DISTRIBUTION AND POPULATION SYSTEMS OF GREEN FROGS (Pelophylax esculentus COMPLEX) IN KALININGRAD OBLAST’, RUSSIA (BALTIC SEA REGION) S. N. Litvinchuk, 1 L. J. Borkin, 2 Y. S. Litvinchuk, 3 and J. M. Rosanov 1 Submitted July 30, 2015. The occurrence of three green frog taxa (Pelophylax lessonae, P. ridibundus, and P. esculentus) in Kaliningrad oblast’ of Russia was evidenced by DNA flow cytometry analysis. Almost all frogs were diploid. Only one young specimen of P. esculentus proved to be a triploid with two genomes of P. ridibundus and one genome of P. lessonae. The triploidy occurrence among hybrids was equal to 1.1%. Two species (P. lessonae and P. esculen- tus) seem to be distributed over all territory of Kaliningrad oblast’. The third species (P. ridibundus) predomi- nantly inhabits the western part of the region, where it can be found in brackish water bodies (4.7 – 6.7‰). Six population systems were revealed; with a single species (R, L, and, probably, E) and with mixed species popula- tions (L-E, R-E, and R-E-L). The study of P. esculentus sperm in males taken from the R-E, L-E, and R-E-L sys- tems demonstrated that all of them produced gametes with the ridibundus genome. Keywords: Ranidae; DNA flow cytometry; genome size; hybrids; triploidy; water salinity tolerance; color devia- tions. INTRODUCTION European green (or water) frogs of the Pelophylax esculentus complex, formerly assigned to the genus Rana Linnaeus, 1758, demonstrate unusual genetic phenomena associated with hybridization, clonality and polyploidy (Plötner, 2005). The complex consists of three species (Fig. 1). Pelophylax esculentus (Linnaeus, 1758) has a hybrid origin, and its parental species are P. lessonae (Camerano, 1882) and P. ridibundus (Pallas, 1771). Hybrid frogs are characterized by special mode of clonal inheritance. As a rule, during gametogenesis in diploid hybrid, the entire genome of a parental species is selec- tively eliminated, and the hybrid passes the genome of another parental species to progeny without crossing- over or segregation. Hybridogenetic frogs (P. esculentus) are widely known across temperate Europe from France in the west to Volga River in the east (Günther, 1997; Borkin et al., 2003). Diploid hybrids occur in almost all parts of the species’ range. Triploid and, rarely, tetraploid hybrids have been recorded in some populations distributed in the western and central parts of Europe, as well as in the Severskiy Donets River drainage in eastern Ukraine and adjacent Russia (Günther, 1975; Mikulícek and Kotlík, 2001; Ogielska et al., 2001; Rybacki and Berger, 2001; Borkin et al., 2004, 2006; Plötner, 2005; Jakob, 2007). Unfortunately, up to present day, detailed information about the distribution of species of the P. esculentus com- plex remains insufficient and fragmented. Kaliningrad oblast’ is the westernmost exclave part of Russian Federation located in the Baltic region be- tween Poland and Lithuania, in the northern part of his- torical East Prussia. The first records of green frogs in this region seemed to be published by P. Muhling (1898). He studied hel- minths of “Rana esculenta” from Rossitten (now Ryba- chiy) and “Rana esculenta var. ridibunda” from Pillau (now Baltiysk). Later, based on large size of some speci- mens, O. le Roi (1903) suggested that green frogs from Rossitten might belong to “Rana esculenta var. ridibun- da.” F. Pagast (1941) mentioned records of “Rana ridi- bunda” from Pregel (now Pregolya) River in the vicini- ties of Königsberg (now Kaliningrad), and Frischen (= Vistula) and Kurischen (= Kuronian) lagoons. 1026-2296/2015/2203-0188 © 2015 Folium Publishing Company Russian Journal of Herpetology Vol. 22, No. 3, 2015, pp. 188 – 196 1 Institute of Cytology, Russian Academy of Sciences, Tikhoretsky pr. 4, St. Petersburg 194064, Russia; e-mail: [email protected] 2 Zoological Institute, Russian Academy of Sciences, Universitetskaya nab. 1, St. Petersburg 199034, Russia. 3 St. Petersburg State University, Universitetskaya nab. 7, St. Peters- burg 199034, Russia.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DISTRIBUTION AND POPULATION SYSTEMS
OF GREEN FROGS (Pelophylax esculentus COMPLEX)
IN KALININGRAD OBLAST’, RUSSIA (BALTIC SEA REGION)
S. N. Litvinchuk,1
L. J. Borkin,2
Y. S. Litvinchuk,3
and J. M. Rosanov1
Submitted July 30, 2015.
The occurrence of three green frog taxa (Pelophylax lessonae, P. ridibundus, and P. esculentus) in Kaliningrad
oblast’ of Russia was evidenced by DNA flow cytometry analysis. Almost all frogs were diploid. Only one young
specimen of P. esculentus proved to be a triploid with two genomes of P. ridibundus and one genome of
P. lessonae. The triploidy occurrence among hybrids was equal to 1.1%. Two species (P. lessonae and P. esculen-
tus) seem to be distributed over all territory of Kaliningrad oblast’. The third species (P. ridibundus) predomi-
nantly inhabits the western part of the region, where it can be found in brackish water bodies (4.7 – 6.7‰). Six
population systems were revealed; with a single species (R, L, and, probably, E) and with mixed species popula-
tions (L-E, R-E, and R-E-L). The study of P. esculentus sperm in males taken from the R-E, L-E, and R-E-L sys-
tems demonstrated that all of them produced gametes with the ridibundus genome.
Keywords: Ranidae; DNA flow cytometry; genome size; hybrids; triploidy; water salinity tolerance; color devia-
tions.
INTRODUCTION
European green (or water) frogs of the Pelophylax
esculentus complex, formerly assigned to the genus Rana
Linnaeus, 1758, demonstrate unusual genetic phenomena
associated with hybridization, clonality and polyploidy
(Plötner, 2005). The complex consists of three species
(Fig. 1). Pelophylax esculentus (Linnaeus, 1758) has a
hybrid origin, and its parental species are P. lessonae
(Camerano, 1882) and P. ridibundus (Pallas, 1771).
Hybrid frogs are characterized by special mode of clonal
inheritance. As a rule, during gametogenesis in diploid
hybrid, the entire genome of a parental species is selec-
tively eliminated, and the hybrid passes the genome of
another parental species to progeny without crossing-
over or segregation.
Hybridogenetic frogs (P. esculentus) are widely
known across temperate Europe from France in the west
to Volga River in the east (Günther, 1997; Borkin et al.,
2003). Diploid hybrids occur in almost all parts of the
species’ range. Triploid and, rarely, tetraploid hybrids
have been recorded in some populations distributed in the
western and central parts of Europe, as well as in the
Severskiy Donets River drainage in eastern Ukraine and
adjacent Russia (Günther, 1975; Mikulícek and Kotlík,
2001; Ogielska et al., 2001; Rybacki and Berger, 2001;
Borkin et al., 2004, 2006; Plötner, 2005; Jakob, 2007).
Unfortunately, up to present day, detailed information
about the distribution of species of the P. esculentus com-
plex remains insufficient and fragmented.
Kaliningrad oblast’ is the westernmost exclave part
of Russian Federation located in the Baltic region be-
tween Poland and Lithuania, in the northern part of his-
torical East Prussia.
The first records of green frogs in this region seemed
to be published by P. Muhling (1898). He studied hel-
minths of “Rana esculenta” from Rossitten (now Ryba-
chiy) and “Rana esculenta var. ridibunda” from Pillau
(now Baltiysk). Later, based on large size of some speci-
mens, O. le Roi (1903) suggested that green frogs from
Rossitten might belong to “Rana esculenta var. ridibun-
da.” F. Pagast (1941) mentioned records of “Rana ridi-
bunda” from Pregel (now Pregolya) River in the vicini-
ties of Königsberg (now Kaliningrad), and Frischen
(= Vistula) and Kurischen (= Kuronian) lagoons.
1026-2296�2015�2203-0188 © 2015 Folium Publishing Company
Russian Journal of Herpetology Vol. 22, No. 3, 2015, pp. 188 – 196
1Institute of Cytology, Russian Academy of Sciences, Tikhoretsky pr.
4, St. Petersburg 194064, Russia; e-mail: [email protected]
2Zoological Institute, Russian Academy of Sciences, Universitetskaya
nab. 1, St. Petersburg 199034, Russia.
3St. Petersburg State University, Universitetskaya nab. 7, St. Peters-
burg 199034, Russia.

Later, several Russian authors published some re-
cords of green frogs in Kaliningrad oblast’ (Alimov et al.,
1976; Litvinchuk, 1996; Grishanov and Belyakov, 2000;
Aleksandrova, 2003; Lada, 2012; Kuzmin, 2013; Alekse-
ev, 2014; Sokolov, 2014).
Usually, in many previous papers, the identification
of species was based on external morphological charac-
ters. However, these species of the P. esculentus complex
are relatively similar and, moreover, diagnostic features
vary geographically (Nekrasova and Morozov-Leonov,
2001; Pisanets, 2007). Therefore, reliable molecular or
cytological markers should be used for their correct iden-
tification (Plötner, 2005).
Using biochemical methods, we started to study the
species composition of green frogs in Kaliningrad oblast’
in 1979 (Borkin et al., 1986, 2004; Borkin, 2001; Litvin-
chuk et al., 2007).
The aim of present paper was to summarize our data
on geographic distribution and the composition of popu-
lation systems of green frogs in Kaliningrad oblast’,
based on application of DNA flow cytometry.
MATERIAL AND METHODS
Green frogs (228 individuals) were collected in
1988 – 2014 in 19 localities of Kaliningrad oblast’
(Table 1; Fig. 2). The content of DNA per nucleus (or ge-
nome size) in erythrocytes was measured (in pg) by DNA
flow cytometry (Table 1). This method allows to identify
each individual reliably in terms of parental species,
hybridity, ploidy level as well as genome composition in
polyploids. Erythrocytes of Rana temporaria Linnaeus,
1758 collected in north-western Russia were used as a
reference standard. The details of technique have been
published previously (e.g., Vinogradov et al., 1990; Bor-
kin et al. 2001).
The 53 individuals collected in 1988 – 1990 were
examined by A. E. Vinogradov (St. Petersburg) with use
of DNA flow cytometry. The staining conditions and ac-
Distribution of Green Frogs in Kaliningrad Oblast’ 189
Fig. 1. Pelophylax lessonae: male (A) and female (B), the village Romanovo; P. esculentus female, Romanovo (C); P. ridibundus: female, the
Southern park, Kaliningrad (D).

curacy of measurements in these studies were different in
comparison with later studies performed by J. M. Rosa-
nov. Therefore, genome size estimations of 1988 – 1990
were not included in Table 2 and Fig. 3.
Genome types in sperm from testes cell suspensions
in males of P. esculentus were identified by DNA flow
cytometry (two individuals from Strel’nya) or by means
of protein electrophoresis (eight individuals from Ryba-
190 S. N. Litvinchuk et al.
TABLE 1. Distribution of Various Population Systems (PS) of Green Frogs and Frequency of Individuals of Each Species per Locality (in %),
Based on Application of DNA Flow Cytometry*
N Locality Year PS Latitude Longitude n les esc rid
1 Baltiysk 2002, 2008, 2014 R 54.6355 19.8739 10 — — 100
2 Mamonovo 2014 R 54.4490 19.9520 5 — — 100
3 Ladushkin 2011 L-E 54.5667 20.1833 11 27 73 —
4 Ul’yanovka 2014 L 54.6000 20.2050 1 100 — —
5 Ladygino 2014 R 54.6080 20.2040 1 — — 100
6 Ushakovo 2014 R 54.6120 20.2430 1 — — 100
7 Southern park, Kaliningrad 2014 R 54.6910 20.5120 4 — — 100
8 Strel’nya 1989, 1990, 2002 L-E 54.4667 20.6000 57 61 39 —
9 Bagrationovsk 1989, 1990, 2008, 2014 L-E 54.3833 20.6167 31 77 23 —
10 Svetlogorsk 2014 L-E 54.9360 20.1530 14 7 93 —
11 Pionerskiy 2002 L-E 54.9333 20.2167 10 10 90 —
12 Romanovo 2014 L-E 54.8980 20.2770 13 77 23 —
13 Zelenogradsk 2008 R-E 54.9524 20.4836 9 — 44 56
14 Rybachiy 2008, 2014 R-E 55.1542 20.8354 25 — 60 40
15 Gvardeysk 2014 L 54.6550 21.1040 6 100 — —
16 Sovetsk 2014 R-E-L 55.0940 21.8440 22 5 27 68
17 Nesterov 1999 L-E 54.6333 22.5500 2 50 50 —
18 Botanical garden, Kaliningrad 1989 L-E 54.7377 20.5198 3 33 67 —
19 Svoboda 1988 E 54.5400 21.7300 2 — 100 —
* les, P. lessonae; esc, P. esculentus; rid, P. ridibundus.
Fig. 2. Distribution of green frog population systems in Kaliningrad oblast’. Green circles are the L type, red circles are the R type, blue squares are
the L-E type, black squares are the R-E type, red squares are the R-E-L type; and a green square is the E type. Numbers for localities are given in
Table 1. Localities 20 and 21 are Pravdinskoe and Vyshtinets Lake, respectively.
chiy, Svetlogorsk, and Sovetsk). In the second case, we
used Lactate dehydrogenase previously found useful in
discriminating P. ridibundus and P. lessonae genomes
(Uzzell and Berger, 1975; Uzzell et al., 1980; Mikulíèek
et al., 2015).
In water bodies studied in 2014, we measured salinity
with use of tester Hanna HI98312 (Dist 6).
RESULTS AND DISCUSSION
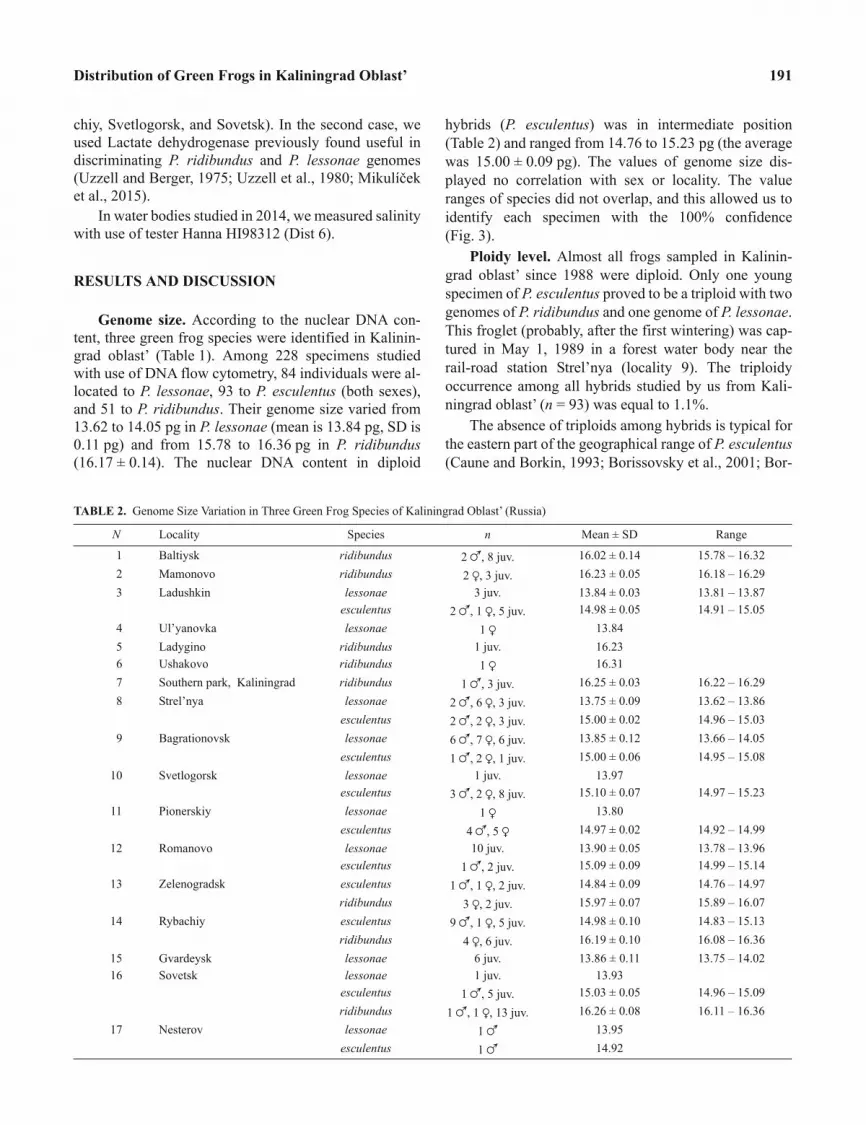
Genome size. According to the nuclear DNA con-
tent, three green frog species were identified in Kalinin-
grad oblast’ (Table 1). Among 228 specimens studied
with use of DNA flow cytometry, 84 individuals were al-
located to P. lessonae, 93 to P. esculentus (both sexes),
and 51 to P. ridibundus. Their genome size varied from
13.62 to 14.05 pg in P. lessonae (mean is 13.84 pg, SD is
0.11 pg) and from 15.78 to 16.36 pg in P. ridibundus
(16.17 ± 0.14). The nuclear DNA content in diploid
hybrids (P. esculentus) was in intermediate position
(Table 2) and ranged from 14.76 to 15.23 pg (the average
was 15.00 ± 0.09 pg). The values of genome size dis-
played no correlation with sex or locality. The value
ranges of species did not overlap, and this allowed us to
identify each specimen with the 100% confidence
(Fig. 3).
Ploidy level. Almost all frogs sampled in Kalinin-
grad oblast’ since 1988 were diploid. Only one young
specimen of P. esculentus proved to be a triploid with two
genomes of P. ridibundus and one genome of P. lessonae.
This froglet (probably, after the first wintering) was cap-
tured in May 1, 1989 in a forest water body near the
rail-road station Strel’nya (locality 9). The triploidy
occurrence among all hybrids studied by us from Kali-
ningrad oblast’ (n = 93) was equal to 1.1%.
The absence of triploids among hybrids is typical for
the eastern part of the geographical range of P. esculentus
(Caune and Borkin, 1993; Borissovsky et al., 2001; Bor-
Distribution of Green Frogs in Kaliningrad Oblast’ 191
TABLE 2. Genome Size Variation in Three Green Frog Species of Kaliningrad Oblast’ (Russia)
N Locality Species n Mean ± SD Range
1 Baltiysk ridibundus 2 �, 8 juv. 16.02 ± 0.14 15.78 – 16.32
2 Mamonovo ridibundus 2 �, 3 juv. 16.23 ± 0.05 16.18 – 16.29
3 Ladushkin lessonae 3 juv. 13.84 ± 0.03 13.81 – 13.87
esculentus 2 �, 1 �, 5 juv. 14.98 ± 0.05 14.91 – 15.05
4 Ul’yanovka lessonae 1 � 13.84
5 Ladygino ridibundus 1 juv. 16.23
6 Ushakovo ridibundus 1 � 16.31
7 Southern park, Kaliningrad ridibundus 1 �, 3 juv. 16.25 ± 0.03 16.22 – 16.29
8 Strel’nya lessonae 2 �, 6 �, 3 juv. 13.75 ± 0.09 13.62 – 13.86
esculentus 2 �, 2 �, 3 juv. 15.00 ± 0.02 14.96 – 15.03
9 Bagrationovsk lessonae 6 �, 7 �, 6 juv. 13.85 ± 0.12 13.66 – 14.05
esculentus 1 �, 2 �, 1 juv. 15.00 ± 0.06 14.95 – 15.08
10 Svetlogorsk lessonae 1 juv. 13.97
esculentus 3 �, 2 �, 8 juv. 15.10 ± 0.07 14.97 – 15.23
11 Pionerskiy lessonae 1 � 13.80
esculentus 4 �, 5 � 14.97 ± 0.02 14.92 – 14.99
12 Romanovo lessonae 10 juv. 13.90 ± 0.05 13.78 – 13.96
esculentus 1 �, 2 juv. 15.09 ± 0.09 14.99 – 15.14
13 Zelenogradsk esculentus 1 �, 1 �, 2 juv. 14.84 ± 0.09 14.76 – 14.97
ridibundus 3 �, 2 juv. 15.97 ± 0.07 15.89 – 16.07
14 Rybachiy esculentus 9 �, 1 �, 5 juv. 14.98 ± 0.10 14.83 – 15.13
ridibundus 4 �, 6 juv. 16.19 ± 0.10 16.08 – 16.36
15 Gvardeysk lessonae 6 juv. 13.86 ± 0.11 13.75 – 14.02
16 Sovetsk lessonae 1 juv. 13.93
esculentus 1 �, 5 juv. 15.03 ± 0.05 14.96 – 15.09
ridibundus 1 �, 1 �, 13 juv. 16.26 ± 0.08 16.11 – 16.36
17 Nesterov lessonae 1 � 13.95
esculentus 1 � 14.92

kin et al., 2002, 2003, 2004, 2005, 2006, 2008; Ruchin et
al., 2005, 2010; Zamaletdinov et al., 2005; Lada et al.,
2009, 2011a, 2011b; Svinin et al., 2013, 2015; our data).
Moreover, among 707 individuals of P. esculentus from
adjacent territory of Poland triploids were lacking.
However, in the vicinities of Gdansk (Poland) their share
provided 21% among 162 P. esculentus (Rybatsky and
Berger, 2001).
Species distribution. According to previously pub-
lished maps (Kuzmin, 2013: supplementary disc), the
ranges of all three species of P. esculentus complex cover
the territory of Kaliningrad oblast’. Based on the map for
P. ridibundus, we could suggest that the northern limit of
native range of the species coincides with the Nemunas
River valley, which forms the northern border of
Kaliningrad oblast’. Introduced populations of the spe-
cies are known in more northern territories in Latvia, Es-
tonia, Finland, and Leningradskaya oblast’ of Russia
(Caune, 1987; Milto, 2007; Hoogesteger et al., 2013; our
data). Interestingly, P. ridibundus is lacking in adjacent
regions of Poland (Rybacki and Berger, 1994, 2001;
Glowaciñski and Rafiñski, 2003).
In addition to DNA flow cytometry data, some previ-
ous records could be used in analysis of the distribution
of green frog species in Kaliningrad oblast’ as well. For
example, using protein electrophoresis (serum albumins)
L. J. Borkin et al. (1986) revealed one individual of P. ri-
dibundus and four P. esculentus in Pravdinskoe fish farm
(54°25�48� N 21°1�12�� E; locality 20 in Fig. 2). In July
15, 1999, we visited the western shore of Vyshtinets Lake
(54°26�24�� N 22°42� E; locality 21 in Fig. 2) and col-
lected there three green frogs, which were assigned to
P. lessonae according to morphological characters. One
specimen of them is kept in herpetological collections of
the Zoological Institute (ZISP.6696). In July 31, 2002, we
collected a specimen (ZISP.7044) in Pionerskiy settle-
ment (locality 11), which was identified as P. ridibundus.
G. A. Lada (2012) also ascribed this specimen to that spe-
cies. Therefore, taking into consideration genome size
data (Tables 1 and 2), we can consider that this locality
was settled by green frogs of the R-E-L population
system.
Two species (P. lessonae and P. esculentus) seem to
be distributed over all territory of Kaliningrad oblast’.
However, the first species prefers to occupy forested ar-
eas (Fig. 4A), whereas the second was mostly recorded in
border zones (so called ecotones) between forested and
open landscapes (Fig. 4B – D). The third species (P. ridi-
bundus) lives in open and urban landscapes near rivers,
lakes, lagoons, and large reservoirs (Fig. 4C – E). This
species was predominantly found in the western part of
Kaliningrad oblast’, where it inhabits lower parts of Ne-
munas and Pregolya rivers, Pravdinskoe Reservoir, as
well as the shores of Kuronian and Vistula lagoons. Such
landscape differences between these three species were
previously described for other parts of their geographical
ranges in European Russia (Ruchin et al., 2009; Lada et
al., 2011b).
Water salinity. Sometimes, in Kaliningrad oblast’
P. ridibundus can be registered in brackish water bodies.
So, in August 24, 2014, we observed a small population
of this species in the Vistula Spit shore of Baltic Sea
(Baltiysk; locality 1; Fig. 4F), where frogs inhabited
the moat of old fort with water salinity equal to 6.7‰.
In this day, we met there numerous adults only (about
two animals per a meter of waterside). No juveniles and
larvae were found. In contrast, juvenile frogs were abun-
dant in our previous visits to this place (08�02�2002 and
08�22�2008).
The second brakish water population of P. ridibundus
was observed in moats connected with Baltic Sea (salin-
ity 5.8‰ in 08�24�2014) situated in another part of
Baltiysk Town. The third population was revealed in the
shore of Vistula Lagoon near Ladygino village covered
with dense reeds (locality 5; Fig. 3E). Here we found four
adult and a recently metamorphosed individuals of P. ri-
dibundus (salinity 4.7‰).
The maximum water salinity for P. esculentus
(0.8‰) was registered in a small concrete fire pond in the
settlement Rybachiy located in Kuronian Spit (locality
14; Fig. 4C) and for P. lessonae (0.3‰) in desolated
quarry in the Nemunas River valley near the town So-
vetsk (locality 16; Fig. 4D).
Previously, K. D. Milto (2008) mentioned a case of
the breeding of introduced P. ridibundus in the Neva Bay,
Baltic Sea (Leningradskaya oblast’, Russia). However,
the salinity in this water body was low (about 0.1‰).
In Bulgaria, N. Natchev et al. (2011) wrote about the
192 S. N. Litvinchuk et al.
esculentus
lessonae ridibundus
13.5 14 14.5 15 15.5 16 16.5
The nuclear DNA content, pg
Num
ber
of
specim
ens
25
20
15
10
5
0
Fig. 3. The nuclear DNA content variation in three green frog species
from Kaliningrad oblast’.

breeding of the same frog species in Shablenska Tuzla
Lagoon (the western part of Black Sea), with the salinity
2.2 – 2.9‰. In southern Ukraine, I. B. Dotsenko (2006)
recorded a small population of P. ridibundus inhabiting
Khadzhibeyskyi Liman (4.7‰) in the northern part of
Black Sea.
Distribution of Green Frogs in Kaliningrad Oblast’ 193
Fig. 4. Habitats of green frogs in Kaliningrad oblast’: A, Ul’yanovka (Pelophylax lessonae); B, Ladushkin (L-E type); C, Rybachiy (R-E type); D,
Sovetsk (R-E-L type); E, Ladygino; F, Baltiysk (P. ridibundus).

Population systems. In Kaliningrad oblast’, we
found six types of population systems (Fig. 2). Among
21 localities studied by DNA flow cytometry and other
techniques, the L-E systems were most frequent (33%).
The mixed system of the R-E type (14%) and systems
with a single parental species (R type 24% and L type
14%) were also relatively common. Water bodies with all
three species together (R-E-L) were found only twice
(10%). Two specimens of P. esculentus from Svoboda
might belong to so called pure E system, which contains
of hybridogenetic frogs only. However, the sample size
was too small to support this suggestion.
Hybrid gametes. The study of P. esculentus sperm in
four males taken from the R-E system in Rybachiy, three
males from the L-E system in Svetlogorsk, two males
from the L-E system in Strel’nya, and one male from the
R-E-L system in Sovetsk demonstrated that all of them
produced gametes with the ridibundus (R) genome.
Based on genome size data, gametes in two P. esculentus
males from Strel’nya were haploid. This situation is typi-
cal for reproduction of hybrids in L-E systems (Vino-
gradov et al., 1990; Svinin et al., 2013). However, it is
important to note that P. lessonae is quite rare in the
Svetlogorsk population (Table 2). We found only one
metamorphosed froglet of this species among 14 individ-
uals studied. The record of R genome gametes trans-
ferred by a hybrid male in the R-E-L system (Sovetsk)
allow us to suggest that local male P. esculentus mate
with female P. lessonae (Table 2). That gamete transfer in
hybrids is very common for such type of mixed popula-
tions (Borkin et al., 1987; Mezhzherin et al., 2005;
Provust et al., 2013; Svinin et al., 2013, 2015).
The record of R genome only in gametes of four
males from the R-E system (Rybachiy) was unexpected,
because, theoretically, in such system P. esculentus frogs
should eliminate R genome and pass haploid lessonae (L)
gametes for successful reproduction. Some evidences
supporting that were previously published (e.g., Vinogra-
dov et al., 1990; Mezhzherin et al., 2005). However, in
many cases the hybrids in R-E systems provided sperm
with R genome (Berger, 1994; Mezhzherin et al., 2005;
our data). Perhaps, reproduction of P. esculentus in such
system depends on females producing haploid oocytes
with the L genome.
Coloration. In Kaliningrad oblast’ some green frogs
had unusual coloration with blue tint (Fig. 5). We re-
vealed such “blue” animals in two species from three
neighboring localities (numbers 10 – 12 in Fig. 2). In Ro-
manovo among ten juveniles of P. lessonae we found one
individual with such coloration (Fig. 5). In the Svetlo-
gorsk sample, two “blue” animals (male and female)
were among 13 hybrids. Finally, in Pionerskiy nine P. es-
culentus included one specimen with blue coloration.
Previously, such color (perhaps, insufficient quantity of
yellow pigment) was found in P. esculentus in France
(Bogaerts, 2003) and in all three species of the P. esculen-
tus complex in Ukraine (Nekrasova, 2014).
Acknowledgments. L. F. Litvinchuk (St. Petersburg, Rus-
sia) contributed greatly during the field trips. We also thanks to
S. P. Gruodis (Vilnius, Lithuania) and A. E. Vinogradov (St. Pe-
tersburg, Russia) for kind assistance. The work was partially
funded by grants RFBR (15-04-0506815 and 15-29-02546) and
NSH (3553.2014.4).
REFERENCES
Aleksandrova M. B. (2003), Fauna of Amphibians of Kalinin-
grad City. Diploma Thesis, Kaliningrad State University,
Kaliningrad [in Russian].
Alekseev F. E. (2014), “Natural site Vistula Spit,” in: Nature of
Kaliningrad oblast’. Key Natural Sites, Istok, Kaliningrad,
136 – 146 [in Russian].
Alimov A. F., Vinogradova N. B., Golikov A. N., Dol-
nik V. R., Dolnik T. V., Nitsenko A. A., and Paev-
skiy V. A. (1976), Biological Station of Zoological Institute
of Russian Academy of Sciences “Rybachiy,” Izd. Zool. Inst.
AN USSR, Leningrad [in Russian].
Berger L. (1994), “Some peculiar phenomena in European
water frogs,” Zool. Poloniae, 39(3 – 4), 267 – 280.
Bogaerts S. (2003), “Blue ‘green frogs’ (Rana esculenta) and
other herpetofauna on the Drulon estate (dépt. Cher),
France,” Podarcis, 4(1), 1 – 7.
Borissovsky A. G., Borkin L. J., Litvinchuk S. N., and Rosa-
nov J. M. (2001), “Distribution of green frogs (Rana escu-
lenta complex) in Udmurtia,” Vestn. Udm. Univ., 6, 51 – 63
[in Russian].
Borkin L. J. (2001), “Speciation, hybridization, and polyploi-
dy in amphibians of the Palearctic,” in: Ananjeva N. B. et
194 S. N. Litvinchuk et al.
Fig. 5. A “blue” juvenile Pelophylax lessonae, the village Roma-
novka.

al. (eds.), The Problems of Herpetology, Pushchino – Mos-
cow, pp. 46 – 48 [in Russian].
Borkin L. J., Caune I., Pikulik M., and Sokolova T. (1986),
“Distribution and structure of the green frog complex in the
USSR,” in: Roèek Z. (ed.), Studies in Herpetology, Charles
University, Prague, pp. 675 – 678.
Borkin L. J., Vinogradov A. E., Rosanov J. M., and
Tsaune I. A. (1987), “Hemiclonal inheritance in the hybri-
dogenetic complex Rana esculenta: evidence by method of
DNA flow cytometry,” Dokl. AN SSSR, 295(5), 1261 –
1264 [in Russian].
Borkin L. J., Litvinchuk S. N., Rosanov J. M., and Milto K. D.
(2001), “Cryptic speciation in Pelobates fuscus (Anura, Pe-
lobatidae): evidence from DNA flow cytometry,” Amphi-
bia–Reptilia, 22(4), 387 – 396.
Borkin L. J., Litvinchuk S. N., Mannapova E. I., Pes-
tov M. V., and Rosanov J. M. (2002), “The distribution of
green frogs (Rana esculenta complex) in Nizhny Novgorod
Province, central European Russia,” Russ. J. Herpetol.,
9(3), 195 – 207.
Borkin L. J., Litvinchuk S. N., Rosanov J. M., Lada G. A.,
Ruchin A. B., Fayzulin A. I., and Zamaletdinov R. I.
(2003), “Hybridogenetic complex Rana esculenta: would
exist ‘Volga paradox’?” in: Bakiev A. G., Malenev A. L.,
Noskova O. L. (eds.), Materials of the Third Conference of
Volga Herpetologists, Tol’yatti, pp. 7 – 12 [in Russian].
Borkin L. J., Korshunov A. V., Lada G. A., Litvin-
chuk S. N., Rosanov J. M., Shabanov D. A., and Zinen-
ko A. I. (2004), “Mass occurrence of polyploid green frogs
(Rana esculenta complex) in eastern Ukraine,” Russ. J.
Herpetol., 11(3), 194 – 213.
Borkin L. J., Zinenko A. I., Korshunov A. V., Lada G. A.,
Litvinchuk S. N., Rosanov J. M., and Shabanov D. A.
(2005), “Mass polyploidy in Rana esculenta hybridogenic
complex (Ranidae, Anura, Amphibia) in eastern Ukraine,”
in: Pisanets E. (ed.), Proc. of the First Conf. of the Ukrai-
nian Herpetol. Soc., Izd. Zoomuseum NMNH NAS of
Ukraine, Kyiv, pp. 23 – 26 [in Russian].
Borkin L. J., Lada G. A., Litvinchuk S. N., Melnikov D. A.,
and Rosanov J. M. (2006), “The first record of mass tri-
ploidy in hybridogenic green frog Rana esculenta in Russia
(Rostov oblast’),” Russ. J. Herpetol., 13(1), 77 – 82.
Borkin L. J., Bezman-Moseiko O. S., Mazepa G. A., Zinen-
ko A. I., Korshunov A. V., Lada G. A., Shabanov D. A.,
Litvinchuk S. N., and Rozanov J. M. (2008), “On the
southern limit of range of hybrid Rana esculenta (Ranidae,
Anura, Amphibia) in Ukraine and Moldova: DNA flow
cytometry evidence,” Proc. Ukr. Herpetol. Soc., 1, 5 – 10
[in Russian].
Caune I. A. (1987), Systematics and Distribution of Hybrido-
genetic Complex Rana esculenta in the Territory of Latvian
SSR. Ph.D. Thesis, Izd. Zool. Inst. AN SSSR, Leningrad
[in Russian].
Caune I. A. and Borkin L. J. (1993), “A new kind of unisex-
ual-bisexual population systems in European green frogs
(Rana esculenta complex),” in: Rossolimo O. L. (ed.), Hy-
bridization and Species Problem in Vertebrates, Izd. MGU,
Moscow, pp. 34 – 52 (Archives of Zoological Museum,
Moscow State University, 30) [in Russian].
Dotsenko I. B. (2006), “The lake frog (Rana ridibunda) salt-
water populations in vicinities of Odessa,” Zbirnyk Prac’
Zool. Mus. Kyiv, 38, 80 – 83 [in Russian].
Glowaciñski Z. and Rafiñski J. (eds.) (2003), Atlas P³azów i
Gadów Polski. Status — Rozmieszczenie — Ochrona,
Kraków.
Grishanov G. V. and Belyakov V. V. (2000), Land Vertebrates
of Kaliningrad Oblast’, Izd. KGU, Kaliningrad [in
Russian].
Günther R. (1975), “Untersuchungen der Meiose bei Män-
chen von Rana ridibunda Pall., Rana lessonae Cam. und
der Bastardform “Rana esculenta” L. (Anura),” Biol. Zen-
tralblatt, 94(3), 277 – 294.
Günther R. (1997), “Rana kl. esculenta Linnaeus, 1758,”
in: J.-P. Gasc et al. (eds.), Atlas of Amphibians and Reptiles
in Europe, Soc. Eur. Herpetol. and Mus. Natl. d’Hist. Nat.,
Paris, pp. 138 – 139.
Heppich S., Tunner H. G., and Greilhuber J. (1982), “Pre-
meiotic chromosome doubling after genome elimination
during spermatogenesis of the species hybrid Rana escu-
lenta,” Theor. Appl. Genet., 61, 101 – 104.
Hoogesteger T., Rahkonen J., and Karhilahti A. (2013),
“Pool frog (Pelophylax lessonae) Camerano 1882 (Anura,
Ranidae), an addition to the Finnish amphibian fauna,”
Mem. Soc. Fauna Flora Fenn., 89, 25 – 31.
Jakob C. (2007), Structure and Dynamics of Pure Hybridoge-
netic Water Frog Populations of Rana esculenta in Southern
Sweden. Ph.D. Thesis, Universität Zürich.
Kuzmin S. L. (2013), The Amphibians of the Former Soviet
Union, Sofia – Moscow, Pensoft.
Lada G. A., Borkin L. J., Litvinchuk S. N., and Rosanov J. M.
(2009), “Species composition of green frogs (the Rana es-
culenta complex) in Dnepropetrovsk Province (eastern Uk-
raine),” Proc. Ukr. Herpetol. Soc., 2, 37 – 44 [in Russian].
Lada G. A., Borkin L. J., Litvinchuk S. N., and Rosanov J. M.
(2011a), “Species composition and population systems of
green frogs (Rana esculenta complex) in Psyol River drain-
age,” Proc. Ukr. Herpetol. Soc., 3, 76 – 83 [in Russian].
Lada G. A., Borkin L. J., Litvinchuk S. N., and Rosanov J.
M. (2011b), “Types of population systems of green frogs
(Rana esculenta complex) in the territory of the Russian
Plain,” in: Ananjeva N. B. (ed.), Problems of Herpetology,
St. Petersburg, pp. 142 – 148 [in Russian].
Lada G. A. (2012), Anurans of the Russian Plain: Variability,
Speciation, Ranges, Conservation. Doctoral Thesis, Kazan’
[in Russian].
le Roi O. (1903), “Ornithologischer Bericht über die Monate
März bis October 1902 vom südlichen Teile der Kurischen
Nehrung,” J. Ornitol., 51, 231 – 256.
Litvinchuk S. N. (1996), “On records of Salamandra sala-
mandra in the south-eastern part of the Baltic region,” Russ.
J. Herpetol., 3(2), 196 – 198.
Litvinchuk S. N., Pashkova I. M., Rosanov J. M., and Bor-
kin L. J. (2007), “Heat resistance of the muscle in West-Pa-
learctic green frogs of Rana esculenta complex,” Biol.
Bull., 34(1), 61 – 66.
Mezhzherin S. V., Morozov-Leonov S. Yu., Nekrasova O. D.,
Kurtyak F. F., and Zhalay E. I. (2005), “Spatial structure
of hybrid complex of green frogs Rana esculenta (Anura,
Ranidae) on the territory of Ukraine,” in: Pisanets E. (ed.),
Distribution of Green Frogs in Kaliningrad Oblast’ 195

Proc. of the First Conf. of the Ukr. Herpetol. Soc., Izd. Zoo-
museum NMNH NAS of Ukraine, Kyiv, pp. 110 – 114 [in
Russian].
Mikulícek P. and Kotlík P. (2001), “Two water frog popula-
tions from western Slovakia consisting of diploid females
and diploid and triploid males of the hybridogenetic hybrid
Rana esculenta (Anura, Ranidae),” Mitt. Mus. Naturkund.
Berlin. Zool. Reiche, 77(1), 59 – 64.
Mikulíèek P., Kautman M., Kautman J., and Pruvost N. B. M.
(2015), “Mode of hybridogenesis and habitat preferences
influence population composition of water frogs (Pelophy-
lax esculentus complex, Anura: Ranidae) in a region of
sympatric occurrence (western Slovakia),” J. Zool. Syst.
Evol. Res., 53(2), 124 – 132.
Milto K. D. (2007), Amphibians and Reptiles of Northwestern
Russia: Biodiversity Estimation. Ph.D. Thesis, Izd. Zool.
Inst. RAN, St. Petersburg [in Russian].
Milto K. D. (2008), “Amphibians breed in the Baltic Sea,”
Russ. J. Herpetol., 15(1), 8 – 10.
Muhling P. (1898), “Die Helminthen-Fauna der Wirbeltiere
Ostpreussens,” Arch. Naturgesch. Berlin, 64(1), 1 – 118.
Natchev N., Tzankov N., and Gemel R. (2011), “Green frog
invasion in the Black Sea: habitat ecology of the Pelophylax
esculentus complex (Anura, Amphibia) population in the
region of Shablenska Tuzla lagoon in Bulgaria,” Herpetol.
Notes, 4, 347 – 351.
Nekrasova O. D. (2014), “Some aspects of anomaly’s mani-
festation in amphibian coloration,” in: Anomalies and Pa-
thologies of Amphibians and Reptiles: Methodology, Evolu-
tionary Impact, Possibilities for Estimation of Environmen-
tal Health, Yekaterinburg, pp. 144 – 149 [in Russian].
Nekrasova O. D. and Morozov-Leonov S. Yu. (2001), “Diag-
nostics of green frogs complex representatives Rana escu-
lenta L., 1758 (Amphibia, Ranidae) of the hybrid popula-
tions of the Dnepr basin,” Vestn. Zool., 35(5), 47 – 52 [in
Russian].
Ogielska M., Kazana K., and Kierzkowski P. (2001), “DNA
content in erythrocyte nuclei of water frogs from a pure
Rana esculenta population in Dêbki (Gdañsk district, Po-
land),” Mitt. Mus. Naturkund. Berlin. Zool. Reiche, 77(1),
65 – 70.
Pagast F. (1941), “Über die Lurch- und Kriechtierfauna Ost-
preußens,” Physik.-ökon. Gesellschaft Königsberg, 27(1),
173 – 197.
Pisanets E. M. (2007), Amphibians of Ukraine (Guide to Am-
phibians of Ukraine and Adjacent Territories), Zoological
Museum of NSNM NAS of Ukraine, Kyiv [in Russian].
Plötner J. (2005), Die westpalaarktischen Wasserfrosche, Lau-
renti-Verlag, Bielefeld.
Pruvost N. B. M., Hoffmann A., and Reyer H.-U. (2013),
“Gamete production patterns, ploidy, and population genet-
ics reveal evolutionary significant units in hybrid water
frogs (Pelophylax esculentus),” Ecol. Evol., 3(9), 2933 –
2946.
Ruchin A. B., Borkin L. J., Lada G. A., Litvinchuk S. N.,
Rosanov Yu. M., and Ryzhov M. K. (2005), “Morphologi-
cal variation, genome size and population systems of the
green frogs (Rana esculenta complex) of Mordovia,” Byull.
Mosk. Obshch. Ispyt. Prir. Ser. Biol., 111(2), 3 – 10 [in
Russian].
Ruchin A. B., Lada G. A., Borkin L. J., Litvinchuk S. N.,
Rosanov J. M., Ryzhov M. K., and Zamaletdinov R. I.
(2009), “On habitat distribution of three green frog species
of the Rana esculenta complex in the Volga River basin,”
Povolzh. Ékol. Zh. Saratov, 2, 137 – 147 [in Russian].
Ruchin A. B., Borkin L. J., Lada G. A., Litvinchuk S. N.,
Rosanov J. M., and Ryzhov M. K. (2010), “On fauna of
green frogs (Rana esculenta complex) in Chuvashia,”
Nauch. Trudy Nats. Parka “Chavash Varmane,” Chebok-
sary, 3, 102 – 110 [in Russian].
Rybacki M. and Berger L. (1994), “Distribution and ecology
of water frogs in Poland,” Zool. Polon., 39(3�4), 293 – 303.
Rybacki M. and Berger L. (2001), “Types of water frog popu-
lations (Rana esculenta complex) in Poland,” Mitt. Mus.
Naturkund. Berlin. Zool. Reiche, 77(1), 51 – 57.
Sokolov A. A. (2014), “Natural site Romintskaya Puscha,” in:
Nature of Kaliningrad oblast’. Key Natural Sites, Istok, Ka-
liningrad, 30 – 69 [in Russian].
Svinin A. O., Litvinchuck S. N., Borkin L. J., and Rosa-
nov J. M. (2013), “Distribution and population system
types of green frogs (Pelophylax Fitzinger, 1843) in Mari El
Republic,” Curr. Studies Herpetol. Saratov, 13(3�4),
137 – 147 [in Russian].
Svinin A. O., Litvinchuk S. N., Ermakov O. A., Iva-
nov A. Yu., and Rosanov J. M. (2015), “New data about
distribution of the edible frog Pelophylax esculentus (Lin-
naeus, 1758) in Mariy El,” in: Osmanova G. O. and Zhuko-
va L. A. (eds.), Principles and Methods of Conservation of
Biodiversity, Izd. MariGU, Yoshkar-Ola, pp. 108 – 111 [in
Russian].
Tunner H. G. (1974), “Die klonale Struktur einer Wasser-
froschpopulation,” Zeitschr. Zool. Syst. Evolut. Forschung.,
12(4), 309 – 314.
Tunner H. G. and Heppich S. (1981), “Premeiotic genome
exclusion during oogenesis in the common edible frog,
Rana esculenta,” Die Naturwissenschaften, 68, 207 – 208.
Uzzell T. and Berger L. (1975), “Electrophoretic phenotypes
of Rana ridibunda, Rana lessonae, and their hybridogenetic
associate, Rana esculenta,” Proc. Acad. Nat. Sci. Philadel-
phia, 127(2), 13 – 24.
Uzzell T., Hotz H., and Berger L. (1980), “Genome exclusion
in gametogenesis by an interspecific Rana hybrid: evidence
from electrophoresis of individual oocytes,” J. Exp. Zool.,
214(3), 251 – 259.
Vinogradov A. E., Rosanov J. M., Tsaune I. A., and Bor-
kin L. J. (1988), “Elimination of genome of one of parental
species before premeiotic synthesis of DNA in hybridoge-
netic species Rana esculentus,” Tsitologiya, 30(6), 691 –
698 [in Russian].
Vinogradov A. E., Borkin L. J., Günther R., and Rosanov J. M.
(1990), “Genome elimination in diploid and triploid Rana
esculenta males: cytological evidence from DNA flow
cytometry,” Genome, 33(5), 619 – 627.
Zamaletdinov R. I., Borkin L. J., Litvinchuk S. N., and Ro-
sanov J. M. (2005), “On the green frogs complex structure
in Volga-Kama Reserve Raifa part,” Trudy. Volzhsko-Kam-
sky Nats. Prir. Zapovednik. Kazan, 6, 326 – 333 [in Rus-
sian].
196 S. N. Litvinchuk et al.
Related Documents











