doi:10.1182/blood-2005-09-3623 Prepublished online December 13, 2005; 2006 107: 2993-3001 Samuel Strober and Defu Zeng Chunyan Zhang, Ivan Todorov, Zhifang Zhang, Yinping Liu, Fouad Kandeel, Stephen Forman, graft-versus-host disease with autoimmune manifestations T and B cells in transplants induce chronic + Donor CD4 (1710 articles) Transplantation (4651 articles) Immunobiology Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: Copyright 2011 by The American Society of Hematology; all rights reserved. Washington DC 20036. by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.org From

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1182/blood-2005-09-3623Prepublished online December 13, 2005;2006 107: 2993-3001
Samuel Strober and Defu ZengChunyan Zhang, Ivan Todorov, Zhifang Zhang, Yinping Liu, Fouad Kandeel, Stephen Forman, graft-versus-host disease with autoimmune manifestations
T and B cells in transplants induce chronic+Donor CD4
(1710 articles)Transplantation � (4651 articles)Immunobiology �
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Washington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom

TRANSPLANTATION
Donor CD4� T and B cells in transplants induce chronic graft-versus-host diseasewith autoimmune manifestationsChunyan Zhang, Ivan Todorov, Zhifang Zhang, Yinping Liu, Fouad Kandeel, Stephen Forman, Samuel Strober, and Defu Zeng
Chronic graft-vs-host disease (GVHD) is amajor cause of morbidity and mortality oflong-term survivors of allogeneic hemato-poietic cell transplantation (HCT). ChronicGVHD can have features of an autoim-mune collagen vascular disease with clini-cal manifestations similar to autoimmunescleroderma and systemic lupus erythem-atosus (SLE). However, the pathogenesisof chronic GVHD is poorly understood. Itis unclear how autoreactive T and B cellsare generated in chronic GVHD recipi-ents. We have recently developed a newchronic GVHD model by transplantation
of donor DBA/2 (H-2d) spleen cells intomajor histocompatibility complex (MHC)–matched but minor antigen–mismatchedsublethally irradiated BALB/c (H-2d) recipi-ents as well as athymic BALB/cnu/nu andadult-thymectomized BALB/c recipients.Both euthymic and athymic BALB/c recipi-ents developed high levels of serum IgGautoantibodies, sclerodermatous skindamage, and glomerulonephritis. Dis-ease induction required both donorCD25�CD4� T and B cells in transplants.In contrast, donor CD25�CD4� T regula-tory (Treg) cells prevented the disease
induction. These results indicate that hostthymus is not required for induction ofchronic GVHD and that quiescent autore-active T and B cells in transplants fromnonautoimmune donors may be activatedand expanded to cause chronic GVHDwith autoimmune manifestations in alloge-neic recipients, and donor Treg cells cansuppress this process. (Blood. 2006;107:2993-3001)
© 2006 by The American Society of Hematology
Introduction
Chronic graft-versus-host disease (GVHD) is a serious and com-mon long-term complication of allogeneic hematopoietic celltransplantation (HCT) occurring in 20% to 70% of patientssurviving more than 100 days after HCT.1-4 Despite improvementsin the practice of allogeneic HCT over the last 25 years, there hasbeen little change in the incidence, morbidity, and mortality of thiscomplication.3 One of the difficulties in combating chronic GVHDis the poor understanding of its pathogenesis.
Chronic GVHD differs from acute GVHD in many aspects.First, the onset of acute GVHD is usually at 1 to 2 monthsfollowing transplantation but the onset of chronic GVHD is usuallydelayed until 4 to 6 months after transplantation.3 Second, althoughthe target organ tissues of chronic GVHD are significantly over-lapped with that of acute GVHD (ie, skin, gut, liver, and lung), thehistopathology is distinguishably different.3 While acute GVHDshows donor lymphocyte infiltration and host tissue-cell apoptosisand necrosis in target organs, chronic GVHD is featured by amarked increase in collagen deposition and a lack of T-lymphocyteinfiltration in the target organ tissues.3,4 Third, up to 70% of chronicGVHD patients have elevated levels of serum autoantibodies (eg,antinuclear, anti-dsDNA, and anti–smooth-muscle antibodies),1,5,6
and depletion of B cells ameliorates refractory chronic GVHD insome patients.3,7,8 Therefore, chronic GVHD has features similar toautoimmune collagen vascular disease such as scleroderma andsystemic lupus erythematosus (SLE).1,9 However, it is unclear howthe autoimmune responses develop in chronic GVHD.
Several murine HCT models have been used to study thepathogenesis of chronic GVHD. The first type of model istransplantation of parental lymphocytes into nonirradiated majorhistocompatibility complex (MHC)–mismatched F1 recipients.10,11
In those models, the F1 recipients developed high levels of serumanti-dsDNA and glomerulonephritis, and the production of autoan-tibodies is a result of a cognate interaction between donor CD4� Tcells and host B cells.10-15 However, it is not clear whether themechanisms revealed in those models reflect the pathogenesis ofchronic GVHD in the irradiated HC transplant recipients.
The second type of model is transplantation of donorlymphocytes into MHC-matched but minor antigen–mis-matched irradiated recipients. In one model, donor LP/J (H-2b)bone marrow and spleen cells were transplanted into lethallyirradiated C57BL/6 (H-2b) recipients and the recipients developedacute and chronic forms of GVHD.16 Clonal analysis of T cells
From the Beckman Research Institute, City of Hope National Medical Center,Duarte, CA; and Stanford University School of Medicine, Stanford, CA.
Submitted September 9, 2005; accepted December 2, 2005. Prepublishedonline as Blood First Edition Paper, December 13, 2005; DOI 10.1182/blood-2005-09-3623.
Supported by the Leslie and Susan Gonda (Goldschmied) Foundation, anArthritis Foundation Investigator Award, and National Institutes of Health grantR01-AI66008 (D.Z.).
C.Z. designed research, performed experiments, analyzed data, andprepared the manuscript; I.T. performed experiments and analyzed datain immunohistology; Z.Z. performed experiments in cell transfer and flowcytometry analysis; Y.L. performed experiments in autoantibody production;F.K. advised on experimental design and reviewing of manuscript; S.S. advised
on experimental design and manuscript preparation; S.F. advised onexperimental design and manuscript preparation; and D.Z. designed research,analyzed data, and prepared the manuscript.
An Inside Blood analysis of this article appears at the front of this issue.
The online version of this article contains a data supplement.
Reprints: Defu Zeng, The Beckman Research Institute, Gonda Building,R2017, City of Hope National Medical Center, 1500 East Duarte Rd, Duarte,CA 91010; e-mail: [email protected].
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. section 1734.
© 2006 by The American Society of Hematology
2993BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom

from the acute and chronic GVHD C57BL/6 recipients indicatedthat acute GVHD was due primarily to recipient-specific cytotoxicdonor T lymphocytes, whereas chronic GVHD was due to autoreac-tive CD4� T lymphocytes.16 However, it was unknown how theautoreactive CD4� T cells were generated.
In another model, B10D2 (H-2d) donor spleen cells weretransplanted into lethally irradiated BALB/c recipients and therecipients developed sclerodermatous skin damage.17-19 DonorCD4� T cells were required for the initiation of the disease,20 but itwas unclear how donor CD4� T cells induced the disease.
In the current study, we have developed a new model of chronicGVHD with autoimmune manifestations in which donor DBA/2(H-2d) spleen cells were transplanted into sublethally irradiated,MHC-matched but minor antigen–mismatched, euthymic BALB/c(H-2d) recipients as well as athymic BALB/cnu/nu and adult-thymectomized BALB/c recipients. The recipients developed anautoimmune lupus and scleroderma overlap syndrome manifestedwith high levels of serum autoantibodies, sclerodermatous skindamage, proteinuria, and glomerulonephritis. Disease inductionrequired both donor conventional CD25�CD4� T and B cells intransplants. In contrast, donor CD4�CD25� T regulatory (Treg)cells in transplants suppressed the disease induction. Our studiesdemonstrate that host thymus is not required for induction ofchronic GVHD and that both donor CD4� T and B cells intransplants from nonautoimmune donors are required for thedisease induction in this model.
Materials and methods
Mice
Male DBA/2(H-2d), B10D2/nSnJ (H-2d), BALB/c (H-2d), and BALB/cnu/nu
and adult-thymectomized (aged 6 weeks) BALB/c mice were purchasedfrom The Jackson Laboratory (Bar Harbor, ME) and maintained in apathogen-free room at City of Hope Research Animal Facilities (Duarte,CA). Mice aged 8 to 12 weeks were used in the current studies.
Monoclonal antibodies, flow cytometric analysis,and cell sorting
The FITC-, phycoerythrin-, APC-, and Cy7-APC–conjugated monoclonalantibodies (mAbs) to mouse T-cell receptor (TCR), B220, CD4, CD8, andCD25 were all purchased from BD Pharmingen (San Diego, CA).Multiple-color fluorescence-activated cell sorter (FACS) analysis andsorting were performed at City of Hope FACS facility using a 4-laserMOFLO immunocytometry system (Dako Cytomation, Fort Collins, CO),and data was analyzed using FLOWJO software (Tree Star, San Carlos,CA).21,22 The FITC-, APC-conjugated mAbs to mouse IL-2, IL-4, andIFN-� were all purchased from BD Pharmingen. CD8� and CD4� T cellswere purified with a magnetic purification system from Miltenyi Biotec(Auburn, CA) as described previously.21 The purity of positively selectedCD8� or CD4� T cells was greater than 96%. Depletion of CD4�, CD8�,and B220� cells was performed with magnetic microbeads and Miltenyidepletion columns. The residual target cells were about 1% of totalmononuclear cells.
Proliferation assays
Purified CD25�CD4� T cells (1 � 105) together with 2 � 105 irradiatedsyngeneic T-cell–depleted spleen cells were cultured in a U-bottomed96-well plate for 72 hours in the presence of 0.5 �g/mL anti-CD3 mAb.3H-TdR (1 �Ci/mL [0.037 MBq/mL]) was added 8 hours before harvest.Results are expressed as the mean of triplicate cultures.
Histopathology and immunofluorescence microscopy
Hematoxylin and eosin (HE) staining of skin tissues and immunofluores-cent staining of IgG immune complex in skin and kidney tissues of BALB/crecipients were described previously.21,23,24 For collagen I staining, cryosec-tions were first incubated with rabbit anti–mouse collagen I at 1:200dilution (Chemicon International, Temecula, CA) and then with Texasred–conjugated donkey anti–rabbit IgG (Jackson Immunoresearch Labora-tories). After extensive washes, the samples were counterstained with4,6-diamidino-2-phenylindole (DAPI; Sigma, St Louis, MO) and embed-ded in Vectashield (Vector Laboratories, Burlingame, CA). Visualizationand image preparation procedures were described previously.21 Briefly, theHE staining slides and immunofluorescent staining slides were examinedat 400 � magnification. The samples were visualized with an OlympusBX51 fluorescent microscope equipped with Olympus 20 �/0.70 NA and40 �/0.90 Plan Apo objectives (Olympus America, Melville, NY) and aPixera (600CL) cooled CCD camera (Pixera, Los Gatos, CA). Fluorescentimages relative to each marker were collected using a corresponding filter set andPixera viewfinder acquisition software 3.0. Color composite images weregenerated using Adobe Photoshop 7.0 (Adobe Systems, San Jose, CA).
Measurement of cytokines and antibodies in serumand culture supernatants
Cytokines were measured using the Luminex LabMap system and enzyme-linked immunosorbent assay (ELISA) kits (Biosource International, Cama-rillo, CA) as described previously.21,23 Total IgG and anti-dsDNA IgG weremeasured with ELISA as described previously.24,25 For determination of theallotype of IgG2a anti-dsDNA, plates coated with calf thymus ds-DNAwere used to capture anti-dsDNA, then biotin-conjugated anti–mouseIgG2aa mAb (BD Pharmingen clone 8.3) was applied to the plates, andstreptavidin-conjugated HRP (BD Pharmingen) was used to develop theELISA.
Statistical analysis
Proteinuria incidences and survival in different groups were comparedusing the log-rank test with GraphPad Prism Version 3.0 (Graph PadSoftware, San Diego, CA). Comparison of 2 means was analyzed usingunpaired 2-tailed Student t test.
Results
DBA/2 spleen cells induced GVHD in sublethally irradiatedBALB/c recipients in a dose-dependent manner
Since parental donor DBA/2 alloreactive CD4� T cells inducedlupuslike GVHD in nonirradiated (DBA/2 x C57BL/6) F1 recipi-ents via activation of host B cells and DBA/2 donor CD8� T cellswere unable to induce GVHD,12,26 we tested whether transplanta-tion of DBA/2 (H-2d) spleen cells into sublethally irradiatedBALB/c (H-2d) recipients induced GVHD with autoimmune mani-festations. Accordingly, graded numbers (3.125 � 106 to 100 � 106)of DBA/2 spleen cells were injected intravenously into sublethallyirradiated (6.5 Gy [650 rad]) BALB/c recipients. The recipientswere monitored for clinical signs of GVHD such as diarrhea,weight loss, hair loss, and death and were also checked forproteinuria twice a week. As shown in Figure 1A, all the recipientsgiven irradiation alone or syngeneic BALB/c spleen cells(100 � 106) showed no signs of GVHD and survived for more than50 days. In contrast, the recipients injected with DBA/2 spleen cellsshowed mild acute GVHD for the first 2 weeks and then showedchronic GVHD with proteinuria and hair loss in a donor-celldose-dependent manner. The recipients given 100 � 106 spleencells all developed proteinuria (4�), ascites, and hair loss by day22 and all died by day 42 after cell injection, but none of the
2994 ZHANG et al BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom
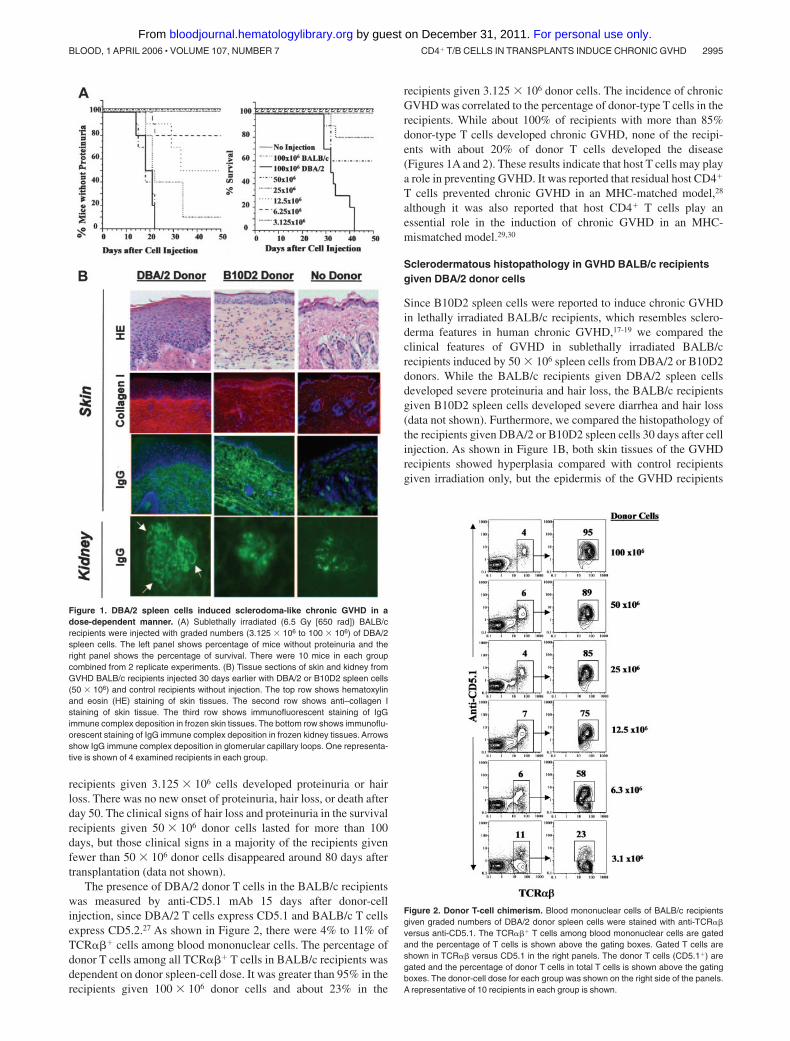
recipients given 3.125 � 106 cells developed proteinuria or hairloss. There was no new onset of proteinuria, hair loss, or death afterday 50. The clinical signs of hair loss and proteinuria in the survivalrecipients given 50 � 106 donor cells lasted for more than 100days, but those clinical signs in a majority of the recipients givenfewer than 50 � 106 donor cells disappeared around 80 days aftertransplantation (data not shown).
The presence of DBA/2 donor T cells in the BALB/c recipientswas measured by anti-CD5.1 mAb 15 days after donor-cellinjection, since DBA/2 T cells express CD5.1 and BALB/c T cellsexpress CD5.2.27 As shown in Figure 2, there were 4% to 11% ofTCR��� cells among blood mononuclear cells. The percentage ofdonor T cells among all TCR��� T cells in BALB/c recipients wasdependent on donor spleen-cell dose. It was greater than 95% in therecipients given 100 � 106 donor cells and about 23% in the
recipients given 3.125 � 106 donor cells. The incidence of chronicGVHD was correlated to the percentage of donor-type T cells in therecipients. While about 100% of recipients with more than 85%donor-type T cells developed chronic GVHD, none of the recipi-ents with about 20% of donor T cells developed the disease(Figures 1A and 2). These results indicate that host T cells may playa role in preventing GVHD. It was reported that residual host CD4�
T cells prevented chronic GVHD in an MHC-matched model,28
although it was also reported that host CD4� T cells play anessential role in the induction of chronic GVHD in an MHC-mismatched model.29,30
Sclerodermatous histopathology in GVHD BALB/c recipientsgiven DBA/2 donor cells
Since B10D2 spleen cells were reported to induce chronic GVHDin lethally irradiated BALB/c recipients, which resembles sclero-derma features in human chronic GVHD,17-19 we compared theclinical features of GVHD in sublethally irradiated BALB/crecipients induced by 50 � 106 spleen cells from DBA/2 or B10D2donors. While the BALB/c recipients given DBA/2 spleen cellsdeveloped severe proteinuria and hair loss, the BALB/c recipientsgiven B10D2 spleen cells developed severe diarrhea and hair loss(data not shown). Furthermore, we compared the histopathology ofthe recipients given DBA/2 or B10D2 spleen cells 30 days after cellinjection. As shown in Figure 1B, both skin tissues of the GVHDrecipients showed hyperplasia compared with control recipientsgiven irradiation only, but the epidermis of the GVHD recipients
Figure 2. Donor T-cell chimerism. Blood mononuclear cells of BALB/c recipientsgiven graded numbers of DBA/2 donor spleen cells were stained with anti-TCR��versus anti-CD5.1. The TCR��� T cells among blood mononuclear cells are gatedand the percentage of T cells is shown above the gating boxes. Gated T cells areshown in TCR�� versus CD5.1 in the right panels. The donor T cells (CD5.1�) aregated and the percentage of donor T cells in total T cells is shown above the gatingboxes. The donor-cell dose for each group was shown on the right side of the panels.A representative of 10 recipients in each group is shown.
Figure 1. DBA/2 spleen cells induced sclerodoma-like chronic GVHD in adose-dependent manner. (A) Sublethally irradiated (6.5 Gy [650 rad]) BALB/crecipients were injected with graded numbers (3.125 � 106 to 100 � 106) of DBA/2spleen cells. The left panel shows percentage of mice without proteinuria and theright panel shows the percentage of survival. There were 10 mice in each groupcombined from 2 replicate experiments. (B) Tissue sections of skin and kidney fromGVHD BALB/c recipients injected 30 days earlier with DBA/2 or B10D2 spleen cells(50 � 106) and control recipients without injection. The top row shows hematoxylinand eosin (HE) staining of skin tissues. The second row shows anti–collagen Istaining of skin tissue. The third row shows immunofluorescent staining of IgGimmune complex deposition in frozen skin tissues. The bottom row shows immunoflu-orescent staining of IgG immune complex deposition in frozen kidney tissues. Arrowsshow IgG immune complex deposition in glomerular capillary loops. One representa-tive is shown of 4 examined recipients in each group.
CD4� T/B CELLS IN TRANSPLANTS INDUCE CHRONIC GVHD 2995BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom
given DBA/2 spleen cells was much thicker than that of GVHDrecipients given B10D2 spleen cells. In addition, it appeared thatthere was less mononuclear-cell infiltration in the skin dermis ofGVHD recipients given DBA/2 donor cells compared with therecipients given B10D2 donor cells.
Excess collagen deposition in the dermis is one of the character-istic features of human chronic GVHD.31 Thus, we also stained theskin tissues with anti–collagen I as described previously.19 Asshown in Figure 1B (second row), there was excess collagendeposition in the dermis of both GVHD recipients compared withcontrol recipients, but it appeared that there was more collagendeposition in the dermis of recipients given DBA/2 donor cellscompared with the recipients given B10D2 donor cells.
Human chronic GVHD was reported to have IgG immunecomplex deposition in dermis,32 and human and murine systemiclupus was reported to have IgG immune complex deposition inglomerular capillary loops.33,34 Thus, skin and kidney tissues ofGVHD recipients given DBA/2 or B10D2 donor spleen cells werechecked for IgG immune complex deposition 30 days afterdonor-cell injection. As shown in Figure 1B (bottom 2 rows), IgGimmune complex deposition was found in the dermis of bothGVHD recipients. It was also found in glomerular capillary loopsand mesangium of GVHD recipients given DBA/2 donor cells butonly in the glomerular mesangium of GVHD recipients givenB10D2 donor cells. The control recipients showed a little IgGdeposition in either skin dermis or glomerular mesangium. Takentogether, similar to BALB/c recipients given B10D2 donor cells,BALB/c recipients given DBA/2 donor cells developed a scleroder-matous syndrome. In addition, the latter recipients also developed alupuslike syndrome.
High levels of serum anti-dsDNA antibodies of DBA/2 donororigin in the BALB/c recipients with chronic GVHD
Since BALB/c recipients given DBA/2 donor cells showed immunecomplex glomerulonephritis (Figure 1B), serum levels of total IgG1 andIgG2a and anti-dsDNA IgG1 and IgG2a of BALB/c recipients given50 � 106 DBA/2 spleen cells were measured with ELISA 15 days aftercell injection. As shown in Table 1, compared with recipients givenirradiation only, the recipients given additional DBA/2 spleen cells had10-fold higher levels of serum IgG1, 100-fold higher levels of IgG2a,and more than 100-fold higher levels of anti-dsDNA IgG1 and IgG2a(P � .001). BALB/c or DBA/2 recipients given syngeneic BALB/c orDBA/2 spleen cells did not show significant increase of total oranti-dsDNA IgG (data not shown).
Next, we determined which B cells (donor or host) were thesource of autoantibodies in the irradiated BALB/c recipients givenDBA/2 spleen cells. IgG2a antibodies secreted by DBA/2 B cells
express allotype c, but those secreted by BALB/c B cells expressallotype a.35 Since there is anti–allotype a, but no anti–allotype c,mAb available, we used anti–allotype a mAb to identify the originof anti-dsDNA IgG2a in the serum of BALB/c recipients. Inaddition, because IgG2a secreted by New Zealand Black (NZB)B cells is allotype a, we use serum from sublethally irradiatedBALB/c recipients given NZB donor spleen cells as the positivecontrol of anti–allotype a. As shown in Table 1, almost all of serumanti-dsDNA IgG2a (366 79 U/mL) of BALB/c recipients givenNZB donor cells was allotype a (293 32 U/mL; P .1). Incontrast, no allotype a anti-dsDNA IgG2a was detectable in theserum of recipients given DBA/2 donor cells with high levels ofanti-dsDNA IgG2a (1459 163 U/mL), indicating that thoseanti-dsDNA IgG2a antibodies are not secreted by the B cells ofirradiated BALB/c recipients with allotype a but are insteadsecreted by the B cells of DBA/2 donors. B cells are very sensitiveto irradiation.36-38 Thus, it is not surprising that autoantibodies inthe sublethally irradiated BALB/c recipients given DBA/2 donorcells are from donor B cells, although autoantibodies in thenonirradiated (C57BL/6 x DBA/2) F1 recipients given parentalDBA/2 donor cells are from host B cells.26
Transplanted donor CD4� but not CD8� T cells were requiredfor the induction of chronic GVHD with autoimmunemanifestations
As shown in Figure 3A, while all the recipients given 50 � 106
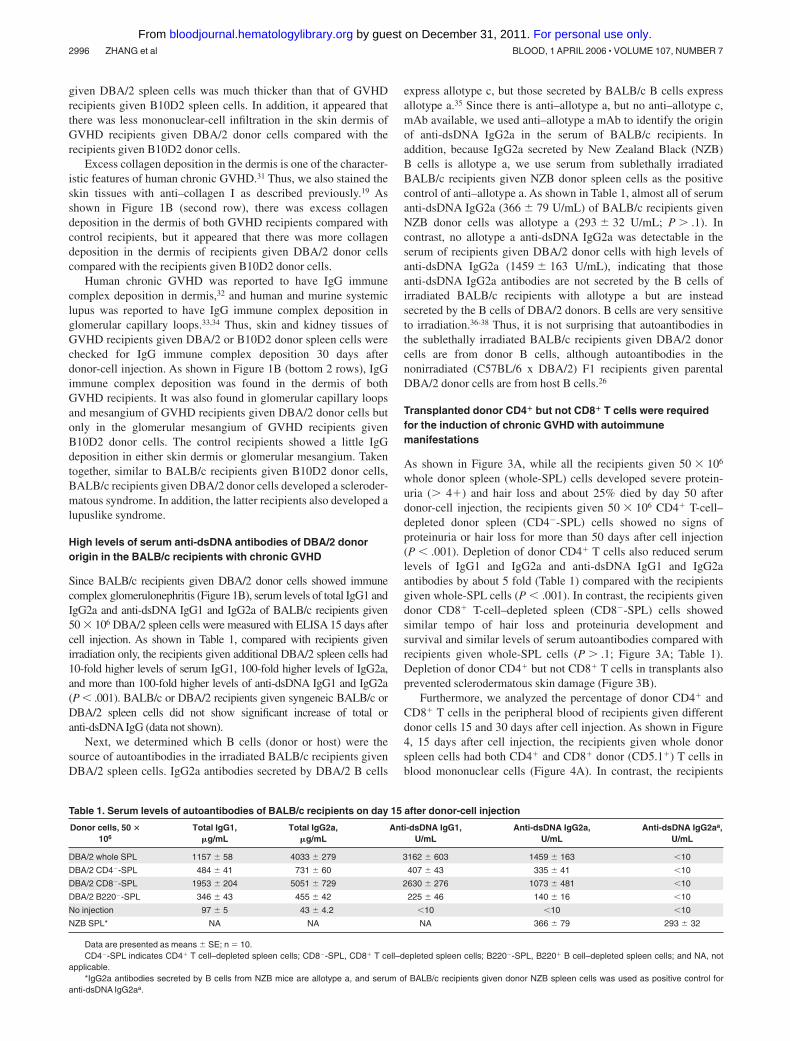
whole donor spleen (whole-SPL) cells developed severe protein-uria ( 4�) and hair loss and about 25% died by day 50 afterdonor-cell injection, the recipients given 50 � 106 CD4� T-cell–depleted donor spleen (CD4�-SPL) cells showed no signs ofproteinuria or hair loss for more than 50 days after cell injection(P � .001). Depletion of donor CD4� T cells also reduced serumlevels of IgG1 and IgG2a and anti-dsDNA IgG1 and IgG2aantibodies by about 5 fold (Table 1) compared with the recipientsgiven whole-SPL cells (P � .001). In contrast, the recipients givendonor CD8� T-cell–depleted spleen (CD8�-SPL) cells showedsimilar tempo of hair loss and proteinuria development andsurvival and similar levels of serum autoantibodies compared withrecipients given whole-SPL cells (P .1; Figure 3A; Table 1).Depletion of donor CD4� but not CD8� T cells in transplants alsoprevented sclerodermatous skin damage (Figure 3B).
Furthermore, we analyzed the percentage of donor CD4� andCD8� T cells in the peripheral blood of recipients given differentdonor cells 15 and 30 days after cell injection. As shown in Figure4, 15 days after cell injection, the recipients given whole donorspleen cells had both CD4� and CD8� donor (CD5.1�) T cells inblood mononuclear cells (Figure 4A). In contrast, the recipients
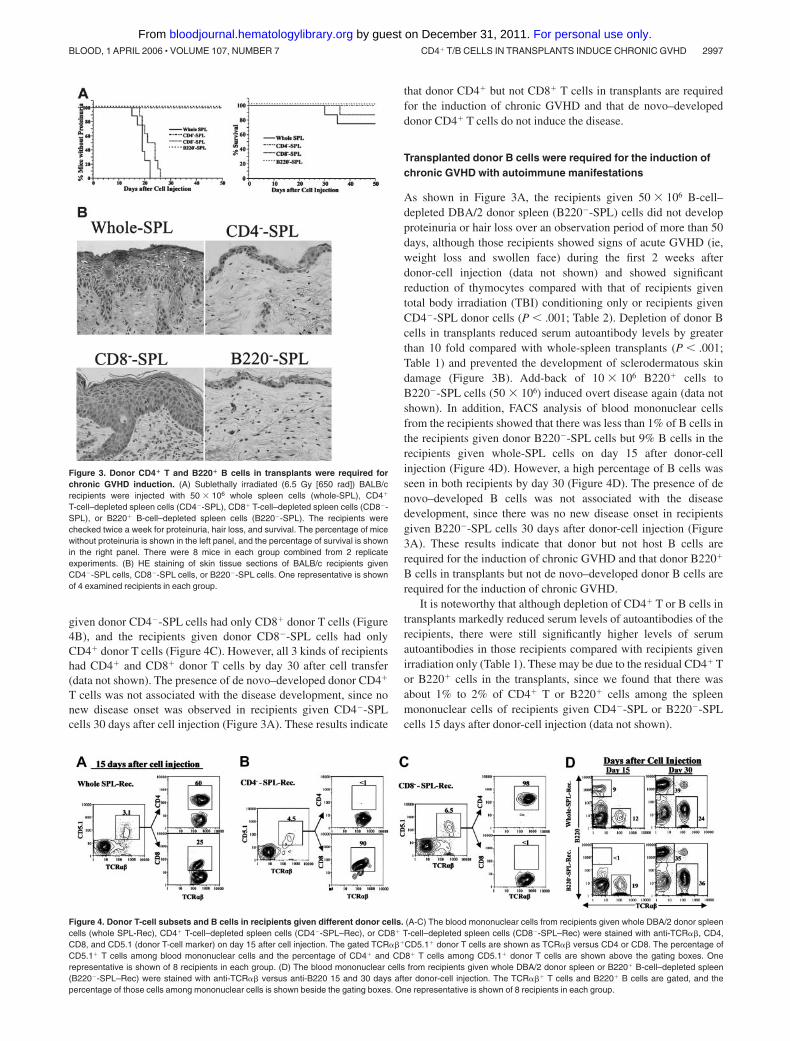
Table 1. Serum levels of autoantibodies of BALB/c recipients on day 15 after donor-cell injection
Donor cells, 50 �
106Total IgG1,
�g/mLTotal IgG2a,
�g/mLAnti-dsDNA IgG1,
U/mLAnti-dsDNA IgG2a,
U/mLAnti-dsDNA IgG2aa,
U/mL
DBA/2 whole SPL 1157 58 4033 279 3162 603 1459 163 �10
DBA/2 CD4�-SPL 484 41 731 60 407 43 335 41 �10
DBA/2 CD8�-SPL 1953 204 5051 729 2630 276 1073 481 �10
DBA/2 B220�-SPL 346 43 455 42 225 46 140 16 �10
No injection 97 5 43 4.2 �10 �10 �10
NZB SPL* NA NA NA 366 79 293 32
Data are presented as means SE; n � 10.CD4�-SPL indicates CD4� T cell–depleted spleen cells; CD8�-SPL, CD8� T cell–depleted spleen cells; B220�-SPL, B220� B cell–depleted spleen cells; and NA, not
applicable.*IgG2a antibodies secreted by B cells from NZB mice are allotype a, and serum of BALB/c recipients given donor NZB spleen cells was used as positive control for
anti-dsDNA IgG2aa.
2996 ZHANG et al BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom
given donor CD4�-SPL cells had only CD8� donor T cells (Figure4B), and the recipients given donor CD8�-SPL cells had onlyCD4� donor T cells (Figure 4C). However, all 3 kinds of recipientshad CD4� and CD8� donor T cells by day 30 after cell transfer(data not shown). The presence of de novo–developed donor CD4�
T cells was not associated with the disease development, since nonew disease onset was observed in recipients given CD4�-SPLcells 30 days after cell injection (Figure 3A). These results indicate
that donor CD4� but not CD8� T cells in transplants are requiredfor the induction of chronic GVHD and that de novo–developeddonor CD4� T cells do not induce the disease.
Transplanted donor B cells were required for the induction ofchronic GVHD with autoimmune manifestations
As shown in Figure 3A, the recipients given 50 � 106 B-cell–depleted DBA/2 donor spleen (B220�-SPL) cells did not developproteinuria or hair loss over an observation period of more than 50days, although those recipients showed signs of acute GVHD (ie,weight loss and swollen face) during the first 2 weeks afterdonor-cell injection (data not shown) and showed significantreduction of thymocytes compared with that of recipients giventotal body irradiation (TBI) conditioning only or recipients givenCD4�-SPL donor cells (P � .001; Table 2). Depletion of donor Bcells in transplants reduced serum autoantibody levels by greaterthan 10 fold compared with whole-spleen transplants (P � .001;Table 1) and prevented the development of sclerodermatous skindamage (Figure 3B). Add-back of 10 � 106 B220� cells toB220�-SPL cells (50 � 106) induced overt disease again (data notshown). In addition, FACS analysis of blood mononuclear cellsfrom the recipients showed that there was less than 1% of B cells inthe recipients given donor B220�-SPL cells but 9% B cells in therecipients given whole-SPL cells on day 15 after donor-cellinjection (Figure 4D). However, a high percentage of B cells wasseen in both recipients by day 30 (Figure 4D). The presence of denovo–developed B cells was not associated with the diseasedevelopment, since there was no new disease onset in recipientsgiven B220�-SPL cells 30 days after donor-cell injection (Figure3A). These results indicate that donor but not host B cells arerequired for the induction of chronic GVHD and that donor B220�
B cells in transplants but not de novo–developed donor B cells arerequired for the induction of chronic GVHD.
It is noteworthy that although depletion of CD4� T or B cells intransplants markedly reduced serum levels of autoantibodies of therecipients, there were still significantly higher levels of serumautoantibodies in those recipients compared with recipients givenirradiation only (Table 1). These may be due to the residual CD4� Tor B220� cells in the transplants, since we found that there wasabout 1% to 2% of CD4� T or B220� cells among the spleenmononuclear cells of recipients given CD4�-SPL or B220�-SPLcells 15 days after donor-cell injection (data not shown).
Figure 4. Donor T-cell subsets and B cells in recipients given different donor cells. (A-C) The blood mononuclear cells from recipients given whole DBA/2 donor spleencells (whole SPL-Rec), CD4� T-cell–depleted spleen cells (CD4�-SPL–Rec), or CD8� T-cell–depleted spleen cells (CD8�-SPL–Rec) were stained with anti-TCR��, CD4,CD8, and CD5.1 (donor T-cell marker) on day 15 after cell injection. The gated TCR���CD5.1� donor T cells are shown as TCR�� versus CD4 or CD8. The percentage ofCD5.1� T cells among blood mononuclear cells and the percentage of CD4� and CD8� T cells among CD5.1� donor T cells are shown above the gating boxes. Onerepresentative is shown of 8 recipients in each group. (D) The blood mononuclear cells from recipients given whole DBA/2 donor spleen or B220� B-cell–depleted spleen(B220�-SPL–Rec) were stained with anti-TCR�� versus anti-B220 15 and 30 days after donor-cell injection. The TCR��� T cells and B220� B cells are gated, and thepercentage of those cells among mononuclear cells is shown beside the gating boxes. One representative is shown of 8 recipients in each group.
Figure 3. Donor CD4� T and B220� B cells in transplants were required forchronic GVHD induction. (A) Sublethally irradiated (6.5 Gy [650 rad]) BALB/crecipients were injected with 50 � 106 whole spleen cells (whole-SPL), CD4�
T-cell–depleted spleen cells (CD4�-SPL), CD8� T-cell–depleted spleen cells (CD8�-SPL), or B220� B-cell–depleted spleen cells (B220�-SPL). The recipients werechecked twice a week for proteinuria, hair loss, and survival. The percentage of micewithout proteinuria is shown in the left panel, and the percentage of survival is shownin the right panel. There were 8 mice in each group combined from 2 replicateexperiments. (B) HE staining of skin tissue sections of BALB/c recipients givenCD4�-SPL cells, CD8�-SPL cells, or B220�-SPL cells. One representative is shownof 4 examined recipients in each group.
CD4� T/B CELLS IN TRANSPLANTS INDUCE CHRONIC GVHD 2997BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom
DBA/2 spleen cells induced chronic GVHD in BALB/cnu/nu andadult-thymectomized BALB/c recipients
It was proposed that chronic GVHD resulted from generation ofautoreactive T cells in the GVHD recipient’s thymus.31,39,40 We alsoobserved a marked reduction of thymocytes in the chronic GVHDrecipients given donor whole-SPL cells compared with GVHD-freerecipients given CD4�-SPL cells (P � .01; Table 2). To testwhether thymus is required for induction of chronic GVHD,50 � 106 DBA/2 spleen cells were injected into sublethally irradi-ated athymic BALB/cnu/nu as well as adult-thymectomized andsham-thymectomized BALB/c recipients. As shown in Table 3, allof the recipients given DBA/2 spleen cells developed severeproteinuria (4�) and high levels of serum anti-dsDNA IgGautoantibodies by day 20 after donor-cell injection. The recipientsalso showed hyperplasia in epidermis and proliferative glomerulo-nephritis (data not shown). There was no significant differencebetween thymectomized and sham-thymectomized recipients inclinical features and serum autoantibody levels (Table 3; P .1).These results indicate that induction of autoimmune-like chronicGVHD dose not require the host thymus or de novo–developeddonor T cells from the recipient thymus.
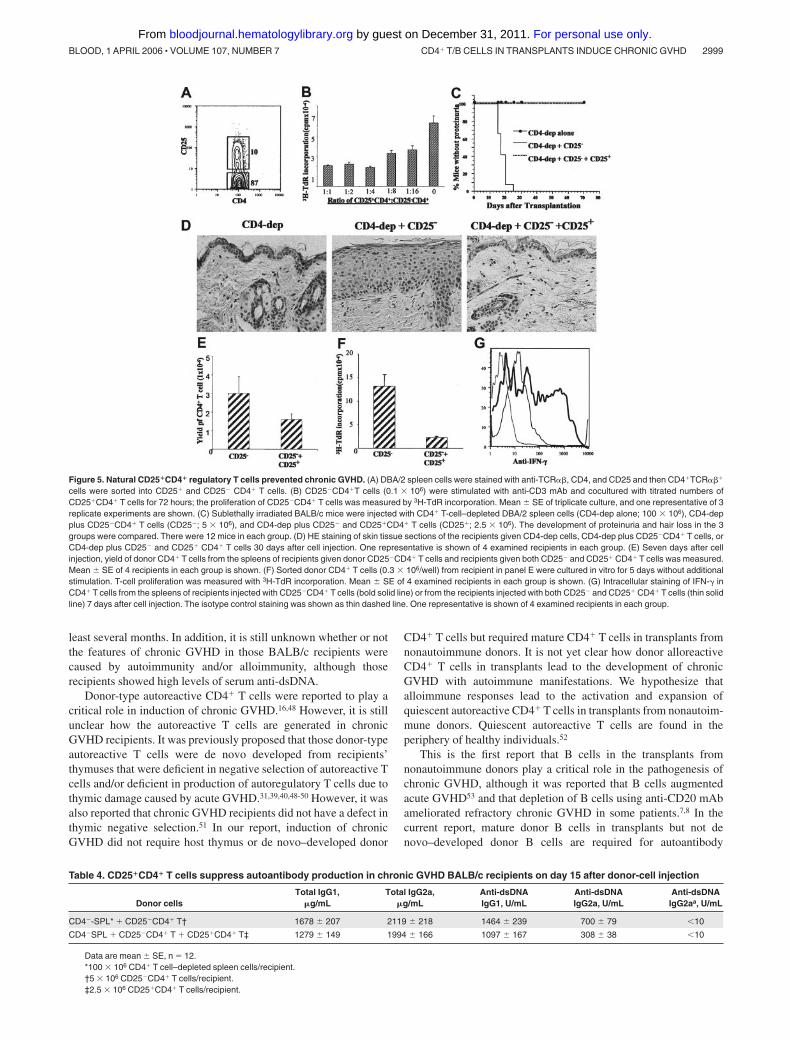
DBA/2 donor splenic CD25�CD4� T cells prevent chronic GVHDinduced by CD25�CD4� T cells
Donor CD25�CD4� T cells prevent acute GVHD in a variety ofanimal models.28,41-44 We tested whether CD25�CD4� T cells indonor DBA/2 spleen prevented chronic GVHD. DBA/2 spleenCD4� T cells were sorted into CD25� and CD25� subsets (Figure5A). We first found that CD25�CD4� T cells suppressed theproliferation of CD25�CD4� T cells in vitro in a cell-dose–dependent manner (Figure 5B). We then observed that 12 of 12 ofBALB/c recipients given 100 � 106 CD4� T-cell–depleted (CD4�-SPL) cells and 5 � 106 CD25�CD4� T cells developed proteinuriaand hair loss by day 30 after cell injection and 3 of 12 of them diedby day 40; the clinical signs of hair loss and proteinuria in the
survival recipients lasted for more than 100 days after cell injection(data not shown). In contrast, 0 of 12 of the BALB/C recipientsgiven the same numbers of donor CD4�-SPL and CD25�CD4� Tcells and additional 2.5 � 106 CD25�CD4� T cells developed thedisease during an observation period of more than 50 days (Figure5C). Compared with recipients given donor CD4�-SPL andCD25�CD4� T cells, the recipients given the same numbers ofdonor CD4�-SPL and CD25�CD4� T cells and additional 2.5 � 106
CD25�CD4� T cells showed a more than 100-fold increase inthymocyte yield (P � .001; Table 2), a 2-fold reduction in serumlevels of anti-dsDNA IgG2a (P � .01; Table 4), and a markedimprovement in sclerodermatous skin damage (Figure 5D).
In additional experiments, we attempted to study the mecha-nisms whereby donor CD25�CD4� T cells prevent chronic GVHD.Seven days after donor-cell injection, the yield of donor CD4� Tcells in spleens of recipients and in vitro spontaneous proliferationand IFN-� secretion of donor CD4� T cells were measured. Wefound that coinjection of donor CD25�CD4� T cells reduced theyield of donor CD4� T cells by 2 fold (Figure 5E; P � .01),suppressed the in vitro spontaneous proliferation of donor CD4� Tcells by 5 fold (Figure 5F; P � .01), reduced the percentage ofIFN-�–secreting donor CD4� T cells in spleen from 56% 7% to28% 3% (mean SE, n � 8; P � .01; Figure 5G), and reducedserum levels of IFN-� from 818 69 to 377 38 pg/mL (mean SE, n � 8; P � .01). These results indicate that donor CD25�CD4�
T cells may suppress the expansion and IFN-� secretion of donorCD25�CD4� T cells to reduce the production of autoantibodiesand prevent the development of chronic GVHD with autoimmunemanifestations. IFN-� was reported to play an important role inautoantibody isotype switch to IgG2a45,46 and augmentation ofcollagen deposition in chronic GVHD.31
Discussion
We have demonstrated here that CD4� T and B cells in transplantsfrom nonautoimmune DBA/2 donors induced chronic GVHD withhigh levels of serum autoantibodies, sclerodermatous skin damage,and glomerulonephritis in sublethally irradiated MHC-matched butminor antigen–mismatched euthymic and athymic BALB/c recipi-ents. The disease induction did not require host thymus but didrequire both donor CD4� T and B cells. In contrast, donorCD25�CD4� T cells suppressed the disease induction.
The scleroderma and lupus manifestations with high levels ofserum autoantibodies in the BALB/c recipients given DBA/2 donorcells reflect important clinical features of human chronicGVHD.1,31,47 However, the relevance of this model to humanchronic GVHD is unclear because the temporal time course ofchronic GVHD in those BALB/c recipients does not mirror that inhuman chronic GVHD, which can have a long latency period of at
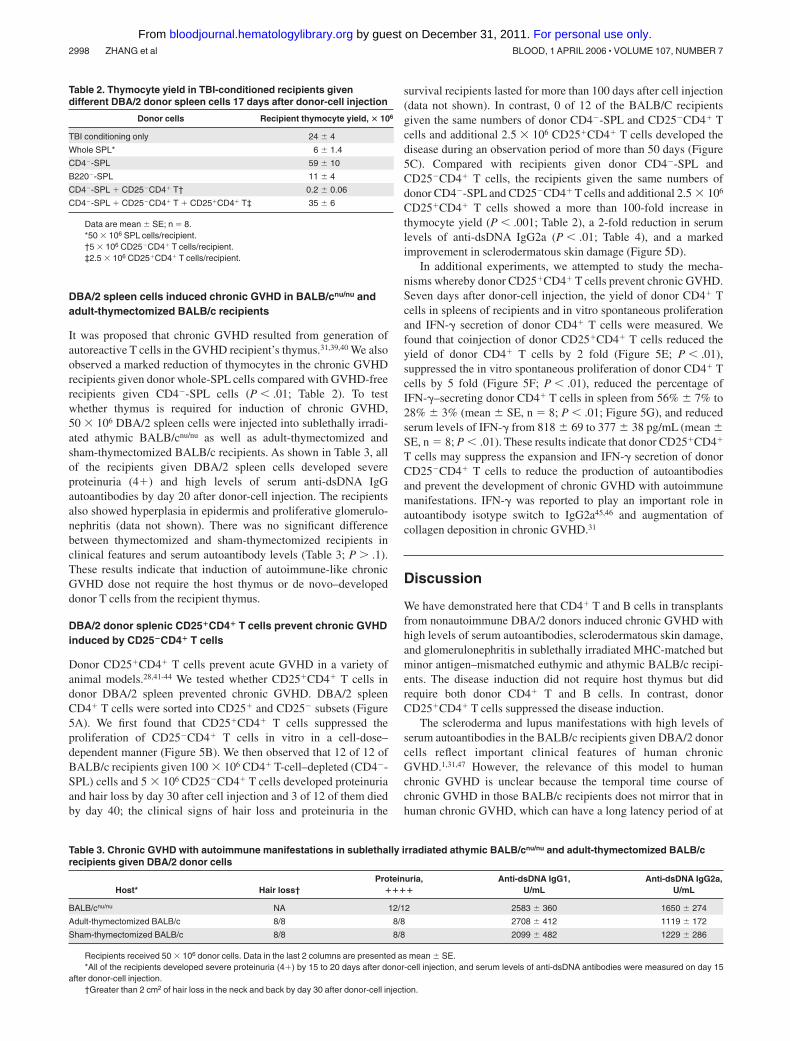
Table 2. Thymocyte yield in TBI-conditioned recipients givendifferent DBA/2 donor spleen cells 17 days after donor-cell injection
Donor cells Recipient thymocyte yield, � 106
TBI conditioning only 24 4
Whole SPL* 6 1.4
CD4�-SPL 59 10
B220�-SPL 11 4
CD4�-SPL � CD25�CD4� T† 0.2 0.06
CD4�-SPL � CD25�CD4� T � CD25�CD4� T‡ 35 6
Data are mean SE; n � 8.*50 � 106 SPL cells/recipient.†5 � 106 CD25�CD4� T cells/recipient.‡2.5 � 106 CD25�CD4� T cells/recipient.
Table 3. Chronic GVHD with autoimmune manifestations in sublethally irradiated athymic BALB/cnu/nu and adult-thymectomized BALB/crecipients given DBA/2 donor cells
Host* Hair loss†Proteinuria,
����
Anti-dsDNA IgG1,U/mL
Anti-dsDNA IgG2a,U/mL
BALB/cnu/nu NA 12/12 2583 360 1650 274
Adult-thymectomized BALB/c 8/8 8/8 2708 412 1119 172
Sham-thymectomized BALB/c 8/8 8/8 2099 482 1229 286
Recipients received 50 � 106 donor cells. Data in the last 2 columns are presented as mean SE.*All of the recipients developed severe proteinuria (4�) by 15 to 20 days after donor-cell injection, and serum levels of anti-dsDNA antibodies were measured on day 15
after donor-cell injection.†Greater than 2 cm2 of hair loss in the neck and back by day 30 after donor-cell injection.
2998 ZHANG et al BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom
least several months. In addition, it is still unknown whether or notthe features of chronic GVHD in those BALB/c recipients werecaused by autoimmunity and/or alloimmunity, although thoserecipients showed high levels of serum anti-dsDNA.
Donor-type autoreactive CD4� T cells were reported to play acritical role in induction of chronic GVHD.16,48 However, it is stillunclear how the autoreactive T cells are generated in chronicGVHD recipients. It was previously proposed that those donor-typeautoreactive T cells were de novo developed from recipients’thymuses that were deficient in negative selection of autoreactive Tcells and/or deficient in production of autoregulatory T cells due tothymic damage caused by acute GVHD.31,39,40,48-50 However, it wasalso reported that chronic GVHD recipients did not have a defect inthymic negative selection.51 In our report, induction of chronicGVHD did not require host thymus or de novo–developed donor
CD4� T cells but required mature CD4� T cells in transplants fromnonautoimmune donors. It is not yet clear how donor alloreactiveCD4� T cells in transplants lead to the development of chronicGVHD with autoimmune manifestations. We hypothesize thatalloimmune responses lead to the activation and expansion ofquiescent autoreactive CD4� T cells in transplants from nonautoim-mune donors. Quiescent autoreactive T cells are found in theperiphery of healthy individuals.52
This is the first report that B cells in the transplants fromnonautoimmune donors play a critical role in the pathogenesis ofchronic GVHD, although it was reported that B cells augmentedacute GVHD53 and that depletion of B cells using anti-CD20 mAbameliorated refractory chronic GVHD in some patients.7,8 In thecurrent report, mature donor B cells in transplants but not denovo–developed donor B cells are required for autoantibody
Figure 5. Natural CD25�CD4� regulatory T cells prevented chronic GVHD. (A) DBA/2 spleen cells were stained with anti-TCR��, CD4, and CD25 and then CD4�TCR���
cells were sorted into CD25� and CD25� CD4� T cells. (B) CD25�CD4�T cells (0.1 � 106) were stimulated with anti-CD3 mAb and cocultured with titrated numbers ofCD25�CD4� T cells for 72 hours; the proliferation of CD25�CD4� T cells was measured by 3H-TdR incorporation. Mean SE of triplicate culture, and one representative of 3replicate experiments are shown. (C) Sublethally irradiated BALB/c mice were injected with CD4� T-cell–depleted DBA/2 spleen cells (CD4-dep alone; 100 � 106), CD4-depplus CD25�CD4� T cells (CD25�; 5 � 106), and CD4-dep plus CD25� and CD25�CD4� T cells (CD25�; 2.5 � 106). The development of proteinuria and hair loss in the 3groups were compared. There were 12 mice in each group. (D) HE staining of skin tissue sections of the recipients given CD4-dep cells, CD4-dep plus CD25�CD4� T cells, orCD4-dep plus CD25� and CD25� CD4� T cells 30 days after cell injection. One representative is shown of 4 examined recipients in each group. (E) Seven days after cellinjection, yield of donor CD4� T cells from the spleens of recipients given donor CD25�CD4� T cells and recipients given both CD25� and CD25� CD4� T cells was measured.Mean SE of 4 recipients in each group is shown. (F) Sorted donor CD4� T cells (0.3 � 106/well) from recipient in panel E were cultured in vitro for 5 days without additionalstimulation. T-cell proliferation was measured with 3H-TdR incorporation. Mean SE of 4 examined recipients in each group is shown. (G) Intracellular staining of IFN-� inCD4� T cells from the spleens of recipients injected with CD25�CD4� T cells (bold solid line) or from the recipients injected with both CD25� and CD25� CD4� T cells (thin solidline) 7 days after cell injection. The isotype control staining was shown as thin dashed line. One representative is shown of 4 examined recipients in each group.
Table 4. CD25�CD4� T cells suppress autoantibody production in chronic GVHD BALB/c recipients on day 15 after donor-cell injection
Donor cellsTotal IgG1,
�g/mLTotal IgG2a,
�g/mLAnti-dsDNAIgG1, U/mL
Anti-dsDNAIgG2a, U/mL
Anti-dsDNAIgG2aa, U/mL
CD4�-SPL* � CD25�CD4� T† 1678 207 2119 218 1464 239 700 79 �10
CD4�SPL � CD25�CD4� T � CD25�CD4� T‡ 1279 149 1994 166 1097 167 308 38 �10
Data are mean SE, n � 12.*100 � 106 CD4� T cell–depleted spleen cells/recipient.†5 � 106 CD25�CD4� T cells/recipient.‡2.5 � 106 CD25�CD4� T cells/recipient.
CD4� T/B CELLS IN TRANSPLANTS INDUCE CHRONIC GVHD 2999BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom

production and induction of chronic GVHD, since depletion ofmature donor B cells in transplants prevented the autoantibodyproduction and disease induction. The mechanisms whereby donorB cells lead to the development of autoimmune-like chronicGVHD are unclear. We hypothesize that quiescent autoreactivedonor B cells in transplants from nonautoimmune donors areactivated and expanded by alloreactive donor CD4� T cells viarecognition of host minor alloantigens presented by donor B cells.Autoreactive B cells are commonly found in the peripherallymphoid tissues of healthy individuals.54,55
We observed that both donor CD4� T and B cells intransplants were required for induction of chronic GVHD andthat depletion of either one prevented the disease induction. Themechanisms whereby donor CD4� T and B cells interact toinduce chronic GVHD are not yet clear. We recently found thatCD4� T cells from chronic GVHD recipients given wholeDBA/2 spleen cells augmented the in vitro and in vivoautoantibody production by B cells from the chronic GVHDrecipients, however, CD4� T cells from the nonchronic GVHDrecipients given B-cell–depleted DBA/2 spleen cells did notaugment the autoantibody production by the B cells from thechronic GVHD recipients (Supplemental Figure S1, available onthe Blood website; click on the Supplemental Figure link at thetop of the online article). Although further investigation isneeded, these results may indicate that there are autoreactiveCD4� T cells in the chronic GVHD recipients given donorDBA/2 spleen cells and the activation and expansion of donorautoreactive CD4� T cells in the recipients required donor Bcells. It was previously reported that activation of autoreactive Bcells led to activation of autoreactive T cells.56 It has also beenproposed that autoreactive B cells are important antigen-presenting cells that play a central role in amplifying autoim-mune responses and epitope spreading of autoreactive T and Bcells due to the fact that B-cell surface Ig receptors are veryefficient at capturing soluble antigens and one B cell can presentmultiple antigens that activate multiple CD4� T-cell clones.57-60
We observed that the recipients given B-cell–depleted DBA/2 donorspleen cells showed clinical signs of acute GVHD and marked reductionof thymocytes early after donor-cell injection but did not go on todevelop chronic GVHD. In contrast, recipients given whole donorspleen cells developed signs of acute GVHD early after donor-cellinjection and went on to develop chronic GVHD with autoimmunemanifestations. We also found that transplantation of DBA/2 spleen cellsinto sublethally irradiated syngenic DBA/2 recipients did not inducechronic GVHD or autoantibody production. These results indicate thatalloreactive CD4� T cells are required but not sufficient for induction ofchronic GVHD. Our observation is consistent with clinical observationsthat chronic GVHD is not simply a consequence of tissue damagecaused by acute GVHD.
CD25�CD4� Treg cells have been reported to prevent GVHDand autoimmune disease in a variety of models.28,41-44,61,62 Consis-tent with previous reports, we observed that donor CD25�CD4�
Treg cells suppressed donor CD25�CD4� T-cell proliferation and
expansion and IFN-� secretion when coinjected into allogeneicrecipients. The CD25�CD4� Treg cells also suppressed autoanti-body production and prevented induction of chronic GVHD. Theseresults indicate that the breakdown of peripheral tolerance of donorlymphocytes triggered by HCT procedures and alloimmune re-sponses can be restored by infusion of large numbers of donorCD25�CD4� T cells. It is noteworthy that our data showed thatadd-back of donor CD25�CD4� Treg cells to CD25�CD4� T cellsmarkedly reduced the incidence of proteinuria and chronic GVHDtissue damage, but the reduction of anti-dsDNA was relativelylimited. These results may indicate that CD25�CD4� T cells do notcompletely suppress the in vivo function of donor CD25�CD4� Tcells so that some of the latter CD4� T cells can still activate donorB cells to secrete autoantibodies. These results may also indicatethat autoantibody alone may not play a critical role in the tissuedamage of chronic GVHD. Shlomchik and colleagues (Chan etal59) had demonstrated that secreted autoantibodies are not requiredfor tissue damage in autoimmune lupus mice.
Taken together, the current experimental results can be explained bythe following hypotheses. After injection of DBA/2 donor spleen cellsinto sublethally irradiated BALB/c recipients, host/donor dendritic cellsactivate the alloreactive CD4� T cells, which subsequently activatedonor B cells by recognizing the host alloantigens presented by donor Bcells, including autoreactive B cells that present both alloantigens andnuclear autoantigens. The activated autoreactive B cells then activate theautoreactive CD4� T cells; in turn, the activated autoreactive CD4� Tcells augment the antibody secretion and isotype switch of autoreactiveB cells via costimulatory molecules and their secretion of cytokines (ie,IFN-�). The autoreactive T- and B-cell interaction also leads to theepitope spreading of T and B cells and the amplification of autoimmuneresponse. Finally, autoreactive CD4� T infiltration and/or autoantibody-immune complex deposition in the tissues cause chronic GVHD. Incontrast, Treg cells suppress the autoimmune response at the abovedifferent steps.
In conclusion, our studies indicate that alloreactive CD4� Tcells and quiescent autoreactive donor CD4� T and B cells intransplants from nonautoimmune donors induce chronic GVHDwith autoimmune manifestations and that donor CD25�CD4� Tregcells can prevent the disease induction. Therefore, depletion ofCD25�CD4� T and B cells in transplants and infusion of donorTreg cells may be an effective approach in preventing chronicGVHD in patients.
Acknowledgments
We thank Tammy Huang, Stephen Scott, and Noe Gonzales in ourlaboratory; Lucy Brown and Claudia Spalla at City of Hope (COH)Flow Cytometry Facility; and Sofia Loera at COH AnatomicPathology Laboratory for their excellent technical assistance. Wethank Dr Richard Ermel and his staff at COH Research AnimalFacility for providing excellent animal care and Dr RobertsonParkman for critical review of our manuscript.
References
1. Sullivan KM. Graft-vs-host disease. In: BlumeKG, Forman SJ, Appelbaum FR, eds. Thomas’Hematopoietic Cell Transplantation. Malden, MA:Blackwell Science; 2004:635-664.
2. Lee SJ, Vogelsang G, Flowers ME. Chronic graft-versus-host disease. Biol Blood Marrow Trans-plant. 2003;9:215-233.
3. Lee SJ. New approaches for preventing and
treating chronic graft-versus-host disease. Blood.2005;105:4200-4206.
4. Farag SS. Chronic graft-versus-host disease:where do we go from here? Bone Marrow Trans-plant. 2004;33:569-577.
5. Rouquette-Gally AM, Boyeldieu D, Prost AC,Gluckman E. Autoimmunity after allogeneic bonemarrow transplantation: a study of 53 long-term-
surviving patients. Transplantation. 1988;46:238-240.
6. Lister J, Messner H, Keystone E, Miller R, FritzlerMJ. Autoantibody analysis of patients with graftversus host disease. J Clin Lab Immunol. 1987;24:19-23.
7. Ratanatharathorn V, Ayash L, Reynolds C, etal. Treatment of chronic graft-versus-host
3000 ZHANG et al BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom

disease with anti-CD20 chimeric monoclonalantibody. Biol Blood Marrow Transplant. 2003;9:505-511.
8. Canninga-van Dijk MR, van der Straaten HM,Fijnheer R, Sanders CJ, van den Tweel JG, Ver-donck LF. Anti-CD20 monoclonal antibody treat-ment in 6 patients with therapy-refractory chronicgraft-versus-host disease. Blood. 2004;104:2603-2606.
9. Ferrara J, Antin, J. The pathophysiology of graft-vs-host disease. In: Blume KG, Forman SJ, Ap-pelbaum FR, eds. Thomas’ Hematopoietic CellTransplantation. Malden, MA: Blackwell Science;2004:353-368.
10. van der Veen FM, Rolink AG, Gleichmann E. Au-toimmune disease strongly resembling systemiclupus erythematosus (SLE) in F1 mice undergo-ing graft-versus-host reaction (GVHR). Adv ExpMed Biol. 1982;149:669-677.
11. Morris SC, Cheek RL, Cohen PL, Eisenberg RA.Autoantibodies in chronic graft versus host resultfrom cognate T-B interactions. J Exp Med. 1990;171:503-517.
12. Via CS, Shearer GM. T-cell interactions in auto-immunity: insights from a murine model of graft-versus-host disease. Immunol Today. 1988;9:207-213.
13. Morris SC, Cheek RL, Cohen PL, Eisenberg RA.Allotype-specific immunoregulation of autoanti-body production by host B cells in chronic graft-versus host disease. J Immunol. 1990;144:916-922.
14. Sekiguchi DR, Jainandunsing SM, Fields ML, etal. Chronic graft-versus-host in Ig knockin trans-genic mice abrogates B cell tolerance in anti-double-stranded DNA B cells. J Immunol. 2002;168:4142-4153.
15. Sekiguchi DR, Eisenberg RA, Weigert M. Sec-ondary heavy chain rearrangement: a mecha-nism for generating anti-double-stranded DNA Bcells. J Exp Med. 2003;197:27-39.
16. Parkman R. Clonal analysis of murine graft-vs-host disease, I: phenotypic and functional analy-sis of T lymphocyte clones. J Immunol. 1986;136:3543-3548.
17. Jaffee BD, Claman HN. Chronic graft-versus-hostdisease (GVHD) as a model for scleroderma, I:description of model systems. Cell Immunol.1983;77:1-12.
18. Claman HN, Jaffee BD, Huff JC, Clark RA.Chronic graft-versus-host disease as a model forscleroderma, II: mast cell depletion with deposi-tion of immunoglobulins in the skin and fibrosis.Cell Immunol. 1985;94:73-84.
19. Zhang Y, McCormick LL, Desai SR, Wu C, GilliamAC. Murine sclerodermatous graft-versus-hostdisease, a model for human scleroderma: cutane-ous cytokines, chemokines, and immune cell acti-vation. J Immunol. 2002;168:3088-3098.
20. Hamilton BL. L3T4-positive T cells participate inthe induction of graft-vs-host disease in responseto minor histocompatibility antigens. J Immunol.1987;139:2511-2515.
21. Liang Y, Huang T, Zhang C, et al. Donor CD8� Tcells facilitate induction of chimerism and toler-ance without GVHD in autoimmune NOD miceconditioned with anti-CD3 mAb. Blood. 2005;105:2180-2188.
22. Zeng D, Liu Y, Sidobre S, Kronenberg M, StroberS. Activation of natural killer T cells in NZB/Wmice induces Th1-type immune responses exac-erbating lupus. J Clin Invest. 2003;112:1211-1222.
23. Zeng D, Lewis D, Dejbakhsh-Jones S, et al. Bonemarrow NK1.1(-) and NK1.1(�) T cells recipro-cally regulate acute graft versus host disease. JExp Med. 1999;189:1073-1081.
24. Zeng D, Dick M, Cheng L, et al. Subsets of trans-genic T cells that recognize CD1 induce or pre-vent murine lupus: role of cytokines. J Exp Med.1998;187:525-536.
25. Zeng D, Lee MK, Tung J, Brendolan A, Strober S.Cutting edge: a role for CD1 in the pathogenesisof lupus in NZB/NZW mice. J Immunol. 2000;164:5000-5004.
26. Via CS, Sharrow SO, Shearer GM. Role of cyto-toxic T lymphocytes in the prevention of lupus-likedisease occurring in a murine model of graft-vs-host disease. J Immunol. 1987;139:1840-1849.
27. Pierres M, Rebouah JP, Liabeuf A, Dosseto M.Mouse monoclonal antibody against Lyt-1.1 al-loantigen. J Immunogenet. 1981;8:67-71.
28. Anderson BE, McNiff JM, Matte C, Athanasiadis I,Shlomchik WD, Shlomchik MJ. Recipient CD4�T cells that survive irradiation regulate chronicgraft-versus-host disease. Blood. 2004;104:1565-1573.
29. Chen F, Maldonado MA, Madaio M, EisenbergRA. The role of host (endogenous) T cells inchronic graft-versus-host autoimmune disease.J Immunol. 1998;161:5880-5885.
30. Choudhury A, Maldonado MA, Cohen PL, Eisen-berg RA. The role of host CD4 T cells in thepathogenesis of the chronic graft-versus-hostmodel of systemic lupus erythematosus. J Immu-nol. 2005;174:7600-7609.
31. Parkman R. Chronic graft-versus-host disease.Curr Opin Hematol. 1998;5:22-25.
32. Tsoi MS, Storb R, Jones E, et al. Deposition ofIgM and complement at the dermoepidermaljunction in acute and chronic cutaneous graft-vs-host disease in man. J Immunol. 1978;120:1485-1492.
33. Hahn BH. Antibodies to DNA. N Engl J Med.1998;338:1359-1368.
34. Kotzin BL. Systemic lupus erythematosus. Cell.1996;85:303-306.
35. Huang CM, Parsons M, Oi VT, Huang HJ, Her-zenberg LA. Genetic characterization of mouseimmunoglobulin allotypic determinants (allotopes)defined by monoclonal antibodies. Immunogenet-ics. 1983;18:311-321.
36. Anderson RE, Olson GB, Autry JR, Howarth JL,Troup GM, Bartels PH. Radiosensitivity of T andB lymphocytes, IV: effect of whole body irradia-tion upon various lymphoid tissues and numbersof recirculating lymphocytes. J Immunol. 1977;118:1191-1200.
37. Durum SK, Gengozian N. The comparative radio-sensitivity of T and B lymphocytes. Int J RadiatBiol Relat Stud Phys Chem Med. 1978;34:1-15.
38. Pazdernik TL, Nishimura T. Radiosensitivity ofdifferent B and T subpopulations of lymphocytesin the mouse spleen. Agents Actions. 1978;8:229-237.
39. Fukushi N, Arase H, Wang B, et al. Thymus: adirect target tissue in graft-versus-host reactionafter allogeneic bone marrow transplantation thatresults in abrogation of induction of self-toler-ance. Proc Natl Acad Sci U S A. 1990;87:6301-6305.
40. Desbarats J, Lapp WS. Thymic selection and thy-mic major histocompatibility complex class II ex-pression are abnormal in mice undergoing graft-versus-host reactions. J Exp Med. 1993;178:805-814.
41. Cohen JL, Trenado A, Vasey D, Klatzmann D,Salomon BL. CD4(�)CD25(�) immunoregulatoryT cells: new therapeutics for graft-versus-hostdisease. J Exp Med. 2002;196:401-406.
42. Hoffmann P, Ermann J, Edinger M, Fathman CG,Strober S. Donor-type CD4(�)CD25(�) regula-tory T cells suppress lethal acute graft-versus-host disease after allogeneic bone marrow trans-plantation. J Exp Med. 2002;196:389-399.
43. Edinger M, Hoffmann P, Ermann J, et al.CD4�CD25� regulatory T cells preserve graft-versus-tumor activity while inhibiting graft-versus-host disease after bone marrow transplantation.Nat Med. 2003;9:1144-1150.
44. Zeng D, Lan F, Hoffmann P, Strober S. Suppres-
sion of graft-versus-host disease by naturally oc-curring regulatory T cells. Transplantation. 2004;77:S9-S11.
45. Jacob CO, van der Meide PH, McDevitt HO. Invivo treatment of (NZB X NZW)F1 lupus-like ne-phritis with monoclonal antibody to gamma inter-feron. J Exp Med. 1987;166:798-803.
46. Datta SK, Patel H, Berry D. Induction of a cationicshift in IgG anti-DNA autoantibodies: role of Thelper cells with classical and novel phenotypesin three murine models of lupus nephritis. J ExpMed. 1987;165:1252-1268.
47. Ratanatharathorn V, Ayash L, Lazarus HM, Fu J,Uberti JP. Chronic graft-versus-host disease:clinical manifestation and therapy. Bone MarrowTransplant. 2001;28:121-129.
48. Tivol E, Komorowski R, Drobyski WR. Emergentautoimmunity in graft versus host disease. Blood.2005;105:4885-4891.
49. Hollander GA, Widmer B, Burakoff SJ. Loss ofnormal thymic repertoire selection and persis-tence of autoreactive T cells in graft vs host dis-ease. J Immunol. 1994;152:1609-1617.
50. Miura Y, Thoburn CJ, Bright EC, et al. Associationof Foxp3 regulatory gene expression with graft-versus-host disease. Blood. 2004;104:2187-2193.
51. Hakim FT, Payne S, Shearer GM. Recovery of Tcell populations after acute graft-vs-host reaction.J Immunol. 1994;152:58-64.
52. Klein L, Klugmann M, Nave KA, Tuohy VK,Kyewski B. Shaping of the autoreactive T-cell rep-ertoire by a splice variant of self protein ex-pressed in thymic epithelial cells. Nat Med. 2000;6:56-61.
53. Schultz KR, Paquet J, Bader S, HayGlass KT.Requirement for B cells in T cell priming to minorhistocompatibility antigens and development ofgraft-versus-host disease. Bone Marrow Trans-plant. 1995;16:289-295.
54. Bendelac A, Bonneville M, Kearney JF. Autoreac-tivity by design: innate B and T lymphocytes. NatRev Immunol. 2001;1:177-186.
55. Melchers F, Rolink, A. B-lymphocyte developmentand biology. In: Paul W, ed. Fundamental Immu-nology. Philadelphia, PA: Lippincott-Raven; 1998:367-409.
56. Lin RH, Mamula MJ, Hardin JA, Janeway CA Jr.Induction of autoreactive B cells allows priming ofautoreactive T cells. J Exp Med. 1991;173:1433-1439.
57. Roosnek E, Lanzavecchia A. Efficient and selec-tive presentation of antigen-antibody complexesby rheumatoid factor B cells. J Exp Med. 1991;173:487-489.
58. Singh RR, Hahn BH. Reciprocal T-B determinantspreading develops spontaneously in murine lu-pus: implications for pathogenesis. Immunol Rev.1998;164:201-208.
59. Chan OT, Hannum LG, Haberman AM, MadaioMP, Shlomchik MJ. A novel mouse with B cellsbut lacking serum antibody reveals an antibody-independent role for B cells in murine lupus. JExp Med. 1999;189:1639-1648.
60. Shlomchik MJ, Craft JE, Mamula MJ. From T to Band back again: positive feedback in systemicautoimmune disease. Nat Rev Immunol. 2001;1:147-153.
61. La Cava A, Ebling FM, Hahn BH. Ig-reactiveCD4�CD25� T cells from tolerized (New Zea-land Black x New Zealand White)F1 mice sup-press in vitro production of antibodies to DNA.J Immunol. 2004;173:3542-3548.
62. Zheng SG, Wang JH, Koss MN, Quismorio F Jr,Gray JD, Horwitz DA. CD4� and CD8� regula-tory T cells generated ex vivo with IL-2 and TGF-beta suppress a stimulatory graft-versus-host dis-ease with a lupus-like syndrome. J Immunol.2004;172:1531-1539.
CD4� T/B CELLS IN TRANSPLANTS INDUCE CHRONIC GVHD 3001BLOOD, 1 APRIL 2006 � VOLUME 107, NUMBER 7
For personal use only. by guest on December 31, 2011. bloodjournal.hematologylibrary.orgFrom
Related Documents











