Louisiana State University LSU Digital Commons LSU Historical Dissertations and eses Graduate School 1997 DNA Wrapping and the Nature of Interaction Between E. Coli RNA Polymerase and Promoter DNA. Parisa Jazbi Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: hps://digitalcommons.lsu.edu/gradschool_disstheses is Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and eses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. Recommended Citation Jazbi, Parisa, "DNA Wrapping and the Nature of Interaction Between E. Coli RNA Polymerase and Promoter DNA." (1997). LSU Historical Dissertations and eses. 6571. hps://digitalcommons.lsu.edu/gradschool_disstheses/6571

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Louisiana State UniversityLSU Digital Commons
LSU Historical Dissertations and Theses Graduate School
1997
DNA Wrapping and the Nature of InteractionBetween E. Coli RNA Polymerase and PromoterDNA.Parisa JazbiLouisiana State University and Agricultural & Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion inLSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please [email protected].
Recommended CitationJazbi, Parisa, "DNA Wrapping and the Nature of Interaction Between E. Coli RNA Polymerase and Promoter DNA." (1997). LSUHistorical Dissertations and Theses. 6571.https://digitalcommons.lsu.edu/gradschool_disstheses/6571

INFORMATION TO USERS
This manuscript has been reproduced from the microfilm master. UMI
films the text directly from the original or copy submitted. Thus, some
thesis and dissertation copies are in typewriter face, while others may be
from any type o f computer printer.
The quality of this reproduction is dependent upon the quality of the
copy submitted. Broken or indistinct print, colored or poor quality
illustrations and photographs, print bleedthrough, substandard margins,
and improper alignment can adversely afreet reproduction.
In the unlikely event that the author did not send UMI a complete
manuscript and there are missing pages, these will be noted. Also, if
unauthorized copyright material had to be removed, a note will indicate
the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by
sectioning the original, beginning at the upper left-hand comer and
continuing from left to right in equal sections with small overlaps. Each
original is also photographed in one exposure and is included in reduced
form at the back of the book.
Photographs included in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6” x 9” black and white
photographic prints are available for any photographs or illustrations
appearing in this copy for an additional charge. Contact UMI directly to
order.
UMIA Bell & Howell Information Company
300 North Zeeb Road, Ann Arbor MI 48106-1346 USA 313/761-4700 800/521-0600
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

DNA WRAPPING AND THE NATURE OF INTERACTION BETWEEN K coli RNA POLYMERASE AND PROMOTOR DNA
A Dissertation
Submitted to the Graduate Faculty of the Louisiana State University and
Agricultural and Mechanical College in partial fulfilment of the
requirements for the degree of Doctor of Philosophy
in
The Department of Microbiology
byParisa Jazbi
B.S., Shiraz University, 1991 December 1997
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

UMI Number: 9820725
UMI Microform 9820725 Copyright 1998, by UMI Company. All rights reserved.
This microform edition is protected against unauthorized copying under Title 17, United States Code.
UMI300 North Zeeb Road Ann Arbor, MI 48103
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

In the name of God most merciful most gracious
DEDICATION
To my parents for their love and faith in me
ii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

ACKNOWLEDGMENTS
I am greatly indebted to my major adviser who has been a significant mentor. His
constant support, guidance, and encouregment made this work possible.
I would also like to thank the members of my committee, Dr. Randall Gayda, Dr.
Ronald Siebeling, Dr. Gregory Jarosik, Dr. Gregg Pettis, Dr. Ding Shih, and Dr. David
Senior for their invaluable and sensible advice.
My thanks also to my fellow graduate students in the microbiology department for
their friendship and encouragement.
My brother, Ali deserves a special thank you for being my inspiration and incentive
to finish this work.
Most of all, I am grateful to my husband, without whose love and belief in me I
never could have completed this degree. He believed in me when I had a hard time believing
in myself.
iii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

TABLE OF CONTENTS
DEDICATION................................................................................................................ ii
ACKNOWLEDGMENTS ............................................................................................iii
LIST OF TABLE............................................................................................................vi
LIST OF FIGURES......................................................................................................vii
ABSTRACT.................................................................................................................... ix
INTRODUCTION.......................................................................................................... 1DNA-dependent RNA polymerase....................................................................... 1Promoters............................................................................................................. 5Transcription initiation kinetics............................................................................ 8Regulators of transcription................................................................................. 10DNA curvature................................................................................................... 13DNA curvature in prokaryote promoters............................................................15
MATERIALS AND M ETHODS................................................................................ 18Materials..............................................................................................................18Plasmid transformation and isolation.................................................................. 18Construction of Alul56 derivative promoters.....................................................18Primer labeling .............................................................................................21PCR amplification...............................................................................................21RNA polymerase isolation..................................................................................22DNase I footprinting analysis............................................................................. 22Gel retardation analysis...................................................................................... 23KMn04 footprinting...........................................................................................24Single-round run-off transcription assay............................................................ 24
RESULTS..................................................................................................................... 26DNase I footprint analysis of the interaction between E. coliRNA polymerase and Alul56 and Ball29 and their derivatives..........................26Nature of the interaction between RNA polymerase and DNA upstreamof the -35 region................................................................................................. 41The relationship between open promoter complex formation andDNA wrapping................................................................................................... 54Test for DNA wrapping at different stages of transcription initiation................ 54Effect of AT-rich regions in DNA wrapping and transcription efficiency.......... 63
iv
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Effect of a mutation in the C-terminal domain of a subunit on the interaction between RNA polymerase and DNA upstream of the -35 region.......................74
DISCUSSION................................................................................................................ 79Effect of DNA curvature immediately upstream of the -35 region on DNAwrapping by the RNA polymerase...................................................................... 79Model for the role of DNA curvature in RNA polymerase binding andtranscription initiation........................................................................................80The nature of the interaction between E. coli RNA polymerase and DNAupstream of the -35 region.................................................................................. 82Existence of unwrapped open promoter complexes............................................83DNA wrapping in the absence of open promoter complex formation.................84Comparison of the effect of AT-rich regions and curved DNA on wrappingand transcription initiation...................................................................................85Contribution of the a subunit of RNA polymerase on wrapping........................ 86Summary of the model........................................................................................ 86
REFERENCES .............................................................................................................87
VITA............................................................................................................................. 98
v
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

LIST OF TABLES
1. Promoters................................................................................................................. 21
vi
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

LIST OF FIGURES
1. Nucleotide sequence of the Alul56 and Ball 29 promoters..................................... 27
2. Nucleotide sequence of the Alul56, Ball29, and their derivatives..........................28
3. DNase I footprint analysis of the Alul56 promoter..................................................30
4. DNase I footprinting of the AluExt promoter bound by RNA polymerase..............33
5. The summary of DNase I footprint analysis on both strands o f theAlul56 (A) and the AluExt (B) promoters...............................................................
6. DNase I footprint analysis of Ball29 promoter........................................................ 37
7. DNase I footprint analysis of BalExt promoter........................................................ 39
8. The summary of DNase I footprinting results on both strands ofBall29 (A) and BalExt (B )........................................................................................42
9. DNase I footprint analysis of the Alul56 promoter bound to RNA polymeraseas a function of NaCl concentration........................................................................... 45
10. Effect of NaCl concentration on DNase I footprint pattern of AluExt.....................48
11. DNase I footprint analysis of the Ball29 promoter as a function ofNaCl concentration....................................................................................................50
12. Effect of NaCl concentration on the DNase I footprint analysis of theBalExt promoter...................................................................................................... 52
13. Open promoter complex formation at different salt concentrationsfor Ball29 (A) and Alul56 (B )................................................................................. 55
14. Effect of temperature on open complex formation of Alul56 promoter.................. 57
15. DNase I footprint analysis of Alul56 as a function o f temperature..........................58
16. Effect of heparin on the footprint analysis of the Alul56 promoter boundto RNA polymerase at 37°C and 0 °C .................................................................... 61
17. Nucleotide sequence of the Alul56, AluExt, AluDel, and AT-rich derivatives 64
vii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

18. DNase I footprint analysis of the Alul56, AluUnc, AluUp, andAlu27AT promoters................................................................................................. 65
19. DNase I footprint analysis of the Alul56, AluExt, Alu8AT, andATDel promoters..................................................................................................... 67
20. Gel retardation analysis of Alul56, AluUnc, AluUp, and Alu27AT.........................70
21. Gel retardation analysis of Alul56, AluExt, Alu8AT, and AluDel...........................71
22. Transcribable promoter complexes formed as a function of temperature................. .72
23. DNase I footprint analysis of the AIuPr, BalP^ and APR promoters as afunction of wild-type (lane 2) and mutant (lane 3) RNA polymerase.......................76
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

ABSTRACT
Regulation of transcription most often occurs at the stage of initiation. RNA
polymerase binding to the transcription start site, the promoter, is influenced by many
nucleotide sequence elements. The predominant recognition sequences are those bound by
the a subunit of RNA polymerase located at -10 and -35 relative to initiation site of most
promoters. Another element involved in this regulation is intrinsic DNA curvature. This
study examined the contribution of intrinsically curved DNA immediately upstream of the
promoter to the interaction between Escherichia coli RNA polymerase and this DNA
DNase I footprinting analysis confirmed that RNA polymerase wraps DNA upstream of the
promoter around the enzyme. The nature of interaction between DNA upstream of
promoter and RNA polymerase was explored using addition of NaCl. The wrapped
complex was not observed at NaCl concentration above 150 mM suggesting the
electrostatic, sequence-independent nature of the interaction. Study of the effect of
temperature on DNA wrapping and open promoter complex formation demonstrated the
existence of closed, wrapped complexes. No wrapped complexes survived a 30 second
heparin challenge indicating the absence of wrapped open complexes. The above data
provided evidence that DNA wrapping occurs prior to open complex formation. Promoters
containing an AT-rich region or the UP element of ribosomal RNA promoter rm BPl were
constructed. Using a gel retardation assay, the relative affinity of RNA polymerase for these
promoters was compared to that observed for curved DNA-containing promoter. The
promoter containing curved DNA displayed the highest binding to RNA polymerase. The
ix
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

presence of curved DNA favored the formation of the wrapped complex. A run-off
transcription assay limited to a single round of initiation examined the efficiency of
transcription for these promoters as a function of temperature. Relative to promoters
lacking curved DNA, the promoter with curved DNA formed significantly more heparin-
resistant, closed complexes at low temperature. These complexes could quickly isomerize
to open complex at 37°C. We propose that curved DNA facilitates wrapping of DNA
around RNA polymerase and enhances the transition from a heparin sensitive closed
complex to a heparin resistance closed promoter complex.
x
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

INTRODUCTION
The regulation of gene expression is essential to the efficiency and economy of
bacterial growth. Control of the activity of bacterial genes usually starts at the stage of
transcription. The controlled step in this process is commonly the initiation of transcription.
The least amount of energy and other resources are used by controlling the very first step
of transcription.
Several elements are involved in the multiple step process o f initiation of
transcription. These components are common among several species of bacteria, including
Escherichia coli and Bacillus subtilis. DNA dependent RNA polymerase binds to the
promoter, a specific sequence of DNA, with or without the regulatory proteins that repress
or activate transcription. Most of the interactions between the RNA polymerase and the
promoter DNA are mediated through consensus DNA sequence elements, such as those
commonly found at -10 and -35 relative to the site of transcription initiation. These
interactions are involved in the separation of the DNA strands. Other DNA elements, such
as intrinsically curved DNA, have been shown to affect the initiation of transcription.
Involvement of these elements in regulation of transcription initiation can be further studied
using in vitro systems.
DNA-dependent RNA polymerase. RNA polymerase is the cell's central processing unit.
This enzyme was discovered in 1959 by Weiss and Gladstone. RNA polymerase copies the
information from a DNA template to RNA molecule specifically. It synthesizes an RNA
molecule that is complementary to the DNA template. The RNA polymerase of E. coli is
composed of a core enzyme with the subunit structure of (Chamerlin, 1982; McClure,
1
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

2
198S) and one of the several species of a subunit which are involved in the specific
promoter recognition (Helmann and Chamberlin, 1988). The catalytic site of RNA
polymerase is located on the 3 (150,619 Daltons) subunit (Glass et aL, 1982), while RNA
polymerase binds to the DNA nonspecifically via the 3' (155,162 Daltons) subunit (Fukuda
and Ishihama, 1979).
Alpha (a) subunits are involved in assembly of the RNA polymerase (Ishihama,
1981), and also in protein protein interactions with positive regulators (Ishihama, 1992).
E. coli RNA polymerase with deletion in the C-terminal one third of the a subunit is
enzymatically active, however, some activator dependent promoters cannot be transcribed
by this mutant RNA polymerase. This suggests that the C-terminal region of a interacts
with some transcription factors (Igarashi and Ishihama, 1991). The cyclic AMP receptor
protein (CRP) contacts the a subunit of RNA polymerase when activating transcription at
“class I promoters” in which the CRP binding site is located upstream of the -35 region.
The targets for CRP interaction also reside in the C-terminal domain (CTD) of the a subunit
(Chen et aL 1994; Zhou, et aL, 1994; Zou et aL, 1992). In addition, for a number of other
bacterial activators, such as AraC, OxyR, PhoB, FNR, and integration host factor (IHF), it
has been shown that they mediate their effect via the a CTD (Ishihama, 1992). The function
of the C-terminal domain of the a subunit of E. coli RNA polymerase in basal expression
and integration host factor-mediated activation of the early promoter of bacteriophage MU
has been recently studied by Vanulsen et aL(1997). The results of this study indicates that
interaction of the a CTD with DNA is involved not only in the IHF mediated activation of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

3
early promoter but also in maintaining the basal level of transcription from this promoter.
It was shown in rm B P 1 promoter that sequences between -40 and -60, the so called “UP
element”, increases transcription by interacting with the a subunit of RNA polymerase (Ross
et aL, 1993; Gaal et aL, 1996). RNA polymerase lacking the C-terminal domain of the a
subunit was unable to contact the UP element, and therefore, transcription from rm B PI
promoter was less efficiently initiated (Ross et aL, 1993). In the same study, it was shown
that purified a binds specifically to the UP element. The binding was diminished when
mutated a was used. Therefore, it can be concluded that the UP element represents a third
promoter recognition region and that a acts directly in promoter binding.
Six different a subunits have been found in association with E. coli polymerase. All
these sigma factors play an important role in the specificity of the transcription initiation.
Alternative a subunits control the transcription of coordinately regulated sets of genes
distinct from those recognized by the primary o factor (Doi and Wang, 1986; Losick et aL
1986). In exponentially growing cells, most genes are transcribed by a 70 (70,263 Daltons).
Genes for heat shock proteins are transcribed by a32 (Grossman et aL, 1984; Cowing et aL,
1985; Fujita et aL, 1987). A second heat shock sigma factor, a24, has been identified as a
requirement for survival of high temperatures. The a54 is required for transcription of the
genes which are controlled by the availability of nitrogen source (Garcia et aL, 1977). The
promoters of the genes that are expressed only in stationary growth phase can be recognized
by a 3* (Tanaka et aL, 1993; Heggearonis, 1993). Genes for flagellar synthesis and
chemotaxis are controlled by a28 (Amosti et aL, 1989). Recently, evidence for contact
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

between CRP and a™ subunit of E coli RNA polymerase has been published for class II
promoters (Ruzhonsl et aL, 1995). The data indicates the role of the a 70 subunit in
transcription activation.
Unlike E. coli, B. subtilis undergoes a unique developmental process, called
sporulation. This process is directed by a cascade of sigmas which lead to the temporal
activation of different sets of genes during sporulation (Losick and Pero, 1981; Doi, 1982).
B. subtilis RNA polymerase is very similar in core subunit composition to that of-El coli.
However, B. subtilis also contain an additional polypeptide, the 8 subunit (20,400 Dalton)
(Doi, 1982; Lampe et aL, 1988) and two © subunits. The 8 subunit is responsible in
promoter discrimination. It appears that the 5 subunit allows RNA polymerase to
differentiate between strong and weak promoters (Achberger and Whiteley, 1982; Whiteley
et aL, 1982). The a subunit may be required for rapid recycling of the core RNA polymerase
after termination (Juang and Helmann, 1994). The majority of cellular transcription is
controlled by a*, the major vegetative sigma of B. subtilis (Losick et aL, 1986). The o° is
involved in flagellar synthesis, chemotaxis, and nutrient stress responses (Helmann and
Chamberlin, 1987; Helmann et aL, 1988). In addition to a * , or B, a c(Wiggs et aL, 1981)
and oD (Haldemwang and Losick, 1980) are associated with vegetatively growing cells.
Recently, three minor sigmas, a* , a Y, and o w, involved in the control of gene expression
in response to environmental stresses were discovered (Huang et al., 1997). There are other
sigmas associated with sporulating B. subtilis, including a", a F, o E, a °, and a K factors
(Losick and Stragier, 1992). During sporulation the cell undergoes an asymmetric septation
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

5
that gives rise to a small cell destined to become the endospore and a larger, terminally
differentiated mother cell. The a ” is a minor sigma factor involved in the transcription of
both vegetative and sporulating genes. In this process, the inactive form of c f is transcribed
by o” in the mother cell before the asymmetric cell division. Activation of o* after septation
directs the expression of oG. The (9 is produced in forespore and involved in
transcription of forespore specific genes. The active sigma in mother cell, a 2, is required
for prespore formation. Endospore coat proteins are the product of cot genes. The 0 s is
necessary for expression of these genes in the mother cell.
Alternative sigma factors also have been found in phage SP82 or SP01 infected
B. subtilis cells. During the infection process, phage gene expression is controlled by a
cascade of sigmas. The host a* RNA polymerase is required for expression of early phage
genes including gp28, which encodes a®28. The a®2* displaces a* on the RNA polymerase
shutting down host and early phage gene expression. The middle phage gene promoters are
transcribed by o®28 RNA polymerase. Among the middle genes are the gp33 and gp34, the
gene products of which form a sigma, o®3334, these new sigmas displace <3®28 from the RNA
polymerase shutting down middle phage gene expression and permitting late phage gene
expression. Thus, the temporal gene expression during phage development is regulated by
alternate o factors (Duffy et aL, 1975; Talkington and Pero, 1978; Lee and Pero, 1981;
Geidushik and Ito, 1982).
Promoters. Promoters are specific sequences of DNA located upstream of transcription
start sites. RNA polymerase recognizes and binds to the promoter and initiates
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

6
transcription. The level of expression o f a gene is greatly affected by the sequence of the
promoter (Galas et aL 1985). The sequence comparison of many promoters has generated
consensus sequence for particular RNA polymerase. The consensus sequence for K coli
ct70 holoenzyme is TATAAT (the -10 region) located about 10 base pairs upstream of the
transcription start she and TTGACA (the -35 region) located 17 base pairs upstream of the
-10 region (Rosenberg and Court, 1979; Siebenlist et aL 1980; Hawly and McClure, 1983;
Harley and Reynolds, 1987). The role of -10 and -35 regions in the initiation of
transcription has been studied. The -10 region is involved in DNA melting, while the -35
region plays a role in initial binding of RNA polymerase. In addition to binding affinity, the
rate of open promoter complex formation is also affected by base substitution in -35 region
(Hawley and McClure, 1982; Koboyashi et aL, 1990). Genetic studies have shown that two
regions of most a factors are involved in interaction with the -10 and -35 regions of
promoters (Waldburger et aL, 1990; Siegle, et aL, 1989; Kenney and Moran, 1991). In feet,
amino acid substitution in the conserved sequences of a which interact with the -35 region
can alter promoter specificity. Thus, holoenzyme containing such a mutant a recognizes
promoters which normally are not transcribed by wild-type holoenzyme (Schmidt et al.,
1990). It has been also demonstrated that holoenzyme containing alternate o subunits
recognizes unique promoters with different consensus sequences (Doi and Wang, 1986).
Several elements are responsible for promoter strength or the relative
transcriptional effectiveness. Promoter strength is profoundly affected by mutation in the
-10 and -35 region. Mutations causing divergence from the consensus sequence reduce the
promoter strength, while mutations increasing the level of homology exhibit enhanced
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

7
promoter activity (McClure, 1985; Hawley and McClure, 1983; Harley and Reynold, 1987).
The actual sequence of spacer DNA does not play a major role in promoter strength,
however the distance between two consensus regions is critical for productive interaction
between RNA polymerase and promoter.
Investigators have identified an additional conserved promoter sequence immediately
upstream of the -10 region (Moran et aL, 1982). A sequence around the +1 promoter
region has been also identified to affect transcription in B. subtilis but not in E. coli (Henkin
and Sonenshein, 1987).
Recently it has been established that there is a third important sequence element, in
addition to the -35 and -10 elements, at some E. coli promoters (Ross et aL, 1993). The
exceptional strength of these promoters (ribosomal RNA genes) is due to the UP element,
an AT-rich sequence of 20 base pairs located immediately upstream of the -35 region or the
UP element. Fredrick et al. (1995) also demonstrated that transcription from the B. subtilis
felagellin promoter is stimulated by an UP element both in vivo and in vitro. It is likely that
promoter strength is a function of all promoter elements, with very strong promoters having
near consensus elements while weaker promoters deviate significantly from the consensus.
Although the major forms of the RNA polymerase from E. coli and B. subtilis
recognize the same consensus sequences (Moran et aL,1982; Galas et aL,1985; Graves and
Rabinowitz, 1986), some differences have been reported for B. subtilis RNA polymerase.
B. subtilis RNA polymerase generally requires DNA sequences that are more similar to the
consensus sequence. In addition, inefficient utilization of E. coli promoters has been
observed when they are introduced to B. subtilis (Wigg etal.,\919\ Lee et al., 1980; Henkin
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

8
and Sonenshein,1987; Whipple and Sonenshein,1992). In contrast, B. subtilis promoter can
be utilized efficiently in £ coli. Investigators have identified an AT-rich sequence upstream
of the -35 region in many B. subtilis promoters, including promoters for early gene
expression in phage SPO1, which may be responsible for functional differences between K
coli and B. subtilis (Moran et aL,1982). To be fully functional, many promoters need
additional sequences where regulatory proteins bind. These proteins can act as repressors
or activators and regulate promoter function (Pabo and Sauer, 1984).
Transcription initiation kinetics. E. coli is the traditional system for the study of
transcription initiation kinetics. Transcription initiation by RNA polymerase is a
complicated process comprised of a series of defined biochemical intermediates
(Chamberlin, 1974; VonHipple et aL,1984; McClure, 1985). Two intermediates were
involved in the original model. In this model, after binding of RNA polymerase (R) to the
promoter (P), first a closed promoter complex is formed (RPc). The close complex then is
isomerized to open promoter complex (RP0) in which the DNA strands of the promoter
region are melted (Sienbenlist, 1979; Kirkegaad et aL, 1983), exposing the template strand
to RNA polymerase for RNA synthesis.
R + P '*— *“ RPC T— *“ RP0 ^ R N A synthesis
The existence of at least two intermediate complexes before open promoter complex
was documented (Rosenberg et aL,1982; Roe et aL,1984; Roe, 1985; Buc and McClure,
1985; Duval-valentin and Ehrlich, 1987). In most recent kinetic scheme RNA polymerase
first binds to promoter and forms a closed complex. A conformational change, possibly
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

9
leading to partial unwinding of the helix, forms a second closed complex, PR^ . Although
closed promoter complexes are normally sensitive to heparin, this complex is heparin
resistant. Heparin is a polyanion with high affinity for RNA polymerase (Walter et
aL,1967). Isomerization later on leads to open promoter complex formation in a
temperature-dependent process. Increasing the temperature activates the isomerization and
drives the open promoter complex formation. In & coli abortive RNA synthesis has been
identified as an intermediate step between open promoter complex formation and productive
RNA synthesis (Carpouis and Gralla, 1980). RNA polymerase of£. coli can go through
multiple cycles of abortive initiation. After each cycle, a short RNA oligomer is released
from the transcription complex. Usually after synthesis o f 9 to 11 bases of RNA, promoter
clearance occurs and the RNA polymerase complex enters the productive state (Grachev
and Zaychikov, 1980; Streney and Crother, 1985; Garpouisis and Gralla, 1985). After the
RNA polymerase clears the promoter, the o subunit is released, and the elongation phase
begins. The process ends with RNA chain termination.
There are many useful techniques to study thermodynamics and kinetics of specific
RNA polymerase-promoter interactions. Binding assays, run-off transcription (Strauss et
aL,1980; Rose et aL, 1984) and abortive initiation (Hawley and McClure, 1982) are among
the widely used techniques in this field. Chemical probes have been very useful to
investigate RNA polymerase-DNA complexes. DNA melting has been studied using
potassium permanganate which is known to react prefentially with pyrimidines in single
stranded DNA. This chemical probe can be used to detect DNA melting and open promoter
complex formation both in vitro and in vivo (Sasse-Dwight and Gralla, 1989; Kainz and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

10
Roberts, 1992). Information about DNA conformation and accessibility to solvent and the
presence of single-stranded DNA in open promoter complex has been accumulated using
1,10-phenanthroline-copper. There are some other chemical probes, such as hydroxy radical
which has been used in the investigation. DNase I footprinting analysis has helped to
demonstrate the interaction between RNA polymerase and promoter region and to define
the kinetic intermediates of transcription initiation. DNase I cleavage patterns are different
for open and closed complexes. Gel retardation analysis (Crothers, 1987; Gamer and
Revzin, 1986) also known as gel shift assay can be used to determine the binding affinity of
RNA polymerase for a promoter.
Regulators of transcription. Regulation of transcription initiation can be influenced by
components in addition to the -10 and -35 regions. These components include proteins or
DNA structural factors which either increase or decrease promoter activity and the
initiation of transcription. & coli CRP is a structurally characterized transcription activator
protein (Kolb et aL, 1993). In the presence of the allosteric effector cyclic AMP, CRP binds
to specific DNA sites located near or in CRP-dependent promoters. Indeed, the binding of
RNA polymerase to the promoter is enhanced by contact with bound CRP. CRP also
stimulates transcription by bending the DNA. The bend induced by CRP, is estimated to
be about 100° to 130° (Tompson and Landy,1988; Zinkle and Crothers, 1990). The
binding of CRP and of RNA polymerase are cooperative because they bend DNA in the
same direction. There are two classes of CRP-dependent promoters. In class I , the DNA
site for CRP is located upstream of the DNA site for RNA polymerase. The best
characterized class I promoter is the ZoePl promoter. In class II, however, the binding site
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

11
for CRP overlaps the DNA site for RNA polymerase. The gal? I promoter is an example
of class n. Amino acids 156-164 of CRP constitute an activator region essential for
transcription activation at both class I and class II CRP-dependent promoters, but it is not
essential for DNA binding and DNA bending (Bell et aL,1990; Eschenlauer and
Reznikoff 1991; Zhou et aL, 1993; Niu et al,1994; Niu et aL, 1996). For both class I and
class II promoters, the activating region functions through protein-protein interaction with
RNA polymerase. It is now believed that CRP interacts with the RNA polymerase a subunit
C-terminal domain (aCTD) and facilitates the binding o f aCTD to DNA adjacent to CRP
(Igarashi and Ishihama, 1991; Kolb et aL,1993; Chen et aL, 1994; Belyaeva et aL,1996).
For class I promoters, the interaction between the activating region and aCTD appears to
be the entire basis of transcription activation. The CRP-induced bend in class I tends to
become localized at an apical loop of supercoiled D N A , thus helping the DNA to wrap
around the promoter- bound RNA polymerase. In class n , the transcription activation
requires not only the interaction between the activating region and aCTD, but also an
interaction between a second site in the activating region of CRP and the RNA polymerase
a subunit N-terminal domain (Niu et aL, 1996). This new finding establishes that an
activator can interact with multiple targets within the transcription machinery and thereby
affect multiple steps of transcription initiation.
Another example of a regulatory protein which can bend the DNA is the P4 protein
of B. subtilis phage ((>29 . This protein is responsible for the switch from early to late
transcription in the bacteriophage development. Protein P4 is produced at an early stage
o f infection and activates the transcription from the promoter for late genes called PA3.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

12
This promoter is inactivated in the absence of protein P4. Activation of transcription in the
presence of P4 is via stabilizing the binding of B. subtilis RNA polymerase to the late
promoter as a closed complex (Nuez et aL, 1992). There is also evidence that P4 protein
interacts directly with RNA polymerase at the PA3 promoter (Nuez et aL, 1991; 1992;
Serrano et al,1991).
It has been shown recently that N4ssB, a single stranded DNA binding protein
encoded by bacteriophage N4, interacts with the carboxyl terminus of the RNA polymerase
P' subunit(Miller et aL, 1997). N4ssB activates transcription by the K coli a70 RNA
polymerase at the N4 late promoter.
The level of transcription activation is also influenced by DNA supercoiling. It has
been observed that supercoiling may either increase or decrease transcription activation in
some genes. Nevertheless, many other genes are not influenced by these phenomena (Pruss
and Drelica, 1989). DNA supercoiling may facilitate binding of RNA polymerase to the
promoter and activator protein to the DNA Transcription initiation at the lacPl promoter
of E. coli is assisted by DNA supercoiling in a CRP-dependent fashion (Meikleijohn and
Gralla, 1989).
Negative regulation of transcription can be also mediated through regulatory
proteins. In 1961 Jacob and Monad proposed that a regulator gene produces a repressor
that can interact with a DNA segment called the operator. Upon this interaction, initiation
of transcription is prevented. This type of negative control has been observed for the lac
operon. The product of the lac I gene is an allosteric repressor protein with two binding
sites. One binding site is for the operator region and the other is for the inducer molecule
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

13
(allolactose). Upon binding of the lac repressor to the operator region, RNA polymerase
is prevented from initiating transcription. However, in the presence of an inducer molecule,
this molecule binds to the repressor and alters the structure of the repressor so it no longer
binds to the operator. Thus the operator is unoccupied and the RNA polymerase can bind
the promoter and start transcription.
There are other factors which influence how proteins interact with the DNA during
transcription. DNA looping plays an important role in transcriptional control. This
phenomena is mediated by binding of regulatory proteins to two distinct sites. The gal
operon is one of the best characterized operons involving DNA looping. There are two
operator sequences in gal operon, Oe and O j. 0 E is found upstream of two overlapping
promoters and Ot is located downstream of the promoters (Irani et aL,1983; Adhya,1987;
Mandal et aL,1990). The loop structure is formed when a single repressor complex binds
both operators and consequently RNA polymerase is constrained from binding to the
structured promoters. An analogous mechanism has been observed in lac operon
regulation. However, in the lac operon the operators are located downstream of the
promoter (Mossing and Record, 1986; Kramer et aL, 1987).
Finally, it has been noted that sequence-dependent DNA curvature afreets many
processes in which the DNA is structured, such as DNA bending, wrapping and looping.
DNA curvature has been associated with many promoters.
DNA curvature. There are many unusual forms of DNA Among those, intrinsic DNA
curvature has been most studied the most and its biological significant has been shown.
Intrinsic DNA curvature is a phenomena that plays an important role in a variety of DNA
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

14
transactions. While there are various methods for detecting altered DNA structure, the most
sensitive and convenient method is polyacrylamide gel electrophoresis (Diekmann, 1987).
Curved DNA tends to migrate more slowly in an acrylamide gel than non-curved DNA of
equal length. Aberrant mobility is influenced by acrylamide concentration, temperature, and
salt concentration in the gel (Diekmann and Wang, 1985; Diekmann, 1987).
Curved DNA was first identified in electrophoretic studies of Idnetoplast minicircle
DNA from Leishmtmia tarantolate (Marini et aL, 1982). The first clue that kinetoplast DNA
might be bent came from the observation that a restriction fragment from a Ltaratolate
minicircle migrated anomalously slowly during electrophoresis on a polyacrylamide gel.
It was shown that runs of adenine, or an adenine tract (A-tract), would produce a small
bend in DNA helix. Intrinsically curved DNA is commonly characterized by runs of 4-6
adenine residues in phase with periodicity of B-form DNA This periodicity places the A
tracts on the same side of the helix and allows the angle of deflection from the helical axis
resulting from each A tract to be additive, thus leading to a large overall bend in the DNA
(Hagerman,1985; Diekmann, 1986; Koo et aL,1986).
Several theoretical models have been proposed to explain intrinsic DNA bending on
the molecular level. Among those, the first proposed model still is widely accepted. This
model, called the “wedge model” (Trifonove and Sussman, 1980), assumes smooth global
bending as a result of small additive wedges. The combination of tilt and roll cause
formation of a wedge or angle between adjacent AT base pairs in the DNA helix (Trifonov
and Sussman, 1980; Ulanovsky, 1987; Bolshoy et al., 1991). Such deformation in-phase
with the helical repeat cause a long-range curvature. It has been recently proposed that the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

15
DNA helix in an A-tract is characterized by base inclination in form of a negative roll
(Haran et aL, 1994). Progressive narrowing in the minor groove of the helix is the result
o f this negative roll.
In addition to A-tracts, there are other sequences which cause bending of DNA.
Sequences with specific periodic dinucleotide, such as AG, CG, GA, or GC, have been
shown to contribute in bending of DNA (Bolshoy et aL, 1991). Compared to A-tracts, they
bend the DNA to smaller degree (Fujimura,1988; Milton et aL,1990; Bolshoy et aL,1991).
Strong gel-mobility anomaly has been noticed for GGGCCC-containing DNA in the
presence of divalent ions (Brukner et aL, 1994). Perhaps the sequence dependent dynamic
feature of DNA is influenced by metal ions. Another intrinsically curved sequence appears
to be the GGCC element. This element is bent toward the major groove (Goodsell et al.,
1993).
DNA curvature in prokaryote promoters. DNA curvature generally is generated by an
A-tract sequence located upstream of the -35 region (Tanaka et aL,1991). Promoters
containing this upstream sequence are a well documented phenomena in prokaryotes. It has
been suggested that there is a relationship between intrinsic curved DNA and transcriptional
activity in certain E. coli ribosomal and tRNA promoters (Nishi and Itoh, 1986; Bauer et
aL, 1988). Reduced activity has been observed when the curved DNA is deleted in several
other promoters such as the ompF (Verda et aL, 1981), the bla promoter from pUC19
(Ohyama et aL,1992), the his and 1PP promoters (Verda et aL, 1981), and the Alul56
promoter from 5. subtilis phage SP82 (McAllister and Achberger, 1988).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

16
The effect o f CRP in the activation of gal promoter in E. coli has been well
documented. It has been shown that if the CRP binding site is replaced with synthetic or
natural curved DNA, the transcriptional activity can be restored (Bracco et aL,1989).
Similar results were reported using synthetic curved DNA in the lac promoter (Gartenberg
and Crothers, 1991). The addition of distamycin, a drug which is able to relax the DNA,
to gal p i promoter, which has a curved sequence upstream of promoter, caused a
significant reduction in transcriptional activity in the absence of the cAMP-CAP complex
(Lavigne et aL,1992). These results suggest that similar to protein induced bending,
sequence-specific DNA curvature enhances gene expression.
In studies from our laboratory, it was shown that sequence dependent DNA
curvature immediately upstream of the -35 region can enhance RNA polymerase binding to
promoters. Furthermore, it has been reported that deletion of curved DNA dramatically
decreased promoter utilization by the RNA polymerase from B. subtilis in vivo and in vitro.
It was also documented that one region of curved DNA will substitute for another when
properly aligned, and the rotational orientation (on the helix) of the curve relative to the
promoter was more important to function than the distance between the curved DNA and
the promoter (McAllister and Achberger, 1988, 1989). Hybrid promoters were created by
placing curved DNA from B. subtilis bacteriophage promoters on E. coli phage APL and
PR promoters. The addition of curved DNA influenced the binding of the RNA polymerase
from either B. subtilis or E. coli (McAllister, 1988). Wrapping of promoter DNA around
the E coli RNA polymerase was documented for one series of these promoters (Nickerson
and Achberger, 1995). B. subtilis RNA polymerase also wrapped the DNA upstream of the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

17
promoter around itself (Cheng, 1996). A model was proposed for all these finding. It was
proposed that curved DNA enhances promoter function by facilitating the wrapping of the
DNA around the RNA polymerase. This structured DNA-RNA polymerase complex allows
the DNA helix to be untwisted and the two strands separated for transcription initiation.
The focus of this research was to test for the interaction of E. coli RNA polymerase
with DNA upstream of the promoter consistent with DNA wrapping and to investigate the
nature of this phenomenon. In this study, the Alul56 and Ball29 promoters from the B.
subtilis bacteriophage SP82 were chosen for analysis. For each promoter, the nucleotide
sequence upstream from -35 region contains intrinsic DNA curvature. A DNase I
footprinting assay was used to study the interaction between promoter DNA and RNA
polymerase. In addition, the effect of temperature, salt concentration, and heparin on RNA
polymerase complex formation and wrapping was investigated. This study shows that E.
coli RNA polymerase wraps the DNA upstream of the promoter around itself. A model
for the nature of this interaction and its relation to the initiation of transcription is proposed.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

MATERIALS AND METHODS
Materials. PCR reagents were supplied by Perkin Elmer. Restriction enzymes were
purchased from either Bethesda Research Laboratories or New England Biolabs.
DNase I was purchased from Boehringer Manheim GmbH. Permanganate was purchased
from Sigma Chemical Company. The [y32P] ATP and [c^P] ATP were purchased from
New England Nuclear, Dupont. All other materials used were of the highest quality
available.
Plasmid transformation and isolation. Escherichia coli strain DH5a McR was grown in
LB medium (Miller, 1972) at 37°C. E coli cells were made competent using CaCl2 washes
(Lederberg and Cohen, 1974). Competent cells were mixed with 20-50 ng plasmid in the
test tube, placed on ice for 20 minutes and heat shocked at 42 °C for 2 minutes. The cells
were cooled briefly on ice and diluted with L-broth followed by incubation at 37°C with
shaking for 90 minutes. Between 0.2-1 ml of cells were concentrated and spread onto the
agar plates containing 50 mg/ml ampicillin to select for plasmid containing cells. Selected
colonies were isolated and cultured for plasmid isolation. Alkaline-lysis method of
Bimboim and Doly(1979) was used in order to isolate plasmid DNA To isolate highly
purified plasmid DNA large scale isolation was carried out followed by cesium chloride
density gradient separation in the presence of ethidium bromide. The purified DNA was
quantified by spectrophotometry.
Construction of Alul56 derivative promoters. An Alul56 promoter derivative which
contain uncurved DNA upstream of -35 region was constructed. In order to disrupt the
I S
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

19
intrinsic DNA curvature, Thymines were inserted into the middle of each A tract using site
directed mutagenesis (Chen and Przybyla, 1994). In this method, two rounds of PCR were
performed. In the first round, the AluUnc primer (5'-GCTAATATTCCTGAATA
ATATTGCAATAAGTTGTTGAC-3') and the M13/pUC reverse sequencing primer (-48)
24-mer (New England Biolabs) were used to incorporate the mutations into the promoter.
The template used in this round was plasmid pUC8 containing the original Alul56
promoter. The first PCR amplified fragment was gel purified and used directly as a primer
together with the M13/pUC sequencing primer
(-47) 24-mer (New England Biolabs) to direct a second round of DNA amplification using
the Alu 156 DNA template. Both rounds of PCR were performed in a Perkin Elmer Model
480 thermal cycle for 25 cycle at 94°C for 1 min, 60°C for 2 min, and 72°C for 3 min. The
products from the second round of amplification were digested with EcoRl and HindUI.
The restriction fragments were then gel purified and ligated to pUC8 digested with the same
enzymes. The promoter was named AluUnc.
To study the role of AT rich regions upstream of the promoter, two promoters in
which AT rich DNA was substituted for the curved DNA were constructed using PCR For
the first promoter the AT-rich region immediately upstream of the E. coli rmBP promoter,
called the UP-element (Ross et al., 1993), was inserted upstream of -35 region. To
construct this promoter, the M13/pUC sequencing primer (-47) 24-mer (New England
Biolabs) and AluUp primer (5-GCGAATTCAGAAAATTATTTTAAATTG
TTGTTGACTTTCTCTACGAGGTGTG-3') were used for DNA amplification with the
plasmid pUC8 containing the wild-type Alul56 promoter as template. The amplified
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

20
products were digested with EcoBl and H indSl and gel purified. Purified fragments were
ligated to pUC8 digested with the same enzymes. The promoter was named AluUp.
The second promoter contained 27 nucleotide long AT rich region upstream of the
promoter. The same procedures were used except that the 27AT primer (51-
GCGAATTC AT AATT AAT AATT AATTCGTTGACTTT-3') was used instead o f AluUp
primer. This promoter was named Alu27AT. In order to replace the DNA upstream of
the curved DNA in Alu27AT and AluUP with heterologous DNA in other promoters used
in this study, EcoRI DNA fragment from AluExt promoter DNA was inserted at the EcoRI
site of these promoter. AIu27At, AluUp, and AluUnc primers were synthesized in the Gene
Lab, Louisiana State University.
To study the effect of AT-rich region, two other promoters were constructed one
with a short stretch of AT and the other without any AT-rich region upstream of -3 5 region.
For construction of the first promoter, AluExt which does not possess any curved DNA
upstream of promoter was cut with EcoRI. The S' extension was made flush using Klenow
fragment and the gap was ligated. This promoter was named AT.
The last promoter lacked any AT-rich region upstream of promoter. In order to
construct this promoter, after cutting the AluExt with EcoRI, the S' over hang was digested
with S1 nuclease to remove the single stranded DNA and then ligated. This promoter was
named AluDel.
The promoter constructs were sequenced using the Circumvent Thermal Cycle
Sequencing kit (New England Biolabs) to insure that there were no changes in the promoter
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

21
sequence other than the intended mutations. Other promoters used in this study are listed
in Table 1. All of these promoters are from our laboratory collection.
Table 1. Promoters
Name Description Source
Alu 156Early gene promoter from phage SP82 containing curved DNA, Alul56 is 82% homologous to E. coli promoter consenses sequence
Laboratorycollection
AluExt Alul56 derivatives in which the curved DNA was replaced by a fragment of pBR322, base pairs 376-467
Laboratorycollection
Ball29B. subtilis bacteriophage SP82 promoter containing curved DNA, Ball29 is 90% homologous to E. coli promoter consenses sequence
Laboratorycollection
BalExt Ball29 derivatives in which the curved DNA was replaced by a fragment of pBR322, base pairs 376-467
Laboratorycollection
^Pr Bacteriophage X promoter Laboratorycollection
AlupR XpR derivatives in which curved DNA from Alul56 was inserted upstream of phage promoter
Laboratorycollection
BalpR XpR derivatives in which curved DNA from Ball29 was inserted upstream of phage promoter
Laboratorycollection
Primer labeling. Each primer was 5'end labeled using T4 polynucleotide kinase (New
England Biolabs) and 30 mCi [g-32? ATP] (Dupont, New England Nuclear) at 37°C for 30
min. In each 25 ml reaction 20 pmol of primer and 10 units of kinase were used. The
reaction was then denatured at 95 °C for 5 min and stored at - 20°C.
PCR amplification. Two primers, M13/pUC reverse sequencing primer (-48) 24-mer and
the M13/pUC sequencing primer (-47) 24-mer (New England Biolabs) used for
amplification. Only one of the two primers was labeled with [g-32P] ATP as it was.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

22
Drivitives of pUC8 containing the various promoter constructs were used as templates in
amplification reactions. Each reaction contained, 2.5 ml of supplied 10X PCR buffer, 2ml
of a mixture containing 2.5 mM of each deoxynucleotide substrate, 2 ml 25 mM MgClj, 0.1
ml AmpliTaq DNA polymerase, 4ml 5'end-labeled primer (0.8 mM) and 1ml second primer
(20 mM), 2 ml (0.2 ng/ml) DNA template and 11.5 ml water. PCR was performed in Perkin
Elmer DNA Thermal Cycler Model 480 for 25 cycles of the following, 94 °C for 1 min,
60°C for 2 min, and 72°C for 3 min. Labeled products were purified through a Sephadex
G-50 (Pharmacea Biotech) spin column (Neal and Florini, 1973).
RNA polymerase isolation. RNA polymerase was isolated from E. coli MRE600 as
described by Spiegelman et al. (1978) with minor modifications (Achberger and Whitely,
1980; McAllister and Achberger, 1988). Sonication was used to lyse the cells, and RNA
polymerase was purified by the sequential steps of polyethylene glycol-dextran phase
partitioning, ammonium sulfate precipitation, gel filtration chromatography, and DNA
cellulose chromatography. RNA polymerase activity was examined using an in vitro
transcription assay (Spiegelman et al., 1978). SDS-PAGE electrophoresis was used to
analyze subunit composition and purity of RNA polymerase (Lammli, 1970). The Bio-Rad
protein assay was used to calculate RNA polymerase concentration. RNA polymerase was
stored at -20°C in 35% glycerol.
DNase I footprinting analysis. To examine promoter-RNA polymerase interactions, a 190
pi of reaction mixture containing 40 mM Tris-HCl(pH 8.0), 10 mM MgCl^ 50 mM NaCl,
and 100,000cpm end-labeled DNA was incubated at 37°C with or without 2 mg RNA
polymerase. After 5 min incubation, 0.04 pg of DNase I(Boehringer Manheim Corp.) was
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

23
added to mixture. Following a 30 sec digestion with DNase I, 20 pi of salt solution
containing 3 M sodium acetate and 0.05 pg/pl yeast tRNA was added and two volumes of
phenol/chloroform mixture were added to stop the reaction. After mixing with a vortex
mixer, the phases were separated by centrifugation. The aqueous layer containing the
DNA was ethanol precipitated, rinsed with 80% ethanol, and dried under reduced pressure.
The DNA pellet was resuspended in 5 pi of loading buffer made with 10 ml deionized
formamide, 10 mg xylene cyanol, 10 mg bromophenol blue, and 0.2 ml 0.5 M EDTA, pH
7.0. Samples were heated to 95° C for 5 min and immediately transferred to ice. DNA
banding patterns was visualized by autoradiography after electrophoresing the samples on
6% polyacrylamide (acrylamide to bisacrylamide, 30:1.5) gel containing 7M urea in TBE
buffer at 1700 volts. Following electrophoresis the dried gels were analyzed by
autoradiography.
Gel retardation analysis. To analyze the relative afl&nity of E. coli RNA polymerase for
different promoters, gel retardation analysis was performed as described by Ausubel et al.,
1989 with minor modifications. End-labeled DNA fragments were incubated with or
without RNA polymerase in buffer containing 40 mM Tris-HCl (pH 7.8), 10 mM MgCl^
50 mMNaCl and 1 pg of non-specific competitor DNA at 37°C for 10 min. Following the
addition of 4 pi of loading dye (26% Ficoll, 0.1% Bromophenol blue), DNA-RNA
polymerase complexes were resolved in 4% polyacrylamide gel (acrylamide to
bisacrylamide, 30:2 ) with high ionic strength buffer (50 mM Tris, 400 mM Glycine, and 2
mM EDTA pH 8.5). The the gel was electrophoresed at 150 volts at room temperature for
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

24
two hours. Then gel was transferred to Whatman 3 MM paper , dried and
autoradiographed. Both free and RNA polymerase bound DNA were quantified by
densitometry of autoradiograms.
K M n04 footprinting. To detect open promoter complex formation, the basic reaction
conditions were the same used for DNase I footprinting. After 5 min incubation at 37°C
with or without RNA polymerase, 5 pi of 80 mM KMn04 was added to the mixture. The
reactions were stopped by adding 10 pi 2-mercaptoethanol and 15 pi of 3.0 M sodium
acetate after 5 min. The samples were extracted with two volume of a phenol/chloroform
mature and precipitated with ethanol. DNA pellets were washed with 80% ethanol, dried,
and resuspended in 5 pi formamide loading buffer as described before. After heating the
samples to 95 °C for 5 min, they were analyzed by polyacrylamide gel electrophoresis as
described for DNase I footprinting. Following electrophoresis the dried gels were analyzed
by autoradiography.
Single-round run-off transcription assay. All DNA templates used for the experiment
were synthesized by PCR amplification. For each reaction, an equal amount of DNA
template was used as determined by densitometry of ethidium bromide stained
polyacrylamide gels. In addition to template DNA, each reaction contained 0.5 pi of K coli
RNA polymerase (1.5 mg/ml), 2 pi of 10X transcription buffer (400 mM Tris-HCl, 100 mM
MgCl, 500 mMNaCl2), and water for the total reaction volume of 17.5 pi. The reactions
were mixed on ice and then transferred in duplicate to desired temperature for 5 min. Then
2 pi of nucleotides (20 mM GTP, 20 mM CTP, 20 mM UTP, 4 mM ATP, and 0.5 mCi/ml,
3000 Ci/mmol [a32?] ATP) and 1 pi of heparin (1 mg/ml) was added to each reaction at the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

25
same time. Half of the reactions were shifted to 37°C after 30 second and the rest remained
at incubated temperature. After 5 min., all of the reactions were stopped by adding urea to
a final concentration of 5M.
Transcription products were analyzed by electrophoresing the samples on 6%
polyacrylamide (acrylamide to bisacrylamide, 30:1.5) gel containing 7M urea in TBE buffer
at 500 volts. After electrophoresis, the gels were dried and exposed to X-Ray film (Kodak,
X-OMAT). Resulting bands were quantified by densitometry.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

RESULTS
The present work focuses on the role of intrinsic DNA curvature upstream of -35
promoter region in DNA wrapping and investigates the nature of interaction between £ coli
RNA polymerase and curved DNA The Alul56 and Ball29 promoters from the B. subtilis
bacteriophage SP82 are 83% and 90% homologous to the consensus sequence ofE. coli
promoters, respectively. These two promoters are recognized by £ coli RNA polymerase
and both have intrinsically curved DNA sequences upstream of -35 region (Figure 1).
Previous studies have demonstrated that upstream DNA curvature is required for efficient
utilization o f these promoters by B. subtilis RNA polymerase (McAllister and Achberger,
1988:1989). Curved DNA also appears to play a role in DNA structuring by £ coli RNA
polymerase. When curved DNA was spliced onto the phage XPR promoter, £ coli RNA
polymerase wrapped the curved DNA around the enzyme (Nickerson and Achberger,
1995). To study the phenomena of DNA wrapping, theAlul56andBall29 promoters
and their derivatives have been used in this study.
DNase I footprint analysis of the interaction between £ coli RNA polymerase and
Alul56 and Ball29 and their derivatives. It was of interest to determine if DNA
wrapping could occur in promoters other than those used in a previous study in our
laboratory. To investigate this problem, Akil56 and Ball29 and their derivatives was used.
The AluExt and BalExt are derivatives of Alul56 and Ball29, respectively, in which the
curved DNA upstream of promoter was replaced by DNA with no curvature (Figure 2).
The DNase I footprint analysis on both strands of all these promoters was performed. This
26
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
-60 -50 -40 -30 -20 -10 +1A 1u 156 I I I I I I ICTGCTAAAATTCCTGAAAAATTTTGCAAAAAGTTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
-60 -50 -40 -30 -20 -10 +1Ball29 I I I I I I IAAGAAAAAATATCTACAGAAAATATGAAAAAGTTGTTGACATTTCTTCCCATCCATGCTATAATAAAGTCA
Figure 1: Nucleotide sequence of the Alu 156 and Bal 129 promoters. Bases representing the +1 transcription start site,the -35 and the -10 regions are in bold type. The runs of adenine in the DNA upstream of the -35 region are underlined.
to-o

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
-35 -10 +1Alu156 | | |CTGCTAAAATTCCTGAAAAATTTTGCAAAAAGTTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
AluExtTGGGGAAGATCCCCGGGAATTCCCCCAGTGAATTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
-35 -10 +1Ball 29 | | |AAGAAAAAATATCTACAGAAAATATGAAAAAGTTGTTGACATTTCTTCCCATCCATGCTATAATAAAGTCA
BalExtCGATGGGGAAGATCCCCGGGAATTCCCCCAGTGAATTGACATTTCTTCCCATCCATGCTATAATAAAGTCA
Figure 2: Nucleotide sequence of the Alul56, Ball 29, and their derivatives. Bases representing the +1 transcription start,the -35 and -10 regions are in bold type. The runs of adenine in the DNA upstream of the -35 region are underlined. Nucleotide sequence downstream of the -35 region is the same for each wild-type promoters and its derivative.

29
assay can locate the protein binding site. Bound protein will protect the DNA from cleavage
with DNase I. DNase I cutting is also sensitive to protein-induced changes in the DNA
conformation. DNA conformation changes are often observed as DNase I hypersensitive
sites. DNase I digestion pattern indicative of wrapping is identified as a series of enhanced
cleavages (dark bands) followed by protection (light bands) with a 10 base pair periodicity.
Figure 3 corresponds to an autoradiogram after DNase I footprint analysis of both strands
of Alul56. Lanes marked by a minus sign are control reactions where the DNA fragment
containing the promoter was digested with DNase I in the absence of RNA polymerase.
When RNA polymerase was bound to the Alul56 promoter (lanes marked by plus sign), a
large protection area typical of results with prokaryote RNA polymerases was observed
from +24 to -55 in lower strand of DNA (Figure 3 A). The nucleotides are numbered
relative to the transcription start site at position +1. Hypersensitive sites, represented by
dark bands relative to control lane, were observed at -58, -59, -68, -69, -71, -92, -97, -101,
-102, and from -108 to -110. The curved DNA region from -40 to -65 is weakly cleaved
by DNase I because of the altered DNA conformation in this region. Small regions of
protection from DNase I were observed following these hypersensitive areas. For the upper
strand o f this promoter, the pronounced protected region was observed from +21 to -56
(Figure 3B). This region was interrupted with hypersensitive sites at -24, -44, and -47.
DNase I hypersensitive area upstream of promoter were around -57, -58, -59, -60, -67, -68,
-78, -79, -80, -81, -86, -87, -98, -99, -112 and -113 followed by protected regions. This
pattern of alternating protection and enhanced cleavages with 10 base pair periodicity in
both strands of Alul56 was indicative of wrapping.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

30
Figure 3: DNase I footprint analysis of the Alul56 promoter. Panel A represents thefootprinting of the lower (template) strand and panel B correspond the upper strand DNA footprint. DNase I digestion patterns in the absence (lanes with minus sign) and presence (lanes with plus sign) of RNA polymerase are shown. The bands are numbered relative to the transcription start site at position+1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
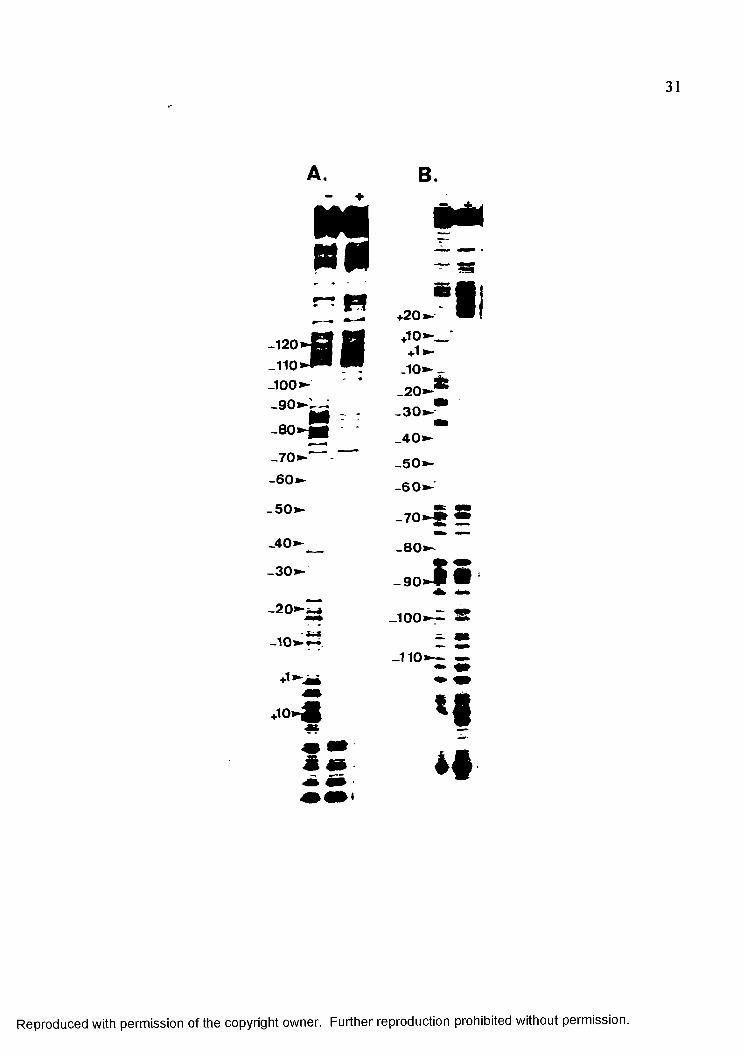
31
A. B.saW
-120 *»K
_100»-
-90**'.—m
-Q o ^m- 70 * - '
- 6 0 * -
- 5 0 * -
-40»~_
_30»-
—10 >- •
jo * M
■
~ ISm
m- -* +20 ►' I+1 ►
_10* - _
afe_ 20« -«i -
-30*-’
_40>-
_50>-
- 6 0 * -
-70**jgf 5
> J S
_80*-
-90«
_ioo»-^ S
-110^1 ~
4
i f
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

32
The DNase I pattern for AluExt, which lacks the curved DNA, was similar to that
of Alul56 promoter in the protected region from DNase I cleavage from +24 to -46 in
lower strand (Figure 4A) and +20 to -56 in upper strand (Figure 4B). Enhanced cleavages
were observed around the -60 region of both strands. Evidence of protein-DNA interaction
consistent with DNA wrapping was less obvious for this promoter. Figure 5 represents the
summary of the DNase I footprinting results on both strands o f Alul56 and AluExt
promoters compiled from three separate experiments.
Analysis of E. coli RNA polymerase binding to the Ball29 promoter has also
provided evidence for DNA wrapping. The footprint for lower strand of Ball29 extended
from +17 to -47, which was typical for other E. coli promoters (Figure 6A). Enhanced
cleavages upstream of promoter were observed at -49, -48, -58, -59, -60, -69, -70, -71,
-73 and -93, and -94. For upper strand of Ball29 (Figure 6B) the footprint region was from
+20 to -43 with enhanced cleavages at -23 and a gap at -21, and -22 . Hypersensitive areas
to DNase I were observed at -44 to -46, and -53. Protected regions were observed around
-50, -60, and -70. For technical reasons, footprint data upstream of -72 was unavailable for
the upper strand. This pattern of DNase I cleavage for both strands of Ball29 was in
agreement with presence of DNA wrapping.
The general footprint for BalExt was the same as Ball29 promoter downstream of
-40 except enhancement cleavages at -32, and -37 to -39 in upper strand and -37 in lower
strand (Figure 7). Almost no upstream interaction was detected for this promoter.
Evidence of DNA wrapping for BalExt was not obvious. This suggests that the curved
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

33
Figure 4: DNase I footprinting of the AluExt promoter bound by RNA polymerase.Footprint of the lower (template) strand of DNA is shown in panel A and panel B represents the footprinting of upper strand of the promoter. Presence and absence of RNA polymerase are indicated by plus and minus signs, respectively. The DNA bands are numbered relative to the transcription start site at position +1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

34
A. B.
- +
_100i.90* .80* _70»-| .60*
_50H
_40*
.30*
_20»-|
.10*
+ 1 *
+ 10*
+20.
+20*- +10^ +1^
I
I'i
. 10*- 20*
-30>
-40*-
- 5 0 » - #
— r iC f > - J ?I*
_70»- *
-80*
-90*-
. 100*
. 110*
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

35
Figure 5: The summary of DNase I footprint analysis on both strands of the Alul56(A) and the AluExt (B) promoters. The -10 and -35 regions are in bold type. The line over the upper strands and below the lower strands represent the area protected from DNase I cleavage. Hypersensitive regions are marked by arrows. The results were compiled from three separate experiments.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
- 1 2 0 - 1 1 0 - 1 0 0 - 9 0 - 8 0 - 7 01
GAAAGATATCCTAACAGCACAAGAGCGGAAACACGfl’TfGTTCTACATcjtGAACAACCTCTicT CTTTCTATAGGATTGTCGTGTTCTCGCCTTTGTGCAAAACAAGATGTAGGTCTTGTTGGAGACGA AAA AA A A------------- A A A
- 6 0 - 5 0 - 4 0 - 3 0 - 2 0 - 1 0 +1_ l w _ _ _ _ _ _ _ I_ _ _ f f T T - - - J_ _ _ _ _ _ _ _ _ _ _ _ I_ _ _ _ _ _ T _ _ _ _ I_ _ _ _ _ _ _ _ _ _ _ _ I_ _ _ _ _ _ _ _ _ _ _ _ It t c c t g a a a a a t t t t g c a a Aa a g ttg ttg a c tttc tc ta c g a g g t g tg g c a ta a ta a tc ttaAAAA'
t t t t a aggact t t t t a a a a c g t t t t t c a a ca actg aaa g a g a tg c tc c a c a c c g ta tta tta g a a t
+10 + 20 +301__________ Lt t t t t t t t tIttt
a c a a c a g c a g g a c g c t a g g a c g g a t c c g g g g a a t t cTGTTGTCGTCCTGCGATCCTGCCTAGGCCCCTTAAG ----------------A A
B - 6 0 - 5 0 - 4 0 - 3 0 - 2 0 - 1 0 +1
TCACCGATGgGGAAGATCCCCGGGAiTTCGTTGACTTTCTCTACGAGGtfGtfGGCATAMAATCTTAAGT^CTACCCCTTCTAGGGGCCCTTAAGCAACTGAAAGAGATGCTCCACACCGTATTATTAGAAT
+10 +20 +30________ I______ ^ I t t t t t t t t tItttACAACAGCAGGACGCTAGGACGGATCCGGGGAATTC TGTTGTCGTCCTGCGATCCTGCCTAGGCCCCTTAAG ------------------------------AA
u>On

37
Figure 6: DNase I footprint analysis o f Ball29 promoter. Lower (template) strand(panel A) and upper strand (panel B) of the promoter are footprinted in the absence (lanes with a minus sign) and presence (lanes with a plus sign) of RNA polymerase. The DNA bands are numbered relative to transcription start she a t+1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

39
Figure 7: DNase I footprint analysis of BalExt promoter. Lower (template) strand(panel A) and upper strand (panel B) of the promoter are footprinted in the absence (lanes with a minus sign) and presence (lanes with a plus sign) of RNA polymerase. The DNA bands are numbered relative to transcription start site a t+1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

40
A. B.
1 0 0 ^
m
_ 10j
+1»
+.10*Ks
, 2 0w «
r l
-90>
> 110*
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

41
DNA contributes to the interaction of RNA polymerase with DNA upstream of the -35
region. Figure 8 represents the summary of footprint analysis for both strands of BaI129
and BalExt.
There is an excellent agreement between the results for both promoters. DNase I
digestion pattern indicative of DNA wrapping was more obvious for both wild-type
promoters relative to the altered promoter lacking curved DNA There was a periodicity
of approximately 10 base pairs in the pattern of enhanced cleavages and protection in both
wild-type promoters. This pattern provided evidence that the upstream curvature of Alul5 6
and Ball29 was bent when wrapped around the RNA polymerase. It should be noted that
the possibility of second RNA polymerase binding to upstream region was ruled out by
footprinting the promoters at different RNA polymerase concentrations. At the lowest
concentration of RNA polymerase where binding to the promoter was observed, wrapping
was present. The DNase I pattern indicative of wrapping disappeared at the same point as
the main footprint (+20 to -50 region). This indicates that wrapping was not the result of
binding of another RNA polymerase to upstream sequence.
Nature of the interaction between RNA polymerase and DNA up stream of the -35
region. Electrostatic interactions between RNA polymerase and the backbone of DNA are
essential for sequence-independent DNA binding. Salts such as NaCl affect sequence-
independent binding between RNA polymerase and promoter DNA by competing with RNA
polymerase for charged phosphate residues on the DNA backbone (Roe and Record, 1985;
Suh et al., 1992). To explore the nature of interaction between RNA polymerase and DNA
upstream of the curved DNA, NaCl was used to disrupt electrostatic interactions. DNase
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

42
Figure 8: The summary ofDNase I footprinting results on both strands of Ball29 (A)and BalExt (B). The -10 and -35 regions are in bold type. The lines over the upper strand and below the lower strand represent the area protected from DNase I cleavage. Hypersensitive regions are marked by arrows. The results were compiled from at least two experiments for each strand.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
- 9 0 - 8 0 - 7 0
GAATTCCCCGGATCCGTCACCCCTAAGAA CTTAAGGGGCCTAGGCAGTGGGGATTCTT M - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - A A A - - - - -
- 6 0 - 5 0 - 4 0 - 3 0 - 2 0 - 1 0 +1
d^TACAGAAlATATGLuuJ^GTTGTTGACATTTCTTCCCATCCATGCTA' 'AATAAACTclAAAATATCTACAGITTTTA^AGATGTCTTTTATACTTTTTCAACAACTGTAAAGAAGGGTAGGTACGATATTATTTGAGT
+10 +20 +30
ta g a g a a c a a c a c ta tc aI atg a a^ J a^ I^ a t tATCTCTTGTTGTGATAGTTTACTTACCTCTCTAA - - - - - - - - - - - - - - - - - - - - - - - - - - - - - A A A - - - - - - A A A A A A A
B- 6 0 - 5 0 - 4 0 - 3 0 - 2 0 - 1 0 +1
GGGAAGATCCCCGGGAATTCCCCCAGTGAATTGACATTTCTTCCCATCCATGCTATAATAAACTCACCCTTCTAGGGGCCCTTAAGGGGGTCACTTAACTGTAAAGAAGGGTAGGTACGATATTATTTGAGT
+10 +20 +30
t A c J A g a a c I a c a c t a t c a I a t g a a t g g a g a g a t t a t c t c t t g t t g t g a t a g t t t a c t t a c c t c t c t a a
- - - - - - - - - - - - - - - - - - - A A A A A A A A A A

44
I footprinting analysis was performed for both wild-type promoters and their derivatives
using binding buffer containing various concentrations of NaCl. Since the results of DNase
I footprinting analysis were in agreement for both strands of each promoter, only the
interactions with lower strands were analyzed in this study.
Figure 9 demonstrates the effect of different salt concentration on DNA wrapping
for the lower strand of Alul56. Lane 1 and 7 represent control reactions which DNase I
footprinting was performed in the absence o f RNA polymerase. In lanes 2 to 6, salt
concentrations were incrementally increased from 50 mM to 200 mM, respectively. The
major footprint area (i.e., +20 to -56) consistent with the RNA polymerase bound to the
promoter, was observed at every salt concentration tested. Some differences were detected
among various salt concentrations. For example, bases -38 and -58 were not protected from
DNase I at salt concentrations greater than 100 mM. This indicates that RNA polymerase-
DNA complexes formed at lower salt concentrations were different from those at higher salt
concentrations. Evidence of DNA wrapping progressively faded with increasing NaCl
concentration. Visible changes were observed at different regions upstream of promoter.
For example, enhanced cleavages at -61, and -62 disappeared by increasing the salt
concentration to 100 mM. Obvious differences were displayed at the regions around -70,
-80, -90, -100, -110, and -120. Even bases very far upstream exhibit evident differences
among various salt concentration. Basically, enhanced cleavages and protection areas
indicative of DNA wrapping were lost gradually by increasing the salt concentration in
binding buffer.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

45
Figure 9: DNase I footprint analysis of the Alul56 promoter bound to RNApolymerase as a function o f NaCl concentration. Lanes 1 and 7 represent the DNase I digestion pattern in the absence o f RNA polymerase at 50 and 200 mM NaCl concentration. Lanes 2 to 6 correspond to the footprint of the promoter bound by RNA polymerase at 50, 75, 100, 150, and 200 mM NaCl respectively.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
46
i
Reproduced with permission o f the copyright owner. Further reproduction prohibited without permission.

47
The AluExt promoter was also footprinted as a function of salt concentration
(Figure 10). Footprint area from +20 to -53 was observed in all lanes in Figure 10 except
two control lanes. Minor protection around -50 and enhanced cleavages around -60
disappeared as the salt concentration increased. Even in the absence of detectable DNA
wrapping, the RNA polymerase-promoter complex that predominate at 50 mM NaCl
differed from those at 150 to 250 mM.
Effect of salt concentration on Ball29 promoter was also examined (Figure 11). As
observed for last two promoters, the large footprint area from +19 to -50 remained the same
at all salt concentrations except for changes at -40 and -48 regions consistent with the
presence of different complexes at various salt concentrations. Upstream interactions
indicative of wrapping were diminished by increasing the salt concentration. Enhanced
cleavages around -60, -71, and -91 disappeared at lane 5, which represents 150mMNaCl
concentration. Protected areas around -50, -60, -80, and -100 were diminished at the same
salt concentration. At 150 mM NaCl or greater, no changes in the DNase I pattern were
observed upstream of -59.
In the case of the BalExt promoter, as observed for AluExt, the limits of the
footprint at the promoter region were similar at all salt concentrations (Figure 12). Minor
changes observed upstream of promoter around -60 were diminished when salt
concentration was increased. For each promoter tested, protection at +16 to +20 increased
with increase of NaCl concentration.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

48
Figure 10: Effect of NaCl concentration on the DNase I footprint pattern o f AluExt.DNase I digestion patterns in the absence of RNA polymerase at SO and 250 mM NaCl concentration are shown in lanes 1 and 8. Lanes 2 to 7 correspond to the footprint of the promoter at SO, 75, 100, ISO, 200, and 250 mMNaCl, respectively.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

50
Figure 11: DNase I footprint analysis of the Ball29 promoter as a function of NaClconcentration. DNase I digestion patterns in the absence of RNA polymerase at 50 and 250 mMNaCl concentration are shown in lanes 1 and 8. Lanes 2 to 7 correspond to the footprint of the promoter at 50,75,100, 150, 200, and 250 mMNaCl, respectively.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

51
a 4 5 6 7 _8_
120 ►
t i l S_110 —
M M_ 9 0 > - iia_60*"
1
Ifi l lm m
Reproduced with permission o f the copyright owner. Further reproduction prohibited without permission.

52
Figure 12: Effect ofNaCl concentration on the DNase I footprint analysis of the BalExtpromoter. DNase I digestion pattern in the absence o f RNA polymerase at 50 and 250 mM NaCl concentration are shown in lanes 1 and 8. Lanes 2 to 7 correspond to the footprint o f the promoter at 50, 75, 100,150,200, and 250 mMNaCl.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

54
In general, as the salt concentration increased, wrapping decreased. DNase I
digestion patterns consistent with the RNA polymerase bound to the promoter were
observed at every NaCl concentration tested. Based on our results it can be concluded that
wrapping is more favorable at low salt concentration suggesting the electrostatic nature of
the phenomenon. This would be expected if these DNA-RNA polymerase interactions were
sequence independent.
The relationship between open promoter complex formation and DNA wrapping.
Potassium permanganate was used to probe the RNA polymerase-promoter complexes
formed at various NaCl concentration for both Alul56 and Ball29 (Figure 13). Potassium
permanganate is an oxidizing agent, which preferentially nicks at T and C residues in single
stranded DNA (Sasse-Dwight and Gralla, 1988). It was noted that while open promoter
complexes decreased at high salt concentration (i.e., 200 mM), they were still observed.
DNA wrapping as visualized by DNase I footprinting was lost at high NaCl concentrations.
This indicates that there are open complexes in which the DNA is no longer wrapped.
Test for DNA wrapping at different stages of transcription initiation. It is known that
at low temperature, the conformational changes leading to the open promoter complex do
not occur (Cowing et al., 1989; Mecsas et al., 1991). When Alul56 was tested for open
promoter complex formation at different temperatures, it was observed that at 0°C almost
no open complexes were formed (Figure 14). Upon raising the temperature, significant
increase in the amount of open promoter complex formation was observed. RNA
polymerase-promoter complexes at 0°, 5°, 15°, 22°, and 37°C were examined by DNase
I footprinting (Figure 15). The results were consistent with the presence of DNA wrapping
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

55
Figure 13: Open promoter complex formation at different salt concentrations forBall29 (A) and Alul56 (B). Potassium permanganate probe was used to detect open promoter complexes formed in the absence (lane with minus sign) and presence of RNA polymerase. The salt concentration used in each reaction is marked above the lines. The sequence of the open promoter complex regions are marked.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

56
A .
N a C l , o i n o in o o( m M ) I « ® w w g
' T
I*G
B .
i 8K s- r
f t f I fT AT T A T T
\ G
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

57
Figure 14: Effect of temperature on open promoter complex formation for the Alu 156promoter. Absence and presence of RNA polymerase in each reaction are shown by minus and plus sign respectively. The temperature at which each reaction was performed is shown. The sequence of open complex region is marked.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

58
Figure 15: DNase I footprint analysis of Alul56 as a function of temperature. Presence or absence of RNA polymerase is represented by plus and minus sign. The temperature of each reaction is shown.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

59
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

60
of all temperatures. Basically, the protected region indicative of sequence-specific RNA
polymerase-promoter interaction was detected at every temperature. Wrapping occurred
at low temperature in the absence of open promoter complexes. RNA polymerase footprint
was shorter at 0°C. Protection from the +1 region to +20 is significantly reduced at 0°C.
This is a signature footprint for closed promoter complexes.
To confirm the presence of wrapped closed complexes, heparin was used in DNase
I footprinting analysis. Heparin is strong DNA competitor that rapidly binds free RNA
polymerase but not enzyme stably bound to the DNA or enzyme engaged in RNA synthesis
(Walter et al., 1967). Closed complexes with short half lives are sensitive to effect of
heparin (i.e., they irreversibly dissociate in the presence of heparin). The Alul56 promoter
was footprinted at 37°C and 0°C (Figure 16) with and without a heparin challenge. RNA
polymerase was allowed to bind the AlulS6 promoter at the indicated temperature. To one
sample at each temperature, heparin was added for 30 second prior to DNase I treatment.
At 37° C, addition of heparin virtually eliminated the DNase I pattern consistent with DNA
wrapping. Since heparin eliminates sensitive closed promoter complexes, this indicated that
unwrapped heparin resistant open complexes were formed at 37°C. In addition, wrapped
complexes appeared to be closed complexes. It was previously demonstrated that at 0°C
almost no open promoter complexes were formed. DNase I digestion pattern consistent
with wrapping was observed in the absence of heparin at 0°C. Upon addition of heparin,
in addition to loss o f wrapping, the footprint was diminished too. This suggests that all
wrapped complexes were closed complexes.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

61
Figure 16: Effect of heparin on the footprint analysis of the Alul56 promoter bound toRNA polymerase at 37°C and 0°C. The minus sign represents the DNase I digestion pattern in the absence of RNA polymerase. Lane marked with the plus sign (+) represent the binding of RNA polymerase without the heparin challenge. Lanes in which the reactions were challenged with heparin are designated by “+H”.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

62
3 7 ° c 0 ° c
- + + H
100**_90»-
_80*-
_60>-
%T' ss*<- 3
- + + H
#
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

63
Effect of AT-rich regions in DNA wrapping and transcription efficiency. The a
subunit of RNA polymerase is known to bind an AT-rich sequence upstream of the -35
region (Ross et al., 1991). Since curved DNA is AT rich, the contribution o f the AT-rich
sequence was examined. To compare the role of AT-rich region upstream of promoter and
curved DNA in DNA wrapping and transcription efficiency, five different promoter-
containing fragments were constructed. In one promoter, AluUp, the DNA upstream of -35
region in AluExt promoter was replaced with the a subunit binding site, the so called “UP
element”, of.E. coli ribosomal RNA promoter rmB PI. In second promoter, AluUnc, a
non-curved AT-rich region was constructed by insertion of T’s in the middle of each A-tract
upstream of Alul56 promoter. These mutations eliminate curvature while preserving the
AT-rich nature of this promoter. Both Alu8AT and Alu27AT promoters contained AT-rich
regions upstream of the -35 region of AluExt promoter. However, the length of this region
in Alu8AT was shorter than that of Alu27AT. The AluDel promoter was constructed as a
control with no AT-rich region upstream of AluExt promoter. The relevant nucleotide
sequences of these promoters are listed in Figure 17 in comparison with the sequence of the
Alul56 and AluExt promoters.
To examine the specific interactions between E. coli RNA polymerase and the AT-
rich promoters, DNase I footprinting analysis was performed. This approach was used to
determine the role of AT-rich region in DNA wrapping. Each promoter sequence was
digested with DNase I in the absence and presence of RNA polymerase. Figures 18 and 19
illustrate the DNase I digestion patterns for the lower strands of each promoter. The DNase
I digestion pattern indicative of wrapping that was observed for Alul56 was reduced in the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
-35 -10 +1AluI56 | | |TAAAATTCCTGAAAAATTTTGCAAAAAGTTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
AluUncTAATATTCCTGAATAATATTGCAATAAGTTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
* * * *AluExtGAAGATCCCCGGGAATTCCCCCAGTGAATTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
AluUpCGAATTCAGAAAATTATTATTTTAAATTGTTGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
AluSATCACAGCTATGACATGATTACGAATTAATTCGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
Alu27ATTATCGCGAATTCATAATTAATAATTAATTCGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
AluDelCAACAGCAGGACAGCTATGACCATGATTACGTTGACTTTCTCTACGAGGTGTGGCATAATAATCTTA
Figure 17: Nucleotide sequence of the Alu 156, AluExt, AluDel, and AT-rich drivatives. AT-rich regions areunderlined. The -10, -35, and +1 regions are in bold type. An asterisk is placed under the bases in AluUnc that differ from Alu 156.

65
Figure 18: DNase I footprint analysis of the Alu 156, AluUnc, AluUp, and Alu27ATpromoters. Lanes depicting DNase I digestion in the presence and absence of RNA polymerase are indicated by plus and minus sign. The DNA bands are numbered relative to the transcription start site at position +1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

the copyright owner. Further
R e p ro d u c e d Witt' p e rm iss io n
reproduction prohibited without permission.

67
Figure 19: DNase I footprint analysis of the Alul56, AluExt, Alu8AT, and ATDelpromoters. DNase I patterns generated in the presence and absence of RNA polymerase are indicated by plus and minus sign. The DNA bands are numbered relative to the transcription start site at position +1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

69
AluUnc promoter (Figure 18). For the rest of promoters, evidence of protein-DNA
interactions consistent with DNA wrapping were not obvious. The DNase I footprinting
pattern for all promoters was similar to that of the original Alul56 in the region protected
from DNase I cleavage from +20 to -57. The AluUp promoter displayed a strongly
protected region between -37 and -58 corresponding to the UP element. The AluExt and
AluDel, which lacked AT-rich regions and curved DNA, displayed no significant DNA
wrapping. Likewise, DNA wrapping was not observed for Alu8AT promoter (Figure 19).
Gel retardation analysis was performed to determine the relative binding affinity of
the RNA polymerase for the original Alul56 and each of the promoters listed in Figure 17.
The binding reactions were carried out in binding buffer with equivalent amount of 32P end-
labeled promoter. For each concentration of RNA polymerase used, the DNA fragment
containing the Alu 156 promoter was efficiently bound by E. coli RNA polymerase (Figure
20 and 21). This suggest that RNA polymerase binding is enhanced by DNA curvature
upstream of promoter. An unexpected result was observed for AIu27AT promoter. RNA
polymerase displayed a dramatically low affinity for this promoter.
In order to compare the promoter strength in vitro, the single-round run-off
transcription assay was performed. This assay measures the number of the transcribable
complexes at the time of assay. Equal amount of DNA for each promoter was incubated
in the presence of RNA polymerase at each temperature tested. Heparin and mixture of all
four nucleotides triphosphates including [a 32P] ATP were added to each reaction. After
10 minutes, the reactions were stopped by adding urea and heating to 95°C for 5 minutes,
the sample were analyzed by polyacrylamide gel electrophoresis. The relative amount of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

70
Alu156 UncRNA
BO UN
27 AT
FREE
m
-r.—X-tV?}. ... / ^ «. £*^.
Figure 20: Gel retardation analysis of Alul56, AluUnc, AluUp, and Alu27AT.Equivalent amounts of32? end-labeled DNA fragments were incubated with various amounts of RNA polymerase at 37°C for 10 minutes, followed by electrophoresis. Lanes marked with a minus sign represent the absence of RNA polymerase. The RNA polymerase concentration was increased as it is marked by the symbol. The RNA polymerase amounts were 0.005, 0.016, and 0.05 pg. The DNA bands representing the RNA polymerase bound complexes and free DNA are marked.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

71
A l u 1 5 6 A l u E x t 8 A T A T D e l
RNAn
BOUND-
« • • - | * * * * # *4 * - | *
FREE —
Figure 21: Gel retardation analysis of Alu 156, AluExt, Alu8AT, and AluDel.Equivalent amounts of32? end-labeled DNA fragments were incubated with various amounts of RNA polymerase at 37°C for 10 minutes, followed by electrophoresis. Lanes marked with a minus sign represent the absence of RNA polymerase. The RNA polymerase concentration was increased as it is marked by the symbol. The RNA polymerase amounts were 0.02,0.07, and 0.2 pg. The DNA bands representing the RNA polymerase bound complexes and free DNA are marked.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

72
Figure 22: Transcribable promoter complexes formed as a function of temperature.Relative number of transcripts for Alu 156(9), AluExt(■), AluUnc(A), AluUp(V), and Alu27AT(^) without (A) and with (B) temperature shift were determined by densitometry from an autoradiograph of a polyacrylamide gel containing 32P labeled transcripts. Both panels contain representative data for single-round, run-off transcription in the presence of heparin.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

73
B:
COQ-O03CCO
<D.OE3z<D.>
JOa>o '
10003Q.O03CCO
<u.QE3z<u_>JSa>a:
10 15 20 25 30Temperature (°C)
15 20 25Temperature ( C)
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

74
transcripts formed for each promoter was determined by densitometry. The results are
shown in Figure 22 A. As expected, very little transcription was observed at 0°C, with
increased temperature, transcription increased for all promoters. At low temperature, the
Alu 156 displayed slightly better transcription relative to the other promoters. The Alu27AT
displayed very low transcription at all temperatures. For second set of reactions, 30 seconds
after addition of heparin and nucleotides, they were transferred to 37°C for 10 minutes prior
to stopping the reactions. This temperature shift allows the heparin resistance complexes
to form open complexes and start transcription. The relative number of transcripts as a
function of temperature are shown in Figure 22B. The Alu27AT was the weakest promoter
in all temperatures. This result confirms the result from gel retardation assay where RNA
polymerase exhibited a very low affinity for this promoter. In the temperature shift
reactions, significant transcription at low temperature was observed for the original Alu 156
promoter in contrast to the results observed for panel A, where small number of transcripts
where formed even at 10°C for all promoters tested. At temperatures above 30°C no
significant difference was observed among promoters (data not shown). This indicates that
the step in transcription initiation aided by curved DNA is normally inefficient at low
temperatures. The presence of curved DNA stimulates the formation of a heparin resistant,
closed complex. In the single round transcription assay, open complex formation (i.e.,
strand separation) appears to be the overall rate limiting step for Alul56 and its derivatives.
Effect of a mutation in the C-terminal domain of a subunit on the interaction between
RNA polymerase and DNA upstream of the -35 region. To study the role of a subunit
in the DNA wrapping, RNA polymerase with a deletion starting at position 235 of C-
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

75
terminal domain (CTD) of a subunit was used. RNA polymerase with and without the CTD
mutation were a gracious gift from Dr. Richard Gourse at University of Wisconsin-Madison.
It has been shown that protection of UP element DNA by the CTD mutant RNA polymerase
is severely reduced in footprinting experiment (Ross et al., 1993). Three hybrid promoters
A1uPr , BalPR, and AP R (McAllister, 1988) were used in footprinting study with mutant
RNA polymerase. The phenomenon of DNA wrapping has been documented for both
A1uPr and BalPR (Nickerson and Achberger, 1995). These two promoters contain the
curved region of Alul56 and BaI129 upstream of APR promoter. APR lacks the curved
DNA immediately upstream of the promoter and no obvious evidence of wrapping was
observed for this promoter.
The results of footprinting with the mutant RNA polymerase are illustrated in Figure
23. For each promoter, the DNase I digestion pattern in the absence of RNA polymerase
is shown in lane 1. Lane 2 represents the footprint of each promoter using wild-type RNA
polymerase and lane 3 shows the footprinting analysis using the CTD mutant RNA
polymerase. Evidence of DNA wrapping was observed for AluPR and BalPR when wild-
type RNA polymerase was used. For APR no obvious wrapping was observed. This result
was in agreement with published data. Addition of mutant RNA polymerase to APr did not
change the pattern except that an enhanced region was observed around -50 region, which
is part o f a binding site. Since mutation in a prevents binding of the subunit to this region,
mutant RNA polymerase can not protect this region from DNase I cutting. This “a
signature” was observed for other two promoters as well. DNA wrapping was diminished
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

76
Figure 23: DNase I footprint analysis of the AluP*, BalPR, and promoters as afunction of wild-type (lane 2) and mutant (lane 3) RNA polymerase. The nucleotide are numbered approximately relative to transcriptiion start sit at position+1.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

BalPB Xi77
-1 0
m *
-9o*-II £2 - 8 0 ^ 2 r ; -70*- i s
_ 6 0 » - ) ~ s *
_50»- — • »
_40>-
-30> -
_ 20* -
- 10 :
.1 ►
s a x
• •
- M i
—80i
—70i —60*
-50*
—401
a »
m
**— ( •fc_
-3 0 ►
_ 2 0 * - f
—101
+ 1 ;
j - -
1 1 •••£ w »
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

78
for both AIuPr and BaIPR when mutant RNA polymerase was used. This indicates that the
curved DNA is an a binding site and binding of a subunit to its binding site aids DNA
wrapping in the promoters containing curved DNA
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

DISCUSSION
Effect of DNA curvature immediately upstream of the -35 region on DNA wrapping
by the RNA polymerase. In a previous study using hybrid promoters containing curved
DNA, R coli RNA polymerase was shown to wrap the curved DNA around the enzyme
(Nickerson and Achberger, 1995). The hybrid promoters were made by splicing the curved
DNA found upstream of the Alu 156 and Ball 29 promoters of I?, subtilis phage SP82 onto
X PR promoter (McAllister, 1988). hi this limited study, the phenomenon of wrapping was
examined on one strand of those promoters. One of the first questions asked in the present
study was whether DNA wrapping could be observed for other promoters containing
curved DNA. To explore this question, the Alul56 and Ball29 promoters were used.
Both of these promoters were isolated from B. subtilis bacteriophage SP82 and contain
curved DNA immediately upstream of -35 region. DNase I footprinting was performed to
investigate the interaction between the R coli RNA polymerase and these promoters. Large
region of protection demonstrating the tight binding of RNA polymerase to -10 and -35
regions was observed for both promoters. The DNase I footprint patterns for these two
promoters exhibited enhanced cleavages and sites protected from DNase I repeated almost
every 10 base pair in the DNA upstream of promoters. This pattern was observed for both
strands of both promoters. This feature is indicative of DNA wrapping around RNA
polymerase. A similar pattern was observed when DNA is wrapped around proteins such
as nucleosomes (Shaw et al., 1976; Prunell et al., 1984). The DNase I patterns of both
strands combine to describe interactions characterized by region of DNA wrapped around
79
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

so
the enzyme and larger region of protection from -70 to -90 consistent with the DNA bound
within a fold or cleft of the RNA polymerase. For derivatives of Alu 156 and Ball29 which
lack curved DNA, the footprint of the promoter region was the same as wild-type
promoters. However, there was no obvious evidence of the DNase I cleavage pattern
consistent with DNA wrapping upstream o f these promoters. This indicates that curved
DNA facilitated the wrapping of DNA upstream of the promoter around the RNA
polymerase. Similar results were observed using the RNA polymerase from B. subtilis
(Cheng, 1996).
Model for the role of DNA curvature in RNA polymerase binding and transcription
initiation. Sequence-directed and protein-induced DNA curvature has been found in
promoters of both prokaryotes and eukaryotes. It has been documented that intrinsically
bent DNA plays a role in modulation of transcription initiation (Lamond and Travers, 1983;
Bossi and Smith, 1984; Gourse et al., 1986; McAllister and Achberger, 1988). Based on
previous studies in our laboratory, a model was proposed for the role of curved DNA in
transcription initiation. It was proposed that curved DNA enhances promoter function by
facilitating the wrapping of the DNA around the RNA polymerase. The structured DNA-
RNA polymerase complex allows the DNA helix to be untwisted and the two strands
separated for transcription initiation.
The model addressed the fact that there are multiple RNA polymerase promoter
complexes identified with conformational changes in the DNA and enzyme. Among these
complexes are the initial closed RNA polymerase-promoter complexes (RPCi)> a second
closed complex in which the strands of the DNA have been partially untwisted but remain
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

81
base paired (RPq), and the open promoter complex in which DNA strand separation has
occurred between the -10 region and +1 start site (RP0 ). Previous studies with B. subtilis
RNA polymerase have demonstrated that curved DNA associated with promoters aids
binding and the formation of open promoter complex (McAllister and Achberger, 1988;
Stemke, 1993). More recently DNase I footprinting studies with B. subtilis RNA-
polymerase demonstrated the presence of a closed complex in which the upstream DNA is
wrapped around the RNA polymerase (Cheng, 1996). This suggested that curved DNA
stimulates a step prior to RP0. Prior to the present study, little evidence, other than the
existence of DNA wrapping for hybrid promoters, was available to address this model to the
E. coli RNA polymerase.
The model proposed in our laboratory is similar to a model proposed for the role of
E. coli CRP, a DNA bending regulatory protein, in transcription activation. It has been
demonstrated that curved DNA can replace the CRP binding she for the lacP l promoter and
gcdpl promoters (Barco et al., 1989; Gartenberg and Crothers, 1991). Based on the model
proposed by Gartenberg and Crothers (1991), curved DNA localizes at the end of a
superhelical domain and facilitates the wrapping of DNA around RNA polymerase bound
to the promoter. This structure favors the transcription initiation.
Based on present the study we were able to relate K coli RNA polymerase to the
existing model and advance it. The presence of an additional closed promoter complex
between the initial closed complex and the open promoter complex has been identified in
many studies (Kadesch et al. 1982; Buc and McClure, 1985; Straney and Crothers, 1985).
The RPq is stable at low temperature (i.e., the predominant complex formed below 20°C)
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

82
and resistant to RNA polymerase competitors such as heparin and poly d(AT) (Buc and
McClure, 1985; Spascky et al. 1985; Schickor et al., 1990). The transition step from RPq
to RP0 became the rate limiting step below 20°C (Buc and McClure, 1985; Spassky et al.
1985). We now have evidence that the presence of curved DNA affects the formation of
RPq. A single-round transcription assay was used in a temperature shift format. RNA
polymerase was allowed to form complexes with various promoters at 0°, 10°, 20°, and
30 °C prior to the addition of a mixture of the four nucleotide triphosphates and heparin.
The nucleotides allow RNA syntheses from RPq, and the heparin binds free RNA
polymerase. RPC1 is said to be heparin sensitive since it dissociates rapidly and free RNA
polymerase is bound by heparin. If the reactions were shifted to 37°C after addition of
heparin and nucleotides, transcription from the low temperature complexes increased
dramatically for Alul56 relative to promoters without curved DNA. Curved DNA on
Alu 156 allowed formation of RPq at low temperatures (0° and 10°C) which quickly
isomerized to RP0 at 37°C. Taking this result in consideration, our updated model
proposes that by facilitating the wrapping of DNA around RNA polymerase (i.e., assisting
the RNA polymerase to structure the DNA), curved DNA enhances the transition from
heparin sensitive closed complex (RPC]) to a heparin-resistant, closed complex (RI^ ),
which is easily isomerized to form the open promoter complex. This allows more efficient
transcription initiation when this step is rate limiting. Since the DNA in RPa is believed
to be partially untwisted, curved DNA appears to aid DNA untwisting by RNA polymerase.
The nature of the interaction between K coli RNA polymerase and DNA upstream of
the -35 region. The interaction between RNA polymerase and DNA upstream of the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

83
curved DNA would be sequence independent if salt can be used to disrupt these
interactions. In other words, addition of salts such as NaCl interfere with the electrostatic
interactions between RNA polymerase and the phosphate charges on the DNA As the salt
concentration increased to ISO or 200 mM, wrapping diminished for both Alu 156 and
Ball29 promoters. The DNase I pattern consistent with RNA polymerase bound to the
promoter was observed at every salt concentration. This indicates that the population of
DNA-RNA polymerase complexes formed at low salt concentration were different to those
formed at high salt concentration. The wrapped DNA complex was formed at low salt
concentration suggesting that electrostatic interactions predominant.
Existence of unwrapped open promoter complexes. Since presence of different DNA-
RNA polymerase complexes were shown in various salt concentrations, it was important to
show if any of these complexes represent open promoter complexes. Potassium
permanganate was used to test for open promoter complex formation as a function of salt
concentration. While open promoter complexes were formed at all salt concentrations
tested, open promoter complex formation decreased with increasing the NaCl concentration.
Considering that no wrapped DNA complexes were observed above ISO mMNaCl there
were open promoter complexes in which the DNA is no longer wrapped. In order for RNA
polymerase to engage in active transcription wrapping must be released at some point.
Presence of unwrapped open promoter complexes are in agreement with this proposal.
Consistent with the proposed model, this indicates that wrapping must occur in steps prior
to open promoter complex formation.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

84
DNA wrapping in the absence of open promoter complex formation. According to our
model, for DNA wrapping to aid strand separation, wrapping must exist prior to formation
of the open promoter complex in which the strands are separated. The step of strand
separation is rate limiting at low temperatures. Thus the effect of temperature on the
structured complex of Alul56 was studied. At 0°C, as expected, almost no open promoter
complexes were found. Open promoter complexes were observed at S°C, and the
formation of these complexes increased when temperature was raised. Although minor
differences were observed in the DNase I digestion patterns upstream of the curved DNA
at each temperature, DNA wrapping was obvious at all temperatures. The minor changes
in the footprint indicates that there were different complexes formed at various
temperatures. The evidence supports the presence of DNA wrapping in the absence of open
promoter complex formation. This suggests that wrapping can occur in complexes
preceding the open promoter complex.
In our study, we went one step further and confirmed the presence of closed,
wrapped promoter complexes. This would be expected if indeed wrapping occurs prior to
strand separation. Many closed complexes are sensitive to effect of heparin. Heparin
challenge was used for complexes formed at 37°C and 0°C. Results indicate that at 37°C
all wrapped complexes were eliminated upon addition of heparin. Heparin challenge at 0°C
confirmed the idea that wrapped complexes were heparin sensitive, closed complexes. Most
o f the complexes formed at 0°C dissociated in the presence of heparin including all the
wrapped complexes. Since few open complexes were observed at 0°C, all wrapped
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

85
complexes were closed, and heparin sensitive. This agrees well with the model that the
wrapped complex is the RPC1 complex.
Comparison of the effect of AT-rich regions and curved DNA on wrapping and
transcription initiation. It has been established that the a subunit of & coli RNA
polymerase binds a specific AT-rich region upstream of the -35 region. It has been
suggested that A-tract curvature simply functions as an a binding site (Ross et al., 1993).
A collection of promoters containing AT-rich region upstream of promoter were studied and
compared to Alu 156 promoter. Based on the results of DNase I footprints from these
promoters, wrapping was more evident for wild-type Alu 156. However, small degree of
wrapping was observed for AluUnc and AluUp promoters. This supports the idea that the
curved DNA may stabilize a transient complex, formed by RNA polymerase with all
promoters, long enough to be detected by DNase I footprinting. The results from gel
retardation assay suggested that binding of E. coli RNA polymerase was strongest with the
promoter with curved DNA This suggests that there are more tight binding complexes in
the population of RNA polymerase-promoter complexes formed in the presence of curved
DNA. Some researchers have suggested that A-tract curved DNA is simply an AT-rich
binding site for a subunit of RNA polymerase. Although we agree with part of that
suggestion that curved DNA could act as an a binding site, our study showed that none
of the AT-rich DNA containing promoters, even the AluUp promoter, which contain the
so called UP element, were able to perform as well as Alul56. In addition, we showed that
not all AT-rich regions are beneficial. In our Alu27AT promoter, the long stretch of AT
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

86
seemed to be detrimental to promoter function. Perhaps for an AT-rich region to be good
a binding site, proper alignment of the a binding site relative to the promoter is necessary.
Contribution of a subunit of RNA polymerase on wrapping. The £ coli RNA
polymerase containing C-terminal truncation of 94 amino acids is stable in vivo and
assembles into holoenzyme. In other studies, this enzyme failed to protect the UP element
DNA of ribosomal RNA promoter rm BPl. hi other words, mutant a subunit could not bind
to its AT-rich binding site (Ross et al., 1993). We used this enzyme to explore the effect
of a subunit on DNA wrapping. DNase I footprints of this enzyme to the promoter region
was the same as that of wild-type enzyme. However, DNA wrapping for promoters
containing curved DNA was diminished when the mutant enzyme was used. This indicates
that A tracts involved in DNA curvature may function as a subunit binding site and
contribute to DNA wrapping.
Summary of the model. Based on the present study, we concluded that the C-terminal
domain of a contacts the A-tract curvature and promoters DNA structuring by the RNA
polymerase. Wrapping appears to be limited to RPC1, a heparin sensitive, closed complex.
The curved DNA promotes the transition from RPC1 to RPo, a heparin resistant, closed
complex. The RPq can quickly isomerize to RPq, an open promoter complex lacking DNA
wrapping. The effect of DNA curvature on overall transcription will only be observed under
conditions in which the RPC1 to RP^ transition is the rate-limiting reaction.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

REFERENCES
Achberger, E. C., M. D. Hilton, and H. R. Whiteley. 1982. The effect of the delta subunit on the interaction of Bacillus subtilis RNA polymerase with bases in a SP82 early gene promoter. Nucleic Acids Res. 10:2893-2910.
Adhya, S. 1987. The galactose operon, p. 1503-1512. In Neidhardt, F. C. (ed.), Escherichia coli and Salmonella typhimurium. America Society for Microbiology, Washington, D. C.
Amosti, D. N., and M. J. Chamberlin. 1989. Secondary a factor controls transcription of flagellar and chemotaxis genes in Escherichia coli. Proc. Natl. Acad. Sci. USA. 86: 830- 834.
Bauer, B. F., E. G. Kar, R. M. Elford, and W. M. Holmes. 1988. Sequence determinants for promoter strength in the leuV operon of Escherichia coli. Gene 63: 123-134.
Bell, A., K. Gaston, R. Williams, K. Chapman, A. Colb, H. Buc, S. Minchin, J. Williams, and S. Busby. 1990. Mutation that alterthe ability of the Escherichiacoli cyclic AMP receptor protein to activate transcription. Nucl. Acids Res. 18: 7243-7250.
Belyard, T., J. Bown, N. Fujita, A. Ishihama, and S. Busby. 1996. Location of the C- terminal domain of E.coli galactose operon regulatory region. Nucl. Acids res. 24: 2243- 2251.
Bossi, L., and D. M Smith. 1984. Conformational change in the DNA associated with an unusual promoter mutation in a tRNA operon of salmonella. Cell 39: 643-652.
Bolshoy, A., P. McNamara, R. E. Harrington, and E. N. Trifonov. 1991. Curved DNA without A-A: experimental estimation of all 16 DNA wedge angles. Proc. Natl. Acad. Sci. USA 88:2312-2316.
Bracco, L., D. Kotlarz, A. Kolb, S. Diekmann, and H. Buc. 1989. Synthetic curved DNA sequences can act as transcriptional activators in Escherichia coli. EMBO J. 8: 4289-4296.
Brukner, I., Susie, S., Dlakic, M., Savic, A., and Pongor, S. 1994. Physiological concentration of magnesium ions induces a strong macroscopic curvature in GGGCCC- containing DNA. J. Mol. Biol. 236: 26-32.
Buc, H., and R. McClure. 1985. Kinetics of open complex formation between Escherichia coli RNA polymerase and the lac UV5 promoter. Evidence for a sequential mechanism involving three steps. Biochemistry 24:2712-2723.
87
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

88
Carpousis, A. J., and J. D. Gralla. 1980. Cycling of ribonucleic acid polymerase to produce oligonucleotides during initiation in vitro at the lacUVS promoter. Biochemistry 19: 3245- 3253.
Carpousis, A. J., and J. D. Gralla. 1985. Interaction of RNA polymerase with lacUVS promoter DNA during mRNA initiation and elongation. Footprinting, methylation and rifampidn-sensitrvity changes accompanying transcription initiation. J. Mol. Biol. 183: 165- 177.
Chamberlin, M J. 1974. The selectivity of transcription. Annu. Rev. Biochem. 43: 721- 775.
Chamberlin, M. J. 1982. Bacterial DNA-dependent RNA polymerase, p.61-68. In P. D. Boyer (ed.), the enzymes XV-Nucleic Acids Part B. Academic press, New York.
Chen, B. and A. E. Przybyla. 1994. An efficient site-directed mutagenesis method based on PCR. BioTechniques . 17: 657-659.
Cheng, B. 1996. Role of intrinsic DNA curvature in DNA wrapping and transcription initiation by B.subtilis RNA polymerase. Ph.D. Dissertation, Auburn university.
Crothers, D. M 1987. Gel electrophoresis of protein-DNA complexes. Nature 325: 464- 465.
deCrombrugghe, B., Busby S. and Buc, H. 1984. Activation of transcription by the cyclic AMP receptor protein. Science 224: 831-837.
Diekmann, S. 1987. Temperature and salt dependence of the gel migration anomaly of curved DNA fragments. Nucleic Acids Res. 15: 247-265.
Diekmann, S., and J. C. Wang. 1985. On the sequence determinants and flexibility of the kinetoplast DNA fragment with abnormal gel electrophoretic mobilities. J. Mol. Biol. 186: 1- 11.
Doi, R. H. 1982. Multiple RNA polymerase holoenzymes exert transcriptional specificity in Bacillus subtilis. Arch. Biochem. Biophys. 214:772-781.
Doi, R. HL, and L. F. Wang. 1986. Multiple procaryotic ribonucleic acid polymerase sigma factors. Microbiol. Rev. 50:227-243.
Duffy, J. J., R. L. Petrusek, and E. P. Geiduschek. 1975. Conversion of Bacillus subtilis RNA polymerase activity in vitro by a protein induced by phage SP01. Proc. Natl. Acad. Sci. USA 72: 2366-2370.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

89
Duval-Valentin, G., and R Ehrlich. 1987. Dynamic and structural characterization of multiple steps during complex formation between K coli RNA polymerase and the tetR promoter from pSClOl. Nucleic Acids Res. 15:575-594.
Eschnelauer, A , and W. Reznikoffi 1991. Escherichia coli catabolite gene activator protein mutants defective in the controle of lac operon transcription. J. Bacteriol. 173: 5024-5029.
Fredric, R , T. Caramori, Y. Chen, A Galizzi, J. Helmann. 1995. Promoter architecture in the flagellar region of B.subtilis: high level expression of flagellin by the o15 RNA polymerase requires an upstream promoter element. Proc. Natl. Acad. Sci.USA.92:2582- 2586.
Fujimura, F. K. 1988. Point mutation in the polymavirus enhancer alters local DNA conformation. Nucleic. Acids Res. 16: 1987-1997.
Fujita, Y., and T. Fujita. 1987. The gluconate operon gnt of Bacillus subtilis encoded its own transcriptional negative regulator. Proc. Natl. Acad. Sci. USA 84:4524-4528.
Fukuda, R , and Alshihama. 1979. Subunits of RNA polymerase in function and structure. J. Mol. Biol. 87: 523-540.
Galas, D. J., M. Eggert, M., and M. Waterman. 1985. Rigorous pattem-recognition methods for DNA sequences. J. Mol. Biol. 186:117-128.
Garcia, E., S. Bancroft, S. G. Rhee, and S. Kustu. 1977. The product of the newly identified gene, glnE, is required for synthesis of glutamine synthetase in Salmonella. Proc. Natl. Acad. Sci. USA 74: 1662-1666.
Gamer, M. M., and A Revzin. 1986. The use of gel electrophoresis to detect and study nucleic acis-protein interactions. Trends Biochem. Sci. 11: 395-396.
Gartenberg, M R , and D. M Crothers. 1991. Synthetic DNA bending sequences increase the rate of in vitro transcription initiation at the Escherichia coli lac promoter. J. Mol. Biol. 219: 217-230.
Geidushik and Ito. 1982. Regulatory mechanisms in the development of lytic bacteriophages in Bacillus subtilis, p. 203-246. In Dubnar, D. A (ed.), the molecular biology of the Bacilli. Academic Press, New York.
Glass, RE., V. Nade, and M.G. Hunter. 1982. Informational suppression as a tool for the investigation of gene structure and function. Biochem. J. 203: 1-3.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

90
Goodsell, D. S., M. L. Kopka, D. Cascio, and R. E. Dickerson. 1993. Crystal structure of CATGGCCATG and its implications for A-tract bending models. Proc. Natl. Acad. Sci. USA. 90: 2930-2934.
Gourse, R.L., H.A deBoer, and M. Nomura. 1988. DNA determinants for RNA synthesis in Kcoli: growth rate dependent regulation, feedback inhibition, upstream activation, antitermination. Cell. 44: 197-205.
Grachev, M. A , and E. F. Zaychikov. 1980. Initiation by Escherichia coli RNA polymerase: transformation of abortive to productive complex. FEBS Iett. 115:23-26.
Graves, M. C., and J. C. Rabinowitz. 1986. In vitro and in vivo transcription of the Clostridiumpasteurianum ferredoxin gene. J. Biol. Chem. 261:11409-11415.
Hagerman, P. J. 1985. Sequence dependence of the curvature of DNA: A test of the phasing hypothesis. Biochemistry 24: 7033-7037.
Haldenwang, W. G., and R. Losick. 1980. Novel RNA polymerase a factor from Bacillus subtilis. Proc. Natl. Acad. Sci. USA 77: 7000-7004.
Haran, T. E., Kahn, J. D., and Crothers, D. M. 1994. Sequence elements responsible for DNA curvature. J. Mol. Biol. 244: 135-143.
Harley, C. B., andR. P. Reynolds. 1987. Analysis of E coli promoter sequences. Nucleic Acids Res. 15:2343-2360.
Hawley, D. K., and W. R. McClure. 1982. Mechanism of activation of transcription initiation from the XPm promoter. J. Mol. Biol. 157: 493-525.
Hawley, D. K., and W. R. McClure. 1983. Compilation and analysis of Escherichia coli promoter sequences. Nucleic Acids Res. 11:2237-2255.
Hegge-Aronis, R. 1993. Survival of hunger and stress: the rol of rpos in early stationary phage gene regulation in Escherichia coli. Cell 72: 165-168.
Henkin, T. M., and A. L. Sonenshein. 1987. Mutations in Escherichia coli lacUVS promoter resulting in increased expression in Bacillus subtilis. Mol. Gen. Genet. 209:467- 474.
Huang, X., A Decatur, A Sorokin, and J. D. Helmann. 1997. The Bacillus subtilis o* protein is an extracytoplasmic function o factor contributing to survival at high temperature. J.Bac. 179:2915-2921.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

91
Igarashi, K , and A. Ishihama. 1991. Bipartite functional map of E.coli RNA polymerase a subunit: involvement of the C-terminal region in transcription activation by cAMP-CRP. Cell. 65: 1015-1022.
Irani, M., L. Orosz, and S. Adhya. 1983. A control element within a structural gene: the gal operon of Escherichia coli. Cell 57:783-788.
Ishihama, A. 1992. Role of the RNA polymerase a subunit in transcription activation. Mol. Micro. 6:3283-3288.
Ishihama, A. 1981. Subunit assembly oiEscherichia coli RNA polymerase. Adv. Biophys. 14: 1-35.
Jacob, F., and J. Monod. 1961. Genetic regulatory mechanisms in synthesis of proteins. J. Mol. Biol. 3:318-356.
Juang, Y-L., and JJD.Helmann. 1994. Apromoter melting region in the primary a factor of B.subtilis. Identification of functionally important aromatic amino acids. J.Mol. Biol. 235: 1471-1488.
Kenny, T.J., and C.P. Moran. 1991. Genetic evidence for interaction of oA with two promoters in B.subtilis. J. Bactriol. 173: 3282-3290.
Kirkegaard, K., H. Buc, A. Spassky, and J. C. Wang. 1983. Mapping of single-stranded regions in duplex DNA at the sequence level: Single-stranded-specific cytosine methylation in RNA polymerase-promoter complexes. Proc. Natl. Acad. Sci. USA 80: 2544-2548.
Kobayashi, M , K Nagata, and A. Ishihama. 1990. Promoter selectivity of Escherichia coli RNA polymerase: effect of base substitutions in the promoter -35 region on promoter strength. Nucleic Acids Res. 18: 7367-7372.
Kolb, A., S. Busby, H. Buc, S. Garges, and S. Adhya. 1993. Transcriptional regulation by cAMP and its receptor protein. Ann. Rev. Biochem. 62: 749-795.
Kolb, A., K. Igarashi, A. Ishihama, M Levigne, M. Buckle, and H. Buc. 1993. E coli RNA polymerase, deleted in C-terminal part of its Alpha-subunit, intracts differently with the cAMP-CRP complex at lacpl and at the galpl promoter. Nucl. Acid Res. 21:319-326.
Koo, H-S., H-M. We, and D. M Crothers. 1986. DNA bending at adenine, thymine tracts. Nature 320: 501-506.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

92
Kramer, H., M. Niemoller, M. Amouyal, B. Revet, B. V. Wilcken-Bergmann, and H. Muller. 1987. lac repressor forms loops with linear DNA carrying two suitably spaced lac operator. EMBOJ. 6:1481-1491.
Lamond, A. I., and A A Travers. 1983. Requirement for an upstream element for optimal transcription of a bacterial tRNA gene. Nature 305: 248-250.
Lampe, M., C. Binnie, R. Schmidt, and R. Losick. 1988. Cloned gene encoding the delta subunit o f Bacillus subtilis RNA polymerase. Gene 67: 13-19.
Lavigne, M., Herbert, M., Kolb, A., and Buc, H. 1992. Upstream curved sequences influence the initiation of transcription at the Escherichia coli galactose operon. J. Mol. Biol. 224:293-306.
Lee, G., C. Talkington, and J. Pero. 1980. Nucleotide sequence of a promoter recognized by Bacillus subtilis RNA Polymerase. Mol. Gen. Genet. 180:57-65.
Li, Y., and JD . Helmann. 1994. The a subunit of B.subtillis RNA polymerase an allosteric effector o f the initiation and core-recycling phases of transcription. J.Mol.Biol. 239:1-14.
Losick, R., and J. Pero. 1981. Cascades o f sigma factors. Cell 25: 582-584.
Losick, R., and P. Stragier. 1992. Crisscross regulation of cell-type-specific gene expression during development in B. subtilis. Nature 355: 601-604.
Losick, R_, P. Youngman, P. J. Piggot. 1986. Genetics of endospore formation in Bacillus subtilis. Ann. Rev. Genet. 20: 625-669.
Mandal, N., S. Wen, R. Haber, S. Adhya, and H. Echols. 1990. DNA looping in cellular repression o f transcription of the galactose operon. Genes and Develop. 4: 410-418.
Marini, J. C., S. D. Levene, D. M. Crothers, and P. T. Englund. 1982. Bent helical structure in kinetoplast DNA Proc. Natl. Acad. Sci. USA 79: 7664-7668.
McAllister, C.F. Ph.D dissertation LSU. 1988.
McAllister, C. F., and E. C. Achberger. 1988. Effect of polyadenine-containing curved DNA on promoter utilization in Bacillus subtilis. J. Biol. Chem. 263: 11743-11749.
McAllister, C. F., and E. C. Achberger. 1989. Rotational orientation of upstream curved DNA affects promoter function in Bacillus subtilis. J. Biol. Chem. 264: 10451-10456.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

93
McClure, W. R 1985. Mechanism and control of transcription initiation in prokaryotes. Ann. Rev. Biochem. 54: 171-204.
Meiklejohn, A. L., and J. D. Gralla. 1989. Activation at the lac promoter and its variants, synergistic effects o f catabolite activator protein and supercoiling in vitro. J. Mol. Biol. 207: 661-673.
Miller, A., D. Wood, RH. Ebright, L.B. Rothman-Denes. 1997. RNA polymerase P’ subunit: A target of DNA binding-independent activation. Science. 275:1655-1657.
Moran, C. P. Jr., N. Lang, S. F. J. Le Grice, G. Lee, M. Stephens, A. L. Sonenshein, J. Pero, and R Losick. 1982. Nucleotide sequences that signal the initiation of transcription and translation in Bacillus subtilis. Mol. Gen. Genet. 186: 339-346.
Mossing, M. C., and M. T. Record, Jr. 1986. Upstream operators enhance repression of the lac promoter. Science 233:889-892.
Nickerson, C.A., E.C. Achberger. 1995. Role of curved DNA in binding o f E .coli RNA polymerase to promoters. J. Bacteriol. 177: 5756-5761.
Niu, W., K. Young, T. Gregory, H. Tomasz, and R H. Ebright. 1996. Transcription activation of class II CAP-dependent promoters: Two interactions between CAP and RNA polymerase. Cell. 87: 1123-1134.
Nuez, B., F. Rojo, I. Barthelemy, and M. Salas. 1991. Identification of the sequences recognized by phage ({>29 transcriptional activator: possible interaction between the activator and the RNA polymerase. Nucleic Acids Res. 19: 2337-2342.
Nuez, B., F. Rojo, and M. Salas. 1992. Phage ({>29 regulatory protein P4 stabilizes the binding of RNA polymerase to late promoter in a process involving direct protein-protein contacts. Pro. Natl. Acad. Sci. USA 89: 11401-11405.
Ohyama, T., M Nagumo, Y. Hirota, and S. Sakuma. 1992. Alteration of the curved helical structure located in the upstream region of the P-lactomase promoter of plasmid pUC19 and its effect on transcription. Nucleic Acids Res. 20: 1617-1622.
Pabo, C.O., and RT. Sauer. 1984. Protein-DNA recognition. Arm. Rev. biochem. 53: 293- 321.
Prunell, A., I. Goulet, Y. Jacob, and F. Goutorbe. 1984. The smaller helical repeat of poly(dA)-poly(dT) relative to DNA may reflect the wedge property of the dA-dT base pair. Eur. J. Biochem. 138: 253-257.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

94
Pruss, G. J., and K. Drlica. 1989. DNA supercoiling and prokaryotic transcription. Cell 56: 521-523.
Roe, J.-R , R. R. Burgess, and M. T. Record, Jr. 1984. Kinetics and mechanism of the interaction ofEscherichia coli RNA polymerase with the APR promoter. J. Mol. Biol. 176: 495-521.
Roe, J.-R , R. R. Burgess, M. T. Record. 1985. Temperature dependence of the rate constants of the Escherichia coli RNA polymerase APR promoter interaction. Assignment of the kinetic steps corresponding to protein conformational change and DNA opening. J. Mol. Biol. 184:441-453.
Roe, J .-R , and M. T. Record Jr. 1985. Regulation of the kinetics of the interaction of Escherichia coli RNA polymerase with the APR promoter by salt concentration. Biochemistry 24:4721-4726.
Rosenberg, M , and D. Court. 1979. Regulatory sequences involved in the promotion and termination of RNA transcription. Ann. Rev. Genet. 13: 319-353.
Rosenberg, S., T. K. Kadesch, and M. J. Chamberlin. 1982. Binding of Escherichia coli RNA polymerase holoenzyme to bacteriophage T7 DNA Measurements of the rate of open complex formation at T7 promoter. J. Mol. Biol. 155: 31-51.
Ross, W., K. K. Gosink, J. Salomon, K. Igarashi, C. Zou, A Ishihama, K. Severinov, and R Gourse. 1993. A third recognition element in bacterial promoters:DNA binding by the a subunit of RNA polymerase. Science 262: 1407-1413.
Ruzhong, J., K.A Shari£ and J.S.Krakow. 1995. Evidence for contact between the cyclic AMP receptor protein and the a70 subunit of E.coli RNA polymerase. J. Biol. Chem. 270: 19213-19216.
Sasse-Dwight, S., and J. D. Gralla. 1989. KMn04 as a probe for lac promoter DNA melting and mechanism in vivo. J. Biol. Chem. 264: 8074-8081.
Schickor, P., W. Metzger, W. Weler, R Lederer, and R Heumann. 1990. Topography of intermediates in transcription initiation oiE co li. EMBO. J. 9: 2215-2220.
Schmidt, R., P. Margolis, L.Duncan, R. Coppolecchia, C. Moran, and R. Losick. 1990. Controle of developmental transcripton factor aF by spomlation regulatory proteins SPOHA and SPOIIAB in B. Subtilis. Proc. Natl. Acad. Sci. USA 87: 9221-9225.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

95
Serrano, M , I. Barthelemy, and M. Salas. 1991. Transcription activation at a distance by phage <f>29 protein P4 effect of bent and nonbent intervening DNA sequences. J. Mol. Biol. 219: 403-414.
Shaw, B. R., T. M. Herman, R. T. Kovacic, G. S. Beaudreau, and K. E. Van Hold. 1976. Analysis of subunit organization in chicken erythrocyte chromatin. Proc. Natl. Acad. Sci. USA 73: 505-509.
Siebenlist, U., R. B. Simpson, and W. Gilbert. 1980. K coli RNA polymerase interacts homologously with two different promoters. Cell 20:269-281.
Siegele, D.A, J.C. Hu, W. A. Walter, C. A. Gross. 1989. Altered promoter recognition by mutant forms of the a70 subunit of E xoli RNA polymerase. J. Mol. Biol. 206: 591-603.
Spassky, A , and D. Sigman. 1985. Nuclease activity of 1,10-phenanthroline-copper ion. Conformational analysis and footprinting of the lac operon. Biochemistry 24: 8050-8056.
Stemke, D. J. 1993. Transcription initiation studies with B.subtilis promoters containing curved DNA Ph.D. Dissertation, LSU.
Straney, D. C., and D. M. Crothers. 1985. Intermediates in transcription initiation from the E. coli lac UV5 promoter. Gene 43: 449-459.
Talkington, C., and J. Pero. 1978. Promoter recognition by phage SPOl-modified RNA polymerase. Proc. Natl. Acad. Sci. USA 75: 1185-1189.
Tanaka, K., S. Muramatsu, H. Yamada, and T. Mizuno. 1991. Systematic characterization of curved DNA segments randomly cloned from Escherichia coli and their functional significance. Mol. Gen. Genet. 226: 367-376.
Tanaka, K., Y. Takayanagi, N. Fujita, A Ishihama, and H. Takahashi. 1993. Heterogeneity of the principal o factor in Escherichia coli: the rpoS gene product, o3*, is a second principal a factor of RNA polymerase in statinary-phase Escherichia coli. Proc. Natl. Acad. Sci. USA 99:3511-3515.
Thompson, J. F., and A Landy. 1988. Empirical estimation of protein-induced DNA bending angles: application to X site-specific recombination complexes. Nucleic Acid Res. 16: 9687-9705.
Trifonov, E. N., and J. L. Sussman. 1980. The pitch of chromatin DNA is reflected in its nucleotide sequence. Proc. Natl. Acad. Sci. USA 77: 3816-3820.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

96
Ulanovsky, L. E., and E. N. Trifonov. 1987. Estimation of wedge components in curved DNA. Nature 326: 720-721.
Van Ulsen, P., M.H. Lebrand, M. Kainz, R. Collard, L. Zulianello, P. Van Deputte, R. L. Gourse, and N. Goosen. 1997. Function of the C-terminal domain of Alpha subunit o f Exoli and integration host factor-mediated activation of the early promoter of bacteriophage Mu. J. Bac. 179: 530-537.
Verde, P., R. Frunzio, P. P. di Nocera, F. Blasi, and C. P. Bruni. 1981. Identification, nucleotide sequence and expression of the regulatory region of the histidine operon of Escherichia coli K12. Nucleic Acids Res. 9: 2075-2086.
von Hippel, P. H., D. G. Bear, W. D. Morgan, and J. A. McSwiggen. 1984. Protein- nucleic acid interactions in transcription: A molecular analysis. Ann. Rev. Biochem. 53: 389-446.
Waldberger, C., T. Gardella, R. Wang, and M.M. Susskind.1990. Change in conserved region 2 of E xoli o70 affecting promoter recognition. J. Mol. Biol. 215: 267-276.
Walter, G., W. Zillig, and E. Fuchs. 1967. Initiation of DNA synthesis and the effect of heparin on RNA polymerase. Eur. J. Biochem. 3: 194-201.
Weiss, S. B., and L. Gladstone. 1959. A mammalian system for the incorporation of cytidine triphosphate into ribonucleic acid. J.Am Chem. Soc. 81:4118-4119.
Whiteley, H. R., E. C. Achberger, and M D. Hamilton. 1982. The effect of the delta subunit on the interaction of B.subtilis RNA polymerase with SP82 early gene promoters. Promoters: structure and function. 267-282.
Whipple, F. W. and Sonenshien, A L. 1992. Mechanism of initiation of transcription by Bacillus subtilis RNA polymerase at several promoters. J. Mol. Biol. 223: 399-414.
Wiggs, J. L., J. W. Bush, and M. J. Chamberlin. 1979. Utilization of promoter and termination sites on bacteriophage T7 DNA by RNA polymerase from a variety of bacterial orders. Cell 16:97-109.
Wiggs, J. L., M. Z. Gilman, M. J. Chamberlin. 1981. Heterogeneity of RNA polymrease in Bacillus subtilis: Evidence for an additional o factor in vegetative cells. Proc. Natl. Acad. Sci. USA 78: 2762-2766.
Zinkel, S. S., and D. M Crothers. 1990. Comparative gel electrophoresis measurement of the DNA bend angle induced by the catabolite activator protein. Biopolymers 29: 28-38.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

97
Zhou, Y., S.Busby, and R. Ebright. 1993. Identification of the functional subunit of a dimeric transcription activator protein by use of oriented heterodimers. Cell. 73: 375-379.
Zhou, Y., T. Markel, and R. Ebright. 1994. Characterization of the activating region of E .coli catabolite gene activator protein (CAP):II. Role of calss I and class II CAP- dependent promoters. J. Mol. Biol. 243: 603-610.
Zhou, Y., P. S. Pendergrast, A. Bell, R Williams, S. Busby, and R. Ebright. 1994. The functional subunit of a dimeric transcription activator protein depends on promoter architecture. EMBO J. 13:4549-4557.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

VITA
Parisa Jazbi received her bachelor of science degree in Zoology from Shiraz
University in Shiraz, Iran. In August 1993, she joined the graduate program in the
Department of Microbiology at Louisiana State University. She has worked as a teaching
assistant for past four years. Her research focuses on investigating the role o f DNA
wrapping in initiation of transcription using K coli RNA polymerase. Parisa is currently
attending Louisiana State University, where she is a candidate for the doctor of philosophy
degree in Microbiology with a minor in Biochemistry.
98
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

DOCTORAL EXAMINATION AND DISSERTATION REPORT
Candidate: Parisa Jazbi
Major Field: Microbiology
Title of Dissertation: DNA Wrapping and the Nature of Interaction
between E.coli RNA Polymerase and Promoter DNA
Approved:IM
Major Professor and Chairman
EXAMINING COMMITTEE:
/ c f * ~Cj~cc
A . J . i M l
1 1
Date of Examination:
October 17, 1997
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
IMAGE EVALUATION TEST TARGET (Q A -3 )
1.0
l.l
13.2
J 2.2
*" E Hill 2-0
.8
1.25 1.4 1.6
150mm
V
<P;/
A P P L I E D IIWIGE . Inc1653 East Main Street
- ■ Rochester. NY 14609 USA = Phone: 716/482-0300
-= = r -= = Fax: 716/288-5989
0 1993. Applied Image. Inc.. All Rights Resetved
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Related Documents











