Does Context-dependent Memory Reactivation Work in the Waking State? A Music EEG Study Thesis Submitted in partial fulfillment of the requirements for the degree Master of Science Master program in Neurosciences Faculty of Medicine and Faculty of Mathematics and Natural Sciences Rheinische Friedrich-Wilhelms University of Bonn Presented by Alexander Seldon Chockley from Cleveland, USA Bonn, September 2014

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Does Context-dependent Memory Reactivation Work in the Waking State? A Music EEG Study
Thesis Submitted in partial fulfillment of the requirements for the degree
Master of Science
Master program in Neurosciences Faculty of Medicine and
Faculty of Mathematics and Natural Sciences Rheinische Friedrich-Wilhelms University of Bonn
Presented by Alexander Seldon Chockley
from Cleveland, USA
Bonn, September 2014

2
This work has been performed at the Klinik für Epileptologie, Universitätsklinikum Bonn, Universität Bonn, Bonn, Germany In the team of PD Dr. Juergen Fell
1. Referee: PD Dr. Juergen Fell
Cortical Oscillations Group
Klinik für Epileptologie
Universitätsklinikum Bonn
Sigmund-Freud-Straße 25
D-53105 Bonn, Germany
Phone: ++49 0228 / 287-1-9343
Fax: ++49 0228 / 287-1-1559
Email: [email protected]
2. Referee: Prof. Dr. phil. Christoph Helmstaedter
Department of Neuropsychology
Klinik für Epileptologie
Universitätsklinikum Bonn
Sigmund-Freud-Straße 25
D-53105 Bonn, Germany
Phone: ++49 0228 / 287-16108
Fax: ++49 0228 / 287-16294
Email: [email protected]

Index 3
Index
INDEX ......................................................................................................................................................................... 3
1 LIST OF ABBREVIATIONS ............................................................................................................................ 4
2 INTRODUCTION ................................................................................................................................................. 5 2.1 MEMORY CONSOLIDATION ....................................................................................................................................... 5 2.2 MEMORY REACTIVATION .......................................................................................................................................... 6 2.3 CONTEXT-DEPENDENT MEMORY .......................................................................................................................... 7 2.4 CONTEXT-DEPENDENT MEMORY REACTIVATION ............................................................................................. 8 2.5 MUSIC AND THE BRAIN ............................................................................................................................................... 9 2.6 MUSIC AND BRAIN OSCILLATIONS ....................................................................................................................... 10
3 AIMS OF THE PRESENT WORK ............................................................................................................. 13
4 MATERIALS AND METHODS ................................................................................................................... 14 4.1 PARADIGM .................................................................................................................................................................... 14 4.2 STIMULUS PRESENTATION ..................................................................................................................................... 17 4.4 PILOTING ....................................................................................................................................................................... 18 4.5 EEG COLLECTION ..................................................................................................................................................... 19 4.6 DATA ANALYSIS .......................................................................................................................................................... 19
4.6.1 Behavioral data ....................................................................................................................................................... 19 4.6.2 EEG Data ................................................................................................................................................................... 20
5 RESULTS ........................................................................................................................................................... 21 5.1 BEHAVIORAL RESULTS ............................................................................................................................................ 21
5.1.1 Experimental Conditions ................................................................................................................................... 21 5.1.2 Recognition versus Familiarity ...................................................................................................................... 22 5.1.3 Effects of Study, Notice, or Training .......................................................................................................... 23 5.1.4 Testing versus Piloting ....................................................................................................................................... 24
5.2. EEG RESULTS .......................................................................................................................................................... 25
6 DISCUSSION .................................................................................................................................................... 29 6.1 RECOLLECTION VS. FAMILIARITY ......................................................................................................................... 30 6.2 EFFECTS OF MUSIC ON PERFORMANCE ........................................................................................................... 30 6.3 EFFECT OF STUDY ON PERFORMANCE ............................................................................................................. 31 6.4 EEG POWER ANALYSIS .......................................................................................................................................... 32 6.5 CONCLUSIONS AND FUTURE STUDIES .............................................................................................................. 33
7 SUMMARY ......................................................................................................................................................... 36
8 REFERENCES ................................................................................................................................................. 37
9 ACKNOWLEDGEMENTS ............................................................................................................................. 44
10 APPENDICES................................................................................................................................................. 45 A. STUDY ADVERTISEMENT FOR PARTICIPANT RECRUITMENT ....................................................................... 45 B. POST-EXPERIMENT QUESTIONNAIRE ................................................................................................................... 46
11 AFFIRMATION FOR THE MASTER’S THESIS .................................................................................. 48
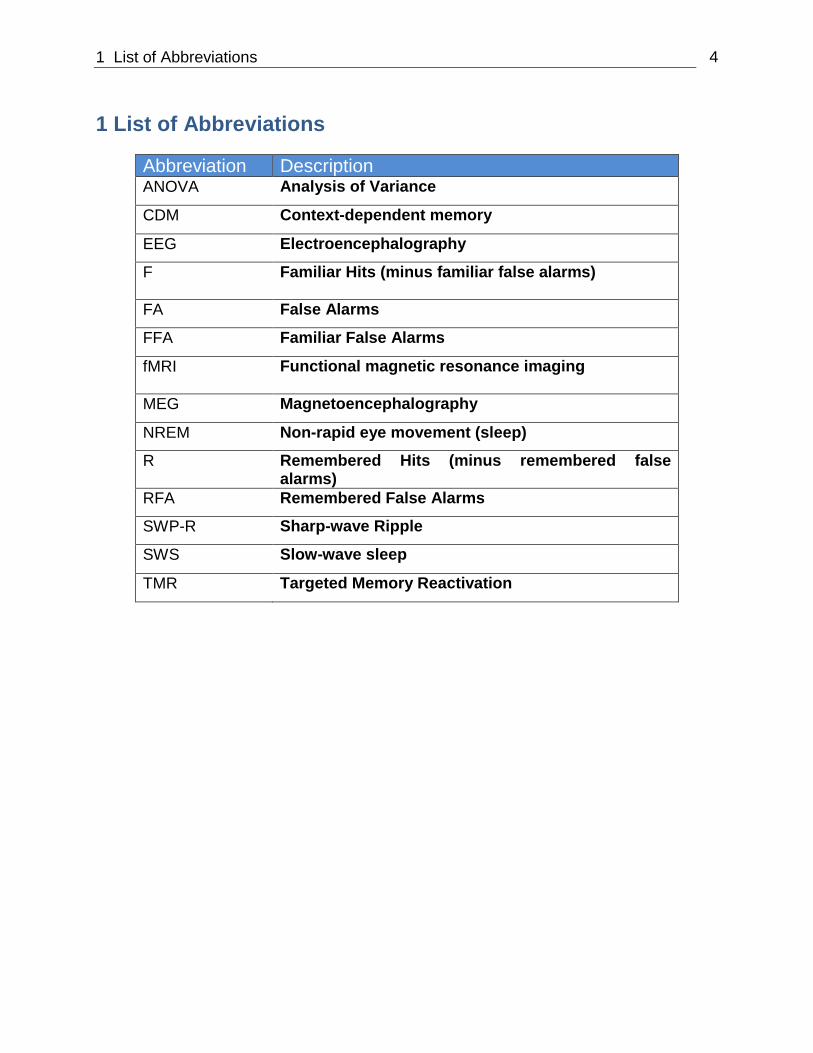
1 List of Abbreviations 4
1 List of Abbreviations
Abbreviation Description ANOVA Analysis of Variance
CDM Context-dependent memory
EEG Electroencephalography
F Familiar Hits (minus familiar false alarms)
FA False Alarms
FFA Familiar False Alarms
fMRI Functional magnetic resonance imaging
MEG Magnetoencephalography
NREM Non-rapid eye movement (sleep)
R Remembered Hits (minus remembered false alarms)
RFA Remembered False Alarms
SWP-R Sharp-wave Ripple
SWS Slow-wave sleep
TMR Targeted Memory Reactivation

2 Introduction 5
2 Introduction
2.1 Memory Consolidation
Memory consolidation is a dynamic process through which memories are strengthened
and made more resistant to interference through reorganization at both the synaptic and
systems levels (Kandel, Schwartz, and Jessell, 2012; Dudai, 2004). Synaptic
consolidation occurs within minutes to hours of learning and involves the stabilization of
changes in synaptic connectivity in localized circuits (for example, through the growth of
new synaptic connections as well as the restructuring of existing ones) (Lynch, 2004).
On a broader scale, system consolidation takes hours to days and is dependent on
reorganization of brain regions supporting memory and their connections (Dudai, 2004;
Kandel, Schwartz, and Jessell, 2012). According to the standard model of memory
consolidation, the creation of new memories is thought to comprise two main steps –
encoding, during which new memories are created, and consolidation, during which
memories are strengthened through increased connectivity (Frankland and Bontempi,
2005). This two-step cycle, however, isn’t the entire story. When previously
consolidated memories are recalled their traces become re-labilized and subsequently
reconsolidated (Tronson and Taylor, 2007). In this way, memory can be thought of as a
dynamic cycle, in which memory traces are consolidated, recalled, and reconsolidated.
Consolidation, whether synaptic or systems, is a largely spontaneous occurrence in the
brain and occurs most commonly during slow-wave sleep (SWS; Stickgold, 2005).
Consolidation during sleep is associated with oscillatory activity in the Delta range (1-4
Hz), likely originating from coordinated activity between the Hippocampus and cortical
areas associated with memory (Hobson and Pace-Schott, 2002). In addition to this
Delta range activity, memory consolidation is closely linked to the occurrence of sharp-
wave ripple complexes (SWP-Rs; Logothetis et al., 2012). These are composed of large
amplitude sharp waves in local field potential and fast field oscillations as first described
by Buzsaki et al. (1992) and occur both during SWS and waking immobility. Disruption
of SWP-Rs in rats by electrical stimulation impairs memory consolidation (Ego-Stengel
and Wilson, 2010), and SWP-Rs are related to behavioral performance in humans
(Axmacher, Elger, and Fell, 2008). These findings suggest that while memory

2 Introduction 6
consolidation mainly occurs during SWS, it also occurs during wakeful rest. The
question remains, however, to what extent waking-state consolidation occurs.
2.2 Memory Reactivation
While it is frequently assumed that slow-wave sleep is necessary for memory
consolidation, it has also been shown that spontaneous reactivation during wakefulness
can occur. Spontaneous reactivation of memory traces during wakefulness has been
shown on the cellular level in rats, (Karlsson and Frank, 2009), monkeys (Skaggs et al,
2007), and in humans (Staresina et al. 2013). While the majority of memory
consolidation is thought to occur spontaneously, many studies have also demonstrated
experimental manipulation of memory reactivation. In these studies, subjects learn
associations between stimuli during an encoding phase. Following this, parts of some
stimulus pairs are replayed during a consolidation phase, and then memory
performance is compared for reactivated and non-reactivated stimulus pairs. This is
aptly named Targeted Memory Reactivation (TMR), and these studies are typically
performed during SWS.
Many TMR studies show that this effect does not work during waking-state
consolidation; however, because memory consolidation has been demonstrated during
wakefulness, it is possible that TMR also works during wakefulness. The reason why
the effect hasn’t been seen thus far could simply be due to relatively weak stimulus
associations or a potential threshold difference between what is required for reactivation
during wakefulness compared to SWS. Diekelmann et al. (2011) suggested that the
main reason waking-state TMR doesn’t work is because when memories are
reactivated during wakefulness they are made labile and are therefore susceptible to
interference (Diekelmann et al. 2011). This conclusion is also supported by their results
that an interference task presented after reactivation weakened spatial memories during
wakefulness and not during SWS. Further, it has been suggested that waking-state
consolidation uses different mechanisms than that during SWS, with glucocorticoid-
mediated consolidation being one of them (Roozendaal, 2000). Plihal and Born (1999),
for example, demonstrated that experimentally raising glucocorticoid levels during
retention sleep (a 3 h post-learning nap) led to reduced episodic memory performance

2 Introduction 7
as compared to placebo controls on a paired associate word task. This effect was
shown to selectively interfere with the consolidation process, and not with acquisition or
retrieval. The glucocorticoid system is generally inhibited during early nocturnal sleep
and during SWS, with a potential reason being related to memory consolidation (Plihal
and Born, 1999). Considering that consolidation of episodic memories has been shown
to occur during wakefulness (when the glucocorticoid system is not inhibited), it is
possible that different mechanisms are responsible for waking and SWS consolidation.
Regardless of the exact mechanisms, studies on TMR have shown that memory
consolidation isn’t necessarily always a spontaneous process and can be
experimentally manipulated (review: Oudiette and Paller, 2013).
2.3 Context-Dependent Memory
Targeted memory reactivation studies generally use specific stimulus-stimulus pairings
and reactivate specific memories (for example, each image learned is paired with a
unique sound; Oudiette et al., 2013). In attempts to understand how memories can be
cued under slightly more natural conditions, many researchers have investigated the
topic of context-dependent memory (CDM; review: Smith, 2001). Considering the fact
that whenever a memory is created in nature it is set in a general environmental
context, by using a broader “stimulus-stimulus” pair researchers can assess what
occurs under more natural conditions. CDM is a phenomenon in which memory
performance can be improved by repeating the context in which stimuli were learned.
This is a very similar phenomenon to that which is manipulated in TMR, but rather than
specific stimulus associations a specific stimulus is encoded together with a more
general context. One of the most common findings of CDM has been that recall of
learned material is better if testing occurs in the original learning context rather than in a
different context (Smith et al., 1978). This phenomenon has been demonstrated in the
1970s and ‘80s for a large variety of contexts, including physical environment (Godden
and Baddeley, 1975), location (Smith, 1979), circadian rhythm state (Holloway and
Wansley, 1978), alcohol/drug states (Eich, 1980), mood states (Bartlett and Santrock,
1979), and music (Smith, 1985), among others. CDM is thought to involve multiple brain
regions, most notably the hippocampus and prefrontal cortex (review: Maren, Phan, and
Liberzon, 2013). The majority of studies on context-dependent memory only test the

2 Introduction 8
effects of repeating context during recall, and not on the effects that the context alone
could have on memory consolidation or reactivation. Further, the majority of studies into
context-dependent memory use a delayed recall of about 48 hours, which means there
can be no conclusions drawn about the wakefulness versus SWS consolidation
question. In an experiment investigating the effects of music context on verbal memory,
Balch, Bowman, and Mohler (1992) used an immediate recall task (with a short delay of
a numerical distraction task) and repeated effects that were shown in the original
instance of the experiment using a 48-hour delay (Smith, 1985). Assuming that the
effects of repeated context on memory performance are due to increased consolidation
of the context-stimulus relationships, this suggests possible manipulations of waking
state consolidation through use of context.
2.4 Context-dependent memory reactivation
In a hallmark study by Rasch et al. (2007), introducing learned context during SWS
improved memory performance. In this study, subjects were presented with a rose-
scented odor while they learned object locations in 2D space. To investigate the
necessity of SWS on memory reconsolidation and the effectiveness of using context for
reactivation, subjects were presented with the odor a second time either during SWS,
non-rapid eye movement (NREM) sleep, or wakefulness. The authors found that
memory performance was significantly improved only in the SWS group, suggesting that
memory reactivation by use of repeated context can only work during SWS, however
testing in the waking-state group occurred after a sleep period, which doesn’t fully test
the effects of waking-state reactivation. In a follow-up fMRI experiment, left hippocampal
activation was higher in the SWS group than in the waking-state group, suggesting that
the effects of reactivation are hippocampus-dependent (Rasch et al. 2007). Although
reactivation during wakefulness did not result in better performance, it is worth
investigating if context-dependent reactivation can work with an immediate recall test
and with different types of context. The present study, therefore, intends to investigate
the effects of context on reconsolidation with music as the main context and without a
sleeping period before testing. Thus far, no studies of a similar design have used music
as context, even though it has been shown to be appropriate for context-dependent
memory effects.

2 Introduction 9
2.5 Music and the brain
Music is a phenomenon shared by all human cultures (Andrade and Bhattacharya,
2003), and recent research suggests that it can provide both health and psychological
benefits to listeners. It has been shown, for example, to alleviate symptoms of anxiety
and depression (Siedlecki and Good, 2006), help language acquisition in impaired
children (Cassileth, Vickers, and Magill 2003), and enhance students’ academic
performance on exams (Rickard, Toukhsati, and field 2005). Additionally, the mere
presence of music has been shown to improve cognitive functioning, as several studies
have shown increases in attention, memory, mental arithmetic, and learning during
music listening (Wallace, 1994; Thompson, Schellenberg, and Husain, 2001; Hallam,
Price, and Katsarou, 2002; Särkämö et al., 2008). The exact mechanisms of how music
enhances cognitive abilities are unknown, but it is suggested to be through aiding
concentration (Hall 1952), exerting calming effects on the listener (Savan, 1999), or by
providing attentional cues (Ferreri et al., 2013). Students are notorious for listening to
music while studying, and it is therefore important that the way in which music affects
learning and cognition is understood.
Music has been shown to have quite global effects on the brain through increasing
activity in multiple areas. Processing of music is distributed across many brain regions
including the brainstem, thalamus, hippocampus, insula, auditory cortex, limbic system,
and multimodal cortical association areas (Andrade and Bhattacharya, 2003; Bamiou,
Musiek, and Luxon, 2003; Klostermann, Loui, and Shimamura, 2009; Watanabe,
Yagishita, and Kikyo, 2008). Of these areas, many are known to be involved in memory,
specifically the hippocampus and widely distributed cortical association networks,
suggesting direct influences of music listening on memory network functioning. In a
study that first demonstrated what is now popularly known as “The Mozart Effect”,
exposure to Mozart’s sonata K448 before learning a spatial reasoning task improved
performance relative to listening to a relaxation tape or silence (Rauscher, Shaw, and
Ky, 1993). This effect was shown to only be short-lived (10-15 minutes), but still
demonstrates the potential usefulness of music with learning tasks. Many researchers

2 Introduction 10
have investigated this effect, and the majority have found that it also exists for non-
Mozart classical music and have confirmed that the effect is only short-lived (review:
Jenkins, 2001). In addition to listening to music before performing a task, hearing music
during a non-spatial verbal memory task has been shown to improve performance. In a
within subjects design, hearing upbeat jazz music during encoding improved
performance over non-music conditions (Ferreri, Aucouturier, and Bugaiska, 2013).
Music has been shown to be sufficient for context-dependent retrieval, has global
effects on the brain, and is associated with increased activation in many brain areas.
Therefore, music is likely sufficient for context-dependent reactivation. Spontaneous
consolidation has been shown to occur during wakefulness, and it is possible that
context-dependent reactivation can work during wakefulness. The evidence for context-
dependent reactivation in the waking state isn’t so compelling, but it also is very limited.
The present study is therefore intended to test the effects of music-context-dependent
memory reactivation during a waking rest period following learning.
2.6 Music and Brain Oscillations
As previously mentioned, music is known to activity many different brain areas involved
with cognition and learning. With most studies using neuroimaging techniques, such as
fMRI, and focusing on specific aspects of music, such as tempo, melody, or beat, there
isn’t very much information available on the temporal nature of brain activity during
music listening or how oscillatory networks in the brain are affected by music
presentation at rest. Rhythmic neural activity as measured by electroencephalography
(EEG) or magnetoencephalography (MEG) reflects communication between brain
areas, such as between neocortex and thalamus (Steriade and Llinas, 1998), and is
modulated by sensory and cognitive processes (Hari and Salmelin, 1997; Pfurtscheller
and Lopes da Silva, 1999). Of particular interest to the present study are rhythms in the
alpha (8-12 Hz) and beta (15-30 Hz) ranges. Alpha activity has historically been
associated with offline activity of the visual cortex, as it is strongly reduced upon visual
stimulation or opening the eyes (Adrian and Matthews, 1934), but is somewhat recently
considered to reflect a level of cortical inhibition (Ray and Cole, 1985; Klimesch, 1996;

2 Introduction 11
Pfurtscheller, 2003; Klimesch et al., 2006). In support of this inhibition hypothesis is a
study by Kay et al. (2012) in which subjects listened to self-selected music at rest. EEG
activity was averaged over occipital and parietal electrodes, and it was found that power
in the alpha range was higher during music listening compared to rest. This could fit
with the inhibition theory of occipital alpha activity, in that during complex auditory
stimulation the visual cortex could be down-modulated. Additionally, alpha activity is
positively correlated with relaxation and introspection (Niedermeyer, 1999), for example
during meditation (Fell, Axmacher, and Haupt, 2010), and music listening is likely to
increase internal concentration (Kay et al., 2012). While alpha rhythms have been a
topic of interest in perception and attention research, beta rhythms have not received as
much attention, specifically in auditory research. Beta activity is associated with normal
waking consciousness (Berge, 1938; Niedermeyer, 1999), as well as with movement
suppression in the motor cortex (Baumeister, Barthel, Geiss, and Weiss, 2008; Zhang,
Chen, Bressler, and Ding, 2008), binding of anatomically distributed feature processing
into coherent perceptual objects (Singer, 1999), and sensorimotor interactions
(Niedermeyer, 1999). In a study similar to that from Kay et al. (2012), Nakamura et al.
(1999) also investigated oscillatory activity during music listening at rest. Rather than
self-selected music, however, subjects were presented with traditional Indonesian
Gamelan music. EEG power was averaged across the “posterior two-thirds” of
electrodes, which included temporal, parietal, and occipital electrodes. Beta power was
found to be increased during music listening as compared to silence, and no differences
in alpha power were found. These results strongly contrast those from Kay et al., as in
that study beta power was found to be decreased during the music conditions. This
discrepancy in results could potentially be due to methodological differences, as Kay et
al. averaged power over only occipital and parietal electrodes, while Nakamura et al.
also included temporal electrodes. Additionally, increased beta power during music
listening has been found in the temporal and frontal electrodes, further supporting the
idea of temporal localization of Beta activity (Sarnthein et al., 1997). It is therefore likely
that Beta power is carried mostly by temporal electrodes, and that the alpha power is
represented in occipital and parietal electrodes.

2 Introduction 12
Taken together, the results of the present study are expected to show that music
listening at rest will show increased alpha range power over the occipital electrodes
(corresponding to visual cortex) and concurrently increased beta range power over
temporal electrodes (as an effect of auditory stimulation). Furthermore, as SWP-Rs are
associated with waking state consolidation, if a music context is sufficient to reactivate
and therefore stimulate consolidation, conditions during which the music context is
repeated during a wakeful rest period should show and increased number of SWP-Rs.

3 Aims of the Present Work 13
3 Aims of the present work The aims of this project are to assess context-dependent memory reactivation in the
waking state and to investigate oscillatory activity in the brain during music listening at
rest. This work is submitted in partial fulfillment of the degree of Master of Science in
Neurosciences at the University of Bonn and, as such, is intended to display the
Master’s student’s ability to use proper scientific methods to assess research questions.
The decision to investigate CDM reactivation during wakefulness stemmed from a lack
of available literature directly investigating the problem. There are currently few studies
on CDM reactivation, and all of them specifically address reactivation in SWS.
Additionally, this is the first study of its kind that attempts to use music as learned
context through which to reactivate memories. Further, there currently exists very little
literature on oscillatory activity in the brain during music listening at rest. While there is
rather extensive literature on music perception, most of the studies focus on specific
aspects of music, such as pitch, tempo, or beat processing. It is important to gain a
holistic view of the effects of music on the brain, as this will certainly provide insight not
only into rhythmic neural activity but also the ways in which music perception can
interact with learning, memory, and cognition. This research spans the subjects of
memory, perception, and learning, and is therefore valuable not only to neuroscience,
but also more broadly to psychology and education as a whole.

4 Materials and Methods 14
4 Materials and Methods 24 healthy subjects (6 female, 21-38 years; mean = 25.42, S.D. = 3.84) were recruited
from the local population through posted flyers and advertisement on a university job
forum (Appendix A). All participants were informed verbally and in writing of the purpose
of the study and the methods involved and gave written informed consent. All
participants were paid for their participation. The Ethics Commission of the Medical
Faculty of the University of Bonn approved all experiments. Testing was performed at
the Deutsches Zentrum für Neurodegenerative Erkrankungen (DZNE) in Bonn,
Germany.
During the experiment, subjects heard four pieces from Nocturnes by Frédéric Chopin
(Opus 27, No. 2; Opus 32, No. 2; Opus 37, No. 2; Opus 62, No. 1) performed by Arthur
Rubinstein (2008, EMI Classics) and purchased through iTunes (Apple Inc., California,
USA). All four songs are composed in major keys and contain similar compositions. The
pieces were shortened to five minutes in length using Audacity (version 2.0.5, The
Audacity Team, USA), given a 5-second fade out, and converted to WAV format. The
songs were played through Sennheiser HD 201 headphones (Sennheiser, Hanover,
Germany) at approximately 50dB.
4.1 Paradigm
A face recognition memory paradigm was presented using Presentation software
(Version 17.1, NeuroBehavioralSystems, California, USA) on a Fujitsu Siemens
Lifebook E8310 (Fujitsu-Siemens, Munich, Germany) running Windows XP Professional
2002 (Microsoft, Washington, USA). The paradigm consisted of three experimental
conditions (Repeat, No Repeat, and No Song), each consisting of three phases
(encoding, consolidation, and retrieval). Each subject went through the three-phase
cycle three times, with slight variations between the experimental conditions. In all
conditions music was played during the encoding phase and never played during the
retrieval phase. During the Repeat condition, the same song was played during the
encoding and consolidation phases, in the No Repeat condition a different song was
played during the consolidation phase, and in the No Song condition, no music was

4 Materials and Methods 15
played during the consolidation phase. This required a total of four songs, and their
presentation order was balanced. An example paradigm for one subject is given in
Figure 1.
Figure 1: An example of the paradigm for one participant; Three different stimulus sets were presented with the three experimental conditions (Repeat, No Repeat, and No Song). Music was presented only during the Encoding phase (in all conditions) and the Consolidation phase (in Repeat and No Repeat conditions), with music never being presented during the Retrieval phase. The presentation order of conditions, stimulus sets, and songs was counterbalanced and randomized.
Before each phase, participants were given verbal and written instructions and
confirmed with the experimenter that they understood the task. During the Encoding
phase, subjects were shown 60 individual faces for 2 seconds each with a 2500-
3500ms inter-stimulus interval during which a fixation cross was presented (Figure 2A).
To maintain their attention, subjects were asked to identify the faces as male or female
by clicking the right or left mouse button, respectively. Music presentation began with
the first face and was continuous until the end of the five-minute phase.
During the Consolidation phase, a fixation cross was presented on the screen and
participants were asked to relax, keep their eyes open, and stay awake. To ensure that
participants stayed awake, the experimenter sat behind and to the side of them and
observed them during this phase. During the conditions in which music was presented,
there was first a minute of silence, followed by five minutes of music presentation,
followed by one minute of silence. In the No Song condition, the consolidation phase
was seven minutes of silence.

4 Materials and Methods 16
During the Retrieval phase, subjects were presented with all 60 previously learned
stimuli and 60 new stimuli in a randomized order. Each face was presented for three
seconds, after which three choices appeared on the screen (“Remembered”, “Familiar”,
and “New”; Figure 2B). Subjects were given 2500-3500 msec to respond, and instructed
to respond as quickly as possible. A “remembered” choice indicated that the participant
was certain that the stimulus had been presented in the encoding phase. A “familiar”
choice indicated that they were not certain, but believed that it had been shown in the
encoding phase. A “new” choice indicated that the stimulus had not been seen before.
Figure 2: Visual depiction of the paradigm; (A) Encoding phase – Each face was presented for two seconds, followed by a fixation cross for three seconds. During the presentation of each face, the subject was required to indicate the sex of the individual by clicking the left mouse button for female and the right button for male. (B) Retrieval phase – subjects were shown a face for three seconds and then asked to report if they remember it, think it is familiar, or think it is new. They were given three seconds to respond and requested to respond as quickly as possible.
Presentation order of the three picture sets, three experimental conditions, and four
songs was counter-balanced, thus requiring 24 participants. Due to the extreme number
of participants that would be required (24*6*6 = 824), only the presentation order was
counterbalanced and not the frequency of each song in each experimental condition
with each stimulus set; however, there was not an ostensibly large bias for any of the
presentation combinations.

4 Materials and Methods 17
4.2 Stimulus Presentation
Face stimuli consisted of 360 individual neutral faces selected from the Park Aging Mind
Laboratory Face Stimulus Set (Minear and Park, 2004). Faces with off-centered gaze or
obvious distinguishing characteristics (e.g., pink hair, facial piercings, etc.) were not
used. Individuals ranged in age from 18 to 69, and comprised three race groups –
Caucasian, African-American, and Other (Asian/Indian), with Caucasian being the
majority group (~70%). Sample stimuli can be seen in Figure 3. Generally speaking, the
ability of people to recognize faces of people outside of their race is lower than within
their race, known as the cross-race effect (review: Meisser and Brigham, 2001). This
effect can be reduced by long-term exposure to other races (Chiroro and Valentine,
2007), as well as by explicitly informing participants of the effect (Hugenberg et al.,
2007). In order to avoid contamination of the results, only participants having lived in a
primarily Caucasian society for at least five years were accepted and all participants
were informed of the cross-race effect prior to beginning the experiment. Stimuli
consisted of 58.33% female faces. Three sets of 180 stimuli were formed and balanced
for gender and age. Each set was then split into two sets of 60 stimuli, which would be
used as the “old” and “new” stimuli for each stimulus set. Color images (640 x 480
pixels) were presented at a resolution of 1400 x 1050 pixels.

4 Materials and Methods 18
Figure 3: Four sample face stimuli; Stimuli were taken from the Park Aging Mind Laboratory Face Stimulus Set (Minear and Park, 2004); here, four stimuli are presented to show variation, but only one face was presented at a time to the subjects.
4.3 Questionnaire
Upon completion of all three conditions, subjects were allowed to shower (to remove
EEG-gel) and then completed a questionnaire. This questionnaire was designed to
gather data about musical training, use of music as a study aid, enjoyment/prevalence
of music, and about the experiment itself (Appendix B).
4.4 Piloting
In order to assess the appropriateness of the stimuli and paradigm, as well as any
possible differences between stimulus sets, the paradigm was piloted with six
individuals. Subjects (N=6, 5 Female) were administered the same experiment as
during testing but without music during any of the phases. EEG data was not collected
during piloting, and testing was conducted in the Department of Epileptology, University
Clinic, Bonn, Germany.

4 Materials and Methods 19
4.5 EEG Collection
The EEG was collected from 32 electrodes mounted in an elastic cap (BrainVision
Professional BrainCap-MR32, EasyCap, Herrsching-Breitbrunn, Germany) and
referenced to FCz during recording. The electrode positions were labeled according to
the extended international 10-20 system (Oostenveld and Praamstra, 2001).
Impedances were kept below 25 kΩ. The EEG was amplified and sampled at 1000Hz
using a BrainAmp MR-plus amplifier (Brain Products, Gilching, Germany) powered by a
BrainVision Professional Powerpack (Brain Products), and sent through a BrainVision
Professional USB 2 Adaptor (Brain Products) to the data-collection PC. Data were
collected in BrainVision Recorder software (Version 1.20.0506, Brain Products) on a
Fujitsu Lifebook U772 (Fujitsu, Tokyo, Japan) running Windows 7 Home Premium
(Microsoft, Washington, USA). Stimulus and response triggers were sent from the
stimulus-delivery PC through the USB Adaptor to the data-collection PC. After
recording, the data were re-referenced to an average reference value.
4.6 Data Analysis
4.6.1 Behavioral data
Behavioral data were analyzed using MATLAB (Version 2013b, Mathworks,
Massachusetts, USA). Statistical analyses were performed with SPSS Statistics
(Version 20, IBM, New York, USA). Signal Detection Theory terminology was used to
define behavioral parameters: Hits (remembered or familiar response to “old” image),
false alarms (FA; remembered or familiar response to “new” images), remembered (R;
hits with “remembered” responses), familiar (F; hits with “familiar” responses),
remembered false alarms (RFA; false alarms for “remembered” responses) and familiar
false alarms (FFA; false alarms for “familiar” responses). All hit percentages
(hits/number of “old” stimuli), as well as R and F percentages, were corrected for
guessing by subtracting the FA rates (FA/number of “new” images) and from this point
on are presented as corrected values. All parameters were subjected to one-way
repeated measures Analysis of Variance (ANOVA) tests with the experimental condition
as the repeated measures factor. To assess whether certain subject characteristics had
an effect on performance, two-way mixed factorial ANOVAs were performed on the six

4 Materials and Methods 20
behavioral parameters with condition as a within-subjects factor and either Study
(whether or not the subject listens to music while studying), Training (if the subject has
had formal musical training), or Notice (if the subject noticed the Repeat condition), as
the between-subjects factor. Further, hits and FAs averaged across conditions were
subjected to a one-way repeated measures ANOVA to compare overall differences.
4.6.2 EEG Data
Ideally, all of the EEG data collected would have been analyzed to investigate, for
example, differential subsequent memory effects between conditions, power differences
during all phases of the experiment, or SWP-R counts as markers of consolidation.
However, due to time constraints, only the continuous data from the consolidation
phases were analyzed. This allowed us to investigate oscillatory activity during music
listening at rest as compared between the three experimental conditions. Data were
preprocessed using BrainVision Analyzer software (Version 1.05, Brain Products). Data
were first re-referenced to the average, and then filtered with a Butterworth zero phase
filter (low cutoff = 0.53 Hz, high cutoff = 70.0 Hz, notch filter at 50 Hz), and segmented
to extract consolidation phase data. Data were then exported to the FieldTrip MATLAB
toolbox for EEG/MEG analysis (Oostenveld et al., 2011). They were then manually
inspected and marked for artifacts. Main sources of artifacts were eye blinks, eye
movements, and tonic muscle activity. The EEG from the five minutes of music
presentation (or the corresponding five minutes of silence in the No Song condition) was
then separated into 2000 msec epochs. Epochs containing electrode or movement
artifacts were discarded. After artifact rejection, a continuous wavelet transform was
applied to single epochs in each electrode using Morlet wavelets at 1 Hz intervals in the
1 to 30 Hz range with a wavelet width of 7. Power was derived from the squared
amplitude coefficient of the wavelet transform of the average waveform. Due to a
suggested spatial localization of alpha (8-12 Hz) and beta (15-30 Hz) activity, power
was averaged over temporal (T7, T8, TP9, and TP10) and occipital (O1, O2, and Oz)
electrodes. Power values for both frequency ranges were then compared between the
three experimental conditions and the two electrode location groups using a two-way
repeated measures ANOVA.
5 Results 21
5 Results
5.1 Behavioral Results
5.1.1 Experimental Conditions
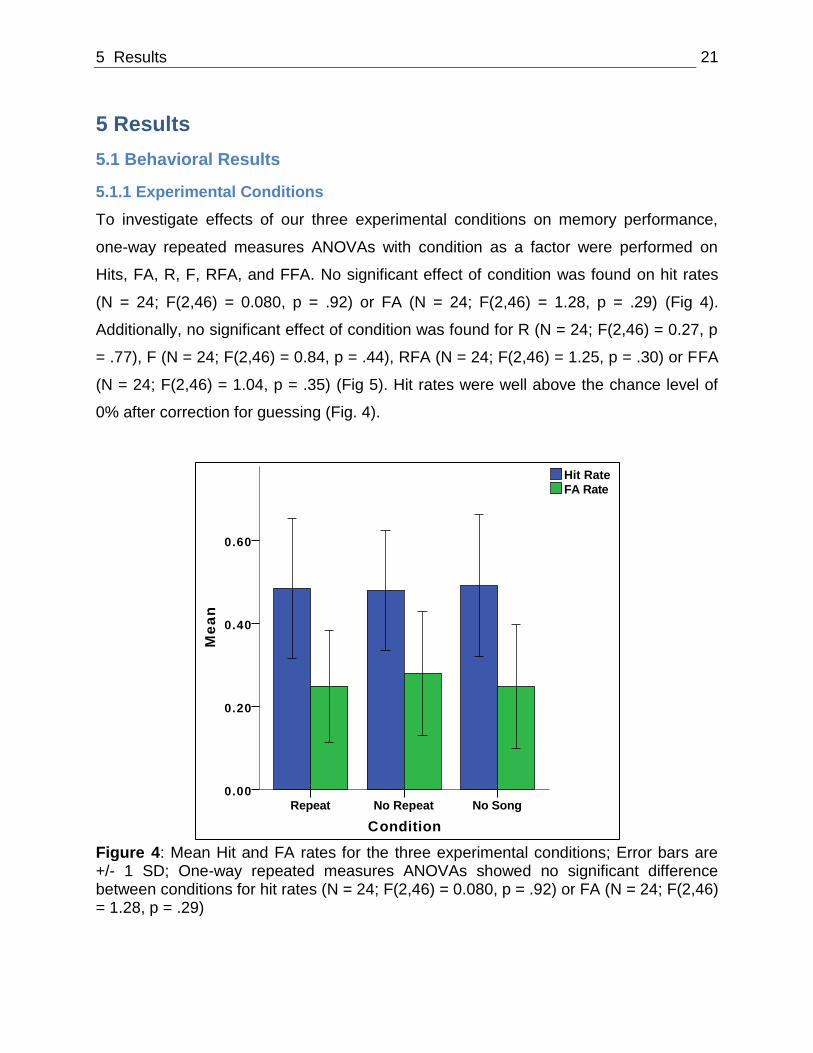
To investigate effects of our three experimental conditions on memory performance,
one-way repeated measures ANOVAs with condition as a factor were performed on
Hits, FA, R, F, RFA, and FFA. No significant effect of condition was found on hit rates
(N = 24; F(2,46) = 0.080, p = .92) or FA (N = 24; F(2,46) = 1.28, p = .29) (Fig 4).
Additionally, no significant effect of condition was found for R (N = 24; F(2,46) = 0.27, p
= .77), F (N = 24; F(2,46) = 0.84, p = .44), RFA (N = 24; F(2,46) = 1.25, p = .30) or FFA
(N = 24; F(2,46) = 1.04, p = .35) (Fig 5). Hit rates were well above the chance level of
0% after correction for guessing (Fig. 4).
Figure 4: Mean Hit and FA rates for the three experimental conditions; Error bars are +/- 1 SD; One-way repeated measures ANOVAs showed no significant difference between conditions for hit rates (N = 24; F(2,46) = 0.080, p = .92) or FA (N = 24; F(2,46) = 1.28, p = .29)
Condition
No SongNo RepeatRepeat
Me
an
0.60
0.40
0.20
0.00
FA Rate
Hit Rate
Page 1

5 Results 22
5.1.2 Recognition versus Familiarity
Averaged across conditions and participants, the number of R responses (N = 24; M =
38.83, SD = 1.57) was higher than F responses (N = 24; M = 21.28, SD = 8.40). A one-
sample t-test showed that this difference was significant (t(23) = 5.32, p < .0001). To
investigate the effectiveness of choosing “remembered” or “familiar”, a two-way
repeated measures ANOVA was conducted with the choice and condition as repeated
measures and found that R rates (M = 0.47, SD = 0.17) were significantly higher than F
rates (M = 0.018, SD = 0.10) ) (Fig 5A, F(1,23) = 7.28, p < ..001). Additionally, RFA (M =
0.10, SD = 0.092) was found to be significantly lower than FFA (M = 0.17, SD = 0.092)
(Fig 5B; F(1,23) = 19.81, p < .001). F rates were not significantly different from the
chance level of 0% (F(1,23) = 1.23, p = .28).
Figure 5: Hit rates (A) and FA rates (B) for “remembered” and “familiar” choices; error bars are +/-1 SD (A) One-way repeated measures ANOVA found no significant effect of condition on R (N = 24; F(2,46) = 0.27, p = .77) or F (N = 24; F(2,46) = 0.84, p = .44). (B) One-way repeated measured ANOVA found no significant effect of condition on RFA (N = 24; F(2,46) = 1.25, p = .30) or FFA (N = 24; F(2,46) = 1.04, p = .35). Two-way repeated measures ANOVAs found R rates (M = 0.47, SD = 0.17) to be significantly higher than F rates (M = 0.018, SD = 0.10) (A; F(1,23) = 7.28, p < .001), and FFAs ( M = 0.17, SD = 0.092) to be significantly higher than RFAs (M = 0.10, SD = 0.092) (B; F(1,23) = 19.81, p < .001). F rates were not significantly different from chance (chance = 0%; F(1,23) = 1.23, p = .28).
5 Results 23
5.1.3 Effects of Study, Notice, or Training
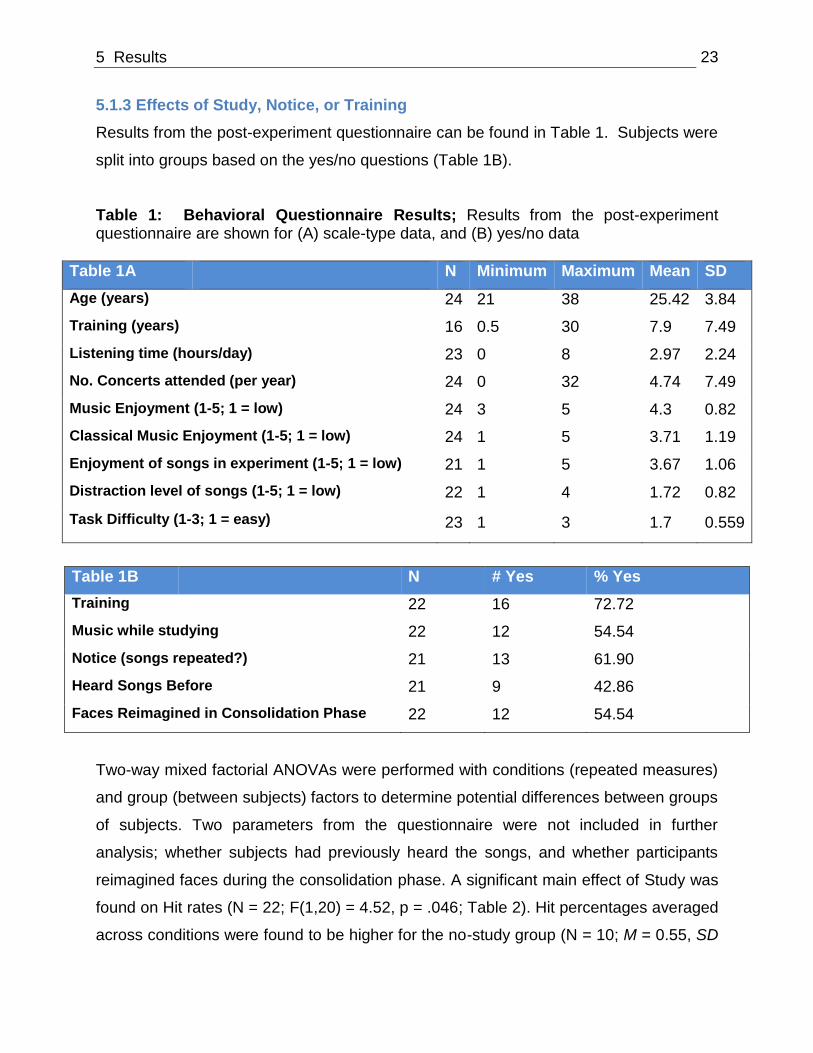
Results from the post-experiment questionnaire can be found in Table 1. Subjects were
split into groups based on the yes/no questions (Table 1B).
Table 1: Behavioral Questionnaire Results; Results from the post-experiment questionnaire are shown for (A) scale-type data, and (B) yes/no data
Table 1A N Minimum Maximum Mean SD
Age (years) 24 21 38 25.42 3.84
Training (years) 16 0.5 30 7.9 7.49
Listening time (hours/day) 23 0 8 2.97 2.24
No. Concerts attended (per year) 24 0 32 4.74 7.49
Music Enjoyment (1-5; 1 = low) 24 3 5 4.3 0.82
Classical Music Enjoyment (1-5; 1 = low) 24 1 5 3.71 1.19
Enjoyment of songs in experiment (1-5; 1 = low) 21 1 5 3.67 1.06
Distraction level of songs (1-5; 1 = low) 22 1 4 1.72 0.82
Task Difficulty (1-3; 1 = easy) 23 1 3 1.7 0.559
Table 1B N # Yes % Yes
Training 22 16 72.72
Music while studying 22 12 54.54
Notice (songs repeated?) 21 13 61.90
Heard Songs Before 21 9 42.86
Faces Reimagined in Consolidation Phase 22 12 54.54
Two-way mixed factorial ANOVAs were performed with conditions (repeated measures)
and group (between subjects) factors to determine potential differences between groups
of subjects. Two parameters from the questionnaire were not included in further
analysis; whether subjects had previously heard the songs, and whether participants
reimagined faces during the consolidation phase. A significant main effect of Study was
found on Hit rates (N = 22; F(1,20) = 4.52, p = .046; Table 2). Hit percentages averaged
across conditions were found to be higher for the no-study group (N = 10; M = 0.55, SD
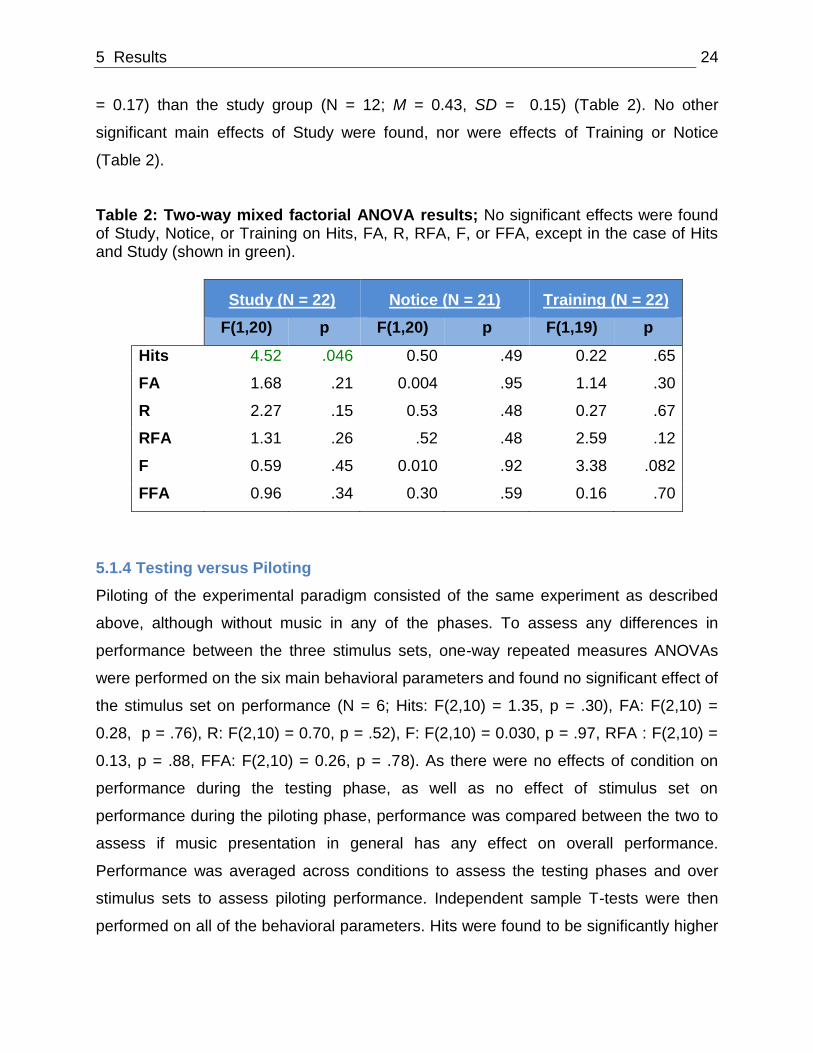
5 Results 24
= 0.17) than the study group (N = 12; M = 0.43, SD = 0.15) (Table 2). No other
significant main effects of Study were found, nor were effects of Training or Notice
(Table 2).
Table 2: Two-way mixed factorial ANOVA results; No significant effects were found of Study, Notice, or Training on Hits, FA, R, RFA, F, or FFA, except in the case of Hits and Study (shown in green).
Study (N = 22) Notice (N = 21) Training (N = 22)
F(1,20) p F(1,20) p F(1,19) p
Hits 4.52 .046 0.50 .49 0.22 .65
FA 1.68 .21 0.004 .95 1.14 .30
R 2.27 .15 0.53 .48 0.27 .67
RFA 1.31 .26 .52 .48 2.59 .12
F 0.59 .45 0.010 .92 3.38 .082
FFA 0.96 .34 0.30 .59 0.16 .70
5.1.4 Testing versus Piloting
Piloting of the experimental paradigm consisted of the same experiment as described
above, although without music in any of the phases. To assess any differences in
performance between the three stimulus sets, one-way repeated measures ANOVAs
were performed on the six main behavioral parameters and found no significant effect of
the stimulus set on performance (N = 6; Hits: F(2,10) = 1.35, p = .30), FA: F(2,10) =
0.28, p = .76), R: F(2,10) = 0.70, p = .52), F: F(2,10) = 0.030, p = .97, RFA : F(2,10) =
0.13, p = .88, FFA: F(2,10) = 0.26, p = .78). As there were no effects of condition on
performance during the testing phase, as well as no effect of stimulus set on
performance during the piloting phase, performance was compared between the two to
assess if music presentation in general has any effect on overall performance.
Performance was averaged across conditions to assess the testing phases and over
stimulus sets to assess piloting performance. Independent sample T-tests were then
performed on all of the behavioral parameters. Hits were found to be significantly higher
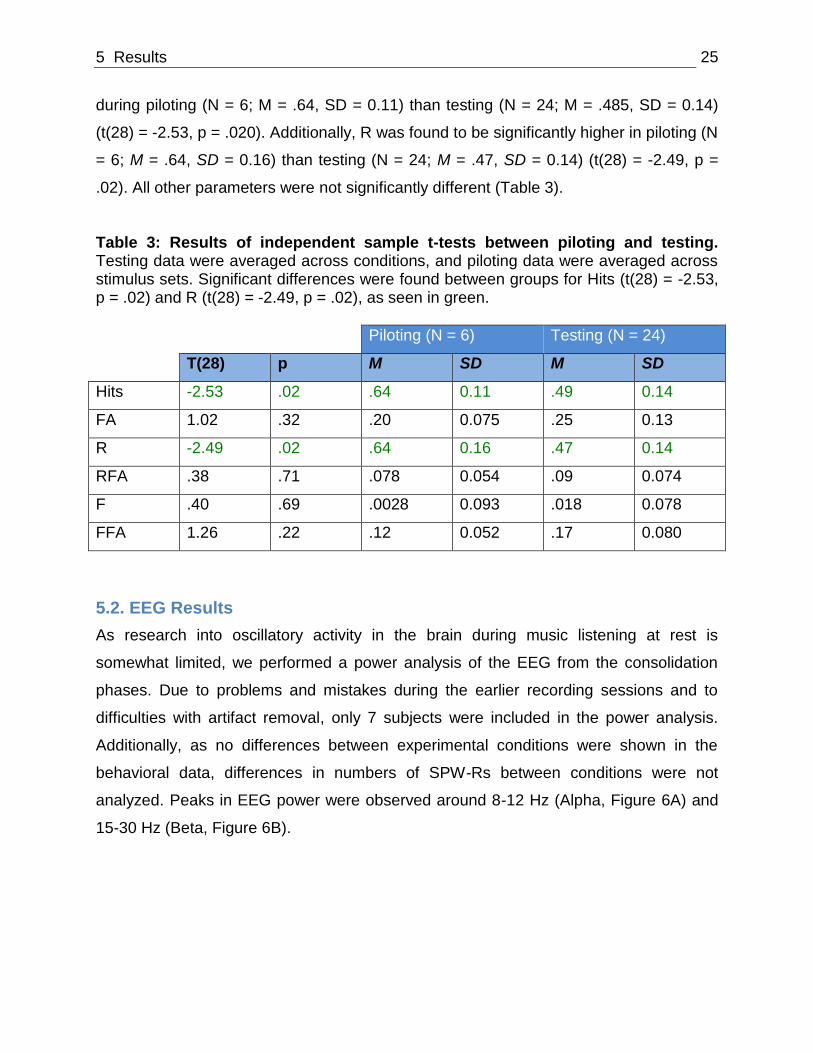
5 Results 25
during piloting (N = 6; M = .64, SD = 0.11) than testing (N = 24; M = .485, SD = 0.14)
(t(28) = -2.53, p = .020). Additionally, R was found to be significantly higher in piloting (N
= 6; M = .64, SD = 0.16) than testing (N = 24; M = .47, SD = 0.14) (t(28) = -2.49, p =
.02). All other parameters were not significantly different (Table 3).
Table 3: Results of independent sample t-tests between piloting and testing. Testing data were averaged across conditions, and piloting data were averaged across stimulus sets. Significant differences were found between groups for Hits (t(28) = -2.53, p = .02) and R (t(28) = -2.49, p = .02), as seen in green.
Piloting (N = 6) Testing (N = 24)
T(28) p M SD M SD
Hits -2.53 .02 .64 0.11 .49 0.14
FA 1.02 .32 .20 0.075 .25 0.13
R -2.49 .02 .64 0.16 .47 0.14
RFA .38 .71 .078 0.054 .09 0.074
F .40 .69 .0028 0.093 .018 0.078
FFA 1.26 .22 .12 0.052 .17 0.080
5.2. EEG Results
As research into oscillatory activity in the brain during music listening at rest is
somewhat limited, we performed a power analysis of the EEG from the consolidation
phases. Due to problems and mistakes during the earlier recording sessions and to
difficulties with artifact removal, only 7 subjects were included in the power analysis.
Additionally, as no differences between experimental conditions were shown in the
behavioral data, differences in numbers of SPW-Rs between conditions were not
analyzed. Peaks in EEG power were observed around 8-12 Hz (Alpha, Figure 6A) and
15-30 Hz (Beta, Figure 6B).
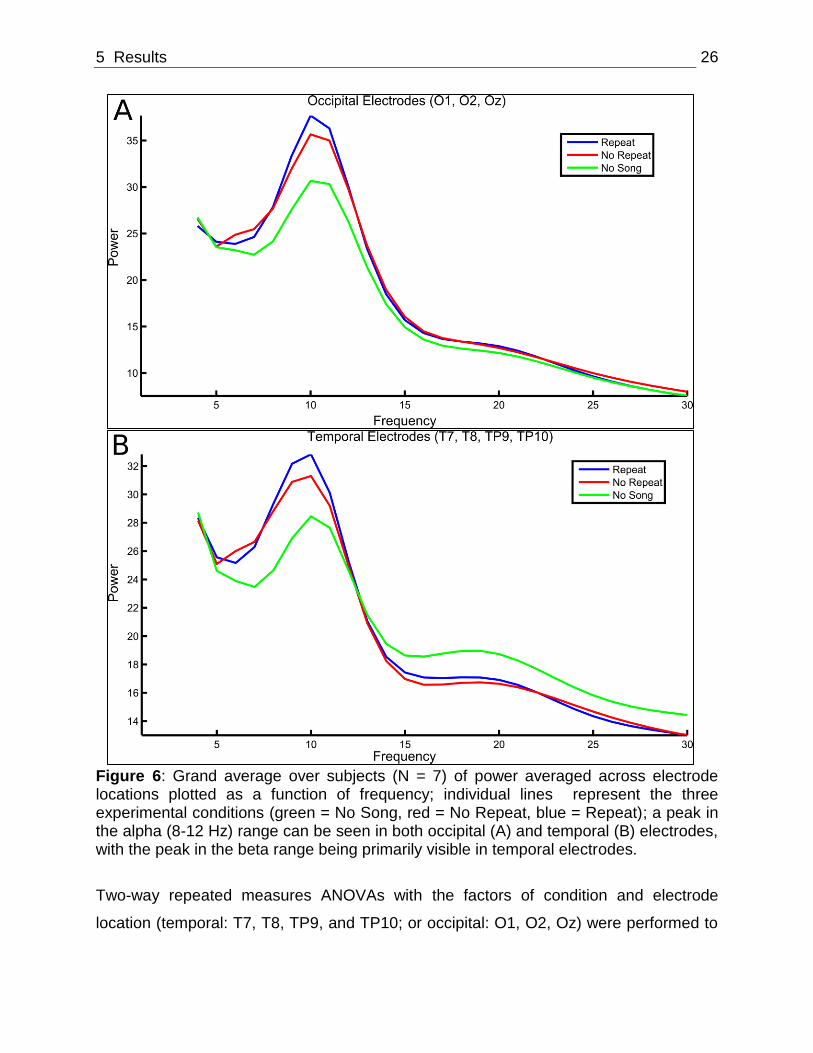
5 Results 26
Figure 6: Grand average over subjects (N = 7) of power averaged across electrode locations plotted as a function of frequency; individual lines represent the three experimental conditions (green = No Song, red = No Repeat, blue = Repeat); a peak in the alpha (8-12 Hz) range can be seen in both occipital (A) and temporal (B) electrodes, with the peak in the beta range being primarily visible in temporal electrodes.
Two-way repeated measures ANOVAs with the factors of condition and electrode
location (temporal: T7, T8, TP9, and TP10; or occipital: O1, O2, Oz) were performed to
5 Results 27
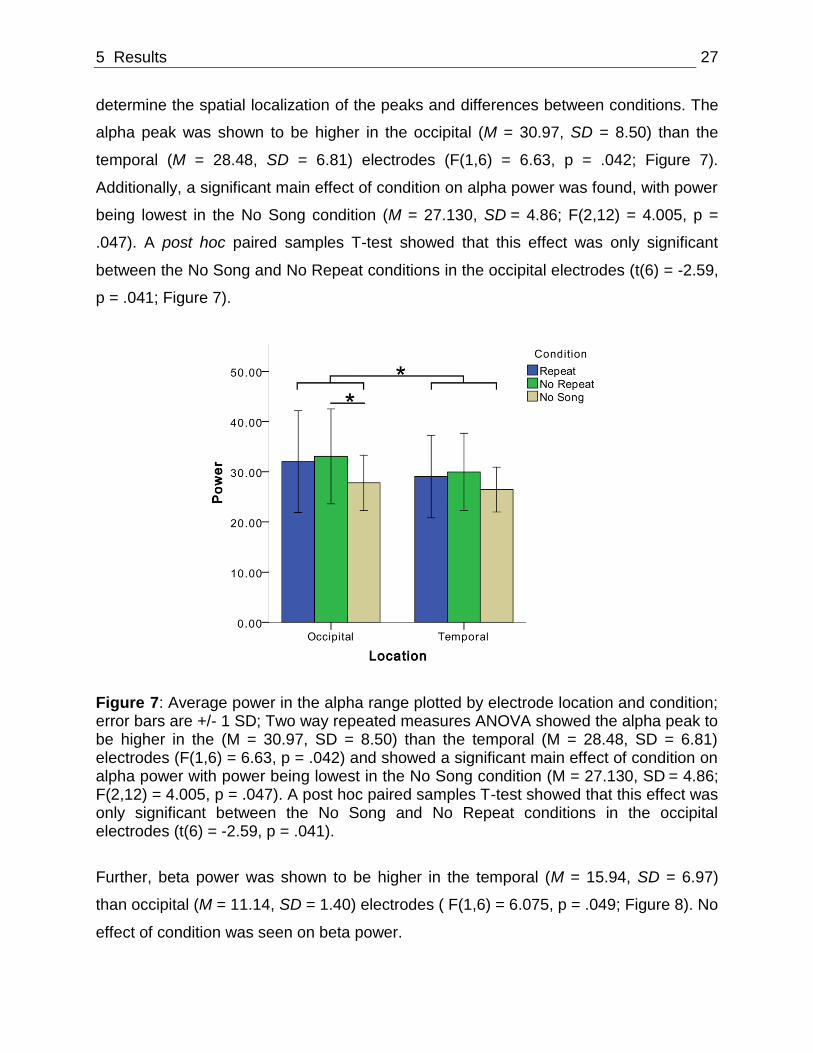
determine the spatial localization of the peaks and differences between conditions. The
alpha peak was shown to be higher in the occipital (M = 30.97, SD = 8.50) than the
temporal (M = 28.48, SD = 6.81) electrodes (F(1,6) = 6.63, p = .042; Figure 7).
Additionally, a significant main effect of condition on alpha power was found, with power
being lowest in the No Song condition (M = 27.130, SD = 4.86; F(2,12) = 4.005, p =
.047). A post hoc paired samples T-test showed that this effect was only significant
between the No Song and No Repeat conditions in the occipital electrodes (t(6) = -2.59,
p = .041; Figure 7).
Figure 7: Average power in the alpha range plotted by electrode location and condition; error bars are +/- 1 SD; Two way repeated measures ANOVA showed the alpha peak to be higher in the (M = 30.97, SD = 8.50) than the temporal (M = 28.48, SD = 6.81) electrodes (F(1,6) = 6.63, p = .042) and showed a significant main effect of condition on alpha power with power being lowest in the No Song condition (M = 27.130, SD = 4.86; F(2,12) = 4.005, p = .047). A post hoc paired samples T-test showed that this effect was only significant between the No Song and No Repeat conditions in the occipital electrodes (t(6) = -2.59, p = .041).
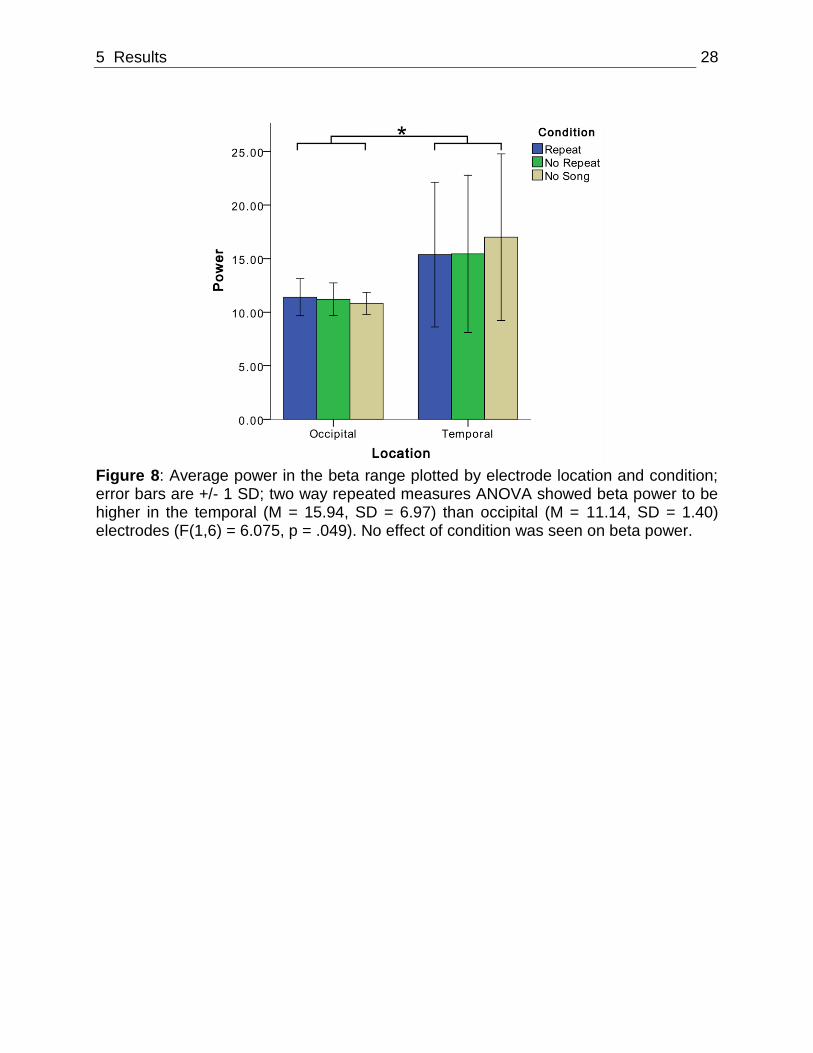
Further, beta power was shown to be higher in the temporal (M = 15.94, SD = 6.97)
than occipital (M = 11.14, SD = 1.40) electrodes ( F(1,6) = 6.075, p = .049; Figure 8). No
effect of condition was seen on beta power.
5 Results 28
Figure 8: Average power in the beta range plotted by electrode location and condition; error bars are +/- 1 SD; two way repeated measures ANOVA showed beta power to be higher in the temporal (M = 15.94, SD = 6.97) than occipital (M = 11.14, SD = 1.40) electrodes (F(1,6) = 6.075, p = .049). No effect of condition was seen on beta power.

6 Discussion 29
6 Discussion Music has been shown to be sufficient for context-dependent retrieval, has global
effects on the brain, and is associated with increased activation in many brain areas.
Therefore, it is likely sufficient for context-dependent reactivation. The present study
intended to test the effectiveness of using music as a context to reactive declarative
memory during wakefulness. The behavioral results presented here demonstrate that
there was no difference between the three experimental conditions (Repeat, No Repeat,
and No Song) for any of the behavioral parameters tested (Hits, FA, R, F, RFA, and
FFA). This is not a surprising result, considering that other studies investigating waking-
state TMR have also not seen positive results (Rasch et al. 2007; Rudoy et al., 2009;
Diekelmann 2011). Additionally, a previous study very similar to the present study also
showed no effects of repeated music context on memory performance in wakefulness
(Rebecca Spencer, personal correspondence). Further, CDM reactivation using an odor
as context also found no effect of reactivating memory traces during a wakeful rest
period (Rasch et al. 2007), and there are several reasons why this might not work. First
of all, it is likely that waking state and SWS consolidation use different mechanisms
(Plihal and Born, 1999). Due to this difference, it is also possible that CDM reactivation
is not sufficient to stimulate the consolidation of memory traces in the waking state.
Second, as reactivating memories in the waking state relabilizes them, thus making
them more susceptible to interference, it is possible that the stimulus-context
relationships were too weak to be resistant to interference after reactivation
(Diekelmann, 2011). This could potentially be improved by increasing the strength of
these relationships, possibly through more stimulus repetitions during encoding. Studies
into TMR and CDM reactivation often require subjects to reach a learning criterion
before proceeding with reactivation (Rasch et al., 2007; Hupbach et al., 2008), likely
leading to memory traces that, even when relabilized, are more resistant to interference.
Moreover, it is possible that the consolidation phase in the present study – at only 7
minutes in length – was simply too short to support reactivation and consolidation of
memory traces. Hupbach et al. (2007), for example, showed that reactivated memories
were more strongly affected by an intrusion task immediately after reactivation than

6 Discussion 30
after a 48h delay. To improve on this, a similar study with longer waking consolidation
phases should be carried out.
Music context has been shown to be sufficient for CDM effects during recall, but not yet
for CDM reactivation. In order to fully test this, it would be useful to perform a study
comparing reactivation during the waking state and during SWS. The evidence for
targeted memory reactivation working only during SWS is overwhelming (Rasch et al.,
2007; Hupbach et al., 2008; Rudoy et al., 2009; Diekelmann, 2011; Alveires et al.,
2012), and it is therefore most likely that this phenomenon does not exist in the waking
state. The resources to carry out such a study were not available here; however, this
would allow for the dissociation between CDM reactivation using music during SWS or
waking state
6.1 Recollection vs. Familiarity
Dual process models of recognition memory (Atkinson and Juola, 1973, 1974; Mandler,
1980; Jacoby, 1991; Yonelinas, 1994) propose that there are two distinct routes with
which one can recognize a previously seen face: familiarity and recollection. Most
people can relate to the experience of meeting someone and finding their face familiar
but being unable to remember details from when or where one might have met them;
this is an example of familiarity-based recognition. Recollection, on the other hand, is
characterized by remembering some form of contextual detail, such as specific previous
encounters. Traditional dual process models propose that familiarity can vary in strength
whereas recollection is usually assumed to be an all-or-nothing, high strength memory.
It is not surprising, then, that in the present study, recollection (“remembered”
responses) produced more correct responses than familiarity (“familiar” responses). In
fact, familiarity memory in this study was roughly as reliable as guessing, as it is not
significantly higher than the chance level. Further, in support of the strength of familiarity
and recollection, familiarity was associated with significantly higher false alarm rates,
suggesting that it is weaker and more susceptible to interference.
6.2 Effects of Music on Performance
As there was no difference during piloting between stimulus sets and no difference

6 Discussion 31
during testing between our experimental conditions, it was possible to compare the
general effects of music on task performance. Piloting showed better performance in
terms of Hits, specifically due to “remembered” responses. As the only difference
between piloting and testing was the lack of music throughout the experiment, it is
possible to draw from this that music adversely affected performance. This is not in line
with many prior studies that have demonstrated the positive effects of music on task
performance, albeit with varied music styles, tasks, and performance measures
(Wallace, 1994; Thompson, Schellenberg, and Husain, 2001; Hallam, Price, and
Katsarou, 2002; Rickard, Toukhsati, and field 2005; Särkämö et al., 2008), with one
study demonstrating improved performance on a non-spatial verbal memory task
(Ferreri, Aucouturier, and Bugaiska, 2013). It can be said here that the background
music somehow was an impediment to the subjects. It is possible that the music acted
as a distractor, though the subjective rating of distraction of the songs was relatively low
(1-5 scale, 1 = “low”; 1.72 +/- 0.82, mean +/- SD). It has also been previously
demonstrated that subjects’ preference for music style does not affect performance in a
serial recall task (Perham and Vizard, 2011), and it is unlikely that enjoyment of the
songs had an effect on performance here; however, subjects in the present study rated
the songs as rather enjoyable (1-5 scale, 1 = “low”; 3.67 +/- 1.06, mean +/- SD). Further,
CDM generally involves background contexts – things that aren’t necessarily the focus
of attention (Smith, 2001) – and it is therefore possible that the music in the present
study was too much in the foreground to facilitate context-dependent memory effects,
although the effects of attention on CDM has not been previously investigated. Future
studies should take all of this into consideration, and for now it can only be said that
classical piano music as context cannot reactivate declarative memory traces during a
wakeful consolidation phase.
6.3 Effect of Study on Performance
To assess if the lack of an effect of condition could be due to differences between
participants, two-way mixed-factorial ANOVAs were performed with between subject
factors of Study, Notice, and Training. A significant effect of Study was found on Hits,
with the no-study group performing better than the study group. This is extremely
counter-intuitive, as the subjects who normally listen to music when they study might

6 Discussion 32
have been expected to perform better considering effects of context on declarative
memory performance (Smith, 1985). There is a severe lack of published studies on the
effects on memory performance of listening to music while studying, but research on
background music and task performance suggests that mathematics, reading
comprehension, and logical reasoning are not severely hindered by background music
(Hall, 1952; Savan, 1999; Hallam and Price, 1997; Hallam, Price, and Katsarou, 2002).
The present finding is, therefore, not in line with the literature. More definitive studies on
the effects of background music specifically on memory performance need to be carried
out before conclusions can be drawn on this effect.
6.4 EEG Power Analysis
Oscillatory activity in the brain during music listening at rest is not strongly represented
in the literature. Most studies into the topic investigate the processing of specific
features of music, but not on holistic music perception. Of two studies into the topic,
unfortunately with slightly incongruent methodologies, the results are not concordant.
However, the results presented here shed some light on the discrepancies between
these studies. First of all, there was a condition-dependent effect on alpha power,
showing that it was lowest in the No Song condition, further supporting the notion that
music listening is associated with increased alpha activity. This is concordant with the
results from Kay et al. (2012), which also showed increased alpha power during music
listening. In this study, activity from only the parietal and occipital electrodes was
analyzed, which supports prior research into alpha activity showing that it is mostly
localized to the occipital cortex (Adrian and Matthews, 1934; Ray and Cole, 1985;
Klimesch, 1996; Pfurtscheller, 2003; Klimesch et al., 2006; Niedermeyer, 1999). While
occipital activity is suggested to represent cortical inhibition, Kay et al. suggest that it
represents auditory information transfer. They found alpha power during music listening
to be associated with increases in regional cerebral blood flow (rCBF) in the
periaqueductal grey (PAG) which likely represents increases in PAG connectivity with
the thalamus. The authors suggested that this is due to enhanced information exchange
between the inferior colliculus, which is adjacent to PAG and auditory cortex, and not
due to cortical inhibition. Due to the present study only using EEG, it cannot clarify the

6 Discussion 33
functional reasons for increased alpha power. In addition to alpha power, Kay et al.
(2012) found EEG power in the beta range to be reduced in the music condition, while a
similar study by Nakamura et al. (1999) found it to be increased during music listening.
Unfortunately, the present study cannot clarify this discrepancy as no effect of condition
on beta power was found. Beta power in the three experimental conditions showed
relatively high variability, possibly due to headphone interference or muscle activity.
Headphones have been shown to distort auditory brainstem responses at frequencies
greater than 20 Hz (De Silva and Schier, 2008). Additionally, EEG signals are prone to
muscle movement artifacts, especially in frequencies above 20 Hz (Brunner et al., 1996;
Nunez and Srinivasan, 2006). It is possible that without this variability in beta power an
effect of condition would be seen. Though there was no effect of condition on beta
power, the present study hints at a compromise between Kay et al. (2012) and
Nakamura et al. (1999). Nakamura et al. averaged power over the “posterior 2/3rds” of
electrodes, which included occipital, parietal, and temporal electrodes, whereas Kay et
al. didn’t include temporal electrodes. It is therefore possible that the difference in
results comes from the difference in electrodes analyzed. The present study
demonstrates that the alpha power was indeed localized to the occipital electrodes, as it
was significantly higher there than in temporal electrodes. Further, beta power was
shown to be higher on the temporal and temporoparietal electrodes than the occipital
electrodes, further supporting the notion that beta power may be localized to the
temporal cortex, corresponding to primary auditory cortex. If beta power is indeed
increased during music listening, it is likely due to increased beta rhythm activity in the
auditory cortex areas. Beta rhythms have been associated with a wide variety of
cognitive processes, but the present results cannot clearly speak to the functionality of
the beta rhythms seen, as there was no difference between experimental conditions.
However, as beta rhythms were present in all conditions during rest, the present results
support the role of beta rhythms in normal waking consciousness as first described by
Berger (1938).
6.5 Conclusions and Future Studies
Taken together, the present study confirms that CDM reactivation during wakefulness is
not effective, as no effect of repeating context during consolidation was seen study. This

6 Discussion 34
is not surprising, as music has been established as a reliable context for memories but
both TMR and CDM reactivation studies commonly show a lack of memory
strengthening when reactivated in the waking state. In addition to the lack of
reactivation, music presentation led to lowered performance as measured by hit rates,
suggesting an interfering effect of music on memory encoding or consolidation, though
this finding is not in line with other studies. Further, it was found that, rather counter-
intuitively, subjects who normally listen to music while studying performed worse on the
task than those who don’t. Research into the effects of background music on declarative
memory is not very extensive, and more studies need to be done to investigate this
effect. The present results clarify previous discrepancies between studies on oscillatory
activity while listening to music at rest, in that it is shown here that alpha range (8-12
Hz) activity stronger in occipital than temporal electrodes and that beta range activity
(15-30 Hz) was stronger in temporal electrodes. Further, alpha power was lowest in the
No Song condition, supporting prior results by Kay et al. (2012). Though beta power has
been previously shown to be increased during music listening (Nakamura et al. 1999),
the present study found no effect of condition on beta power. These results are also not
concordant with Kay et al. (2012), who found decreased beta power during music
listening.
The results presented here should be further investigated. Specifically, although no
difference between conditions was found, it would be interesting to investigate the
numbers of SWP-Rs between the conditions, which was unfortunately not performed
due to time constraints. This would allow for an additional quantification of memory
consolidation besides measures of memory performance. Additionally, in order to truly
investigate the usefulness of music context in CDM reactivation, the present study
should be repeated with reactivation occurring during SWS. Moreover, as waking state
reactivation likely doesn’t work due to memory traces being very susceptible to
interference, it would be useful to increase the length of the consolidation phase or to
ensure the strength of memories through repetitions during encoding or by requiring
subjects to reach a learning criterion. Finally, to further investigate oscillatory activity
during music listening at rest, studies should be carried out investigating multiple types
of music. This would allow for generalization of the effects of music on the brain, and

6 Discussion 35
possibly provide better insight into how background music can affect learning and
memory processes.

7 Summary 36
7 Summary Within the framework of a M.Sc. Neurosciences Master’s thesis, the present study
assessed context-dependent memory (CDM) reactivation in the waking state and
oscillatory activity in the brain during music listening at rest. CDM reactivation is a
phenomenon with which memory consolidation can be experimentally manipulated by
repeating the context in which memories were initially created. Music has been shown
to be sufficient for the effects of context-dependent memory, but experimental
reactivation using music as a context has not yet been shown. Further, many students
use background music as a study aid, and it is therefore relevant to research in
memory, cognition, and education. The present study investigated oscillatory activity in
the brain during music listening at rest, specifically in the alpha (8-12 Hz) and beta (15-
30 Hz) ranges. After performing a non-spatial face memory task with background music
(Chopin’s Nocturnes), memory traces were manipulated through repeating context in a
waking-state consolidation phase. Testing immediately following the consolidation
phase demonstrated no effect of repeating context on performance, suggesting that
CDM reactivation does not work during wakefulness. This is in line with many other
studies, but should further be investigated during slow-wave sleep to determine the
usefulness of music as context on CDM reactivation as a whole. Further, the condition
in which there was no music during consolidatiion exhibited lower power in the alpha (8-
12 Hz) range compared to the music conditions. This is in line with previous research
and supports the idea of alpha activity representing cortical inhibition. Moreover, alpha
activity was localized to the occipital electrodes, suggesting localization to the visual
cortex. No effect of condition was found on beta power, but it was found to be higher
over temporal and temporoparietal electrodes, suggesting localization to temporal
cortical areas, including auditory cortex. The results presented here clarify some
discrepancies in previously published literature, but the question remains of the
effectiveness of CDM reactivation in wakefulness and of the nature of oscillatory neural
activity during music listening and its functional effects on memory task performance.

8 References 37
8 References Adrian ED, Matthews, BH (1934) The interpretation of potential waves in the cortex. J.
Physiol. 81:440-471. Alvares LO, Einarsson EO, Santana F, Crestani AP, Haubrich J, Cassini LF, Nader K,
Quillfeldt JA (2012) Periodically Reactivated Context Memory Retains Its Precision and Dependence on the Hippocampus. Hippocampus 22(5):1092–1095.
Andrade PE, Bhattacharya J (2003) Brain tuned to music. J. R. Soc. Med. 96(6):284–
287. Atkinson RC, Juola JF (1973). Factors influencing speed and accuracy of word
recognition. Atten. Perform. IV:538-612. Atkinson RC, Juola JF (1974) Search and decision processes in recognition memory. In
Contemporary developments in mathematical psychology, Krantz DH, Atkinson RC, Suppes P (eds.) San Francisco: WH Freeman, 243-290.
Axmacher N, Elger C, Fell J (2008) Ripples in the medial temporal lobe are relevant for
human memory consolidation. Brain 131:1806-1817. Balch WR, Bowman K, Mohler LA (1992) Music-dependent memory in immediate and
delayed word recall. Memory & Cognition 20(1):21-28 Bamiou DE, Musiek FE, Luxon LM (2003) The insula (island of reil) and its role in
auditory processing: Literature review. Brain Res. Rev. 42(2):143–154. Bartlett JC, Santrock JW (1979) Affect-dependent episodic memory in young children.
Child Development 50:513-518. Baumeister J, Barthel T, Geiss KR, Weiss M (2008) Influence of phosphatidylserine on
cognitive performance and cortical activity after induced stress. Nutritional Neuroscience 11(3):103-110
Berger, H. (1938) Über das Elektrenkephalogramm des Menschen. Vierzehnte
Mifteilung. Arch. Psychiat. Nervenkr. 108:407-431. Brunner DP, Vasco RC, Detka CS, Monahan JP, Reynolds CF, Kupfer DJ (1996)
Muscle artifacts in the sleep EEG: Automated detection and effect on all-night EEG power spectra. J. Sleep Res. 5:155-164.
Burns EJ, Tree JJ, Weidemann CT (2014) Recognition memory in developmental

8 References 38
prosopagnosia: electrophysiological evidence for abnormal routes to face recognition. Frontiers in Human Neuroscience 8:622.
Buzsaki G, Horvath Z, Urioste R, Hetke J, Wise K (1992) High-frequency network
oscillation in the hippocampus. Science 256:1025–1027. Cassileth BR, Vickers AJ, Magill LA (2003) Music therapy for mood disturbance during
hospitalization for autologous stem cell transplantation: A randomized controlled trial. Cancer 98:2723–2729.
Chiroro P, Valentine T (1995) An investigation of the contact hypothesis of the own-race
bias in face recognition. The Quarterly Journal of Experimental Psychology Section A: Human Experimental Psychology 48(4):879-894.
Cleary A (2011) Face recognition without identification. In Corcoran, P. (Ed.), Reviews,
Refinements and New Ideas in Face Recognition. Winchester: InTech. Available from: http://www.intechopen.com/books/reviews-refinements-and-new-ideas-in-face-recognition/face-recognition-without-identification
De Silva A, Schier M (2008) A feasibility study of commercially available audio
transducers in ABR studies. ICBME Proceedings 23:156-160. Diekelmann S, Büchel C, Born J, Rasch B (2011) Labile or stable: opposing
consequences for memory when reactivated during waking and sleep. Nature Neuroscience 14(3):381-386.
Dudai Y (2004) The neurobiology of consolidations, or, how stable is the engram? Annu
Rev Psychol 55:51-86. Ego-Stengel V, Wilson MA (2010) Disruption of ripple-associated hippocampal activity
during rest impairs spatial learning in the rat. Hippocampus 20(1):1–10. Eich JE (1980) The cue-dependent nature of state-dependent retrieval. Memory
Cognition 8:157-173. Fell J, Axmacher N, Haupt S (2010) From alpha to gamma: Electrophysiological
correlates of meditation-related states of consciousness. Medical Hypotheses 75: 218-224.
Ferreri L, Aucouturier J, Bugaiska A (2013) Music improves verbal memory encoding
while decreasing prefrontal cortex activity: An fNIRS study. Front Hum Neurosci. 7:779.
Frankland PW, Bontempi B (2005) The organization of recent and remote memories.
Nature Reviews Neuroscience 6(2):119–130.

8 References 39
Godden DR, Baddeley AD (1975) Context-dependent memory in two natural environments: On land and underwater. British Journal of Psychology 66:325-331.
Hall J (1952) The effect of background music on the reading comprehension of 278
eighth and ninth grade students. Journal of Educational Research 45:451-458. Hallam S, Price J (1997) Can listening to background music improve children's
behaviour and performance in mathematics? Paper presented at the British Educational Research Association Annual Conference, September 11-14 1997: University of York.
Hallam S, Price J, Katsarou G (2002) The effects of background music on primary
school pupils’ task performance. Educational Studies 28(2):111-122. Hobson J, Pace-Schott E (2002) The cognitive neuroscience of sleep: Neuronal
systems, consciousness and learning. Nature Reviews Neuroscience 3(9):679-693.
Holloway FA, Wansley R (1973) Multiphasic retention deficits at periodic intervals after
passive-avoidance learning. Science 180:208-210. Hugenberg K, Miller J, Claypool HM (2007) Categorization and individuation in the
cross-race recognition deficit: Toward a solution to an insidious problem. Journal of Experimental Social Psychology 43(2):334-340.
Hupbach A, Hardt O, Gomez R, Nadel L (2008) The dynamics of memory: Context-
dependent updating. Learn. Mem. 15:574-579. Jacoby LL (1991) A process dissociation framework separating automatic from
intentional uses of memory. J. Mem. Lan. 30:513-541. Jiang X, Sengupta AK (2011) Effect of music and induced mental load in word
processing task. IEEE SMC 2011:3261-3266. Kandel ER, Schwartz JH, Jessell TM (2012) Principles of neural science. New York:
McGraw-Hill, Health Professions Division. Karlsson MP, Frank LM (2009) Awake replay of remote experiences in the
hippocampus. Nature Neuroscience 12(7):913–918. Klimesch W (1996) Memory processes, brain oscillations and EEG synchronization. Int.
J. Pathophysiol. 24:61-100. Klimesch W, Sauseng P, Hanslmayr S (2006) EEG alpha oscillations: The inhibition-
timing hypothesis. Brain Res. Rev. 53:63-88.

8 References 40
Klostermann EC, Loui P, Shimamura AP, (2009) Activation of right parietal cortex during
memory retrieval of nonlinguistic auditory stimuli. Cogn. Affect. Behav. Neurosci. 9(3):242–248.
Lee JH, Whittington MA, Kopell NJ (2013) Top-Down Beta Rhythms Support Selective
Attention via Interlaminar Interaction: A Model. PLoS Comput Biol 9(8):e1003164.
Logothetis NK, Eschenko O, Murayama Y, Augath M, Steudel T, Evard HC, Besserve
M, Oeltermann A (2012) Hippocampal–cortical interaction during periods of subcortical silence. Nature 491:547–553.
Lynch M (2004) Long-term potentiation and memory. Physiol Rev 84(1):87–136. Mandler G (1980) Recognizing the judgement of previous occurrence. Psychol. Rev.
87:252-271. Maren S, Phan KL, Liberzon I (2013) The contextual brain: Implications for fear
conditioning, extinction, and psychopathology. Nature Reviews Neuroscience 14(6):417–428.
Martin RC, Wogalter MS, Forlano JG (1988) Reading comprehension in the presence of
unattended speech and music. Journal of Memory and Language 27(4):382-398. Meissner CA, Brigham JC (2001) Years of investigating the own-race bias in memory
for faces: A meta-analytic review. Psychology, Public Policy, and Law 7(1):3-35. Minear M, Park DC (2004). A lifespan database of adult facial stimuli. Behavior
Research Methods, Instruments, & Computers 36(4):630-633. Nakamura S, Sadato N, Oohashi T, Nishina E, Fuwamoto Y, Yonekura Y (1999)
Analysis of music-brain interaction with simultaneous measurement of regional cerebral bloodflow and electroencephalogram beta rhythm in human subjects. Neuroscience Letters 275:222-226.
Niedermeyer, E (1999) The normal EEG of the waking adult. In:
Electroencephalography: Basic Principles, Clinical Applications, and Related Fields. Baltimore: Lippincott Williams Wilkins:149-173.
Nunez PL, Srinivasan R (2006) Electric Fields of the Brain: The Neurophysis of EEG,
Second Edition. New York City: Oxford University Press. Oostenveld R, Fries P, Maris E, Schoffelen JM (2011) FieldTrip: Open source software
for advanced analysis of MEG, EEG, and invasive electrophysiological data. Computational Intelligence and Neuroscience 2011.

8 References 41
Oostenveld R, Praamstra P (2001) The five percent electrode system for high-resolution
EEG and ERP measurements Clinical Neurophysiology 112:713-719. Oudiette D, Antony JW, Creery JD, Paller KA (2013) The role of memory reactivation
during wakefulness and sleep in determining which memories endure. The Journal of Neuroscience 33(15):6672-6678.
Oudiette D, Paller KA (2013) Upgrading the sleeping brain with targeted memory
reactivation. Trends in Cognitive Sciences 17(3):142-149. Palva S, Palva JM (2007) New vistas for alpha-frequency band oscillations. Trends
Neurosci 30(4):150-158. Perham N, Vizard J (2011) Can preference for background music mediate the irrelevant
sound effect? Applied Cognitive Psychology 25:625-631. Pfurtscheller G (2003) Induced oscillations in the alpha band: Functional meaning.
Epilepsia 44(12):2-8. Plihal W, Born J (1999) Memory consolidation in human sleep depends on inhibition of
glucocorticoid release. Learning and Memory 10(13):2741-2747. Rasch B, Büchel C, Gais S, Born J (2007) Odor cues during slow-wave sleep prompt
declarative memory consolidation. Science 315:1426-1429. Rauscher FH, Shaw GL, Ky CN (1993) Music and spatial task
performance. Nature 365:611. Ray WJ, Cole HW (1985) EEG alpha activity reflects attentional demands, and beta
activity reflects emotional and cognitive processes. Science 228:750-752. Rickard NS, Toukhsati SR, Field SE (2005) The effect of music on cognitive
performance: Insight from neurobiological and animal studies. Behavioural and Cognitive Neuroscience Reviews 4:235.
Roozendaal B (2000) Glucocorticoids and the regulation of memory
consolidation. Psychoneuroendocrinology 25:213–238. Rudoy JD, Voss JL, Westerberg CE, Paller KA (2009) Strengthening Individual
Memories by Reactivating Them During Sleep. Science 326:1079. Särkämö T, Tervaniemi M, Laitinen S, Forsblom A, Soinila S, Mikkonen M, Autti
T, Silvennoinen HM, Erkkilä J, Laine M, Peretz I, Hietanen M (2008) Music listening enhances cognitive recovery and mood after middle cerebral artery stroke. Brain 131(3):866-76.

8 References 42
Sarnthein J, von Stein A, Rappelsberger P, Petsche H, Rauscher FH, Shaw GL (1997)
Persistent patterns of brain activity: An EEG coherence study of the positive effect of music on spatio-temporal reasoning. Neurol Res 19:107-116.
Savan A (1999) The effect of background music on learning. Psychology of Music
27:138. Siedlecki SL, Good M (2006) Effect of music on power, pain, depression and disability.
Journal of Advanced Nursing 54:553–562. Singer W (1999) Neuronal synchrony: a versatile code for the definition of relations?
Neuron 24:49–65;111–125. Skaggs WE, McNaughton BL, Permenter M, Archibeque M, Vogt J, Amaral DG, Barnes
CA (2007) EEG sharp waves and sparse ensemble unit activity in the macaque hippocampus. Journal of Neurophysiology 98(2):898–910.
Smith SM (1979) Remembering in and out of context. Journal of Experimental
Psychology. Human Learning and Memory 5:460-471. Smith SM (1985) Background music and context-dependent memory. American Journal
of Psychology 98(4):591-603. Smith SM, Vela E (2001) Environmental context-dependent memory: A review and
meta-analysis. Psychon Bull Rev. 8(2):203-220. Staresina BP, Fell J, Do Lam ATA, Axmacher N, Henson RN (2012) Memory signals
are temporally dissociated in and across human hippocampus and perirhinal cortex. Nature Neuroscience 15(8):1167-1173.
Stickgold R (2005) Sleep-dependent memory consolidation. Nature 437:1272-1278. Thompson WF, Schellenberg EG, Husain G (2001) Arousal, mood, and the Mozart
effect. Psychological Science 12:248–251. Tronson NC, Taylor JR (2007) Molecular mechanisms of memory
reconsolidation. Nature Reviews Neuroscience 8(4)262–275. Wallace WT (1994) Memory for music: Effect of melody on recall of text. Journal of
Experimental Psychology: Learning, Memory, and Cognition 20:1471–1485. Watanabe T, Yagishita S, Kikyo H (2008) Memory of music: Roles of right hippocampus
and left inferior frontal gyrus. Neuroimage 39(1):483–491.

8 References 43
Yonelinas AP (1994) Receiver operating characteristics in recognition memory: Evidence for a dual process model. J. Exp. Psychol. Learn. Mem. Cogn. 20:1341-1354.
Zhang Y, Chen Y, Bressler SL, Ding M (2008) Response preparation and inhibition: the
role of the cortical sensorimotor beta rhythm. Neuroscience 156(1):238–246.

9 Acknowledgements 44
9 Acknowledgements First and foremost I would like to thank PD Dr. Juergen Fell for allowing me to do my
thesis in his group and for allowing me a bit of creative freedom and independence on
the project, as well as for the invaluable assistance and feedback. Additionally, thanks
go out to Prof. Dr. Nikolai Axmacher the members of the working groups of AG
Axmacher and AG Fell for immediately welcoming me as a member of the group and
providing me with much help with programming and ideas. Specifically, extra special
thanks go to Marcin Lezcynski – without his help, none of the EEG analysis in this
thesis would’ve been possible. I would also like to thank Jacob Nelson for his
assistance in setting up the EEG equipment (and making it to really early appointments
all the way out in Buschdorf) and additionally for help with technical issues and
entertaining subjects during EEG setup. Further I would like to thank Dr. Ines Heuer for
her support, especially when trying to cope with the stress that a thesis brings upon you.
Also to Yilmaz Sagik – ohne deine Hilfe mit den Probanden, technische Probleme, und
der Finanzierung wäre meine Studie gar nicht möglich. A huge thank you to my parents,
Mark and Lynn Chockley, not only for providing financial support to come to Germany to
work towards this degree, but also for supporting me in every aspect of life and always
pushing me to do what I love and to never give up. Aside from the select few I have
mentioned here, many others deserve thanks from me, including all of my friends and
colleagues here in Bonn and around the world. Without the support of my community
none of this would have been possible. I am very grateful to everyone who has
supported me not only during this thesis, but also during my entire time here in Bonn.

10 Appendices 45
10 Appendices
A. Study Advertisement for Participant Recruitment
CallforParticipants:Theinfluenceofmusiconmemory
Description:Theworkinggroup“CorticalOscillations–AGAxmacher/AGFell”islookingforparticipantsforanEEGstudy.ThestudyinvestigatestheeffectsofmusiconmemoryandEEG(electroencephalogram)signals.ThisstudyispartofaMaster’sThesisintheNeurosciencesM.Sc.program.Task:Participantswillbeaskedtomemorizefacespresentedtothemonacomputerscreenwhilelisteningtomusic.EEGdatawillberecordedthroughoutthesession.Duration:2.5-3hours–1hourforset-up,1.5hoursforthetaskAllparticipantswillbereimbursed10Euros/hour(25-30Eurostotal)Location:CenterforClinicalResearch GermanCenterforNeurodegenerativeDiseases(DZNE)Ernst-Robert-Curtius-Straße1253117Bonn–BuschdorfWhen:Appointmentswillgenerallybeat9a.m.and1p.m.(timesaresomewhatflexible)onweekdaysstartingassoonaspossible.ThisstudyisapprovedbytheethicscommissionoftheMedicalFacultyoftheUniversityofBonnandposesnoriskstotheparticipants’healthorwell-being.Participationinthestudyinentirelyvoluntary,meaningtheparticipantscanwithdrawatanytimewithoutconsequence.Allpersonaldatawillbekeptanonymous.Yourparticipationwillcontributetotheadvancementofresearchinmemoryconsolidation,theneuroscienceofmusic,andeducation.Therefore,wewouldreallyappreciateyourparticipation!
Ifyouareinterestedinparticipating,[email protected]+49(0)177-781-2636
Prof. Dr. C. E. Elger, FRCP Direktor Alexander Chockley
Fon: 0177. 781-2636 [email protected] www.epileptologie-bonn.de

10 Appendices 46
B. Post-experiment Questionnaire

10 Appendices 47
Subject(Nr:(__(Sex:(__(Age:(__(
2((
Was(there(a(strategy(that(you(used(to(memorize(the(faces?__________________________________(
__________________________________________________________________________________
_______________________________________________________________________________(
!
Overall!
Do(you(have(any(further(comments(about(the(experiment?(_________________________________(
__________________________________________________________________________________
__________________________________________________________________________________
__________________________________________________________________________________
__________________________________________________________________________________(
(
Thank(you(very(much(for(your(participation!(It(is(very(much(appreciated.((!(

11 Affirmation for the Master’s Thesis 48
11 Affirmation for the Master’s Thesis I herewith declare that I have independently written this thesis. Only the sources cited
have been used in this work. Parts that are direct quotes or paraphrases are identified
as such.
Bonn , _________________________ _____________________________
(date) (signature)
Related Documents











