For Peer Review Only Diversity and pathogenecity of Vibrio species in cultured bivalve molluscs. Journal: Environmental Microbiology and Environmental Microbiology Reports Manuscript ID: EMI-2009-0874.R1 Manuscript Type: EMIR - Minireview Journal: Environmental Microbiology Reports Date Submitted by the Author: 12-Dec-2009 Complete List of Authors: Hidalgo, Roxana; Universitat Rovira i Virgili, Ciencias Mèdiques Bàsiques Balboa, Sabela; Universidad de Santiago de Compostela, Microbiologia y Parasitologia Romalde, Jesús; Universidad de Santiago de Compostela, Microbiologia y Parasitologia Figueras, Maria; Rovira i Virgili, Microbiology Keywords: bacteria, microbial communities, pathogen ecology Wiley-Blackwell and Society for Applied Microbiology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

For Peer Review O
nly
Diversity and pathogenecity of Vibrio species in cultured bivalve molluscs.
Journal: Environmental Microbiology and Environmental Microbiology
Reports
Manuscript ID: EMI-2009-0874.R1
Manuscript Type: EMIR - Minireview
Journal: Environmental Microbiology Reports
Date Submitted by the Author:
12-Dec-2009
Complete List of Authors: Hidalgo, Roxana; Universitat Rovira i Virgili, Ciencias Mèdiques Bàsiques Balboa, Sabela; Universidad de Santiago de Compostela, Microbiologia y Parasitologia Romalde, Jesús; Universidad de Santiago de Compostela, Microbiologia y Parasitologia Figueras, Maria; Rovira i Virgili, Microbiology
Keywords: bacteria, microbial communities, pathogen ecology
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
1
1
2
3
Diversity and pathogenecity of Vibrio species in cultured bivalve molluscs. 4
5
6
Roxana Beaz-Hidalgo1, Sabela Balboa 2, Jesús L. Romalde2 & Maria José Figueras1* 7
8
1 Departament de Ciències Mèdiques Bàsiques, Facultat de Medicina i Ciències de la Salut. 9
IISPV. Universitat Rovira i Virgili, Reus, Spain. 10
2 Departamento de Microbiología y Parasitología. CIBUS. Facultad de Biología. Universidad de 11
Santiago de Compostela. Santiago de Compostela, Spain. 12
13
14
15
Running title: Vibrio spp., bivalve molluscs, pathogens. 16
17
18
Revised version Ms. EMI-2009-0874. December, 2009 19
20
21
22
23
24
*Corresponding author. Mailing address: Unitat de Biologia y Microbiologia. Facultat de 25
Medicina i Ciencia de la Salut. Universitat Rovira i Virgili. Sant Llorenc 21, 43201 Reus, 26
Barcelona, Spain. Phone: +34-97-759321. E-mail: [email protected] 27
28
29
Page 1 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
2
Summary 30
Shellfish production is seriously affected by bacterial pathogens that cause high losses in 31
hatcheries and in the aquaculture sector. A number of Vibrio species are considered important 32
pathogens and have provoked severe mortality outbreaks. The pathologies caused by vibrios in 33
bivalves have been described since the 1960s; however, over recent years, successive episodes 34
of high mortality have been recorded due to these microorganisms. The present work provides 35
an updated overview of the different studies performed in relation with the diversity of Vibrio 36
spp. associated to bivalves. Special attention is given to the main Vibrio diseases and implicated 37
species affecting the different life stages of cultured bivalves. 38
39
Introduction 40
Coastal and estuarine environments are growing areas of bivalve molluscs with hatcheries for 41
seed production of commercialised species, this being an important industry in many countries. 42
Due to their filter-feeding habit, bivalves normally accumulate a rich bacterial microbiota, 43
composed of various species belonging to different genera like Vibrio, Pseudomonas, 44
Acinetobacter, Photobacterium, Moraxella, Aeromonas, Micrococcus and Bacillus (Murchelano 45
and Brown, 1970; Kueh and Chan, 1985; Prieur et al., 1990). One of the main problems in the 46
culture of bivalve molluscs is the repeated episodes of mortality due to bacterial infections that 47
reduce the production and cause high economical losses. Some members of the genus Vibrio 48
have been described as the main causal agents of diseases affecting all life stages of bivalve 49
molluscs: larval, juveniles and adults (Liu et al., 2000; 2001; Allam et al., 2002; Waechter et 50
al., 2002; Lee et al., 2003; Paillard et al., 2004 and references therein; Anguiano-Beltrán et al., 51
2004; Estes et al., 2004; Gay et al., 2004 a, b; Gómez-León et al., 2005; Prado et al., 2005; 52
Labreuche et al., 2006 a, b; Garnier et al., 2007; 2008; Gómez-León et al., 2008). The diversity 53
of Vibrio species associated with bivalves in different geographical areas has also been the 54
subject of various studies (Montilla et al., 1994; Hariharan et al., 1995; Arias et al., 1999; 55
Pujalte et al., 1999; Maugeri et al., 2000; Caballo and Stabili 2002; Castro et al., 2002; 56
Guisande et al., 2004; Lafisca et al., 2008; Beaz-Hidalgo et al., 2008). It is well known that 57
environmental parameters, such as variations in the water temperature and salinity, can 58
influence the diversity of Vibrio spp. in the environment as well as the physiological state of the 59
bivalve and its susceptibility to bacterial infections (Arias et al., 1999; Pujalte et al., 1999; 60
Maugeri et al., 2000; Paillard et al., 2004; Garnier et al., 2007). 61
Paillard et al. (2004) provided an excellent and complete overview of the bacterial diseases in 62
marine bivalves, including an updated reference of pathogenic Vibrio spp. They recognized that 63
Page 2 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
3
the emergence of vibrios as etiological agents in cultured bivalves is likely to increase over the 64
coming years due to ocean warming. The present study provides an updated overview of studies 65
performed on the diversity of Vibrio species associated with bivalve molluscs. Special emphasis 66
is made on the species that produce disease in the different bivalve mollusc’s life stages (larvae 67
vs. juveniles and adults) describing the characteristic signs and diseases caused by Vibrio spp. in 68
different hosts. 69
70
Diversity of Vibrio species associated with bivalves 71
Studies on the diversity of Vibrio species found that the predominating species associated with 72
bivalves from different geographical locations (Spain, Canada, Italy or Brazil) were either V. 73
splendidus, V. alginolyticus, V. harveyi or any combination of these species. Regarding this, 74
Pujalte et al. (1999) and Arias et al. (1999) found V. splendidus and V. harveyi as the prevailing 75
species in wild bivalves. Montilla et al. (1995) agreed in the dominance of V. splendidus, but 76
also recovered a high proportion of V. tubiashii strains, as did Castro et al. (2002) who also 77
recognized V. harveyi as an abundant species. However, other studies have found V. 78
alginolyticus as the dominant species (Hariharan et al., 1995; Caballo and Stabili, 2002; Lafisca 79
et al., 2008). Other species such as V. fluvialis, V. vulnificus and V. mimicus have also been 80
associated with mussels (Mytilus galloprovincialis) (Maugeri et al., 2000; Caballo and Stabili, 81
2002). In those mentioned studies, the prevalence of Vibrio species was established using 82
phenotypic identification methods which are known to underestimate the real diversity present 83
in bivalves. For instance, the high variability among the phenotypic characteristics within the V. 84
splendidus-like and the V. harveyi-like groups makes it virtually impossible to discriminate the 85
many species masked under these groups (Thompson et al., 2005; Le Roux and Austin, 2006; 86
Pascual et al., 2009). 87
Some of the above mentioned studies described the influence of environmental parameters (i.e. 88
salinity and water temperature) on the diversity of Vibrio species (Kaspar and Tamplin, 1993; 89
Motes et al., 1998; Arias et al., 1999; Pujalte et al., 1999). In wild bivalves (Ostrea edulis, M. 90
edulis, Chaemelea gallina, Donax trincili and D. semistriatus) from the Mediterranean Sea, V. 91
harveyi has been found to be dominant during the warmer months and V. splendidus during 92
winter and spring (Arias et al., 1999; Pujalte et al., 1999). Densities of V. vulnificus in shellfish 93
growing areas of the US Northern Gulf Coast were high and were not changing at temperatures 94
above 26°C and/or at salinity below 25 ppt, but were drastically reduced below this temperature 95
and/or above this salinity (Motes et al., 1998). Previous experiments in sterile microcosms 96
(Kaspar and Tamplin, 1993) demonstrated that salinity greater than 25 ppt reduced the survival 97
Page 3 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
4
of V. vulnificus in seawater. The presence of V. vulnificus in brackish water and mussels was 98
documented by Maugeri et al. (2000), but a low incidence of V. vulnificus was found in bivalves 99
from the western Mediterranean coast where the salinity values fluctuated between 30.7 and 100
38.3 ppt (Arias et al., 1999). These facts suggest that bivalves (oysters, clams, mussels, etc) 101
harvested from areas with high water temperature and low salinity may harbour high densities 102
of V. vulnificus, thereby increasing the risk of transmission to humans. The latter species 103
together with V. parahaemolyticus and V. cholerae are considered the most important human 104
pathogens and have produced important outbreaks associated with the consumption of 105
contaminated shellfish, mainly oysters, and therefore have been the subject of many studies (see 106
reviews by Morris, 2003; Cheng Su and Liu, 2007; Jones and Oliver, 2009). The public health 107
importance of these species will not be evaluated in this study. 108
The introduction of molecular techniques such as the amplified fragment length polymorphism 109
(AFLP) and multilocus sequence analysis (MLSA) have allowed a more precise identification of 110
Vibrio species which were previously masked under other taxa (Thompson et al., 2001; 2005; 111
Beaz-Hidalgo et al., 2008; Pascual et al., 2009). In this sense, phenotypically identified V. 112
harveyi strains were re-classified as V. campbellii by AFLP, DNA-DNA hybridization and 113
MLSA (Gómez-Gill et al., 2004; Thompson et al., 2007). Furthermore, molecular studies have 114
demonstrated the genetic diversity and the polyphyletic nature of V. splendidus (Thompson et 115
al., 2001; Le Roux et al., 2002; Thompson et al., 2005) and have enabled many new species to 116
be described, such as V. kanaloae, V. pomeroyi, V. chagasii and V. gallaecicus (Thompson et 117
al., 2003; Beaz-Hidalgo et al., 2009 c). In a previous study (Beaz-Hidalgo et al., 2008) we 118
analysed the diversity of Vibrio spp. in clams (Ruditapes decussatus and R. philippinarum) 119
cultured in Galicia (north-western coast of Spain) with phenotypic and genotypic methods 120
(AFLP). The latter method enabled us to identify 39% (57/145) of the isolates to the species 121
level. The predominant species that accounted for 66.6% of the total identified strains were V. 122
cyclitrophicus (17/57), V. splendidus (16/57) and V. alginolyticus (5/57), and we found only a 123
29.8% concordance between phenotypic and AFLP results (Beaz-Hidalgo et al., 2008). There 124
were major discrepancies for the V. splendidus-like strains, from which many were genetically 125
identified as V. crassostreae, V. cyclitrophicus, V. chagasii or could not be identified to species 126
level. The latter two species together with V. diabolicus and V. ichthyoenteri were associated for 127
the first time with cultured bivalves (Beaz-Hidalgo et al., 2008). Further studies using the 16S 128
rRNA gene and a MLSA with 4 genes (rpoD, recA, pyrH and atpA) enabled us to recognize 129
three new species within the family Vibrionaceae: Aliivibrio finisterrensis, Vibrio breoganii and 130
the already mentioned Vibrio gallaecicus (Beaz-Hidalgo et al., 2009 a, b, c). Pascual et al. 131
Page 4 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
5
(2009) investigated the usefulness of an MLSA approach with 6 protein genes (recA, pyrH, 132
rpoD, gyrB, rctB and toxR) to discriminate six species, V. harveyi, V. campbellii, V. 133
rotiferianus, V. parahaemolyticus, V. alginolyticus and V. natriegens that are tightly related 134
because they have DNA-DNA reassociation values very close to 70%. They recognized the 135
genes toxR and rpoD as the most reliable for species identification. Furthermore they proposed a 136
threshold value (> 90.3%) for species definition on the basis of the similarities of the 137
concatenated sequences of the 3 most resolving genes (toxR, rpoD and rctB). 138
139
Pathogenic Vibrio species in bivalve larvae 140
Larval development of bivalves includes a planktonic embryonic stage that evolves by 141
metamorphosis into a free-living veliger larva, first developing a velum to swim and during the 142
late stage developing a “foot” to become a pediveliger larva (settling stage). In a hatchery, 143
although all larval stages are vulnerable, during the temporary fixation of the larvae on the 144
bottom of the tank they are exposed to a high concentration of potential pathogenic bacteria 145
associated with the tank surface, moribund larvae or organic detritus (Sutton and Garrick, 1993). 146
Guillard (1959) was the first to report evidence of the involvement of a Vibrio sp. in the 147
disruption of the velum and internal tissues of the clam larvae Mercenaria mercenaria, which 148
produced a mortality of 70% of the population. In the study performed by Tubiash et al. (1965), 149
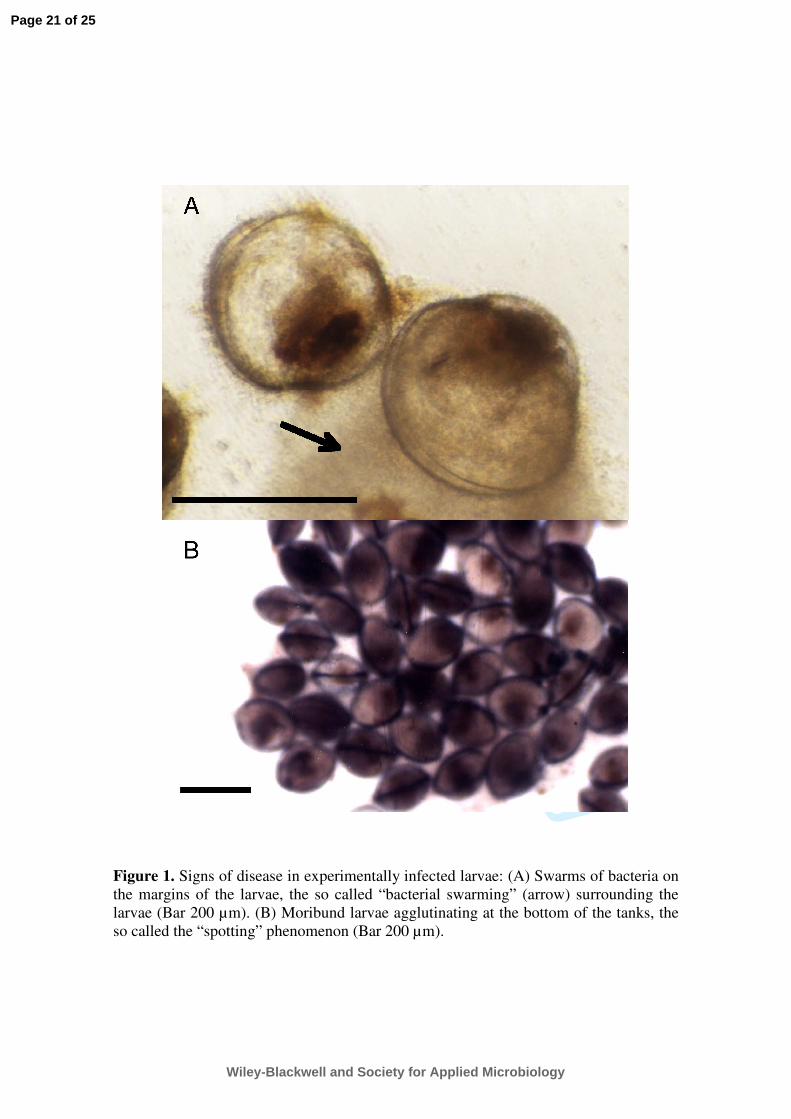
they observed “swarms” of bacteria appearing on the margins of the larvae (Fig. 1A), which 150
became progressively denser and after 8 hours, mortality occurred as a result of granular 151
necrosis. This study was the first one to describe the term “bacillary necrosis” affecting 152
numerous bivalves: Crassostrea virginica, O. edulis, M. mercenaria, Argopecten irradians and 153
Teredo navalis. Typical signs of “bacillary necrosis” included the extension of the velum, 154
motility reduction or erratic movements in circles that appeared after 4-5 hours of exposure to 155
Vibrio spp. The species V. alginolyticus, V. tubiashii and V. anguillarum were recognised as the 156
main causal agents of the “bacillary necrosis” in later studies (Tubiash et al., 1970; Tubiash and 157
Otto, 1986). Elston and Leibovitz (1980) described three patterns of bivalve larvae disease 158
produced by Vibrio spp. termed Pathogenesis I, II and III. Pathogenesis I affects all larval 159
stages, the larvae become sedentary showing signs of colonization of the mantle and invasion of 160
the visceral cavity. Pathogenesis II affects the early stage of the veliger larvae, producing velum 161
disruption and extension and abnormal swimming. The larvae remain active, showing visceral 162
atrophy before invasion into the organs of the digestive tract occurs. Pathogenesis III affects the 163
late veliger larval stage (pediveliger larva) and as in Pathogenesis I the larvae become 164
sedentary. However, in Pathogenesis III there is a progressive and extensive visceral atrophy 165
Page 5 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
6
and lesions in the organs of the digestive tract. Another characteristic sign of larval vibriosis in 166
hatcheries is the appearance of the phenomenon called “spotting” (Fig. 1B), defined as an 167
accumulation of moribund larvae agglutinated at the bottom of the tanks (Di Salvo et al., 1978). 168
All the studies mentioned so far have set up the typical signs produced by vibriosis in bivalve 169
larvae, which are still being used for the recognition and description of other pathogenic Vibrio 170
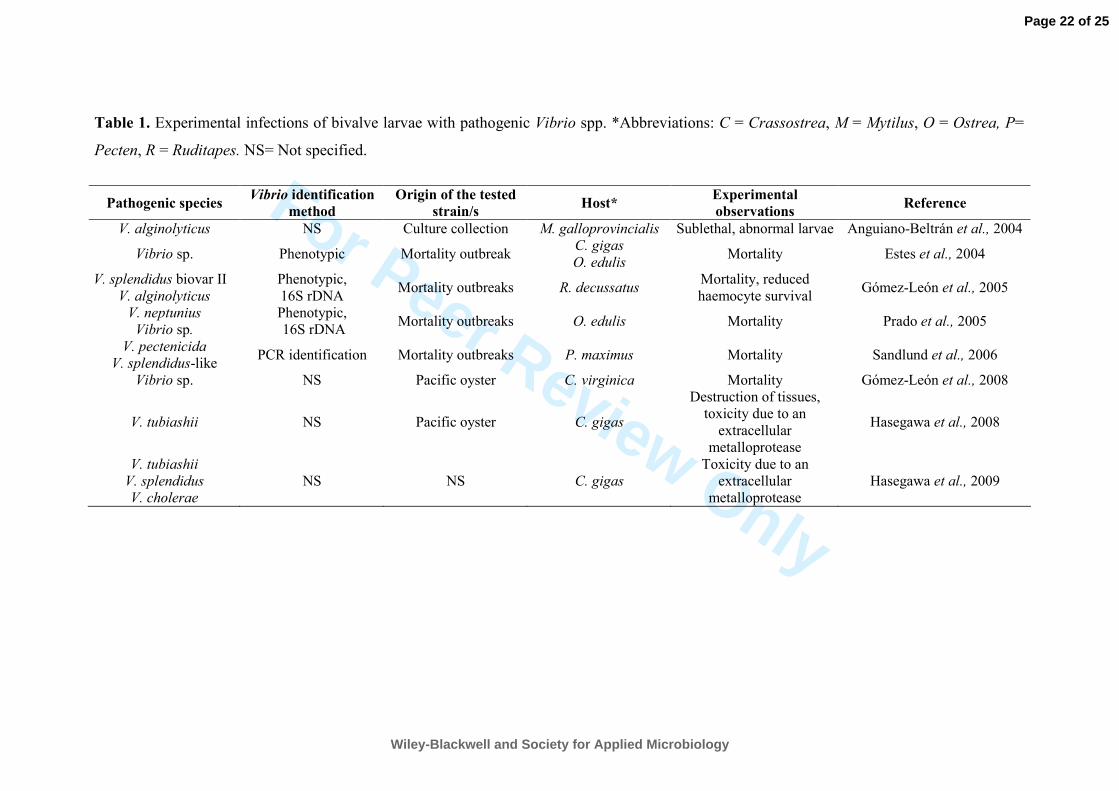
spp. Table 1 provides a summary of the most recent studies on bivalve larvae vibriosis, updating 171
the information presented in an earlier review (Paillard et al., 2004). As commented originally, 172
V. anguillarum, V. alginolyticus, V. tubiashii (Tubiash et al., 1970, Tubiash and Otto 1986) 173
together with V. splendidus (Sugumar et al., 1998) have been recognized as the main pathogenic 174
species associated to larval vibriosis. However, other species have been described in the last two 175
decades (Nicolas et al., 1996; Lambert et al., 1998; Prado et al., 2005). This is the case of V. 176
pectenicida (Lambert et al., 1998), originally described as a Vibrio sp. strain able to produce a 177
high mortality of the scallop larvae Pecten maximus after 48 hours of exposure due to the 178
release of bacteria toxins which interrupt the digestive transit and degrade the tissues of the 179
larvae (Nicolas et al., 1996). It is worth mentioning that V. pectenicida does not grow on the 180
thiosulfate-citrate-bile salts sucrose (TCBS) selective medium classically used for vibrios. A 181
number of studies have been conducted with this species to reproduce experimentally the 182
infection and to evaluate the pathogenesis and the interactions with haemocytes (cited and 183
discussed in the review by Paillard et al., 2004). 184
Prado et al. (2005) investigated 3 Vibrio strains isolated from oyster larvae (O. edulis) in three 185
different hatchery outbreaks in Galicia that had been identified with the 16S rRNA gene 186
sequences. One strain was identified as V. neptunius, and the other two classified as Vibrio sp., 187
because despite showing similarities to V. orientalis and V. vulnificus they could not be 188
unequivocally assigned to these species. Further studies showed that one of the latter Vibrio sp. 189
strains belonged to a new species, still unpublished, named V. ostreicida (S. Prado and J. L. 190
Romalde, personal communication). The experimental challenges on oyster larvae performed 191
with the three mentioned isolates caused a very high mortality (98-100%) in less than 48h. 192
Microscopical examination showed abnormal swimming behaviour, closed valves with internal 193
movement and velum abnormalities (Prado et al., 2005). Interestingly, this is the first study that 194
demonstrated the pathogenicity of V. neptunius in oyster larvae. The disease was compatible 195
with a “bacillary necrosis” and was associated with the Pathogenesis II described by Elston and 196
Leibovitz (1980). Other signs observed by Prado et al. (2005) included the previously described 197
“spotting” phenomenon (Di Salvo et al., 1978, Nottage and Birkbeck 1987) and “swarms” of 198
bacteria around the larvae (Tubiash et al., 1965). 199
Page 6 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
7
The species V. tubiashii reported as one of the causative agents of the “bacillary necrosis” 200
(Tubiash et al., 1970; Tubiash and Otto 1980) has recently been described as a re-emergent 201
pathogen in North America causing a decline of approximately 59% in larvae oyster production 202
(Elston et al., 2008). In addition, Hasegawa et al. (2008) demonstrated that one of the critical 203
factors in the pathogenicity of V. tubiashii in oysters (C. gigas) is the presence of a 204
metalloprotease (VtpA). These authors found that the gene encoding this protease is widespread 205
among Vibrio species and that the protein is regulated by quorum sensing (i.e. the gene is 206
activated at high cell population densities). The purified VtpA protein was found to be more 207
toxic for 6-day-old larvae than for 16-day-old larvae (Hasegawa et al., 2009). 208
It has been suggested that agents (pathogenic or opportunistic) responsible for the development 209
of disease in bivalve larvae seem to act synergistically and infection occurs as a result of 210
stressed larvae (Di Salvo et al., 1978; Paillard et al., 1994; Prado et al., 2005). 211
212
4. Pathogenic Vibrio species in juvenile and adult bivalves. 213
Tubiash et al. (1965) were also the first to perform research studies on pathogenic vibrios in 214
juvenile and adult bivalves. Their studies associated moribund adult bivalves (M. mercenaria, 215
C. virginica, M. edulis and Mya arenaria) with the species V. tubiashii and V. alginolyticus. 216
Elston and Leibovitz (1980) described further cases of vibriosis in oysters, detecting anomalies 217
in the shell and alterations in the function of the ligament and digestive processes. The classical 218
Vibrio infections are “summer mortality” in juvenile oysters and “brown ring disease” in adult 219
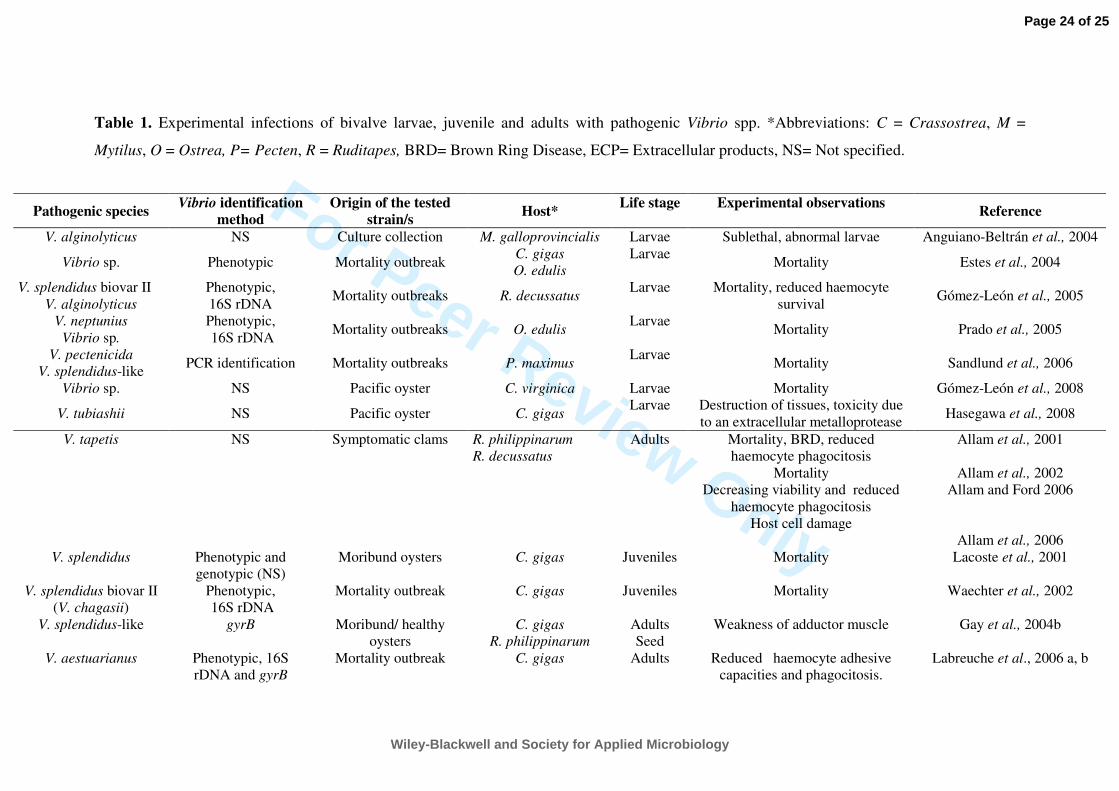
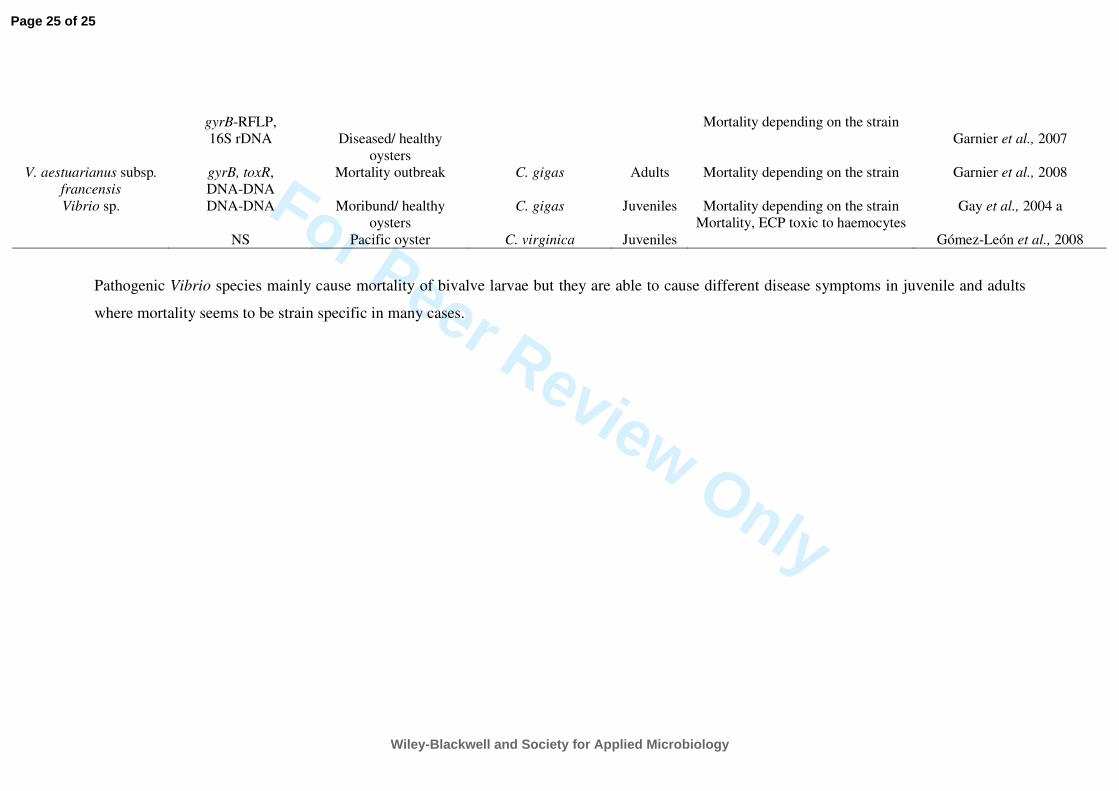
clams. Table 2 lists recent studies of Vibrio spp. that have caused pathologies in juvenile and 220
adult bivalves, completing the information provided by Paillard et al. (2004). 221
222
“Summer mortality” of juvenile oysters 223
“Summer mortality” affects juvenile populations of the Pacific oyster (C. gigas) during the 224
warmer months when the water temperature is ≥18ºC and reproduction takes place (Berthelin et 225
al., 2000). This phenomenon has been associated to stress situations, low dissolved oxygen or 226
presence of toxic substances in the sediment (Cheney et al., 2000). Lipp et al. (1976) were the 227
first to observe that the oyster haemolymph had high levels of vibrios that were causing death. 228
Later studies in France identified V. splendidus as the causal agent of “summer mortality” 229
(Lacoste et al., 2001; Waechter et al., 2002; Gay et al., 2004 a, b). Recently, Garnier et al. 230
(2007) analysed the bacterial populations in the haemolymph of moribund oysters by 231
phenotypic and molecular methods (RFLP of gyrB gene and partial sequences of the 16S rRNA 232
gene) and found that the prevalent species were, among others, V. aestuarianus (56%) and V. 233
Page 7 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
8
splendidus (25%). The pathogenicity of V. aestuarianus for the oyster was demonstrated by 234
Labreuche et al. (2006 b), that described the immunosuppressive activity of the extracellular 235
products of this species on the haemocyte functions. The susceptibility of bivalve microbiota to 236
the bactericidal activity of the haemolymph is one of the factors that regulate their persistence in 237
bivalve tissues (Canesi et al., 2002, Pruzzo et al., 2005). Bacterial surface ligands, soluble 238
haemolymph components and the ability of vibrios to activate distinct stress signalling pathways 239
involved in the haemocyte response are considered the factors that determine their presence 240
within a bivalve host (Pruzzo et al., 2005). 241
Garnier et al. (2008) characterized several strains of V. aestuarianus recovered from oyster (C. 242
gigas) and described them as V. aestuarianus subsps. francensis. They further demonstrated 243
(challenging studies on oysters) that 45% (5/11) of the strains of the new subspecies were highly 244
virulent producing a mortality above 50%, while 27% (3/11) of the strains caused moderate 245
mortality (15-50% mortality) and 27% were innocuous (< 15% mortality), considering virulence 246
to be a strain-specific character. More recently, Allain et al. (2009) suggested the possible role 247
of V. harveyi as aetiological agent of “summer mortality”, since it was detected in most samples 248
of affected oysters during the 2008 warm season, and was able to produce mortality in 249
experimental challenges. 250
The most accepted theory today is that “summer mortality” of the oyster (C. gigas) is not 251
attributed to only one bacterial pathogen but to a complex interaction between the physiological 252
and/or genetic state of the host, environmental factors and the presence of various opportunistic 253
infectious agents (Paillard et al., 2004; Pruzzo et al., 2005; Labreuche et al., 2006 b), among 254
which Vibrio spp. may play an important role. 255
256
Brown Ring Disease 257
Brown Ring Disease (BRD) caused by V. tapetis has been widely studied and the pathogenic 258
aetiology in adult clams (R. phillipinarum and R. decussatus) has been reviewed previously 259
(Paillard et al., 1994; Paillard, 2004). Bivalve’s susceptibility to V. tapetis infections is species 260
specific, causing greater physiological disturbances and mortality in R. philippinarum than in 261
other species of clam (R. decussatus and M. mercenaria) or in the oyster C. virginica (Allam et 262
al., 2001; 2006). This microorganism alters the clam’s haemocytes decreasing their viability and 263
phagocytic activity in experimental studies (Allam and Ford, 2006). The disease is characterized 264
by the alteration of the calcification process of the inner surface of the valves and the 265
appearance of a characteristic brown deposit consisting of conchiolin between the edge of the 266
shell and the pallial line (Paillard and Maes, 1990; Borrego et al., 1996). This brown deposit 267
Page 8 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
9
resembles that which appears in the Juvenile Oyster Disease (or Roseovarius Oyster Disease) 268
produced by the bacterial species Roseovarius crassostreae, although in this case the conchiolin 269
is deposited over the entire mantle and the attachment of the adductor muscle (Paillard et al., 270
2004; Boardman et al., 2008). 271
BRD is considered one of the main limiting factors when culturing Manila clams (R. 272
philippinarum) on many European coasts: France, Italy, Spain, Portugal, England, Ireland and 273
has been recently introduced in Norway (Paillard and Maes, 1990; Figueras et al., 1996; Allam 274
et al., 2000; Drummond et al., 2007; Paillard et al., 2009). The pathogen has also been recently 275
detected in Manila clams cultured in Korea (Park et al., 2006). It has been demonstrated that 276
clam sensitivity to V. tapetis varies according to the geographical origin of the clams. 277
Populations of French Manila clams have been described as more susceptible to BRD than 278
populations in the United States (Allam et al., 2001) as are Manila clams grown in Galicia 279
compared to those of Ireland (Drummond et al., 2007). Environmental factors (i.e. temperature 280
and salinity) play a role in the development of BRD, which tends to be more frequent in the 281
spring and winter as the optimum growth temperature for V. tapetis is 15ºC (Paillard et al., 282
1994; 2004). Reid et al. (2003) demonstrated when challenging BRD infections on Manila 283
clams, that the disease was more severe when performed at 20 ppt salinity than at 40 ppt. The 284
diagnostic of BRD is currently based on the examination of the characteristic brown ring on the 285
inner edge of the shell. For an early diagnosis in the absence of the brown ring, specific PCR 286
detection protocols targeting the 16S rRNA gene of V. tapetis have been designed (Paillard et 287
al., 2006; Park et al, 2006; Romalde et al., 2007). Biochemical, serological and genetic 288
intraspecific variability within V. tapetis had been described as well as three main subgroups 289
that correlate with the host type (Rodríguez et al., 2006). This heterogeneity has been further 290
demonstrated with a MLSA and proteomic analysis by 2D-PAGE (Balboa et al., 2007, 2008). 291
292
Control and preventive measures 293
Control of diseases is a difficult task in the culture of bivalve molluscs, both at the hatcheries 294
and at adult stages, the maintenance of optimum water quality and culture density being 295
important for the prevention of mortality outbreaks in hatcheries (Prado et al., 2009). Once the 296
bivalves begin to die, chemotherapy is the only possible approach, but it has been shown to be 297
inefficient and to have disadvantages, like the development of antibiotic resistances 298
(Karunasagar et al., 1994; Kemper, 2008). On the other hand, vaccines, an alternative useful in 299
fish cultures, are not viable in bivalves (Bachère, 2003). Furthermore none of these methods can 300
be used when bivalve are in the growing beds in the natural environment. 301
Page 9 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
10
One promising approach is the use of probiotics defined in this context as ‘those live microbial 302
adjuncts which have a beneficial effect on the host’ (Riquelme et al., 2000; Verschuere et al., 303
2000; Prado et al., 2009). The use of live bacteria with activity against pathogens is especially 304
interesting in bivalve hatcheries, because the larvae are released and exposed to environmental 305
microbiota in early ontogenic stages. The filter feeding behaviour of these animals increases the 306
influence of the surrounding water. In the last years a number of strains have been proposed as 307
good candidates to be used as probiotics in aquaculture systems. They are usually marine 308
bacteria with a wide spectrum of antibacterial activity, also effective against the main 309
pathogenic Vibrio spp., being able to develop their inhibitory activity in seawater. Their 310
biotechnological potential is based on the ability to control the proliferation of opportunistic 311
pathogens in a hatchery and farm environments. 312
Other research area of increasing interest is the selective breeding of molluscs to obtain genetic 313
families resistant to diseases (Tanguy et al., 2008). The main advantage of breeding selection is 314
that it offers long-term increased resistance, from hatchery through all live stages up to the 315
commercial size. This approach has been developed against parasitic diseases, like QX disease 316
(specific to Sydney Rock oysters caused by the protozoan parasite Marteilia sydneyi) and 317
bonamiasis, where hemocytes of flat oysters are affected (Bezemer et al., 2006; da Silva et al., 318
2008; Simonian et al., 2009). However, in the last years some attemps have been made to get 319
families of bivalves, mainly Pacific oyster (C. virginica), resistant to bacterial pathogens 320
(Gómez-León et al., 2008) with promising results. Thus, some oyster families have been 321
selected with increased resistance, not only to protistan, but also to bacterial pathogens 322
including Roseovarius crassostreae and several Vibrio species. 323
Other promising research line which has been scarcely studied is the genomic characterization 324
through the analysis of EST (Expressed Sequence Tags) collections, which are short sub-325
sequences of a transcribed cDNA sequences that represent the genes expressed in an organism 326
under a given condition. Using this strategy host defence genes have been identified in 327
numerous bivalves including Argopecten irradians, Chlamys farreri, C. gigas, C. virginica, M. 328
edulis or R. decussatus, which may help to understand the host-parasitic relationship and the 329
molecular mechanisms by which these marine bivalves respond to pathogens (Song et al., 2006, 330
Gestal et al., 2007, Guegen et al., 2007, Wang et al., 2009; Fleury et al., 2009 and references 331
therein). In this sense, the expression of several genes implicated in the resistance and 332
susceptibility of C. gigas to the “summer mortality” have been recognized and compiled in a 333
publicly-available database (Fleury et al., 2009 and references therein). Further investigation in 334
Page 10 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
11
this area will help to elucidate of the physiological and genetic basis for the survival of selected 335
progeny. 336
337
Conclusions and future prospects 338
The present study overviews the diversity of Vibrio species associated with bivalve molluscs. 339
Ongoing studies on the disease and pathogenicity of bivalves primarily relies on the use of 340
phenotypic and molecular methods for an exact species identification following challenging in 341
vivo and in vitro tests. However, studies that establish the molecular mechanisms involved in the 342
development of bivalve disease, such as the detection of virulent genes, are scarce and therefore 343
needed in order to better understand the host-pathogen interactions. Major advances are being 344
made in the study of toxicity of bacterial extracellular products to bivalves as well as in the 345
study of the cellular mechanisms in relation to bacteria–hemocyte interactions. These new 346
insights will explain how a bacterial pathogen is able to avoid phagocytic activity and multiply 347
to high levels inside the bivalve, leading to death. A complete description of how the 348
environment influences these bacteria-host interactions and health status of the bivalve may lead 349
towards a better understanding of the infection routes of the pathogen and mode of action in the 350
natural conditions. 351
It remains to be investigated to what extent some of the recently discovered species are 352
commensal, opportunistic or pathogenic organisms. Knowledge of the infection mechanisms 353
used by classical and emerging Vibrio spp. to develop disease in bivalve molluscs will help to 354
establish adequate preventive measures to control the transmission of these pathogens in 355
hatcheries and in coastal growing areas. Furhermore, research on the selection of genetic 356
resistant bivalve populations and on the potential use of probiotic bacteria may be greatly useful 357
for the improvement of cultured bivalve production. 358
359
360
361
362
363
364
365
366
367
368
Page 11 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
12
References 369
Allain, G., Arzul, I., Chollet, B., Cobret, L., de Decker, S., Faury, N., Ferrand, S, et al. (2009). 370
Summer mortality outbreaks of French Pacific oysters, Crassostrea gigas in 2008: research 371
and detection of pathogens. 14th EAFP International Conference. Diseases of Fish and 372
Shellfish. Prague (Czech Republic). pp. 127. 373
Allam, B., Ashton-Alcox, A., and Ford, S.E. (2001). Hemocyte activities associated with 374
resistance to brown ring disease in Ruditapes spp. clams. Dev Comp Immunol 25: 365-375. 375
Allam, B., and Ford, S.E. (2006). Effects of the pathogenic Vibrio tapetis on defence factors of 376
susceptible and non-susceptible bivalve species: I. Haemocyte changes following in vitro 377
challenge. Fish Shellfish Immunol 20: 374-383. 378
Allam, B., Paillard, C., Auffre, M., and Ford, S.E. (2006). Effects of the pathogenic Vibrio 379
tapetis on defence factors of susceptible and non-susceptible bivalves species: II. Cellular and 380
biochemical changes following in vivo challenges. Fish Shellfish Immunol 20: 384-397. 381
Allam, B., Paillard, C., and Ford, S.E. (2002). Pathogenicity of Vibrio tapetis, the etiological 382
agent of brown ring disease in clams. Dis Aquat Organ 48: 221-231. 383
Allam, B., Paillard C., Howard A., and Le Pennec M. (2000). Isolation of the pathogen Vibrio 384
tapetis and defence parameters in brown ring diseased Manila clams Ruditapes philippinarum 385
cultivated in England. Dis Aquat Org 41:105–113. 386
Anguiano-Beltrán, C., Lizárraga-Partida M.L., and Searcy-Bernal R. (2004). Effect of Vibrio 387
alginolyticus on larval survival of the blue mussel Mytilus galloprovincialis. Dis Aquat Org 388
59: 119-123. 389
Arias, C.R., Macián, M.C., Aznar, R., Garay, E., and Pujalte, M.J. (1999). Low incidence of 390
Vibrio vulnificus among Vibrio isolates from sea water and shellfish of the western 391
Mediterranean coast. J Appl Microbiol 86: 125-134. 392
Bachère, E. (2003) Anti-infectious immune effectors in marine invertebrates: potential tools for 393
disease control in larviculture. Aquaculture 227: 427-438. 394
Balboa, S., Bermúdez-Crespo, J., Gianzo, C., Doce, A., López, J.L., and Romalde, J.L. (2007). 395
Proteomic analysis of Vibrio tapetis, the etiological agent of Brown Ring Disease. Vibrio 396
Congress 2007. 397
Balboa, S., Doce, A., Diéguez, A.L., and Romalde, J.L. (2008) Estudio de la variabilidad 398
intraespecífica del patógeno de almeja Vibrio tapetis mediante MLSA. VII Reunión del grupo 399
de Microbiología del Medio Acuático (S.E.M.). 400
Page 12 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
13
Beaz-Hidalgo, R., Cleenwerck, I., Balboa, S., De Watcher, M., Thompson, F.L., Swings, J., De 401
Vos, P., and Romalde, J.L. (2008). Diversity of vibrios with reared clams in Galicia (NW 402
Spain). Syst Appl Microbiol 31: 215-222. 403
Beaz-Hidalgo R., Cleenwerck I., Balboa S., Prado S., de Vos P., and Romalde J.L. (2009a). 404
Vibrio breoganii sp. nov., a non motile alginolytic marine bacterium within the Vibrio 405
halioticoli clade. Int J Syst Evol Microbiol 59: 1589-1594. 406
Beaz-Hidalgo R., Doce A., Balboa S., Barja J.L., and Romalde J.L. (2009b) Aliivibrio 407
finisterrae sp. nov., isolated from Manila clams, Ruditapes philippinarum. Int J Syst Evol 408
Microbiol (in press). 409
Beaz-Hidalgo R., Doce A., Pascual J., Toranzo A.E., and Romalde J.L. (2009c). Vibrio 410
gallaecicus sp. nov., isolated from cultured clams in north-western Spain. Syst Appl Microbiol 411
32: 111-117. 412
Berthelin, C., Kellner, K., and Mathieu, M. (2000). Storage metabolism in the Pacific oyster 413
(Crassostrea gigas) in relation to summer mortalities and reproductive cycle (west coast of 414
France). Comp Biochem Physiol B Biochem Mol Bio 125: 359-169. 415
Bezemer, B., Butt, D., Nell, J., Adlard, R., and Raftos, D. (2006). Breeding for QX disease 416
resistance negatively selects one form of the defensive enzyme, phenoloxidase, in Sidney rock 417
oysters. Fish Shellfish immunol. 20: 627-636. 418
Boardman, C.L., Maloy, A., and Boettcher, K. (2008). Localization of the bacterial agent 419
juvenile oyster disease (Roseovarius crassostreae) within affected eastern oysters 420
(Crassostrea virginica). J Invert Pathol 97: 150-158. 421
Borrego, J.J., Castro, D., Luque, A., Paillard, C., Maes, P., García, M., and Ventosa, A. (1996). 422
Vibrio tapetis sp. nov., the causative agent of the brown ring disease affecting cultured clams. 423
Int J Syst Bacteriol 46: 480-484. 424
Canesi, L., Gallo, G., Gavioli, M., and Pruzzo, C. (2002). Bacteria-hemocyte interactions and 425
phagocytosis in marine bivalves. Micros Res Tech 57: 469-476. 426
Castro, D., Pujalte, M.J., Lopez-Cortes, L. Garay, E., and Borrego, J.J. (2002). Vibrios isolated 427
from the cultured manila clam (Ruditapes philippinarum): numerical taxonomy and 428
antibacterial activities. J Appl Microbiol 93: 438-447. 429
Caballo, R.A., and Stabili, L. (2002). Presence of vibrios in seawater and Mytillus 430
galloprovincialis (Lam.) from the Mar Piccolo of Taranto (Ionian Sea). Water Res 36: 3719-431
3729. 432
Cheng Su, Y., and Liu, C. (2007). Vibrio parahaemolyticus: A concern of seafood safety. Food 433
Microbiol 24: 549-558. 434
Page 13 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
14
Cheney, D.P., McDonald, B.F., and Elston, R.A. (2000). Summer mortality of Pacific oysters, 435
Crassostrea gigas (Thunberg): initial findings on multiple environmental stressors in Puget 436
Sound, Washington, 1998. J Shellfish Res 19: 353-380. 437
da Silva, P.M., Comesaña, P., Fuentes, J., and Villalba, A. (2008). Variability of haemocyte and 438
haemolymph parameters in European flat oyster Ostrea edulis families obtained from brood 439
stocks of different geographical origins and relation with infection by the protozoan Bonamia 440
ostreae. Fish Shellfish Immunol 24: 551-563. 441
Di Salvo, L.H., Blecka J., and Zebal R. (1978). Vibrio anguillarum and larval mortality in a 442
California coastal shellfish hatchery. Appl Environ Microbiol 35: 219-221. 443
Drummond, L.C., Balboa, S., Beaz, R., Mulcahy, M.F, Barja, J.L., Culloty, S.C., and Romalde 444
J.L. (2007). The susceptibility of Irish-grown and Galician-grown Manila clams, Ruditapes 445
philippinarum, to Vibrio tapetis and brown ring disease. J Invertebr Pathol 95:1–8. 446
Elston, R.A., Hasegawa, H., Humphrey, K.L., Polyak, I.K., and Häse, C.C. (2008). Re-447
emergence of Vibrio tubiashii in bivalve shellfish aquaculture: severity, environmental 448
drivers, geographic extent and management. Dis Aquat. Organ 82: 119-134. 449
Elston, R., and Leibovitz L. (1980). Pathogenesis of experimental vibriosis in larval American 450
oysters, Crassostrea virginica. Can J Fish Aquat Sci 37: 964-978. 451
Estes, R.M., Friedman C.S., Elston R.A., and Herwig R.P. (2004). Pathogenicity testing of 452
shellfish hatchery bacterial isolates on Pacific oyster Crassostrea gigas larvae. Dis Aquat Org 453
58: 223-230. 454
Figueras, A., Robledo, J.A.F., and Novoa, B. (1996). Brown ring disease and parasites in clams 455
(Ruditapes decussatus and Ruditapes philippinarum) from Spain and Portugal. J Shellfish Res 456
15: 363-368. 457
Fleury, E., Huvet, A., Lelong, C., de Lorgeril, J., Boulo, V., Gueguen, Y. et al. (2009). 458
Generation and analysis of a 29,745 unique Expressed Sequence Tags from the Pacific oyster 459
(Crassostrea gigas) assembled into a publicly accessible database: the GigasDatabase. BMC 460
Genomics 10:341: doi:10.1186/1471-2164-10-341. 461
Garnier, M., Labreuche, Y., García, C., Robert, M., and Nicolas, J.L. (2007). Evidence for the 462
involvement of pathogenic bacteria in summer mortalities of the Pacific oyster Crassostrea 463
gigas. Microb Ecol 53: 187-196. 464
Garnier, M., Laubreche, Y., and Nicolas, J.L. (2008). Molecular and phenotypic 465
characterization of Vibrio aestuarianus subsp. francensis subsp. nov., a pathogen of the oyster 466
Crassostrea gigas. Syst Appl Microbiol 31: 358-365. 467
Page 14 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
15
Gay, M., Berthe F.C.J., and Le Roux F. (2004a). Screening of Vibrio isolates to develop an 468
experimental infection model in the Pacific oyster Crassostrea gigas. Dis Aquat Org 59: 49-469
56. 470
Gay, M., Renault, T., Pons, A.M., and Le Roux, F. (2004b). Two Vibrio splendidus related 471
strains collaborate to kill Crassostrea gigas: taxonomy and host alterations. Dis Aquat Org 472
62: 65-74. 473
Gestal, C., Costa M., Figueras, A., and Novoa, B. (2007). Analysis of differentially expressed 474
genes in response to bacterial stimulation in hemocytes of the carpt shell clam Ruditapes 475
decussatus: Identification of new antimicrobial peptides. Marine Genomics 406: 134-143. 476
Gómez-Gill, B., Soto-Rodríguez, S., García-Gasca, A., Roque, A., Vázquez-Juárez, A., and 477
Swings, J. (2004). Molecular identification of V. harveyi-related isolates associated with 478
diseased aquatic organisms. Microbiology 150: 1769-1777. 479
Gómez-León, J., Villamil, L., Lemos, M.L., and Novoa, B. (2005). Isolation of Vibrio 480
alginolyticus and Vibrio splendidus from aquacultured carpet shell clam (Ruditapes 481
decussatus) larvae associated with mass mortalities. Appl Environ Microbiol 71: 98-104. 482
Gómez- León, J., Villamil, L., Salger, S.A., Sallum, R.H., Remacha-Triviño, A., Leavitt, D.F., 483
and Gómez-Chiarri, M. (2008). Survival of eastern oysters Crassostrea virginica from three 484
lines following experimental challenge with bacterial pathogens. Dis Aquat Org 79: 95-105. 485
Guegen, Y., Cadoret, J.P., Flament, D., Roumiguière, C.B., Girardot, A.L., Garnier, J. et al. 486
(2003). Immune gene discovery by expressed sequence tags generated from hemocytes of the 487
bacteria-challenged oyster, Crassostrea gigas. Gene 303: 139-145. 488
Guillard, R.R.L. (1959). Further evidence of the destruction of bivalve larvae by bacteria. Biol 489
Bull 117: 258-266, 1959. 490
Guisande, J.A., Montes, M., Farto, R., Armada, S.P., Pérez, M.J., and Nieto T.P. (2004). A set 491
of tests for the phenotypic identification of culturable bacteria associated with Galician 492
bivalve mollusk production. J Shellfish Res 23: 599-610. 493
Hariharan, H., Giles, J.S., Heaney, S.B., Arsenault, G., Mcnair, N., and Rainnie, D.J. (1995). 494
Bacteriological studies on mussels and oysters from six river systems in Prince Edward Island, 495
Canada. J Shellfish Res 14: 527-532. 496
Hasegawa, H., Lind, E.J., Boin, M.A., and Häse, C. (2008). The extracelular metalloprotease of 497
Vibrio tubiashii is a major virulence factor for pacific oyster (Crassostrea gigas) larvae. Appl 498
Environ Microbiol 74: 4101-4110. 499
Page 15 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
16
Hasegawa, H., Gharaibeh, D.N., Lind, E.J., and Häse, C.C. (2009). Virulence of 500
metalloproteases produced by Vibrio species on Pacific oyster Crassostrea gigas larvae. Dis 501
Aquat Org 85: 123-131. 502
Jones, M.K., and Oliver, J.D. (2009). Vibrio vulnificus: Disease and Pathogenesis. Infect Immun 503
77: 1723-1733. 504
Karunasagar, I., Pai, R., Malathi, G.R., and Karunasagar, I. (1994). Mass mortality of Penaeus 505
monodon larvae due to antibiotic resustat Vibrio harveyi infection. Aquaculture 128: 203-209. 506
Kaspar, C.W., and Tamplin, M.L. (1993). Effects of temperature and salinity on the survival of 507
Vibrio vulnificus in seawater and shellfish. Appl Environ Microbiol 59: 2425-2429. 508
Kemper, N. (2008) Veterinary antibiotics in the aquatic and terrestrial environment. Ecol Indic 509
8: 1-13. 510
Kueh, C.S.W., and Chan, K.Y. (1985). Bacteria in bivalve shellfish with special reference to the 511
oyster. J Appl Bacteriol 59: 41-47. 512
Labreuche, Y., Lambert, C., Soudant, P., Boulo, V., Huvet, A., and Nicolas, J.L. (2006a). 513
Cellular and molecular hemocyte responses of the Pacific oyster, Crassostrea gigas, following 514
bacterial infection with Vibrio aestuarianus strain 01/32. Microbes Infect 8: 2715-2724. 515
Labreuche Y., Soudant P., Goncalves M, Lambert C., and Nicolas J.L. (2006b). Effects of 516
extracellular products from the pathogenic Vibrio aestuarianus strain 01/32 on lethality and 517
cellular immune responses of the oyster Crassostrea gigas, Dev Comp Immunol 30: 367–379. 518
Lacoste, A., Jalabert, F., Malham, S., Cueff, A., Gelebart, F., Cordevant, C., et al. (2001). A 519
Vibrio splendidus strain is associated with summer mortality of juvenile oysters Crassostrea 520
gigas in the Bay of Morlaix (North Brittany, France). Dis Aquat Org 46: 139-145. 521
Lafisca, A., Pereira C.S., Giaccone, V., and Rodriguez, D.D.P. (2008). Enzymatic 522
characterization of Vibrio alginolyticus strains isolated from bivalves harvested at Venice 523
lagoon (Italy) and Guanabara Bay (Brazil). Rev Inst Med trop S Paulo 50: 199-202. 524
Lambert C., Nicolas J.L., Cilia V., and Corre S. (1998). Vibrio pectenicida sp. nov., a pathogen 525
of scallop (Pecten maximus) larvae. Int J Syst Bacteriol 48: 481-487. 526
Lee, K.K., Ping-Chung, L., and Huang, C.Y. (2003). Vibrio parahaemolyticus infections for 527
both humans and edible mollusc abalone. Microb Infect 5: 481-485. 528
Le Roux, F., and Austin, B. (2006). Vibrio splendidus. In The Biology of Vibrios. Thompson F. 529
L., Austin, B., and Swings, J. (eds). Washington: ASM Press edition, pp. 285-296. 530
Le Roux, F., Gay, M., Lambert, C., Waechter, M., Poubalanne, S., Chollet, B., Nicolas, J.L., 531
and Berthe, F.C. (2002). Comparative analysis of Vibrio splendidus related strains isolated 532
during Crassostrea gigas mortality events. Aquat Liv Resour 15: 251-258. 533
Page 16 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
17
Lipp, P.R., Brown B., Liston J., and Chew K. (1976). Recent findings on the summer diseases 534
of Pacific oysters. Proc Nat Shellfish Assoc 65: 9-10. 535
Liu, P.C., Chen, C.Y., Huang, C.Y., and Lee, K.K. (2000). Virulence of Vibrio 536
parahaemolyticus isolated from cultured small abalone, Haliotis diversicolor supertexta, with 537
withering syndrome. Lett Appl Microbiol 31: 433-437. 538
Liu, P.C. , Chen, Y.C., and Lee, K.K. (2001). Pathogenicity of Vibrio alginolyticus isolated 539
from diseased small abalone Haliotis diversicolor supertexta. Microbios 104: 71-77. 540
Maugeri, T.L., Caccamo, D., and Gugliandolo, C. (2000). Potentially pathogenic vibrios in 541
brackish waters and mussels. J Appl Microbiol 89: 261-266. 542
Morris, J.G. (2003). Cholera and other types of Vibriosis: A story of human pandemics and 543
oysters on the half shell. Food Safety 37: 272-280. 544
Motes, M.L., De Paola, A., Cook, D.W., Veazey, J.E., Hunsucker, J.C., Garthright, W.E., et al. 545
(1998). Influence of water temperature and salinity on Vibrio vulnificus in Northen Gulf and 546
Atlantic Coast Oysters (Crassostrea virginica). Appl Env Microbiol 64: 1459-1465. 547
Montilla, R., Palomar, J., Santmarti, M., Fuste, C., and Viñas, M. (1994). Isolation and 548
characterization of halophilic Vibrio from bivalves bred in nurseries at the Ebre Delta. J Invert 549
Pathol 63: 178-181. 550
Murchelano, R.A., and Brown, C. (1970). Heterotrophic bacteria in Long Island Sound. Mar 551
Bio 7: 1-6. 552
Nicolas L., Corre S., Gauthier G., Robert R., and Ansquer D. (1996). Bacterial problems 553
associated with scallop Pecten maximus larval culture. Dis Aquat Org 27: 67-76. 554
Nottage, A. S., and Birkbeck T.H. (1987). The role of toxins in Vibrio infections of bivalve 555
molluscs. Aquaculture 67: 244-246. 556
Paillard, C. (2004) A short-review of brown ring disease, a vibriosis affecting clams, Ruditapes 557
philipinarum and Ruditapes decussatus. Aquat Living Resour 17: 467-475. 558
Paillard, C., Gausson, S., Nicolas, J.L., Le Pennec, J.P., and Haras, D. (2006). Molecular 559
identification of Vibrio tapetis, the causative agent of the BRD of Ruditapes philippinarum. 560
Aquaculture 253: 25-38. 561
Paillard, C. and Maes, P. (1990). Étiologie de la maladie de l’anneau brun chez Tapes 562
philippinarum: Pathogenicité d’un Vibrio sp. CR Acad Sci Paris 310: 15–20. 563
Paillard, C., Maes, P., and Oubella, R. (1994). Brown ring disease in clams. Ann Rev Fish Dis 4: 564
1–22. 565
Paillard, C., Le Roux F., and Borrego, J.J. (2004). Bacterial disease in marine bivalves, a 566
review of recent studies: Trends and evolution. Aquat Living Resour 17: 477-498. 567
Page 17 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
18
Paillard, C., Korsnes, K., Le Chevalier, P., Le Boulay, C., Harkestad, L., Eriksen, A.G., et al., 568
(2009). Vibrio tapetis-like strain isolated from introduced Manila clams Ruditapes 569
philippinarum showing symptoms of brown ring disease in Norway. Dis Aquat Organisms 81: 570
153-161. 571
Park, K.I, Paillard, C., Le Chevalier, P., and Choi, K.S. (2006) Report on the occurrence of 572
brown ring disease (BRD) in Manila clam, Ruditapes philippinarum, on the west coast of 573
Korea. Aquaculture 200: 610-613. 574
Pascual, J., Macián, M.C., Arahal, D.R., Garay, E., and Pujalte, M.J. (2009). Multilocus 575
Sequence Analysis of the central clade of the genus Vibrio using the 16S rRNA, recA, pyrH, 576
rpoD, gyrB, rctB and toxR genes. Int J Syst Evol Microbiol (in press) 577
(doi:10.1099/ijs.0.010702-0). 578
Prado, S., Romalde, J.L., Montes, J., and Barja J.L. (2005). Pathogenic bacteria isolated from 579
disease outbreaks in shellfish hatcheries. First description of Vibrio neptunius as an oyster 580
pathogen. Dis Aquat Org 67: 209-215. 581
Prado, S., Montes, J., Romalde, J.L., and Barja, J.L. (2009). Inhibitory activity of Phaeobacter 582
strains against aquaculture pathogenic bacteria. Int Microbiol 12: 107-114. 583
Prieur, D., Mével, G., Nicolas, J.L., Plusquellec, A., and Vigneulle, M. (1990). Aerobic and 584
facultative anaerobic heterotrophic bacteria associated to Mediterranean oysters and seawater. 585
Int Microbiol 2: 259-266. 586
Pruzzo, C., Gallo, G., and Canesi, L. (2005). Persistence of vibrios in marine bivalves: the role 587
of interactions with haemolymph components. Environ Microbiol 6: 761-772. 588
Pujalte, M.J., Ortigosa, M., Macián, M.C., and Garay, E. (1999). Aerobic and facultative 589
anaerobic heterotrophic bacteria associated to Mediterranean oysters and seawater. Int 590
Microbiol 2: 259-266. 591
Reid, H.I., Soudant, P., Lambert, C., Paillard, C., and Birkbeck, T.H. (2003). Salinity effects on 592
immune parameters of Ruditapes philippinarum challenged with Vibrio tapetis. Dis Aquat 593
Org 56: 249–258. 594
Riquelme, C., Araya, R., and Escribano, R. (2000). Selective incorporation of bacteria by 595
Argopecten purpuratus larvae: implications for the use of probiotics in culturing systems of the 596
Chilean scallop. Aquaculture 181: 25-36. 597
Rodríguez, J.M., López-Romalde, S., Beaz, R., Alonso, C., Castro, D., and Romalde, J.L. 598
(2006). Molecular fingerprinting of Vibrio tapetis strains using three PCR-based methods: 599
ERIC-PCR, REP-PCR and RAPD. Dis Aquat Org 69: 175-183. 600
Page 18 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
19
Romalde, J.L., Rodríguez, J.M., and Borrego, J.J. (2007). Protocolo de PCR (reacción en 601
cadena de la polimerasa) para detección e identificación de Vibrio tapetis a partir de cultivos 602
puros del microorganismo y de tejidos de moluscos. Patent ES 2 265 707. Oficina Española de 603
Patentes y Marcas. 604
Sandlund, N., Torkildsen, L., Maqnesen, T., Mortesen, S., and Bergh, O. (2006). 605
Immunohistochemistry of great scallop Pecten maximus larvae experimentally challenged 606
with pathogenic bacteria. Dis Aquat Org 69: 163-173. 607
Simonian, M., Nair, S.V., Nell, J.A., and Raftos, D.A. (2009). Proteomic clues to the 608
identification of QX disease-resistance biomarkers in selectively bred Sydney rock oysters, 609
Saccostrea glomerata. J Proteomics 73: 209-217. 610
Song, L., Xu, W., Li, C., Li, H., Wu, L., Xiang, J., and Guo, X. (2006). Development of 611
expressed sequence tags from the bay scallop, Argopecten irradians irradians. Mar Biotech 8: 612
161-169. 613
Sutton, D.C, and Garrik R. (1993). Bacterial disease of cultured giant clam Tridacna gigas 614
larvae. Dis Aquat Org 16: 47-53. 615
Tanguy, A., Bierne, N., Saavedra, C., Pina, B., Bachérre E., Kube, M., et al., (2008). Increasing 616
genomic information in bivalves through new EST collections in four species: Development 617
of new genetic markers for environmental studies and genome evolution. Gene 408: 27-36. 618
Thompson F.L., Gevers D., Thompson C.C, Dawyndt P., Naser S., Hoste B., et al. (2005). 619
Phylogeny and molecular identification of vibrios on the basis of multilocus sequence 620
analysis. Appl Environ Microbiol 71: 5107-5115. 621
Thompson, F.L., Gómez-Gill, B., Ribeiro Vasconcelos, A.T., and Sawabe, T. (2007). 622
Multilocus sequence analysis reveals that Vibrio harveyi and V. campbelli are distinct species. 623
Appl Environ Microbiol 73: 4279-4285. 624
Thompson, F.L., Hoste B., Vandeneulebroecke K., and Swings J. (2001). Genomic diversity 625
amongst Vibrio isolates from different source determined by fluorescent amplified fragment 626
length polymorphism. Syst Appl Microbiol 24: 520-538. 627
Thompson, F.L., Thompson, C.C., Li, Y., Gómez-Gil, B., Vanderberghe, J., Hoste, B., and 628
Swings, J. (2003). Vibrio kanaloae sp. nov., Vibrio pomeroyi sp. nov. and Vibrio chagasii sp. 629
nov., from sea water and marine animals. Int J Syst Evol Microbiol 53: 753-759. 630
Tubiash, H.S., Chanley, P.E., and Leifson, E. (1965). Bacillary necrosis, a disease of larval and 631
juvenile bivalve mollusks. I. Etiology and epizootiology. J Bacteriol 90: 1036-1044. 632
Tubiash H.S., Colwell R.R., and Sakazaki, R. (1970). Marine vibrios associated with bacillary 633
necrosis, a disease of larval and juvenile bivalve molluscs. J Bacteriol 103: 272-273. 634
Page 19 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review O
nly
20
Tubiash, H.S., and Otto, S.V. (1986). Bacterial problems in oysters. A review. In Pathology in 635
Marine Aquaculture. Vivarès C.P., Bonami J.R., and Jasper E. (eds) Bredene (Belgium): 636
European Aquaculture Society, Spec. Publ. 9, pp. 233-242. 637
Verschuere L., Rombaut G., Sorgeloos P., and Verstraete W. (2000) Probiotic bacteria as 638
biological control agents in aquaculture. Microbiol Mol Biol Rev 64: 655-671. 639
Waechter M., Le Roux F., Nicolas J.L., Marissal E., and Berthe F. (2002). Characterization of 640
pathogenic bacteria of the cupped oyster Crassostrea gigas. C R Biol 325: 231. 641
Wang, L., Song, L., Zhao, J., Qiu, L., Zhang, H., Wei, X. et al. (2009). Expressed sequence tags 642
from the zhikong scallop (Chlamys farreri): Discovery and annotation of host defense genes. 643
Fish & Shellfish Immunol 26: 744-750. 644
Page 20 of 25
Wiley-Blackwell and Society for Applied Microbiology
For Peer Review O
nly
Figure 1. Signs of disease in experimentally infected larvae: (A) Swarms of bacteria on
the margins of the larvae, the so called “bacterial swarming” (arrow) surrounding the
larvae (Bar 200 µm). (B) Moribund larvae agglutinating at the bottom of the tanks, the
so called the “spotting” phenomenon (Bar 200 µm).
Page 21 of 25
Wiley-Blackwell and Society for Applied Microbiology
For Peer Review Only
Table 1. Experimental infections of bivalve larvae with pathogenic Vibrio spp. *Abbreviations: C = Crassostrea, M = Mytilus, O = Ostrea, P=
Pecten, R = Ruditapes. NS= Not specified.
Pathogenic species Vibrio identification
method
Origin of the tested
strain/s Host*
Experimental
observations Reference
V. alginolyticus NS Culture collection M. galloprovincialis Sublethal, abnormal larvae Anguiano-Beltrán et al., 2004
Vibrio sp. Phenotypic Mortality outbreak C. gigas
O. edulis Mortality Estes et al., 2004
V. splendidus biovar II
V. alginolyticus
Phenotypic,
16S rDNA Mortality outbreaks R. decussatus
Mortality, reduced
haemocyte survival Gómez-León et al., 2005
V. neptunius
Vibrio sp.
Phenotypic,
16S rDNA Mortality outbreaks O. edulis Mortality Prado et al., 2005
V. pectenicida
V. splendidus-like PCR identification Mortality outbreaks P. maximus Mortality Sandlund et al., 2006
Vibrio sp. NS Pacific oyster C. virginica Mortality Gómez-León et al., 2008
V. tubiashii NS Pacific oyster C. gigas
Destruction of tissues,
toxicity due to an
extracellular
metalloprotease
Hasegawa et al., 2008
V. tubiashii
V. splendidus
V. cholerae
NS NS C. gigas
Toxicity due to an
extracellular
metalloprotease
Hasegawa et al., 2009
Page 22 of 25
Wiley-Blackwell and Society for Applied Microbiology

For Peer Review Only
Table 2. Experimental infections of juvenile and adult bivalves with pathogenic Vibrio spp. *Abbreviations: C = Crassostrea, R = Ruditapes.
BRD= Brown Ring Disease, ECP= Extracellular products, NS= Not specified.
Pathogenic species
Vibrio
identification
method
Origin of the
tested strain/s
Host* Life
stage
Experimental observations Reference
V. tapetis
NS
Symptomatic
clams
R. philippinarum
R. decussatus
Adults
Mortality, BRD, reduced
haemocyte phagocitosis
Mortality
Decreasing viability and
reduced haemocyte
phagocitosis
Host cell damage
Allam et al., 2001
Allam et al., 2002
Allam and Ford 2006
Allam et al., 2006
V. splendidus Phenotypic and
genotypic (NS)
Moribund
oysters
C. gigas Juveniles Mortality Lacoste et al., 2001
V. splendidus biovar II
(V. chagasii)
Phenotypic,
16S rDNA
Mortality
outbreak
C. gigas Juveniles Mortality Waechter et al., 2002
V. splendidus-like gyrB Moribund/
healthy oysters
C. gigas
R. philippinarum
Adults
Seed
Weakness of adductor
muscle
Gay et al., 2004b
V. aestuarianus Phenotypic, 16S
rDNA and gyrB
gyrB-RFLP,
16S rDNA
Mortality
outbreak
Diseased/
healthy oysters
C. gigas
Adults
Reduced haemocyte
adhesive capacities and
phagocitosis.
Mortality depending on the
strain
Labreuche et al., 2006 a, b
Garnier et al., 2007
V. aestuarianus subsp.
francensis
gyrB, toxR,
DNA-DNA
Mortality
outbreak
C. gigas Adults Mortality depending on the
strain
Garnier et al., 2008
Vibrio sp. DNA-DNA
NS
Moribund/
healthy oysters
Pacific oyster
C. gigas
C. virginica
Juveniles
Juveniles
Mortality depending on the
strain
Mortality, ECP toxic to
haemocytes
Gay et al., 2004 a
Gómez-León et al., 2008
Page 23 of 25
Wiley-Blackwell and Society for Applied Microbiology
For Peer Review Only
Table 1. Experimental infections of bivalve larvae, juvenile and adults with pathogenic Vibrio spp. *Abbreviations: C = Crassostrea, M =
Mytilus, O = Ostrea, P= Pecten, R = Ruditapes, BRD= Brown Ring Disease, ECP= Extracellular products, NS= Not specified.
Pathogenic species Vibrio identification
method
Origin of the tested
strain/s Host*
Life stage Experimental observations Reference
V. alginolyticus NS Culture collection M. galloprovincialis Larvae Sublethal, abnormal larvae Anguiano-Beltrán et al., 2004
Vibrio sp. Phenotypic Mortality outbreak C. gigas
O. edulis
Larvae Mortality Estes et al., 2004
V. splendidus biovar II
V. alginolyticus
Phenotypic,
16S rDNA Mortality outbreaks R. decussatus
Larvae Mortality, reduced haemocyte
survival Gómez-León et al., 2005
V. neptunius
Vibrio sp.
Phenotypic,
16S rDNA Mortality outbreaks O. edulis
Larvae Mortality Prado et al., 2005
V. pectenicida
V. splendidus-like PCR identification Mortality outbreaks P. maximus
Larvae Mortality Sandlund et al., 2006
Vibrio sp. NS Pacific oyster C. virginica Larvae Mortality Gómez-León et al., 2008
V. tubiashii NS Pacific oyster C. gigas Larvae Destruction of tissues, toxicity due
to an extracellular metalloprotease Hasegawa et al., 2008
V. tapetis
NS
Symptomatic clams R. philippinarum
R. decussatus
Adults
Mortality, BRD, reduced
haemocyte phagocitosis
Mortality
Decreasing viability and reduced
haemocyte phagocitosis
Host cell damage
Allam et al., 2001
Allam et al., 2002
Allam and Ford 2006
Allam et al., 2006
V. splendidus Phenotypic and
genotypic (NS)
Moribund oysters C. gigas Juveniles Mortality Lacoste et al., 2001
V. splendidus biovar II
(V. chagasii)
Phenotypic,
16S rDNA
Mortality outbreak C. gigas Juveniles Mortality Waechter et al., 2002
V. splendidus-like gyrB Moribund/ healthy
oysters
C. gigas
R. philippinarum
Adults
Seed
Weakness of adductor muscle Gay et al., 2004b
V. aestuarianus Phenotypic, 16S
rDNA and gyrB
Mortality outbreak
C. gigas
Adults
Reduced haemocyte adhesive
capacities and phagocitosis.
Labreuche et al., 2006 a, b
Page 24 of 25
Wiley-Blackwell and Society for Applied Microbiology
For Peer Review Only
Pathogenic Vibrio species mainly cause mortality of bivalve larvae but they are able to cause different disease symptoms in juvenile and adults
where mortality seems to be strain specific in many cases.
gyrB-RFLP,
16S rDNA
Diseased/ healthy
oysters
Mortality depending on the strain
Garnier et al., 2007
V. aestuarianus subsp.
francensis
gyrB, toxR,
DNA-DNA
Mortality outbreak C. gigas Adults Mortality depending on the strain Garnier et al., 2008
Vibrio sp. DNA-DNA
NS
Moribund/ healthy
oysters
Pacific oyster
C. gigas
C. virginica
Juveniles
Juveniles
Mortality depending on the strain
Mortality, ECP toxic to haemocytes
Gay et al., 2004 a
Gómez-León et al., 2008
Page 25 of 25
Wiley-Blackwell and Society for Applied Microbiology
Related Documents









![有效日期從.. 國別 (zh) Canada 00295 [ZH] SECTION Live bivalve … · 2020. 8. 20. · 1 / 46 國別 [ZH] SECTION (zh) Canada Live bivalve molluscs 有效日期從.. 10/08/2007](https://static.cupdf.com/doc/110x72/5fd99fb0e13f48596642571a/oeoe-oe-zh-canada-00295-zh-section-live-bivalve-2020-8-20.jpg)

