Rapid Publication Distribution of the EP3 Prostaglandin E 2 Receptor Subtype in the Rat Brain: Relationship to Sites of Interleukin-1– Induced Cellular Responsiveness MONICA EK, 1 * CARLOS ARIAS, 2 PAUL SAWCHENKO, 2 AND ANDERS ERICSSON-DAHLSTRAND 1 1 Department of Medicine, Unit of Rheumatology, The Karolinska Institute, S-171 76, Stockholm, Sweden 2 Laboratory of Neuronal Structure and Function, The Salk Institute for Biological Studies, La Jolla, CA 92037 ABSTRACT The activation of neurosecretory neurons that express corticotropin-releasing hormone (CRH) in response to increased circulating levels of interleukin-1b (IL-1b) depends on prosta- glandin E 2 (PGE 2 ) acting locally within the brain parenchyma. To identify potential central targets for PGE 2 relevant to pituitary-adrenal control, the distribution of mRNA encoding the PGE 2 receptor subtype EP3 (EP3R) was analyzed in rat brain. Hybridization histochemistry revealed prominent labeling of cells in discrete portions of the olfactory system, iso- and hip- pocampal cortices, and subcortical telencephalic structures in the septal region and amygdala. Labeling over the midline, intralaminar, and anterior thalamic groups was particularly promi- nent. EP3R expression was enriched in the median preoptic nucleus and adjoining aspects of the medial preoptic area (MPO) implicated in thermoregulatory/febrile responses and sleep induc- tion. EP3R-expressing cells were also prominent in brainstem cell groups involved in nociceptive information processing/modulation (periaqueductal gray, locus coeruleus (LC), parabrachial nu- cleus (PB), caudal raphe ´ nuclei), arousal and wakefulness (LC, midbrain raphe ´ and tuberomam- millary nuclei); and in conveying interoceptive input, including systemic IL-1 signals, to the endocrine hypothalamus (nucleus of the solitary tract (NTS) and rostral ventrolateral medulla [VLM]). Combined hybridization histochemical detection of EP3R mRNA with immunolocaliza- tion of IL-1b–induced Fos protein expression identified cytokine-sensitive, EP3R-positive cells in the medial NTS, rostral VLM, and, to a lesser extent, aspects of the MPO. These findings are consistent with the view that increased circulating IL-1 may stimulate central neural mecha- nisms, including hypothalamic CRH neurons, through an EP3R-dependent mechanism involving PGE 2 -mediated activation of cells in the caudal medulla and/or preoptic region. J. Comp. Neurol. 428:5–20, 2000. © 2000 Wiley-Liss, Inc. Indexing terms: acute-phase response; neuroendocrine; pain; fever; sleep; c-Fos, blood-brain barrier Grant sponsor: The Swedish Medical Research Council; Grant sponsor: The Swedish Cancer Foundation; Grant number: 4095; Grant sponsor: The Wenner-Gren Center Foundation for Scientific Research; Grant sponsor: The Swedish Society for Medicine; Grant sponsor: The Swedish Association of Rheumatology Research; Grant sponsor: The King Gustaf V 80th Year Foundation; Grant sponsor: Foundations of the Karolinska Institute; Grant sponsor: Nanna Swartz; Grant sponsor: Sven and Ebba-Christina Hagberg; Grant sponsor: Harald and Greta Jeansson; Grant sponsor: Sven and Dagmar Sahle ´n; Grant sponsor: Ulla and Gustaf af Uggla; Grant sponsor: Bo ¨rje Dahlin; Grant sponsor: Lars Hierta; Grant sponsor: Åke Wiberg; Grant sponsor: National Institutes of Health; Grant number: NS-21182. *Correspondence to: Monica Ek, Laboratory of Rheumatology, Center for Molecular Medicine, Building L8:04, The Karolinska Hospital, S-171 76, Stockholm, Sweden. E-mail: [email protected] Received 30 May 2000; Revised 14 August 2000; Accepted 15 August 2000 THE JOURNAL OF COMPARATIVE NEUROLOGY 428:5–20 (2000) © 2000 WILEY-LISS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Rapid Publication
Distribution of the EP3 Prostaglandin E2
Receptor Subtype in the Rat Brain:Relationship to Sites of Interleukin-1–
Induced Cellular Responsiveness
MONICA EK,1* CARLOS ARIAS,2 PAUL SAWCHENKO,2
AND ANDERS ERICSSON-DAHLSTRAND1
1Department of Medicine, Unit of Rheumatology, The Karolinska Institute,S-171 76, Stockholm, Sweden
2Laboratory of Neuronal Structure and Function, The Salk Institute for Biological Studies,La Jolla, CA 92037
ABSTRACTThe activation of neurosecretory neurons that express corticotropin-releasing hormone
(CRH) in response to increased circulating levels of interleukin-1b (IL-1b) depends on prosta-glandin E2 (PGE2) acting locally within the brain parenchyma. To identify potential centraltargets for PGE2 relevant to pituitary-adrenal control, the distribution of mRNA encoding thePGE2 receptor subtype EP3 (EP3R) was analyzed in rat brain. Hybridization histochemistryrevealed prominent labeling of cells in discrete portions of the olfactory system, iso- and hip-pocampal cortices, and subcortical telencephalic structures in the septal region and amygdala.Labeling over the midline, intralaminar, and anterior thalamic groups was particularly promi-nent. EP3R expression was enriched in the median preoptic nucleus and adjoining aspects of themedial preoptic area (MPO) implicated in thermoregulatory/febrile responses and sleep induc-tion. EP3R-expressing cells were also prominent in brainstem cell groups involved in nociceptiveinformation processing/modulation (periaqueductal gray, locus coeruleus (LC), parabrachial nu-cleus (PB), caudal raphe nuclei), arousal and wakefulness (LC, midbrain raphe and tuberomam-millary nuclei); and in conveying interoceptive input, including systemic IL-1 signals, to theendocrine hypothalamus (nucleus of the solitary tract (NTS) and rostral ventrolateral medulla[VLM]). Combined hybridization histochemical detection of EP3R mRNA with immunolocaliza-tion of IL-1b–induced Fos protein expression identified cytokine-sensitive, EP3R-positive cells inthe medial NTS, rostral VLM, and, to a lesser extent, aspects of the MPO. These findings areconsistent with the view that increased circulating IL-1 may stimulate central neural mecha-nisms, including hypothalamic CRH neurons, through an EP3R-dependent mechanism involvingPGE2-mediated activation of cells in the caudal medulla and/or preoptic region. J. Comp. Neurol.428:5–20, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: acute-phase response; neuroendocrine; pain; fever; sleep; c-Fos, blood-brain barrier
Grant sponsor: The Swedish Medical Research Council; Grant sponsor:The Swedish Cancer Foundation; Grant number: 4095; Grant sponsor: TheWenner-Gren Center Foundation for Scientific Research; Grant sponsor:The Swedish Society for Medicine; Grant sponsor: The Swedish Associationof Rheumatology Research; Grant sponsor: The King Gustaf V 80th YearFoundation; Grant sponsor: Foundations of the Karolinska Institute;Grant sponsor: Nanna Swartz; Grant sponsor: Sven and Ebba-ChristinaHagberg; Grant sponsor: Harald and Greta Jeansson; Grant sponsor: Sven
and Dagmar Sahlen; Grant sponsor: Ulla and Gustaf af Uggla; Grantsponsor: Borje Dahlin; Grant sponsor: Lars Hierta; Grant sponsor: ÅkeWiberg; Grant sponsor: National Institutes of Health; Grant number:NS-21182.
*Correspondence to: Monica Ek, Laboratory of Rheumatology, Center forMolecular Medicine, Building L8:04, The Karolinska Hospital, S-171 76,Stockholm, Sweden. E-mail: [email protected]
Received 30 May 2000; Revised 14 August 2000; Accepted 15 August 2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 428:5–20 (2000)
© 2000 WILEY-LISS, INC.

The marked changes in autonomic, endocrine, and be-havioral functions that follow local or systemic inflamma-tion are mediated via pro-inflammatory substances suchas IL-1b (Dinarello, 1991). Intravenous (i.v.) administra-tion of IL-1b stimulates secretion of adrenocorticotropichormone (ACTH) through a mechanism dependent on therelease of hypothalamic CRH (Berkenbosch et al., 1987;Sapolsky et al., 1987) and on the integrity of cat-echolaminergic pathways from the lower brainstem to thehypothalamus (Ericsson et al., 1994). Interestingly, sev-eral brain regions, including medullary aminergic cellgroups, display dose-dependent patterns of neuronal acti-vation in response to IL-1b. Since none of these IL-1–sensitive neurons express the transmembrane-signalingtype 1 IL-1 receptor (IL-1R1) (Ericsson et al., 1995), cyto-kine effects on these neurons are likely to be mediatedindirectly. The IL-1R1 is expressed abundantly by cells atthe interface between the systemic circulation and thebrain parenchyma, including cells associated with the ce-rebral vasculature and the area postrema (Cunningham
et al., 1992; Yabuuchi et al., 1994; Ericsson et al., 1995).These structures also exhibit indices of cellular activation(i.e., elevated expression of the immediate-early genesNGFI-B and IkB-a) following i.v. administration of IL-1(Ericsson et al., 1995; Quan et al., 1997).
Activation of IL-1R1–bearing elements at these poten-tial sites of transduction has been proposed as triggeringlocal release of secondary signaling substances, notablyprostaglandins, which in turn may act on neurons regu-lating the hypothalamo-pituitary-adrenal (HPA) axis.Systemic administration of IL-1 results in increased ex-pression of cyclooxygenase (COX)-2 (Cao et al., 1996; La-croix and Rivest, 1998), a key enzyme in the prostanoidbiosynthesis, and of PGE2-immunoreactivity (Van Dam etal., 1996) in the cerebral vasculature. In addition, mRNAsencoding IL-1R1 and COX-2 mRNA colocalize withinendothelial-like cells of the cerebral microvasculature fol-lowing intraperitoneal (i.p.) IL-1b administration (Cao etal., 1996). Pretreatment with indomethacin (an inhibitorof prostaglandin synthesis) attenuates IL-1–stimulated
Abbreviations
AAA anterior amygdaloid areaac anterior commissureAD anterodorsal nucleus thalamusAl anterior lobeAM anteromedial nucleus thalamusAMB nucleus ambiguusAOB accessory olfactory bulbAON anterior olfactory nucleusAP area postremaARH arcuate nucleus hypothalamusAV anteroventral nucleus thalamusAVP anteroventral preoptic nucleusBST bed nuclei stria terminalisbv blood vesselCA1 field CA1 (hippocampal formation)CA2 field CA2 (hippocampal formation)CA3 field CA3 (hippocampal formation)cc corpus callosumCEA central nucleus amygdalaCOAp cortical nucleus amygdala, posterior partCP caudoputamenCSm superior central nucleus raphe, medial partCVLM caudal ventrolateral medullaDCO dorsal cochlear nucleusDG dentate gyrusDMH dorsomedial nucleus hypothalamusDMX dorsal motor nucleus vagus nerveDR dorsal raphe nucleusENT entorhinal areaep ependymal cell layerEPd dorsal endopiriform nucleusfi fimbriaFRP frontal polegr granule cell layerHIP hippocampal regionIC inferior colliculusil intermediate lobeISO isocortexLA lateral nucleus amygdalaLC locus coeruleusLD lateral dorsal nucleus thalamusle external lateral subnucleus (parabrachial nucleus)LGd lateral geniculate nucleus, dorsal partLHA lateral hypothalamic areaLM lateral mammillary nucleusLP lateral posterior nucleus thalamusLSi intermediate lateral septal nucleusmcp middle cerebellar peduncleMD mediodorsal nucleus thalamus
me median eminenceMePO median preoptic nucleusmi mitral cell layer (olfactory bulb)MGm medial geniculate complex, medial partMM medial mammillary nucleusMOB main olfactory bulbMPO preoptic areaMPN medial preoptic nucleusNTS nucleus of the solitary tractoch optic chiasmopt optic tractOT olfactory tubercleOVLT vascular organ of the lamina terminalisPAG periaqueductal grayPAGdl periaqueductal gray, dorsolateral partPB parabrachial nucleusPF parafascicular nucleusPG pontine grayPIR piriform cortexPMv ventral premammillary nucleuspo polymorph layer (dentate gyrus)PS parastrial nucleusPVH paraventricular nucleus hypothalamusPVT paraventricular nucleus thalamusRE nucleus reuniensRH rhomboid nucleusRM nucleus raphe magnusRO nucleus raphe obscurusRPa nucleus raphe pallidusRVLM rostral ventrolateral medullaSC superior colliculusscp superior cerebellar pedunclesg granule cell layer (dentate gyrus)SGN suprageniculate nucleusSN substantia nigraSO supraoptic nucleusTM tuberomammillary nucleusSPV spinal nucleus of the trigeminalV motor nucleus of the trigeminal nerveVAL ventral anterior-lateral complex thalamusv3 third ventriclev4 fourth ventricleVL lateral ventricleVLM ventrolateral medullaVLPO ventral lateral preoptic nucleusVMPO ventral medial preoptic nucleusVn trigeminal nerveVPM ventral posteromedial nucleus thalamusXII hypoglossal nucleus
6 M. EK ET AL.

ACTH release (Katsuura et al., 1988; Morimoto et al.,1989; Watanabe et al., 1990) and IL-1–induced neuronalactivation in the hypothalamus (Ericsson et al., 1997).Local microinjection of PGE2 into the rostral VLM, theprincipal source of cytokine-sensitive aminergic projec-tions to the endocrine hypothalamus, mimics the centralactivational effects of a systemic IL-1 challenge, includingrecruitment activation of CRH-expressing neurosecretoryneurons (Ericsson et al., 1997).
A possible role for PGE2 as an intermediate messengerbetween circulating IL-1 and IL-1–sensitive neuronswithin the brain requires that the latter express PGE2receptors. PGE2 receptors are divided into four subtypes,designated EP1–4, that differ in ligand-binding specific-ity, tissue distribution, and coupling to intracellular sig-nal transduction pathways (reviewed in Negishi et al.,1995). Northern blot and in situ hybridization analyseshave previously demonstrated expression of EP3R mRNAin mouse brain (Sugimoto et al., 1992, 1994) and immu-nohistochemical analyses have revealed EP3R-like immu-noreactivity in rat brain (Nakamura et al., 2000). More-over, Zhang and Rivest (1999) recently demonstratedinduced expression of EP2 and EP4 receptor mRNAs inthe brain of rats challenged with i.v. lipopolysaccharide orIL-1. In the present study we utilize a combination of insitu hybridization and immunohistochemical techniquesto explore the central distribution of the EP3R mRNAwithin the rat central nervous system (CNS), with specialemphasis on IL-1–responsive cell groups.
MATERIALS AND METHODS
Animals
Pathogen-free adult male Sprague-Dawley rats (280–320 g) were housed individually in a room with constanttemperature and on a 12-hour light-dark cycle (lights onat 06.00 h), with food and water freely available. To avoidstress during the injection procedure, the animals werehandled daily for 6 to 7 days before the day of injection. Allanimal experiments were approved and performed in ac-cordance with Swedish and NIH guidelines.
Systemic administration of IL-1b
Recombinant human IL-1b (kindly provided by Dr. S.Gillis, Immunex Research and Development Corp., Seat-tle, WA) corresponding to the 152-residue mature form ofthe protein was utilized. The original specific biologicalactivity of this protein exceeded 1 3 105 units/mg protein(A375 assay; Nakano et al., 1988; 17 pg endotoxin/mgprotein). The IL-1b protein was prepared for i.v. injectionas previously described (Ericsson et al., 1997). The centraleffects of this cytokine preparation have been extensivelycharacterized (Ericsson and Sawchenko, 1993; Ericsson etal., 1994, 1995, 1997).
Systemic administration of IL-1b to awake and freelymoving rats in their home cages was carried out viaan intrajugular venous catheter as previously described(Ericsson and Sawchenko, 1993).
Perfusion and histology
Animals were deeply anesthetized with CO2 or chloralhydrate (350 mg/kg, i.p.) and perfused immediately viathe aorta with 0.9% NaCl followed by 300–400 ml ofice-cold fixative solution (4% paraformaldehyde in 0.1 M
borate buffer, pH 9.5) for 20 to 25 min. Brain, pituitary,and adrenal glands were removed and postfixed in fixativesolution containing 10% sucrose for 3 hours, and thencryoprotected in 0.1 M phosphate buffer (pH 7.4) contain-ing 10% sucrose overnight at 4°C. Brains were frozen andcut using a cryostat into 30-mm-thick coronal sections,which were collected in cold cryoprotectant (0.05 M so-dium phosphate buffer, pH 7.4; 30% ethylene glycol, 20%glycerol) in five 1-in-6 series and then stored at 220°C.Pituitary and adrenal glands were cut into 12-mm-thicksections, which were mounted directly onto Probe On Plusslides (Fischer Scientific, Houston, TX).
Radiolabeled cRNA probes
Preparation of radioactively labeled cRNA probes wasconducted as previously described (Simmons et al., 1989).In brief, a 885-bp cDNA fragment-encoding part of theEP3Ra isoform of the rat PGE2 receptor was generated byreverse transcriptase-PCR using sequence-specific oligo-nucleotide primers (59primer: GAGAGCAAACGCAAAA-AGTC, 39primer: CAGGGCAAGGAGGTAGAGCTG; Takeu-chi et al., 1993) and cloned into a pBluescript II KS1plasmid vector (Stratagene, La Jolla, CA). Identity of thecloned cDNA fragment was confirmed by restriction map-ping and DNA sequencing. The plasmid was subsequentlylinearized with XbaI and HindIII restriction enzymes.Anti-sense and sense (control) cRNA probes were tran-scribed by T3 and T7 RNA polymerases, respectively, inthe presence of (a-33P)-UTP (NEN Life Science Products,Inc., Boston, MA) to a specific activity of 1–3 3 109 cpm/mg. Unincorporated nucleotides were removed by usingQuick Spin columns (Boehringer Mannheim, Indianapo-lis, IN). The restriction enzymes and RNA polymeraseswere obtained from Promega (Madison, WI). The EP3RacDNA fragment encoded a large sequence (838 bp) com-mon to all isoforms of the EP3R and a short (47 bp)sequence unique to the EP3Ra (Takeuchi et al., 1993). Theprobes were therefore expected to hybridize to mRNAsencoding all EP3R isoforms. A Blast search of Genbank(June 29, 2000 release) revealed that this sequence ishighly specific to the rat EP3R with no other sequencesexpected to generate cross-hybridization under the strin-gency conditions utilized in this study. In addition, alarger sized rat EP3Ra cDNA construct, which includedthe entire sequence used in this study, has previouslybeen demonstrated by Northern blot analysis to be specificto the EP3R, with no cross-hybridization to the otherEP-receptor isoforms (Boie et al., 1997).
In situ hybridization
To detect expression of EP3R mRNA in the brain, in situhybridization experiments were performed on coronal tis-sue sections from rat brains. Hybridization and autora-diography were carried out following protocols previouslydescribed (Simmons et al., 1989). Briefly, brain sectionswere mounted on Probe On Plus slides and dried undervacuum overnight. Sections of brain, pituitary, and adre-nal glands were further postfixed in 4% paraformaldehyde(pH 7.4) for 30 minutes, digested in 10 mg/ml proteinase Kat 37°C for 20 min, acetylated for 10 minutes, dehydrated,and then vacuum dried for at least 3 hours. After appli-cation of 120 ml of hybridization solution, containing 106
cpm of the cRNA probes, the slides were mounted withcoverslips and incubated at 60°C for 16 to 20 hours. Slideswere subsequently washed in 43 standard saline citrate
7PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS

(SSC, pH 7.0), digested in 20 mg/ml Ribonuclease A solu-tion at 37°C for 30 minutes, washed in decreasing concen-trations of SSC, ending with 0.13SSC for 30 minutes at 75to 76°C, dehydrated with ethanol, and dried. The slideswere then defatted in ethanol and xylene, and dipped inKodak NTB-2 (Eastman Kodak, Rochester, NY) nucleartrack emulsion. After 14 to 30 days of exposure at 4°C theslides were developed in Kodak D-19 developer for 4 min-utes at 14 to 15°C. Sections were then counterstained with0.1% cresyl violet and coverslipped with coverslips.
Combined immuno- and hybridizationhistochemistry
Expression of the EP3R mRNA in cells displaying IL-1b–induced expression of the cellular activation marker,Fos, was studied by using a protocol for dual immuno- andhybridization histochemical labeling (Watts and Swanson,1989). The immunohistochemical detection of Fos-likeimmunoreactivity (Fos-LI) was performed using anavidin-biotin-immunoperoxidase technique on free-floating tissue sections from both IL-1b- and vehicle(control)-injected rats. Briefly, sections were pretreatedwith 0.3% H2O2 for 10 minutes to block endogenous per-oxidase activity, incubated with primary antibody solution(1:5,000 diluted primary antibody in phosphate-bufferedsaline (PBS), pH 7.4, 0.4% Triton X-100, 2% BSA, 5 mg/mlheparin, and 40 U/ml RNasin) for 48 hours at 4°C. Theprimary antiserum used was an affinity-purified poly-clonal antiserum raised in rabbit against a synthetic pep-tide corresponding to residues 4–17 of the N-terminalportion of the human Fos protein (Oncogene Sciences,Cambridge, MA). This antiserum is not known to cross-react with any identified Fos-related antigen, and previ-ous study has determined that the nuclear staining isabolished by overnight preincubation of the antiserum in50 mM of the synthetic peptide (Ericsson et al., 1994).
Sections were then incubated with secondary antibodysolution (1:200 diluted biotinylated goat anti-rabbit IgGantibody in PBS, 0.4% Triton X-100, 5 mg/ml heparin, and40 U/ml RNasin, at room temperature for 1 hour), andthen with avidin-biotin-peroxidase complex (Elite ABCkit, Vector Laboratory, Burlingame, CA) for 1 hour atroom temperature. After consecutive washes in 0.1 Macetate buffer (pH 6.0) and the same buffer containing 1%CoAc, reaction product was developed in a solution con-taining diaminobenzidine (DAB) (0.5 mg/ml in 0.1 M NaAccontaining 2 mg/ml b-D glucose, 0.2 mg/ml NH4Cl, and0.1% glucose oxidase, at 4°C for 1 to 2 hours. Sections werefinally mounted on Probe On Plus slides for subsequentlocalization of EP3R mRNA by in situ hybridization his-tochemistry as described above.
Imaging
Images were recorded on either Ilford XP-2 (Ilford Im-aging USA Inc., Paramus, NJ) or Kodak Technical Panfilm, and digitized using a Nikon LS-1000 film scanner(Nikon Inc., Melville, NY). Low-magnification darkfieldmacrophotographic imaging (Fig. 1) was carried out usinga Nikon Multiphot system, Kodak Ektapan film, and aKodak RS-3570 film scanner. Digital images were im-ported into Adobe Photoshop (v. 5.0), where they wereadjusted to balance and optimize brightness, contrast, andsharpness. Individual files were exported to Canvas(v. 3.55) for assembly into plates, which were rendered atan initial resolution of 300 dpi.
RESULTS
Distribution of EP3R mRNA in the ratbrain and pituitary
In situ hybridization histochemistry was carried outusing 33P-labeled antisense probes encoding the EP3R on30-mm-thick coronal sections throughout the rat brain andselect peripheral tissues. This revealed a discrete anddistinctive pattern of expression throughout much of theneuraxis, which was fundamentally similar to prior de-scriptions of the EP3R mRNA distribution in mouse brain(Sugimoto et al., 1994). Microscopic analysis at high mag-nifications indicated that intraparenchymal cells express-ing the EP3R mRNA exhibited a neuron-like nuclear sizeand cellular morphology. Whether this is universally thecase remains to be established in double-labeling experi-ments with cell-specific markers. Sense-strand runoffs la-beled to similar specific activities as antisense probes con-sistently failed to provide any suggestion of specificlabeling. The distribution and relative strength of EP3Rexpression in rat brain cell groups is summarized in Table1. Nomenclature follows that is employed in the atlas ofSwanson (1992), except for the identification of the ventralmedial preoptic nucleus (VMPO) and ventral lateral pre-optic nucleus (VLPO) as defined by Elmquist and Saper(1996) and Sherin et al. (1996).
Cerebral cortex. In the isocortex, EP3R mRNA signalappeared most prominently as a continuous band of cellsoccupying the superficial part of layer V. Many fewer andless robustly labeled cells were found to be scattered in thesuperficial aspects of layer II/III. This general pattern oflabeling did not vary appreciably across isocortical fields(Fig. 1D–I). In the hippocampal formation, the strength oflabeling seen over the pyramidal cell layer of Ammon’shorn varied as a function of position along the septo-temporal axis, with labeling predominating over the CA1subfield near the septal pole, CA2 at intermediate levels,and both CA2 and CA3 temporally. By contrast, no suchgradient was apparent in the moderate hybridization sig-nals over the granule cell layer of the dentate gyrus (Figs.1G–I, 2B).
Among olfactory structures, EP3 mRNA signal in theolfactory bulb was most prominent over the mitral celllayer of the accessory bulb (Fig. 1B), with the same layerof the main bulb showing much more sporadic labelingover isolated individual cells (Fig. 1A). Moderately stronglabeling was seen over the anterior olfactory nucleus (Fig.1C), and deeper layers of piriform cortex (Fig. 1D–H).
Subcortical telencephalic structures. In the septalregion, a moderate density of positively hybridized cellswas seen reliably in the intermediate lateral septal nu-cleus. Labeling over the bed nuclei of the stria terminalis(BST) was dominated by a relatively strong and quitediscrete signal over the parastrial nucleus (see Simerly etal., 1984; Fig. 1D–E). Several amygdaloid nuclei exhibitedweak to moderate EP3R hybridization signals, with thoseover the cortical and lateral nuclei predominating. Thecentral and basolateral nuclei, by contrast, failed to dis-play positive labeling (Fig. 1F–I).
The basal ganglia showed little capacity for EP3R ex-pression, though an associated cell group, the reticularpart of the substantia nigra, contained scattered cellsdisplaying a relatively low-level mRNA signal (Fig. 1D–I).
Thalamus. Of all the principal gross divisions of thebrain, EP3R mRNA expression was clearly predominant
8 M. EK ET AL.

in the dorsal thalamus, with an apparent majority of cellsin most of the principal midline, intralaminar, and ante-rior nuclei showing moderate to strong hybridization sig-nals. Labeling over the ventral tier and posterior complexwas generally muted, while discrete aspects of the lateralgeniculate (dorsal nucleus, intergeniculate leaflet) andmedial geniculate (medial and suprageniculate nuclei)groups displayed low to moderate capacities for expressionof EP3R transcripts (Fig. 1F–I).
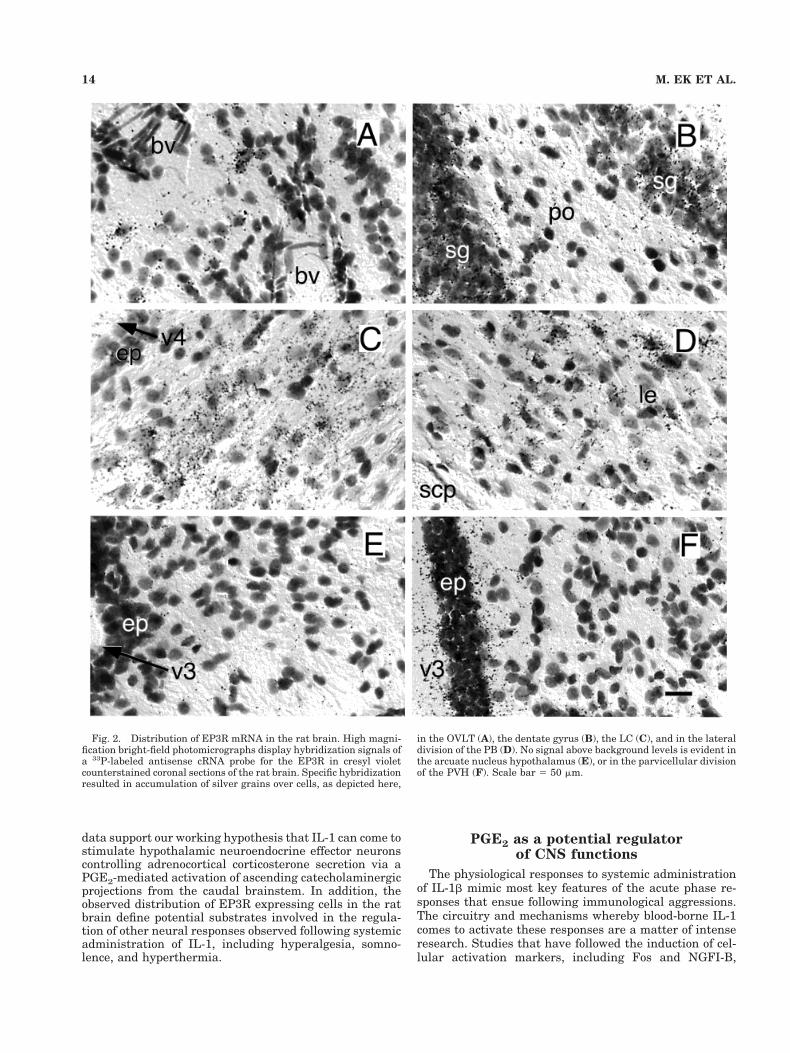
Hypothalamus/preoptic region. The median preop-tic nucleus stood out as the clearly dominant seat of EP3RmRNA signal in preoptic/hypothalamic region (Fig. 1D–E). This appeared at rostral levels as an invertedY-shaped structure, capping the vascular organ of thelamina terminalis (OVLT; Fig. 2A). More scattered cellscontiguous with the tips of the arms of the “Y” extendedventrolaterally and caudally, beyond the generally ac-cepted borders of the median preoptic nucleus, into themedial preoptic nucleus (MPN) and adjoining aspects ofthe MPO, including the caudal aspects of the VMPO,which has been implicated previously in febrile responsesto endotoxic challenges (Scammell et al., 1996, 1998b) andat least the dorsal aspects of the VLPO, which has beenimplicated in sleep induction (Sherin et al., 1996).
Within the hypothalamus, proper, EP3R mRNA expres-sion was generally low, though substantial proportions ofcells in a few discrete cell groups were associated withmoderate hybridization signals (Fig. 1F–I). These in-cluded the dorsomedial nucleus (including its compactdivision), the ventral premammillary, tuberomammillary,median part of the medial mammillary, and lateral mam-millary nuclei. More scattered positively hybridized cellswere seen reliably in the lateral (tuberal part) and poste-rior hypothalamic areas. It is worthy of mention thatapart from isolated cells in the anterior periventricularnucleus, positive hybridization signals were not apparentover cell groups comprising the (neuroendocrine-related)periventricular zone of the hypothalamus, including theparaventricular nucleus hypothalmus (PVH; Fig. 2F) andarcuate nucleus (Fig. 2E).
Brainstem. The most distinctive feature of the EP3RmRNA distribution in the brainstem was its pervasiveassociation with the raphe system (Fig. 1J–N). This in-cluded a major site of expression in the dorsal raphenucleus, somewhat less prominent labeling over the me-dian raphe, and very likely cells in the ventral pons dis-tributed in manner highly reminiscent of the B9 seroto-nergic cell group. Similarly, each of the major brainstemrepresentations, including the nuclei raphe magnus, pon-tis, obscurus, and pallidus, contained substantial numbersof cells displaying moderate to very strong hybridizationsignals, as again, did associated cell groups such as theaspects of the B3 serotonergic cell group that straddle therostral part of the inferior olive.
Among other monoaminergic cell groups, strong EP3Rexpression was seen throughout the LC (Fig. 2C) andsubcoeruleus regions (Fig. 1K). Scattered cells in the A5region of the ventral pons also displayed moderate hybrid-ization signals. In the medulla, populations of positivelyhybridized cells were seen in catecholaminergic regions,being most numerous in the rostral (C1) region of the VLMand caudal (A2) area of the medial NTS (Fig. 1L–N).Hybridization signal over individual cells in medullaryaminergic regions was moderate, and clearly less intensethan that seen over raphe neurons at similar levels.
Other sites of substantial EP3R expression in the brain-stem were noted. In the midbrain (Fig. 1J), these includedthe superior colliculus, where the strongest hybridizationsignals were seen over the intermediate gray, and theperiaqueductal gray matter, where positively hybridizedcells appeared to localize in a highly preferential mannerto the dorsolateral columns (Bandler and Shipley, 1994).In the pons, strong and extensive cellular labeling wasseen reliably in the PB (Fig. 2D), with the most prominentaccumulations of positively hybridized cells seen in thesuperior, central and external lateral subnuclei (Fulwilerand Saper, 1984), and somewhat lesser ones in the medialPB and Kolliker-Fuse nucleus. Apart from the monoam-inergic cell groups alluded to above, EP3R expression inthe medulla was found to be quite limited, though moder-ately strong hybridization signals were seen over the dor-sal motor nucleus of the vagus and the compact formationof the ambiguual complex.
Cerebellum. The deep nuclei comprised the only sitesof EP3R mRNA expression in cerebellum that were evi-dent in our material. Neither the cortex nor the vermisshowed any indication of above-background levels of hy-bridization (Fig. 1K).
Non-neuronal elements. Moderately strong hybrid-ization signals were seen continuously over the ependy-mal lining of the ventricular system throughout the brain,except for tanycytes at the base of the third ventricle atthe level of the arcuate nucleus/median eminence (Fig.1G–H). EP3 mRNA expression was not detected in themeninges, choroid plexus, or in cells associated with thecerebral vasculature.
Pituitary-adrenal axis. In the pituitary, intense la-beling was observed in the intermediate lobe, a prominentsource for the immunomodulatory hormone alpha-melanocyte stimulating hormone, while only weak label-ing was observed over scattered cells in the anterior lobe(Fig. 3A). No specific labeling above background levels wasdetected in cells in the posterior lobe or in the adrenalglands (data not included). In control experiments usingsense probes on adjacent sections of the pituitary (Fig. 3B)and the adrenal glands, no signal above background levelswas detected.
Expression of EP3R mRNA inIL-1b–responsive cells
In line with our principal interest in identifying poten-tial substrates for central prostanoid mediation of neu-roendocrine responses to a systemic cytokine challenge,the central distribution of EP3R mRNA described aboveincluded cell groups identified previously as displayingIL-1–sensitivity and/or as projecting to the PVH. To ex-plicitly identify IL-1–responsive neurons that expressEP3R transcripts, brains from rats injected i.v. with IL-1b(2 mg/kg) 3 hours before sacrifice were prepared for com-bined immuno- and hybridization histochemical demon-stration of Fos-LI and EP3R mRNA. The dose was chosenon the basis of being moderately above the threshold re-quired for activation of CRH-expressing neurosecretoryneurons, and the 3 hour time point was previously foundto yield maximal activational responses in this and otherrelevant cell types in a similar paradigm (Ericsson et al.,1994).
In these experiments, IL-1 injection consistently pro-voked substantial Fos-LI in a number of interconnectedsites involved in central autonomic and neuroendocrine
9PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS
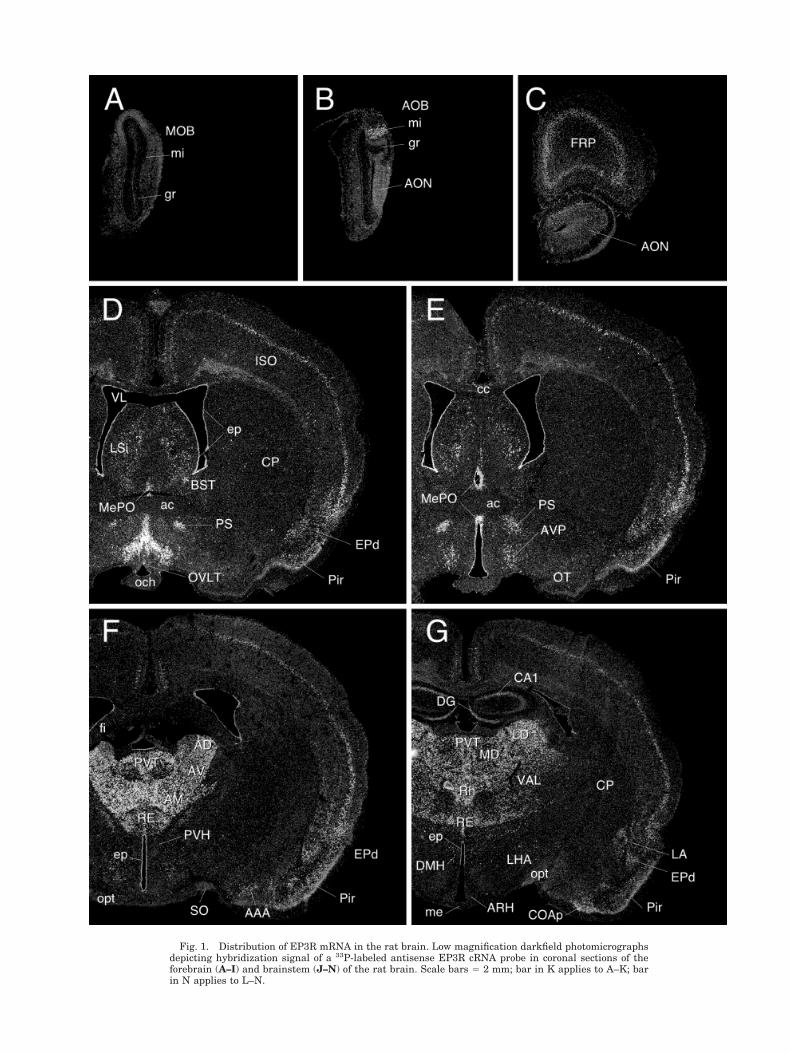
Fig. 1. Distribution of EP3R mRNA in the rat brain. Low magnification darkfield photomicrographsdepicting hybridization signal of a 33P-labeled antisense EP3R cRNA probe in coronal sections of theforebrain (A–I) and brainstem (J–N) of the rat brain. Scale bars 5 2 mm; bar in K applies to A–K; barin N applies to L–N.
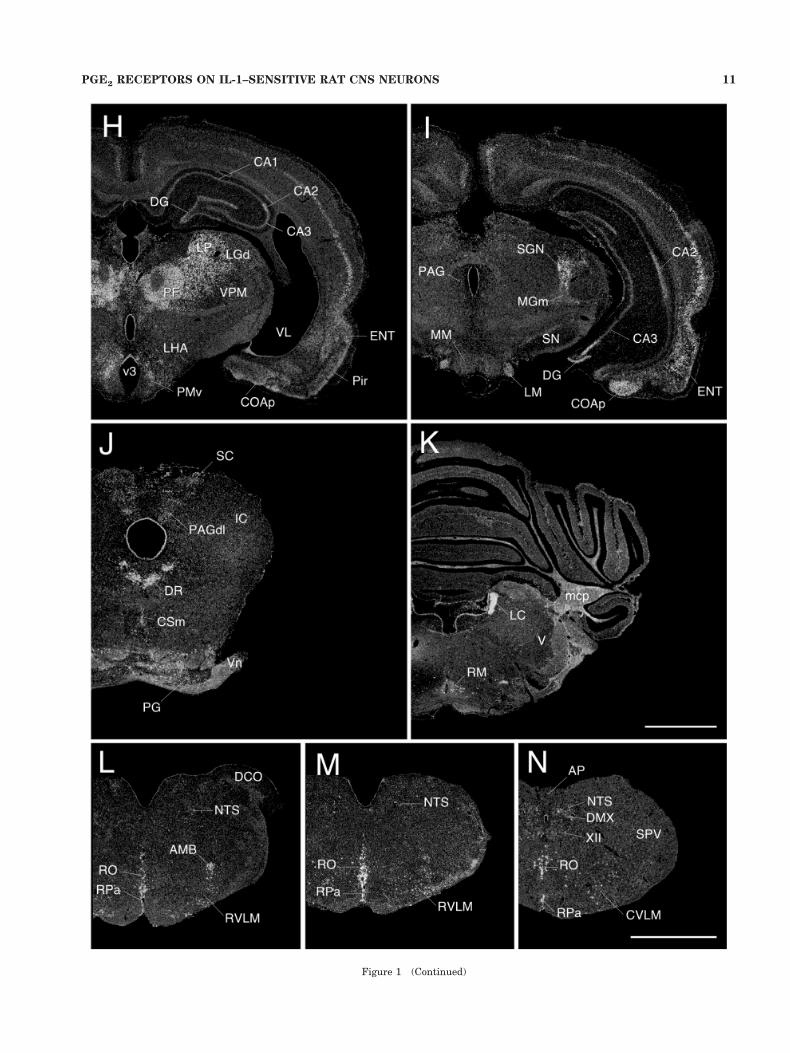
Figure 1 (Continued)
11PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS

TABLE 1. Strength and Distribution of EP3R mRNA Expression in Rat Brain Regions1
Cell group Cell group
I. ForebrainIsocortex
II/III 1IV 2V 111VI 2Claustrum 2
Olfactory regionsMain bulb
Mitral cell layer 1Accessory bulb
Mitral cell layer 111Anterior olfactory nucleus 11Piriform area
I 2II 1III 11Endopiriform nucleus 11
Taenia tecta 2Hippocampal formation
CA1Pyramidal layer 11
CA2Pyramidal layer 11
CA3Pyramidal layer 11
Dentate gyrusGranule cell layer 11
Fasciola cinerea 11Amygdala
Medial nucleusAnterior part 1Posterodorsal part 1
Amygdalohippocampal area 1Cortical nucleus
Anterior part 11Posterior part 111
Nucleus lateral olfactory tract 1Anterior amygdaloid area 2Central nucleus 2Lateral nucleus 11Basolateral nucleus 2Basomedial nucleus 1Intercalated nuclei 2
SeptumLateral nucleus
Dorsal part 2Intermediate part 11Ventral part 2
Medial nucleusNucleus of the diagonal band 1
Bed nuclei stria terminalisRostromedial region 1Parastrial nucleus 11
Bed nucleus anterior commissure 2Septofimbrial nucleus 1Triangular nucleus 2Subfornical organ 11
Basal gangliaCaudoputamen
Posteroventral part 2Nucleus accumbens 2Fundus of striatum 2
Globus pallidusEntopeduncular nucleus 2Substantia innominata 2
Subthalamic nucleus 2Thalamus
Medial habenula 1Lateral habenula 2Anterior group
Anteroventral nucleus 1111Anteromedial nucleus 1111Anterodorsal nucleus 11Interanteromedial nucleus 111Interanterodorsal nucleus 1
Mediodorsal nucleusMedial part 111Central part 1Lateral part 11
Lateral groupLateral dorsal nucleus 11Lateral posterior nucleus 111
Midline groupParaventricular nucleus 111Parataenial nucleus 11Nucleus reuniens 11Rhomboid nucleus 11Nucleus gelatinosa 2
Ventral groupVentral anterior/ventral lateral 11Ventral medial 1Ventral posterior 1
Posterior complex 1Medial geniculate complex
Lateral division 2Medial division 1Suprageniculate nucleus 11
Lateral geniculate complexDorsal lateral geniculate nucleus 11Ventral lateral geniculate nucleus 2Intergeniculate leaflet 1
Intralaminar nucleiCentral medial nucleus 111Paracentral nucleus 11Central lateral nucleus 11
Reticular nucleus 2Zona incerta
Rostral 2Caudal 2
HypothalamusPeriventricular zone
Median preoptic nucleus 1111Anteroventral periventricular nucleus 2Ventral medial preoptic nucleus 1Suprachiasmatic nucleus 2Supraoptic nucleus 2Paraventricular nucleus
Autonomic part 2Parvicellular part 2Magnocellular part, anterior 2
Anterior periventricular nucleus 1Arcuate nucleus 2Posterior periventricular nucleus 2Median eminence
Internal lamina 2External lamina 2
Medial zoneMedial preoptic area 11Medial preoptic nucleus
Lateral part 11Medial part 2Central part 2
Anteroventral preoptic nucleus 11Anterior hypothalamic nucleus
Anterior part 2Central, posterior parts 1
Retrochiasmatic area 2Ventromedial nucleus 2Dorsomedial nucleus 11Tuberomammilary nucleus 11Premammillary nuclei
Dorsal 2Ventral 11
Lateral mammillary nucleus 11Medial mammillary nucleus 2Median mammillary nucleus 11
Lateral zoneLateral preoptic area 2Lateral hypothalamic area 11Posterior hypothalamic area 1
Pituitary glandAnterior lobe 1Intermediate lobe 11Posterior lobe 2
II. BrainstemSensory
VisualSuperior colliculus
Superficial gray 1Intermediate gray 11
Pretectal region 2Parabigeminal nucleus 2Medial terminal nucleus 2
SomatosensoryMesencephalic nucleus V 2Principal sensory nucleus V 2Spinal nucleus V
Oral part 2Interpolar part 2Caudal part 1
Dorsal column nuclei 2External cuneate nucleus 2
AuditoryCochlear nuclei
Dorsal 2Ventral 2
12 M. EK ET AL.

regulation, including the PVH, central nucleus of theamygdala (CEA), BST (oval subnucleus), lateral PB, NTS,and VLM. No more than a few scattered cells displayingnuclear Fos-LI were detected within these brain regions insimilarly prepared rats given injections of the saline ve-hicle alone (n 5 3). These findings are in close agreementwith our previous findings (Ericsson et al., 1994).
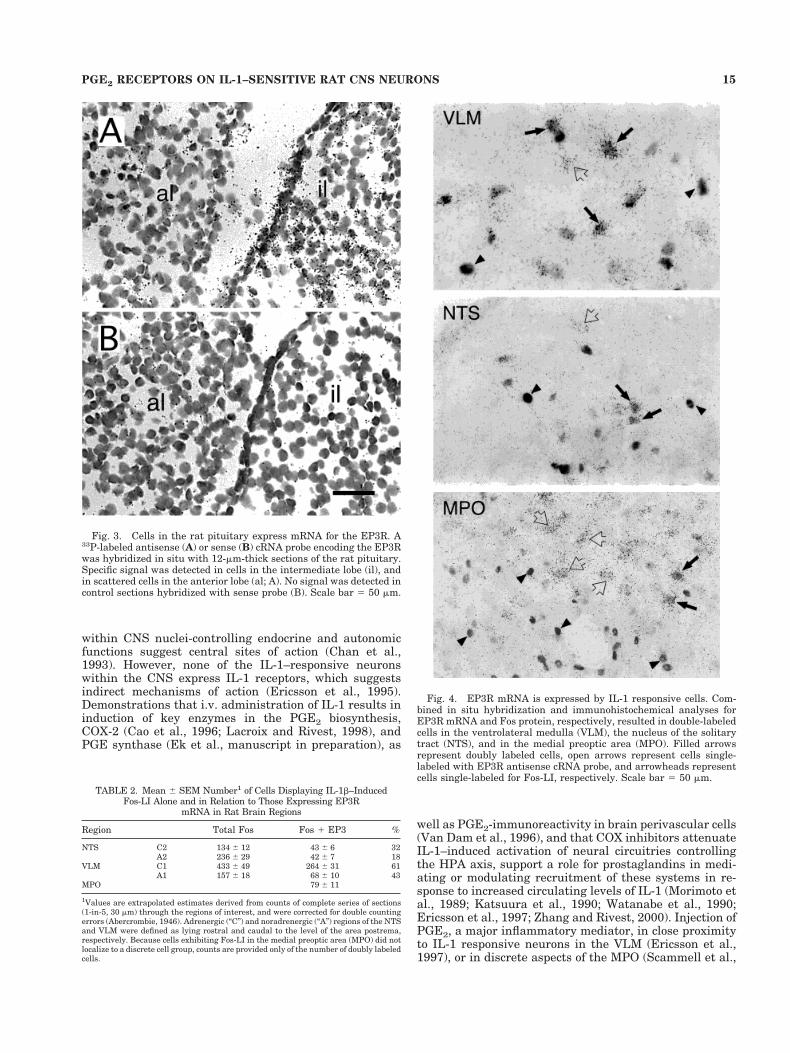
Direct comparison of EP3R mRNA and IL-1b–inducedFos expression in individual neurons identified doublylabeled neurons in discrete aspects of the NTS, VLM,and MPO (Fig. 4; Table 2). These were numericallypredominant in the VLM (Table 2), and were preferen-tially localized to the C1 region of the rostral VLM. Inthe NTS, substantially smaller numbers of IL-1–sensitive neurons associated with EP3R mRNA signalwere roughly evenly distributed over the rostral (C2)and caudal (A2) aspects of the medial NTS. Overall, 43%of all Fos-positive neurons in the NTS and VLM in oursample displayed positive hybridization signals forEP3R mRNA, and it is worthy of mention that substan-tial numbers of Fos-negative, receptor-positive cellswere intermingled with doubly labeled cells, consistentwith the conclusion that overlap between these popula-tions is partial.
Doubly labeled cells were also detected reliably in theMPO (Table 2). Here, the bulk of cells labeled for EP3RmRNA tended to be massed more medially, in andaround the anteroventral preoptic nucleus, while asmall group of cells evincing IL-1–induced Fos-LI wereconcentrated ventrally and laterally to this, encompass-ing at least in part the VMPO of Scammell et al.(1996). While cells labeled singly for receptor andFos-LI tended for the most part to be segregated alongthese lines, doubly labeled neurons were consistentlydetected in the region of partial overlap at the marginsof these distributions. In neither the preoptic region northe medulla were more than occasional isolated exam-ples of doubly labeled cells apparent in vehicle-treatedrats.
Other prominent sites of EP3R mRNA expression, suchas the cerebral cortex, hippocampal formation, thalamus,LC, raphe nuclei, and subfornical organ did not manifestFos-LI under the IL-1b treatment conditions employed inour experiments. Finally, it is noteworthy that no frankdifferences were observed in either the regional distribu-tion or cellular levels of EP3R mRNA expression in com-paring material harvested from IL-1b and vehicle-treatedrats.
DISCUSSION
The present study demonstrates that mRNA encodingthe EP3 subtype of the PGE2 receptor is expressed in ahighly restricted manner in rat brain and pituitary. In-triguingly, and ostensibly complementary to the estab-lished role of prostanoids as local inflammatory mediatorsin the periphery, many of the central sites of EP3R ex-pression conform to cell groups known to be involved inaspects of the acute phase response, including nociceptiveinformation processing and modulation, arousal andwakefulness, as well as febrile and neuroendocrine re-sponses. In line with this generalization, i.v. injection ofIL-1 activated a subset of EP3R expressing cells withinthe caudal aspects of the NTS, the rostral divisions of theVLM as well as in the preoptic hypothalamic area. These
TABLE 1. (continued)
Cell group
Nucleus of the trapezoid body 2Lateral 1Medial 2
Superior olive 2Nucleus of the lateral lemniscus
Ventral 2Dorsal 1
Inferior colliculusExternal 2Dorsal 2Central 1
Nucleus brachium inferior colliculus 2Nucleus sagulum 2
VestibularMedial nucleus 1Lateral nucleus 2Superior nucleus 2Spinal nucleus 1
GustatoryNucleus of the solitary tract, anterior 2
VisceralNucleus of the solitary tract
Medial part 11Commissural part 11Lateral part 1
Area postrema 2Parabrachial nucleus
Lateral 111Medial 11Kolliker-Fuse nucleus 11
MotorJaw
Motor nucleus V 2Face
Facial nucleus (VII) 2Pharynx/larynx
Nucleus ambiguus 11Tongue
Hypoglossal nucleus (XII) 1Viscera
Dorsal motor nucleus X 11Reticular core
Periaqueductal grayDorsolateral column 11Dorsal tegmental nucleus 2Ventral tegmental nucleus 2Laterodorsal tegmental nucleus 2Barrington’s nucleus 2Locus coeruleus 1111
RapheInterfascicular nucleus 2Rostral linear nucleus 2Dorsal nucleus raphe 1Nucleus raphe pontis 11Nucleus raphe magnus 111Nucleus raphe obscurus 1111Nucleus raphe pallidus 1111
Reticular formationPedunculopontine nucleus 2Cuneiform nucleus 2Pontine reticular nucleus 2Linear nucleus medulla 2Parvicellular reticular field 2Gigantocellular reticular nucleus 2Ventrolateral medulla (A1/C1) 11Magnocellular reticular nucleus 111Intermediate reticular field 2Paramedian reticular nucleus 2
Pre- and postcerebellarPontine gray 1Inferior olive 2Nucleus of roller 2Nucleus prepositus 2
III. CerebellumDeep nuclei 11Cortex
Molecular layer 2Purkinje layer 2Granular layer 2
1The relative strength of EP3R mRNA expression was rated by two independentobservers. Ratings reflect primarily the density of positively labeled cells, with (2)representing a complete lack of above-control levels of labeling, (1) isolated positivelylabeled cells, and (1111) labeling in a substantial majority of all cells in a given cellgroup or field. Ratings of mRNA expression were adjusted secondarily on the basis ofthe strength of hybridization signal, but never by more than a single rating point.
13PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS
data support our working hypothesis that IL-1 can come tostimulate hypothalamic neuroendocrine effector neuronscontrolling adrenocortical corticosterone secretion via aPGE2-mediated activation of ascending catecholaminergicprojections from the caudal brainstem. In addition, theobserved distribution of EP3R expressing cells in the ratbrain define potential substrates involved in the regula-tion of other neural responses observed following systemicadministration of IL-1, including hyperalgesia, somno-lence, and hyperthermia.
PGE2 as a potential regulatorof CNS functions
The physiological responses to systemic administrationof IL-1b mimic most key features of the acute phase re-sponses that ensue following immunological aggressions.The circuitry and mechanisms whereby blood-borne IL-1comes to activate these responses are a matter of intenseresearch. Studies that have followed the induction of cel-lular activation markers, including Fos and NGFI-B,
Fig. 2. Distribution of EP3R mRNA in the rat brain. High magni-fication bright-field photomicrographs display hybridization signals ofa 33P-labeled antisense cRNA probe for the EP3R in cresyl violetcounterstained coronal sections of the rat brain. Specific hybridizationresulted in accumulation of silver grains over cells, as depicted here,
in the OVLT (A), the dentate gyrus (B), the LC (C), and in the lateraldivision of the PB (D). No signal above background levels is evident inthe arcuate nucleus hypothalamus (E), or in the parvicellular divisionof the PVH (F). Scale bar 5 50 mm.
14 M. EK ET AL.
within CNS nuclei-controlling endocrine and autonomicfunctions suggest central sites of action (Chan et al.,1993). However, none of the IL-1–responsive neuronswithin the CNS express IL-1 receptors, which suggestsindirect mechanisms of action (Ericsson et al., 1995).Demonstrations that i.v. administration of IL-1 results ininduction of key enzymes in the PGE2 biosynthesis,COX-2 (Cao et al., 1996; Lacroix and Rivest, 1998), andPGE synthase (Ek et al., manuscript in preparation), as
well as PGE2-immunoreactivity in brain perivascular cells(Van Dam et al., 1996), and that COX inhibitors attenuateIL-1–induced activation of neural circuitries controllingthe HPA axis, support a role for prostaglandins in medi-ating or modulating recruitment of these systems in re-sponse to increased circulating levels of IL-1 (Morimoto etal., 1989; Katsuura et al., 1990; Watanabe et al., 1990;Ericsson et al., 1997; Zhang and Rivest, 2000). Injection ofPGE2, a major inflammatory mediator, in close proximityto IL-1 responsive neurons in the VLM (Ericsson et al.,1997), or in discrete aspects of the MPO (Scammell et al.,
Fig. 3. Cells in the rat pituitary express mRNA for the EP3R. A33P-labeled antisense (A) or sense (B) cRNA probe encoding the EP3Rwas hybridized in situ with 12-mm-thick sections of the rat pituitary.Specific signal was detected in cells in the intermediate lobe (il), andin scattered cells in the anterior lobe (al; A). No signal was detected incontrol sections hybridized with sense probe (B). Scale bar 5 50 mm.
TABLE 2. Mean 6 SEM Number1 of Cells Displaying IL-1b–InducedFos-LI Alone and in Relation to Those Expressing EP3R
mRNA in Rat Brain Regions
Region Total Fos Fos 1 EP3 %
NTS C2 134 6 12 43 6 6 32A2 236 6 29 42 6 7 18
VLM C1 433 6 49 264 6 31 61A1 157 6 18 68 6 10 43
MPO 79 6 11
1Values are extrapolated estimates derived from counts of complete series of sections(1-in-5, 30 mm) through the regions of interest, and were corrected for double countingerrors (Abercrombie, 1946). Adrenergic (“C”) and noradrenergic (“A”) regions of the NTSand VLM were defined as lying rostral and caudal to the level of the area postrema,respectively. Because cells exhibiting Fos-LI in the medial preoptic area (MPO) did notlocalize to a discrete cell group, counts are provided only of the number of doubly labeledcells.
Fig. 4. EP3R mRNA is expressed by IL-1 responsive cells. Com-bined in situ hybridization and immunohistochemical analyses forEP3R mRNA and Fos protein, respectively, resulted in double-labeledcells in the ventrolateral medulla (VLM), the nucleus of the solitarytract (NTS), and in the medial preoptic area (MPO). Filled arrowsrepresent doubly labeled cells, open arrows represent cells single-labeled with EP3R antisense cRNA probe, and arrowheads representcells single-labeled for Fos-LI, respectively. Scale bar 5 50 mm.
15PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS

1996), evoke indices of HPA axis activation and hyperther-mia, suggesting that IL-1 recruits these populations bystimulating local release of PGE2. Such a mechanism sug-gests production of PGE2 receptors by IL-1–responsiveneurons in these areas. Interestingly, high levels of PGE2binding sites have been demonstrated within the anteriorwall of the third ventricle and NTS (Matsumura et al.,1992). Recently, mRNA-encoding PGE2 receptors of theEP2 and EP4 subtypes have been demonstrated in selectregions of the rat brain before and after peripheral im-mune challenges (Zhang and Rivest, 1999), including thestructures BST, MPO, PVH, CEA, and the caudal VLM.Here we present data which demonstrate that PGE2 re-ceptors of the EP3 subtype are expressed by a subset ofIL-1–responsive neuron-like cells in the rostral VLM,NTS, and within the preoptic area. Our findings are gen-erally in agreement with a recent immunocytochemicalanalysis of the rat brain that revealed intense EP3R-likeimmunoreactivity within preoptic area (median and me-dial subnuclei), thalamus (anterior, intralaminar andmidline subdivisions), discrete aspects of midbrain andpons (PB, periaqueductal gray and superior colliculus),the raphe nuclear complex, and NTS (Nakamura et al.,2000). These experiments collectively demonstrate thatPGE2 receptors, including the EP3R subtype, are indeedexpressed within the CNS, including regions previouslydemonstrated as nodal points for coordinating central ac-tivation of the HPA axis, among other acute phase re-sponses.
These studies may also collectively provide a potentialbasis for understanding how an omnipresent PGE2 pro-duction by IL-1–stimulated perivascular cells may come toregulate brain functions in a region-specific manner. Thethree PGE2 receptor isoforms exhibit basically distinct,though partly overlapping, patterns of expression—suggesting that they may control different neuronal effec-tor functions as well as, in some cases, modulate oneanother’s activities. For example, EP2 receptor expressionis largely limited to circumventricular organs (area pos-trema and subfornical organ) as well as the CEA and thehypothalamic ventromedial nucleus (Zhang and Rivest,1999). By contrast, the thalamus and raphe nuclei mani-fest a selective expression of the EP3 receptor subtype,whereas the EP4 receptor expression is limited to the PVHand supraoptic nucleus of immune-challenged rats (Zhangand Rivest, 1999). Other regions, such as the preopticarea, LC, and the VLM display partly overlapping pat-terns of expression for all three PGE2 receptor subtypes.The EP2 and EP4 receptor subtypes are considered toactivate intracellular cAMP formation via Gs-stimulatedactivation of adenylate cyclase (Negishi et al., 1995). Bycontrast, the EP3 receptor may both trigger or inhibitadenylate cyclase via the recruitment of Gs or Gi proteins,depending on the utilization of functionally divergentC-terminal domains through alternative splicing (Negishiet al., 1995). The nature of the neuronal response(s) tocentral PGE2 is therefore likely to depend on the localcomplement of EP receptors. The situation is further com-plicated by the fact that individual neurons may altertheir expression of individual EP receptors during thecourse of an inflammatory event (Zhang and Rivest, 1999).Moreover, individual EP receptors exhibit differential af-finities for PGE2 (Boie et al., 1997; Ki values for EP1: 22nM, EP2: 6.8 nM, EP3a: 0.9 nM, and EP4: 1.1 nM), indi-
cating that PGE2-mediated neuronal responses are apt tovary regionally in a dose-dependent manner.
Functional correlates of the EP3Rdistribution in rat brain
Neuroendocrine responses to prostaglandins. TheHPA axis responses to systemic administration of IL-1 isattenuated by prostaglandin synthesis inhibitors such asindomethacin (Morimoto et al., 1989; Katsuura et al.,1990; Watanabe et al., 1990; Ericsson et al., 1997; Zhangand Rivest, 2000). PGE2 has been suggested as the prin-cipal mediator of this response, since intracerebroventric-ular (i.c.v.) or intra-preoptic administration of PGE2 (Mu-rakami and Watanabe, 1989; Rassnick et al., 1995;Parrott and Vellucci, 1996; Zhang and Rivest, 2000) stim-ulate ACTH and glucocorticoid release. In addition, i.c.v.injections of an EP3R selective agonist elicit increases inplasma glucocorticoids (Parrott and Vellucci, 1996), whichindicates that PGE2 can induce HPA axis responses byacting upon EP3R in the brain.
The apparent paucity of EP3R mRNA-expressing cellswithin the parvicellular part of the PVH under basal orcytokine-stimulated conditions suggests that PGE2 doesnot activate CRH-producing neurons directly by way ofthis receptor. However, we have previously demonstratedthat the NTS and the VLM are important structures in-volved in the activation of the central limb of the HPA axisto elevated plasma levels of IL-1b (Chan et al., 1993;Ericsson et al., 1994). A possible role for PGE2 as anintermediate messenger in this pathway is supported byour previous findings that microinjection of PGE2 into theC1 region of the rostral VLM triggers enhanced expressionof Fos-LI and CRH mRNA in the PVH, and essentiallyreplicates the central pattern of neuronal activation seenin response to a systemic IL-1b challenge (Ericsson et al.,1997). In agreement with these earlier findings, thepresent study reveals that IL-1–sensitive cells displayingEP3R mRNA expression are localized within aminergicregions of the VLM and NTS, as well as in the MPO.
While our results define a basis for understanding howdiscrete cell groups relevant to HPA control may come tobe activated by IL-1 in an EP3R-dependent manner, anumber of factors indicate that this is not apt to prove asimple or unitary mechanism. For one, EP3R-expressingcells were identified within each of these regions that didnot exhibit Fos-LI following treatment with IL-1, suggest-ing that some may be refractory to this stimulus. Alter-natively, since the EP3R is generated in different isoformswhich couple to either Gs- or Gi-proteins (Negishi et al.,1995), it is also plausible that PGE2 may instead inhibitsignaling functions in some EP3R-expressing neurons—acellular response that would not be expected to be mani-fest in the form of nuclear induction of Fos. Moreover, thedegree of overlap between IL-1–sensitive and EP3R-positive cells was only partial (Table 2), suggesting theexistence of subpopulations that are not subject to directinfluences of PGE2. Interestingly, the recent findings ofEP4 receptor mRNA expression in IL-1–responsive neu-rons of the NTS and VLM, as well as in the PVH, proposesthat PGE2 may modulate neuronal functions within thesecell group through alternative PGE2 receptor mecha-nisms, as well. However, inconclusive evidences to provethat activation of central command circuitries for neu-roendocrine functions to i.v. IL-1 are indeed mediated viasecretion of perivascular secretion of PGE2 will await the
16 M. EK ET AL.

development and utilization of subtype-specific EP-receptor antagonists.
Fever responses to prostaglandins. PGE2 has beensuggested as mediating febrile responses to endotoxinsand cytokines (Matsumura et al., 1998). Structures liningthe anteroventral aspect of the third ventricle have beenproposed to be a key site for PGE2-mediated febrile re-sponses to immune stimuli (Scammell et al., 1996, 1998b;Blatteis and Sehic, 1997). Local microinjections of PGE2into the VMPO and median preoptic nucleus, regions thatcontain EP3R mRNA-expressing cells, induce fever re-sponses (Scammell et al., 1996). Moreover, Scammell et al.(1998b) have suggested that PGE2 may induce fever byactivating thermoregulatory neurons in the VMPO thatproject to the PVH, which, in turn, is in a position tomodulate autonomic circuitry mediating thermogenesis. Acomplementary mechanism can be forwarded from ourpresent findings of cells expressing EP3R mRNA withinthe nucleus raphe pallidus, a key central site for control-ling sympathetic outflow to brown adipose tissue (Morri-son et al., 1999). Neurons within this region are tonicallyinhibited via local GABAergic interneurons (Morrison etal., 1999), and PGE2-mediated disinhibition of these cellscould be speculated to result in increased thermogenesis.Moreover, recent findings have shown that i.c.v. injectionof an EP3R-specific agonist results in elevated body tem-perature (Parrott and Vellucci, 1996) and mice withgenomic deletions of the EP3R, but not those of the EP1and -2 receptors, fail to elicit a febrile response to PGE2,IL-1b, and lipopolysaccharide (Ushikubi et al., 1998).These findings are consistent with the hypothesis thatcentral PGE2 may trigger fever responses during immunechallenges through EP3R-dependent mechanisms.
Prostaglandins as regulators of sleep-wake states.
The somnogenic effect of IL-1 (for review see Krueger andMajde, 1994) is believed to be under the control of prosta-glandins (in particular prostaglandin D2; PGD2) actingwithin the VLPO. Local infusions of IL-1 into the sub-arachnoid space underlying the ventral surface of the ratVLPO induces slow wave sleep, an effect that can bemimicked by local infusion of PGD2 and abrogated bycoadministration of COX inhibitors (Scammell et al.,1998a; Terao et al., 1998). Neurons in the VLPO are acti-vated during sleep (Sherin et al., 1996) or following sub-arachnoid infusions of the somnogen PGD2 (Scammell etal., 1998a) and project to the tuberomammilary nucleus,the dorsal and median raphe nuclei, and LC, all of whichare additional sites believed to be nodal points in sleep-wake regulation (Sherin et al., 1998). Our present findingsof prominent expression of EP3R mRNA-expressing cellsin preoptic areas overlapping the VLPO as well as withinthe tuberomammilary nucleus, LC, and the dorsal andmedian raphe nuclei provide strong indications that cen-tral PGE2 may influence on sleep-wake states at multiplesites, possibly by modulating the somnogenic effects ofPGD2. Indeed, PGE and PGD have long been implicatedas opposing factors in balancing sleep and arousal(Hayaishi, 1991).
The role of prostaglandins in pain transmission.
Prostaglandins participate in the processing of nociceptivestimuli as evidenced by the analgesic effects of prostaglan-din synthesis inhibitors such as indomethacin and aspirin(Vane, 1971; Ferreira, 1972; Malmberg and Yaksh, 1992).PGE2 sensitizes nociceptors in the periphery (reviewed inDavies et al., 1984), and modulates nociceptive processing
at the level of the spinal cord (Uda et al., 1990; Minami etal., 1994) and the brain (Poddubiuk, 1976; Oka et al.,1994; Hosoi et al., 1997). The nociceptive effect of centrallyadministered PGE2 is dose dependent, causing hyperalge-sia at low, non-pyrogenic, doses and analgesia at high,pyrogenic ones (Oka et al., 1994). Moreover, i.c.v. or intra-preoptic microinjections of an EP3R-specific agonist alsoinduce hyperalgesia, indicating that PGE2 induces hyper-algesia in an EP3R-dependent manner (Oka et al., 1994;Hosoi et al., 1997). Our data demonstrate EP3R mRNAexpression both in nuclei involved in ascending nocicep-tive pathways and descending modulatory pathways (Wil-lis and Westlund, 1997). Cells expressing EP3R mRNAare, for instance, localized in the midline and intralaminarthalamic nuclei, which receive a spinothalamic input (Wil-lis and Westlund, 1997). EP3R mRNA is also expressed innuclei of the spinoreticular nociceptive pathway, includingthe LC, the PB, and the VLM (Willis and Westlund, 1997).Moreover, a recent study has localized EP3R expression tosubregions of the lateral PB that process ascending noci-ceptive information (Engblom et al., 2000). Expression ofEP3R mRNA in cells of the VLM, the dorsal raphe nu-cleus, and the nucleus raphe magnus, key structures inthe descending analgesia pathways from the periaqueduc-tal gray (Willis and Westlund, 1997), indicates that thesemay comprise additional sites where PGE2 may modulatepain perception via the EP3R. These results, along withcomplementary findings of EP3R expression in dorsal rootganglia (Sugimoto et al., 1994) and EP3R-like immunore-activity in lamina 1 and 2 of the spinal cord (Beiche et al.,1998), are in consistent with the hypothesis that PGE2may modulate pain transmission during inflammatory orinfectious conditions in an EP3R-dependent manner.
In conclusion, we have demonstrated that mRNA encod-ing the EP3 subtype of the PGE2 receptor is widely ex-pressed by neuron-like cells throughout the rat brain. Theanatomical distribution of cells expressing this receptorprovides clues as to the mechanisms by which PGE2,which is released from perivascular brain cells followinginflammatory insults, may trigger neural acute phase re-sponses. We exemplify this by demonstrating that theEP3R is expressed by IL-1–responsive neurons in theMPO, VLM, and the NTS. This finding supports our hy-pothesis that circulating IL-1b may come to activate CRH-producing neurons in the hypothalamus via a mechanisminvolving PGE2 stimulation of catecholaminergic neuronsin the caudal medulla. In addition, intense expression ofthe receptor within cell groups, known to regulate painperception, brown adipose tissue, and sleep-wake states,suggests a PGE2/EP3R-mediated regulation of hyperalge-sia, fever, and increased sleep episodes following exposureto immunological aggressors.
ACKNOWLEDGMENTS
We thank Dr. S. Gillis, Immunex Research and Devel-opment Corporation, Seattle, Washington, for generouslyproviding the preparation of IL-1b.
LITERATURE CITEDAbercrombie M. 1946. Estimation of nuclear population from microtome
sections. Anat Rec 94:239–247.Bandler R, Shipley MT. 1994. Columnar organization in the midbrain
17PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS

periaqueductal gray: modules for emotional expression? Trends Neu-rosci 17:379–389.
Beiche F, Klein T, Nusing R, Neuhuber W, Goppelt-Struebe M. 1998.Localization of cyclooxygenase-2 and prostaglandin E2 receptor EP3 inthe rat lumbar spinal cord. J Neuroimmunol 89:26–34.
Berkenbosch F, van Oers J, del Rey A, Tilders F, Besedovsky H. 1987.Corticotropin-releasing factor-producing neurons in the rat activatedby interleukin-1. Science 238:524–526.
Blatteis CM, Sehic E. 1997. Circulating pyrogen signaling of the brain. Anew working hypothesis. Ann N Y Acad Sci 813:445–447.
Boie Y, Stocco R, Sawyer N, Slipetz DM, Ungrin MD, Neuschafer-Rube F,Puschel GP, Metters KM, Abramovitz M. 1997. Molecular cloning andcharacterization of the four rat prostaglandin E2 prostanoid receptorsubtypes. Eur J Pharmacol 340:227–241.
Cao C, Matsumura K, Yamagata K, Watanabe Y. 1996. Endothelial cells ofthe rat brain vasculature express cyclooxygenase-2 mRNA in responseto systemic interleukin-1 beta: a possible site of prostaglandin synthe-sis responsible for fever. Brain Res 733:263–272.
Chan RK, Brown ER, Ericsson A, Kovacs KJ, Sawchenko PE. 1993. Acomparison of two immediate-early genes, c-fos and NGFI-B, as mark-ers for functional activation in stress-related neuroendocrine circuitry.J Neurosci 13:5126–5138.
Cunningham E Jr, Wada E, Carter DB, Tracey DE, Battey JF, De SouzaEB. 1992. In situ histochemical localization of type I interleukin-1receptor messenger RNA in the central nervous system, pituitary, andadrenal gland of the mouse. J Neurosci 12:1101–1114.
Davies P, Bailey PJ, Goldenberg MM, Ford-Hutchinson AW. 1984. The roleof arachidonic acid oxygenation products in pain and inflammation.Annu Rev Immunol 2:335–357.
Dinarello CA. 1991. Interleukin-1 and interleukin-1 antagonism. Blood77:1627–1652.
Elmquist JK, Saper CB. 1996. Activation of neurons projecting to theparaventricular hypothalamic nucleus by intravenous lipopolysaccha-ride. J Comp Neurol 374:315–331.
Engblom D, Ek M, Hallbeck M, Ericsson-Dahlstrand A, Blomqvist A. 2000.Distributions of prostaglandin EP3 and EP4 receptor mRNA in the ratparabrachial nucleus. Neurosci Lett 281:163–166.
Ericsson A, Sawchenko PE. 1993. c-fos–based functional mapping of cen-tral pathways subserving the effects of interleukin-1 on thehypothalamo-pituitary-adrenal axis. In: DeSouza EB, editor. Methodsin neuroscience. The neurobiology of cytokines. New York: AcademicPress, p 155–171.
Ericsson A, Kovacs KJ, Sawchenko PE. 1994. A functional anatomicalanalysis of central pathways subserving the effects of interleukin-1 onstress-related neuroendocrine neurons. J Neurosci 14:897–913.
Ericsson A, Liu C, Hart RP, Sawchenko PE. 1995. Type 1 interleukin-1receptor in the rat brain: distribution, regulation, and relationship tosites of IL-1–induced cellular activation. J Comp Neurol 361:681–698.
Ericsson A, Arias C, Sawchenko PE. 1997. Evidence for an intramedullaryprostaglandin-dependent mechanism in the activation of stress-relatedneuroendocrine circuitry by intravenous interleukin-1. J Neurosci 17:7166–7179.
Ferreira SH. 1972. Prostaglandins, aspirin-like drugs and analgesia. Na-ture New Biol 240:200–203.
Fulwiler CE, Saper CB. 1984. Subnuclear organization of the efferentconnections of the parabrachial nucleus in the rat. Brain Res 319:229–259.
Hayaishi O. 1991. Molecular mechanisms of sleep-wake regulation: roles ofprostaglandins D2 and E2. Faseb J 5:2575–2581.
Hosoi M, Oka T, Hori T. 1997. Prostaglandin E receptor EP3 subtype isinvolved in thermal hyperalgesia through its actions in the preoptichypothalamus and the diagonal band of Broca in rats. Pain 71:303–311.
Katsuura G, Gottschall PE, Dahl RR, Arimura A. 1988. Adrenocortico-tropin release induced by intracerebroventricular injection of recombi-nant human interleukin-1 in rats: possible involvement of prostaglan-din. Endocrinology 122:1773–1779.
Katsuura G, Arimura A, Koves K, Gottschall PE. 1990. Involvement oforganum vasculosum of lamina terminalis and preoptic area ininterleukin-1 beta-induced ACTH release. Am J Physiol 258:E163–171.
Krueger JM, Majde JA. 1994. Microbial products and cytokines in sleepand fever regulation. Crit Rev Immunol 14:355–379.
Lacroix S, Rivest S. 1998. Effect of acute systemic inflammatory responseand cytokines on the transcription of the genes encoding cyclooxygen-ase enzymes (COX-1 and COX-2) in the rat brain. J Neurochem 70:452–466.
Malmberg AB, Yaksh TL. 1992. Hyperalgesia mediated by spinal gluta-mate or substance P receptor blocked by spinal cyclooxygenase inhibi-tion. Science 257:1276–1279.
Matsumura K, Watanabe Y, Imai-Matsumura K, Connolly M, Koyama Y,Onoe H, Watanabe Y. 1992. Mapping of prostaglandin E2 binding sitesin rat brain using quantitative autoradiography. Brain Res 581:292–298.
Matsumura K, Cao C, Watanabe Y, Watanabe Y. 1998. Prostaglandinsystem in the brain: sites of biosynthesis and sites of action undernormal and hyperthermic states. Prog Brain Res 115:275–295.
Minami T, Nishihara I, Uda R, Ito S, Hyodo M, Hayaishi O. 1994. Char-acterization of EP-receptor subtypes involved in allodynia and hyper-algesia induced by intrathecal administration of prostaglandin E2 tomice. Br J Pharmacol 112:735–740.
Morimoto A, Murakami N, Nakamori T, Sakata Y, Watanabe T. 1989.Possible involvement of prostaglandin E in development of ACTHresponse in rats induced by human recombinant interleukin-1.J Physiol 411:245–256.
Morrison SF, Sved AF, Passerin AM. 1999. GABA-mediated inhibition ofraphe pallidus neurons regulates sympathetic outflow to brown adiposetissue. Am J Physiol 276:R290–297.
Murakami N, Watanabe T. 1989. Activation of ACTH release is mediatedby the same molecule as the final mediator, PGE2, of febrile responsein rats. Brain Res 478:171–174.
Nakamura K, Kaneko T, Yamashita Y, Hasegawa H, Katoh H, Negishi M.2000. Immunohistochemical localization of prostaglandin EP3 receptorin the rat nervous system. J Comp Neurol 421:543–569.
Nakano K, Okugawa K, Hayashi H, Abe S, Sohmura Y, Tsuboi T. 1988.Establishment of dye-uptake method (A375 assay) for quantitativemeasurement of IL-1: correlation with LAF assay. Dev Biol Stand69:93–101.
Negishi M, Sugimoto Y, Ichikawa A. 1995. Molecular mechanisms of di-verse actions of prostanoid receptors. Biochim Biophys Acta 1259:109–119.
Oka T, Aou S, Hori T. 1994. Intracerebroventricular injection of prosta-glandin E2 induces thermal hyperalgesia in rats: the possible involve-ment of EP3 receptors. Brain Res 663:287–292.
Parrott RF, Vellucci SV. 1996. Effects of centrally administered prosta-glandin EP receptor agonists on febrile and adrenocortical responses inthe prepubertal pig. Brain Res Bull 41:97–103.
Poddubiuk ZM. 1976. A comparison of the central actions of prostaglandinsA1, E1, E2, F1alpha, and F2alpha in the rat. I. Behavioral, antinoci-ceptive and anticonvulsant actions of intraventricular prostaglandinsin the rat. Psychopharmacology 50:89–94.
Quan N, Whiteside M, Kim L, Herkenham M. 1997. Induction of inhibitoryfactor kappaBalpha mRNA in the central nervous system after periph-eral lipopolysaccharide administration: an in situ hybridization histo-chemistry study in the rat. Proc Natl Acad Sci USA 94:10985–10990.
Rassnick S, Zhou D, Rabin BS. 1995. Central administration of prostaglan-din E2 suppresses in vitro cellular immune responses. Am J Physiol269:R92–97.
Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W. 1987. Interleukin-1stimulates the secretion of hypothalamic corticotropin-releasing factor.Science 238:522–524.
Scammell TE, Elmquist JK, Griffin JD, Saper CB. 1996. Ventromedialpreoptic prostaglandin E2 activates fever-producing autonomic path-ways. J Neurosci 16:6246–6254.
Scammell T, Gerashchenko D, Urade Y, Onoe H, Saper C, Hayaishi O.1998a. Activation of ventrolateral preoptic neurons by the somnogenprostaglandin D2. Proc Natl Acad Sci USA 95:7754–7759.
Scammell TE, Griffin JD, Elmquist JK, Saper CB. 1998b. Microinjection ofa cyclooxygenase inhibitor into the anteroventral preoptic region at-tenuates LPS fever. Am J Physiol 274:R783–789.
Sherin JE, Shiromani PJ, McCarley RW, Saper CB. 1996. Activation ofventrolateral preoptic neurons during sleep. Science 271:216–219.
Sherin JE, Elmquist JK, Torrealba F, Saper CB. 1998. Innervation ofhistaminergic tuberomammillary neurons by GABAergic and galanin-ergic neurons in the ventrolateral preoptic nucleus of the rat. J Neu-rosci 18:4705–4721.
Simerly RB, Swanson LW, Gorski RA. 1984. Demonstration of a sexualdimorphism in the distribution of serotonin-immunoreactive fibers inthe medial preoptic nucleus of the rat. J Comp Neurol 225:151–166.
Simmons DM, Arriza JL, Swanson LW. 1989. A complete protocol for insitu hybridization of messenger RNAs in brain and other tissues withradiolabeled single-stranded RNA probes. J Histotechnol 12:169–181.
18 M. EK ET AL.

Sugimoto Y, Namba T, Honda A, Hayashi Y, Negishi M, Ichikawa A,Narumiya S. 1992. Cloning and expression of a cDNA for mouse pros-taglandin E receptor EP3 subtype. J Biol Chem 267:6463–6466.
Sugimoto Y, Shigemoto R, Namba T, Negishi M, Mizuno N, Narumiya S,Ichikawa A. 1994. Distribution of the messenger RNA for the prosta-glandin E receptor subtype EP3 in the mouse nervous system. Neuro-science 62:919–928.
Swanson LW. 1992. Brain maps: structure of the rat brain. Amsterdam:Elsevier Science Publishers BV.
Takeuchi K, Abe T, Takahashi N, Abe K. 1993. Molecular cloning andintrarenal localization of rat prostaglandin E2 receptor EP3 subtype.Biochem Biophys Res Commun 194:885–891.
Terao A, Matsumura H, Saito M. 1998. Interleukin-1 induces slow-wavesleep at the prostaglandin D2-sensitive sleep-promoting zone in the ratbrain. J Neurosci 18:6599–6607.
Uda R, Horiguchi S, Ito S, Hyodo M, Hayaishi O. 1990. Nociceptive effectsinduced by intrathecal administration of prostaglandin D2, E2, or F2alpha to conscious mice. Brain Res 510:26–32.
Ushikubi F, Segi F, Sugimoto Y, Murata T, Matsuoka T, Kobayashi T,Hizaki H, Tuboi K, Katsuyama M, Ichikawa A, Tanaka T, Yoshida N,Narumiya S. 1998. Impaired febrile response in mice lacking the pros-taglandin E receptor subtype EP3. Nature 395:281–284.
Van Dam AM, De Vries HE, Kuiper J, Zijlstra FJ, De Boer AG, Tilders FJ,
Berkenbosch F. 1996. Interleukin-1 receptors on rat brain endothelialcells: a role in neuroimmune interaction? Faseb J 10:351–356.
Vane JR. 1971. Inhibition of prostaglandin synthesis as a mechanism ofaction for aspirin-like drugs. Nature New Biol 231:232–235.
Watanabe T, Morimoto A, Sakata Y, Murakami N. 1990. ACTH responseinduced by interleukin-1 is mediated by CRF secretion stimulated byhypothalamic PGE. Experientia 46:481–484.
Watts AG, Swanson LW. 1989. The combination of in situ hybridization withimmunohistochemistry and retrograde tract-tracing. In: Conn PM, editor.Methods in neurosciences. New York: Academic Press. p 127–136.
Willis WD, Westlund KN. 1997. Neuroanatomy of the pain system and ofthe pathways that modulate pain. J Clin Neurophysiol 14:2–31.
Yabuuchi K, Minami M, Katsumata S, Satoh M. 1994. Localization of typeI interleukin-1 receptor mRNA in the rat brain. Brain Res Mol BrainRes 27:27–36.
Zhang J, Rivest S. 1999. Distribution, regulation and colocalization of thegenes encoding the EP2- and EP4-PGE2 receptors in the rat brain andneuronal responses to systemic inflammation. Eur J Neurosci 11:2651–2668.
Zhang J, Rivest S. 2000. A functional analysis of EP4 receptor-expressingneurons in mediating the action of prostaglandin E-2 within specificnuclei of the brain in response to circulating interleukin-1 beta. J Neu-rochem 74:2134–2145.
19PGE2 RECEPTORS ON IL-1–SENSITIVE RAT CNS NEURONS
Related Documents











