RESEARCH ARTICLE Distribution and early life history of Kaupichthys leptocephali (family Chlopsidae) in the central Indonesian Seas Tae Won Lee Michael J. Miller Hak Bin Hwang Sam Wouthuyzen Katsumi Tsukamoto Received: 28 March 2007 / Accepted: 23 August 2007 / Published online: 10 October 2007 Ó Springer-Verlag 2007 Abstract Leptocephali of the widely distributed tropical marine eels of the genus Kaupichthys (family Chlopsidae) were collected around Sulawesi Island during a sampling survey in the Indonesian Seas in late September and early October 2002, and the otolith microstructure of 24 of the 59 specimens captured was examined to learn about the larval growth rates and spawning times of these small sized eels. Leptocephali ranging in size from 25 to 60 mm were collected in Makassar Strait and the Celebes Sea, but they were most abundant in the semi-enclosed Tomini Bay of northeast Sulawesi Island. The Kaupichthys leptocephali examined had 39–161 otolith growth increments. Their back-calculated hatching dates indicated that five age groups were present and each group appeared to have been spawned around the full moon of previous months. Aver- age growth rate estimates of the first two age groups were 0.65 and 0.54 mm/day for the 27.4–30.4 and 37.6– 45.6 mm age classes. The growth rates of the oldest three age groups (52.0–60.8 mm) appeared to have slowed down after they reached their approximate maximum size. An increase in increment widths at the outer margin of the otoliths of those larger than 53 mm suggested that the process of metamorphosis had begun even though there were few external morphological changes indicating metamorphosis. It is hypothesized that chlopsid lepto- cephali have an unusually short gut that may not need to move forward during early metamorphosis. The presence of four age classes in Tomini Bay suggests that the Togian Islands region may be productive habitats for Kaupichthys juveniles and adults. Introduction The false morays of the family Chlopsidae are found in shallow water areas in tropical regions worldwide and are one of the most poorly known families of the Anguilli- formes. These eels are much smaller in size than most marine eels because they only reach maximum sizes of about 250 mm (Smith 1989a). They probably hide in holes or crevices primarily in coral reef areas or sea grass beds and very little is known about their reproductive ecology or spawning areas because they are rarely observed or col- lected (Matsubara and Asano 1959; Smith 1969, 1989a). Some information about their life histories can be inferred from the distribution and size of their leptocephali that suggest they do not migrate very far offshore to spawn (Smith 1989b; Miller 1995; Miller and McCleave 2007). Some of the most common chlopsid eels seem to be those of the genus Kaupichthys, which includes about six species worldwide. Kaupichthys hyoproroides appears to be widely distributed in tropical areas of the western North Atlantic and also has been considered to be present in the Pacific and Indian oceans (Smith 1989a). However, outside Communicated by S. Nishida. T. W. Lee H. B. Hwang Department of Oceanography, Chungnam National University, Daejeon 305-764, Korea M. J. Miller (&) K. Tsukamoto Ocean Research Institute, The University of Tokyo, 1-15-1 Minamidai, Nakano-ku, Tokyo 164-8639, Japan e-mail: [email protected] S. Wouthuyzen Research Center for Oceanography, Indonesian Institute of Sciences, Jl. Pasir Putih 1, Ancol Timur, Jakarta 11480, Indonesia 123 Mar Biol (2008) 153:285–295 DOI 10.1007/s00227-007-0804-z

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
Distribution and early life history of Kaupichthys leptocephali(family Chlopsidae) in the central Indonesian Seas
Tae Won Lee Æ Michael J. Miller Æ Hak Bin Hwang ÆSam Wouthuyzen Æ Katsumi Tsukamoto
Received: 28 March 2007 / Accepted: 23 August 2007 / Published online: 10 October 2007
� Springer-Verlag 2007
Abstract Leptocephali of the widely distributed tropical
marine eels of the genus Kaupichthys (family Chlopsidae)
were collected around Sulawesi Island during a sampling
survey in the Indonesian Seas in late September and early
October 2002, and the otolith microstructure of 24 of the
59 specimens captured was examined to learn about the
larval growth rates and spawning times of these small sized
eels. Leptocephali ranging in size from 25 to 60 mm were
collected in Makassar Strait and the Celebes Sea, but they
were most abundant in the semi-enclosed Tomini Bay of
northeast Sulawesi Island. The Kaupichthys leptocephali
examined had 39–161 otolith growth increments. Their
back-calculated hatching dates indicated that five age
groups were present and each group appeared to have been
spawned around the full moon of previous months. Aver-
age growth rate estimates of the first two age groups were
0.65 and 0.54 mm/day for the 27.4–30.4 and 37.6–
45.6 mm age classes. The growth rates of the oldest three
age groups (52.0–60.8 mm) appeared to have slowed down
after they reached their approximate maximum size.
An increase in increment widths at the outer margin of
the otoliths of those larger than 53 mm suggested that
the process of metamorphosis had begun even though
there were few external morphological changes indicating
metamorphosis. It is hypothesized that chlopsid lepto-
cephali have an unusually short gut that may not need to
move forward during early metamorphosis. The presence
of four age classes in Tomini Bay suggests that the Togian
Islands region may be productive habitats for Kaupichthys
juveniles and adults.
Introduction
The false morays of the family Chlopsidae are found in
shallow water areas in tropical regions worldwide and are
one of the most poorly known families of the Anguilli-
formes. These eels are much smaller in size than most
marine eels because they only reach maximum sizes of
about 250 mm (Smith 1989a). They probably hide in holes
or crevices primarily in coral reef areas or sea grass beds
and very little is known about their reproductive ecology or
spawning areas because they are rarely observed or col-
lected (Matsubara and Asano 1959; Smith 1969, 1989a).
Some information about their life histories can be inferred
from the distribution and size of their leptocephali that
suggest they do not migrate very far offshore to spawn
(Smith 1989b; Miller 1995; Miller and McCleave 2007).
Some of the most common chlopsid eels seem to be
those of the genus Kaupichthys, which includes about six
species worldwide. Kaupichthys hyoproroides appears to
be widely distributed in tropical areas of the western North
Atlantic and also has been considered to be present in the
Pacific and Indian oceans (Smith 1989a). However, outside
Communicated by S. Nishida.
T. W. Lee � H. B. Hwang
Department of Oceanography,
Chungnam National University,
Daejeon 305-764, Korea
M. J. Miller (&) � K. Tsukamoto
Ocean Research Institute, The University of Tokyo,
1-15-1 Minamidai, Nakano-ku, Tokyo 164-8639, Japan
e-mail: [email protected]
S. Wouthuyzen
Research Center for Oceanography,
Indonesian Institute of Sciences, Jl. Pasir Putih 1,
Ancol Timur, Jakarta 11480, Indonesia
123
Mar Biol (2008) 153:285–295
DOI 10.1007/s00227-007-0804-z

of the Atlantic, this species is now considered to be a
complex of cryptic species that include K. japonicus in the
Indonesia region and K. diodontus in the central Pacific
region, which appear to be the more common species in the
tropical Indo-Pacific (Froese and Pauly 2007; K. Tighe pers
comm). These eels are found primarily in and around coral
reefs and are rarely collected without the use of ichthyo-
cides (Smith 1989a). Females can reach sexual maturity by
at least a size of 150 mm (Smith 1989a) and K. hyopr-
oroides appears to be one of the more common chlopsid
eels based on the abundance of their leptocephali (Smith
1989b; Miller 1995; Miller and McCleave 2007). Smith
(1989b) reported that K. hyoproroides leptocephali were
collected throughout the year in the western North Atlantic.
The leptocephali of this genus also have been described
from the western North Pacific (Tabeta and Mochioka
1988), however, four species of Kaupichthys appear to be
present in the Indo-Pacific (K. astronasus, K. brachychirus,
K. diodontus, K. japonicus) based on adult specimens
(Matsubara and Asano 1959; Allen and Adrim 2003;
Froese and Pauly 2007). More research is needed on the
morphology and identification of Kaupichthys adults and
leptocephali in the Indonesian Seas region to facilitate
studies on the life histories of these poorly known fishes.
Because aspects of the life history of fishes are recorded
chronologically in otoliths, analysis of otolith microstruc-
ture provides valuable information about the previous
growth history of a species when some life stages are not
easy to study in their native habitats. Although the otolith
microstructure of Kaupichthys or other chlopsid lepto-
cephali have not been previously examined, there have
been studies on the otoliths of leptocephali of anguillid and
a few families of marine eels. The otolith microstructure
and microchemistry of several species of temperate and
tropical anguillid leptocephali (Castonguay 1987; Arai
et al. 2001a; Ishikawa et al. 2001; Kuroki et al. 2005, 2006)
and glass eels have been studied (e.g. Tsukamoto 1990;
Otake et al. 1994; Wang and Tzeng 2000; Arai et al. 1997,
2001b; Marui et al. 2001). There also have been studies on
the otolith microstructure of leptocephali of congrid eels
such as Conger (e.g. Lee and Byun 1996; Otake et al. 1997;
Correia et al. 2002, 2004), Ariosoma, and Paraconger
(Bishop et al. 2000). Other leptocephali of marine eels of
the families Muraenidae and Ophichthidae (Bishop et al.
2000) and Nettatomatidae and Synaphobranchidae (Ma
et al. 2005) also have been aged using their otolith
microstructure. These studies have been based on the
assumption of daily deposition of growth increments in
otoliths after hatching, which has been validated in the
glass eel or elver stages of Anguilla japonica (Tsukamoto
1989), Anguilla rostrata (Cieri and McCleave 2001), and
tropical anguillids (Arai et al. 2000; Sugeha et al. 2001).
Evidence of daily deposition during the leptocephalus stage
also has been found using A. japonica larvae hatched and
reared in the laboratory (Shinoda et al. 2004) and a meta-
morphosing ophichthid (Powles et al. 2006).
Studies on the otolith microstructure of leptocephali and
glass eels can be especially useful because leptocephali
undergo drastic changes in morphology when they meta-
morphose into the glass eel stage, which is reflected in their
otoliths (Otake 2003). During metamorphosis, the end of
the gut moves forward, the larval teeth are absorbed, the
body thickens, and there is an increase in head length
(Castle 1970; Asano et al. 1978; Smith 1989c; Lee and
Byun 1996; Bell et al. 2003; Miller and Tsukamoto 2004).
Otolith studies found that during metamorphosis there is an
increase in the otolith increment widths in anguillid and
congrid leptocephali, which is also accompanied by chan-
ges in the Sr:Ca ratios in the otoliths (Otake et al. 1994,
1997; Arai et al. 1997). Leptocephali differ significantly
from other fish larvae however, because the body is
transparent and is filled with an energy storage material
that is used to form new tissues during metamorphosis
(Pfeiler 1999), but the physiological processes that occur
when the body of leptocephali is transformed into the eel-
like body of glass eels are poorly understood.
This study was designed to provide the first information
about the early life history and spawning times of chlopsid
eels by describing the larval distribution and size of the
Kaupichthys leptocephali collected around Sulawesi Island
in the central Indonesian Seas and by examining their
otolith microstructure. The specific objectives were to use
otolith microstructure to determine the ages, larval growth
rates, and hatching dates using a wide range of sizes of
leptocephali that were collected in various different areas
during a sampling survey that targeted leptocephali.
Materials and methods
Leptocephali were collected in September and October of
2002 during a cruise of the R/V Baruna Jaya VII of the
Research Center for Oceanography of the Indonesian
Institute of Sciences. Sampling occurred at 34 stations
in the Java Sea, Makassar Strait, Celebes Sea, Molucca
(Maluku) Sea, and Tomini Bay (Fig. 1). The oceano-
graphic characteristics of this region have been overviewed
recently by Gordon (2005), Miller et al. (2006) and Susanto
et al. (2006). Sampling for leptocephali at each station
usually consisted of a single tow of the large pelagic trawl,
the Isaacs Kidd Midwater Trawl (IKMT) with a net
opening of 8.7 m2 and 0.5 mm mesh size (Isaacs and Kidd
1953). All sampling was done at nighttime (except for two
stations in Tomini Bay), and each tow consisted of an
approximately 30 min oblique IKMT tow to a depth of
about 200 m in most cases except in the Java Sea, or a
286 Mar Biol (2008) 153:285–295
123

60–80 min IKMT step tow, which towed horizontally for
10 min at five depths of around 30, 60, 90, 120 and 150 m.
Both an oblique and a step tow were made at the same
station (Stn) on two occasions (Stn 9, 31). Stations were
numbered sequentially as sampling occurred along the
cruise track from the Java Sea through Makassar Strait, the
southern Celebes Sea, around the northern tip of Suluawesi
Island, through the Molucca Sea, and into Tomini Bay
where there were 11 stations. The same cruise track was
followed on the return trip back through the Celebes Sea,
Makassar Strait, and the over the continental shelf of the
Java Sea. Except for the stations in the Java Sea (Stn 1, 31,
32, 33, 34) all stations were located over water about
1,000–4,000 m deep.
Specimens were sorted fresh from the plankton, their
lengths measured to the nearest 0.1 mm (total length TL,
predorsal length PDL, preanal length PAL). Among the 59
individuals collected, 24 specimens representative of entire
size range of leptocephali were subsampled for otolith
analysis and they were preserved in 99% ethanol; the other
specimens were preserved in a 10% formalin–seawater
solution. The body shape and distinctive pigment spots
all over the body make Kaupichthys leptocephali easy to
identify (Smith 1969; Tabeta and Mochioka 1988; Smith
1989b), so time constraints resulted in the number of
myomeres (total myomeres TM, predorsal myomyeres
PDM, preanal myomeres PAM) being obtained from only a
subsample of the specimens that were collected (N = 21),
and these counts ranged from 107 to 126. This range is
similar to the overall range of number of vertebrae of the
juveniles and adults of the K. hyoproroides species complex
in the western Atlantic, western Pacific, and Indian oceans,
which was found to be 109–126 (Smith 1989a). The
leptocephali collected around Sulawesi Island appear to be
the K. hyoproroides type, so they may mostly be the larvae
of K. japonicus in the Indonesian Seas region, but there is
presently not enough published information to allow their
identification due to the overlapping ranges of vertebrae
and myomeres of the Indo-Pacific species (Smith 1969).
Therefore, because of the present lack of information about
the morphology or genetic differences in the Indo-Pacific
species of this genus, the species identity of the Kaupichthys
leptocephali in present study were not determined.
The otoliths of 24 Kaupichthys leptocephali were pre-
pared for examination of their microstructure according to
Lee and Byun (1996). Briefly, the otoliths were embedded
in polyester resin, ground to the sagittal plane with a series
of graded silicon carbide papers (600, 800 and 1,000 grit),
and polished with 1 lm alumina powder. Grinding and
polishing was continued until the growth increments were
clearly visible. All specimens were observed under the
scanning electron microscope (SEM). For viewing under
the SEM, ground surfaces of otoliths were etched with
0.3% HCl for 3–5 s. These samples were coated with gold
for 5 min and examined under the SEM.
Total radius (R) was measured along the longest axis
from the core to the anterior margin using a light micro-
scope when the core was clearly visible during grinding.
The ground surface viewed under the SEM did not always
show the exact sagittal plane through the core, so the radii
and the incremental widths measured under the SEM
photographs were calibrated using the radii measured
Tomini Bay
South ChinaSea
Celebes Sea
Borneo
Java
SulawesiIsland
125°E120°E115°E110°E
Flores Sea
Banda SeaJava Sea
MoluccaSea
0°
5°S
5°N
tiartS rassaka
M
1
2
3
45
6 78 92425
2627
28
2930
3132
3334
10 11
12
131415
Fig. 1 Map of the stations in
the Indonesian Seas around
Sulawesi Island, Indonesia that
were sampled from 27
September to 16 October 2002
showing the locations where
Kaupichthys leptocephali were
collected. Specimens used for
otolith analyses are shown with
black squares, other
Kaupichthys leptocephali with
black circles, and stations where
no Kaupichthys leptocephali
were collected with whitecircles. Stations are labeled
1–34, but stations 16–23 in
Tomini Bay are not labeled
Mar Biol (2008) 153:285–295 287
123
under the light microscope. Since daily deposition of
growth increments in fish otoliths has been generally
accepted, and the microstructure of Kaupichthys otoliths
was similar to that of the other anguilliform otoliths for
which the daily deposition has been validated, the age was
estimated from the number of growth increments from the
core, assuming that the growth increments were deposited
daily from hatching.
The growth rates of the leptocephali based on the
examination of their otolith microstructure was determined
using their age and TL data. A growth curve was fitted to
these data that was calculated using the Von Bertalanffy
growth model. For comparison to the growth rates in other
previous studies on leptocephali, the individual growth
rates were calculated for each specimen by first subtracting
an estimated 3 mm for the size at hatching as has been
done in other studies (Arai et al. 2001b; Ma et al. 2005;
Kuroki et al. 2006), and calculating growth as, growth
rate = TL–3/age. The mean individual growth rates for
each age group found by back-calculating their hatching
dates were also calculated.
Results
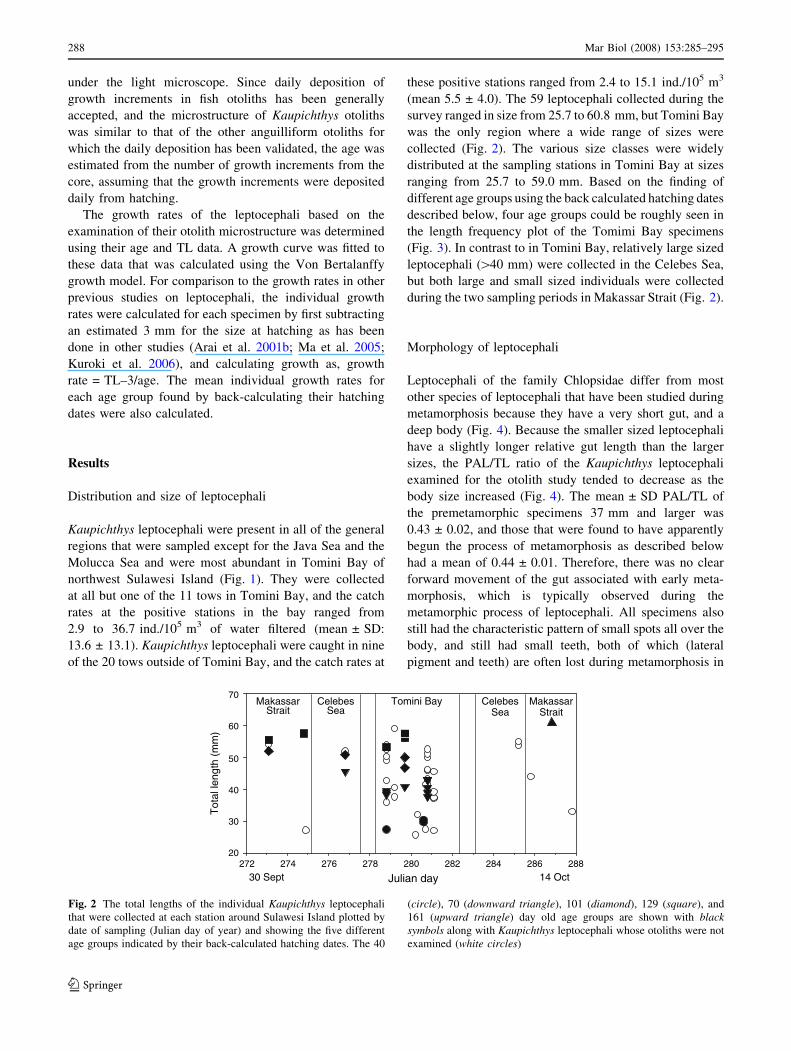
Distribution and size of leptocephali
Kaupichthys leptocephali were present in all of the general
regions that were sampled except for the Java Sea and the
Molucca Sea and were most abundant in Tomini Bay of
northwest Sulawesi Island (Fig. 1). They were collected
at all but one of the 11 tows in Tomini Bay, and the catch
rates at the positive stations in the bay ranged from
2.9 to 36.7 ind./105 m3 of water filtered (mean ± SD:
13.6 ± 13.1). Kaupichthys leptocephali were caught in nine
of the 20 tows outside of Tomini Bay, and the catch rates at
these positive stations ranged from 2.4 to 15.1 ind./105 m3
(mean 5.5 ± 4.0). The 59 leptocephali collected during the
survey ranged in size from 25.7 to 60.8 mm, but Tomini Bay
was the only region where a wide range of sizes were
collected (Fig. 2). The various size classes were widely
distributed at the sampling stations in Tomini Bay at sizes
ranging from 25.7 to 59.0 mm. Based on the finding of
different age groups using the back calculated hatching dates
described below, four age groups could be roughly seen in
the length frequency plot of the Tomimi Bay specimens
(Fig. 3). In contrast to in Tomini Bay, relatively large sized
leptocephali ([40 mm) were collected in the Celebes Sea,
but both large and small sized individuals were collected
during the two sampling periods in Makassar Strait (Fig. 2).
Morphology of leptocephali
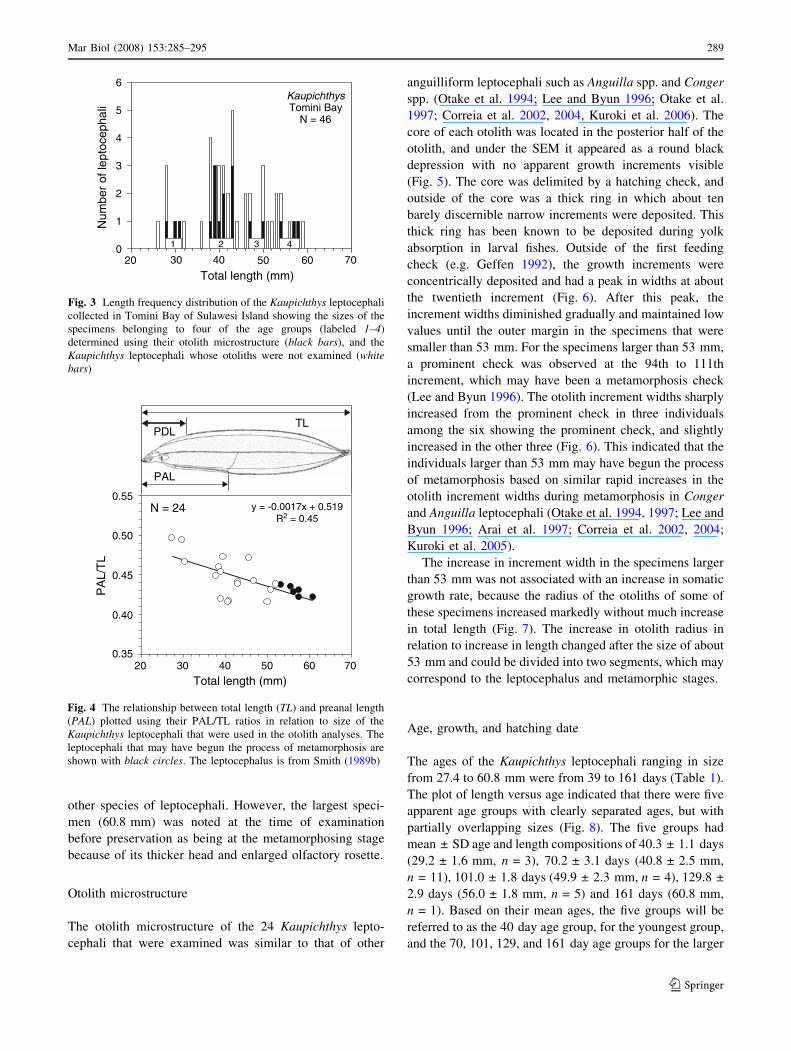
Leptocephali of the family Chlopsidae differ from most
other species of leptocephali that have been studied during
metamorphosis because they have a very short gut, and a
deep body (Fig. 4). Because the smaller sized leptocephali
have a slightly longer relative gut length than the larger
sizes, the PAL/TL ratio of the Kaupichthys leptocephali
examined for the otolith study tended to decrease as the
body size increased (Fig. 4). The mean ± SD PAL/TL of
the premetamorphic specimens 37 mm and larger was
0.43 ± 0.02, and those that were found to have apparently
begun the process of metamorphosis as described below
had a mean of 0.44 ± 0.01. Therefore, there was no clear
forward movement of the gut associated with early meta-
morphosis, which is typically observed during the
metamorphic process of leptocephali. All specimens also
still had the characteristic pattern of small spots all over the
body, and still had small teeth, both of which (lateral
pigment and teeth) are often lost during metamorphosis in
Julian day272 274 276 278 280 282 284 286 288
30 Sept 14 Oct
)m
m( htgnel latoT
20
30
40
50
60
70Makassar
StraitCelebes
SeaTomini Bay Celebes
SeaMakassar
Strait
Fig. 2 The total lengths of the individual Kaupichthys leptocephali
that were collected at each station around Sulawesi Island plotted by
date of sampling (Julian day of year) and showing the five different
age groups indicated by their back-calculated hatching dates. The 40
(circle), 70 (downward triangle), 101 (diamond), 129 (square), and
161 (upward triangle) day old age groups are shown with blacksymbols along with Kaupichthys leptocephali whose otoliths were not
examined (white circles)
288 Mar Biol (2008) 153:285–295
123
other species of leptocephali. However, the largest speci-
men (60.8 mm) was noted at the time of examination
before preservation as being at the metamorphosing stage
because of its thicker head and enlarged olfactory rosette.
Otolith microstructure
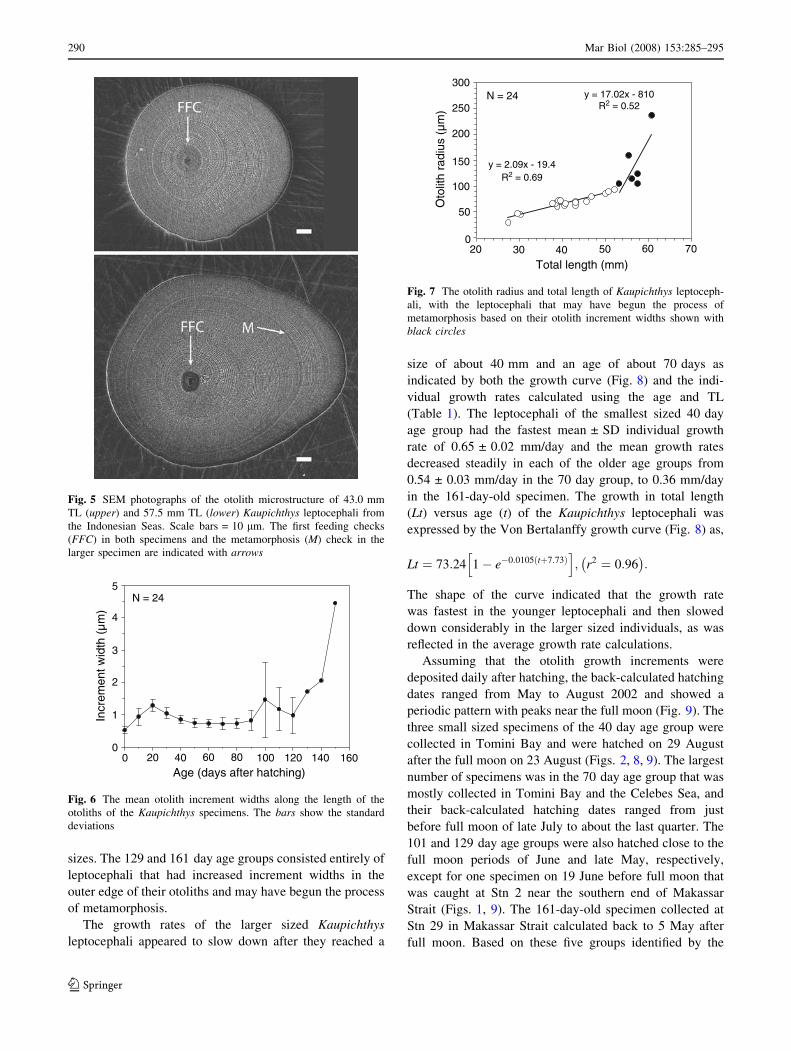
The otolith microstructure of the 24 Kaupichthys lepto-
cephali that were examined was similar to that of other
anguilliform leptocephali such as Anguilla spp. and Conger
spp. (Otake et al. 1994; Lee and Byun 1996; Otake et al.
1997; Correia et al. 2002, 2004, Kuroki et al. 2006). The
core of each otolith was located in the posterior half of the
otolith, and under the SEM it appeared as a round black
depression with no apparent growth increments visible
(Fig. 5). The core was delimited by a hatching check, and
outside of the core was a thick ring in which about ten
barely discernible narrow increments were deposited. This
thick ring has been known to be deposited during yolk
absorption in larval fishes. Outside of the first feeding
check (e.g. Geffen 1992), the growth increments were
concentrically deposited and had a peak in widths at about
the twentieth increment (Fig. 6). After this peak, the
increment widths diminished gradually and maintained low
values until the outer margin in the specimens that were
smaller than 53 mm. For the specimens larger than 53 mm,
a prominent check was observed at the 94th to 111th
increment, which may have been a metamorphosis check
(Lee and Byun 1996). The otolith increment widths sharply
increased from the prominent check in three individuals
among the six showing the prominent check, and slightly
increased in the other three (Fig. 6). This indicated that the
individuals larger than 53 mm may have begun the process
of metamorphosis based on similar rapid increases in the
otolith increment widths during metamorphosis in Conger
and Anguilla leptocephali (Otake et al. 1994, 1997; Lee and
Byun 1996; Arai et al. 1997; Correia et al. 2002, 2004;
Kuroki et al. 2005).
The increase in increment width in the specimens larger
than 53 mm was not associated with an increase in somatic
growth rate, because the radius of the otoliths of some of
these specimens increased markedly without much increase
in total length (Fig. 7). The increase in otolith radius in
relation to increase in length changed after the size of about
53 mm and could be divided into two segments, which may
correspond to the leptocephalus and metamorphic stages.
Age, growth, and hatching date
The ages of the Kaupichthys leptocephali ranging in size
from 27.4 to 60.8 mm were from 39 to 161 days (Table 1).
The plot of length versus age indicated that there were five
apparent age groups with clearly separated ages, but with
partially overlapping sizes (Fig. 8). The five groups had
mean ± SD age and length compositions of 40.3 ± 1.1 days
(29.2 ± 1.6 mm, n = 3), 70.2 ± 3.1 days (40.8 ± 2.5 mm,
n = 11), 101.0 ± 1.8 days (49.9 ± 2.3 mm, n = 4), 129.8 ±
2.9 days (56.0 ± 1.8 mm, n = 5) and 161 days (60.8 mm,
n = 1). Based on their mean ages, the five groups will be
referred to as the 40 day age group, for the youngest group,
and the 70, 101, 129, and 161 day age groups for the larger
KaupichthysTomini Bay
N = 46
Total length (mm)70
ilahpecotpel fo rebmu
N
0
1
2
3
4
5
6
1 2 3 4
20 30 40 50 60
Fig. 3 Length frequency distribution of the Kaupichthys leptocephali
collected in Tomini Bay of Sulawesi Island showing the sizes of the
specimens belonging to four of the age groups (labeled 1–4)
determined using their otolith microstructure (black bars), and the
Kaupichthys leptocephali whose otoliths were not examined (whitebars)
Total length (mm)20 30 40 50 60 70
PA
L/T
L
0.55
0.50
0.45
0.40
0.35
y = -0.0017x + 0.519R2 = 0.45
N = 24
TL
PAL
PDL
Fig. 4 The relationship between total length (TL) and preanal length
(PAL) plotted using their PAL/TL ratios in relation to size of the
Kaupichthys leptocephali that were used in the otolith analyses. The
leptocephali that may have begun the process of metamorphosis are
shown with black circles. The leptocephalus is from Smith (1989b)
Mar Biol (2008) 153:285–295 289
123
sizes. The 129 and 161 day age groups consisted entirely of
leptocephali that had increased increment widths in the
outer edge of their otoliths and may have begun the process
of metamorphosis.
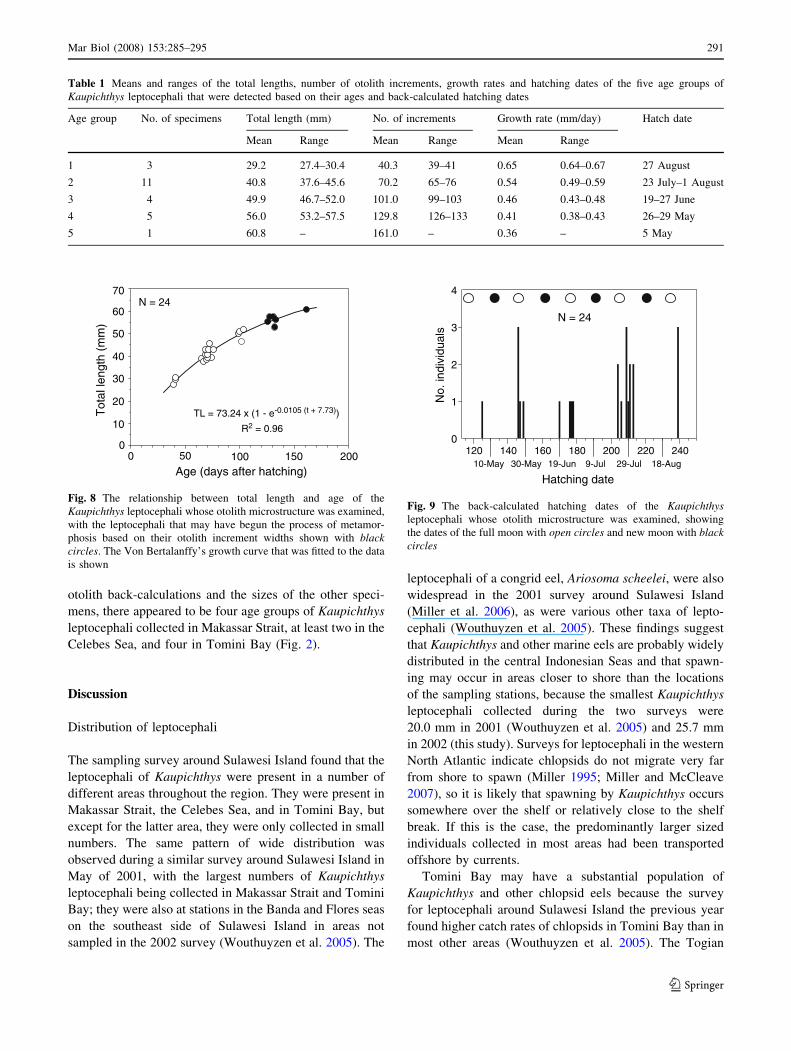
The growth rates of the larger sized Kaupichthys
leptocephali appeared to slow down after they reached a
size of about 40 mm and an age of about 70 days as
indicated by both the growth curve (Fig. 8) and the indi-
vidual growth rates calculated using the age and TL
(Table 1). The leptocephali of the smallest sized 40 day
age group had the fastest mean ± SD individual growth
rate of 0.65 ± 0.02 mm/day and the mean growth rates
decreased steadily in each of the older age groups from
0.54 ± 0.03 mm/day in the 70 day group, to 0.36 mm/day
in the 161-day-old specimen. The growth in total length
(Lt) versus age (t) of the Kaupichthys leptocephali was
expressed by the Von Bertalanffy growth curve (Fig. 8) as,
Lt ¼ 73:24 1� e�0:0105 tþ7:73ð Þh i
; r2 ¼ 0:96� �
:
The shape of the curve indicated that the growth rate
was fastest in the younger leptocephali and then slowed
down considerably in the larger sized individuals, as was
reflected in the average growth rate calculations.
Assuming that the otolith growth increments were
deposited daily after hatching, the back-calculated hatching
dates ranged from May to August 2002 and showed a
periodic pattern with peaks near the full moon (Fig. 9). The
three small sized specimens of the 40 day age group were
collected in Tomini Bay and were hatched on 29 August
after the full moon on 23 August (Figs. 2, 8, 9). The largest
number of specimens was in the 70 day age group that was
mostly collected in Tomini Bay and the Celebes Sea, and
their back-calculated hatching dates ranged from just
before full moon of late July to about the last quarter. The
101 and 129 day age groups were also hatched close to the
full moon periods of June and late May, respectively,
except for one specimen on 19 June before full moon that
was caught at Stn 2 near the southern end of Makassar
Strait (Figs. 1, 9). The 161-day-old specimen collected at
Stn 29 in Makassar Strait calculated back to 5 May after
full moon. Based on these five groups identified by the
Fig. 5 SEM photographs of the otolith microstructure of 43.0 mm
TL (upper) and 57.5 mm TL (lower) Kaupichthys leptocephali from
the Indonesian Seas. Scale bars = 10 lm. The first feeding checks
(FFC) in both specimens and the metamorphosis (M) check in the
larger specimen are indicated with arrows
)mµ( htdi
w tneme rcnI
Age (days after hatching)
0
1
2
3
4
5
1400 160
N = 24
20 40 60 10080 120
Fig. 6 The mean otolith increment widths along the length of the
otoliths of the Kaupichthys specimens. The bars show the standard
deviations
Total length (mm)
300
250
200
150
100
50
03020 40 50 60 70
y = 17.02x - 810R2 = 0.52)
mµ( suidar htilotO
y = 2.09x - 19.4R2 = 0.69
N = 24
Fig. 7 The otolith radius and total length of Kaupichthys leptoceph-
ali, with the leptocephali that may have begun the process of
metamorphosis based on their otolith increment widths shown with
black circles
290 Mar Biol (2008) 153:285–295
123
otolith back-calculations and the sizes of the other speci-
mens, there appeared to be four age groups of Kaupichthys
leptocephali collected in Makassar Strait, at least two in the
Celebes Sea, and four in Tomini Bay (Fig. 2).
Discussion
Distribution of leptocephali
The sampling survey around Sulawesi Island found that the
leptocephali of Kaupichthys were present in a number of
different areas throughout the region. They were present in
Makassar Strait, the Celebes Sea, and in Tomini Bay, but
except for the latter area, they were only collected in small
numbers. The same pattern of wide distribution was
observed during a similar survey around Sulawesi Island in
May of 2001, with the largest numbers of Kaupichthys
leptocephali being collected in Makassar Strait and Tomini
Bay; they were also at stations in the Banda and Flores seas
on the southeast side of Sulawesi Island in areas not
sampled in the 2002 survey (Wouthuyzen et al. 2005). The
leptocephali of a congrid eel, Ariosoma scheelei, were also
widespread in the 2001 survey around Sulawesi Island
(Miller et al. 2006), as were various other taxa of lepto-
cephali (Wouthuyzen et al. 2005). These findings suggest
that Kaupichthys and other marine eels are probably widely
distributed in the central Indonesian Seas and that spawn-
ing may occur in areas closer to shore than the locations
of the sampling stations, because the smallest Kaupichthys
leptocephali collected during the two surveys were
20.0 mm in 2001 (Wouthuyzen et al. 2005) and 25.7 mm
in 2002 (this study). Surveys for leptocephali in the western
North Atlantic indicate chlopsids do not migrate very far
from shore to spawn (Miller 1995; Miller and McCleave
2007), so it is likely that spawning by Kaupichthys occurs
somewhere over the shelf or relatively close to the shelf
break. If this is the case, the predominantly larger sized
individuals collected in most areas had been transported
offshore by currents.
Tomini Bay may have a substantial population of
Kaupichthys and other chlopsid eels because the survey
for leptocephali around Sulawesi Island the previous year
found higher catch rates of chlopsids in Tomini Bay than in
most other areas (Wouthuyzen et al. 2005). The Togian
Table 1 Means and ranges of the total lengths, number of otolith increments, growth rates and hatching dates of the five age groups of
Kaupichthys leptocephali that were detected based on their ages and back-calculated hatching dates
Age group No. of specimens Total length (mm) No. of increments Growth rate (mm/day) Hatch date
Mean Range Mean Range Mean Range
1 3 29.2 27.4–30.4 40.3 39–41 0.65 0.64–0.67 27 August
2 11 40.8 37.6–45.6 70.2 65–76 0.54 0.49–0.59 23 July–1 August
3 4 49.9 46.7–52.0 101.0 99–103 0.46 0.43–0.48 19–27 June
4 5 56.0 53.2–57.5 129.8 126–133 0.41 0.38–0.43 26–29 May
5 1 60.8 – 161.0 – 0.36 – 5 May
)m
m( htgnel latoT
Age (days after hatching)
20
10
00
30
40
50
50 100 150 200
60
70
TL = 73.24 x (1 - e-0.0105 (t + 7.73))
N = 24
R2 = 0.96
Fig. 8 The relationship between total length and age of the
Kaupichthys leptocephali whose otolith microstructure was examined,
with the leptocephali that may have begun the process of metamor-
phosis based on their otolith increment widths shown with blackcircles. The Von Bertalanffy’s growth curve that was fitted to the data
is shown
N = 24
Hatching date
120 140 160 180 200 220 240slaudividni .o
N0
1
2
3
4
18-Aug29-Jul9-Jul19-Jun10-May 30-May
Fig. 9 The back-calculated hatching dates of the Kaupichthysleptocephali whose otolith microstructure was examined, showing
the dates of the full moon with open circles and new moon with blackcircles
Mar Biol (2008) 153:285–295 291
123

Islands are the large group of coral reef islands that can be
seen in Tomini Bay in Fig. 1, and they support a rich coral
reef fauna that includes an unusually high diversity of
corals and some endemic fish species (Wallace 1999; Allen
and Adrim 2003). These coral reef and other habitats may
also support many chlopsid eels, and Allen and Adrim
(2003) reported that both K. brachychirus, K. diodontus
(possibly K. japonicus) were collected in the Togian
Islands. The leptocephali spawned near these islands and
other reef habitats would likely remain within the bay due
to the potentially limited amount of water exchange with
outside areas (Hatayama et al. 1996). Tidal currents and
eddies appear to have caused the leptocephali of four dif-
ferent spawning groups to mix in Tomini Bay where they
were collected at sizes ranging from 25.7 to 59.0 mm.
Coral reef areas are widespread around Sulawesi Island
(Wouthuyzen et al. 2005; Miller et al. 2006) so there may
be suitable habitat for Kaupichthys juveniles and adults
throughout the region.
Otolith microstructure and growth of leptocephali
The otolith microstructure of Kaupichthys leptocephali
consisted of a series of concentric rings from the core to the
edge of the otolith (Fig. 5) that was essentially the same as
other species of leptocephali that have been observed. Each
otolith had a central core, an area deposited during yolk
absorption, and concentric rings extending out to the edge
of the otolith. These same characteristics have also been
observed in the otoliths of the leptocephali of both tem-
perate (Castonguay 1987; Otake et al. 1994; Arai et al.
1997) and tropical (Kuroki et al. 2006) anguillids, species
of Conger (Lee and Byun 1996; Otake et al. 1997; Correia
et al. 2002, 2004, 2006), and nettastomatid and synapho-
branchid species (Ma et al. 2005). Based on the total
number of increments, the ages of the Kaupichthys lepto-
cephali (27.4–60.8 mm) that were examined ranged from
39 to 161 days old (Table 1).
The growth rates of the Kaupichthys leptocephali stea-
dily decreased as the larvae grew older based on the growth
curve of their body sizes and ages. Their mean individual
growth rates varied from 0.65 mm/day in the youngest age
group, to 0.36 mm/day in the oldest. The mean individual
growth rate of the 70 day age group (37.6–45.6 mm) was
0.54 mm/day, which is similar to estimates of the mean
growth rates (0.44–0.56 mm/day) calculated the same way
for four species of anguillid leptocephali (8.0–54.1 mm)
from the Indo-Pacific region including the Indonesian Seas
(Kuroki et al. 2006). Similar individual growth rates of
0.43–0.83 mm mm/day for Sarenchelys stylura leptoceph-
ali (10.0–48.6 mm) and 0.20–0.53 mm/day for Dysomma
sp. leptocephali (8.4–33.5 mm) were found in specimens
from the East China Sea (Ma et al. 2005). Faster growth
rates for species of the Congridae, Muraenidae, or Oph-
ichthidae that were suggested to be greater than 1.0 mm/day
(Bishop et al. 2000), but these faster growth rates have yet
to be confirmed in other samples of leptocephali.
The otolith increment width and otolith radius of
Kaupichthys leptocephali grew slowly before increasing
abruptly from the age of 94–111 days in the six oldest
specimens. This rapid increase in increment width has been
considered to occur during metamorphosis from the lep-
tocephalus stage to the glass eel stage (Otake et al. 1994,
1997; Arai et al. 1997, 2001; Wang and Tzeng 2000; Marui
et al. 2001; Correia et al. 2006). Further support for this
hypothesis was recently found in Anguilla marmorata in a
study that included leptocephali, metamorphosing lepto-
cephali, oceanic glass eels and glass eels collected after
recruitment to coastal areas (Kuroki et al. 2005). The rapid
increase in increment widths was observed to have begun
in the metamorphosing leptocephali and glass eels, but not
in the premetamorphic leptocephali. An increase in otolith
increment widths was also observed to occur during
metamorphosis in tarpon leptocephali (Chen and Tzeng
2006).
Onset of metamorphosis
During the transition from larvae to juveniles, fish larvae
undergo various morphological and osteological modifi-
cations related to changes in swimming and feeding
function. During the metamorphosis of leptocephali the
head thickens, the teeth are lost, the body length is reduced,
and the anus, and the anterior base of the dorsal and anal
fins usually move forward (Smith 1989c; Otake 2003;
Miller and Tsukamoto 2004). These changes in morphol-
ogy during metamorphosis of leptocephali have been
documented in a variety of species and families of eels
(Castle 1970; Raju 1974; Asano et al. 1978; Lieby 1979;
Bell et al. 2003). Interestingly, five of the six largest
Kaupichthys specimens that were examined in this study
showed no obvious external morphological evidence of
metamorphosis, but they showed an increase in the incre-
ment widths at the outer edge of their otoliths, which
suggested the onset of metamorphosis was occurring.
The forward movement of the gut as measured by an
abrupt decrease of the PAL/TL or PAM/TM ratios has been
used as a criterion for defining the onset of metamorphosis
of conger eel leptocephali, because it is difficult to deter-
mine the relative stage of metamorphosis based on other
morphometric characteristics (Tanaka et al. 1987; Lee
and Byun 1996; Otake et al. 1997). The leptocephali of
Anguilla, Ariosoma and Conger have long guts at the onset
of metamorphosis, so their PAL/TL ratios decrease during
292 Mar Biol (2008) 153:285–295
123

metamorphosis, because it is necessary for the gut to move
forward to the eventual position of the anus in the glass eel
and juvenile eel stage (Asano et al. 1978; Lee and Byun
1996; Bell et al. 2003; Otake 2003; Miller et al. 2006).
However, the PAL/TL ratio of Kaupichthys leptocephali
had not changed in the specimens whose otolith increment
widths showed abrupt increases. The obvious possible
reason for this lack of a decrease in the PAL/TL ratios of
the largest Kaupichthys leptocephali is that the position of
the end of the gut in chlopsid leptocephali is located much
farther forward than most other taxa of leptocephali
(Fig. 4; Tabeta and Mochioka 1988; Smith 1989b; Miller
and Tsukamoto 2004). Therefore, because of this anterior
position, the gut does not have to move much to be in its
final position at the end of metamorphosis. Some species of
ophichthid leptocephali, such as Myrophis puntatus (Lieby
1979; Powles et al. 2006) and M. platyrhynchus (Lieby
1989), also have very short guts, which may not move very
much more forward during metamorphosis.
The present study suggests that in some species the rapid
increase in otolith increment widths may occur before there
are major changes in external morphology. It is possible
the rapid increase in otolith increment widths may be
associated with physiological changes linked to the meta-
morphic process, such as breaking down the transparent
internal glycosaminoglycan (GAG) material for use in
building new tissues in the head and body. The oldest
Kaupichthys specimen examined here was 28 days older
than the next oldest specimen, and it showed clear changes
in the morphology of its head, with a thickening of the
tissue and an enlargement of the olfactory rosette. Smith
(1969) described a glass eel and a metamorphosing lepto-
cephalus of Kaupichthys hyoproroides from near the
northern Bahamas in which the gut had moved forward, so
it is likely that as metamorphosis progresses further, the gut
eventually moves forward in chlopsid leptocephali. The
body length of leptocephali also decreases markedly during
metamorphosis from the leptocephalus stage to the glass
eel stage, but the Kaupichthys leptocephali examined here
were possibly not at enough of an advanced stage of
metamorphosis to show any size decrease however. Further
research is needed to determine if the process of meta-
morphosis proceeds differently in chlopsid leptocephali by
examining a greater number of large sized specimens
including those that show clear external evidence of
metamorphosis.
Spawning periodicity
The back-calculated hatching dates of the Kaupichthys
leptocephali examined during this study were mostly
around one of the five full moons from early May to late
August of 2002. This finding suggested that this genus of
marine eels has a lunar cycle of reproduction and spawn
during or shortly after full moon periods around Sulawesi
Island. Four of these spawning groups appeared to be
collected in more than one sea area based on the otolith
analyses or the distinctiveness of the smallest size class
of leptocephali. This suggested that synchronous lunar
spawning may have been occurring in many areas, and so
this could be a life history characteristic of Kaupichthys
eels in the central Indonesian Seas region.
Lunar periodicity has not been reported in marine eels,
but their reproductive ecologies are diverse and very poorly
known (Thresher 1984; Fishelson 1992, 1994). Two ripe
Kaupichthys females (238, 239 mm TL) with full-grown
ova were collected from a shallow coral head in the Amami
Islands near Okinawa Japan during the new moon of
July 1958 (Matsubara and Asano 1959), but it is unclear
when they would have eventually spawned. However, the
catadromous anguillid eel, A. japonica, appears to spawn
during new moon periods in its offshore spawning area in
the western North Pacific (Ishikawa et al. 2001; Tsukamoto
et al. 2003; Tsukamoto 2006). Analyses of the otolith
microstructure of A. japonica leptocephali collected near
the spawning area during several different years have
shown that their back-calculated hatching dates were all
centered on the new moon periods of previous months,
suggesting that this species synchronizes its spawning with
the lunar cycle.
Various other species of teleost fishes such as groupers,
snappers, rabbitfishes and other coral reef fishes also have
lunar spawning periodicities that serve to synchronize their
spawning activities (Thresher 1984). This subject has been
reviewed recently (Takemura et al. 2004) and spawning
associated with either full or new moon has been reported
in a wide variety of families of marine fishes (Rhodes
and Sadovy 2002; Heyman et al. 2005; Johannes 1978;
Domeier and Colin 1997). A primary purpose of this
behavior appears to be to synchronize the formation of
spawning aggregations in a particular location with all
participants becoming ripe at the same time for successful
fertilization of eggs. Other possible functions of lunar
synchronization include predator avoidance, defense of
demersal egg clutches from nocturnal predators, or the
release of eggs during periods of strong tidal flow to
facilitate transport of eggs offshore away from the reef
(Takemura et al. 2004).
Very little is known about the reproductive ecology of
chlopsid and other marine eels however, so the function of
full moon spawning is difficult to determine. Kaupichthys
eels have polycyclic ovaries (Fishelson 1994), but it is
unknown if they spawn multiple times per year as is possible
with this type of ovary, or if they have any form of a spawning
aggregation or migration. Garden eels (Congridae) and some
Mar Biol (2008) 153:285–295 293
123

species of moray eels (Muraenidae) spawn within their
shallow water habitats (Moyer and Zaiser 1982; Thresher
1984; Ferraris 1985), whereas some species of congrids
migrate offshore to spawn (McCleave and Miller 1994;
Miller 2002). Various species of reproductively mature
ophichthids have been observed at the surface at night
(Ross and Rohde 2003) or to make spawning migrations
towards the edge of the shelf (Cohen and Dean 1970). The
only reported observation of spawning related behavior by
chlopsid eels was by a fisherman who observed many small
eels swarming around his light at night over about 900 m of
water (Smith 1989a). The eels had enlarged abdomens and
the one specimen that was collected was a ripe female. This
species was given the name Powellichthys ventriosus, but it
has never been collected again. If Kaupichthys eels also form
spawning aggregations, then spawning during full moon
could allow them to use visual cues to locate mates at night,
or it could serve to synchronize spawning during a period of
strong tides that could transport their eggs and larvae further
offshore.
This finding of apparent spawning during full moon
around Sulawesi Island should be confirmed using larger
sample sizes, and research on the back-calculated hatching
dates of Kaupichthys leptocephali in different seasons and
in other tropical areas is needed to determine if lunar
spawning during full moon periods is characteristic of the
eels of this genus. Other chlopsids in the tropical areas of
the Indonesian Seas also should be examined to see if this
type of spawning strategy is used by other species of the
family.
References
Allen GR, Adrim M (2003) Coral reef fishes of Indonesia. Zool Stud
42:1–72
Arai T, Otake T, Tsukamoto K (1997) Drastic changes in otolith
microstructure and microchemistry accompanying the onset of
metamorphosis in the Japanese eel Anguilla japonica. Mar Ecol
Prog Ser 161:17–22
Arai T, Limbong D, Tsukamoto K (2000) Validation of otolith daily
increments in the tropical eel Anguilla celebesensis. Can J Zool
78:1078–1084
Arai T, Aoyama J, Ishikawa S, Miller MJ, Otake T, Inagaki T,
Tsukamoto K (2001a) Early life history of tropical Anguillaleptocephali in the western Pacific Ocean. Mar Biol 138:887–895
Arai T, Limbong D, Otake T, Tsukamoto K (2001b) Recruitment
mechanisms of tropical eels Anguilla spp. implications for the
evolution of oceanic migration in the genus Anguilla. Mar Ecol
Prog Ser 216:253–264
Asano H, Kubo Y, Yoshimatsu S (1978) On the morphological
change and the behavior of the leptocephali of Conger myriasterduring the period of rearing experiment. Mem Fac Agr Univ
Kinki 11:25–31
Bell GW, Witting DA, Able KW (2003) Aspects of metamorphosis
and habitat use in the conger eel, Conger oceanicus. Copeia
2003:544–552
Bishop RE, Torres JJ, Crabtree RE (2000) Chemical composition
and growth indices in leptocephalus larvae. Mar Biol 137:205–
214
Castle PHJ (1970) Distribution, larval growth, and metamorphosis of
the eel Derichthys serpentinus Gill, 1884 (Pisces, Derichthyi-
dae). Copeia 1970:444–452
Castonguay M (1987) Growth of American eel leptocephali as
revealed by otolith microstructure. Can J Zool 4:875–878
Chen HL, Tzeng WN (2006) Daily growth increment formation in
otoliths of Pacific tarpon Megalops cyprinoides during meta-
morphosis. Mar Ecol Prog Ser 312:255–263
Cieri MD, McCleave JD (2001) Validation of daily otolith increments
in glass-phase American eels Anguilla rostrata (Lesueur) during
estuarine residency. J Exp Mar Biol Ecol 257:219–227
Cohen DM, Dean D (1970) Sexual maturity and migratory behaviour
of the tropical eel, Ahlia egmontis. Nature 227:189–190
Correia AT, Antunes C, Coimbra J (2002) Aspects of the early life
history of the European conger eel (Conger conger) inferred
from the otolith microstructure of metamorphic larvae. Mar Biol
140:165–173
Correia AT, Able KW, Antunes CE, Coimbra J (2004) Early life history
of the American conger eel (Conger oceanicus) as revealed by
otolith microstructure and microchemistry of metamorphosing
leptocephali. Mar Biol 145:477–488
Correia AT, Antunes C, Wilson JM, Coimbra J (2006) An evaluation
of the otolith characteristics of Conger conger during metamor-
phosis. J Fish Biol 68:99–119
Domeier ML, Colin PL (1997) Tropical reef fish spawning aggrega-
tions defined and reviewed. Bull Mar Sci 60:698–726
Ferraris CJ (1985) Redescription and spawning behavior of the
muraenid eel Gymnothorax herrei. Copeia 1985:518–520
Fishelson L (1992) Comparative gonad morphology and sexuality of
the Muraenidae (Pisces, Teleostei). Copeia 1992:197–209
Fishelson L (1994) Comparative internal morphology of deep-sea
eels, with particular emphasis on gonads and gut structure. J Fish
Biol 44:75–101
Froese R, Pauly D (eds) (2007) FishBase. World Wide Web electronic
publication. http://www.fishbase.org, version (01/2007)
Geffen AJ (1992) Validation of otolith increment deposition rate. In:
Stevensen DK, Campana SE (eds) Otolith microstructure
examination and analysis, Can Spec Pub Fish Aquat Sci 117,
pp 101–113
Gordon AL (2005) Oceanography of the Indonesian Seas and their
throughflow. Oceanography 18:14–27
Hatayama T, Awaji T, Akitomo K (1996) Tidal currents in the
Indonesian Seas and their effect on transport and mixing. J
Geophys Res 101:12,353–12,373
Heyman WD, Kjerfve B, Graham RT, Rhodes KL, Garbutt L (2005)
Spawning aggregations of Lutjanus cyanopterus (Cuvier) on the
Belize Barrier Reef over a 6 year period. J Fish Biol 67:83–101
Isaacs JD, Kidd LW (1953) Isaacs-Kidd midwater trawl. Scripps Inst
Oceanogr Ref 53–3, Oceanogr Equip Rep No 1
Ishikawa S, Suzuki K, Inagaki T, Watanabe S, Kimura Y, Okamura
A, Otake T, Mochioka N, Suzuki Y, Hasumoto H, Oya M, Miller
MJ, Lee TW, Fricke H, Tsukamoto K (2001) Spawning time and
place of the Japanese eel Anguilla japonica in the North
Equatorial Current of the western North Pacific Ocean. Fish Sci
67:1097–1103
Johannes RE (1978) Reproductive strategies of coastal marine fishes
in the tropics. Environ Biol Fish 3:65–84
Kuroki M, Aoyama J, Miller MJ, Arai T, Sugeha HY, Minagawa G,
Wouthuyzen S, Tsukamoto K (2005) Correspondence between
otolith microstructual changes and early life history events in
Anguilla marmorata leptocephali and glass eels. Coastal Mar Sci
29:154–161
294 Mar Biol (2008) 153:285–295
123

Kuroki M, Aoyama J, Miller MJ, Wouthuyzen S, Arai T, Tsukamoto
K (2006) Contrasting patterns of growth and migration of
tropical anguillid leptocephali in the western Pacific and
Indonesian Seas. Mar Ecol Prog Ser 309:233–246
Lee TW, Byun JS (1996) Microstructural growth in otoliths of conger
eel (Conger myriaster) leptocephali during the metamorphic
stage. Mar Biol 125:259–268
Leiby MM (1979) Morphological development of the eel Myrophispunctatus (Ophichthidae) from hatching to metamorphosis, with
emphasis on the developing head skeleton. Bull Mar Sci 29:509–
521
Leiby MM (1989) Family Ophichthidae: Leptocephali. In: Bohlke EB
(ed) Fishes of Western North Atlantic, Part 9, vol 2, Mem Sears
Fdn Mar Res, New Haven, pp 764–897
Ma T, Miller MJ, Shinoda A, Minagawa G, Aoyama J, Tsukamoto K
(2005) Age and growth of Saurenchelys (Nettastomatidae) and
Dysomma (Synaphobranchidae) leptocephali in the East China
Sea. J Fish Biol 67:1619–1630
Marui M, Arai T, Miller MJ, Jellyman DJ, Tsukamoto K (2001)
Comparison of the early life history between New Zealand
temperate eels and Pacific tropical eels revealed by otolith
microstructure and microchemistry. Mar Ecol Prog Ser 213:
273–284
Matsubara K, Asano H (1959) A new eel of the genus Kaupichthys.
Copeia 1959:293–297
McCleave JD, Miller MJ (1994) Spawning of Conger oceanicus and
Conger triporiceps (Congridae) in the Sargasso Sea and subsequent
distribution of leptocephali. Environ Biol Fish 39:339–355
Miller MJ (1995) Species assemblages of leptocephali in the Sargasso
Sea and Florida Current. Mar Ecol Prog Ser 121:11–26
Miller MJ (2002) Distribution and ecology of Ariosoma balearicum(Congridae) leptocephali in the western North Atlantic. Environ
Biol Fish 63:235–252
Miller MJ, Tsukamoto K (2004) An introduction to leptocephali:
biology and identification. Ocean Research Institute, University
of Tokyo, Tokyo
Miller MJ, McCleave JD (2007) Species assemblages of leptocephali in
the southwestern Sargasso Sea. Mar Ecol Prog Ser 344:197–212
Miller MJ, Wouthuyzen S, Minagawa G, Aoyama J, Tsukamoto K
(2006) Distribution and ecology of leptocephali of the congrid
eel, Ariosoma scheelei, around Sulawesi Island, Indonesia. Mar
Biol 148:1101–1111
Moyer JT, Zaiser MJ (1982) Reproductive behavior of moray eels at
Miyade-jima. Japan J Ichthyol 28:466–468
Otake T (2003) Metamorphosis. In: Aida K, Tsukamoto K, Yamauchi
K (eds) Eel biology. Springer, Tokyo, pp 61–74
Otake T, Ishii T, Nakahara M, Nakamura R (1994) Drastic changes in
otolith strontium/calcium ratios in leptocephali and glass eels of
Japanese eel Anguilla japonica. Mar Ecol Prog Ser 112:189–193
Otake T, Ishii T, Ishii T, Nakahara M, Nakamura R (1997) Changes in
otolith strontium:calcium ratios in metamorphosing Congermyriaster leptocephali. Mar Biol 128:565–572
Pfeiler E (1999) Developmental physiology of elopomorph lepto-
cephali. Comp Biochem Physiol A Mol Integr Physiol 123:113–
128
Powles PM, Hare JA, Laban EH, Warlen SM (2006) Does eel
metamorphosis cause a breakdown in the tenets of otolith
applications? A case study using the speckled worm eel (Myrophispunctatus, Ophichthidae). Can J Fish Aquat Sci 63:1460–1468
Raju SN (1974) Distribution, growth and metamorphosis of lepto-
cephali of the garden eels, Taenioconger sp. and Gorgasia sp.
Copeia 1974:494–500
Rhodes KL, Sadovy Y (2002) Temporal and spatial trends in
spawning aggregations of camouflage grouper, Epinepheluspolyphekadion, in Pohnpei, Micronesia. Environ Biol Fish
63:27–39
Ross SW, Rohde FC (2003) Collections of ophichthid eels on the
surface at night off North Carolina. Bull Mar Sci 72:241–246
Shinoda A, Tanaka H, Kagawa H, Ohta H, Tsukamoto K (2004)
Otolith microstructural analysis of reared larvae of the Japanese
eel Anguilla japonica. Fish Sci 70:340–342
Smith DG (1969) Xenocongrid eel larvae in the western North
Atlantic. Bull Mar Sci 19:377–408
Smith DG (1989a) Chlopsidae. In: Bohlke EB (ed) Fishes of Western
North Atlantic, Part 9, vol 2, Mem Sears Fdn Mar Res, New
Haven, pp 1639–1640
Smith DG (1989b) Family Chlopsidae: Leptocephali. In: Bohlke EB
(ed) Fishes of Western North Atlantic, Part 9, vol 2, Mem Sears
Fdn Mar Res, New Haven, pp 933–942
Smith DG (1989c) Introduction to leptocephali. In: Bohlke EB (ed)
Fishes of Western North Atlantic, Part 9, vol 2, Mem Sears Fdn
Mar Res, New Haven, pp 657–668
Sugeha HY, Shinoda A, Marui M, Arai T, Tsukamoto K (2001)
Validation of otolith daily increments in the tropical eel Anguillamarmorata. Mar Ecol Prog Ser 220:291–294
Susanto RD, Moore TS, Mara J (2006) Ocean color variability in the
Indonesian Seas during the SeaWiFS era. Geochem Geophys
Geosyst 7:Q05021, doi:10.1029/2005GC001009
Tabeta O, Mochioka N (1988) Leptocephali. In: Okiyama M (ed) An
atlas of the early stage fishes in Japan. Tokai Univ Press, Tokyo,
pp 15–64 (in Japanese)
Takemura A, Rahman MS, Nakamura S, Park YJ, Takano K (2004)
Lunar cycles and reproductive activity in reef fishes with
particular attention to rabbitfishes. Fish Fisheries 5:317–328
Tanaka K, Tabeta O, Mochioka N, Yamada J, Kakuda S (1987)
Otolith microstructure and ecology of the conger eel (Congermyriaster) larvae collected in the Seto Inland Sea, Japan. Nippon
Suisan Gakk 53:543–549
Thresher RE (1984) Reproduction in reef fishes. Tropical Fish
Hobbyist Publications, Neptune City
Tsukamoto K (1989) Otolith daily increments in the Japanese eel.
Bull Jpn Soc Sci Fish 55:1017–1021
Tsukamoto K (1990) Recruitment mechanism of the eel, Anguillajaponica, to the Japanese coast. J Fish Biol 36:659–671
Tsukamoto K 2006 Spawning of eels near a seamount. Nature
439:929
Tsukamoto K, Otake T, Mochioka N, Lee TW, Fricke H, Inagaki T,
Aoyama J, Ishikawa S, Kimura S, Miller MJ, Hasumoto H, Oya
M, Suzuki Y (2003) Seamounts, new moon and eel spawning:
the search for the spawning site of the Japanese eel. Environ Biol
Fish 66:221–229
Wallace CC (1999) The Togian Islands: coral reefs with a unique
coral fauna and an hypothesized Tethys Sea signature. Coral
Reefs 18:162
Wang CH, Tzeng WN (2000) The timing of metamorphosis and
growth rates of American and European eel leptocephali: a
mechanism of larval segregative migration. Fish Res 46:191–205
Wouthuyzen S, Miller MJ, Aoyama J, Minagawa G, Sugeha YH,
Suharti S, Inagaki T, Tsukamoto K (2005) Biodiversity of
anguilliform leptocephali in the central Indonesian Seas. Bull
Mar Sci 77:209–224
Mar Biol (2008) 153:285–295 295
123
Related Documents


![Untitled-2 []...Eben Coffin4 Eben Coffin John Darling7 paul Painter Bell Coffin John Darling King Grounds South Seas South Seas South Seas South Seas Brazil South Seas Production 550](https://static.cupdf.com/doc/110x72/60e8bb276bd41c47e12f2272/untitled-2-eben-coffin4-eben-coffin-john-darling7-paul-painter-bell-coffin.jpg)








