Neuron, Vol. 38, 291–303, April 24, 2003, Copyright 2003 by Cell Press Dopaminergic Supersensitivity in G Protein-Coupled Receptor Kinase 6-Deficient Mice and GRK5 (Benovic and Gomez, 1993; Premont et al., 1999). Multiple GRK enzymes (GRK2, GRK3, GRK5, and GRK6) are found throughout the brain (Gainetdinov et al., 2000; Erdtmann-Vourliotis et al., 2001), but the relative physiological importance of each GRK to the regulation Raul R. Gainetdinov, 1,2 Laura M. Bohn, 1,2 Tatyana D. Sotnikova, 1,2 Michel Cyr, 1,2 Aki Laakso, 1,2 Alexander D. Macrae, 1,3,5 Gonzalo E. Torres, 1,2 Kyeong-Man Kim, 1,2 Robert J. Lefkowitz, 1,3,4, * Marc G. Caron, 1,2,3, * and Richard T. Premont 3 of any given neurotransmitter receptor is unclear. The catecholamine dopamine plays a critical role in 1 Howard Hughes Medical Institute Laboratories 2 Department of Cell Biology several brain functions, including movement control, emotion, and affect (Carlsson, 2001; Greengard, 2001). 3 Department of Medicine 4 Department of Biochemistry Physiological responses to dopamine are controlled by a family of five G protein-coupled dopamine receptors Duke University Medical Center Durham, North Carolina 27710 (D1-D5), which are classified into D1-like (D1, D5) and D2-like (D2, D3, D4) subfamilies based on functional and pharmacological properties (Grandy and Civelli, 1992; Schwartz et al., 1993; Seeman and Van Tol, 1994; Picetti Summary et al., 1997; Missale et al., 1998; Sibley, 1999). Sensitivity of dopamine receptors to endogenous and exogenous Brain dopaminergic transmission is a critical compo- ligands is known to be an important modulator of dopa- nent in numerous vital functions, and its dysfunction mine-related functions in physiology and pathology. Su- is involved in several disorders, including addiction persensitivity of dopamine signaling has been postu- and Parkinson’s disease. Responses to dopamine are lated in several brain disorders (Pandey et al., 1977; mediated via G protein-coupled dopamine receptors Jenner and Marsden, 1987; Kirkpatrick et al., 1992; (D1-D5). Desensitization of G protein-coupled recep- Singer, 1994; Willner, 1997), including addiction (Nestler tors is mediated via phosphorylation by members of and Aghajanian, 1997; Nestler, 2001; Hyman and Ma- the family of G protein-coupled receptor kinases lenka, 2001; Laakso et al., 2002). Particularly, it is be- (GRK1-GRK7). Here we show that GRK6-deficient lieved that chronic psychostimulant-induced sensitiza- mice are supersensitive to the locomotor-stimulating tion, an early biochemical and behavioral manifestation effect of psychostimulants, including cocaine and am- of cellular plasticity leading to addiction, is associated phetamine. In addition, these mice demonstrate an with long-term changes in dopamine receptor respon- enhanced coupling of striatal D2-like dopamine recep- siveness (Nestler, 2001; Hyman and Malenka, 2001; tors to G proteins and augmented locomotor response Laakso et al., 2002). Previous in vitro studies have shown to direct dopamine agonists both in intact and in dopa- that the D1, D2, and D3 dopamine receptors can be mine-depleted animals. The present study indicates phosphorylated by some GRKs (Tiberi et al., 1996; Ito that postsynaptic D2-like dopamine receptors are et al., 1999; Kim et al., 2001), but the role of GRK6 in physiological targets for GRK6 and suggests that this dopamine receptor regulation has never been investi- regulatory mechanism contributes to central dopa- gated. Furthermore, no data on physiological significance minergic supersensitivity. of receptor regulation or the in vivo specificity of dopa- mine receptor/GRK interaction are currently available. Introduction To understand the role of GPCR desensitization mechanisms in the regulation of dopamine function, we G protein-coupled receptors (GPCRs) are regulated via have begun to examine mice bearing inactivated GRK activation-dependent phosphorylation by a family of G genes for alterations in cocaine responses and dopa- protein-coupled receptor kinases (GRKs) (Premont et mine receptor properties. In a previous investigation, al., 1995; Pitcher et al., 1998; Penn et al., 2000). This we found no evidence for dopaminergic supersensitivity process, which modulates the coupling state of GPCRs in mice lacking GRK5 (Gainetdinov et al., 1999). Here to their G proteins, is critically involved in GPCR desensi- we report that mice lacking GRK6 are supersensitive to tization (Pitcher et al., 1998; Penn et al., 2000). GRK- locomotor stimulating effects of several psychostimu- phosphorylated receptors bind to an arrestin protein, lants, display enhanced coupling of D2-like receptors which prevents the receptor from activating more G to their G proteins, and show enhanced behavioral ef- proteins despite the continued binding of agonist fects of direct dopamine agonists. (Pitcher et al., 1998). Seven distinct GRK genes are known, named GRK1 through GRK7, that are classified Results into three distinct groups (Pitcher et al., 1998; Premont et al., 1999; Penn et al., 2000). GRK6 is a member of the Generation of GRK6-KO Mice GRK4 subfamily of GRKs, which also contains GRK4 To address the functions of GRK6 in vivo, the GRK6 gene was targeted by homologous recombination in ES *Correspondence: [email protected] (M.G.C.), lefko001@ cells, as shown in Figures 1A–1C. The targeting con- receptor-biol.duke.edu (R.J.L.) struct introduces into the mouse GRK6 gene two loxP 5 Current address: Clinical Research and Development, Glaxo- SmithKline, Harlow, Essex, United Kingdom. sites, flanking exons 3 through 9, as well as a selection

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neuron, Vol. 38, 291–303, April 24, 2003, Copyright 2003 by Cell Press
Dopaminergic Supersensitivityin G Protein-Coupled ReceptorKinase 6-Deficient Mice
and GRK5 (Benovic and Gomez, 1993; Premont et al.,1999). Multiple GRK enzymes (GRK2, GRK3, GRK5, andGRK6) are found throughout the brain (Gainetdinov et al.,2000; Erdtmann-Vourliotis et al., 2001), but the relativephysiological importance of each GRK to the regulation
Raul R. Gainetdinov,1,2 Laura M. Bohn,1,2
Tatyana D. Sotnikova,1,2 Michel Cyr,1,2
Aki Laakso,1,2 Alexander D. Macrae,1,3,5
Gonzalo E. Torres,1,2 Kyeong-Man Kim,1,2
Robert J. Lefkowitz,1,3,4,* Marc G. Caron,1,2,3,*and Richard T. Premont3 of any given neurotransmitter receptor is unclear.
The catecholamine dopamine plays a critical role in1Howard Hughes Medical Institute Laboratories2 Department of Cell Biology several brain functions, including movement control,
emotion, and affect (Carlsson, 2001; Greengard, 2001).3 Department of Medicine4 Department of Biochemistry Physiological responses to dopamine are controlled by
a family of five G protein-coupled dopamine receptorsDuke University Medical CenterDurham, North Carolina 27710 (D1-D5), which are classified into D1-like (D1, D5) and
D2-like (D2, D3, D4) subfamilies based on functional andpharmacological properties (Grandy and Civelli, 1992;Schwartz et al., 1993; Seeman and Van Tol, 1994; PicettiSummaryet al., 1997; Missale et al., 1998; Sibley, 1999). Sensitivityof dopamine receptors to endogenous and exogenousBrain dopaminergic transmission is a critical compo-ligands is known to be an important modulator of dopa-nent in numerous vital functions, and its dysfunctionmine-related functions in physiology and pathology. Su-is involved in several disorders, including addictionpersensitivity of dopamine signaling has been postu-and Parkinson’s disease. Responses to dopamine arelated in several brain disorders (Pandey et al., 1977;mediated via G protein-coupled dopamine receptorsJenner and Marsden, 1987; Kirkpatrick et al., 1992;(D1-D5). Desensitization of G protein-coupled recep-Singer, 1994; Willner, 1997), including addiction (Nestlertors is mediated via phosphorylation by members ofand Aghajanian, 1997; Nestler, 2001; Hyman and Ma-the family of G protein-coupled receptor kinaseslenka, 2001; Laakso et al., 2002). Particularly, it is be-(GRK1-GRK7). Here we show that GRK6-deficientlieved that chronic psychostimulant-induced sensitiza-mice are supersensitive to the locomotor-stimulatingtion, an early biochemical and behavioral manifestationeffect of psychostimulants, including cocaine and am-of cellular plasticity leading to addiction, is associatedphetamine. In addition, these mice demonstrate anwith long-term changes in dopamine receptor respon-enhanced coupling of striatal D2-like dopamine recep-siveness (Nestler, 2001; Hyman and Malenka, 2001;tors to G proteins and augmented locomotor responseLaakso et al., 2002). Previous in vitro studies have shownto direct dopamine agonists both in intact and in dopa-that the D1, D2, and D3 dopamine receptors can bemine-depleted animals. The present study indicatesphosphorylated by some GRKs (Tiberi et al., 1996; Itothat postsynaptic D2-like dopamine receptors areet al., 1999; Kim et al., 2001), but the role of GRK6 inphysiological targets for GRK6 and suggests that thisdopamine receptor regulation has never been investi-regulatory mechanism contributes to central dopa-gated. Furthermore, no data on physiological significanceminergic supersensitivity.of receptor regulation or the in vivo specificity of dopa-mine receptor/GRK interaction are currently available.Introduction
To understand the role of GPCR desensitizationmechanisms in the regulation of dopamine function, weG protein-coupled receptors (GPCRs) are regulated viahave begun to examine mice bearing inactivated GRKactivation-dependent phosphorylation by a family of Ggenes for alterations in cocaine responses and dopa-protein-coupled receptor kinases (GRKs) (Premont etmine receptor properties. In a previous investigation,al., 1995; Pitcher et al., 1998; Penn et al., 2000). Thiswe found no evidence for dopaminergic supersensitivityprocess, which modulates the coupling state of GPCRsin mice lacking GRK5 (Gainetdinov et al., 1999). Hereto their G proteins, is critically involved in GPCR desensi-we report that mice lacking GRK6 are supersensitive totization (Pitcher et al., 1998; Penn et al., 2000). GRK-locomotor stimulating effects of several psychostimu-phosphorylated receptors bind to an arrestin protein,lants, display enhanced coupling of D2-like receptorswhich prevents the receptor from activating more Gto their G proteins, and show enhanced behavioral ef-proteins despite the continued binding of agonistfects of direct dopamine agonists.(Pitcher et al., 1998). Seven distinct GRK genes are
known, named GRK1 through GRK7, that are classifiedResultsinto three distinct groups (Pitcher et al., 1998; Premont
et al., 1999; Penn et al., 2000). GRK6 is a member of theGeneration of GRK6-KO MiceGRK4 subfamily of GRKs, which also contains GRK4To address the functions of GRK6 in vivo, the GRK6gene was targeted by homologous recombination in ES
*Correspondence: [email protected] (M.G.C.), lefko001@cells, as shown in Figures 1A–1C. The targeting con-receptor-biol.duke.edu (R.J.L.)struct introduces into the mouse GRK6 gene two loxP5 Current address: Clinical Research and Development, Glaxo-
SmithKline, Harlow, Essex, United Kingdom. sites, flanking exons 3 through 9, as well as a selection
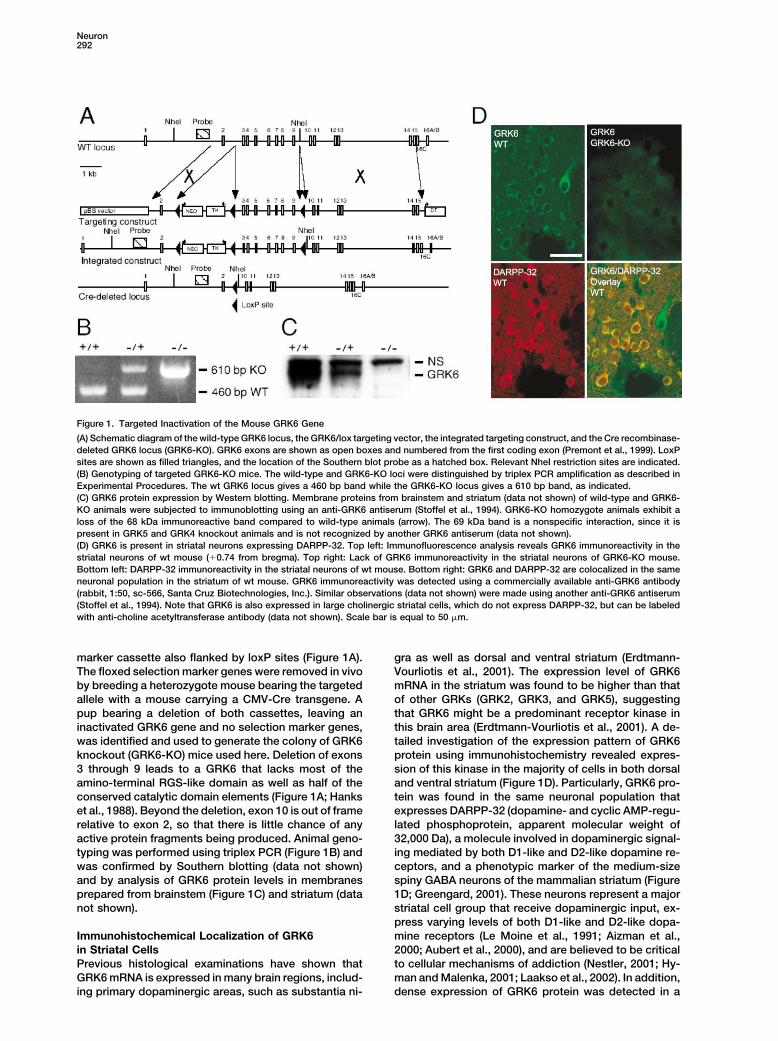
Neuron292
Figure 1. Targeted Inactivation of the Mouse GRK6 Gene
(A) Schematic diagram of the wild-type GRK6 locus, the GRK6/lox targeting vector, the integrated targeting construct, and the Cre recombinase-deleted GRK6 locus (GRK6-KO). GRK6 exons are shown as open boxes and numbered from the first coding exon (Premont et al., 1999). LoxPsites are shown as filled triangles, and the location of the Southern blot probe as a hatched box. Relevant NheI restriction sites are indicated.(B) Genotyping of targeted GRK6-KO mice. The wild-type and GRK6-KO loci were distinguished by triplex PCR amplification as described inExperimental Procedures. The wt GRK6 locus gives a 460 bp band while the GRK6-KO locus gives a 610 bp band, as indicated.(C) GRK6 protein expression by Western blotting. Membrane proteins from brainstem and striatum (data not shown) of wild-type and GRK6-KO animals were subjected to immunoblotting using an anti-GRK6 antiserum (Stoffel et al., 1994). GRK6-KO homozygote animals exhibit aloss of the 68 kDa immunoreactive band compared to wild-type animals (arrow). The 69 kDa band is a nonspecific interaction, since it ispresent in GRK5 and GRK4 knockout animals and is not recognized by another GRK6 antiserum (data not shown).(D) GRK6 is present in striatal neurons expressing DARPP-32. Top left: Immunofluorescence analysis reveals GRK6 immunoreactivity in thestriatal neurons of wt mouse (�0.74 from bregma). Top right: Lack of GRK6 immunoreactivity in the striatal neurons of GRK6-KO mouse.Bottom left: DARPP-32 immunoreactivity in the striatal neurons of wt mouse. Bottom right: GRK6 and DARPP-32 are colocalized in the sameneuronal population in the striatum of wt mouse. GRK6 immunoreactivity was detected using a commercially available anti-GRK6 antibody(rabbit, 1:50, sc-566, Santa Cruz Biotechnologies, Inc.). Similar observations (data not shown) were made using another anti-GRK6 antiserum(Stoffel et al., 1994). Note that GRK6 is also expressed in large cholinergic striatal cells, which do not express DARPP-32, but can be labeledwith anti-choline acetyltransferase antibody (data not shown). Scale bar is equal to 50 �m.
marker cassette also flanked by loxP sites (Figure 1A). gra as well as dorsal and ventral striatum (Erdtmann-Vourliotis et al., 2001). The expression level of GRK6The floxed selection marker genes were removed in vivo
by breeding a heterozygote mouse bearing the targeted mRNA in the striatum was found to be higher than thatof other GRKs (GRK2, GRK3, and GRK5), suggestingallele with a mouse carrying a CMV-Cre transgene. A
pup bearing a deletion of both cassettes, leaving an that GRK6 might be a predominant receptor kinase inthis brain area (Erdtmann-Vourliotis et al., 2001). A de-inactivated GRK6 gene and no selection marker genes,
was identified and used to generate the colony of GRK6 tailed investigation of the expression pattern of GRK6protein using immunohistochemistry revealed expres-knockout (GRK6-KO) mice used here. Deletion of exons
3 through 9 leads to a GRK6 that lacks most of the sion of this kinase in the majority of cells in both dorsaland ventral striatum (Figure 1D). Particularly, GRK6 pro-amino-terminal RGS-like domain as well as half of the
conserved catalytic domain elements (Figure 1A; Hanks tein was found in the same neuronal population thatexpresses DARPP-32 (dopamine- and cyclic AMP-regu-et al., 1988). Beyond the deletion, exon 10 is out of frame
relative to exon 2, so that there is little chance of any lated phosphoprotein, apparent molecular weight of32,000 Da), a molecule involved in dopaminergic signal-active protein fragments being produced. Animal geno-
typing was performed using triplex PCR (Figure 1B) and ing mediated by both D1-like and D2-like dopamine re-ceptors, and a phenotypic marker of the medium-sizewas confirmed by Southern blotting (data not shown)
and by analysis of GRK6 protein levels in membranes spiny GABA neurons of the mammalian striatum (Figure1D; Greengard, 2001). These neurons represent a majorprepared from brainstem (Figure 1C) and striatum (data
not shown). striatal cell group that receive dopaminergic input, ex-press varying levels of both D1-like and D2-like dopa-mine receptors (Le Moine et al., 1991; Aizman et al.,Immunohistochemical Localization of GRK6
in Striatal Cells 2000; Aubert et al., 2000), and are believed to be criticalto cellular mechanisms of addiction (Nestler, 2001; Hy-Previous histological examinations have shown that
GRK6 mRNA is expressed in many brain regions, includ- man and Malenka, 2001; Laakso et al., 2002). In addition,dense expression of GRK6 protein was detected in aing primary dopaminergic areas, such as substantia ni-
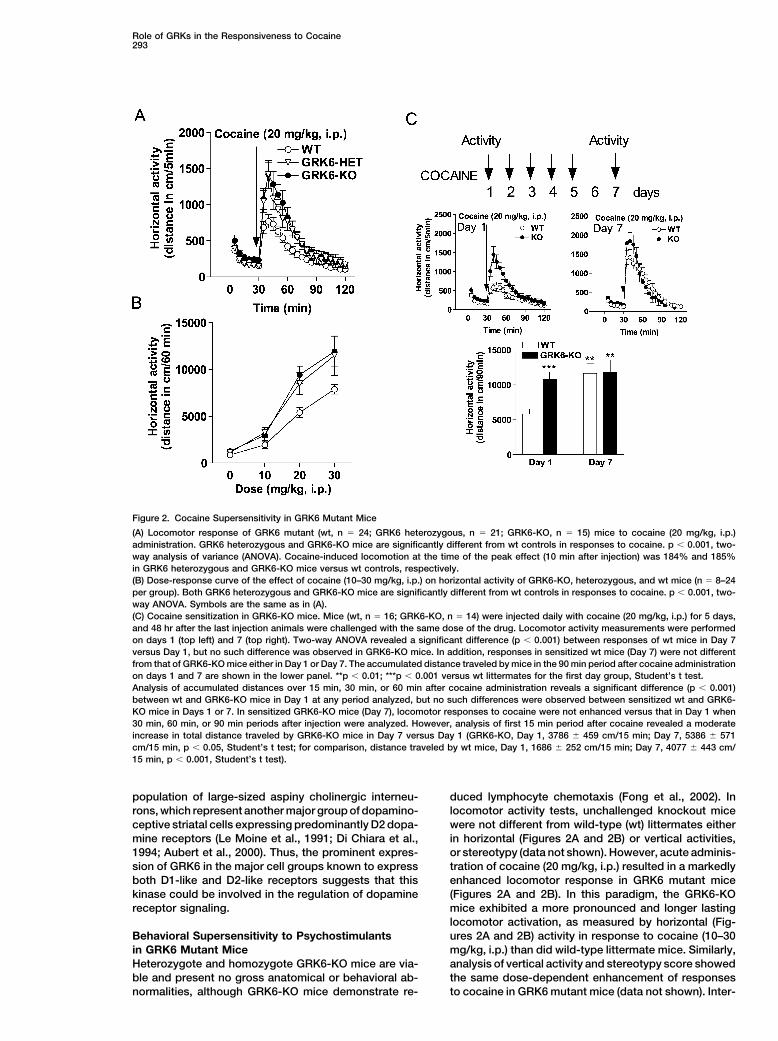
Role of GRKs in the Responsiveness to Cocaine293
Figure 2. Cocaine Supersensitivity in GRK6 Mutant Mice
(A) Locomotor response of GRK6 mutant (wt, n � 24; GRK6 heterozygous, n � 21; GRK6-KO, n � 15) mice to cocaine (20 mg/kg, i.p.)administration. GRK6 heterozygous and GRK6-KO mice are significantly different from wt controls in responses to cocaine. p � 0.001, two-way analysis of variance (ANOVA). Cocaine-induced locomotion at the time of the peak effect (10 min after injection) was 184% and 185%in GRK6 heterozygous and GRK6-KO mice versus wt controls, respectively.(B) Dose-response curve of the effect of cocaine (10–30 mg/kg, i.p.) on horizontal activity of GRK6-KO, heterozygous, and wt mice (n � 8–24per group). Both GRK6 heterozygous and GRK6-KO mice are significantly different from wt controls in responses to cocaine. p � 0.001, two-way ANOVA. Symbols are the same as in (A).(C) Cocaine sensitization in GRK6-KO mice. Mice (wt, n � 16; GRK6-KO, n � 14) were injected daily with cocaine (20 mg/kg, i.p.) for 5 days,and 48 hr after the last injection animals were challenged with the same dose of the drug. Locomotor activity measurements were performedon days 1 (top left) and 7 (top right). Two-way ANOVA revealed a significant difference (p � 0.001) between responses of wt mice in Day 7versus Day 1, but no such difference was observed in GRK6-KO mice. In addition, responses in sensitized wt mice (Day 7) were not differentfrom that of GRK6-KO mice either in Day 1 or Day 7. The accumulated distance traveled by mice in the 90 min period after cocaine administrationon days 1 and 7 are shown in the lower panel. **p � 0.01; ***p � 0.001 versus wt littermates for the first day group, Student’s t test.Analysis of accumulated distances over 15 min, 30 min, or 60 min after cocaine administration reveals a significant difference (p � 0.001)between wt and GRK6-KO mice in Day 1 at any period analyzed, but no such differences were observed between sensitized wt and GRK6-KO mice in Days 1 or 7. In sensitized GRK6-KO mice (Day 7), locomotor responses to cocaine were not enhanced versus that in Day 1 when30 min, 60 min, or 90 min periods after injection were analyzed. However, analysis of first 15 min period after cocaine revealed a moderateincrease in total distance traveled by GRK6-KO mice in Day 7 versus Day 1 (GRK6-KO, Day 1, 3786 � 459 cm/15 min; Day 7, 5386 � 571cm/15 min, p � 0.05, Student’s t test; for comparison, distance traveled by wt mice, Day 1, 1686 � 252 cm/15 min; Day 7, 4077 � 443 cm/15 min, p � 0.001, Student’s t test).
population of large-sized aspiny cholinergic interneu- duced lymphocyte chemotaxis (Fong et al., 2002). Inlocomotor activity tests, unchallenged knockout micerons, which represent another major group of dopamino-
ceptive striatal cells expressing predominantly D2 dopa- were not different from wild-type (wt) littermates eitherin horizontal (Figures 2A and 2B) or vertical activities,mine receptors (Le Moine et al., 1991; Di Chiara et al.,
1994; Aubert et al., 2000). Thus, the prominent expres- or stereotypy (data not shown). However, acute adminis-tration of cocaine (20 mg/kg, i.p.) resulted in a markedlysion of GRK6 in the major cell groups known to express
both D1-like and D2-like receptors suggests that this enhanced locomotor response in GRK6 mutant mice(Figures 2A and 2B). In this paradigm, the GRK6-KOkinase could be involved in the regulation of dopamine
receptor signaling. mice exhibited a more pronounced and longer lastinglocomotor activation, as measured by horizontal (Fig-ures 2A and 2B) activity in response to cocaine (10–30Behavioral Supersensitivity to Psychostimulants
in GRK6 Mutant Mice mg/kg, i.p.) than did wild-type littermate mice. Similarly,analysis of vertical activity and stereotypy score showedHeterozygote and homozygote GRK6-KO mice are via-
ble and present no gross anatomical or behavioral ab- the same dose-dependent enhancement of responsesto cocaine in GRK6 mutant mice (data not shown). Inter-normalities, although GRK6-KO mice demonstrate re-
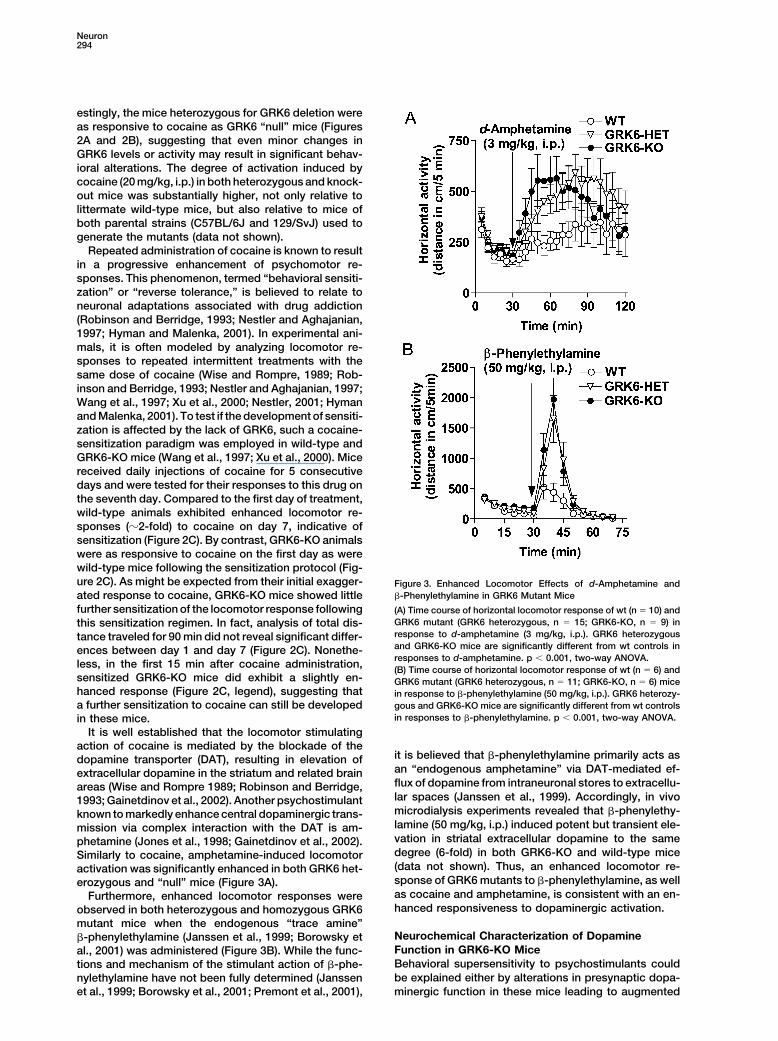
Neuron294
estingly, the mice heterozygous for GRK6 deletion wereas responsive to cocaine as GRK6 “null” mice (Figures2A and 2B), suggesting that even minor changes inGRK6 levels or activity may result in significant behav-ioral alterations. The degree of activation induced bycocaine (20 mg/kg, i.p.) in both heterozygous and knock-out mice was substantially higher, not only relative tolittermate wild-type mice, but also relative to mice ofboth parental strains (C57BL/6J and 129/SvJ) used togenerate the mutants (data not shown).
Repeated administration of cocaine is known to resultin a progressive enhancement of psychomotor re-sponses. This phenomenon, termed “behavioral sensiti-zation” or “reverse tolerance,” is believed to relate toneuronal adaptations associated with drug addiction(Robinson and Berridge, 1993; Nestler and Aghajanian,1997; Hyman and Malenka, 2001). In experimental ani-mals, it is often modeled by analyzing locomotor re-sponses to repeated intermittent treatments with thesame dose of cocaine (Wise and Rompre, 1989; Rob-inson and Berridge, 1993; Nestler and Aghajanian, 1997;Wang et al., 1997; Xu et al., 2000; Nestler, 2001; Hymanand Malenka, 2001). To test if the development of sensiti-zation is affected by the lack of GRK6, such a cocaine-sensitization paradigm was employed in wild-type andGRK6-KO mice (Wang et al., 1997; Xu et al., 2000). Micereceived daily injections of cocaine for 5 consecutivedays and were tested for their responses to this drug onthe seventh day. Compared to the first day of treatment,wild-type animals exhibited enhanced locomotor re-sponses (�2-fold) to cocaine on day 7, indicative ofsensitization (Figure 2C). By contrast, GRK6-KO animalswere as responsive to cocaine on the first day as werewild-type mice following the sensitization protocol (Fig-ure 2C). As might be expected from their initial exagger- Figure 3. Enhanced Locomotor Effects of d-Amphetamine and
�-Phenylethylamine in GRK6 Mutant Miceated response to cocaine, GRK6-KO mice showed littlefurther sensitization of the locomotor response following (A) Time course of horizontal locomotor response of wt (n � 10) and
GRK6 mutant (GRK6 heterozygous, n � 15; GRK6-KO, n � 9) inthis sensitization regimen. In fact, analysis of total dis-response to d-amphetamine (3 mg/kg, i.p.). GRK6 heterozygoustance traveled for 90 min did not reveal significant differ-and GRK6-KO mice are significantly different from wt controls inences between day 1 and day 7 (Figure 2C). Nonethe-responses to d-amphetamine. p � 0.001, two-way ANOVA.
less, in the first 15 min after cocaine administration, (B) Time course of horizontal locomotor response of wt (n � 6) andsensitized GRK6-KO mice did exhibit a slightly en- GRK6 mutant (GRK6 heterozygous, n � 11; GRK6-KO, n � 6) micehanced response (Figure 2C, legend), suggesting that in response to �-phenylethylamine (50 mg/kg, i.p.). GRK6 heterozy-a further sensitization to cocaine can still be developed gous and GRK6-KO mice are significantly different from wt controls
in responses to �-phenylethylamine. p � 0.001, two-way ANOVA.in these mice.It is well established that the locomotor stimulating
action of cocaine is mediated by the blockade of theit is believed that �-phenylethylamine primarily acts asdopamine transporter (DAT), resulting in elevation ofan “endogenous amphetamine” via DAT-mediated ef-extracellular dopamine in the striatum and related brainflux of dopamine from intraneuronal stores to extracellu-areas (Wise and Rompre 1989; Robinson and Berridge,lar spaces (Janssen et al., 1999). Accordingly, in vivo1993; Gainetdinov et al., 2002). Another psychostimulantmicrodialysis experiments revealed that �-phenylethy-known to markedly enhance central dopaminergic trans-lamine (50 mg/kg, i.p.) induced potent but transient ele-mission via complex interaction with the DAT is am-vation in striatal extracellular dopamine to the samephetamine (Jones et al., 1998; Gainetdinov et al., 2002).degree (6-fold) in both GRK6-KO and wild-type miceSimilarly to cocaine, amphetamine-induced locomotor(data not shown). Thus, an enhanced locomotor re-activation was significantly enhanced in both GRK6 het-sponse of GRK6 mutants to �-phenylethylamine, as wellerozygous and “null” mice (Figure 3A).as cocaine and amphetamine, is consistent with an en-Furthermore, enhanced locomotor responses werehanced responsiveness to dopaminergic activation.observed in both heterozygous and homozygous GRK6
mutant mice when the endogenous “trace amine”Neurochemical Characterization of Dopamine�-phenylethylamine (Janssen et al., 1999; Borowsky etFunction in GRK6-KO Miceal., 2001) was administered (Figure 3B). While the func-Behavioral supersensitivity to psychostimulants couldtions and mechanism of the stimulant action of �-phe-be explained either by alterations in presynaptic dopa-nylethylamine have not been fully determined (Janssen
et al., 1999; Borowsky et al., 2001; Premont et al., 2001), minergic function in these mice leading to augmented
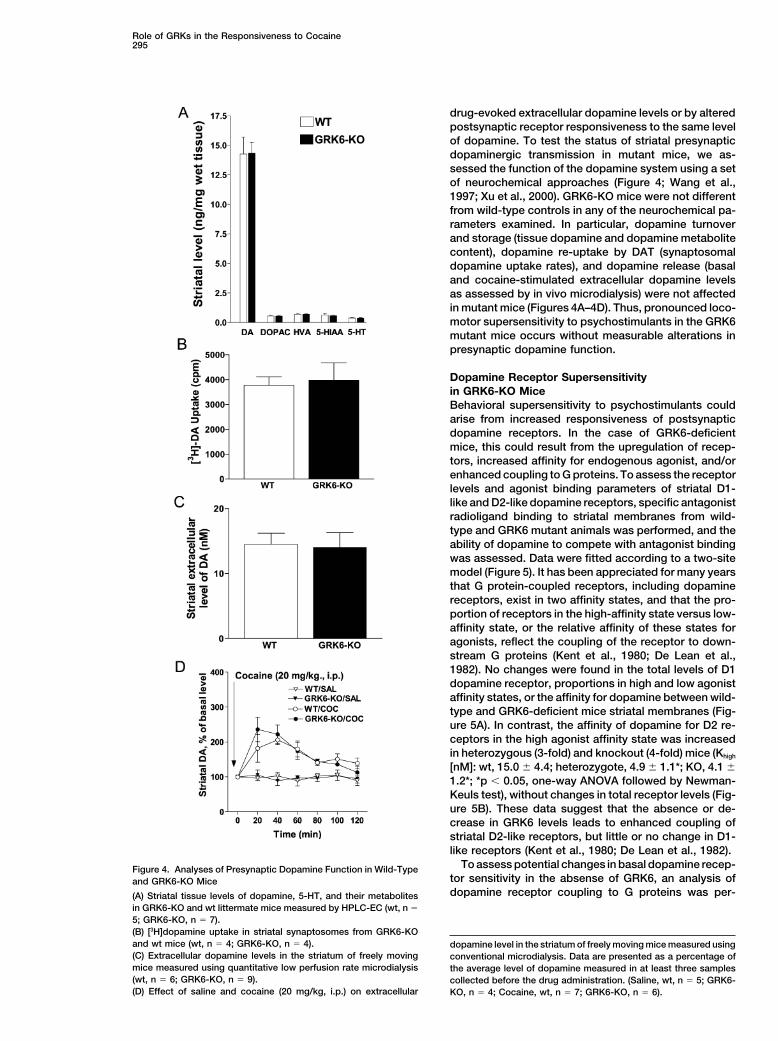
Role of GRKs in the Responsiveness to Cocaine295
drug-evoked extracellular dopamine levels or by alteredpostsynaptic receptor responsiveness to the same levelof dopamine. To test the status of striatal presynapticdopaminergic transmission in mutant mice, we as-sessed the function of the dopamine system using a setof neurochemical approaches (Figure 4; Wang et al.,1997; Xu et al., 2000). GRK6-KO mice were not differentfrom wild-type controls in any of the neurochemical pa-rameters examined. In particular, dopamine turnoverand storage (tissue dopamine and dopamine metabolitecontent), dopamine re-uptake by DAT (synaptosomaldopamine uptake rates), and dopamine release (basaland cocaine-stimulated extracellular dopamine levelsas assessed by in vivo microdialysis) were not affectedin mutant mice (Figures 4A–4D). Thus, pronounced loco-motor supersensitivity to psychostimulants in the GRK6mutant mice occurs without measurable alterations inpresynaptic dopamine function.
Dopamine Receptor Supersensitivityin GRK6-KO MiceBehavioral supersensitivity to psychostimulants couldarise from increased responsiveness of postsynapticdopamine receptors. In the case of GRK6-deficientmice, this could result from the upregulation of recep-tors, increased affinity for endogenous agonist, and/orenhanced coupling to G proteins. To assess the receptorlevels and agonist binding parameters of striatal D1-like and D2-like dopamine receptors, specific antagonistradioligand binding to striatal membranes from wild-type and GRK6 mutant animals was performed, and theability of dopamine to compete with antagonist bindingwas assessed. Data were fitted according to a two-sitemodel (Figure 5). It has been appreciated for many yearsthat G protein-coupled receptors, including dopaminereceptors, exist in two affinity states, and that the pro-portion of receptors in the high-affinity state versus low-affinity state, or the relative affinity of these states foragonists, reflect the coupling of the receptor to down-stream G proteins (Kent et al., 1980; De Lean et al.,1982). No changes were found in the total levels of D1dopamine receptor, proportions in high and low agonistaffinity states, or the affinity for dopamine between wild-type and GRK6-deficient mice striatal membranes (Fig-ure 5A). In contrast, the affinity of dopamine for D2 re-ceptors in the high agonist affinity state was increasedin heterozygous (3-fold) and knockout (4-fold) mice (Khigh
[nM]: wt, 15.0 � 4.4; heterozygote, 4.9 � 1.1*; KO, 4.1 �1.2*; *p � 0.05, one-way ANOVA followed by Newman-Keuls test), without changes in total receptor levels (Fig-ure 5B). These data suggest that the absence or de-crease in GRK6 levels leads to enhanced coupling ofstriatal D2-like receptors, but little or no change in D1-like receptors (Kent et al., 1980; De Lean et al., 1982).
To assess potential changes in basal dopamine recep-Figure 4. Analyses of Presynaptic Dopamine Function in Wild-Typetor sensitivity in the absense of GRK6, an analysis ofand GRK6-KO Micedopamine receptor coupling to G proteins was per-(A) Striatal tissue levels of dopamine, 5-HT, and their metabolites
in GRK6-KO and wt littermate mice measured by HPLC-EC (wt, n �
5; GRK6-KO, n � 7).(B) [3H]dopamine uptake in striatal synaptosomes from GRK6-KOand wt mice (wt, n � 4; GRK6-KO, n � 4). dopamine level in the striatum of freely moving mice measured using(C) Extracellular dopamine levels in the striatum of freely moving conventional microdialysis. Data are presented as a percentage ofmice measured using quantitative low perfusion rate microdialysis the average level of dopamine measured in at least three samples(wt, n � 6; GRK6-KO, n � 9). collected before the drug administration. (Saline, wt, n � 5; GRK6-(D) Effect of saline and cocaine (20 mg/kg, i.p.) on extracellular KO, n � 4; Cocaine, wt, n � 7; GRK6-KO, n � 6).
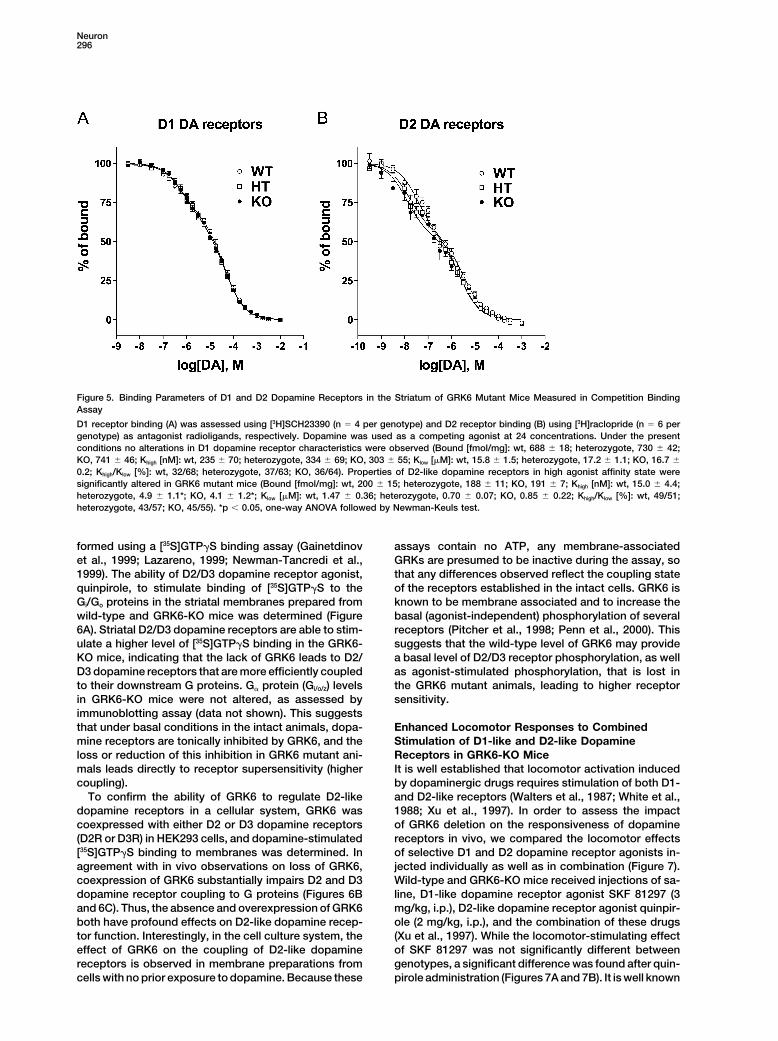
Neuron296
Figure 5. Binding Parameters of D1 and D2 Dopamine Receptors in the Striatum of GRK6 Mutant Mice Measured in Competition BindingAssay
D1 receptor binding (A) was assessed using [3H]SCH23390 (n � 4 per genotype) and D2 receptor binding (B) using [3H]raclopride (n � 6 pergenotype) as antagonist radioligands, respectively. Dopamine was used as a competing agonist at 24 concentrations. Under the presentconditions no alterations in D1 dopamine receptor characteristics were observed (Bound [fmol/mg]: wt, 688 � 18; heterozygote, 730 � 42;KO, 741 � 46; Khigh [nM]: wt, 235 � 70; heterozygote, 334 � 69; KO, 303 � 55; Klow [�M]: wt, 15.8 � 1.5; heterozygote, 17.2 � 1.1; KO, 16.7 �
0.2; Khigh/Klow [%]: wt, 32/68; heterozygote, 37/63; KO, 36/64). Properties of D2-like dopamine receptors in high agonist affinity state weresignificantly altered in GRK6 mutant mice (Bound [fmol/mg]: wt, 200 � 15; heterozygote, 188 � 11; KO, 191 � 7; Khigh [nM]: wt, 15.0 � 4.4;heterozygote, 4.9 � 1.1*; KO, 4.1 � 1.2*; Klow [�M]: wt, 1.47 � 0.36; heterozygote, 0.70 � 0.07; KO, 0.85 � 0.22; Khigh/Klow [%]: wt, 49/51;heterozygote, 43/57; KO, 45/55). *p � 0.05, one-way ANOVA followed by Newman-Keuls test.
formed using a [35S]GTP�S binding assay (Gainetdinov assays contain no ATP, any membrane-associatedGRKs are presumed to be inactive during the assay, soet al., 1999; Lazareno, 1999; Newman-Tancredi et al.,
1999). The ability of D2/D3 dopamine receptor agonist, that any differences observed reflect the coupling stateof the receptors established in the intact cells. GRK6 isquinpirole, to stimulate binding of [35S]GTP�S to the
Gi/Go proteins in the striatal membranes prepared from known to be membrane associated and to increase thebasal (agonist-independent) phosphorylation of severalwild-type and GRK6-KO mice was determined (Figure
6A). Striatal D2/D3 dopamine receptors are able to stim- receptors (Pitcher et al., 1998; Penn et al., 2000). Thissuggests that the wild-type level of GRK6 may provideulate a higher level of [35S]GTP�S binding in the GRK6-
KO mice, indicating that the lack of GRK6 leads to D2/ a basal level of D2/D3 receptor phosphorylation, as wellas agonist-stimulated phosphorylation, that is lost inD3 dopamine receptors that are more efficiently coupled
to their downstream G proteins. G� protein (Gi/o/z) levels the GRK6 mutant animals, leading to higher receptorsensitivity.in GRK6-KO mice were not altered, as assessed by
immunoblotting assay (data not shown). This suggeststhat under basal conditions in the intact animals, dopa- Enhanced Locomotor Responses to Combined
Stimulation of D1-like and D2-like Dopaminemine receptors are tonically inhibited by GRK6, and theloss or reduction of this inhibition in GRK6 mutant ani- Receptors in GRK6-KO Mice
It is well established that locomotor activation inducedmals leads directly to receptor supersensitivity (highercoupling). by dopaminergic drugs requires stimulation of both D1-
and D2-like receptors (Walters et al., 1987; White et al.,To confirm the ability of GRK6 to regulate D2-likedopamine receptors in a cellular system, GRK6 was 1988; Xu et al., 1997). In order to assess the impact
of GRK6 deletion on the responsiveness of dopaminecoexpressed with either D2 or D3 dopamine receptors(D2R or D3R) in HEK293 cells, and dopamine-stimulated receptors in vivo, we compared the locomotor effects
of selective D1 and D2 dopamine receptor agonists in-[35S]GTP�S binding to membranes was determined. Inagreement with in vivo observations on loss of GRK6, jected individually as well as in combination (Figure 7).
Wild-type and GRK6-KO mice received injections of sa-coexpression of GRK6 substantially impairs D2 and D3dopamine receptor coupling to G proteins (Figures 6B line, D1-like dopamine receptor agonist SKF 81297 (3
mg/kg, i.p.), D2-like dopamine receptor agonist quinpir-and 6C). Thus, the absence and overexpression of GRK6both have profound effects on D2-like dopamine recep- ole (2 mg/kg, i.p.), and the combination of these drugs
(Xu et al., 1997). While the locomotor-stimulating effecttor function. Interestingly, in the cell culture system, theeffect of GRK6 on the coupling of D2-like dopamine of SKF 81297 was not significantly different between
genotypes, a significant difference was found after quin-receptors is observed in membrane preparations fromcells with no prior exposure to dopamine. Because these pirole administration (Figures 7A and 7B). It is well known
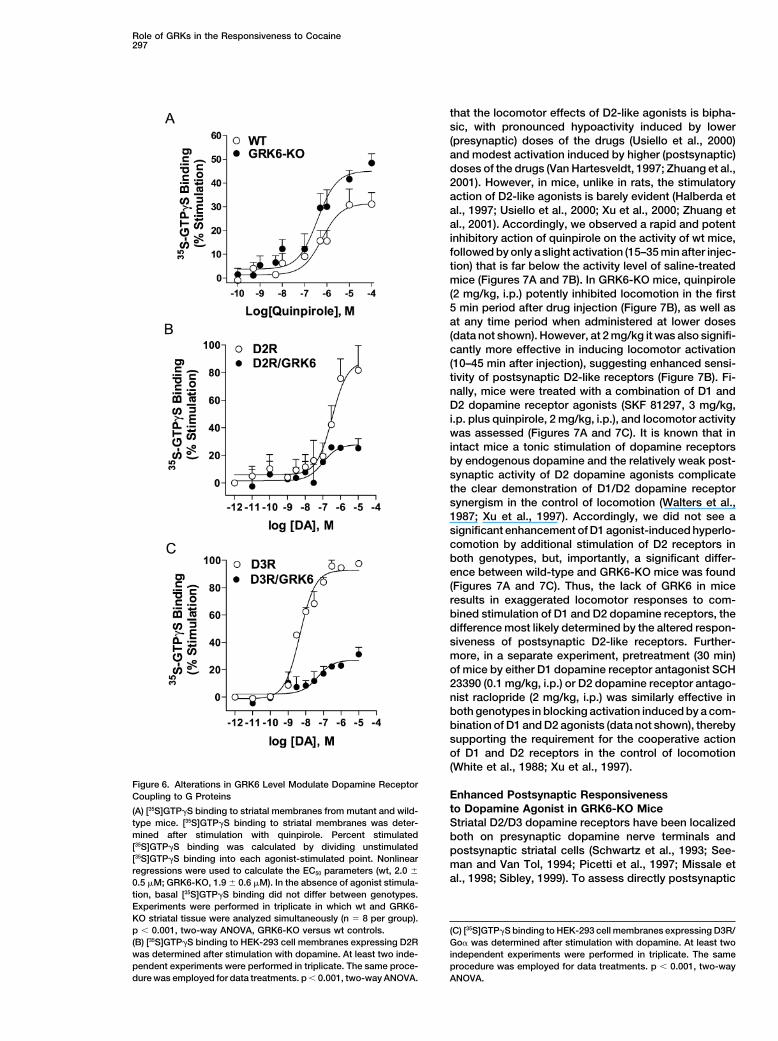
Role of GRKs in the Responsiveness to Cocaine297
that the locomotor effects of D2-like agonists is bipha-sic, with pronounced hypoactivity induced by lower(presynaptic) doses of the drugs (Usiello et al., 2000)and modest activation induced by higher (postsynaptic)doses of the drugs (Van Hartesveldt, 1997; Zhuang et al.,2001). However, in mice, unlike in rats, the stimulatoryaction of D2-like agonists is barely evident (Halberda etal., 1997; Usiello et al., 2000; Xu et al., 2000; Zhuang etal., 2001). Accordingly, we observed a rapid and potentinhibitory action of quinpirole on the activity of wt mice,followed by only a slight activation (15–35 min after injec-tion) that is far below the activity level of saline-treatedmice (Figures 7A and 7B). In GRK6-KO mice, quinpirole(2 mg/kg, i.p.) potently inhibited locomotion in the first5 min period after drug injection (Figure 7B), as well asat any time period when administered at lower doses(data not shown). However, at 2 mg/kg it was also signifi-cantly more effective in inducing locomotor activation(10–45 min after injection), suggesting enhanced sensi-tivity of postsynaptic D2-like receptors (Figure 7B). Fi-nally, mice were treated with a combination of D1 andD2 dopamine receptor agonists (SKF 81297, 3 mg/kg,i.p. plus quinpirole, 2 mg/kg, i.p.), and locomotor activitywas assessed (Figures 7A and 7C). It is known that inintact mice a tonic stimulation of dopamine receptorsby endogenous dopamine and the relatively weak post-synaptic activity of D2 dopamine agonists complicatethe clear demonstration of D1/D2 dopamine receptorsynergism in the control of locomotion (Walters et al.,1987; Xu et al., 1997). Accordingly, we did not see asignificant enhancement of D1 agonist-induced hyperlo-comotion by additional stimulation of D2 receptors inboth genotypes, but, importantly, a significant differ-ence between wild-type and GRK6-KO mice was found(Figures 7A and 7C). Thus, the lack of GRK6 in miceresults in exaggerated locomotor responses to com-bined stimulation of D1 and D2 dopamine receptors, thedifference most likely determined by the altered respon-siveness of postsynaptic D2-like receptors. Further-more, in a separate experiment, pretreatment (30 min)of mice by either D1 dopamine receptor antagonist SCH23390 (0.1 mg/kg, i.p.) or D2 dopamine receptor antago-nist raclopride (2 mg/kg, i.p.) was similarly effective inboth genotypes in blocking activation induced by a com-bination of D1 and D2 agonists (data not shown), therebysupporting the requirement for the cooperative actionof D1 and D2 receptors in the control of locomotion(White et al., 1988; Xu et al., 1997).
Figure 6. Alterations in GRK6 Level Modulate Dopamine ReceptorEnhanced Postsynaptic ResponsivenessCoupling to G Proteinsto Dopamine Agonist in GRK6-KO Mice(A) [35S]GTP�S binding to striatal membranes from mutant and wild-
type mice. [35S]GTP�S binding to striatal membranes was deter- Striatal D2/D3 dopamine receptors have been localizedmined after stimulation with quinpirole. Percent stimulated both on presynaptic dopamine nerve terminals and[35S]GTP�S binding was calculated by dividing unstimulated postsynaptic striatal cells (Schwartz et al., 1993; See-[35S]GTP�S binding into each agonist-stimulated point. Nonlinear man and Van Tol, 1994; Picetti et al., 1997; Missale etregressions were used to calculate the EC50 parameters (wt, 2.0 �
al., 1998; Sibley, 1999). To assess directly postsynaptic0.5 �M; GRK6-KO, 1.9 � 0.6 �M). In the absence of agonist stimula-tion, basal [35S]GTP�S binding did not differ between genotypes.Experiments were performed in triplicate in which wt and GRK6-KO striatal tissue were analyzed simultaneously (n � 8 per group).p � 0.001, two-way ANOVA, GRK6-KO versus wt controls. (C) [35S]GTP�S binding to HEK-293 cell membranes expressing D3R/(B) [35S]GTP�S binding to HEK-293 cell membranes expressing D2R Go� was determined after stimulation with dopamine. At least twowas determined after stimulation with dopamine. At least two inde- independent experiments were performed in triplicate. The samependent experiments were performed in triplicate. The same proce- procedure was employed for data treatments. p � 0.001, two-waydure was employed for data treatments. p � 0.001, two-way ANOVA. ANOVA.
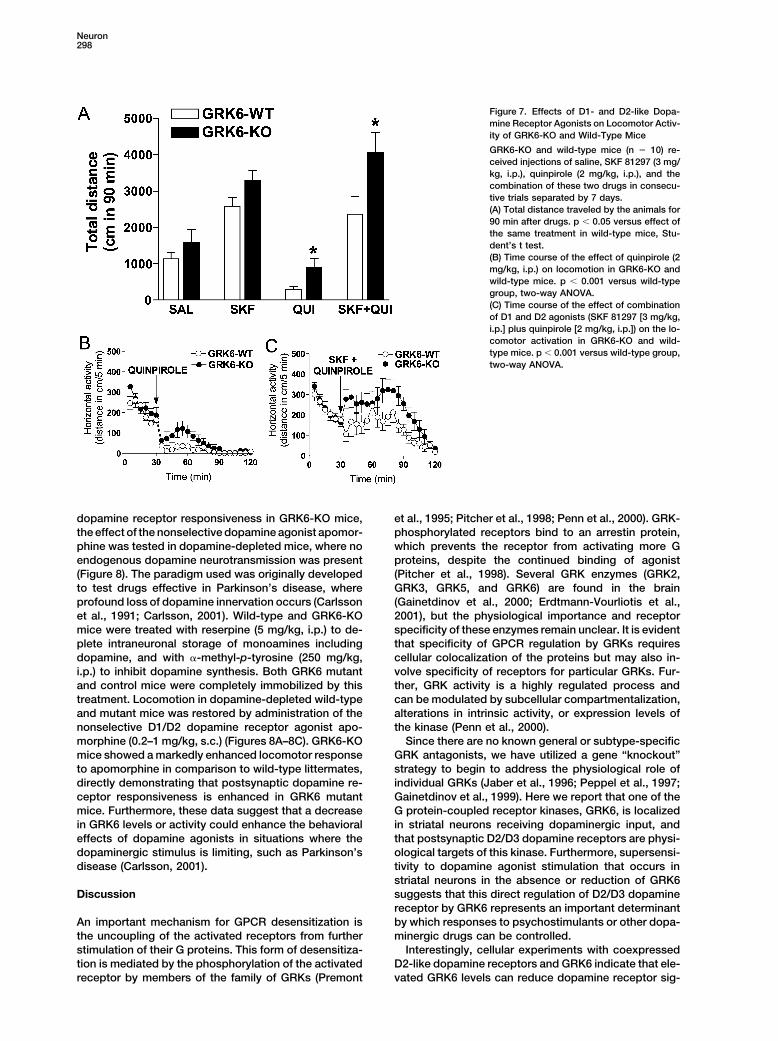
Neuron298
Figure 7. Effects of D1- and D2-like Dopa-mine Receptor Agonists on Locomotor Activ-ity of GRK6-KO and Wild-Type Mice
GRK6-KO and wild-type mice (n � 10) re-ceived injections of saline, SKF 81297 (3 mg/kg, i.p.), quinpirole (2 mg/kg, i.p.), and thecombination of these two drugs in consecu-tive trials separated by 7 days.(A) Total distance traveled by the animals for90 min after drugs. p � 0.05 versus effect ofthe same treatment in wild-type mice, Stu-dent’s t test.(B) Time course of the effect of quinpirole (2mg/kg, i.p.) on locomotion in GRK6-KO andwild-type mice. p � 0.001 versus wild-typegroup, two-way ANOVA.(C) Time course of the effect of combinationof D1 and D2 agonists (SKF 81297 [3 mg/kg,i.p.] plus quinpirole [2 mg/kg, i.p.]) on the lo-comotor activation in GRK6-KO and wild-type mice. p � 0.001 versus wild-type group,two-way ANOVA.
dopamine receptor responsiveness in GRK6-KO mice, et al., 1995; Pitcher et al., 1998; Penn et al., 2000). GRK-phosphorylated receptors bind to an arrestin protein,the effect of the nonselective dopamine agonist apomor-
phine was tested in dopamine-depleted mice, where no which prevents the receptor from activating more Gproteins, despite the continued binding of agonistendogenous dopamine neurotransmission was present
(Figure 8). The paradigm used was originally developed (Pitcher et al., 1998). Several GRK enzymes (GRK2,GRK3, GRK5, and GRK6) are found in the brainto test drugs effective in Parkinson’s disease, where
profound loss of dopamine innervation occurs (Carlsson (Gainetdinov et al., 2000; Erdtmann-Vourliotis et al.,2001), but the physiological importance and receptoret al., 1991; Carlsson, 2001). Wild-type and GRK6-KO
mice were treated with reserpine (5 mg/kg, i.p.) to de- specificity of these enzymes remain unclear. It is evidentthat specificity of GPCR regulation by GRKs requiresplete intraneuronal storage of monoamines including
dopamine, and with �-methyl-p-tyrosine (250 mg/kg, cellular colocalization of the proteins but may also in-volve specificity of receptors for particular GRKs. Fur-i.p.) to inhibit dopamine synthesis. Both GRK6 mutant
and control mice were completely immobilized by this ther, GRK activity is a highly regulated process andcan be modulated by subcellular compartmentalization,treatment. Locomotion in dopamine-depleted wild-type
and mutant mice was restored by administration of the alterations in intrinsic activity, or expression levels ofthe kinase (Penn et al., 2000).nonselective D1/D2 dopamine receptor agonist apo-
morphine (0.2–1 mg/kg, s.c.) (Figures 8A–8C). GRK6-KO Since there are no known general or subtype-specificGRK antagonists, we have utilized a gene “knockout”mice showed a markedly enhanced locomotor response
to apomorphine in comparison to wild-type littermates, strategy to begin to address the physiological role ofindividual GRKs (Jaber et al., 1996; Peppel et al., 1997;directly demonstrating that postsynaptic dopamine re-
ceptor responsiveness is enhanced in GRK6 mutant Gainetdinov et al., 1999). Here we report that one of theG protein-coupled receptor kinases, GRK6, is localizedmice. Furthermore, these data suggest that a decrease
in GRK6 levels or activity could enhance the behavioral in striatal neurons receiving dopaminergic input, andthat postsynaptic D2/D3 dopamine receptors are physi-effects of dopamine agonists in situations where the
dopaminergic stimulus is limiting, such as Parkinson’s ological targets of this kinase. Furthermore, supersensi-tivity to dopamine agonist stimulation that occurs indisease (Carlsson, 2001).striatal neurons in the absence or reduction of GRK6suggests that this direct regulation of D2/D3 dopamineDiscussionreceptor by GRK6 represents an important determinantby which responses to psychostimulants or other dopa-An important mechanism for GPCR desensitization is
the uncoupling of the activated receptors from further minergic drugs can be controlled.Interestingly, cellular experiments with coexpressedstimulation of their G proteins. This form of desensitiza-
tion is mediated by the phosphorylation of the activated D2-like dopamine receptors and GRK6 indicate that ele-vated GRK6 levels can reduce dopamine receptor sig-receptor by members of the family of GRKs (Premont
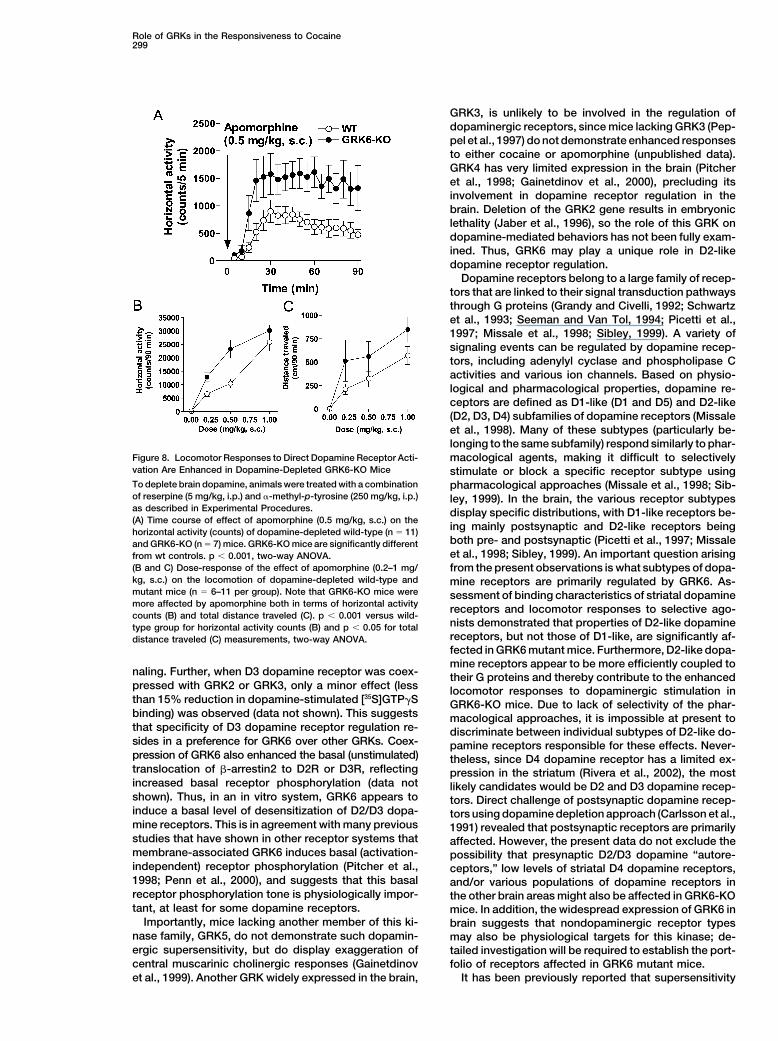
Role of GRKs in the Responsiveness to Cocaine299
GRK3, is unlikely to be involved in the regulation ofdopaminergic receptors, since mice lacking GRK3 (Pep-pel et al., 1997) do not demonstrate enhanced responsesto either cocaine or apomorphine (unpublished data).GRK4 has very limited expression in the brain (Pitcheret al., 1998; Gainetdinov et al., 2000), precluding itsinvolvement in dopamine receptor regulation in thebrain. Deletion of the GRK2 gene results in embryoniclethality (Jaber et al., 1996), so the role of this GRK ondopamine-mediated behaviors has not been fully exam-ined. Thus, GRK6 may play a unique role in D2-likedopamine receptor regulation.
Dopamine receptors belong to a large family of recep-tors that are linked to their signal transduction pathwaysthrough G proteins (Grandy and Civelli, 1992; Schwartzet al., 1993; Seeman and Van Tol, 1994; Picetti et al.,1997; Missale et al., 1998; Sibley, 1999). A variety ofsignaling events can be regulated by dopamine recep-tors, including adenylyl cyclase and phospholipase Cactivities and various ion channels. Based on physio-logical and pharmacological properties, dopamine re-ceptors are defined as D1-like (D1 and D5) and D2-like(D2, D3, D4) subfamilies of dopamine receptors (Missaleet al., 1998). Many of these subtypes (particularly be-longing to the same subfamily) respond similarly to phar-
Figure 8. Locomotor Responses to Direct Dopamine Receptor Acti- macological agents, making it difficult to selectivelyvation Are Enhanced in Dopamine-Depleted GRK6-KO Mice stimulate or block a specific receptor subtype usingTo deplete brain dopamine, animals were treated with a combination pharmacological approaches (Missale et al., 1998; Sib-of reserpine (5 mg/kg, i.p.) and �-methyl-p-tyrosine (250 mg/kg, i.p.) ley, 1999). In the brain, the various receptor subtypesas described in Experimental Procedures. display specific distributions, with D1-like receptors be-(A) Time course of effect of apomorphine (0.5 mg/kg, s.c.) on the
ing mainly postsynaptic and D2-like receptors beinghorizontal activity (counts) of dopamine-depleted wild-type (n � 11)both pre- and postsynaptic (Picetti et al., 1997; Missaleand GRK6-KO (n � 7) mice. GRK6-KO mice are significantly differentet al., 1998; Sibley, 1999). An important question arisingfrom wt controls. p � 0.001, two-way ANOVA.
(B and C) Dose-response of the effect of apomorphine (0.2–1 mg/ from the present observations is what subtypes of dopa-kg, s.c.) on the locomotion of dopamine-depleted wild-type and mine receptors are primarily regulated by GRK6. As-mutant mice (n � 6–11 per group). Note that GRK6-KO mice were sessment of binding characteristics of striatal dopaminemore affected by apomorphine both in terms of horizontal activity receptors and locomotor responses to selective ago-counts (B) and total distance traveled (C). p � 0.001 versus wild-
nists demonstrated that properties of D2-like dopaminetype group for horizontal activity counts (B) and p � 0.05 for totalreceptors, but not those of D1-like, are significantly af-distance traveled (C) measurements, two-way ANOVA.fected in GRK6 mutant mice. Furthermore, D2-like dopa-mine receptors appear to be more efficiently coupled to
naling. Further, when D3 dopamine receptor was coex- their G proteins and thereby contribute to the enhancedpressed with GRK2 or GRK3, only a minor effect (less locomotor responses to dopaminergic stimulation inthan 15% reduction in dopamine-stimulated [35S]GTP�S GRK6-KO mice. Due to lack of selectivity of the phar-binding) was observed (data not shown). This suggests macological approaches, it is impossible at present tothat specificity of D3 dopamine receptor regulation re- discriminate between individual subtypes of D2-like do-sides in a preference for GRK6 over other GRKs. Coex- pamine receptors responsible for these effects. Never-pression of GRK6 also enhanced the basal (unstimulated) theless, since D4 dopamine receptor has a limited ex-translocation of �-arrestin2 to D2R or D3R, reflecting pression in the striatum (Rivera et al., 2002), the mostincreased basal receptor phosphorylation (data not likely candidates would be D2 and D3 dopamine recep-shown). Thus, in an in vitro system, GRK6 appears to tors. Direct challenge of postsynaptic dopamine recep-induce a basal level of desensitization of D2/D3 dopa- tors using dopamine depletion approach (Carlsson et al.,mine receptors. This is in agreement with many previous 1991) revealed that postsynaptic receptors are primarilystudies that have shown in other receptor systems that affected. However, the present data do not exclude themembrane-associated GRK6 induces basal (activation- possibility that presynaptic D2/D3 dopamine “autore-independent) receptor phosphorylation (Pitcher et al., ceptors,” low levels of striatal D4 dopamine receptors,1998; Penn et al., 2000), and suggests that this basal and/or various populations of dopamine receptors inreceptor phosphorylation tone is physiologically impor- the other brain areas might also be affected in GRK6-KOtant, at least for some dopamine receptors. mice. In addition, the widespread expression of GRK6 in
Importantly, mice lacking another member of this ki- brain suggests that nondopaminergic receptor typesnase family, GRK5, do not demonstrate such dopamin- may also be physiological targets for this kinase; de-ergic supersensitivity, but do display exaggeration of tailed investigation will be required to establish the port-central muscarinic cholinergic responses (Gainetdinov folio of receptors affected in GRK6 mutant mice.
It has been previously reported that supersensitivityet al., 1999). Another GRK widely expressed in the brain,

Neuron300
LoxL vector with NotI and Sse8387I and ligated into those sites ofto light is present in GRK1-KO animals (Chen et al.,the LoxC/TK-NEO vector. The long recombination arm, floxed gene1999), cardiac supersensitivity to �-adrenergic stimula-fragment, and floxed TK-NEO marker cassette were excised fromtion is present in heterozygous GRK2-KO mice (Rock-this vector with NotI and AscI and inserted into those sites in the
man et al., 1998), supersensitivity to olfactory stimuli LoxR/DT vector to create the final targeting construct (Figure 1A).(Peppel et al., 1997) and enhanced muscarinic choliner- Growth and selection of targeted ES cells and creation of chimeric
mice was performed essentially as described (Hogan et al., 1994).gic airway responsiveness (Walker et al., 1999) are pres-The targeting DNA was linearized by digestion with NotI and electro-ent in GRK3-KO animals, and central muscarinic recep-porated into AK7 ES cells. Cells were selected for growth in mediator supersensitivity is observed in GRK5-KO micecontaining 200 �g/ml G418, and surviving clones were tested for(Gainetdinov et al., 1999). These observations as wellproper integration by amplification of a DNA band from the PGK
as present findings strongly suggest a generalized prin- promoter of the NEO marker gene to a region near exon 2 adjacentciple that abrogation of GRK-mediated receptor desen- to the targeting construct. The identity of positive clones was con-
firmed by Southern blotting of genomic DNA isolated from the cellssitization can enhance responses to both endogenousand digested with SpeI, using a probe from adjacent to exon 2and exogenous ligands and thus may represent a poten-(Figure 1A). A targeted cell clone with a normal karyotype was ex-tial mechanism for behavioral or physiological supersen-panded and microinjected into day 3.5 C57BL/6J mouse blasto-sitivity.cysts, which were then injected into the uterus of a day 2.5 pseudo-
One example of behavioral supersensitivity is chronic pregnant B6SJLF1/J mouse. Chimeric offspring were crossed withpsychostimulant-induced enhancement of dopaminer- C57BL/6J mice to generate agouti pups that carried the targeted
“lox” GRK6 gene. F1 heterozygote animals were bred with trans-gic responses (Wise and Rompre, 1989; Robinson andgenic mice bearing CMV-Cre (backcrossed to a C57Bl/6J geneticBerridge, 1993), which is believed to be associated withbackground; Nagy et al., 1998) to induce deletion of the floxedchanges in several signaling molecules downstream ofcassettes. From offspring of these crosses, GRK6-KO animals wereDA receptors (Nestler and Aghajanian, 1997; Hyman andobtained, in which both the exon 3–9 cassette and the TK-NEO
Malenka, 2001; Nestler, 2001; Laakso et al., 2002). The marker gene cassettes were deleted (i.e., the gene is inactive). Geno-present observatons raise the possibility that GRK6- typing was routinely performed on tail tip DNA using a PCR method
utilizing three primers to simultaneously detect the wild-type andregulated desensitization of dopamine receptors mightmutant loci (Figure 1B).represent a novel component in the development of
such sensitization. The observation that single-allele in-Western Blotsactivation of GRK6 in mice produces a phenotype identi-Mouse brain regions were dissected on ice and immediately frozencal to the complete knockout of the gene raises thein liquid nitrogen. Crude membranes were prepared from mouse
possibility that even subtle allelic variations in the human brain regions by polytron homogenization in buffer (20 mM Tris, 1GRK6 gene or drug-induced alterations in GRK6 activity mM EDTA, 100 mM NaCl [pH 7.4]) followed by centrifugation atmight contribute to individual sensitivity to psychostim- 200 g for 2 min and then at 21,000 g for 30 min. Aliquots
(60 �g) of each sample were solubilized by addition of SDS-PAGEulants and/or other drugs affecting dopaminergic func-sample buffer and separated by 10% SDS-PAGE. Transferred pro-tion. Furthermore, since supersensitivity of DA receptorsteins were blotted with polyclonal anti-GRK6 (Stoffel et al., 1994) andhas been implicated in the pathogenesis or adversevisualized using enhanced chemiluminescent development (ECL,
reactions associated with treatment of schizophrenia, Amersham, Piscataway, NJ).Tourette Syndrome, and Parkinson’s disease (Pandeyet al., 1977; Kirkpatrick et al., 1992; Jenner and Marsden, Immunohistochemistry1987; Singer, 1994), a role for GRK6-mediated DA recep- Wild-type or GRK6-KO mice (n � 3) were anesthetized with chloral
hydrate (400 mg/kg, i.p.) and perfused transcardially with 0.9% sa-tor regulation in these conditions would be of interestline, followed by 4% paraformaldehyde in 0.1 M borax buffer (pHto consider. Particularly, a pharmacological strategy9.5 at 4C). Brains were post-fixed for 1–3 days and cryoprotectedbased on blockade of GRK6-mediated desensitizationin 10% sucrose. Free floating coronal or saggital sections of 25 �m
may be effective in enhancing the potency of the low were incubated 32 hr at 4C with a mixture of antibodies againstlevels of residual dopamine found in parkinsonian pa- DARPP-32 (mouse, 1:5000, D97520, BD Transduction Laboratories)tients, or augmentating the efficacy of L-DOPA replace- and GRK6 (rabbit, 1:50, sc-566, Santa Cruz Biotechnologies, Inc.).
The second immunoreaction step was performed by incubation (1ment therapy.hr at room temperature) with each of the following antibodies: fluo-rescein-isothiocyanate-labeled goat anti-rabbit IgG and Texas Red-Experimental Procedureslabeled goat anti-mouse IgG (Vector Laboratories). Slides wereviewed on a laser scanning Zeiss confocal microscope (LSM-510)Targeted Deletion of the Mouse GRK6 Locususing the Roper Scientific Cooled CCD digital camera (Coolsnap-The three Triple-Lox vectors described for the GRK5 knockoutFX, BioVision Technologies, Inc., PA) and the IPLab software for(Gainetdinov et al., 1999) were modified as follows. The LoxL vectorWindows v3.0 for image processing (BioVision Technologies, Inc).was modified to have a new multiple cloning site containing NotIImages were acquired separately in each channel (dual scan mode)and NheI sites, a loxP element, and SpeI, XhoI, and Sse8387I sites.to eliminate the possibility of signal bleed-over from one channelThe LoxR/DT vector was modified to contain a multiple cloningto the other.region containing NotI, AscI, XbaI, XhoI, EcoRI, and PmeI sites.
Phage � carrying fragments of the mouse GRK6 gene from the129/SVJ strain were obtained and sequenced as described (Premont Animal Treatment/Drugs/Behavior
3- to 4-month-old littermate wild-type and GRK6 mutant miceet al., 1999). The 2.75 kb XbaI-NheI fragment containing exons 3through 9 (the gene fragment to be flanked by loxP sites, or floxed) (C57BL/6J 129/SvJ) were used in these experiments. In all experi-
ments, wild-type littermates served as controls for mutant mice,and the 6.8 kb NheI-NotI fragment containing exons 10 through 15(the long recombination arm) were ligated into the SpeI and NotI- and all the genotypes were evaluated concurrently. Horizontal, verti-
cal, and stereotypical activities of littermate wild-type, heretozygote,NheI sites of the LoxL vector, respectively. The 1.3 kb XbaI genefragment containing exon 2 (the short recombination arm) was pre- and knockout mice of both genders were measured in an Omnitech
Digiscan activity monitor (42 cm2). Locomotor activity was measuredpared by partial digestion and ligated into the XbaI site in the LoxR/DT vector. The cassette containing the long recombination arm, at 5 min intervals and cumulative counts were taken for data analysis
(Wang et al., 1997; Xu et al., 2000). To evaluate the effects of cocaine,loxP site, and gene fragment to be floxed were excised from the

Role of GRKs in the Responsiveness to Cocaine301
amphetamine, and �-phenylethylamine on locomotor behavior, mice 7.4), 120 mM NaCl, and 1 mM EDTA, and centrifuged for 20 min at40,000 g at 4C. Pellets were resuspended in the same buffer andwere placed in activity monitor, 30–60 min later they were injected
with drugs or vehicle i.p., and locomotor activity was monitored for incubated for 15 min at 37C to facilitate the removal of endogenousDA. After incubation, membranes were centrifuged for 10 min atthe following 90 min. In cocaine sensitization (Wang et al., 1997; Xu
et al., 2000) experiments, mice were treated chronically with cocaine 40,000 g at 4C, resuspended in ice-cold buffer, and centrifugedagain. The resulting pellet was then homogenized in the binding(20 mg/kg, i.p.) for 5 days and their responses to challenging dose
of cocaine were analyzed at day 7. To analyze the effect of direct buffer containing 50 mM Tris-HCl (pH 7.4), 120 mM NaCl, 5 mM KCl,2 mM CaCl2, 1 mM MgCl2, and 0.1% ascorbic acid. D1 receptordopamine agonist in dopamine-depleted mice (Carlsson et al.,
1991), animals were pretreated with a combination of reserpine (5 binding was assessed using 1 nM [3H]SCH23390 in the presence of100 nM ketanserin (to prevent binding to 5-HT2A receptors), andmg/kg, i.p. 20 hr before the experiment) and �-methyl-p-tyrosine
(250 mg/kg, i.p., 1 hr before the experiment). This treatment resulted D2 receptor binding was assessed using 3 nM [3H]raclopride (NewEngland Nuclear/Perkin Elmer Life Sciences). Dopamine was usedin depletion of striatal dopamine to less than 0.75% in both wild-
type and GRK6-KO mice (data not shown). Mice were completely as the competing agonist (24 concentration points in duplicate, from10�8.5 to 10�2 M for D1 receptor, and from 10�9.5 to 10�3 M for D2immobilized by this treatment. Dopamine-depleted mice were
treated with vehicle or D1/D2 dopamine receptor agonist apomor- receptor). Protein concentration was measured using the Lowryassay, and 40 �g and 60 �g of membrane protein in 200 �l reactionphine (0.2–1 mg/kg, s.c.), and locomotor activity was immediately
analyzed as described above. A nonrandomized repeated measures volume was used for each point for D1 and D2 receptor binding,respectively. Binding reaction was incubated for 45 min at roomdesign was employed to assess the locomotor effects of selective
and combined stimulation of D1-like and D2-like dopamine recep- temperature, and bound ligand was separated from free by rapidfiltration onto Whatman GF/C filters washed with ice-cold Tris buffer.tors (Xu et al., 1997). Once a week each mouse was habituated to
locomotor activity chambers (30 min) and received saline, R-(�)- Two-site competition curves were fitted individually for every animal(striata from two animals were pooled for each “n” in D2 bindingSKF-81297 (SKF 81297, 3 mg/kg, i.p.), quinpirole (2 mg/kg, i.p.), or
combination of SKF 81297 (3 mg/kg, i.p.) plus quinpirole (2 mg/kg, assay) by nonlinear regression using GraphPad Prism software(GraphPad Software Inc., San Diego, CA). Values of KHigh, KLow, frac-i.p.), consecutively. In all other experiments (with exception of the
sensitization paradigm), each animal received only a single injection tion of receptors in KHigh, and the amount of specifically bound ra-dioligand generated by nonlinear regression algorithm were sub-with a tested drug. All the data presented in this study are expressed
as means � SEM. jected to statistical analysis. Statistical significances were assessedwith one-way ANOVA followed by Newman-Keuls test for post hocanalyses.Neurochemical Assessments
For monoamine analyses, brain regions were dissected and mono-Acknowledgmentsamines extracted and analyzed for levels of dopamine, serotonin
(5-hydroxytryptamine, 5-HT), and metabolites 3,4-dihydroxyphenyl-This work was supported in part by grants from the National Insti-acetic acid (DOPAC), homovanillic acid (HVA), and 5-hydroxyindole-tutes of Health (DA-14600 to L.M.B., NS-19576 and MH-40159 toacetic acid (5-HIAA) using high performance liquid chromatographyM.G.C., and HL-16037 to R.J.L.) and unrestricted grants from Bristol-with electrochemical detection as described (Wang et al., 1997; XuMyers Squibb (Neuroscience [M.G.C.]; Cardiovascular [R.J.L.]). M.C.et al., 2000). To perform in vivo microdialysis experiments (Wang etwas supported by FRSQ fellowship. A.L. was supported in part byal., 1997; Xu et al., 2000), mice were anesthetized and dialysis probesAcademy of Finland. We thank Cheryl Bock and the Transgenic Corewere implanted into the right striatum. Twenty-four hours after sur-Facility of the Duke Comprehensive Cancer Center for assistance ingery, the dialysis probe was connected to a syringe pump andgenerating the GRK6-KO mice; Dr. Mark Bender (Fred Hutchisonperfused with artificial CSF. Quantitative “low perfusion” rate (70 nl/Cancer Research Center) for the C57Bl/6J-backcrossed CMV-Cremin) microdialysis experiments (Wang et al., 1997; Xu et al., 2000)transgenic mouse strain; and Sandy Duncan, Rachel McAdams,were conducted in freely moving mice for determination of basalKristina Riebe, and Susan Suter for animal care. M.G.C. and R.J.L.extracellular dopamine levels in striatum. To analyze the effects ofare Investigators of the Howard Hughes Medical Institute.cocaine on the extracellular dopamine levels in striatum, “conven-
tional” microdialysis method (perfusion flow rate 1 �l/min) in freelymoving animals was employed (Wang et al., 1997; Xu et al., 2000). Received: September 17, 2002
To measure [3H]dopamine uptake in striatal synaptosomes, stria- Revised: February 13, 2003tal tissue from wild-type and GRK6-KO mice were analyzed as de- Accepted: March 8, 2003scribed (Sandoval et al., 2001). Published: April 23, 2003
ReferencesAnalyses of Dopamine Receptor Coupling by [35S]GTP�SBinding In Vivo and In Vitro
Aizman, O., Brismar, H., Uhlen, P., Zettergren, E., Levey, A.I., Forss-In in vivo experiments, D2/D3 dopamine receptor agonist quinpirole-berg, H., Greengard, P., and Aperia, A. (2000). Anatomical and physi-stimulated [35S]GTP�S binding to striatal membranes from GRK6-ological evidence for D1 and D2 dopamine receptor colocalizationKO and wild-type mice was assessed as previously describedin neostriatal neurons. Nat. Neurosci. 3, 226–230.(Gainetdinov et al., 1999). To directly assess in vitro the role of GRK6
in D2 and D3 dopamine receptor regulation, dopamine-stimulated Aubert, I., Ghorayeb, I., Normand, E., and Bloch, B. (2000). Phenotyp-[35S]GTP�S binding to cultured cell membranes was used (Newman- ical characterization of the neurons expressing the D1 and D2 dopa-Tancredi et al., 1999). HEK-293 cells were transfected with D2R or mine receptors in the monkey striatum. J. Comp. Neurol. 418, 22–32.D3R/Go� with and without GRK6. For D2R, 20 �g of cell membrane Benovic, J.L., and Gomez, J. (1993). Molecular cloning and expres-proteins were incubated in a buffer containing 20 mM HEPES (pH sion of GRK6. A new member of the G protein-coupled receptor7.4), 10 mM MgCl2, 150 mM NaCl, 3 �M GDP, and 0.1 nM [35S]GTP�S kinase family. J. Biol. Chem. 268, 19521–19527.for 1 hr at room temperature. For D3R, 20 �g of cell membrane
Borowsky, B., Adham, N., Jones, K.A., Raddatz, R., Artymyshyn, R.,proteins were incubated in a buffer containing 25 mM HEPES (pHOgozalek, K.L., Durkin, M.M., Lakhlani, P.P., Bonini, J.A., Pathirana,7.4), 120 mM NaCl, 1.8 mM KCl, 20 mM MgCl2, 20 �M GDP, 0.2S., et al. (2001). Trace amines: identification of a family of mammaliannM [35S]GTP�S, and 1 mM sodium deoxycholate for 2 hr at 30C.G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 98, 8966–Incubation mixtures were filtered with GF/B filter and washed with8971.10 mM sodium phosphate buffer.Carlsson, A. (2001). A paradigm shift in brain research. Science 294,1021–1024.Agonist Competition Binding Assay
Striata from GRK6 wild-type, heterozygote, and knockout mice were Carlsson, M., Svensson, A., and Carlsson, A. (1991). Synergisticinteractions between muscarinic antagonists, adrenergic agonistsrapidly dissected on ice and frozen in liquid nitrogen. Tissue was
homogenized in an ice-cold buffer containing 50 mM Tris-HCl (pH and NMDA antagonists with respect to locomotor stimulatory effects

Neuron302
in monoamine-depleted mice. Naunyn Schmiedebergs Arch. Phar- Kim, K.M., Valenzano, K.J., Robinson, S.R., Yao, W.D., Barak, L.S.,and Caron, M.G. (2001). Differential regulation of the dopamine D2macol. 343, 568–573.and D3 receptors by G protein-coupled receptor kinases and beta-Chen, C.K., Burns, M.E., Spencer, M., Niemi, G.A., Chen, J., Hurley,arrestins. J. Biol. Chem. 276, 37409–37414.J.B., Baylor, D.A., and Simon, M.I. (1999). Abnormal photoresponsesKirkpatrick, B., Alphs, L., and Buchanan, R.W. (1992). The conceptand light-induced apoptosis in rods lacking rhodopsin kinase. Proc.of supersensitivity psychosis. J. Nerv. Ment. Dis. 180, 265–270.Natl. Acad. Sci. USA 96, 3718–3722.
Laakso, A., Mohn, A.R., Gainetdinov, R.R., and Caron, M.G. (2002).De Lean, A., Kilpatrick, B.F., and Caron, M.G. (1982). DopamineExperimental genetic approaches to addiction. Neuron 36, 213–228.receptor of the porcine anterior pituitary gland. Evidence for two
affinity states discriminated by both agonists and antagonists. Mol. Lazareno, S. (1999). Measurement of agonist-stimulated [35S]GTPPharmacol. 22, 290–297. gamma S binding to cell membranes. In Receptor Binding Tech-
niques, Volume 106, M. Keen, ed. (Totowa, NJ: Humana Press), pp.Di Chiara, G., Morelli, M., and Consolo, S. (1994). Modulatory func-231–247.tions of neurotransmitters in the striatum: ACh/dopamine/NMDA
interactions. Trends Neurosci. 17, 228–233. Le Moine, C., Normand, E., and Bloch, B. (1991). Phenotypical char-acterization of the rat striatal neurons expressing the D1 dopamineErdtmann-Vourliotis, M., Mayer, P., Ammon, S., Riechert, U., andreceptor gene. Proc. Natl. Acad. Sci. USA 88, 4205–4209.Hollt, V. (2001). Distribution of G-protein-coupled receptor kinase
(GRK) isoforms 2, 3, 5 and 6 mRNA in the rat brain. Brain Res. Mol. Missale, C., Nash, S.R., Robinson, S.W., Jaber, M., and Caron, M.G.Brain Res. 95, 129–137. (1998). Dopamine receptors: from structure to function. Physiol. Rev.
78, 189–225.Fong, A.M., Premont, R.T., Richardson, R.M., Yu, Y.R., Lefkowitz,R.J., and Patel, D.D. (2002). Defective lymphocyte chemotaxis in Nagy, A., Moens, C., Ivanyi, E., Pawling, J., Gertsenstein, M., Hadjan-beta-arrestin2- and GRK6-deficient mice. Proc. Natl. Acad. Sci. USA tonakis, A.K., Pirity, M., and Rossant, J. (1998). Dissecting the role99, 7478–7483. of N-myc in development using a single targeting vector to generate
a series of alleles. Curr. Biol. 8, 661–664.Gainetdinov, R.R., Bohn, L.M., Walker, J.K., Laporte, S.A., Macrae,A.D., Caron, M.G., Lefkowitz, R.J., and Premont, R.T. (1999). Musca- Nestler, E.J. (2001). Molecular basis of long-term plasticity underly-rinic supersensitivity and impaired receptor desensitization in G ing addiction. Nat. Rev. Neurosci. 2, 119–128.protein-coupled receptor kinase 5-deficient mice. Neuron 24, 1029– Nestler, E.J., and Aghajanian, G.K. (1997). Molecular and cellular1036. basis of addiction. Science 278, 58–63.Gainetdinov, R.R., Premont, R.T., Caron, M.G., and Lefkowitz, R.J. Newman-Tancredi, A., Cussac, D., Audinot, V., Pasteau, V., Gavau-(2000). Reply: receptor specificity of G-protein-coupled receptor dan, S., and Millan, M.J. (1999). G protein activation by human dopa-kinases. Trends Pharmacol. Sci. 21, 366–367. mine D3 receptors in high-expressing Chinese hamster ovary cells:Gainetdinov, R.R., Sotnikova, T.D., and Caron, M.G. (2002). Mono- a guanosine-5 -O-(3-[35S]thio)- triphosphate binding and antibodyamine transporter pharamacology and mutant mice. Trends Phar- study. Mol. Pharmacol. 55, 564–574.macol. Sci. 23, 367–373. Pandey, G.N., Garver, D.L., Tamminga, C., Ericksen, S., Ali, S.I., andGrandy, D.K., and Civelli, O. (1992). G-protein-coupled receptors: the Davis, J.M. (1977). Postsynaptic supersensitivity in schizophrenia.new dopamine receptor subtypes. Curr. Opin. Neurobiol. 2, 275–281. Am. J. Psychiatry 134, 518–522.Greengard, P. (2001). The neurobiology of slow synaptic transmis- Penn, R.B., Pronin, A.N., and Benovic, J.L. (2000). Regulation ofsion. Science 294, 1024–1030. G protein-coupled receptor kinases. Trends Cardiovasc. Med. 10,
81–89.Hanks, S.K., Quinn, A.M., and Hunter, T. (1988). The protein kinasefamily: conserved features and deduced phylogeny of the catalytic Peppel, K., Boekhoff, I., McDonald, P., Breer, H., Caron, M.G., anddomains. Science 241, 42–52. Lefkowitz, R.J. (1997). G protein-coupled receptor kinase 3 (GRK3)
gene disruption leads to loss of odorant receptor desensitization.Halberda, J.P., Middaugh, L.D., Gard, B.E., and Jackson, B.P. (1997).J. Biol. Chem. 272, 25425–25428.DAD1- and DAD2-like agonist effects on motor activity of C57 mice:
differences compared to rats. Synapse 26, 81–92. Picetti, R., Saiardi, A., Abdel Samad, T., Bozzi, Y., Baik, J.H., andBorrelli, E. (1997). Dopamine D2 receptors in signal transductionHogan, B., Beddington, R., Constantini, F., and Lacy, E. (1994).and behavior. Crit. Rev. Neurobiol. 11, 121–142.Manipulation of the Mouse Embryo: A Laboratory Manual, Second
Edition (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Pitcher, J.A., Freedman, N.J., and Lefkowitz, R.J. (1998). G protein-Press). coupled receptor kinases. Annu. Rev. Biochem. 67, 653–692.
Hyman, S.E., and Malenka, R.C. (2001). Addiction and the brain: the Premont, R.T., Inglese, J., and Lefkowitz, R.J. (1995). Protein kinasesneurobiology of compulsion and its persistence. Nat. Rev. Neurosci. that phosphorylate activated G protein-coupled receptors. FASEB2, 695–703. J. 9, 175–182.
Ito, K., Haga, T., Lameh, J., and Sadee, W. (1999). Sequestration of Premont, R.T., Macrae, A.D., Aparicio, S.A., Kendall, H.E., Welch,dopamine D2 receptors depends on coexpression of G-protein- J., and Lefkowitz, R.J. (1999). The GRK4 subfamily of G protein-coupled receptor kinases 2 or 5. Eur. J. Biochem. 260, 112–119. coupled receptor kinases: alternative splicing, gene organization
and sequence conservation. J. Biol. Chem. 274, 29381–29389.Jaber, M., Koch, W.J., Rockman, H., Smith, B., Bond, R.A., Sulik,K.K., Ross, J., Jr., Lefkowitz, R.J., Caron, M.G., and Giros, B. (1996). Premont, R.T., Gainetdinov, R.R., and Caron, M.G. (2001). FollowingEssential role of beta-adrenergic receptor kinase 1 in cardiac devel- the trace of elusive amines. Proc. Natl. Acad. Sci. USA 98, 9474–opment and function. Proc. Natl. Acad. Sci. USA 93, 12974–12979. 9475.
Janssen, P.A., Leysen, J.E., Megens, A.A., and Awouters, F.H. (1999). Rivera, A., Cuellar, B., Giron, F.J., Grandy, D.K., de la Calle, A., andDoes phenylethylamine act as an endogenous amphetamine in Moratalla, R. (2002). Dopamine D4 receptors are heterogeneouslysome patients? Int. J. Neuropsychopharmacol. 2, 229–240. distributed in the striosomes/matrix compartments of the striatum.
J. Neurochem. 80, 219–229.Jenner, P., and Marsden, C.D. (1987). Chronic pharmacological ma-nipulation of dopamine receptors in brain. Neuropharmacology 26, Robinson, T.E., and Berridge, K.C. (1993). The neural basis of drug931–940. craving: an incentive-sensitization theory of addiction. Brain Res.
Brain Res. Rev. 18, 247–291.Jones, S.R., Gainetdinov, R.R., Wightman, R.M., and Caron, M.G.(1998). Mechanisms of amphetamine action revealed in mice lacking Rockman, H.A., Choi, D.J., Akhter, S.A., Jaber, M., Giros, B., Lefko-the dopamine transporter. J. Neurosci. 18, 1979–1986. witz, R.J., Caron, M.G., and Koch, W.J. (1998). Control of myocardial
contractile function by the level of beta-adrenergic receptor kinaseKent, R.S., De Lean, A., and Lefkowitz, R.J. (1980). A quantitative1 in gene-targeted mice. J. Biol. Chem. 273, 18180–18184.analysis of beta-adrenergic receptor interactions: resolution of high
and low affinity states of the receptor by computer modeling of Sandoval, V., Riddle, E.L., Ugarte, Y.V., Hanson, G.R., and Fleck-enstein, A.E. (2001). Methamphetamine-induced rapid and revers-ligand binding data. Mol. Pharmacol. 17, 14–23.

Role of GRKs in the Responsiveness to Cocaine303
ible changes in dopamine transporter function: an in vitro model. J.Neurosci. 21, 1413–1419.
Schwartz, J.C., Levesque, D., Martres, M.P., and Sokoloff, P. (1993).Dopamine D3 receptor: basic and clinical aspects. Clin. Neurophar-macol. 16, 295–314.
Seeman, P., and Van Tol, H.H. (1994). Dopamine receptor pharma-cology. Trends Pharmacol. Sci. 15, 264–270.
Sibley, D.R. (1999). New insights into dopaminergic receptor func-tion using antisense and genetically altered animals. Annu. Rev.Pharmacol. Toxicol. 39, 313–341.
Singer, H.S. (1994). Neurobiological issues in Tourette syndrome.Brain Dev. 16, 353–364.
Stoffel, R.H., Randall, R.R., Premont, R.T., Lefkowitz, R.J., and In-glese, J. (1994). Palmitoylation of G protein-coupled receptor kinase,GRK6. Lipid modification diversity in the GRK family. J. Biol. Chem.269, 27791–27794.
Tiberi, M., Nash, S.R., Bertrand, L., Lefkowitz, R.J., and Caron, M.G.(1996). Differential regulation of dopamine D1A receptor respon-siveness by various G protein-coupled receptor kinases. J. Biol.Chem. 271, 3771–3778.
Usiello, A., Baik, J.H., Rouge-Pont, F., Picetti, R., Dierich, A., LeMeur,M., Piazza, P.V., and Borrelli, E. (2000). Distinct functions of the twoisoforms of dopamine D2 receptors. Nature 408, 199–203.
Van Hartesveldt, C. (1997). Temporal and environmental effects onquinpirole-induced biphasic locomotion in rats. Pharmacol. Bio-chem. Behav. 58, 955–960.
Walters, J.R., Bergstrom, D.A., Carlson, J.H., Chase, T.N., and Braun,A.R. (1987). D1 dopamine receptor activation required for postsyn-aptic expression of D2 agonist effects. Science 236, 719–722.
Walker, J.K., Peppel, K., Lefkowitz, R.J., Caron, M.G., and Fisher,J.T. (1999). Altered airway and cardiac responses in mice lacking Gprotein-coupled receptor kinase 3. Am. J. Physiol. 276, R1214–R1221.
Wang, Y.M., Gainetdinov, R.R., Fumagalli, F., Xu, F., Jones, S.R.,Bock, C.B., Miller, G.W., Wightman, R.M., and Caron, M.G. (1997).Knockout of the vesicular monoamine transporter 2 gene results inneonatal death and supersensitivity to cocaine and amphetamine.Neuron 19, 1285–1296.
Willner, P. (1997). The mesolimbic dopamine system as a target forrapid antidepressant action. Int. Clin. Psychopharmacol. 12 (Suppl3), S7–S14.
Wise, R.A., and Rompre, P.P. (1989). Brain dopamine and reward.Annu. Rev. Psychol. 40, 191–225.
White, F.J., Bednarz, L.M., Wachtel, S.R., Hjorth, S., and Brooderson,R.J. (1988). Is stimulation of both D1 and D2 receptors necessaryfor the expression of dopamine-mediated behaviors? Pharmacol.Biochem. Behav. 30, 189–193.
Xu, M., Koeltzow, T.E., Santiago, G.T., Moratalla, R., Cooper, D.C.,Hu, X.T., White, N.M., Graybiel, A.M., White, F.J., and Tonegawa,S. (1997). Dopamine D3 receptor mutant mice exhibit increasedbehavioral sensitivity to concurrent stimulation of D1 and D2 recep-tors. Neuron 19, 837–848.
Xu, F., Gainetdinov, R.R., Wetsel, W.C., Jones, S.R., Bohn, L.M.,Miller, G.W., Wang, Y.M., and Caron, M.G. (2000). Mice lacking thenorepinephrine transporter are supersensitive to psychostimulants.Nat. Neurosci. 3, 465–471.
Zhuang, X., Oosting, R.S., Jones, S.R., Gainetdinov, R.R., Miller,G.W., Caron, M.G., and Hen, R. (2001). Hyperactivity and impairedresponse habituation in hyperdopaminergic mice. Proc. Natl. Acad.Sci. USA 98, 1982–1987.
Related Documents











![[18F]Fluorodopa PETshows striatal dopaminergic dysfunction ...](https://static.cupdf.com/doc/110x72/628e71a806be7c7a267428b6/18ffluorodopa-petshows-striatal-dopaminergic-dysfunction-.jpg)