Dopamine action in the substantia nigra pars reticulata: iontophoretic studies in awake, unrestrained rats Franc ¸ois Windels and Eugene A. Kiyatkin Cellular Neurobiology Branch, National Institute on Drug Abuse ) Intramural Research Program, National Institutes of Health, DHHS, 333 Cassell Drive, Baltimore, MD 21224, USA Keywords: dendritic release, freely moving rats, iontophoresis, presynaptic modulation, single-unit recording Abstract Dopamine (DA) neurons located in the substantia nigra pars compacta release DA not only via axonal terminals, affecting neurotransmission within the striatum, but also via dendrites, some of which densely protrude into the substantia nigra pars reticulata (SNr). Although the interaction of dendritically released DA with somatodendritic autoreceptors regulates DA cell activity, released DA may also affect SNr neurons. These cells, however, lack postsynaptic DA receptors, making it unclear how locally released DA modulates their activity. Although previous work in brain slices suggests that DA might modulate the activity of GABA inputs, thus affecting SNr neurons indirectly, it remains unclear how increased or decreased DA release might affect these cells exposed to normal afferent inputs. To explore this issue, we examined the effects of iontophoretic DA and amphetamine on SNr neurons in awake, unrestrained rats. DA had no consistent effects on SNr cells but amphetamine, known to induce DA release, dose- dependently inhibited most of them. This effect was blocked by SCH23390, a selective D1 receptor blocker, which itself strongly increased neuronal discharge rate. As GABA input is a major factor regulating the activity of SNr neurons, our data suggest that dendritically released DA, by interacting with D1 receptors on striato-nigral and pallido-nigral afferents, is able to decrease this input, thus releasing SNr neurons from tonic, GABA-mediated inhibition. Surprisingly, a full DA receptor blockade (SCH23390 + eticlopride) did not result in the expected increase in SNr discharge rate, suggesting that other mechanisms are responsible for behavioral abnormalities following acute disruption of DA transmission. Introduction Midbrain dopamine (DA) neurons release DA not only via axonal terminals, affecting neurotransmission within their target areas, but also via dendrites (Nieoullon et al., 1977), affecting their neighbor- ing environment. Although the importance of axonally released DA in modulating neural transmission is well appreciated, the functional role of dendritic DA release remains less clear. As most DA neurons possess somatodendritic autoreceptors, dendritically released DA, by interacting with these receptors, may play a crucial role in self- regulating their activity (Groves et al., 1975; Aghajanian & Bunney, 1977). Because of this arrangement, most DA neurons have a low and stable discharge rate in various environmental conditions (Steinfels et al., 1983; Bunney et al., 1991) and show weak, essentially biphasic responses to phasic excitatory inputs (Strecker & Jacobs, 1985; Schultz, 1986; Kiyatkin, 1988). In contrast to the ventral tegmental area, where DA neurons are intermixed with GABA cells, most substantia nigra DA cells are located in the substantia nigra pars compacta with their dendrites extending ventrally into the substantia nigra pars reticulata (SNr), which is composed primarily of GABA projection neurons (Bjo ¨rklund & Lindvall, 1984; Fallon & Loughlin, 1995; Gonzalez-Hernandez & Rodriguez, 2000). This anatomical arrangement makes it possible for dendritically released DA to interact with SNr neurons, thus affecting the activity of this key output structure of the basal ganglia. However, the mechanisms mediating the action of DA on SNr neurons remain unclear. Early attempts to examine the effects of DA on SNr neurons produced conflicting results, with both weak excitations and inhibitions as a response to iontophoretic DA (Ruffieux & Schultz, 1981; Waszczak & Walters, 1983). These effects, moreover, were strongly dependent upon the anesthetic agent used (Ruffieux & Schultz, 1981) and they could not be easily characterized because subsequent work revealed that SNr neurons lack postsynaptic DA receptors (Yung et al., 1995). D1 DA receptors, however, were identified on GABA (Yung et al., 1995) and presumably glutamate (GLU) terminals (Rosales et al., 1997) synapsing on SNr neurons. Therefore, the effects of DA on SNr neurons may be indirect, resulting from presynaptic modulation of GABA and ⁄ or GLU inputs, the two opposing forces regulating the activity of these cells. Although this presynaptic mechanism of DA action on SNr cells has been confirmed in vitro with respect to both GLU (Wittmann et al., 2002) and GABA (Miyazaki & Lacey, 1998; Radnikow & Misgeld, 1998) afferents, these findings were obtained from deafferentated and pharmacologi- cally treated cells at room temperature, usually with evoked activity and using DA agonists and antagonists or DA at large doses. These in vitro findings, moreover, are controversial. Radnikow & Misgeld (1998), for example, suggested that somatodendritically released DA acting via presynaptic D1 receptors on striato-nigral afferents might enhance GABA release but Miyazaki & Lacey (1998), based on their findings, suggested that DA decreases evoked GABA release via the same mechanism. Therefore, it remains unclear what type of modulation occurs in the whole intact brain, when neurons are Correspondence: Dr Eugene A. Kiyatkin, as above. E-mail: [email protected] Received 21 March 2006, revised 5 June 2006, accepted 26 June 2006 European Journal of Neuroscience, Vol. 24, pp. 1385–1394, 2006 doi:10.1111/j.1460-9568.2006.05015.x ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Dopamine action in the substantia nigra pars reticulata:iontophoretic studies in awake, unrestrained rats
Francois Windels and Eugene A. KiyatkinCellular Neurobiology Branch, National Institute on Drug Abuse ) Intramural Research Program, National Institutes of Health,DHHS, 333 Cassell Drive, Baltimore, MD 21224, USA
Keywords: dendritic release, freely moving rats, iontophoresis, presynaptic modulation, single-unit recording
Abstract
Dopamine (DA) neurons located in the substantia nigra pars compacta release DA not only via axonal terminals, affectingneurotransmission within the striatum, but also via dendrites, some of which densely protrude into the substantia nigra pars reticulata(SNr). Although the interaction of dendritically released DA with somatodendritic autoreceptors regulates DA cell activity, released DAmay also affect SNr neurons. These cells, however, lack postsynaptic DA receptors, making it unclear how locally released DAmodulates their activity. Although previous work in brain slices suggests that DA might modulate the activity of GABA inputs, thusaffecting SNr neurons indirectly, it remains unclear how increased or decreased DA release might affect these cells exposed tonormal afferent inputs. To explore this issue, we examined the effects of iontophoretic DA and amphetamine on SNr neurons inawake, unrestrained rats. DA had no consistent effects on SNr cells but amphetamine, known to induce DA release, dose-dependently inhibited most of them. This effect was blocked by SCH23390, a selective D1 receptor blocker, which itself stronglyincreased neuronal discharge rate. As GABA input is a major factor regulating the activity of SNr neurons, our data suggest thatdendritically released DA, by interacting with D1 receptors on striato-nigral and pallido-nigral afferents, is able to decrease this input,thus releasing SNr neurons from tonic, GABA-mediated inhibition. Surprisingly, a full DA receptor blockade (SCH23390 + eticlopride)did not result in the expected increase in SNr discharge rate, suggesting that other mechanisms are responsible for behavioralabnormalities following acute disruption of DA transmission.
Introduction
Midbrain dopamine (DA) neurons release DA not only via axonalterminals, affecting neurotransmission within their target areas, butalso via dendrites (Nieoullon et al., 1977), affecting their neighbor-ing environment. Although the importance of axonally released DAin modulating neural transmission is well appreciated, the functionalrole of dendritic DA release remains less clear. As most DA neuronspossess somatodendritic autoreceptors, dendritically released DA, byinteracting with these receptors, may play a crucial role in self-regulating their activity (Groves et al., 1975; Aghajanian & Bunney,1977). Because of this arrangement, most DA neurons have a lowand stable discharge rate in various environmental conditions(Steinfels et al., 1983; Bunney et al., 1991) and show weak,essentially biphasic responses to phasic excitatory inputs (Strecker &Jacobs, 1985; Schultz, 1986; Kiyatkin, 1988). In contrast to theventral tegmental area, where DA neurons are intermixed withGABA cells, most substantia nigra DA cells are located in thesubstantia nigra pars compacta with their dendrites extendingventrally into the substantia nigra pars reticulata (SNr), which iscomposed primarily of GABA projection neurons (Bjorklund &Lindvall, 1984; Fallon & Loughlin, 1995; Gonzalez-Hernandez &Rodriguez, 2000).
This anatomical arrangement makes it possible for dendriticallyreleased DA to interact with SNr neurons, thus affecting the activity of
this key output structure of the basal ganglia. However, themechanisms mediating the action of DA on SNr neurons remainunclear. Early attempts to examine the effects of DA on SNr neuronsproduced conflicting results, with both weak excitations andinhibitions as a response to iontophoretic DA (Ruffieux & Schultz,1981; Waszczak & Walters, 1983). These effects, moreover, werestrongly dependent upon the anesthetic agent used (Ruffieux &Schultz, 1981) and they could not be easily characterized becausesubsequent work revealed that SNr neurons lack postsynaptic DAreceptors (Yung et al., 1995). D1 DA receptors, however, wereidentified on GABA (Yung et al., 1995) and presumably glutamate(GLU) terminals (Rosales et al., 1997) synapsing on SNr neurons.Therefore, the effects of DA on SNr neurons may be indirect, resultingfrom presynaptic modulation of GABA and ⁄ or GLU inputs, the twoopposing forces regulating the activity of these cells. Although thispresynaptic mechanism of DA action on SNr cells has been confirmedin vitro with respect to both GLU (Wittmann et al., 2002) and GABA(Miyazaki & Lacey, 1998; Radnikow & Misgeld, 1998) afferents,these findings were obtained from deafferentated and pharmacologi-cally treated cells at room temperature, usually with evoked activityand using DA agonists and antagonists or DA at large doses. Thesein vitro findings, moreover, are controversial. Radnikow & Misgeld(1998), for example, suggested that somatodendritically released DAacting via presynaptic D1 receptors on striato-nigral afferents mightenhance GABA release but Miyazaki & Lacey (1998), based on theirfindings, suggested that DA decreases evoked GABA release via thesame mechanism. Therefore, it remains unclear what type ofmodulation occurs in the whole intact brain, when neurons are
Correspondence: Dr Eugene A. Kiyatkin, as above.E-mail: [email protected]
Received 21 March 2006, revised 5 June 2006, accepted 26 June 2006
European Journal of Neuroscience, Vol. 24, pp. 1385–1394, 2006 doi:10.1111/j.1460-9568.2006.05015.x
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd

exposed to natural afferent inputs and animals are under physiolog-ically relevant conditions, and what the resulting consequences forSNr neuronal activity may be.To clarify this issue, single-unit recording was combined with
iontophoresis in awake, unrestrained rats. Taking into account theprofound effects of general anesthesia on impulse activity andresponses of SNr neurons (Windels & Kiyatkin, 2004), we firsttested the effects of iontophoretic DA on these cells and comparedthese effects with those induced by DA in chloral hydrate-anesthetized animals. As iontophoretically applied DA rapidlyundergoes uptake and metabolic destruction, we then usedamphetamine (AMPH) as a tool to induce endogenous DA releaseand examine the neuronal response to this ‘over-DA’ intervention.As DA may act on D1 receptors located on presynaptic GABAterminals, we also tested the effects of SCH23390, a selective D1-like DA antagonist, on the impulse activity of SNr neurons and theirresponses to iontophoretic AMPH. This test was essential to verifythe role of DA in mediating the neuronal effects of AMPH, examinethe role of DA in modulating GABA input and clarify thecontribution of this input to the activity state of SNr neurons.Finally, using a mixture of SCH23390 and eticlopride, a selectiveD2-like DA antagonist, we evaluated how an acute blockade of DAtransmission affects the activity of SNr neurons.
Materials and methods
Animals and surgery
Data were obtained from 33 male Long-Evans rats (400 ± 50 g)supplied by Charles River Laboratories (Greensboro, NC, USA). Allanimals were housed individually under standard laboratory condi-tions (12-h light cycle beginning at 07:00 h) with free access to foodand water. Protocols were performed in compliance with the Guidefor the Care and Use of Laboratory Animals (NIH, Publication 865-23) and were approved by the National Institute on DrugAbuse ) Intramural Research Program Animal Care and UseCommittee. The surgery procedures used have been describedpreviously (Windels & Kiyatkin, 2004). Briefly, under generalanesthesia (Equithesin 0.33 mL/100 g i.p.; dose of sodium pento-barbital 32.5 mg/kg and chloral hydrate 145 mg/kg), rats wereimplanted with a plastic, cylindrical hub, designed to mate with amicroelectrode holder (Rebec et al., 1993) during recording. Thishub was centred over a hole drilled above the substantia nigra(4.8–6.0 mm posterior and 1.4–2.6 mm lateral to the bregma). Aftera 3–4-day period of recovery and habituation to the experimentalchamber, recording sessions were held once daily over the next1–3 days. A separate group of rats (n ¼ 3), prepared as describedabove, underwent a recording session under chloral hydrateanesthesia (400 mg/kg, i.p. followed by 120 mg/kg/h). In theseexperiments body temperature was maintained automatically at37.2 ± 0.2 �C with an electric heating pad and feedback rectalthermal probe.In some experiments, animals were administered with either a D1-
like DA receptor blocker (SCH23390; 1 mg/kg, Sigma, St Louis, MO,USA) or a mixture of SCH23390 with eticlopride, a D2-like DAreceptor blocker (1 mg/kg for each drug). These drugs were freshlydissolved in saline and injected subcutaneously. Neuronal dataobtained between 15 min and 3 h following the injection of DAantagonists were accepted as either D1 receptor blockade or full DAreceptor blockade as confirmed in our previous study (Kiyatkin &Rebec, 1999a) by the antagonism of striatal neuronal responses toiontophoretic DA.
Single-unit recording and iontophoresis
Four-barrelled, microfilament-filled glass pipettes (Omega Dot 50744,Stoelting, Wood Dale, IL, USA), pulled and broken to a diameter of5 ± 1 lm, were used for single-unit recording and iontophoresis. Therecording barrel contained 2% Pontamine sky blue (BDH ChemicalsLtd, Poole, UK) in 3 m NaCl and the balance barrel contained 0.25 m
NaCl. The remaining barrels were filled with solutions of d-AMPHsulfate (0.1 m in distilled water, pH 5.5, National Institute on DrugAbuse) and DA hydrochloride (0.25 m in distilled water, pH 4.5,Sigma). The resistance of the recording channel was 3–5 MW(measured at 100 Hz) and the drug-containing barrels ranged between10 and 35 MW. Retaining ()8 to )10 nA) and ejecting (+10–60 nA)currents were applied with a constant current generator (Ion 100T,Dagan, Minneapolis, MN, USA). Each multibarrel pipette was filledwith fresh solution less than 1 h before use and fixed in a microdriveassembly that was later inserted into the skull-mounted hub. Theelectrode was then advanced 7.8 mm below the skull surface to thestarting point of unit recording.Neuronal discharge signals were sent to a head-mounted preamplifier
(OPA 404KP, Burr Brown, Tucson, AZ, USA) and then additionallyamplified and filtered (band pass 300–3000 Hz) with a NeurologSystem (Digitimer, Hertfordshire, UK). The filtered signal was thenrecorded using a Micro 1401 MK2 interface (Cambridge ElectronicDesign, Cambridge, UK). Spike activity was monitored with a digitaloscilloscope and audio amplifier, and analysed using a Spike2 interface(Cambridge Electronic Design). After the isolation of single-unitdischarges and identification of the spike waveform, data collection foreach neuron typically lasted 10–30 min. Our protocol typicallyincluded several 20-s applications of DA or AMPH performed at90–120-s intervals with either constant or increasing ejection currents.During the experiment the rat’s activity was recorded using a wide-angle camera (Creative Technology, Milpitas, CA, USA). All iontoph-oretic applications used for statistical analysis were performed when theanimals were at rest with no sign of overt movements.
Histology
After the last recording session, animals were deeply anesthetized withEquithesin and Pontamine sky blue was deposited by current injection()20 lA for 20 min) at the last recording site. Rats were perfusedtranscardially with saline solution followed by 4% paraformaldehydesolution. The brain was removed and placed overnight at 4 �C in a20% sucrose solution and then frozen in dry ice. Coronal 30-lm tissuesections were prepared at )20 �C using a microtome cryostat. Theatlas of Paxinos & Watson (1998) served as the basis for histologicalanalyses.
Immunohistochemistry
In some experiments, a precise anatomic differentiation of thesubstantia nigra pars compacta and SNr was needed to establish thelocation of the cells recorded. We therefore performed an immunos-taining of tyrosine hydroxylase, a DA synthesis enzyme abundant inthe pars compacta but not the pars reticulata. Sections were rinsedtwice in phosphate buffer before being incubated for 60 min inphosphate buffer with 0.3% triton and 4% bovine serum albumin.Tissue sections were then incubated overnight at 4 �C in buffer withmonoclonal mouse tyrosine hydroxylase antibody (dilution 1/500,Chemicon International, Temecula, CA, USA). Slices were rinsed intwo buffer baths (10 min each), transferred in a solution of fluorescein-conjugated anti-mouse antibody (1/500, Jackson ImmunoResearch,
1386 F. Windels and E. A. Kiyatkin
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394
West Grove, PA,USA) for 2 h, rinsed four times in buffer again and thenmounted on gelatin-coated slides.
Data analysis
The impulse activity of individual SNr neurons was characterized bythe mean rate based on 20 1-s values of discharge rate. These valueswere grouped together and further analysed by using standardstatistical procedures (i.e. mean and modal group values, variabilityand distributions). As discharge rates of individual units in otherstructures (i.e. medial thalamus, striatum and ventral tegmental area)are distributed based on the ln-normal law (Werner & Mountcastle,1963; Nakahama et al., 1971; Kiyatkin, 1983; Kiyatkin & Rebec,1998, 1999b), this type of distribution pattern was verified for SNrneurons by using ln-transformation of statistical parameters of impulseactivity.
Each iontophoretic test was statistically evaluated and the responsewas accepted (i.e. excitation or inhibition) if the mean firing rateduring iontophoresis differed significantly (P < 0.05; two-tailedStudent’s t-test) from an equivalent period of baseline activityimmediately preceding the iontophoretic application. These responseswere also assessed in terms of absolute and relative magnitude andonset and offset latencies. As the duration of each neuronal recordingin freely moving rats varied from 5 to 30 min and the testing programfor each unit was different, it was impossible to assess responsethresholds and dose–response relationships in each individual unit.Therefore, our data are reported as the number of both units andiontophoretic responses. Various relationships between impulse activ-
ity and iontophoretic responses were assessed with Student’s t-testsand anova that were followed by a Fisher posthoc test.
Results
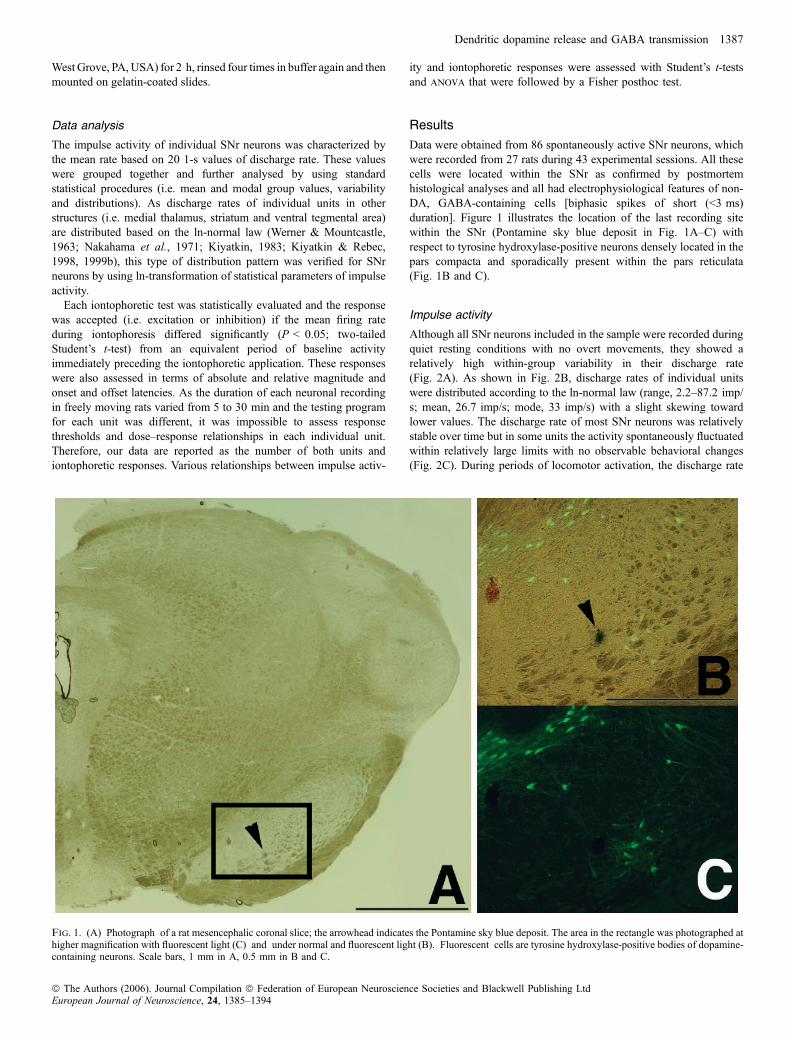
Data were obtained from 86 spontaneously active SNr neurons, whichwere recorded from 27 rats during 43 experimental sessions. All thesecells were located within the SNr as confirmed by postmortemhistological analyses and all had electrophysiological features of non-DA, GABA-containing cells [biphasic spikes of short (<3 ms)duration]. Figure 1 illustrates the location of the last recording sitewithin the SNr (Pontamine sky blue deposit in Fig. 1A–C) withrespect to tyrosine hydroxylase-positive neurons densely located in thepars compacta and sporadically present within the pars reticulata(Fig. 1B and C).
Impulse activity
Although all SNr neurons included in the sample were recorded duringquiet resting conditions with no overt movements, they showed arelatively high within-group variability in their discharge rate(Fig. 2A). As shown in Fig. 2B, discharge rates of individual unitswere distributed according to the ln-normal law (range, 2.2–87.2 imp/s; mean, 26.7 imp/s; mode, 33 imp/s) with a slight skewing towardlower values. The discharge rate of most SNr neurons was relativelystable over time but in some units the activity spontaneously fluctuatedwithin relatively large limits with no observable behavioral changes(Fig. 2C). During periods of locomotor activation, the discharge rate
Fig. 1. (A) Photograph of a rat mesencephalic coronal slice; the arrowhead indicates the Pontamine sky blue deposit. The area in the rectangle was photographed athigher magnification with fluorescent light (C) and under normal and fluorescent light (B). Fluorescent cells are tyrosine hydroxylase-positive bodies of dopamine-containing neurons. Scale bars, 1 mm in A, 0.5 mm in B and C.
Dendritic dopamine release and GABA transmission 1387
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394
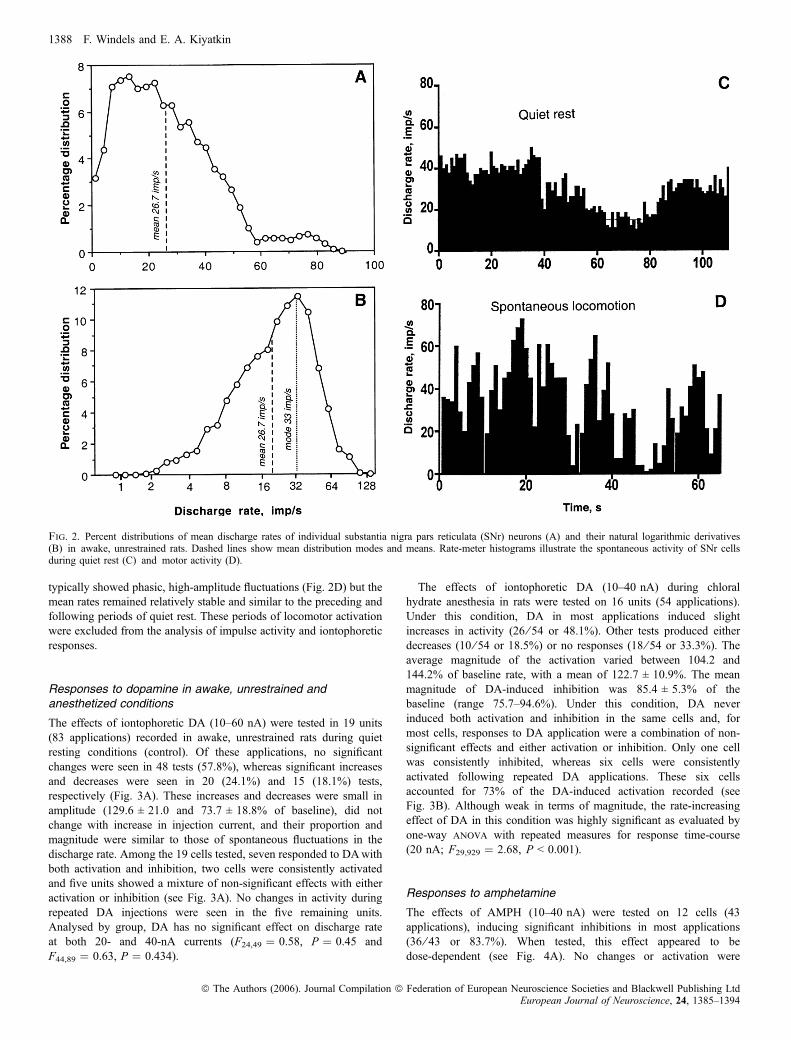
typically showed phasic, high-amplitude fluctuations (Fig. 2D) but themean rates remained relatively stable and similar to the preceding andfollowing periods of quiet rest. These periods of locomotor activationwere excluded from the analysis of impulse activity and iontophoreticresponses.
Responses to dopamine in awake, unrestrained andanesthetized conditions
The effects of iontophoretic DA (10–60 nA) were tested in 19 units(83 applications) recorded in awake, unrestrained rats during quietresting conditions (control). Of these applications, no significantchanges were seen in 48 tests (57.8%), whereas significant increasesand decreases were seen in 20 (24.1%) and 15 (18.1%) tests,respectively (Fig. 3A). These increases and decreases were small inamplitude (129.6 ± 21.0 and 73.7 ± 18.8% of baseline), did notchange with increase in injection current, and their proportion andmagnitude were similar to those of spontaneous fluctuations in thedischarge rate. Among the 19 cells tested, seven responded to DAwithboth activation and inhibition, two cells were consistently activatedand five units showed a mixture of non-significant effects with eitheractivation or inhibition (see Fig. 3A). No changes in activity duringrepeated DA injections were seen in the five remaining units.Analysed by group, DA has no significant effect on discharge rateat both 20- and 40-nA currents (F24,49 ¼ 0.58, P ¼ 0.45 andF44,89 ¼ 0.63, P ¼ 0.434).
The effects of iontophoretic DA (10–40 nA) during chloralhydrate anesthesia in rats were tested on 16 units (54 applications).Under this condition, DA in most applications induced slightincreases in activity (26 ⁄ 54 or 48.1%). Other tests produced eitherdecreases (10 ⁄ 54 or 18.5%) or no responses (18 ⁄ 54 or 33.3%). Theaverage magnitude of the activation varied between 104.2 and144.2% of baseline rate, with a mean of 122.7 ± 10.9%. The meanmagnitude of DA-induced inhibition was 85.4 ± 5.3% of thebaseline (range 75.7–94.6%). Under this condition, DA neverinduced both activation and inhibition in the same cells and, formost cells, responses to DA application were a combination of non-significant effects and either activation or inhibition. Only one cellwas consistently inhibited, whereas six cells were consistentlyactivated following repeated DA applications. These six cellsaccounted for 73% of the DA-induced activation recorded (seeFig. 3B). Although weak in terms of magnitude, the rate-increasingeffect of DA in this condition was highly significant as evaluated byone-way anova with repeated measures for response time-course(20 nA; F29,929 ¼ 2.68, P < 0.001).
Responses to amphetamine
The effects of AMPH (10–40 nA) were tested on 12 cells (43applications), inducing significant inhibitions in most applications(36 ⁄ 43 or 83.7%). When tested, this effect appeared to bedose-dependent (see Fig. 4A). No changes or activation were
Fig. 2. Percent distributions of mean discharge rates of individual substantia nigra pars reticulata (SNr) neurons (A) and their natural logarithmic derivatives(B) in awake, unrestrained rats. Dashed lines show mean distribution modes and means. Rate-meter histograms illustrate the spontaneous activity of SNr cellsduring quiet rest (C) and motor activity (D).
1388 F. Windels and E. A. Kiyatkin
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394

observed during five (11.62%) and two (4.65%) tests, respectively.The range of inhibitions at 40 nA varied from 32.6 to 89.7% with amean of 60.6 ± 15.5%. As shown in Fig. 4B, the discharge ratefollowing AMPH iontophoresis (40 nA, n ¼ 40) significantlydecreased (anova with repeated measures, F39,1239 ¼ 8.366;P < 0.001) with about 6 s onset latency and slowly restored tothe preapplication baseline within 8–10 s after the ejecting currentwas switched off.
Responses to amphetamine under D1 receptor blockade
The effects of AMPH during D1 receptor blockade were tested on 11cells (38 applications). Under this condition, no changes wereobserved in 26 tests (68.42%), whereas activations and inhibitionswere seen in two (5.26%) and 10 (26.3%) tests. Although inhibitionswere rare, their average magnitude varied between 34.7 and 88.9% ofthe baseline rate, with a mean of 70.9 ± 18.4% that was similar tothat seen in control conditions (60.6 ± 15.5%; see above). As shownin Fig. 4C, under D1 receptor blockade, AMPH (40 nA, n ¼ 37) hadno significant effect on discharge rate (anova with repeated measure,F37,1146 ¼ 1.33, P ¼ 0.11). Between-group differences were equallystrong as both absolute and relative change in discharge rate(Fig. 4D).
Changes in substantia nigra pars reticulata discharge ratefollowing SCH23390 and SCH23390 + eticlopride
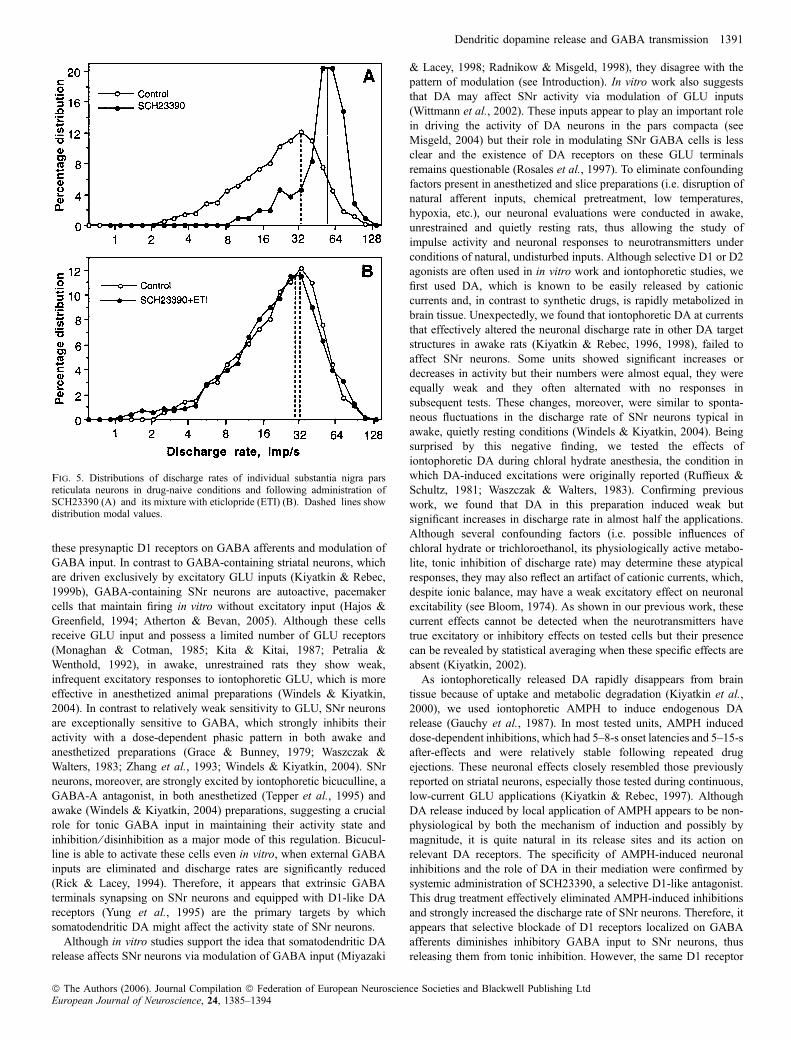
The SNr units recorded after SCH23390 administration had asignificantly higher discharge rate (range, 21.5–111.7 imp ⁄ s; mean,59.57 imp ⁄ s) than in control conditions (range, 2.20–87.36 imp ⁄ s;mean, 26.69 imp ⁄ s; t ¼ 8.91, P < 0.001; compare baselines inFig. 4B and C). The coefficient of variation of impulse activity, anindex of relative regularity of impulse flow, after SCH23390 wassignificantly lower (range, 5.0–49.8%; mean, 17.67%) than in thecontrol group (range, 5.6–139.9%; mean, 28.66%; t ¼ 2.43,P < 0.05). This difference, however, was much less pronouncedthan the two-fold difference in discharge rate. As shown in Fig. 5A,the distribution of SNr discharge rates following D1 receptorblockade was quite different from that in control, drug-freeconditions, with an almost two-fold increase of modal values (55vs. 33 imp/s).The effects of full DA receptor blockade (SCH23390 + eticlopride
at 1 mg ⁄ kg each) on SNr discharge rates were tested in 28 neurons (11rats). In contrast to SCH23390 alone, the mean discharge rate(25.80 imp ⁄ s) and its distribution following this treatment (seeFig. 5B) were surprisingly similar to those in the control, drug-naivecondition (26.69 imp ⁄ s). The modal values of distributions were alsoalmost superimposable (31 vs. 32 imp ⁄ s).
Fig. 3. Rate-meter histogram illustrating responses of an individual substantia nigra pars reticulata cells to iontophoretic dopamine in awake unrestrained rat(A) and during chloral hydrate anesthesia (B). Numbers above horizontal lines refer to ejection current (nA). *Significant differences (P < 0.05; Student’s t-test).
Dendritic dopamine release and GABA transmission 1389
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394

Discussion
Although it is known that DA released from the dendrites of DA-containing substantia nigra pars compacta cells plays a crucial role intheir self-regulation (Groves et al., 1975; Aghajanian & Bunney,1977), the role of somatodendritic DA release in regulating SNractivity in both physiological and pathological conditions remainsunclear. Early work in anesthetized animals revealed that iontophoreticDA often increases the discharge rate of SNr neurons (Ruffieux &
Schultz, 1981; Waszczak & Walters, 1983) but the nature of theseresponses is difficult to reconcile with the lack of postsynaptic DAreceptors on these cells (Yung et al., 1995). D1-like DA receptors,however, were found on striato-nigral and pallido-nigral GABAafferents (Yung et al., 1995) that provide a massive inhibitory input toSNr neurons (Smith & Bolam, 1989; Aceves et al., 1995). Muchneuropharmacological evidence (Gauchy et al., 1987; Timmerman &Abercrombie, 1996; Trevitt et al., 2001, 2002; Correa et al., 2003)suggests that the action of DA on SNr neurons is indirect, involving
Fig. 4. (A) Rate-meter histogram illustrating responses of an individual cell to iontophoretic amphetamine (AMPH) at 40 and 20 nA (*significant inhibition). Peri-stimulus time histogram of all AMPH applications at 40 nA in control condition (B) (n ¼ 40) and after systemic injection of SCH23390 (C) (n ¼ 37); horizontalline represents the 20-s duration of iontophoretic application. Filled bars indicate values significantly lower than pre-AMPH baseline (P < 0.05). (D) Differences inAMPH-induced responses of substantia nigra pars reticulata neurons shown as a change relative to preapplication baseline (¼ 100%). Filled circles indicate valuessignificantly lower than pre-AMPH baseline (P < 0.05).
1390 F. Windels and E. A. Kiyatkin
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394
these presynaptic D1 receptors on GABA afferents and modulation ofGABA input. In contrast to GABA-containing striatal neurons, whichare driven exclusively by excitatory GLU inputs (Kiyatkin & Rebec,1999b), GABA-containing SNr neurons are autoactive, pacemakercells that maintain firing in vitro without excitatory input (Hajos &Greenfield, 1994; Atherton & Bevan, 2005). Although these cellsreceive GLU input and possess a limited number of GLU receptors(Monaghan & Cotman, 1985; Kita & Kitai, 1987; Petralia &Wenthold, 1992), in awake, unrestrained rats they show weak,infrequent excitatory responses to iontophoretic GLU, which is moreeffective in anesthetized animal preparations (Windels & Kiyatkin,2004). In contrast to relatively weak sensitivity to GLU, SNr neuronsare exceptionally sensitive to GABA, which strongly inhibits theiractivity with a dose-dependent phasic pattern in both awake andanesthetized preparations (Grace & Bunney, 1979; Waszczak &Walters, 1983; Zhang et al., 1993; Windels & Kiyatkin, 2004). SNrneurons, moreover, are strongly excited by iontophoretic bicuculline, aGABA-A antagonist, in both anesthetized (Tepper et al., 1995) andawake (Windels & Kiyatkin, 2004) preparations, suggesting a crucialrole for tonic GABA input in maintaining their activity state andinhibition ⁄ disinhibition as a major mode of this regulation. Bicucul-line is able to activate these cells even in vitro, when external GABAinputs are eliminated and discharge rates are significantly reduced(Rick & Lacey, 1994). Therefore, it appears that extrinsic GABAterminals synapsing on SNr neurons and equipped with D1-like DAreceptors (Yung et al., 1995) are the primary targets by whichsomatodendritic DA might affect the activity state of SNr neurons.
Although in vitro studies support the idea that somatodendritic DArelease affects SNr neurons via modulation of GABA input (Miyazaki
& Lacey, 1998; Radnikow & Misgeld, 1998), they disagree with thepattern of modulation (see Introduction). In vitro work also suggeststhat DA may affect SNr activity via modulation of GLU inputs(Wittmann et al., 2002). These inputs appear to play an important rolein driving the activity of DA neurons in the pars compacta (seeMisgeld, 2004) but their role in modulating SNr GABA cells is lessclear and the existence of DA receptors on these GLU terminalsremains questionable (Rosales et al., 1997). To eliminate confoundingfactors present in anesthetized and slice preparations (i.e. disruption ofnatural afferent inputs, chemical pretreatment, low temperatures,hypoxia, etc.), our neuronal evaluations were conducted in awake,unrestrained and quietly resting rats, thus allowing the study ofimpulse activity and neuronal responses to neurotransmitters underconditions of natural, undisturbed inputs. Although selective D1 or D2agonists are often used in in vitro work and iontophoretic studies, wefirst used DA, which is known to be easily released by cationiccurrents and, in contrast to synthetic drugs, is rapidly metabolized inbrain tissue. Unexpectedly, we found that iontophoretic DA at currentsthat effectively altered the neuronal discharge rate in other DA targetstructures in awake rats (Kiyatkin & Rebec, 1996, 1998), failed toaffect SNr neurons. Some units showed significant increases ordecreases in activity but their numbers were almost equal, they wereequally weak and they often alternated with no responses insubsequent tests. These changes, moreover, were similar to sponta-neous fluctuations in the discharge rate of SNr neurons typical inawake, quietly resting conditions (Windels & Kiyatkin, 2004). Beingsurprised by this negative finding, we tested the effects ofiontophoretic DA during chloral hydrate anesthesia, the condition inwhich DA-induced excitations were originally reported (Ruffieux &Schultz, 1981; Waszczak & Walters, 1983). Confirming previouswork, we found that DA in this preparation induced weak butsignificant increases in discharge rate in almost half the applications.Although several confounding factors (i.e. possible influences ofchloral hydrate or trichloroethanol, its physiologically active metabo-lite, tonic inhibition of discharge rate) may determine these atypicalresponses, they may also reflect an artifact of cationic currents, which,despite ionic balance, may have a weak excitatory effect on neuronalexcitability (see Bloom, 1974). As shown in our previous work, thesecurrent effects cannot be detected when the neurotransmitters havetrue excitatory or inhibitory effects on tested cells but their presencecan be revealed by statistical averaging when these specific effects areabsent (Kiyatkin, 2002).As iontophoretically released DA rapidly disappears from brain
tissue because of uptake and metabolic degradation (Kiyatkin et al.,2000), we used iontophoretic AMPH to induce endogenous DArelease (Gauchy et al., 1987). In most tested units, AMPH induceddose-dependent inhibitions, which had 5–8-s onset latencies and 5–15-safter-effects and were relatively stable following repeated drugejections. These neuronal effects closely resembled those previouslyreported on striatal neurons, especially those tested during continuous,low-current GLU applications (Kiyatkin & Rebec, 1997). AlthoughDA release induced by local application of AMPH appears to be non-physiological by both the mechanism of induction and possibly bymagnitude, it is quite natural in its release sites and its action onrelevant DA receptors. The specificity of AMPH-induced neuronalinhibitions and the role of DA in their mediation were confirmed bysystemic administration of SCH23390, a selective D1-like antagonist.This drug treatment effectively eliminated AMPH-induced inhibitionsand strongly increased the discharge rate of SNr neurons. Therefore, itappears that selective blockade of D1 receptors localized on GABAafferents diminishes inhibitory GABA input to SNr neurons, thusreleasing them from tonic inhibition. However, the same D1 receptor
Fig. 5. Distributions of discharge rates of individual substantia nigra parsreticulata neurons in drug-naive conditions and following administration ofSCH23390 (A) and its mixture with eticlopride (ETI) (B). Dashed lines showdistribution modal values.
Dendritic dopamine release and GABA transmission 1391
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394

blockade makes released DA ineffective at increasing GABA input,thus attenuating AMPH-induced neuronal inhibitions. Based on thesedata, it may be suggested that increased somatodendritic DA release,by increasing GABA input, will tend to decrease the activity of SNrneurons. Conversely, a decreased DA release, by decreasing GABAinput, will remove tonic inhibitory influence, thus inducing SNrhyperactivity.This pattern of DA action on SNr neurons has been suggested by
previous in vitro studies (Radnikow & Misgeld, 1998) and it issupported by experiments employing intra-SNr AMPH injections(Gauchy et al., 1987; Timmerman & Abercrombie, 1996) as well asintra-SNr injections of SCH23390 and bicuculline (Trevitt et al., 2001,2002). Intra-SNr administration of both AMPH and SKF82958, aselective D1 agonist, produced behavioral activation, which was fullyblocked by SCH23390 (Timmerman & Abercrombie, 1996) andbicuculline (Trevitt et al., 2002). In contrast, both SCH23390 andbicuculline, locally applied in the SNr, dose-dependently and withhigh potency decreased locomotor activity and inhibited food-reinforced operant behaviors (Trevitt et al., 2001; Correa et al., 2003).If the conclusion based on the effects of D1 receptor blocker is
applicable to DA, hyperactivity of SNr neurons may be expected tooccur when the action of DA at all relevant receptor sites is interruptedand these cells become acutely DA denervated. In contrast to ourexpectations, however, no changes in discharge rate of SNr neuronswere found following full DA receptor blockade, induced by a mixtureof SCH23390 and eticlopride at high doses. Although the reasons forthis unexpected stability of SNr discharge rate and its possible receptormechanisms will be discussed in our next report (Windels & Kiyatkin,2006), this finding as well as similar data obtained in anesthetized ratsboth with DA receptor blockade (Degos et al., 2005) and 6-hydroxy-dopamine-induced DA denervation (MacLeod et al., 1990) maysuggest a more complex picture of modulation of SNr activity bysomatodendritically released DA.
Functional implications and speculations
The SNr is usually considered a major output structure of the basalganglia, a complex of interconnected and inter-related structures,essential for sensorimotor integration and organization of behavior.Although GLU and GABA are the primary transmitters within thebasal ganglia, DA is known to be involved in modulating neurotrans-mission within the entire system of basal ganglia structures. Within thestriatum, axonally released DA appears to play an important role inmodulating neuronal responsiveness to excitatory, GLU-mediatedstimuli (for review see Nicola et al., 2000; Kiyatkin, 2002), thusshaping an inhibitory, GABA-mediated output signal to the pallidumand SNr. The same GABA-mediated signal, which is a primary inputfor SNr neurons, may also be modulated by dendritically released DAvia another presynaptic mechanism. Although the literature and ourpresent data suggest that DA acting via this mechanism may modulatethe efficiency of this functionally important GABA-mediated signal, itis unclear whether this modulation occurs following natural fluctua-tions in dendritic DA release under behavioral conditions and what itsrole might be. We still do not know how dendritic DA release isregulated and how extracellular DA levels fluctuate under physiolo-gical conditions. Microdialysis data suggest that DA levels in the SNror ventral tegmental area are much lower and their increases inducedby environmental stimuli or drugs are much weaker than in theirprimary target areas (Santiago & Westerink, 1992; Cobb &Abercrombie, 2003). These endogenous DA levels, moreover, are100 to 1000 times lower than DA concentrations that reliably affectinhibitory postsynaptic currents evaluated on SNr neurons in slice
preparations (Miyazaki & Lacey, 1998). Although it is evident that themodulatory action of DA may be especially important with respect tophasic GABA input, manifesting as a phasic fluctuation in SNractivity, it is quite difficult to reveal this action experimentally in thewhole brain. Based on recent data that full DA receptor blockadeattenuates the inhibitory component of the SNr evoked neuronalresponse induced by cortical stimulation (Degos et al., 2005), it maybe suggested that dendritic DA may potentiate the GABA inhibitorysignal, making phasic inhibitions of SNr neurons more pronounced. Itis known, however, that axonally released DA appears to enhance therelative magnitude of the GLU response of striatal neurons (Kiyatkin& Rebec, 1996) which, at the level of the SNr as a GABA input, is asubstrate of additional modulation by dendritic DA. Therefore, thetransmission of functionally important somatosensory signals foradequate motor output is the subject of multiple modulationsinvolving different neurotransmitters. At its input level, the cortico-striatal synapses, the modulatory action of DA involves its interactionwith GLU, whereas interaction with GABA appears to be the primaryissue at the output SNr level. As GABA-mediated inhibition ⁄ disin-hibition of SNr neurons is a critical event for movement regulation(Chevalier & Deniau, 1990), dendritically released DA, by modulatingGABA input, appears to be involved in both the phasic and tonicmodulations of their activity, thus affecting the initiation andperformance of movement acts under behavioral conditions.Although the literature and our present data suggest that SNr
neurons are inhibited following an AMPH-induced increase in DArelease and this mechanism, via decreased GABA input to premotornuclei (Chevalier & Deniau, 1990), may be important in mediatingmovement hyperactivity, SNr neuronal activity assessed in freelymoving rats did not follow the same pattern after systemic drugadministration (Olds, 1988; Gulley et al., 2004). In contrast toexpectations and data obtained in anesthetized animals (Bashore et al.,1978), many cells in behaving rats maintained high and fluctuatingdischarge rates with no evident decrease. This difference paralleledneuronal responses to AMPH in the striatum, where most units wereinhibited in anesthetized (Bashore et al., 1978) but clearly activated inawake, behaving conditions (Rebec et al., 1997). These differencesonce more stress the importance of the awake animal preparation inrevealing neuronal perturbations occurring in the whole brain underbehaviorally relevant conditions.Finally, diminished DA input to SNr neurons appears to be the
primary factor underlying behavioral abnormalities following DA celldegeneration. Although acute pharmacological interruption of DAtransmission cannot properly model the DA deficit resulting from slowand progressive loss of DA cells, behavioral changes in bothconditions are generally similar. According to traditional models ofbasal ganglia organization, both conditions should also result inhyperactivity of SNr neurons (Albin et al., 1989; DeLong, 1990). Thischange, however, was not found in previous studies in anesthetizedanimals after both 6-hydroxydopamine lesions of DA terminals(MacLeod et al., 1990) and acute pharmacological blockade of DAtransmission (Degos et al., 2005), and it was not shown in our presentstudy. Although this unexpected stability of SNr discharge ratesremains puzzling and we are unable to explain why selective D1antagonist and full DA receptor blockade induce similar behavioraldeficits but sharply different SNr neuronal outcomes, these results maysuggest that other mechanisms, not simple hyperactivity, are involvedin mediating behavioral abnormalities following the acute interruptionof DA transmission and progressive DA deficit associated withParkinson’s disease. Further studies using different models andexperimental techniques are necessary to clarify the mechanismsunderlying this clinically important human condition.
1392 F. Windels and E. A. Kiyatkin
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394

Acknowledgements
We thank Dr Barry Hoffer for valuable comments regarding this manuscriptand Paul Brown for editorial assistance. This research was supported by theIntramural Research Program of the NIH, National Institute on Drug Abuse.
Abbreviations
AMPH, amphetamine; DA, dopamine; GLU, glutamate; SNr, substantia nigrapars reticulata.
References
Aceves, J., Floran, B., Sierra, A. & Mariscal, S. (1995) D-1 receptor mediatedmodulation of the release of gamma-aminobutyric acid by endogenousdopamine in the basal ganglia of the rat. Prog. Neuropsychopharmacol. Biol.Psychiat., 19, 727–739.
Aghajanian, G.K. & Bunney, B.S. (1977) Dopamine ‘autoreceptors’: pharma-cological characterization by microiontophoretic single cell recordingstudies. Naunyn Schmiedeberg’s Arch. Pharmacol., 297, 1–7.
Albin, R.L., Young, A.B. & Penny, J.B. (1989) The functional anatomy of basalganglia disorders. Trends Neurosci., 12, 366–375.
Atherton, J.F. & Bevan, M.D. (2005) Ionic mechanisms underlyingautonomous action potential generation in the somata and dendrites ofGABAergic substantia nigra pars reticulata neurons in vitro. J. Neurosci., 25,8272–8281.
Bashore, T.R., Rebec, G.V. & Groves, P.M. (1978) Alterations of spontaneousneuronal activity in the caudate-putamen, nucleus accumbens and amygda-loid complex of rats produced by d-amphetamine. Pharmacol. Biochem.Behav., 8, 467–474.
Bjorklund, A. & Lindvall, O. (1984) Dopamine containing system. InBjorklund, A. & Hokfelt, T. (Eds), Handbook of Chemical Neuroanatomy.Elsevier, New York, pp. 55–122.
Bloom, F.E. (1974) To spritz or not to spritz: the doubtful value of aimlessiontophoresis. Life Sci., 14, 1819–1834.
Bloom, F.E., Costa, E. & Salmoiraghi, G.C. (1965) Anesthesia and theresponsiveness of individual neurons of the caudate nucleus of the cat toacetylcholine, norepinephrine and dopamine administered by microelec-trophoresis. J. Pharmacol. Exp. Ther., 150, 244–252.
Bunney, B.S., Chiodo, L.A. & Grace, A.A. (1991) Midbrain dopamine systemelectrophysiological functioning: a review and new hypothesis. Synapse, 9,79–94.
Chevalier, G. & Deniau, J.M. (1990) Disinhibition as a basic process in theexpression of striatal functions. Trends Neurosci., 13, 187–197.
Cobb, W.S. & Abercrombie, E.D. (2003) Differential regulation of somatoden-dritic and nerve terminal dopamine release by serotoninergic innervation ofsubstantia nigra. J. Neurochem., 84, 576–584.
Correa, M., Mingote, S., Betz, A., Wisniecki, A. & Salamone, J.D. (2003)Substantia nigra pars reticulata GABA is involved in the regulation ofoperant lever pressing: pharmacological and microdialysis studies. Neu-roscience, 119, 759–766.
Degos, B., Deniau, J.-M., Thierry, A.-M., Glowinski, J., Pezard, L. & Maurice,N. (2005) Neuroleptic-induced catalepsy; electrophysiological mechanismsof functional recovery induced by high-frequency stimulation of thesubthalamic nucleus. J. Neurosci., 25, 7687–7696.
DeLong, M.R. (1990) Primate models of movement disorders of basal gangliaorigin. Trends Neurosci., 13, 281–285.
Fallon, J.H. & Loughlin, S.E. (1995) Substantia nigra. In Paxinos, G. (Ed), TheRat Nervous System. Academic Press, San Diego, pp. 215–237.
Gauchy, C., Kemel, M.L., Desban, M., Romo, R., Glowinski, J. & Besson, M.J.(1987) The role of dopamine released from distal and proximal dendrites ofnigrostriatal dopaminergic neurons in the control of GABA transmission inthe thalamic nucleus ventralis medialis in the cat. Neuroscience, 22, 935–946.
Gonzalez-Hernandez, T. & Rodriguez, M. (2000) Compartmental organizationand chemical profile of dopaminergic and GABAergic neurons in thesubstantia nigra of the rat. J. Comp. Neurol., 421, 107–135.
Grace, A.A. & Bunney, B.S. (1979) Paradoxical GABA excitation of nigraldopaminergic cells; Indirect mediation through reticulata inhibitory neurons.Eur. J. Pharmacol., 59, 211–218.
Groves, P.M., Wilson, C.J., Young, S.J. & Rebec, G.V. (1975) Self-inhibitionby dopaminergic neurons. Science, 190, 522–528.
Gulley, J.M., Reed, J.L., Kuwajima, M. & Rebec, G.V. (2004) Amphetamine-induced behavioral activation is associated with variable changes in basal
ganglia output neurons recorded from awake, behaving rats. Brain Res.,1012, 108–118.
Hajos, M. & Greenfield, S.A. (1994) Synaptic connections between parscompacta and pars reticulata neurones: electrophysiological evidence forfunctional modules within the substantia nigra. Brain Res., 660, 216–224.
Kita, H. & Kitai, S.T. (1987) Efferent projections of the subthalamic nucleus inthe rat: light and electron microscopic analysis with the PHA-L method.J. Comp. Neurol., 260, 435–452.
Kiyatkin, E.A. (1983) Statistical analysis of impulse activity of diencephalicneurons during restraint stress in rats.Neurosci. Behav. Physiol., 13, 426–431.
Kiyatkin, E.A. (1988) Functional properties of presumed dopamine-containingand other ventral tegmental area neurons in conscious rats. Int. J. Neurosci.,42, 21–43.
Kiyatkin, E.A. (2002) Dopamine in the nucleus accumbens: cellular actions,drug- and behavior-associated fluctuations, and a possible role in anorganism’s adaptive activity. Behav. Brain. Res., 137, 27–46.
Kiyatkin, E.A. & Rebec, G.V. (1996) Dopaminergic modulation of glutamate-induced excitations of neurons in the neostriatum and nucleus accumbens ofawake, unrestrained rats. J. Neurophysiol., 75, 142–153.
Kiyatkin, E.A. & Rebec, G.V. (1997) Iontophoresis of amphetamine in theneostriatum and nucleus accumbens of awake, unrestrained rats. Brain Res.,771, 14–24.
Kiyatkin, E.A. & Rebec, G.V. (1998) Heterogeneity of ventral tegmental areaneurons: single-unit recording and iontophoresis in awake, unrestrained rats.Neuroscience, 85, 1285–1309.
Kiyatkin, E.A. & Rebec, G.V. (1999a) Striatal neuronal activity andresponsiveness to dopamine and glutamate after selective blockade of D1and D2 dopamine receptors in freely moving rats. J. Neurosci., 19, 3594–3609.
Kiyatkin, E.A. & Rebec, G.V. (1999b) Modulation of striatal neuronal activityby glutamate and GABA: iontophoresis in awake, unrestrained rats. BrainRes., 822, 88–106.
Kiyatkin, E.A., Kiyatkin, D.E. & Rebec, G.V. (2000) Phasic inhibition ofdopamine uptake in nucleus accumbens induced by intravenous cocaine infreely behaving rats. Neuroscience, 98, 729–741.
MacLeod, N.K., Ryman, A. & Arbuthnott, G.W. (1990) Electrophysiologicalproperties of nigrothalamic neurons after 6-hydroxydopamine lesions in therat. Neuroscience, 38, 447–456.
Misgeld, U. (2004) Innervation of the substantia nigra. Cell Tissue Res., 318,107–114.
Miyazaki, T. & Lacey, M.G. (1998) Presynaptic inhibition by dopamine of adiscrete component of GABA release in rat substantia nigra pars reticulata.J. Physiol., 513, 805–817.
Monaghan, D.T. & Cotman, C.W. (1985) Distribution of N-methyl-D-aspartate-sensitive L-[3H]glutamate-binding sites in rat brain. J. Neurosci., 5, 2909–2919.
Nakahama, H., Ishii, N., Yamamoto, M. & Saito, H. (1971) Stochasticproperties of spontaneous impulse activity in single central neurons. TohokuJ. Exp. Med., 104, 371–391.
Nicola, S.M., Surmeier, J. & Malenka, R.C. (2000) Dopaminergic modulationof neuronal excitability in the striatum and nucleus accumbens. Annu. Rev.Neurosci., 23, 185–215.
Nieoullon, A., Cheramy, A. & Glowinski, J. (1977) Release of dopamine invivo from cat substantia nigra. Nature, 266, 375–377.
Olds, M.E. (1988) The response of non-dopamine neurons in substantia nigraand ventral tegmental area to amphetamine and apomorphine duringhypermotility: the striatal influence. Brain Res., 452, 237–254.
Paxinos, G. & Watson, C. (1998) The Rat Brain in Stereotaxic Coordinates, 4thedn. Academic Press, San Diego.
Petralia, R.S. & Wenthold, R.J. (1992) Light and electron immunocytochemicallocalization of AMPA-selective glutamate receptors in the rat brain. J. Comp.Neurol., 318, 329–354.
Radnikow, G. & Misgeld, U. (1998) Dopamine D1 receptor facilitate GABA-Asynaptic currents in the rat substantia nigra pars reticulata. J. Neurosci., 18,2009–2016.
Rebec, G.V., Langley, P.E., Pierce, R.C., Wang, Z. & Heidenreich, B.A. (1993)A simple micromanipulator for multiple uses in freely moving rats:electrophysiology, voltammetry, and simultaneous intracerebral infusions.J. Neurosci. Meth., 47, 53–59.
Rebec, G.V., White, I.M. & Puotz, J.K. (1997) Responses of neurons in dorsalstriatum during amphetamine-induced focused stereotypy. Psychopharma-cology, 130, 343–351.
Rick, C.E. & Lacey, M.G. (1994) Rat substantia nigra pars reticulata neuronsare tonically inhibited via GABAA, but not GABAB, receptors in vitro.Brain Res., 659, 133–137.
Dendritic dopamine release and GABA transmission 1393
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394

Rosales, M.G., Martinez-Fong, D., Morales, R., Nunez, A., Flores, G.,Gongora-Alfaro, J.L., Floran, B. & Aceves, J. (1997) Reciprocal interactionbetween glutamate and dopamine in the pars reticulata of the rat substantianigra: a microdialysis study. Neuroscience, 80, 803–810.
Ruffieux, A. & Schultz, W. (1981) Influence of dopamine on pars reticulataneurones of substantia nigra. J. Physiol. (Paris), 77, 63–69.
Santiago, M. & Westerink, B.H. (1992) Simultaneous recording of the releaseof nigral and striatal dopamine in the awake rat. Neurochem. Int., 20(Suppl.), 107S–110S.
Schultz, W. (1986) Responses of midbrain dopamine neurons to behavioraltrigger stimuli in the monkey. J. Neurophysiol., 56, 1439–1461.
Smith, Y. & Bolam, J.P. (1989) Neurons of the substantia nigra reticulatareceive a dense GABA-containing input from the globus pallidus in the rat.Brain Res., 493, 160–167.
Steinfels, G.F., Heym, J., Strecker, R.E. & Jacobs, B.L. (1983) Response ofdopaminergic neurons in cat to auditory stimuli presented across the sleep-waking cycle. Brain Res., 277, 150–154.
Strecker, R.E. & Jacobs, B.L. (1985) Substantia nigra dopaminergic unitactivity in behaving cats: effect of arousal on spontaneous discharge andsensory evoked activity. Brain Res., 361, 339–350.
Tepper, J.M., Martin, L.P. & Anderson, D.R. (1995) GABA-A receptor-mediated inhibition of rat substantia nigra dopaminergic neurons byreticulata projection neurons. J. Neurosci., 15, 3092–3103.
Timmerman, W. & Abercrombie, E.D. (1996) Amphetamine-induced release ofdendritic dopamine in substantia nigra pars reticulata: D1-mediatedbehavioral and electrophysiological effects. Synapse, 23, 280–291.
Trevitt, J.T., Carlson, B.B., Nowend, K. & Salamone, J.D. (2001) Substantianigra pars reticulata is a highly potent site of action for the behavioral effects
of the D1 antagonist SCH 23390 in the rat. Psychopharmacology, 156,32–41.
Trevitt, T., Carlson, B., Correa, M., Keene, A., Morales, M. & Salamone, J.D.(2002) Interactions between dopamine D1 receptors and gamma-aminobu-tyric acid mechanisms in substantia nigra pars reticulata of the rat:neurochemical and behavioral studies. Psychopharmacology, 159, 229–237.
Waszczak, B.L. & Walters, J.R. (1983) Dopamine modulation of the effects ofgamma-aminobutyric acid on substantia nigra pars reticulata neurons.Science, 220, 218–221.
Werner, G. & Mountcastle, V.B. (1963) The variability of central neural activityin a sensory system, and its implications for the central reflection of sensoryevents. J. Neurophysiol., 29, 958–977.
Windels, F. & Kiyatkin, E.A. (2004) GABA, not glutamate, controls theactivity of substantia nigra reticulata neurons in awake, unrestrained rats.J. Neurosci., 24, 6751–6754.
Windels, F. & Kiyatkin, E.A. (2006) Stability of substantia nigra pars reticulataneuronal discharge rates during dopamine receptor blockade and its possiblemechanisms. Neuroreport, 17, 1071–1075.
Wittmann, M., Marino, M.J. & Conn, P.J. (2002) Dopamine modulates thefunction of group II and group III metabotropic glutamate receptors in thesubstantia nigra pars reticulata. J. Pharmacol. Exp. Ther., 302, 433–441.
Yung, K.K., Bolam, J.P., Smith, A.D., Hersch, S.M., Ciliax, B.J. & Levey, A.I.(1995) Immunocytochemical localization of D1 and D2 dopamine receptorsin the basal ganglia of the rat: light and electron microscopy. Neuroscience,65, 709–730.
Zhang, H., Weng, X. & Rosenberg, H.C. (1993) Characterization of substantianigra pars reticulata neurons based on response to iontophoretically appliedGABA and flurazepam. Life Sci., 53, 1911–1919.
1394 F. Windels and E. A. Kiyatkin
ª The Authors (2006). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 24, 1385–1394
Related Documents











