Distribution and diversity of type III secretion system-like genes in saprophytic and phytopathogenic fluorescent pseudomonads Sylvie Mazurier, M elanie Lemunier, S everine Siblot, Christophe Mougel, Philippe Lemanceau * INRA-CMSE UMR ‘‘Microbiologie et G eochimie des Sols’’, 17 rue Sully, BP86510, 21065 Dijon cedex, France Received 2 March 2004; received in revised form 28 April 2004; accepted 29 April 2004 First published online 20 May 2004 Abstract Type three secretion systems (TTSSs) are protein translocation mechanisms associated with bacterial pathogenicity in host plants, and hypersensitive reactions in non-host plants. Distribution and diversity of TTSS-like genes within a collection of sap- rophytic and phytopathogenic fluorescent pseudomonads were characterized. This collection included 16 strains belonging to 13 pathogenic species, and 87 strains belonging to five saprophytic species isolated from plant rhizosphere and soil. Presence of conserved hypersensitive reaction/pathogenicity (hrp) genes (hrcRST) was assessed both by PCR using primers designed to amplify the corresponding sequence and by dot-blot hybridization using a PCR-amplified hrcRST fragment as a probe. PCR allowed the detection of TTSS-like genes in 75% and 32% of the phytopathogenic and saprophytic strains, respectively, and dot-blot hybrid- ization in 100% and 49% of the phytopathogenic and saprophytic strains, respectively. The restriction fragment length polymor- phism (RFLP) of 26 amplified hrcRST fragments revealed a considerable diversity. Twenty-one distinct RFLP types were identified and one hrcRST fragment was sequenced per RFLP type. The obtained hrcRST sequences clustered into three groups. Two of these groups included both phytopathogenic and saprophytic strains. The diversity of 16S rRNA genes, commonly used as an evolution marker, was characterized using PCR-RFLP. Polymorphism of the 16S rRNA genes corresponded to that of hrcRST genes, sug- gesting that these genes have followed a similar evolution. However, the occurrence of few mismatches suggests that sometimes TTSS-like genes might have undergone horizontal genetic transfer. Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Type III secretion system; hrp genes; hrcRST; Rhizosphere; Fluorescent pseudomonads 1. Introduction A type III protein secretion system (TTSS) is de- scribed as a macromolecular translocation apparatus that enables a number of gram-negative pathogenic bacteria to inject virulence determinants directly into the cells of their eukaryotic hosts [1]. The importance of TTSSs in bacterial pathogenicity has been demonstrated for various genera of animal pathogens (Escherichia, Pseudomonas, Salmonella, Shigella, and Yersinia) and of plant pathogens (Erwinia, Pantoea, Pseudomonas, Rals- tonia, and Xanthomonas) [2]. In phytopathogenic bacteria, the TTSS is encoded by hypersensitive reaction/pathogenicity (hrp) genes, which are associated with the ability to elicit the hypersensitive reaction (HR) on non-host plants and to cause disease in the host plant [3]. Depending on their genetic organi- zation and mode of regulation, these genes are found in clusters of type I for Erwinia spp., Pantoea spp. and Pseudomonas syringae, and in clusters of type II for Ralstonia and Xanthomonas [4]. Nine hrp genes, that code for the core part of the TTSS and are conserved among all animal and plant pathogens, are called hrc (for hrp conserved genes) [5]. Eight of the Hrc proteins (HrcV, N, Q 1 ,Q 2 , R, S, T, U) have homologues in * Corresponding author. Tel.: +33-3-80-69-30-56; fax: +33-3-80-69- 32-24/26. E-mail address: [email protected] (P. Lemanceau). 0168-6496/$22.00 Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsec.2004.04.019 FEMS Microbiology Ecology 49 (2004) 455–467 www.fems-microbiology.org

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

FEMS Microbiology Ecology 49 (2004) 455–467
www.fems-microbiology.org
Distribution and diversity of type III secretion system-like genesin saprophytic and phytopathogenic fluorescent pseudomonads
Sylvie Mazurier, M�elanie Lemunier, S�everine Siblot, Christophe Mougel,Philippe Lemanceau *
INRA-CMSE UMR ‘‘Microbiologie et G�eochimie des Sols’’, 17 rue Sully, BP86510, 21065 Dijon cedex, France
Received 2 March 2004; received in revised form 28 April 2004; accepted 29 April 2004
First published online 20 May 2004
Abstract
Type three secretion systems (TTSSs) are protein translocation mechanisms associated with bacterial pathogenicity in host
plants, and hypersensitive reactions in non-host plants. Distribution and diversity of TTSS-like genes within a collection of sap-
rophytic and phytopathogenic fluorescent pseudomonads were characterized. This collection included 16 strains belonging to 13
pathogenic species, and 87 strains belonging to five saprophytic species isolated from plant rhizosphere and soil. Presence of
conserved hypersensitive reaction/pathogenicity (hrp) genes (hrcRST) was assessed both by PCR using primers designed to amplify
the corresponding sequence and by dot-blot hybridization using a PCR-amplified hrcRST fragment as a probe. PCR allowed the
detection of TTSS-like genes in 75% and 32% of the phytopathogenic and saprophytic strains, respectively, and dot-blot hybrid-
ization in 100% and 49% of the phytopathogenic and saprophytic strains, respectively. The restriction fragment length polymor-
phism (RFLP) of 26 amplified hrcRST fragments revealed a considerable diversity. Twenty-one distinct RFLP types were identified
and one hrcRST fragment was sequenced per RFLP type. The obtained hrcRST sequences clustered into three groups. Two of these
groups included both phytopathogenic and saprophytic strains. The diversity of 16S rRNA genes, commonly used as an evolution
marker, was characterized using PCR-RFLP. Polymorphism of the 16S rRNA genes corresponded to that of hrcRST genes, sug-
gesting that these genes have followed a similar evolution. However, the occurrence of few mismatches suggests that sometimes
TTSS-like genes might have undergone horizontal genetic transfer.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Type III secretion system; hrp genes; hrcRST; Rhizosphere; Fluorescent pseudomonads
1. Introduction
A type III protein secretion system (TTSS) is de-
scribed as a macromolecular translocation apparatus
that enables a number of gram-negative pathogenic
bacteria to inject virulence determinants directly into the
cells of their eukaryotic hosts [1]. The importance ofTTSSs in bacterial pathogenicity has been demonstrated
for various genera of animal pathogens (Escherichia,
Pseudomonas, Salmonella, Shigella, and Yersinia) and of
* Corresponding author. Tel.: +33-3-80-69-30-56; fax: +33-3-80-69-
32-24/26.
E-mail address: [email protected] (P. Lemanceau).
0168-6496/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsec.2004.04.019
plant pathogens (Erwinia, Pantoea, Pseudomonas, Rals-
tonia, and Xanthomonas) [2].
In phytopathogenic bacteria, the TTSS is encoded by
hypersensitive reaction/pathogenicity (hrp) genes, which
are associated with the ability to elicit the hypersensitive
reaction (HR) on non-host plants and to cause disease in
the host plant [3]. Depending on their genetic organi-zation and mode of regulation, these genes are found in
clusters of type I for Erwinia spp., Pantoea spp. and
Pseudomonas syringae, and in clusters of type II for
Ralstonia and Xanthomonas [4]. Nine hrp genes, that
code for the core part of the TTSS and are conserved
among all animal and plant pathogens, are called hrc
(for hrp conserved genes) [5]. Eight of the Hrc proteins
(HrcV, N, Q1, Q2, R, S, T, U) have homologues in
. Published by Elsevier B.V. All rights reserved.

456 S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467
flagella biogenesis and are thought to form a specific
inner membrane transport channel [6]. The remaining
Hrc protein (HrcC) belongs to the large family of sec-
retins, which is involved in various processes of trans-
port [7]. HrcC is thought to constitute the outermembrane transport channel [8].
TTSSs have recently been reported in non-pathogenic
bacteria. TTSSs were described in the symbiotic bacte-
ria, Rhizobium sp. parasponia NGR234, Sinorhizobium
fredii, Mesorhizobium loti and Bradyrhizobium japoni-
cum, responsible for biological nitrogen fixation in the
roots of legumes [9]. In these bacteria, TTSSs have been
implicated in the plant-bacteria molecular dialoguemediating the establishment and the activity of the
symbiotic association [9,10].
Fluorescent pseudomonads include strains that im-
prove plant growth and health [11], and strains patho-
genic to plants [12]. Among the pathogenic fluorescent
pseudomonads, other than the species P. syringae, the
presence of a TTSS has only been reported for P. viri-
diflava on the basis of the presence of hrp genes [13,14].The homologues of hrp genes were also described in few
strains of P. fluorescens known to be non-phytopatho-
genic or even beneficial for plant growth and health:
PfG32R [15], C7 [16] and SBW25 [17]. Four of the nine
hrc genes were described in C7 [16]. This strain, previ-
ously reported to suppress fusarium crown and root rot
of tomato [18], was shown to induce a hypersensitive
reaction in this plant species after leaf infiltration [16]. Atype III gene cluster (Rsp) resembling the type III gene
cluster of P. syringae (Hrp) was identified in SBW25
[17]. Using homologues of some of the rsc genes that
encode the TTSS described in P. fluorescens SBW25,
Preston et al. [17] have shown that these genes were also
present in other strains of P. fluorescens and P. putida.
However, the number of strains tested was limited and
these strains only belonged to two saprophytic species.Therefore, data on the distribution of TTSS-like genes
in phytopathogenic and saprophytic fluorescent pseu-
domonads appears to be restricted to few species and,
except for P. syringae, to a limited number of strains.
Furthermore, no information on the polymorphism of
TTSS-like genes in saprophytic fluorescent pseudomo-
nads is available.
The objectives of this study were to evaluate thedistribution of TTSSs in saprophytic and phytopatho-
genic fluorescent pseudomonads, and to describe the
diversity of the corresponding TTSS-like genes. We
studied 103 strains of fluorescent pseudomonads. This
collection included 16 strains belonging to 13 species
pathogenic to plants, and 87 strains belonging to five
saprophytic species. Presence of the hrcRST sequence in
these strains, corresponding to a specific hrc gene suc-cession typical of type I hrp clusters was evaluated by
PCR with specific primers and by dot-blot hybridization
using the P. fluorescens C7 hrcRST sequence as a probe.
The polymorphism of the hrcRST sequence was assessed
by restriction fragment length polymorphism (RFLP)
and by sequencing one fragment per RFLP type. The
genetic diversity of the 16S rRNA genes, commonly
used as an evolution marker, was studied in parallel byPCR-RFLP in order to analyse the relationships be-
tween that gene and the hrcRST sequence.
2. Materials and methods
2.1. Bacterial strains and culture conditions
Bacterial stains used in this work are presented in
Table 1. The collection included (i) 17 type-strains, (ii)
five reference strains, (iii) 65 strains isolated from vari-
ous plant rhizospheres (28 from corn, 24 from tomato,
eight from flax, and five from different plant species),
and (iv) 16 strains isolated from bulk soils. The type-
strains included 14 phytopathogenic strains and two
strains isolated from soils. The reference strains included(i) three PGPR and biocontrol agents P. fluorescens
CFBP2392 (syn. A6) [18,19], CFBP5759 (syn. C7) [20]
and CFBP5935 (syn. F113) [21–23], and (ii) two phy-
topathogenic strains (P. syringae SD68 and 2027-37)
used as PCR controls. The type strain P. lini
CFBP5737T, 14 of the rhizosphere strains (CFBP5733 to
CFBP5736, CFBP5738, CFBP5743 to CFBP5746,
CFBP5755 to CFBP5758, and CFBP5760) and the six-teen soil-borne strains were previously shown to be
representative of the diversity of a larger bacterial col-
lection (340 strains) isolated from flax and tomato cul-
tivated in soils of Dijon and Chateaurenard (France),
and from these soils kept uncultivated [24,25]. The 52
remaining rhizosphere strains were described by Bossis
[26] and kindly provided by L. Gardan (UMR77 PaV�eINRA-INH-Universit�e, Angers, France). Altogetherthis collection included 16 phytopathogenic strains be-
longing to 13 species and 87 saprophytic strains be-
longing to five species.
Pseudomonas strains were grown at 25 �C in LB
medium [27]. For solid medium, 15 g l�1 of agar were
added.
2.2. Extraction of total genomic DNA and PCR ampli-
fications
Total DNA extractions were performed using stan-
dard methods [28]. PCR reactions were conducted in a
programmable thermal cycler (PTC-200, MJ Research)
and primers were synthesized by Eurogentec (Angers,
France). DNA sequences of two conserved hrp genes
(hrcR and hrcT) of phytopathogenic bacteria harbour-ing hrp clusters of type I [4] and belonging to different
species (P. syringae, Erwinia amylovora, and Pantoea
agglomerans formerly E. herbicola), were aligned using
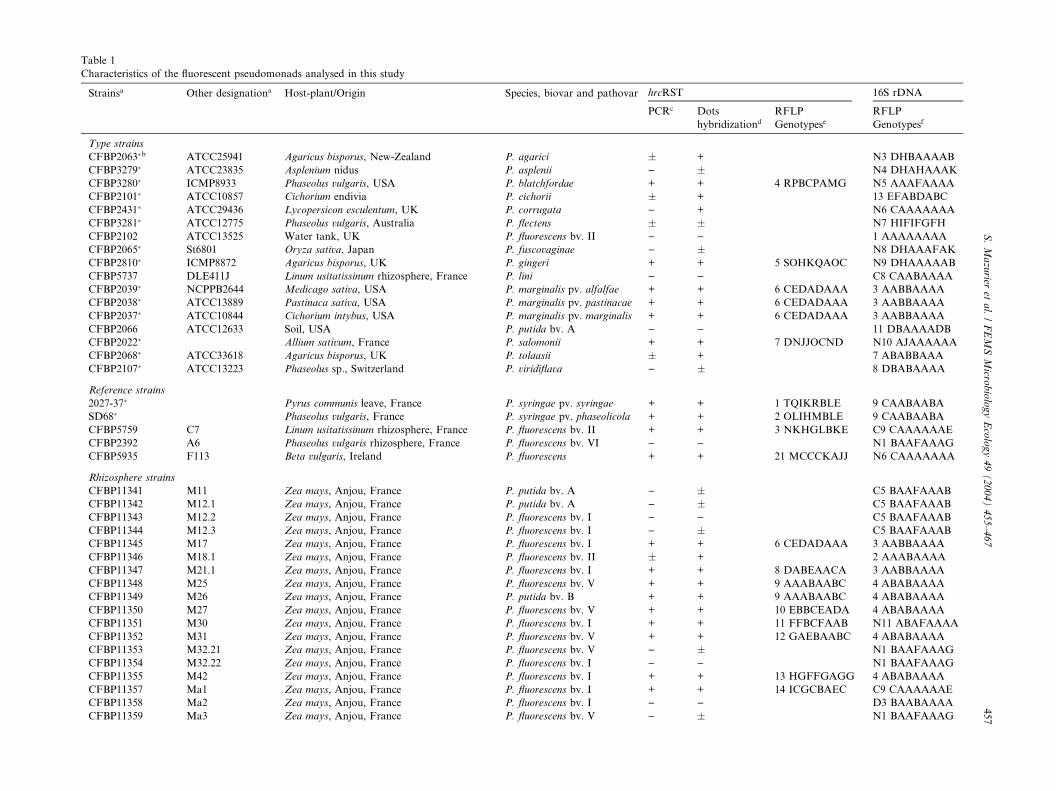
Table 1
Characteristics of the fluorescent pseudomonads analysed in this study
Strainsa Other designationa Host-plant/Origin Species, biovar and pathovar hrcRST 16S rDNA
PCRc Dots
hybridizationd
RFLP
GenotypeseRFLP
Genotypesf
Type strains
CFBP2063�b ATCC25941 Agaricus bisporus, New-Zealand P. agarici � + N3 DHBAAAAB
CFBP3279� ATCC23835 Asplenium nidus P. asplenii ) � N4 DHAHAAAK
CFBP3280� ICMP8933 Phaseolus vulgaris, USA P. blatchfordae + + 4 RPBCPAMG N5 AAAFAAAA
CFBP2101� ATCC10857 Cichorium endivia P. cichorii � + 13 EFABDABC
CFBP2431� ATCC29436 Lycopersicon esculentum, UK P. corrugata ) + N6 CAAAAAAA
CFBP3281� ATCC12775 Phaseolus vulgaris, Australia P. flectens � � N7 HIFIFGFH
CFBP2102 ATCC13525 Water tank, UK P. fluorescens bv. II ) ) 1 AAAAAAAA
CFBP2065� St6801 Oryza sativa, Japan P. fuscovaginae ) � N8 DHAAAFAK
CFBP2810� ICMP8872 Agaricus bisporus, UK P. gingeri + + 5 SOHKQAOC N9 DHAAAAAB
CFBP5737 DLE411J Linum usitatissinum rhizosphere, France P. lini ) ) C8 CAABAAAA
CFBP2039� NCPPB2644 Medicago sativa, USA P. marginalis pv. alfalfae + + 6 CEDADAAA 3 AABBAAAA
CFBP2038� ATCC13889 Pastinaca sativa, USA P. marginalis pv. pastinacae + + 6 CEDADAAA 3 AABBAAAA
CFBP2037� ATCC10844 Cichorium intybus, USA P. marginalis pv. marginalis + + 6 CEDADAAA 3 AABBAAAA
CFBP2066 ATCC12633 Soil, USA P. putida bv. A ) ) 11 DBAAAADB
CFBP2022� Allium sativum, France P. salomonii + + 7 DNJJOCND N10 AJAAAAAA
CFBP2068� ATCC33618 Agaricus bisporus, UK P. tolaasii � + 7 ABABBAAA
CFBP2107� ATCC13223 Phaseolus sp., Switzerland P. viridiflava ) � 8 DBABAAAA
Reference strains
2027-37� Pyrus communis leave, France P. syringae pv. syringae + + 1 TQIKRBLE 9 CAABAABA
SD68� Phaseolus vulgaris, France P. syringae pv. phaseolicola + + 2 OLIHMBLE 9 CAABAABA
CFBP5759 C7 Linum usitatissinum rhizosphere, France P. fluorescens bv. II + + 3 NKHGLBKE C9 CAAAAAAE
CFBP2392 A6 Phaseolus vulgaris rhizosphere, France P. fluorescens bv. VI ) ) N1 BAAFAAAG
CFBP5935 F113 Beta vulgaris, Ireland P. fluorescens + + 21 MCCCKAJJ N6 CAAAAAAA
Rhizosphere strains
CFBP11341 M11 Zea mays, Anjou, France P. putida bv. A ) � C5 BAAFAAAB
CFBP11342 M12.1 Zea mays, Anjou, France P. putida bv. A ) � C5 BAAFAAAB
CFBP11343 M12.2 Zea mays, Anjou, France P. fluorescens bv. I ) ) C5 BAAFAAAB
CFBP11344 M12.3 Zea mays, Anjou, France P. fluorescens bv. I ) � C5 BAAFAAAB
CFBP11345 M17 Zea mays, Anjou, France P. fluorescens bv. I + + 6 CEDADAAA 3 AABBAAAA
CFBP11346 M18.1 Zea mays, Anjou, France P. fluorescens bv. II � + 2 AAABAAAA
CFBP11347 M21.1 Zea mays, Anjou, France P. fluorescens bv. I + + 8 DABEAACA 3 AABBAAAA
CFBP11348 M25 Zea mays, Anjou, France P. fluorescens bv. V + + 9 AAABAABC 4 ABABAAAA
CFBP11349 M26 Zea mays, Anjou, France P. putida bv. B + + 9 AAABAABC 4 ABABAAAA
CFBP11350 M27 Zea mays, Anjou, France P. fluorescens bv. V + + 10 EBBCEADA 4 ABABAAAA
CFBP11351 M30 Zea mays, Anjou, France P. fluorescens bv. I + + 11 FFBCFAAB N11 ABAFAAAA
CFBP11352 M31 Zea mays, Anjou, France P. fluorescens bv. V + + 12 GAEBAABC 4 ABABAAAA
CFBP11353 M32.21 Zea mays, Anjou, France P. fluorescens bv. V ) � N1 BAAFAAAG
CFBP11354 M32.22 Zea mays, Anjou, France P. fluorescens bv. I ) ) N1 BAAFAAAG
CFBP11355 M42 Zea mays, Anjou, France P. fluorescens bv. I + + 13 HGFFGAGG 4 ABABAAAA
CFBP11357 Ma1 Zea mays, Anjou, France P. fluorescens bv. I + + 14 ICGCBAEC C9 CAAAAAAE
CFBP11358 Ma2 Zea mays, Anjou, France P. fluorescens bv. I ) ) D3 BAABAAAA
CFBP11359 Ma3 Zea mays, Anjou, France P. fluorescens bv. V ) � N1 BAAFAAAG
S.Mazurier
etal./FEMSMicro
biologyEcology49(2004)455–467
457
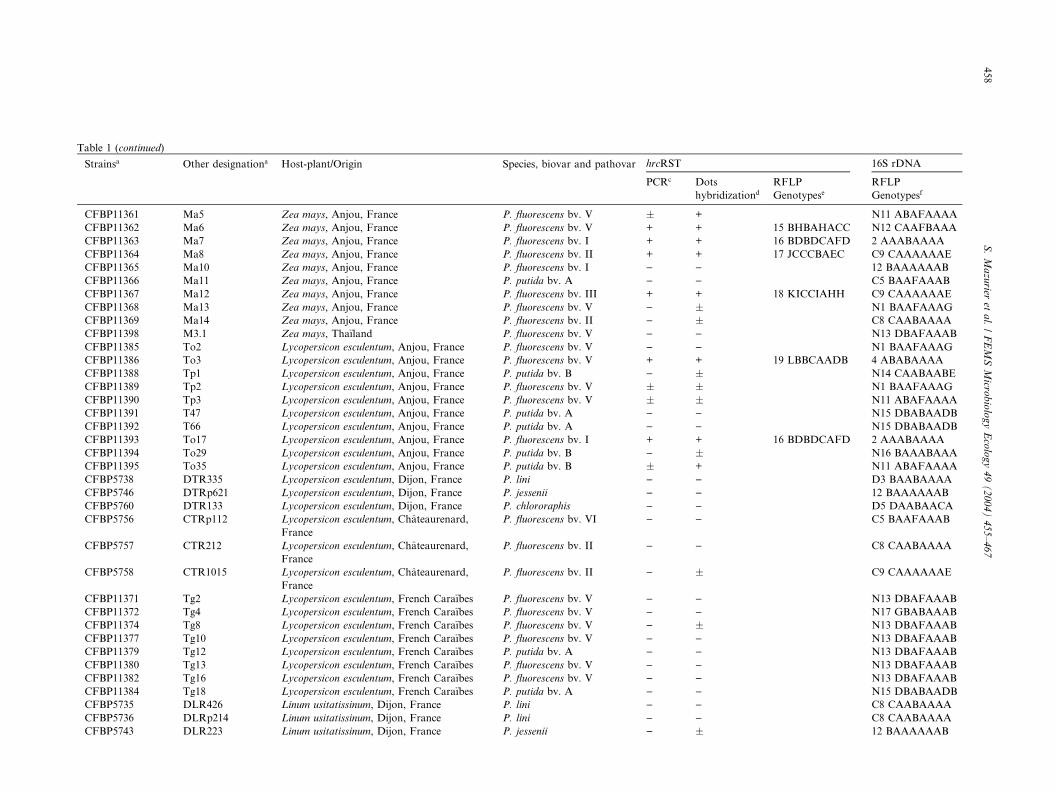
Table 1 (continued)
Strainsa Other designationa Host-plant/Origin Species, biovar and pathovar hrcRST 16S rDNA
PCRc Dots
hybridizationd
RFLP
GenotypeseRFLP
Genotypesf
CFBP11361 Ma5 Zea mays, Anjou, France P. fluorescens bv. V � + N11 ABAFAAAA
CFBP11362 Ma6 Zea mays, Anjou, France P. fluorescens bv. V + + 15 BHBAHACC N12 CAAFBAAA
CFBP11363 Ma7 Zea mays, Anjou, France P. fluorescens bv. I + + 16 BDBDCAFD 2 AAABAAAA
CFBP11364 Ma8 Zea mays, Anjou, France P. fluorescens bv. II + + 17 JCCCBAEC C9 CAAAAAAE
CFBP11365 Ma10 Zea mays, Anjou, France P. fluorescens bv. I ) ) 12 BAAAAAAB
CFBP11366 Ma11 Zea mays, Anjou, France P. putida bv. A ) ) C5 BAAFAAAB
CFBP11367 Ma12 Zea mays, Anjou, France P. fluorescens bv. III + + 18 KICCIAHH C9 CAAAAAAE
CFBP11368 Ma13 Zea mays, Anjou, France P. fluorescens bv. V ) � N1 BAAFAAAG
CFBP11369 Ma14 Zea mays, Anjou, France P. fluorescens bv. II ) � C8 CAABAAAA
CFBP11398 M3.1 Zea mays, Tha€ıland P. fluorescens bv. V ) ) N13 DBAFAAAB
CFBP11385 To2 Lycopersicon esculentum, Anjou, France P. fluorescens bv. V ) ) N1 BAAFAAAG
CFBP11386 To3 Lycopersicon esculentum, Anjou, France P. fluorescens bv. V + + 19 LBBCAADB 4 ABABAAAA
CFBP11388 Tp1 Lycopersicon esculentum, Anjou, France P. putida bv. B ) � N14 CAABAABE
CFBP11389 Tp2 Lycopersicon esculentum, Anjou, France P. fluorescens bv. V � � N1 BAAFAAAG
CFBP11390 Tp3 Lycopersicon esculentum, Anjou, France P. fluorescens bv. V � � N11 ABAFAAAA
CFBP11391 T47 Lycopersicon esculentum, Anjou, France P. putida bv. A ) ) N15 DBABAADB
CFBP11392 T66 Lycopersicon esculentum, Anjou, France P. putida bv. A ) ) N15 DBABAADB
CFBP11393 To17 Lycopersicon esculentum, Anjou, France P. fluorescens bv. I + + 16 BDBDCAFD 2 AAABAAAA
CFBP11394 To29 Lycopersicon esculentum, Anjou, France P. putida bv. B ) � N16 BAAABAAA
CFBP11395 To35 Lycopersicon esculentum, Anjou, France P. putida bv. B � + N11 ABAFAAAA
CFBP5738 DTR335 Lycopersicon esculentum, Dijon, France P. lini ) ) D3 BAABAAAA
CFBP5746 DTRp621 Lycopersicon esculentum, Dijon, France P. jessenii ) ) 12 BAAAAAAB
CFBP5760 DTR133 Lycopersicon esculentum, Dijon, France P. chlororaphis ) ) D5 DAABAACA
CFBP5756 CTRp112 Lycopersicon esculentum, Chateaurenard,
France
P. fluorescens bv. VI ) ) C5 BAAFAAAB
CFBP5757 CTR212 Lycopersicon esculentum, Chateaurenard,
France
P. fluorescens bv. II ) ) C8 CAABAAAA
CFBP5758 CTR1015 Lycopersicon esculentum, Chateaurenard,
France
P. fluorescens bv. II ) � C9 CAAAAAAE
CFBP11371 Tg2 Lycopersicon esculentum, French Cara€ıbes P. fluorescens bv. V ) ) N13 DBAFAAAB
CFBP11372 Tg4 Lycopersicon esculentum, French Cara€ıbes P. fluorescens bv. V ) ) N17 GBABAAAB
CFBP11374 Tg8 Lycopersicon esculentum, French Cara€ıbes P. fluorescens bv. V ) � N13 DBAFAAAB
CFBP11377 Tg10 Lycopersicon esculentum, French Cara€ıbes P. fluorescens bv. V ) ) N13 DBAFAAAB
CFBP11379 Tg12 Lycopersicon esculentum, French Cara€ıbes P. putida bv. A ) ) N13 DBAFAAAB
CFBP11380 Tg13 Lycopersicon esculentum, French Cara€ıbes P. fluorescens bv. V ) ) N13 DBAFAAAB
CFBP11382 Tg16 Lycopersicon esculentum, French Cara€ıbes P. fluorescens bv. V ) ) N13 DBAFAAAB
CFBP11384 Tg18 Lycopersicon esculentum, French Cara€ıbes P. putida bv. A ) ) N15 DBABAADB
CFBP5735 DLR426 Linum usitatissinum, Dijon, France P. lini ) ) C8 CAABAAAA
CFBP5736 DLRp214 Linum usitatissinum, Dijon, France P. lini ) ) C8 CAABAAAA
CFBP5743 DLR223 Linum usitatissinum, Dijon, France P. jessenii ) � 12 BAAAAAAB
458
S.Mazurier
etal./FEMSMicro
biologyEcology49(2004)455–467

CFBP5744 DLR228 Linum usitatissinum, Dijon, France P. jessenii ) ) C5 BAAFAAAB
CFBP5745 DLE3216 Linum usitatissinum, Dijon, France P. jessenii ) ) 12 BAAAAAAB
CFBP5733 CLRp812 Linum usitatissinum, Chateaurenard, France P. lini ) ) 9 CAABAABA
CFBP5734 CLE513 Linum usitatissinum, Chateaurenard, France P. lini ) ) 9 CAABAABA
CFBP5755 CLR711 Linum usitatissinum, Chateaurenard, France P. fluorescens bv. VI ) ) C5 BAAFAAAB
CFBP11397 L26.1 Lactuca satvia, Anjou, France P. fluorescens bv. V ) ) N1 BAAFAAAG
CFBP11400 M114 Glycine max, Ireland P. fluorescens bv. I ) ) N1 BAAFAAAG
CFBP11401 CTQMP26 Glycine max, Ireland P. fluorescens bv. V � � C5 BAAFAAAB
CFBP11402 Persica vulgaris, Algeria Pseudomonas sp. + + 20 QJBCJAII N11 ABAFAAAA
CFBP11403 Prunus armeniaca, Algeria Pseudomonas sp. ) ) N13 DBAFAAAB
Soil strains
CFBP5741 DS131 Dijon, France P. jessenii ) � 12 BAAAAAAB
CFBP5742 DS1026 Dijon, France P. jessenii ) ) 12 BAAAAAAB
CFBP5747 DS824 Dijon, France P. jessenii ) ) 12 BAAAAAAB
DS134 Dijon, France P. chlororaphis ) ) D5 DAABAACA
DS222 Dijon, France P. chlororaphis � � D5 DAABAACA
DS321 Dijon, France P. putida bv. A ) ) 12 BAAAAAAB
DS624 Dijon, France P. putida bv. A � � 12 BAAAAAAB
DS924 Dijon, France P. chlororaphis ) ) D5 DAABAACA
CFBP5732 CS611 Chateaurenard, France P. lini ) ) 9 CAABAABA
CFBP5739 CS111 Chateaurenard, France P. putida bv. A ) ) C1 BAABAAAB
CFBP5740 CS413 Chateaurenard, France P. putida bv. A ) ) C1 BAABAAAB
CS215 Chateaurenard, France P. putida bv. A ) ) C1 BAABAAAB
CS411 Chateaurenard, France P. fluorescens bv. II ) � 9 CAABAABA
CS511 Chateaurenard, France P. fluorescens bv. II � � 9 CAABAABA
CS712 Chateaurenard, France P. putida bv. A ) � C1 BAABAAAB
CS2114 Chateaurenard, France P. putida bv. A � � C1 BAABAAABaCFBP, Collection Franc�aise des Bacteries Phytopathog�enes, INRA Beaucouz�e, France; ATCC, American Type Culture Collection, U ersity Boulevard, Manassas, VA, USA; ICMP, In-
ternational Collection of Micro-organisms from Plants, Landcare Research, Auckland, New Zealand; NCPPB, National Collection of Plant P ogenic Bacteria, Central Science Laboratory, York,
UK; other strain names are laboratory designations.b Strains belonging to phytopathogenic species are indicated with an asterisk.cþ, strong PCR product of expected size and hybridizing with the probe C7hrcRST; �, weak PCR product of expected size hybridizing h the probe C7hrcRST; ), no PCR product and no
hybridization signal.dDot blot hybridization of total DNA with the probe C7hrcRST; þ, hybridization signal of intensity comparable to that of the C7 DNA ed as a control; �, hybridization signal of intensity
significantly weakest than that of the control; ), no visible hybridization signal.eNumbers designate the hrcRST types, and letters designate the patterns obtained with the restriction enzymes AluI, AvaII, DdeI, HinfI, spI, PstI, RsaI, and SphI, respectively.f Numbers designate the 16S rDNA types as described by Laguerre et al. [31], numbers following ‘‘C’’ and ‘‘D’’ designate 16S rDNA types as scribed by Latour et al. [25], numbers following ‘‘N’’
designate new 16S rDNA types described in this study, and letters designate the patterns obtained with the restriction enzymes AluI, DdeI, H II, MseI, MspI, NdeII, RsaI, and TaqI, respectively.
S.Mazurier
etal./FEMSMicro
biologyEcology49(2004)455–467
459
niv
ath
wit
us
M
de
aeI

460 S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467
CLUSTALW 1.6 [29]. Based on these alignments a pair
of oligonucleotide primers, HRCR8092 50-CCITT(C/T)ATCGT(C/T)AT(C/T)GA(C/T)(C/T)T-30 and HRCT
8986 50-CTGTCCCAGATIAICTGIGT-30 (where I in-
dicates inosine), was designed to amplify a part of theoperon U of the hrp cluster of type I including the 30 endof hrcR (26%), hrcS (100%), and the 50 end of hrcT
(42%) (Fig. 1). PCRs were conducted in a 25 ll reactionvolume. Reaction mixtures contained 150 ng of purified
DNA and 1.25 U of Taq DNA polymerase (Q-Biogen,
Illkirch, France) in the corresponding buffer (10 mM
Tris–HCl, pH 9.0 at 25 �C, 50 mM KCl, 1.5 mMMgCl2,
0.1% Triton X-100, 0.2 mgml�1 BSA, Q-Biogen, Ill-kirch, France). Final concentrations of each primer and
of dNTPs were 0.5 and 200 lM, respectively. Thermal
cycling consisted of an initial denaturation step at 95 �Cfor 3 min followed by 35 cycles of denaturation at 94 �Cfor 1 min, annealing at 41 �C for 1 min, and elongation
at 72 �C for 2 min, with a final elongation step at 72 �Cfor 3 min.
16S rRNA genes were amplified with primer pair fD1(50-AGAGTTTGATCCTGGCTCAG-30) and rD1 (50-AAGGAGGTGATCCAGCC-30) [30]. Reactions were
performed in a total volume of 50 ll, by mixing 50 ng of
DNA with 0.2 lM of each primer, 20 lM each of dATP,
dCTP, dGTP, dTTP, 2.5 U of Taq DNA polymerase in
the corresponding buffer (see above). Amplification re-
actions started with an initial denaturation step (3 min
at 95 �C) followed by 35 cycles of 1 min at 94 �C, 1 minat 55 �C, and 2 min at 72 �C, with a final extension of 3
min at 72 �C.Aliquots (5 ll) of the PCR products were analysed by
electrophoresis in 0.9% agarose gels stained with ethi-
dium bromide and photographed underUV illumination.
2.3. DNA hybridization
Hybridizations were performed on Pall Biodyne Plus
membranes (VWR, France). For DNA dot blots, 1 lg oftotal DNA was spotted onto the membrane after a de-
naturation of 10 min in a boiling water bath. For PCR
products, DNA was transferred from agarose gels by
vacuum blotting. DNA was fixed on membranes during
30 min at 80 �C. The C7hrcRST probe used for dot-blot
HRCR8092
HRCT8986
hrcR hrcThrcS
897 bp
Fig. 1. Scheme showing the position and orientation of the primers
HRCR8092 and HRCT8986 (black arrowheads) with respect to hrcR,
hrcS, and hrcT of P. syringae pv. syringae NPS3121 (GenBank Ac-
cession No. AF043444).
hybridization and for checking the specificity of PCR
amplifications was obtained by labeling the hrcRST
fragment yielded by PCR amplification of DNA from P.
fluorescens C7 (syn. CFBP5759). Digoxigenin labeling of
the DNA probe, DNA hybridization and probe detectionwere done using a non-radioactive DNA labeling and
detection kit (Roche Molecular Biochemicals, Meylan,
France) and applying high stringency conditions (hy-
bridization at 68 �C, first washing step in 0.1% SDS and
SSC2�, secondwashing step in 0.1%SDSandSSC0.1�).
2.4. RFLP of hrcRST and 16S rRNA genes
Aliquots (6 ll) of PCR products were digested over
night with 10 U of the following restriction endonuc-
leases purchased from Q-Biogen (Illkirch, France): AluI,
AvaII, DdeI, HinfI, MspI, PstI, RsaI, and SphI for
hrcRST, and AluI, DdeI, HaeIII, MseI, MspI, NdeII,
RsaI, and TaqI for 16S rDNA. The eight enzymes used
for hrcRST PCR fragments were selected on the basis of
restriction maps. They were chosen to discriminate allknown hrcRST sequences of the phytopathogens
aligned for primer design and the hrcRST sequence of
C7 (syn. CFBP5759) (EMBL Accession No. AJ271105).
The eight enzymes used for the amplified 16S rRNA
gene fragments were selected among the 13 used by
Laguerre et al. [31] for their ability to discriminate
strains belonging to different species of fluorescent
pseudomonads. The restriction fragments were sepa-rated by electrophoresis in TAE buffer (40 mM Tris–
HCl, pH 7.9, containing 4 mM sodium acetate and 1
mM EDTA) using 3% (w/v) Small Fragment agarose
(Appligene, Illkirch, France). Electrophoresis was car-
ried out at 100 V. After staining with ethidium bromide,
gels were photographed under UV illumination. For
each gene, each strain was assigned a composite type
defined by the combination of the patterns obtainedwith the different restriction endonucleases.
The computer program TREECON for Windows
1.3b [32] was used to estimate the relationships between
the hrcRST sequences on the basis of CLUSTALW 1.6
[29] alignments and for bootstrap analysis. Distance
estimation was computed according to Jukes and Can-
tor [33]. The matrix of distances was displayed as a
dendrogram using the UPGMA method [34]. Thecomputer program NTSYSpc 2.02k [35] was used to
compare the 16S rDNA types from the fragments ob-
tained with the different restriction endonucleases. The
pairwise Jaccard coefficients of similarity were computed
[36]. The matrix of similarities was displayed as a den-
drogram using the UPGMA method.
2.5. Cloning and sequencing PCR fragments
Before cloning, PCR fragments were purified by
electrophoresis in a 0.9% agarose gel in TAE, excised

S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467 461
and extracted from the gel by electro-elution using a
Biotrap BT 1000 (Schleicher & Schuell, Dassel, Ger-
many) according to the instructions of the manufac-
turer. The pGEM-T Easy Vector System II (Promega,
Charbonni�eres, France) was used for cloning as rec-ommended. Nucleotide sequences of the cloned frag-
ments were determined by Genome Express (Meylan,
France). Nucleotide sequence homology searches
against major sequence databases were done with pro-
grams BLAST 2.0 and FASTA version 2.0X.
2.6. Nucleotide sequence accession numbers
Newly obtained sequences of hrcRST fragments were
deposited in EMBL under the following accession num-
bers: P. syringae pv. syringae 2027-37 (AJ605515), P. sy-
ringae pv. phaseolicola SD68 (AJ605516), P. salomonii
CFBP2022T (AJ605517), P. gingeri CFBP2810T
(AJ605518), P. blatchfordae CFBP3280T (AJ605519), P.
fluorescens bv. II reference strain CFBP5759 (syn. C7)
(AJ271105), P. fluorescens reference strain CFBP5935(syn. F113) (AJ605533), P. fluorescens bv. I CFBP11345
(AJ605520), P. fluorescens bv. I CFBP11347 (AJ605521),
P. fluorescens bv. V CFBP11348 (AJ605522), P. fluores-
cens bv. V CFBP11350 (AJ605523), P. fluorescens bv. I
CFBP11351 (AJ605524),P. fluorescens bv. VCFBP11352
(AJ605525), P. fluorescens bv. I CFBP11355 (AJ605526),
P. fluorescens bv. I CFBP11357 (AJ605527), P. fluores-
cens bv. V CFBP11362 (AJ605528), P. fluorescens bv. ICFBP11363 (AJ605529),P. fluorescensbv. IICFBP11364
(AJ605530), P. fluorescens bv. III CFBP11367
(AJ605531),P. fluorescens bv. VCFBP11386 (AJ605532),
Pseudomonas sp. CFBP11402 (AJ605534).
3. Results
3.1. Distribution of hrp genes homologues in fluorescent
pseudomonads
The distribution of hrcRST sequences in the 103
strains was assessed by PCR with the primers
HRCR8092 and HRCT8986, and by hybridization of
total DNA dot blots with the C7hrcRST probe (Table 1).
PCR allowed the amplification of a single DNAfragment of the expected size (ca. 897 bp) in 40 of the
103 strains tested, these fragments being further showed
to be specific by hybridization with the C7hrcRST
probe. Positive dot-blot hybridization was recorded in
59 of the 103 strains. PCR products and positive hy-
bridization by dot blot were obtained in 75% and 100%
of the phytopathogenic strains, and in 32% and 49% of
the saprophytic strains, respectively. Among the sapro-phytic strains, PCR products and positive hybridization
by dot blot were obtained in 35% and 52% of the rhi-
zosphere strains, and in 22% and 39% of the non-rhi-
zosphere strains, respectively. All strains, for which a
PCR fragment was obtained, gave a positive signal by
dot-blot hybridization.
3.2. Polymorphism of hrcRST PCR fragment and
sequence information
Fourteen of the 40 positive strains gave PCR products
that were too weak to allow RFLP analysis. Analysis
of the PCR fragments was performed with the 26 re-
maining strains that included (i) eight of the 16 phyto-
pathogenic strains (P. blatchfordae CFBP3280T, P.
gingeri CFBP2810T, P. salomonii CFBP2022T, the threeP. marginalis strains CFBP2037T, CFBP2038T,
CFBP2039T, and both P. syringae reference strains 2027-
37 and SD68), and (ii) 18 of the 87 saprophytic strains all
isolated from plant rhizospheres including the reference
strains P. fluorescensC7 (syn. CFBP5759) and F113 (syn.
CFBP5935). Restriction digests were performed with 8
endonucleases on hrcRST PCR fragments. The com-
bined RFLP patterns obtained for the 26 strains studiedgave 21 distinct hrcRST types (Table 1). Each hrcRST
type was represented by a single strain except the types 9,
6 and 16 which included 2, 4 and 2 strains, respectively.
For each hrcRST type, a PCR fragment has been cloned
and sequenced. The strains for which a hrcRST fragment
was sequenced included 16 of the 87 saprophytic strains
and five of the 16 phytopathogenic strains.
The analysis of the Jukes and Cantor [33] pairwisedistance matrix (data not shown) indicated that genetic
distances between the hrcRST sequences ranged be-
tween 0.003 and 0.50, showing a high polymorphism of
hrcRST among the strains. The dendrogram presented
in Fig. 2 indicates the relationships between the se-
quences of the 21 hrcRST fragments and of those ex-
tracted from databases for P. syringae pv. syringae
NSP3121 (Accession No. AF043444), P. fluorescens
SBW25 (Accession No. AF292566), E. amylovara 321
(Accession No. L25828), Yersinia enterolitica A127/90
(Accession No. AY150843), and Rhizobium sp. NGR
234 (Accession No. AE000107).
According to their level of similarity, hrcRST se-
quences were clustered in three groups (Fig. 2). The first
group was made of sequences showing a maximum of
0.25 nucleotide substitution per site, and included threephytopathogenic strains and the biocontrol strain P.
fluorescens C7 (syn. CFBP5759). The second group was
made of sequences of four saprophytic strains isolated
from rhizospheres, including the biocontrol strain P.
fluorescens F113 (syn. CFBP5935), showing a maximum
of 0.12 nucleotide substitution per site. The third group
was made of sequences showing a maximum of 0.29
nucleotide substitution per site, and included 11 sapro-phytic isolates from plant rhizospheres and the biocon-
trol strain P. fluorescens SBW25, plus five
phytopathogenic strains.

Fig. 2. Dendrogram showing the relationships among the hrcRST sequences. Distance estimations according to the coefficient of Jukes and Cantor
[33] were clustered using the UPGMA method of TREECON for windows. hrcRST types are indicated in front of the strain designations. Strains
followed by an accession number under brackets have been included after extraction of the corresponding hrcRST fragment from the sequence cited.
Bootstrap analyses were performed with 1000 resamplings, percent values are shown at the branching points. For each hrcRST type, only the
fragment amplified from the first strain has been sequenced, the other strains belonging to this hrcRST type are indicated after a slash. R, reference
strain; T, type-strain; *, phytopathogenic strain; **, animal pathogenic strain, P. gi. ¼ P. gingeri; P. fl.¼P. fluorescens; P. sy.¼P. syringae; P.
pu.¼P. putida; P. sp¼P. species; P. ma.¼P. marginalis; P. bl.¼P. blatchfordae; P. sa.¼P. salomonii.
462 S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467
3.3. Restriction fragment length polymorphism of 16S
rRNA genes
The combined RFLP patterns of the 16S rRNA genes
obtained with eight restriction endonucleases for the 103
studied strains plus for the sequences of E. coli S17.1, P.
aeruginosa PAO1 and ATCC10145T, and P. fluorescens
SBW25 (from accessions V00348, NC_002516,
AF094713 and NC_002948, respectively) digested in sil-
ico gave 36 distinct ribotypes that are presented in the
dendrogram in Fig. 3. Twelve ribotypes (1, 2, 3, 4, 7, 8, 9,11, 12, 13, 18 and 20) were previously described by La-
guerre et al. [31] and 6 (C8,D3, C9, C5, C1,D5) byLatour
et al. [25]. According to the Jaccard pairwise similarity
matrix, the level of similarity between the different ribo-
types ranged from 0.17 to 0.97 indicating a high poly-
morphism of the 16S rRNA genes. The ribotypes 20, N7,
18 andN2 includingE. coli S17.1, the phytopathogenicP.
flectens CFBP3281T, and the human pathogenic P.
aeruginosa PAO1 and ATCC10145T, respectively, were
the most distant from the others and were then used as
outgroups to root the dendrogram. The ribotype 13 (P.
cichorii CFBP2101T) was also distant from the others.
The level of similarity among the remaining 31 ribo-
types (from 102 strains) was between 0.72 and 0.97, in-
dicating a low polymorphism among the corresponding
strains of fluorescent pseudomonads. These 102 strainswere clustered into three groups (Fig. 3). A first group
(a) showing a level of similarity at least equal to 0.83
included 13 ribotypes representing: (i) three phytopath-
ogenic strains, (ii) 42 saprophytic strains from variousrhizospheres, and (iii) all the soil isolates except P.
putida CFBP2066T. A second group (b) of nine ribo-
types showing a level of similarity at least equal to 0.83
was delineated. This group clustered six phytopatho-
genic strains including, P. blatchfordae CFBP3280T, P.
marginalis CFBP2037T-2038T-2039T, P. salomonii
CFBP2022T, and P. tolaasii CFBP2068T, and 18 sap-
rophytic strains including P. fluorescens CFBP2102T
plus 17 rhizosphere strains. Finally, the nine remaining
ribotypes, encompassing 18 isolates and sharing a sim-
ilarity at least equal to 0.82, were described in a third
group (c); this group included six phytopathogenic
strains, a saprophytic strain isolated from soil (P. putida
CFBP2066T) and 12 saprophytic strains isolated from
rhizospheres.
3.4. Relations between TTSS-like genes and 16S rRNA
gene polymorphism
Strains with TTSS-like genes as detected by PCR and/
or by dot-blot hybridization were distributed in 28 of the
36 ribotypes. Strains harbouring TTSS-like genes were
randomly distributed in the different ribotypes (Fig. 3).
Ten ribotypes included both strains with and withoutthe hrcRST sequence.
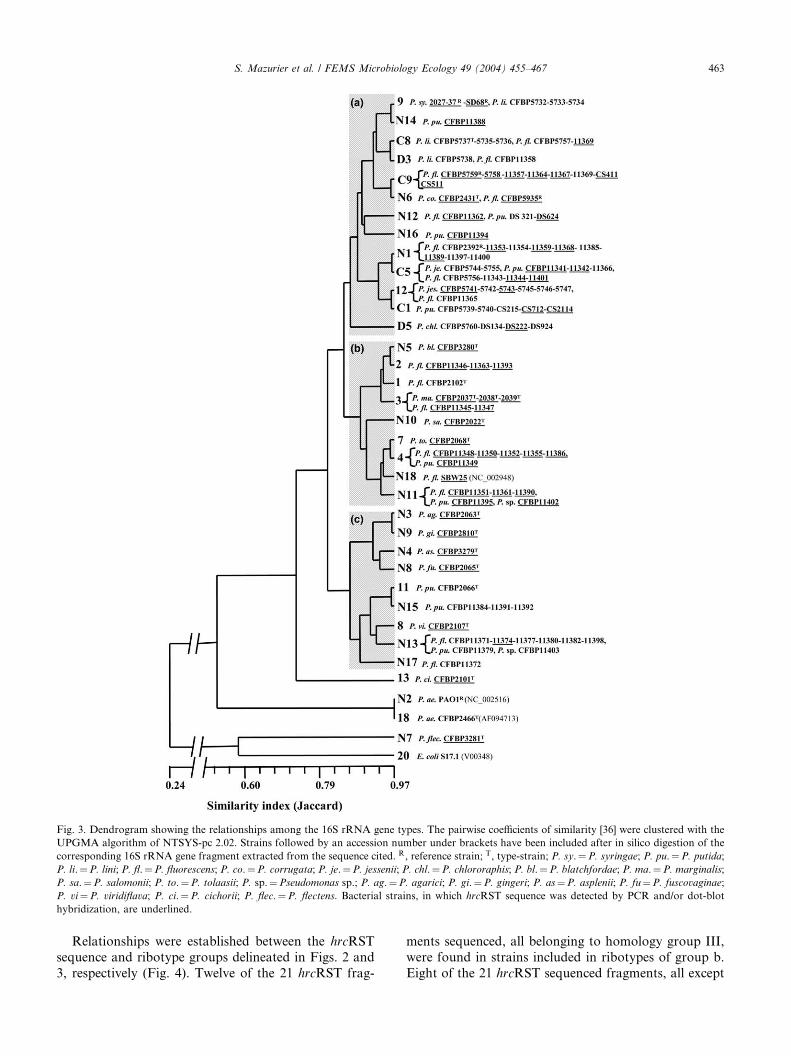
Fig. 3. Dendrogram showing the relationships among the 16S rRNA gene types. The pairwise coefficients of similarity [36] were clustered with the
UPGMA algorithm of NTSYS-pc 2.02. Strains followed by an accession number under brackets have been included after in silico digestion of the
corresponding 16S rRNA gene fragment extracted from the sequence cited. R, reference strain; T, type-strain; P. sy.¼P. syringae; P. pu.¼P. putida;
P. li.¼P. lini; P. fl.¼P. fluorescens; P. co.¼P. corrugata; P. je.¼P. jessenii; P. chl.¼P. chlororaphis; P. bl.¼P. blatchfordae; P. ma.¼P. marginalis;
P. sa.¼P. salomonii; P. to.¼P. tolaasii; P. sp.¼Pseudomonas sp.; P. ag.¼P. agarici; P. gi.¼P. gingeri; P. as¼P. asplenii; P. fu¼P. fuscovaginae;
P. vi¼P. viridiflava; P. ci.¼P. cichorii; P. flec.¼P. flectens. Bacterial strains, in which hrcRST sequence was detected by PCR and/or dot-blot
hybridization, are underlined.
S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467 463
Relationships were established between the hrcRST
sequence and ribotype groups delineated in Figs. 2 and
3, respectively (Fig. 4). Twelve of the 21 hrcRST frag-
ments sequenced, all belonging to homology group III,
were found in strains included in ribotypes of group b.
Eight of the 21 hrcRST sequenced fragments, all except

Fig. 4. Correspondence between the homology groups of hrcRST sequences (1) and of the ribotypes (2) as delineated in Figs. 2 and 3, respectively.
Dashed lines link hrcRST sequences and ribotypes of strains belonging to corresponding groups of homology (hrcRST sequence groups I and II/
ribotype group a, and hrcRST sequence groups III/ribotype group b). Black lines indicate mismatches in the correspondence between the homology
groups of hrcRST sequences and ribotypes referred above.
464 S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467
one belonging to homology groups I and II, were foundin strains included in ribotypes of the group a. Mis-
matches were only recorded for two strains: P. gingeri
CFBP2810T belonged to ribotype group c and to
hrcRST homology group I and P. fluorescens
CFBP11362 belonged to ribotype group a and to
hrcRST homology group III (Fig. 4).
4. Discussion
4.1. Distribution of TTSS-like genes
TTSSs have been extensively studied in P. syringae
[3,13] and hrp genes were described in P. viridiflava [14].
Presence of TTSS-like genes has been recently reported
in few strains that belonged only to two saprophyticspecies (P. fluorescens and P. putida) [15–17]. However
within the fluorescent pseudomonad group, further in-
formation on the distribution of TTSS-like genes in
other phytopathogenic species and in a larger number of
strains belonging to various saprophytic species clearly
remained to be gained. In the present study, we have
then assessed the distribution of these genes in a large
collection of fluorescent pseudomonads (103 strains)belonging to 13 phytopathogenic and five saprophytic
species.
The presence of TTSS-like genes in these strains was
confirmed by the detection of hrcRST sequences chosen
for their high level of conservation since they are a part
of the nine hrc genes coding for the core part of TTSS.
The presence of the hrcRST sequence detected by dot-
blot hybridization of total DNA using C7hrcRST as aprobe was revealed in 100% of the phytopathogenic
strains. These data allowed extending to a wider range
of phytopathogenic species the observation of TTSS-like
genes previously made in P. syringae and P. viridiflava
[13,14]. Furthermore, these data showing the presence of
TTSS-like genes in all tested phytopathogenic strains are
in agreement with the previous demonstration in several
phytopathogenic bacteria that TTSS is implicated in thepathogenesis process [2], and then validate our strategy
based on dot blot using C7hrcRST as a probe for de-
tecting TTSS-like genes in fluorescent pseudomonads.
TTSS-like genes in fluorescent pseudomonads were
found at a lower frequency by PCR amplification than
by dot-blot hybridization. This type of discrepancy be-
tween the data obtained by the two methods has been

S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467 465
commonly reported and may be ascribed to mismatches
between the template and the primers.
Dot-blot hybridization of total DNA allowed the
detection of TTSS-like genes in 49% of the strains dis-
tributed in the five saprophytic species tested. Theseresults indicate that, although the frequency of TTSS
genes is significantly lower in saprophytic than in phy-
topathogenic species, they remain widely distributed
among saprophytic species of fluorescent pseudomo-
nads. These data support and extend those obtained
previously with a small number of strains belonging
only to P. fluorescens and to P. putida [15–17]. This wide
distribution of TTSS-like genes in these strains raisesquestions about their function in saprophytic species,
since TTSSs are known to be dedicated to the translo-
cation of bacterial effector proteins into the cytosol of
eukaryotic cells during the hypersensitive reaction on
non-host plants and disease expression in host plants [3].
The saprophytic strains harbouring TTSS-like genes
included three (P. fluorescens C7, F113 and SBW25) of
the four strains of fluorescent pseudomonads, known asbiocontrol agents, confirming the previous observation
made for P. fluorescens SBW25 [17]. This observation
could raise the hypothesis of the possible involvement of
TTSS in the elicitation of the defence reactions known
to play a major role in the disease suppression by fluo-
rescent pseudomonads [37]. More generally, one may
suggest that TTSS might be involved in the crosstalk
between bacteria and plant as showed for plant-bacterialmolecular dialogue mediating the establishment of
symbiotic associations [9,10]. This is supported by rather
high frequency of the TTSS-like genes in rhizosphere
strains (52%). However, these genes were also found in
non-rhizosphere strains (39%), although at a lower fre-
quency, and their presence was not restricted to specific
bacterial genotypes since they were distributed ran-
domly in most of the ribotypes.Further studies should be performed to determine if
the soil strains harbouring TTSS-like genes, although
they were not isolated from plants, show a higher ability
to elicit plant defence reactions and are more rhizo-
sphere competent than the soil strains without TTSS-
like genes.
4.2. Diversity of TTSS-like genes
The PCR method developed in the present study
provided a series of hrcRST fragments, which allowed
us to characterize their diversity in eight and 18 strains
belonging to various phytopathogenic and saprophytic
species. Comparable studies have so far been performed
for P. syringae and not for saprophytic Pseudomonas
species.PCR-RFLP analysis of the hrcRST sequence allowed
the identification of 21 hrcRST types. One fragment per
type was sequenced, and their diversity was analysed
together with five sequences extracted from a data bank.
The hrcRST sequences have shown a high level of di-
versity. This level was possibly even underestimated
since only 44% of the sequences detected by dot-blot
hybridization gave a PCR product that was strong en-ough for RFLP analysis. Furthermore, only 36% of the
detected hrcRST genes were sequenced. A notably high
diversity was recorded within strains belonging to P.
fluorescens, which contrasts the hypothesis of Preston
et al. [17]. Conversely, a low diversity was recorded
within the two phytopathogenic species P. syringae and
P. marginalis. The three strains of P. marginalis be-
longed to the same hrcRST type despite their distribu-tion in three different pathovars, and the three strains of
P. syringae (pathovars syringae and phaseolicola) har-
boured TTSS-like genes that were closely related. The
low diversity of TTSS-like genes in P. syringae recorded
in the present study is in agreement with the study of
Sawada et al. [38].
The hrcRST sequences were clustered in three groups
according to their level of similarity. The strains be-longing to saprophytic species were distributed
throughout the groups, and the strains belonging to
phytopathogenic species were distributed in the groups I
and III, that included most of the saprophytic strains.
These data indicate that hrcRST sequences in strains
belonging to different phytopathogenic species are dis-
tant from each other, and are even more distant from
each other than from some of the strains belonging tosaprophytic species.
Alfano et al. [39] have shown that in P. syringae the
hrp genes encoding the TTSS machinery are the con-
served center region (CCR) of a tripartite pathogenicity
island that includes exchangeable (EEL) and conserved
(CEL) effector loci. According to these authors, Hrp-
mediated pathogenicity in P. syringae seems to be
dependent on a set of genes that are universal amongdivergent pathovars and on another set that varies
among strains in the same pathovar. This observation
suggests that strains, presenting various pathogenic
properties, would carry a universal TTSS. This sugges-
tion was recently supported by further studies based on
a mutational approach [40,41]. In the present study,
TTSS-like genes belonging to the same homology group
were described in distinct bacterial species and in bac-teria having different effects on plants; either negative
(pathogenic), neutral (saprophytic) or positive (biocon-
trol). These observations suggest that the proposal by
Alfano et al. [39] for P. syringae could possibly be ex-
tended to saprophytic and beneficial fluorescent pseu-
domonads. This hypothesis could be assessed by
comparing the genomic regions of hrc genes of these
types of fluorescent pseudomonads together with thoseknown in P. syringae.
The polymorphism of hrcRST genes within the fluo-
rescent pseudomonads was related to that of the 16S

466 S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467
rRNA genes. 16S rRNA genes are commonly used for
diversity analyses of bacterial populations [25,31,42–44].
A correspondence was established between hrcRST se-
quences and the ribotypes for all the strains, except for
two, when comparing the topology of the correspondingdendrograms. This observation suggests that TTSS-like
and 16S rRNA genes have followed a similar evolution.
According to our data this would apply both to the
saprophytic and phytopathogenic strains. This sugges-
tion is in agreement with the report of Sawada et al. [38]
on the evolutionary stability of hrp gene cluster in P.
syringae when analysing the phylogeny of hrpS and hrpL
of strains belonging to different pathovars. However,two mismatches were observed in the relationship be-
tween 16S rDNA types and hrcRST sequences. These
mismatches involved a phytopathogenic (P. gingeri
CFBP2810T) and a saprophytic strain (P. fluorescens
CFBP11362), and suggest that some TTSS-like genes
might have experienced horizontal gene transfer. This
hypothesis would be consistent with the implication of
horizontal transfer in the evolution of TTSS-like geneclusters, which was previously shown for phytopatho-
genic pseudomonads [1,39,45,46]. Our results also sug-
gest that TTSS gene transfer might not only occur in
phytopathogenic strains but also in saprophytic strains
of fluorescent pseudomonads. Further work on phy-
logeny would be required to further assess the evolution
history of TTSS-like genes within phytopathogenic and
saprophytic strains.
5. Conclusions
TTSS-like genes were distributed in all tested phy-
topathogenic strains belonging to 13 species of fluo-
rescent pseudomonads and were also widely distributed
in strains belonging to saprophytic species. In fact,phytopathogenic and saprophytic strains could be dis-
criminated neither on the basis of the presence of
TTSS-like genes nor on the polymorphism of the cor-
responding sequences. Within saprophytic strains,
TTSS-like genes were more frequently detected in
strains associated with plants but were also common in
strains isolated from soil. Collectively, these data
question the role of these genes in saprophytic strainsincluding strains not associated with eukaryotes.
Comparison of the polymorphisms of hrcRST and of
16S rRNA genes suggests that they have followed a
similar evolution even if horizontal transfer might have
occurred. Further studies are required to support these
hypotheses and to evaluate the possible implication of
the TTSS in plant–microbe interactions. These studies
would be based on phylogeny analysis and on thecomparison of plant–microbe interactions when using
strains harbouring these sequences and the corre-
sponding targeted mutants.
Acknowledgements
The authors are grateful to K. Klein for input to the
English and to S. Delorme, V. Edel, and J. Raaijmakers
for helpful discussions.
References
[1] Galan, J.E. and Collmer, A. (1999) Type III secretion machines:
bacterial devices for protein delivery into host cells. Science 284,
1322–1328.
[2] Hueck, C.J. (1998) Type III protein secretion systems in bacterial
pathogens of animal and plants. Microbiol. Mol. Biol. Rev. 62,
379–433.
[3] Jin, Q., Thilmony, R., Zwiesler-Vollick, J. and He, S.-Y. (2003)
Type III protein secretion in Pseudomonas syringae. Microb.
Infect. 5, 301–310.
[4] Alfano, J.R. and Collmer, A. (1997) Type III (Hrp) secretion
pathway of plant pathogenic bacteria: trafficking harpins, Avr
proteins, and death. J. Bacteriol. 179, 5655–5662.
[5] Bogdanove, A.J., Beer, S.V., Bonas, U., Boucher, C.A., Collmer,
A., Coplin, D.L., Cornelis, G.R., Huang, H.-C., Hutcheson, S.W.,
Panopoulos, N.J. and Van Gijsegem, F. (1996) Unified nomen-
clature for broadly conserved hrp genes of phytopathogenic
bacteria. Mol. Microbiol. 20, 681–683.
[6] He, S.Y. (1998) Type III protein secretion systems in plant and
animal pathogenic bacteria. Annu. Rev. Phytopathol. 36, 363–
392.
[7] Russel, M. (1994) Phage assembly: a paradigm for bacterial
virulence factor export? Science 265, 612–614.
[8] Collmer, A., Badel, J.L., Charkowski, A.O., Deng, W.-L., Fouts,
D.E., Ramos, A.R., Rehm, A.H., Anderson, D.M., Schneewind,
O., van Dijk, K. and Alfano, J.R. (2000) Pseudomonas syringae
Hrp type III secretion system and effector proteins. Proc. Natl.
Acad. Sci. USA 97, 8770–8777.
[9] Marie, C., Broughton, W.J. and Deakin, W.J. (2001) Rhizobium
type III secretion systems: legume charmers or alarmers. Curr.
Opin. Plant Biol. Infect. 4, 336–342.
[10] Krishnan, H.B. (2002) NolX of Sinorhizobium fredii USDA257, a
type III-secreted protein involved in host range determination, is
localized in the infection threads of cowpea (Vigna unguiculata [L.]
Walp) and soybean (Glycine max [L.] Merr.) nodules. J. Bacteriol.
184, 831–839.
[11] Walsh, U.F., Morrissey, J.P. and O’Gara, F. (2001) Pseudomonas
for biocontrol of phytopathogens: from functional genomics to
commercial exploitation. Curr. Opin. Biotechnol. 12, 289–295.
[12] Cao, H., Baldini, R.L. and Rahme, L.G. (2001) Common
mechanism for pathogens of plants and animals. Annu. Rev.
Phytopathol. 39, 259–284.
[13] Charkowski, A.O., Alfano, J.R., Preston, G., Yuang, G., He, S.Y.
and Collmer, A. (1998) The Pseudomonas syringae pv. tomato
HrpW protein has domains similar to harpins and pectate lyases
and can elicit the plant hypersensitive response and bind to
pectate. J. Bacteriol. 180, 5211–5217.
[14] Jakob, K., Goss, E.M., Araki, H., Van, T., Kreitman, M. and
Bergelson, J. (2002) Pseudomonas viridiflava and P. syringae-
natural pathogens of Arabidopsis thaliana. Mol. Plant Microbe
Interact. 15, 1195–1203.
[15] Mulya, K., Takikawa, Y. and Tsuyumu, S. (1996) The presence of
regions homologous to hrp cluster in Pseudomonas fluorescens
PfG32R. Ann. Phytopathol. Soc. Jpn. 62, 355–359.
[16] Mazurier, S., Delorme, S., Siblot, S., Lemanceau, P. (2000)
Presence of DNA sequences characteristic of type III secretion
systems in biocontrol Pseudomonas fluorescens strain C7, p. 77. In:

S. Mazurier et al. / FEMS Microbiology Ecology 49 (2004) 455–467 467
Proceedings of the 5th International PGPR Workshop, Cordoba,
Argentina, October 3–November 3, 2000.
[17] Preston, G., Bertrand, N. and Rainey, P.B. (2001) Type III
secretion in plant growth-promoting Pseudomonas fluorescens
SBW25. Mol. Microbiol. 41, 999–1014.
[18] Lemanceau, P., Samson, R. (1983) Relations entre quelques
caract�eristiques in vitro de 10 Pseudomonas fluorescents et leur
effet sur la croissance du haricot (Phaseolus vulgaris), p. 327. In
‘‘Les antagonismes microbiens’’, 24�eme colloque de la SFP,
Bordeaux, 360 pp.
[19] Gamalero, E., Martinotti, M.G., Trotta, A., Lemanceau, P. and
Berta, G. (2002) Morphogenetic modifications induced by Pseu-
domonas fluorescens A6RI and Glomus mosseae BEG12 in the root
system of tomato differ according to the plant growth conditions.
New Phytol. 155, 293–300.
[20] Lemanceau, P. and Alabouvette, C. (1991) Biological control of
fusarium diseases by fluorescent Pseudomonas and non-pathogenic
Fusarium. Crop Protect. 10, 279–286.
[21] Fenton, A.M., Stephens, P.M., Crowley, J., O’Callaghan, M. and
O’Gara, F. (1992) Exploitation of gene(s) involved in 2,4-
diacetylphloroglucinol biosynthesis to confer a new biocontrol
capability to a Pseudomonas strain. Appl. Environ. Microbiol. 58,
3873–3878.
[22] Cronin, D., Mo€enne-Loccoz, Y., Fenton, A., Dunne, C., Dowling,
D.N. and O’Gara, F. (1997) Role of 2,4-diacetylphloroglucinol in
the interactions of the biocontrol pseudomonad F113 with the
potato cyst nematode Globodera rostochiensis. Appl. Environ.
Microbiol. 63, 1357–1361.
[23] Cronin, D., Mo€enne-Loccoz, Y., Fenton, A., Dowling, D.N. and
O’Gara, F. (1997) Ecological interaction of a biocontrol Pseudo-
monas fluorescens strain producing 2,4-diacetylphloroglucinol with
the soft rot potato pathogen Erwinia carotovora subsp. atroseptica.
FEMS Microbiol. Ecol. 23, 95–106.
[24] Lemanceau, P., Corberand, T., Gardan, L., Latour, X., Laguerre,
G., Boeufgras, J.M. and Alabouvette, C. (1995) Effect of two plant
species flax (Linum usitatissinum L.) and tomato (Lycopersicon
esculentum Mill.) on the diversity of soilborne populations of
fluorescent pseudomonads. Appl. Environ. Microbiol. 61, 1004–
1012.
[25] Latour, X., Corberand, T., Laguerre, G., Allard, F. and Leman-
ceau, P. (1996) The composition of fluorescent pseudomonad
population associated with roots is influenced by plant and soil
type. Appl. Environ. Microbiol. 62, 2449–2556.
[26] Bossis, E. (1995) Les Pseudomonas fluorescents de la rhizosph�ere:�etude taxonomique et effets sur la croissance la tomate et du ma€ıs,
de la germination �a la lev�ee. Th�ese de doctorat, 143 pp. Universit�e
de Nantes, France.
[27] Miller, J.H. (1972) Experiments in Molecular Genetics, pp. 132–
135 Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
[28] Brenner, D.J., McWhorter, A.C., Knuston, J.K. and Steigerwalt,
A.G. (1982) Escherichia vulneris: a new species of Enterobacteri-
aceae associated with human wounds. J. Clin. Microbiol. 15,
1133–1140.
[29] Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUS-
TALW: improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, positions-specific gap
penalties and weight matrix choice. Nucleic Acids Res. 22,
4673–4680.
[30] Weisburg, W.G., Barns, S.M., Pelletier, D.A. and Lane, D.J.
(1991) 16S ribosomal DNA amplification for phylogenetic study.
J. Bacteriol. 137, 697–703.
[31] Laguerre, G., Rigottier-Gois, L. and Lemanceau, P. (1994)
Fluorescent Pseudomonas species categorized by using Polymerase
Chain Reaction (PCR)/restriction fragment analysis of 16S
rDNA. Mol. Ecol. 3, 479–487.
[32] Van de Peer, Y. and De Wachter, R. (1994) TREECON for
Windows: a software package for the construction and drawing of
evolutionary trees for the Microsoft Windows environment.
Comput. Appl. Biosci. 10, 569–570.
[33] Jukes, T.H. and Cantor, C.R. (1969) Evolution of protein
molecules. In: Mammalian Protein Metabolism (Munro, H.H.,
Ed.), pp. 21–132. Academic Press, New York.
[34] Sneath, P.H.A. and Sokal, R.R. (1973) Numerical Taxonomy, the
Principles and Practice of Numerical Classification. Freeman, San
Francisco.
[35] Rohlf, F.J. (1998) NTSYS: Numerical Taxonomy and Multivar-
iate Analysis System, second ed. Exeter Software, State University
of New York, Stany Brook, NY.
[36] Jaccard, P. (1908) Nouvelles recherches sur la distribution florale.
Bull. Soc. Vaud. Sci. Nat. 44, 223–270.
[37] Van Loon, L.C., Bakker, P.A.H.M. and Pieterse, C.M.J. (1998)
Systemic resistance induced by rhizosphere bacteria. Ann. Rev.
Phytopathol. 36, 453–483.
[38] Sawada, H., Suzuki, F., Matsuda, I. and Saitou, N. (1999)
Phylogenetic analysis of Pseudomonas syringae pathovars
suggests the horizontal gene transfer of argK and the
evolutionary stability of hrp gene cluster. J. Mol. Evol. 49,
627–644.
[39] Alfano, J.R., Charkowski, A.O., Deng, W.-L., Badel, J.L.,
Petnicki-Ocwieja, T., van Dijk, K. and Collmer, A. (2000)
The Pseudomonas syringae Hrp pathogenicity island has a
tripartite mosaic structure composed of a cluster of type III
secretion genes bounded by exchangeable effector and con-
served effector loci that contribute to parasitic fitness and
pathogenicity in plants. Proc. Natl. Acad. Sci. USA 98, 4856–
4861.
[40] Deng, W.-L., Rehm, A.H., Charkowski, A.O., Clemencia, M.R.
and Collmer, A. (2003) Pseudomonas syringae exchangeable
effector loci: sequence diversity in representative pathovars and
virulence function in P. syringae pv. syringae B728a. J. Bacteriol.
185, 2592–2602.
[41] Fouts, D.E., Badel, J.L., Ramos, A.R., Rapp, R.A. and
Collmer, A. (2003) A Pseudomonas syringae pv. tomato
DC3000 Hrp (type III secretion) deletion mutant expressing
the Hrp system of bean pathogen P. syringae pv. syringae 61
retains normal host specificity for tomato. Mol. Plant Microbe
Interact. 16, 43–52.
[42] Oger, P., Dessaux, Y., Petit, A., Gardan, L., Manceau, C.,
Chomel, C. and Nesme, X. (1998) Validity, sensitivity and
resolution limit of the PCR-RFLP analysis of the rrs (16S rRNA
gene) as a tool to identify soil-borne and plant-associated bacterial
populations. Genet. Sel. Evol. S1, S311–321.
[43] Delorme, S., Philippot, L., Edel-Hermann, V., Deulvot, C.,
Mougel, C. and Lemanceau, P. (2003) Comparative genetic
diversity of the narG, nosZ, and 16S rRNA genes in
fluorescent pseudomonads. Appl. Environ. Microbiol. 69,
1004–1012.
[44] Keel, C., Weller, D.M., Natsch, A., D�efago, G., Cook, R.J. and
Thomashow, L.S. (1996) Conservation of the 2,4-diacetylphloro-
glucinol biosynthesis locus among fluorescent Pseudomonas strains
from diverse geographic locations. Appl. Environ. Microbiol. 62,
552–563.
[45] Charity, J.C., Pak, K., Delwiche, C.F. and Hutcheson, S.W.
(2003) Novel exchangeable effector loci associated with the
Pseudomonas syringae hrp pathogenicity island: evidence for
integron-like assembly from transposed gene cassettes. Mol. Plant
Microbe Interact. 16, 495–507.
[46] Gophna, U., Ron, E.Z. and Graur, D. (2003) Bacterial type III
secretion systems are ancient and evolved by multiple horizontal-
transfer events. Gene 312, 151–163.
Related Documents











