight were male and eightfemale, were captured in the Bay of Naples and bought fromthe local fish market, transported to our animal facility, and keptin captivity following the protocol of our ethical committee(2012/0124283-08/11/2012). The test specimens were fed freshcrabs (Carcinus aestuarii) for 1 week before the experiments.Octopi that fully recovered from the experimental procedureswere kept in the Department of Biology’s animal facility andlater employed in other experiments.
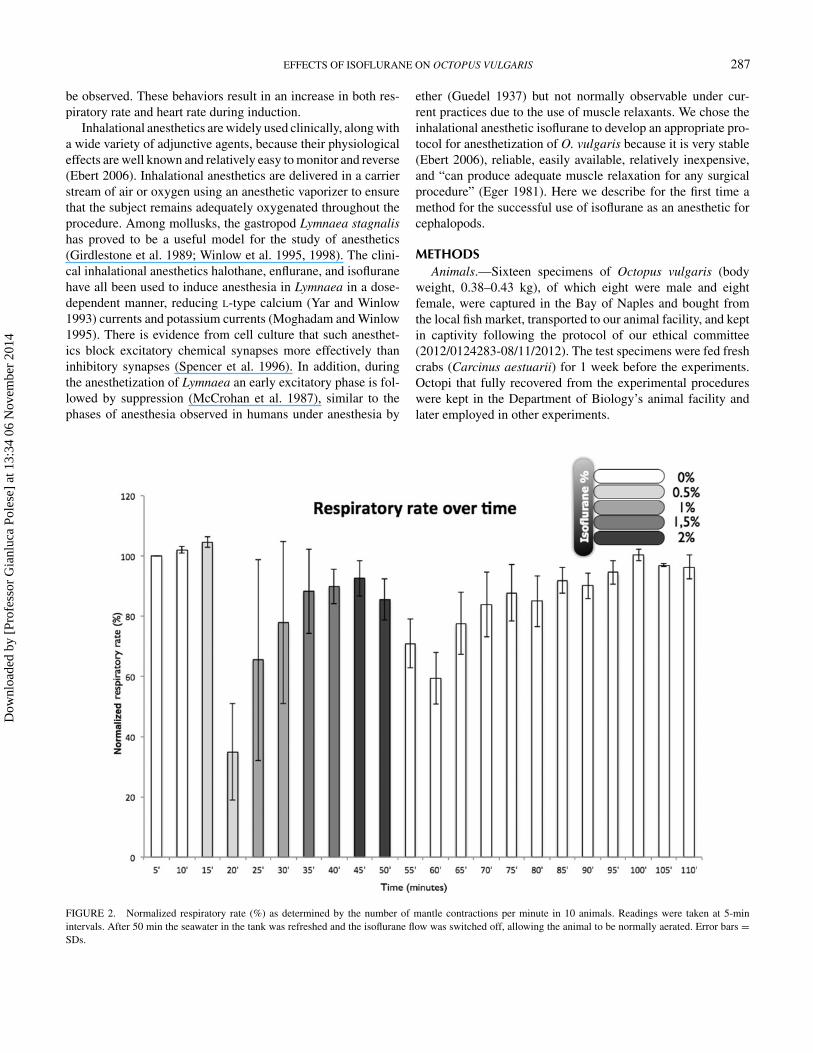
FIGURE 2. Normalized respiratory rate (%) as determined by the number of mantle contractions per minute in 10 animals. Readings were taken at 5-minintervals. After 50 min the seawater in the tank was refreshed and the isoflurane flow was switched off, allowing the animal to be normally aerated. Error bars =SDs.
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
288 POLESE ET AL.
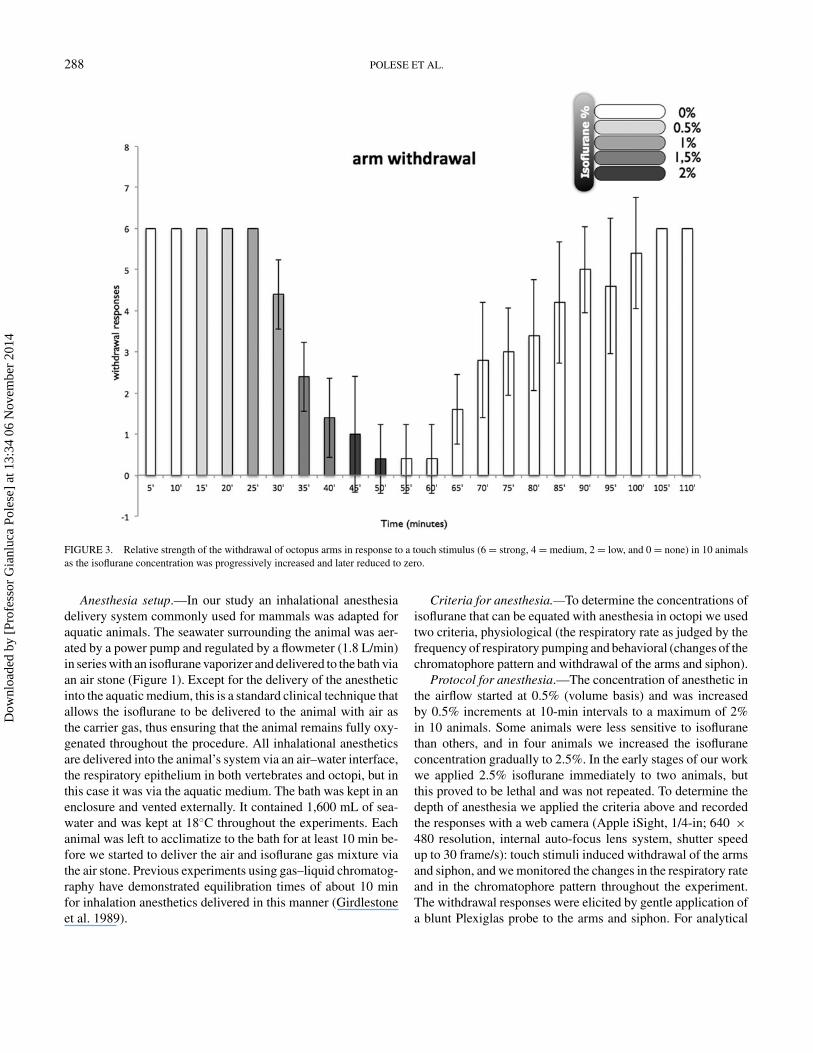
FIGURE 3. Relative strength of the withdrawal of octopus arms in response to a touch stimulus (6 = strong, 4 = medium, 2 = low, and 0 = none) in 10 animalsas the isoflurane concentration was progressively increased and later reduced to zero.
Anesthesia setup.—In our study an inhalational anesthesiadelivery system commonly used for mammals was adapted foraquatic animals. The seawater surrounding the animal was aer-ated by a power pump and regulated by a flowmeter (1.8 L/min)in series with an isoflurane vaporizer and delivered to the bath viaan air stone (Figure 1). Except for the delivery of the anestheticinto the aquatic medium, this is a standard clinical technique thatallows the isoflurane to be delivered to the animal with air asthe carrier gas, thus ensuring that the animal remains fully oxy-genated throughout the procedure. All inhalational anestheticsare delivered into the animal’s system via an air–water interface,the respiratory epithelium in both vertebrates and octopi, but inthis case it was via the aquatic medium. The bath was kept in anenclosure and vented externally. It contained 1,600 mL of sea-water and was kept at 18◦C throughout the experiments. Eachanimal was left to acclimatize to the bath for at least 10 min be-fore we started to deliver the air and isoflurane gas mixture viathe air stone. Previous experiments using gas–liquid chromatog-raphy have demonstrated equilibration times of about 10 minfor inhalation anesthetics delivered in this manner (Girdlestoneet al. 1989).
Criteria for anesthesia.—To determine the concentrations ofisoflurane that can be equated with anesthesia in octopi we usedtwo criteria, physiological (the respiratory rate as judged by thefrequency of respiratory pumping and behavioral (changes of thechromatophore pattern and withdrawal of the arms and siphon).
Protocol for anesthesia.—The concentration of anesthetic inthe airflow started at 0.5% (volume basis) and was increasedby 0.5% increments at 10-min intervals to a maximum of 2%in 10 animals. Some animals were less sensitive to isofluranethan others, and in four animals we increased the isofluraneconcentration gradually to 2.5%. In the early stages of our workwe applied 2.5% isoflurane immediately to two animals, butthis proved to be lethal and was not repeated. To determine thedepth of anesthesia we applied the criteria above and recordedthe responses with a web camera (Apple iSight, 1/4-in; 640 ×480 resolution, internal auto-focus lens system, shutter speedup to 30 frame/s): touch stimuli induced withdrawal of the armsand siphon, and we monitored the changes in the respiratory rateand in the chromatophore pattern throughout the experiment.The withdrawal responses were elicited by gentle application ofa blunt Plexiglas probe to the arms and siphon. For analytical
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
EFFECTS OF ISOFLURANE ON OCTOPUS VULGARIS 289
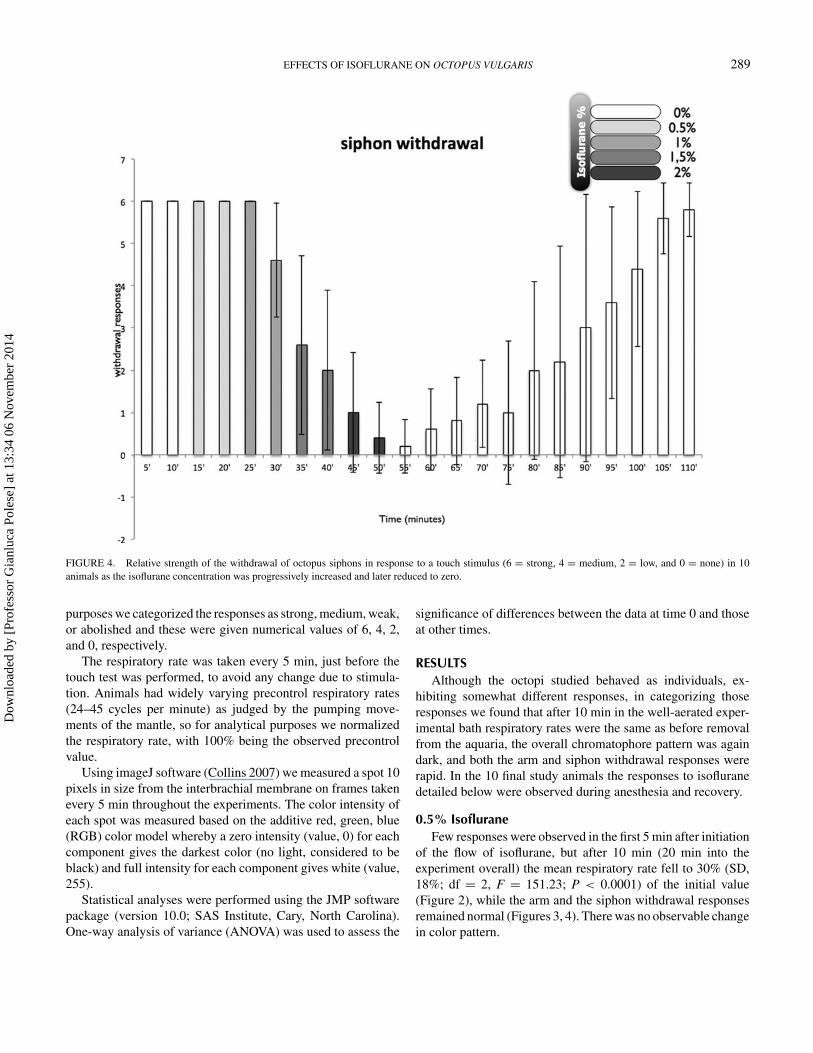
FIGURE 4. Relative strength of the withdrawal of octopus siphons in response to a touch stimulus (6 = strong, 4 = medium, 2 = low, and 0 = none) in 10animals as the isoflurane concentration was progressively increased and later reduced to zero.
purposes we categorized the responses as strong, medium, weak,or abolished and these were given numerical values of 6, 4, 2,and 0, respectively.
The respiratory rate was taken every 5 min, just before thetouch test was performed, to avoid any change due to stimula-tion. Animals had widely varying precontrol respiratory rates(24–45 cycles per minute) as judged by the pumping move-ments of the mantle, so for analytical purposes we normalizedthe respiratory rate, with 100% being the observed precontrolvalue.
Using imageJ software (Collins 2007) we measured a spot 10pixels in size from the interbrachial membrane on frames takenevery 5 min throughout the experiments. The color intensity ofeach spot was measured based on the additive red, green, blue(RGB) color model whereby a zero intensity (value, 0) for eachcomponent gives the darkest color (no light, considered to beblack) and full intensity for each component gives white (value,255).
Statistical analyses were performed using the JMP softwarepackage (version 10.0; SAS Institute, Cary, North Carolina).One-way analysis of variance (ANOVA) was used to assess the
significance of differences between the data at time 0 and thoseat other times.
RESULTSAlthough the octopi studied behaved as individuals, ex-
hibiting somewhat different responses, in categorizing thoseresponses we found that after 10 min in the well-aerated exper-imental bath respiratory rates were the same as before removalfrom the aquaria, the overall chromatophore pattern was againdark, and both the arm and siphon withdrawal responses wererapid. In the 10 final study animals the responses to isofluranedetailed below were observed during anesthesia and recovery.
0.5% IsofluraneFew responses were observed in the first 5 min after initiation
of the flow of isoflurane, but after 10 min (20 min into theexperiment overall) the mean respiratory rate fell to 30% (SD,18%; df = 2, F = 151.23; P < 0.0001) of the initial value(Figure 2), while the arm and the siphon withdrawal responsesremained normal (Figures 3, 4). There was no observable changein color pattern.
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
290 POLESE ET AL.
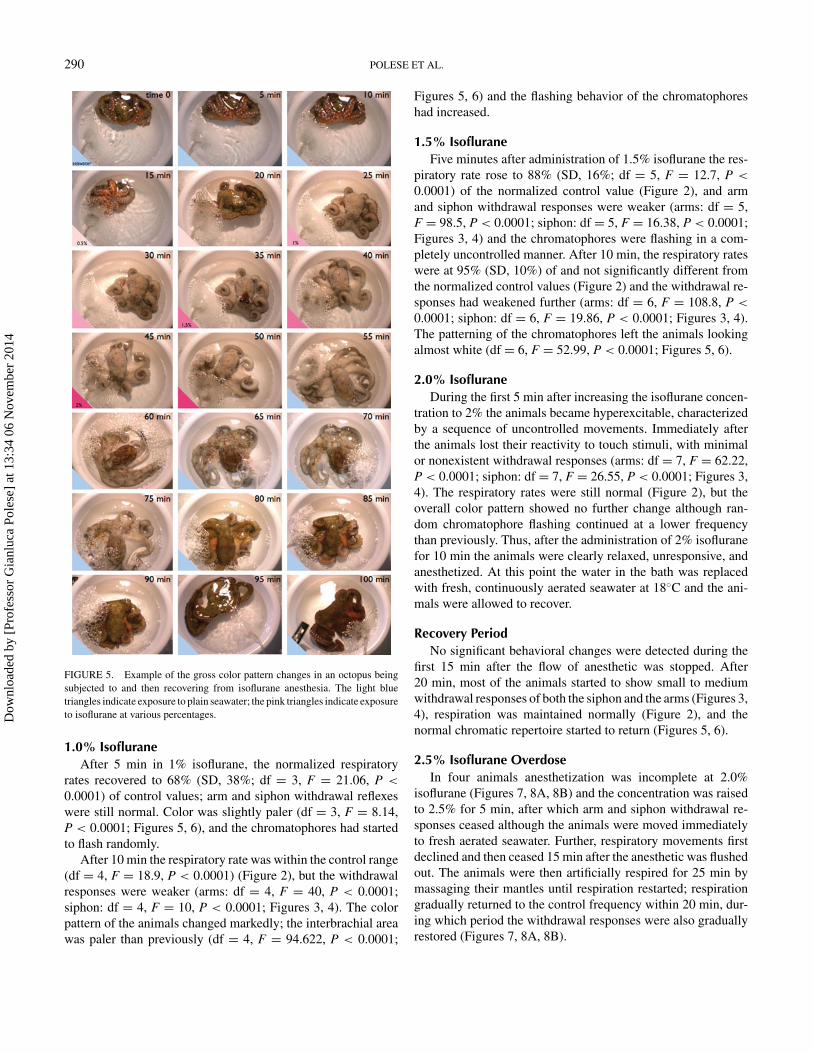
FIGURE 5. Example of the gross color pattern changes in an octopus beingsubjected to and then recovering from isoflurane anesthesia. The light bluetriangles indicate exposure to plain seawater; the pink triangles indicate exposureto isoflurane at various percentages.
1.0% IsofluraneAfter 5 min in 1% isoflurane, the normalized respiratory
rates recovered to 68% (SD, 38%; df = 3, F = 21.06, P <
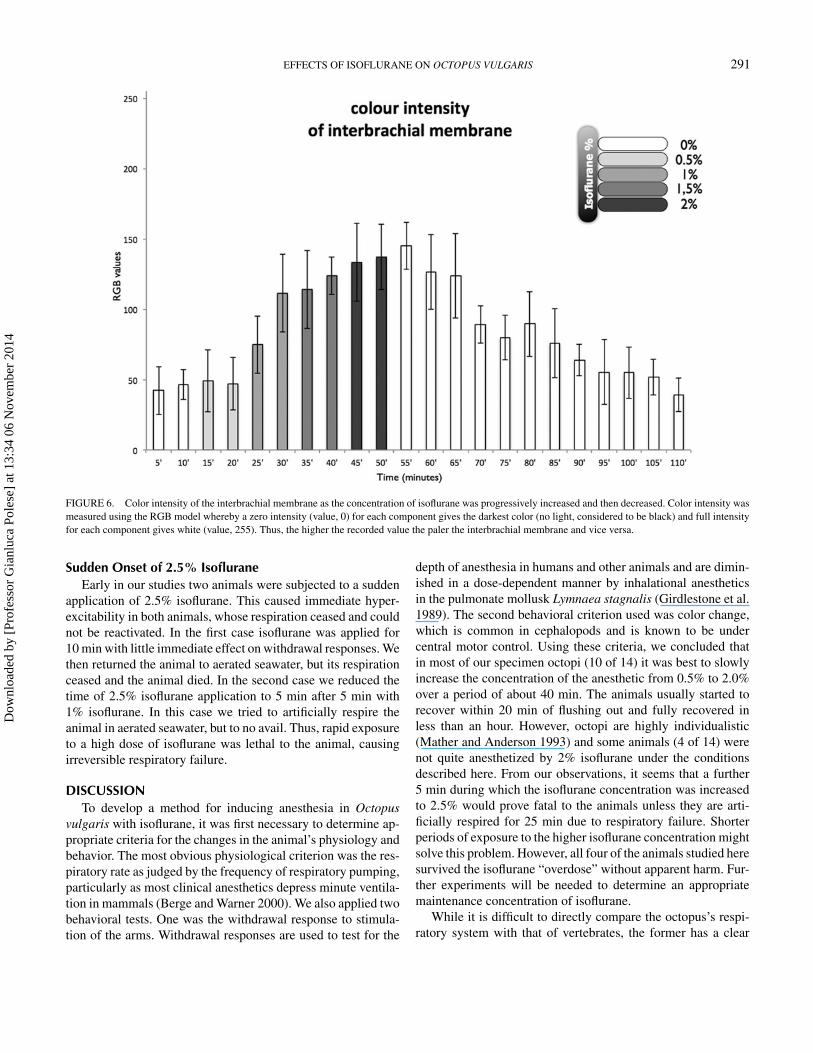
0.0001) of control values; arm and siphon withdrawal reflexeswere still normal. Color was slightly paler (df = 3, F = 8.14,P < 0.0001; Figures 5, 6), and the chromatophores had startedto flash randomly.
After 10 min the respiratory rate was within the control range(df = 4, F = 18.9, P < 0.0001) (Figure 2), but the withdrawalresponses were weaker (arms: df = 4, F = 40, P < 0.0001;siphon: df = 4, F = 10, P < 0.0001; Figures 3, 4). The colorpattern of the animals changed markedly; the interbrachial areawas paler than previously (df = 4, F = 94.622, P < 0.0001;
Figures 5, 6) and the flashing behavior of the chromatophoreshad increased.
1.5% IsofluraneFive minutes after administration of 1.5% isoflurane the res-
piratory rate rose to 88% (SD, 16%; df = 5, F = 12.7, P <
0.0001) of the normalized control value (Figure 2), and armand siphon withdrawal responses were weaker (arms: df = 5,F = 98.5, P < 0.0001; siphon: df = 5, F = 16.38, P < 0.0001;Figures 3, 4) and the chromatophores were flashing in a com-pletely uncontrolled manner. After 10 min, the respiratory rateswere at 95% (SD, 10%) of and not significantly different fromthe normalized control values (Figure 2) and the withdrawal re-sponses had weakened further (arms: df = 6, F = 108.8, P <
0.0001; siphon: df = 6, F = 19.86, P < 0.0001; Figures 3, 4).The patterning of the chromatophores left the animals lookingalmost white (df = 6, F = 52.99, P < 0.0001; Figures 5, 6).
2.0% IsofluraneDuring the first 5 min after increasing the isoflurane concen-
tration to 2% the animals became hyperexcitable, characterizedby a sequence of uncontrolled movements. Immediately afterthe animals lost their reactivity to touch stimuli, with minimalor nonexistent withdrawal responses (arms: df = 7, F = 62.22,P < 0.0001; siphon: df = 7, F = 26.55, P < 0.0001; Figures 3,4). The respiratory rates were still normal (Figure 2), but theoverall color pattern showed no further change although ran-dom chromatophore flashing continued at a lower frequencythan previously. Thus, after the administration of 2% isofluranefor 10 min the animals were clearly relaxed, unresponsive, andanesthetized. At this point the water in the bath was replacedwith fresh, continuously aerated seawater at 18◦C and the ani-mals were allowed to recover.
Recovery PeriodNo significant behavioral changes were detected during the
first 15 min after the flow of anesthetic was stopped. After20 min, most of the animals started to show small to mediumwithdrawal responses of both the siphon and the arms (Figures 3,4), respiration was maintained normally (Figure 2), and thenormal chromatic repertoire started to return (Figures 5, 6).
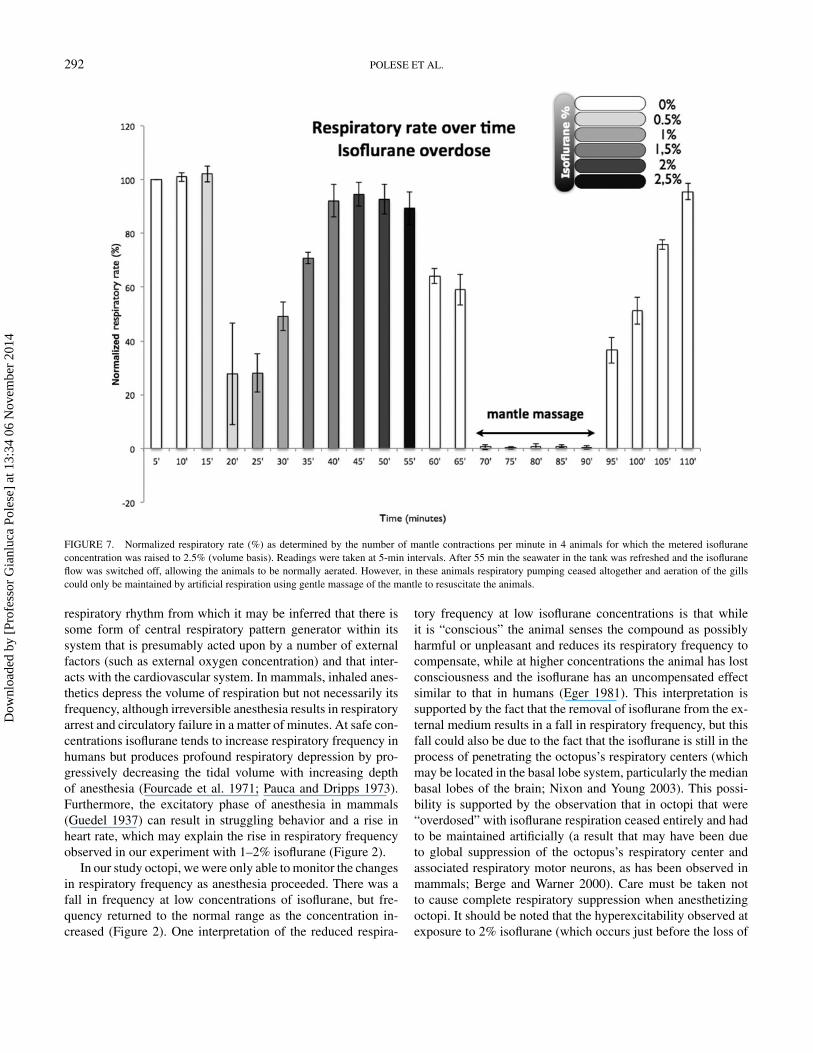
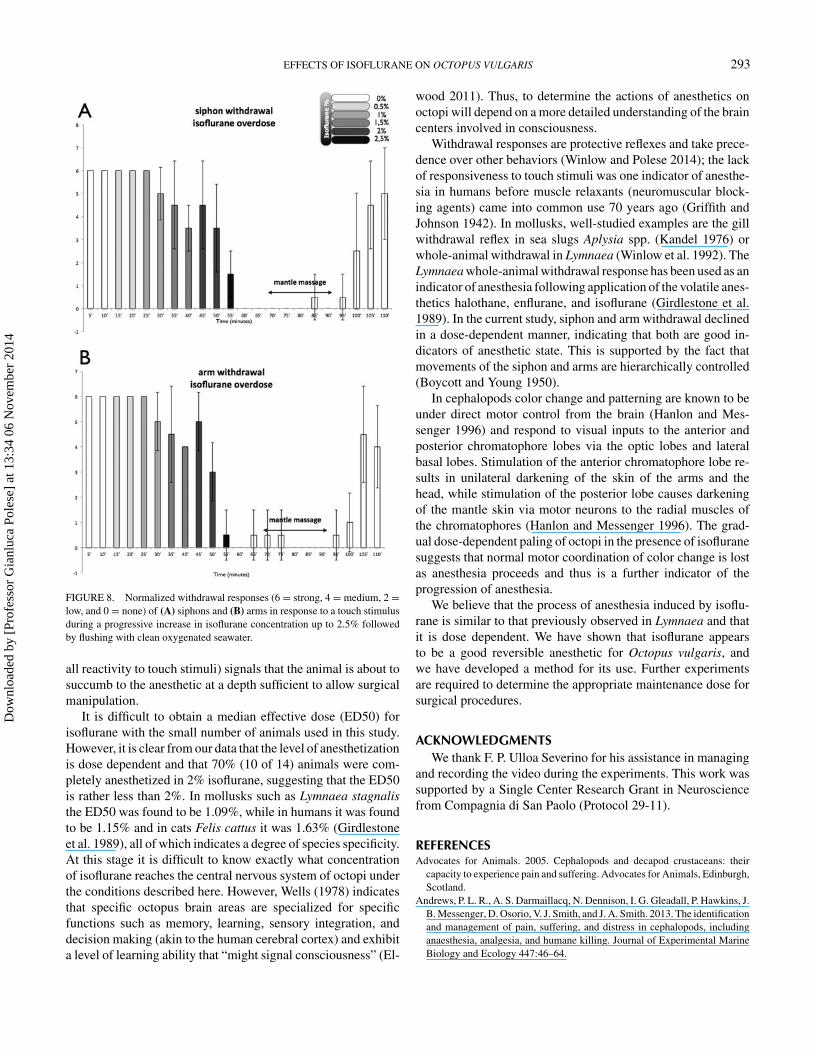
2.5% Isoflurane OverdoseIn four animals anesthetization was incomplete at 2.0%
isoflurane (Figures 7, 8A, 8B) and the concentration was raisedto 2.5% for 5 min, after which arm and siphon withdrawal re-sponses ceased although the animals were moved immediatelyto fresh aerated seawater. Further, respiratory movements firstdeclined and then ceased 15 min after the anesthetic was flushedout. The animals were then artificially respired for 25 min bymassaging their mantles until respiration restarted; respirationgradually returned to the control frequency within 20 min, dur-ing which period the withdrawal responses were also graduallyrestored (Figures 7, 8A, 8B).
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
EFFECTS OF ISOFLURANE ON OCTOPUS VULGARIS 291
FIGURE 6. Color intensity of the interbrachial membrane as the concentration of isoflurane was progressively increased and then decreased. Color intensity wasmeasured using the RGB model whereby a zero intensity (value, 0) for each component gives the darkest color (no light, considered to be black) and full intensityfor each component gives white (value, 255). Thus, the higher the recorded value the paler the interbrachial membrane and vice versa.
Sudden Onset of 2.5% IsofluraneEarly in our studies two animals were subjected to a sudden
application of 2.5% isoflurane. This caused immediate hyper-excitability in both animals, whose respiration ceased and couldnot be reactivated. In the first case isoflurane was applied for10 min with little immediate effect on withdrawal responses. Wethen returned the animal to aerated seawater, but its respirationceased and the animal died. In the second case we reduced thetime of 2.5% isoflurane application to 5 min after 5 min with1% isoflurane. In this case we tried to artificially respire theanimal in aerated seawater, but to no avail. Thus, rapid exposureto a high dose of isoflurane was lethal to the animal, causingirreversible respiratory failure.
DISCUSSIONTo develop a method for inducing anesthesia in Octopus
vulgaris with isoflurane, it was first necessary to determine ap-propriate criteria for the changes in the animal’s physiology andbehavior. The most obvious physiological criterion was the res-piratory rate as judged by the frequency of respiratory pumping,particularly as most clinical anesthetics depress minute ventila-tion in mammals (Berge and Warner 2000). We also applied twobehavioral tests. One was the withdrawal response to stimula-tion of the arms. Withdrawal responses are used to test for the
depth of anesthesia in humans and other animals and are dimin-ished in a dose-dependent manner by inhalational anestheticsin the pulmonate mollusk Lymnaea stagnalis (Girdlestone et al.1989). The second behavioral criterion used was color change,which is common in cephalopods and is known to be undercentral motor control. Using these criteria, we concluded thatin most of our specimen octopi (10 of 14) it was best to slowlyincrease the concentration of the anesthetic from 0.5% to 2.0%over a period of about 40 min. The animals usually started torecover within 20 min of flushing out and fully recovered inless than an hour. However, octopi are highly individualistic(Mather and Anderson 1993) and some animals (4 of 14) werenot quite anesthetized by 2% isoflurane under the conditionsdescribed here. From our observations, it seems that a further5 min during which the isoflurane concentration was increasedto 2.5% would prove fatal to the animals unless they are arti-ficially respired for 25 min due to respiratory failure. Shorterperiods of exposure to the higher isoflurane concentration mightsolve this problem. However, all four of the animals studied heresurvived the isoflurane “overdose” without apparent harm. Fur-ther experiments will be needed to determine an appropriatemaintenance concentration of isoflurane.
While it is difficult to directly compare the octopus’s respi-ratory system with that of vertebrates, the former has a clear
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
292 POLESE ET AL.
FIGURE 7. Normalized respiratory rate (%) as determined by the number of mantle contractions per minute in 4 animals for which the metered isofluraneconcentration was raised to 2.5% (volume basis). Readings were taken at 5-min intervals. After 55 min the seawater in the tank was refreshed and the isofluraneflow was switched off, allowing the animals to be normally aerated. However, in these animals respiratory pumping ceased altogether and aeration of the gillscould only be maintained by artificial respiration using gentle massage of the mantle to resuscitate the animals.
respiratory rhythm from which it may be inferred that there issome form of central respiratory pattern generator within itssystem that is presumably acted upon by a number of externalfactors (such as external oxygen concentration) and that inter-acts with the cardiovascular system. In mammals, inhaled anes-thetics depress the volume of respiration but not necessarily itsfrequency, although irreversible anesthesia results in respiratoryarrest and circulatory failure in a matter of minutes. At safe con-centrations isoflurane tends to increase respiratory frequency inhumans but produces profound respiratory depression by pro-gressively decreasing the tidal volume with increasing depthof anesthesia (Fourcade et al. 1971; Pauca and Dripps 1973).Furthermore, the excitatory phase of anesthesia in mammals(Guedel 1937) can result in struggling behavior and a rise inheart rate, which may explain the rise in respiratory frequencyobserved in our experiment with 1–2% isoflurane (Figure 2).
In our study octopi, we were only able to monitor the changesin respiratory frequency as anesthesia proceeded. There was afall in frequency at low concentrations of isoflurane, but fre-quency returned to the normal range as the concentration in-creased (Figure 2). One interpretation of the reduced respira-
tory frequency at low isoflurane concentrations is that whileit is “conscious” the animal senses the compound as possiblyharmful or unpleasant and reduces its respiratory frequency tocompensate, while at higher concentrations the animal has lostconsciousness and the isoflurane has an uncompensated effectsimilar to that in humans (Eger 1981). This interpretation issupported by the fact that the removal of isoflurane from the ex-ternal medium results in a fall in respiratory frequency, but thisfall could also be due to the fact that the isoflurane is still in theprocess of penetrating the octopus’s respiratory centers (whichmay be located in the basal lobe system, particularly the medianbasal lobes of the brain; Nixon and Young 2003). This possi-bility is supported by the observation that in octopi that were“overdosed” with isoflurane respiration ceased entirely and hadto be maintained artificially (a result that may have been dueto global suppression of the octopus’s respiratory center andassociated respiratory motor neurons, as has been observed inmammals; Berge and Warner 2000). Care must be taken notto cause complete respiratory suppression when anesthetizingoctopi. It should be noted that the hyperexcitability observed atexposure to 2% isoflurane (which occurs just before the loss of
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
EFFECTS OF ISOFLURANE ON OCTOPUS VULGARIS 293
FIGURE 8. Normalized withdrawal responses (6 = strong, 4 = medium, 2 =low, and 0 = none) of (A) siphons and (B) arms in response to a touch stimulusduring a progressive increase in isoflurane concentration up to 2.5% followedby flushing with clean oxygenated seawater.
all reactivity to touch stimuli) signals that the animal is about tosuccumb to the anesthetic at a depth sufficient to allow surgicalmanipulation.
It is difficult to obtain a median effective dose (ED50) forisoflurane with the small number of animals used in this study.However, it is clear from our data that the level of anesthetizationis dose dependent and that 70% (10 of 14) animals were com-pletely anesthetized in 2% isoflurane, suggesting that the ED50is rather less than 2%. In mollusks such as Lymnaea stagnalisthe ED50 was found to be 1.09%, while in humans it was foundto be 1.15% and in cats Felis cattus it was 1.63% (Girdlestoneet al. 1989), all of which indicates a degree of species specificity.At this stage it is difficult to know exactly what concentrationof isoflurane reaches the central nervous system of octopi underthe conditions described here. However, Wells (1978) indicatesthat specific octopus brain areas are specialized for specificfunctions such as memory, learning, sensory integration, anddecision making (akin to the human cerebral cortex) and exhibita level of learning ability that “might signal consciousness” (El-
wood 2011). Thus, to determine the actions of anesthetics onoctopi will depend on a more detailed understanding of the braincenters involved in consciousness.
Withdrawal responses are protective reflexes and take prece-dence over other behaviors (Winlow and Polese 2014); the lackof responsiveness to touch stimuli was one indicator of anesthe-sia in humans before muscle relaxants (neuromuscular block-ing agents) came into common use 70 years ago (Griffith andJohnson 1942). In mollusks, well-studied examples are the gillwithdrawal reflex in sea slugs Aplysia spp. (Kandel 1976) orwhole-animal withdrawal in Lymnaea (Winlow et al. 1992). TheLymnaea whole-animal withdrawal response has been used as anindicator of anesthesia following application of the volatile anes-thetics halothane, enflurane, and isoflurane (Girdlestone et al.1989). In the current study, siphon and arm withdrawal declinedin a dose-dependent manner, indicating that both are good in-dicators of anesthetic state. This is supported by the fact thatmovements of the siphon and arms are hierarchically controlled(Boycott and Young 1950).
In cephalopods color change and patterning are known to beunder direct motor control from the brain (Hanlon and Mes-senger 1996) and respond to visual inputs to the anterior andposterior chromatophore lobes via the optic lobes and lateralbasal lobes. Stimulation of the anterior chromatophore lobe re-sults in unilateral darkening of the skin of the arms and thehead, while stimulation of the posterior lobe causes darkeningof the mantle skin via motor neurons to the radial muscles ofthe chromatophores (Hanlon and Messenger 1996). The grad-ual dose-dependent paling of octopi in the presence of isofluranesuggests that normal motor coordination of color change is lostas anesthesia proceeds and thus is a further indicator of theprogression of anesthesia.
We believe that the process of anesthesia induced by isoflu-rane is similar to that previously observed in Lymnaea and thatit is dose dependent. We have shown that isoflurane appearsto be a good reversible anesthetic for Octopus vulgaris, andwe have developed a method for its use. Further experimentsare required to determine the appropriate maintenance dose forsurgical procedures.
ACKNOWLEDGMENTSWe thank F. P. Ulloa Severino for his assistance in managing
and recording the video during the experiments. This work wassupported by a Single Center Research Grant in Neurosciencefrom Compagnia di San Paolo (Protocol 29-11).
REFERENCESAdvocates for Animals. 2005. Cephalopods and decapod crustaceans: their
capacity to experience pain and suffering. Advocates for Animals, Edinburgh,Scotland.
Andrews, P. L. R., A. S. Darmaillacq, N. Dennison, I. G. Gleadall, P. Hawkins, J.B. Messenger, D. Osorio, V. J. Smith, and J. A. Smith. 2013. The identificationand management of pain, suffering, and distress in cephalopods, includinganaesthesia, analgesia, and humane killing. Journal of Experimental MarineBiology and Ecology 447:46–64.
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14

294 POLESE ET AL.
Andrews, P. L. R., and E. M. Tansey. 1981. The effects of some anaestheticagents in Octopus vulgaris. Comparative Biochemistry and Physiology C70:241–247.
Berge, K. H., and D. O. Warner. 2000. Drugs that affect the respiratory system.Pages 491–498 in H. Hemmings and P. Hopkins, editors. Foundations ofanesthesia. Basic and clinical sciences. Mosby, London.
Boycott, B. B., and J. Z. Young. 1950. The comparative study of learning.Symposia of the Society for Experimental Biology 4:432–453.
Collins, T. J. 2007. ImageJ for microscopy. Biotechniques 43:25–30.Cooper, J. E. 2011. Anesthesia, analgesia, and euthanasia of invertebrates. ILAR
(Institute for Laboratory Animal Research) Journal 52:196–204.Crook, R. J., and E. T. Walters. 2011. Nociceptive behavior and physiology
of molluscs: animal welfare implications. ILAR (Institute for LaboratoryAnimal Research) Journal 52:185–195.
De Lisa, E., A. De Maio, L. L. Moroz, F. Moccia, M. R. F. Mennella, and A.Di Cosmo. 2012. Characterization of novel cytoplasmic PARP in the brain ofOctopus vulgaris. Biological Bulletin 222:176–181.
Ebert, T. J. 2006. Inhalation anesthesia. Pages 384–420 in G. Barash, B. F.Cullen, and R. K. Stoelting, editors. Clinical anesthesia. Lippincott Williamsand Wilkins, Philadelphia.
Eger, E. I. 1981. Isoflurane: a review. Anesthesiology 55:559–576.Elwood, R. W. 2011. Pain and suffering in invertebrates? ILAR (Institute for
Laboratory Animal Research) Journal 52:175–184.European Parliament. 2010. Council on the protection of animals used for scien-
tific purposes. Directive 2010/63/EU L276. Official Journal of the EuropeanUnion 53:33–79.
Field, K. J., and C. M. Lang. 1988. Hazards of urethane (ethyl carbamate): areview of the literature. Laboratory Animals 22:255–262.
Fourcade, H. E., W. C. Stevens, C. P. Larson Jr., T. H. Cromwell, S. H. Bahlman,R. F. Hickey, M. J. Halsey, and E. Eger Jr. 1971. Ventilatory effects of Forane,new inhaled anesthetic. Anesthesiology 35:26–31.
Frazier, D. T., and T. Narahashi. 1975. Tricaine (MS-222): effects on ionicconductances of squid axon membranes. European Journal of Pharmacology33:313–317.
Froesch, D., and H.-J. Marthy. 1975. The structure and function of the ovidu-cal gland in octopods (Cephalopoda). Proceedings of the Royal Society ofLondon B 188:95–101.
Girdlestone, D., S. G. H. Cruickshank, and W. Winlow. 1989. The actionsof 3 volatile general-anesthetics on withdrawal responses of the pond snailLymnaea stagnalis (L). Comparative Biochemistry and Physiology C 92:39–43.
Goncalves, R. A., C. Aragao, P. A. Frias, and A. V. Sykes. 2012. The use ofdifferent anaesthetics as welfare promoters during short-term human ma-nipulation of European cuttlefish (Sepia officinalis) juveniles. Aquaculture370-371:130–135.
Gray, E. G. 1970. The fine structure of the vertical lobe of octopus brain.Philosophical Transactions of the Royal Society of London B 258:379–394.
Griffith, H. R., and G. E. Johnson. 1942. The use of curare in general anesthesia.Anesthesiology 3:418–420.
Guedel, A. E. 1937. Inhalational anesthesia; a fundamental guide. Macmillan,New York.
Hanlon, R. T., and J. B. Messenger. 1996. Cephalopod behaviour. CambridgeUniversity Press, Cambridge, UK.
Harms, C. A., G. A. Lewbart, R. McAlarney, L. S. Christian, and C. Lemons.2006. Surgical excision of mycotic (Cladosporium sp.) granulomas from themantle of a cuttlefish (Sepia officinalis). Journal of Zoo and Wildlife Medicine37:524–530.
Horvath, K., D. Angeletti, G. Nascetti, and C. Carere. 2013. Invertebrate welfare:an overlooked issue. Annali dell’Istituto Superiore di Sanita 49:9–17.
Kandel, E. R. 1976. Cellular basis of behavior: an introduction to behavioralneurobiology. Freeman, San Francisco.
Latham, S. R. 2012. U.S. law and animal experimentation: a critical primer.Hastings Center Report 42:S35–S39.
Magee, B., and R. W. Elwood. 2013. Shock avoidance by discrimination learningin the shore crab (Carcinus maenas) is consistent with a key criterion for pain.Journal of Experimental Biology 216:353–358.
Mather, J. A. 2011. Philosophical background of attitudes toward and treatmentof invertebrates. ILAR (Institute for Laboratory Animal Research) Journal52:205–212.
Mather, J. A., and R. C. Anderson. 1993. Personalities of octopuses(Octopus rubescens). Journal of Comparative Psychology 107:336–340.
McCrohan, C. R., D. Girdlestone, and W. Winlow. 1987. Effects of halothaneon feeding motor-activity in the snail Lymnaea stagnalis. Comparative Bio-chemistry and Physiology C 86:55–62.
Messenger, J. B. 1968. The visual attack of the cuttlefish, Sepia officinalis.Animal Behaviour 16:342–357.
Messenger, J. B., M. Nixon, and K. P. Ryan. 1985. Magnesium chloride asan anesthetic for cephalopods. Comparative Biochemistry and Physiology C82:203–205.
Moghadam, H. F., and W. Winlow. 1995. Pentobarbital inhibits K + current incultured neurons. Journal of Physiology (London) 483:191.
Moltschaniwskyj, N. A., K. Hall, M. R. Lipinski, J. E. A. R. Marian, M.Nishiguchi, M. Sakai, D. J. Shulman, B. Sinclair, D. L. Sinn, M. Staudinger,R. van Gelderen, R. Villanueva, and K. Warnke. 2007. Ethical and welfareconsiderations when using cephalopods as experimental animals. Reviews inFish Biology and Fisheries 17:455–476.
Mooney, T. A., W. J. Lee, and R. T. Hanlon. 2010. Long-duration anestheti-zation of squid (Doryteuthis pealeii). Marine and Freshwater Behaviour andPhysiology 43:297–303.
Nixon, M. J., and J. Z. Young. 2003. The brains and lives of cephalopods. OxfordUniversity Press, New York.
Palmer, L. M., and A. F. Mensinger. 2004. Effect of the anaesthetic tricaine(MS-222) on nerve activity in the anterior lateral line of the oyster toadfish,Opsanus tau. Journal of Neurophysiology 92:1034–1041.
Pauca, A. L., and R. D. Dripps. 1973. Clinical experience with isoflu-rane (Forane): preliminary communication. British Journal of Anaesthesia45:697–703.
Rotava, P., I. L. Cavalcanti, L. Barrucand, L. A. Vane, and N. Vercosa. 2013.Effects of magnesium sulphate on the pharmacodynamics of rocuronium inpatients aged 60 years and older, a randomised trial. European Journal ofAnaesthesiology 30:599–604.
Sharman, K. 2004. Animal law in Australia. Available: http://animallaw.info/article/animal-law-australia. (October 2014).
Spencer, G. E., N. I. Syed, K. Lukowiak, and W. Winlow. 1996. Halothaneaffects both inhibitory and excitatory synaptic transmission at a singleidentified molluscan synapse, in vivo and in vitro. Brain Research 714:38–48.
UK Statutory Instruments. 1993. The Animals (Scientific Procedures) Act(Amendment) Order 1993 Number 2103. UK Government, London.
Wells, M. J. 1978. Octopus: physiology and behaviour of an advanced inverte-brate. Chapman and Hall, London.
Winlow, W., P. M. Hopkins, H. F. Moghadam, I. A. Ahmed, and T. Yar. 1995.Multiple cellular and subcellular actions of general anaesthetics on culturedmolluscan neurones. Acta Biologica Hungarica 46:381–393.
Winlow, W., L. L. Moroz, and N. I. Syed. 1992. Mechanisms of behaviouralselection in Lymnaea stagnalis. Pages 52–72 in J. Kien, C. R. McCrohan, andW. Winlow, editors. Neurobiology of motor programme selection. Pergamon,Oxford, UK.
Winlow, W., and G. Polese. 2014. A neuroplastic network underlying behaviourand seasonal change in Lymnaea stagnalis: a neuroecological standpoint.Pages 145–176 in A. Di Cosmo and W. Winlow, editors. Neuroecology andneuroethology in molluscs: the interface between behaviour and environment.Nova Science Publishers, New York.
Winlow, W., G. E. Spencer, N. I. Syed, and M. M. Qazzaz. 1998. Modula-tion of reconstructed peptidergic synapses and electrical synapses by generalanaesthetics. Toxicology Letters 100/101:77–84.
Yar, T., and W. Winlow. 1993. Halothane depressed a high-voltage-activated cal-cium current in identified cultured neurons from Lymnaea stagnalis. Journalof Physiology (London) 459:12.
Young, J. Z. 1971. The anatomy of the nervous system of Octopus vulgaris.Clarendon Press, Oxford, UK.
Dow
nloa
ded
by [
Prof
esso
r G
ianl
uca
Pole
se]
at 1
3:34
06
Nov
embe
r 20
14
Related Documents











