International Journal of Salt Lake Research 8: 23-40, 1999. 1999 KluwerAcademic Publishers. Printed in the Netherlands. Distribution and characterization of Chilean populations of the brine shrimp Artemia (Crustacea, Branchiopoda, Anostraca) OSCAR ZUlqIGA t, RODOLFO WILSON 1 , FRANCISCO AMAT 2 and FRANCISCO HONTORIA 2 1Departamento de Acuicultura, Facultad de Recursos del Mar, Universidad de Antofagasta, Antofagasta, Chile; 21nstituto de Acuicultura de Torre de la Sal ( CS1C), 12595 Ribera de Cabanes, Castell6n, Espaha Abstract. Hypersaline habitats in Chile, from marine solar salt pans to saline lagoons and pools in Andean salars, were prospected in search of Artemia populations. These saline ecosystemswere characterizedthrough physico-chemical parameters and ionic compositionof their brines. Biometrics of cysts and nauplii, as well as morphometrics by using multivariate discriminant analysis for adult specimens evidenced that the Chilean populations of brine shrimp belong to A. franciscana. Cross-breeding results supported the former hypothesis of conspecific Chilean populations, and their differences with A. persimilis, also endemic to the New World, but restricted to Argentinean sites. Key words: Artemia, Chilean populations, hypersaline ecosystems, cysts, nauplii, adult morphology, crossbreeding Introduction South America is a continent endowed with a great diversity of hypersaline ecosystems, i.e. solar saltworks, salt lakes and lagoons, saline deposits and evaporitic basins named salars ("salares"). In South America these biotopes have been scarcely studied until recently (Hammer, 1986), except for very specific topics related to their origin and chemical characteristics (Chong, 1988; Drago and Quir6s, 1996), and biological characteristics like palaeo- biology (Servant and Mello, 1993), limnology (Dejoux, 1993; Williams et al., 1995), water birds distribution and the populations of invertebrates on which they feed (Hurlbert et al., 1984), and this information is even more sparse for some relatively inaccessible hypersaline biotopes, specially for the lakes and lagoons located in or near the Altiplano (Williams et al., 1995). Previously updated lists of Artemia sites (Vanhaecke et al., 1987; Trianta- phyllidis et al., 1998) pay little attention to South America; the information

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

International Journal of Salt Lake Research 8: 23-40, 1999. �9 1999 KluwerAcademic Publishers. Printed in the Netherlands.
Distribution and characterization of Chilean populations of the brine shrimp Artemia (Crustacea, Branchiopoda, Anostraca)
OSCAR ZUlqIGA t, RODOLFO WILSON 1 , FRANCISCO AMAT 2 and FRANCISCO HONTORIA 2 1Departamento de Acuicultura, Facultad de Recursos del Mar, Universidad de Antofagasta, Antofagasta, Chile; 21nstituto de Acuicultura de Torre de la Sal ( CS1C), 12595 Ribera de Cabanes, Castell6n, Espaha
Abstract. Hypersaline habitats in Chile, from marine solar salt pans to saline lagoons and pools in Andean salars, were prospected in search of Artemia populations. These saline ecosystems were characterized through physico-chemical parameters and ionic composition of their brines. Biometrics of cysts and nauplii, as well as morphometrics by using multivariate discriminant analysis for adult specimens evidenced that the Chilean populations of brine shrimp belong to A. franciscana. Cross-breeding results supported the former hypothesis of conspecific Chilean populations, and their differences with A. persimilis, also endemic to the New World, but restricted to Argentinean sites.
Key words: Artemia, Chilean populations, hypersaline ecosystems, cysts, nauplii, adult morphology, crossbreeding
In troduc t ion
South America is a continent endowed with a great diversity of hypersaline ecosystems, i.e. solar saltworks, salt lakes and lagoons, saline deposits and evaporitic basins named salars ("salares"). In South America these biotopes have been scarcely studied until recently (Hammer, 1986), except for very specific topics related to their origin and chemical characteristics (Chong, 1988; Drago and Quir6s, 1996), and biological characteristics like palaeo- biology (Servant and Mello, 1993), limnology (Dejoux, 1993; Williams et al., 1995), water birds distribution and the populations of invertebrates on which they feed (Hurlbert et al., 1984), and this information is even more sparse for some relatively inaccessible hypersaline biotopes, specially for the lakes and lagoons located in or near the Altiplano (Williams et al., 1995).
Previously updated lists o f A r t e m i a sites (Vanhaecke et al., 1987; Trianta- phyllidis et al., 1998) pay little attention to South America; the information

24
referring to the South Cone, especially Chile and Argentina, is scarce, and only a few recent papers or symposia communications refer to well documented data about the geographical distribution and taxonomy for A. franciscana Kellogg and A. persimilis Piccinelli and Prosdocimi, autochtho- nous in the southern continent (Gajardo et al., 1992, 1995; Wilson et al., 1993; Gajardo and Beardmore, 1993; Z6fiiga et al., 1994; Amat et al., 1994).
The great diversity of climatic, hydrobiological and geological conditions present in Chile, where the Central Andes, together with the Pre-Andean Range, the Coastal Range and one of the most arid deserts in the world promote the development of a great number of hypersaline biotopes, prompted this attempt to update the geographical distribution of Artemia in Chile.
In this paper brine shrimp populations found were characterized through the morphology of their females, the mode of reproduction, the biometrics of cysts and instar-I nauplii, and by interpopulational mating experiments, aiming to join these results with previous information (Gajardo and Beard- more, 1993) on phenotypical and geneticai characterization of Artemia strains from South America.
Materials and methods
Between 1994 and early 1997 Chile was prospected in search of hypersaline biotopes (Table 1) from North to South, and from the Pacific coast to the Central depression and the Pre-Andean saline plains or salars, all along the 4,000 km between the Salar de Surire (Regirn I) to the North-East, bordering on Bolivia (Amat et al., 1994).
In October 1995 the prospection focused on the Atacama region (Regi6n III), in the Pre-Andean and Andean ranges of the Chafiaral and Copiap6 provinces, bordering on Argentina (Figure 1). The saline ecosystems visited were: Laguna de Santa Rosa in the Salar de Maricunga (3,700 m above sea level), Laguna Verde (3,700 m.a.s.1.), Laguna del Negro Francisco (3,800 m.a.s.1.) and Salar de Pedernales (3,700 m.a.s.1.). In March 1997, Laguna Amarga in Torres del Paine National Park (Regirn XII) was prospected following information about a previous report on the existence of Artemia populations there (Campos et al., 1996).
This information on Artemia sites can hardly be considered as exhaustive, because in Chile there are about 100 recorded salt lakes and lagoons, most of them in remote places with limited accessibility (Chong, 1984, 1988).
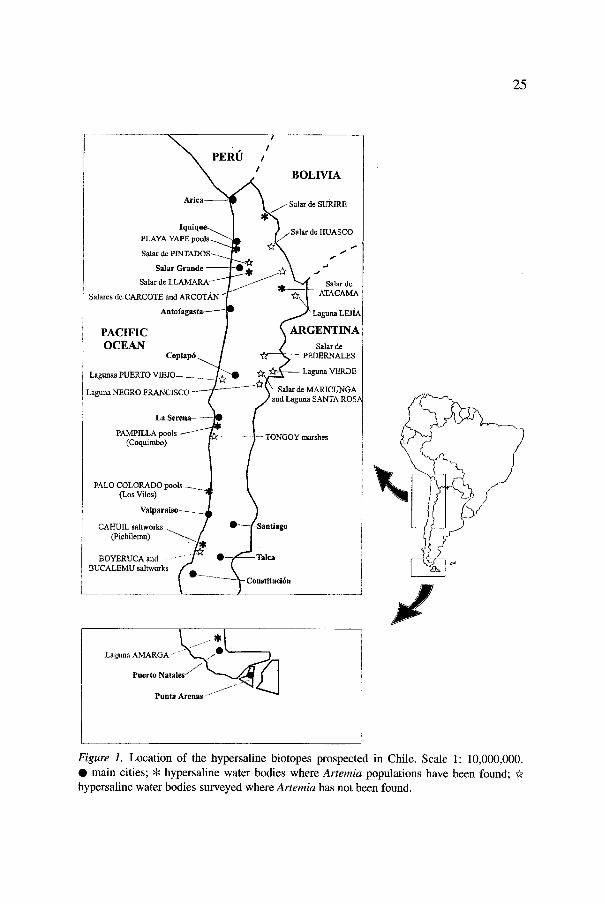
25
PERIJ
PLAYA YAPE pools
Salares de CARCOTE and ARCOT.~N
PACIFIC OCEAN
Lagunas PUERTO VIEIO-.__.
PAMPILLA po~ (Coquimbo)
PALO COLORADO (Los Vilos)
CAHUIL saltworks (Pichilemu)
BOYERUCA and ...... "~[ BUCALEMU saltworks / -
/ /
/ /
BOLIVIA
~ ~ S a | a r de SURIRE
Salar de IIUASCO
* . " I / ~____.'~__ Salarde / -'- r ATACAMA / ~ , j ~ Lagur, a LEJ[A ~) ARGENTINA
~i/~ Sa ar de . ~ PEDERNALES
Laguna VERDE ~5~/'~ Salar de MAKICUNGA ~ and Laguna SANTA ROS/ ~ ~
- TONGOY marshes
S a n t i a g o
- - T a l c a
- C o n s t i t u c i 6 n I
" J - * /
Figure 1. Location of the hypersaline biotopes prospected in Chile. Scale 1: 10,000,000. �9 main cities; * hypersaline water bodies where Artemia populations have been found; ~r hypersaline water bodies surveyed where Artemia has not been found.
26
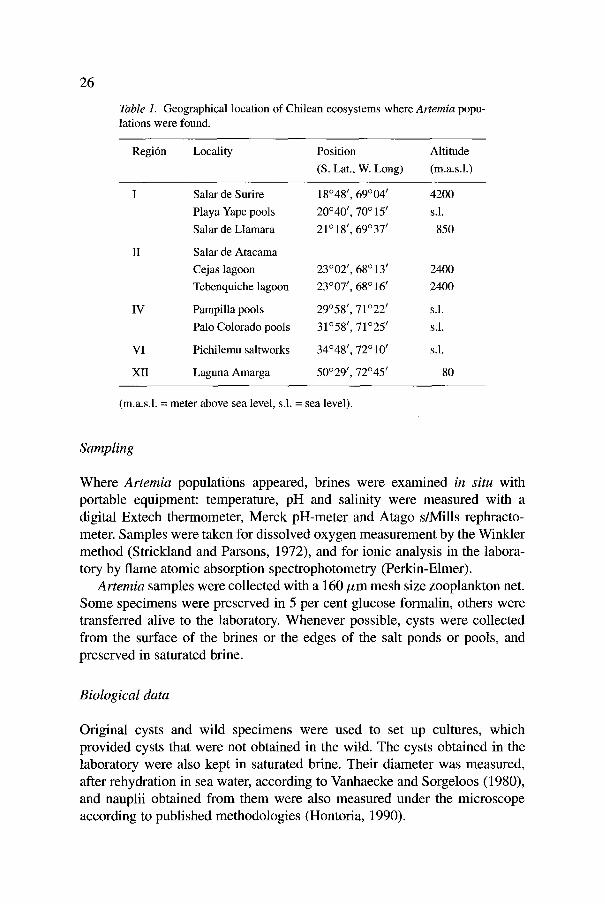
Table 1. Geographical location of Chilean ecosystems where Artemia popu- lations were found.
Regi6n Locality Position Altitude (S. Lat., W. Long) (m.a.s.1.)
I Salar de Surire 18~ 69 ~ 04 t 4200 Playa Yape pools 20~ I, 70 ~ 151 s.1.
Salar de Llamara 21 ~ 18/, 69~ t 850
II Salar de Atacama Cejas lagoon 23002 I, 68 ~ 13 f 2400 Tebenquiche lagoon 23~ t, 68 ~ 161 2400
IV Pampilla pools 29 ~ 58 t, 71 ~ 221 s.1.
Palo Colorado pools 31~ ~, 71~ ~ s.1.
VI Pichilemu saltworks 34048 I, 72 ~ 101 s.1.
XII Laguna Amarga 50~ ~, 72~ t 80
(m.a.s.1. = meter above sea level, s.1. = sea level).
Sampling
Where Artemia populations appeared, brines were examined in situ with portable equipment: temperature, pH and salinity were measured with a digital Extech thermometer, Merck pH-meter and Atago s/Mills rephracto- meter. Samples were taken for dissolved oxygen measurement by the Winkler method (Strickland and Parsons, 1972), and for ionic analysis in the labora- tory by flame atomic absorption spectrophotometry (Perkin-Elmer).
Artemia samples were collected with a 160/ t in mesh size zooplankton net. Some specimens were preserved in 5 per cent glucose formalin, others were transferred alive to the laboratory. Whenever possible, cysts were collected from the surface of the brines or the edges of the salt ponds or pools, and preserved in saturated brine.
Biological data
Original cysts and wild specimens were used to set up cultures, which provided cysts that were not obtained in the wild. The cysts obtained in the laboratory were also kept in saturated brine. Their diameter was measured, after rehydration in sea water, according to Vanhaecke and Sorgeloos (1980), and nauplii obtained from them were also measured under the microscope according to published methodologies (Hontoria, 1990).

27
The adult morphometry was performed according to methodologies previ- ously described (Hontoria and Amat, 1992 a,b) and was used to differentiate species and strains in the genus Artemia (Prrez et al., 1994; Pilla and Beardmore, 1994).
Nauplii from the Chilean populations at Pichilemu or Cahuil saltworks, Playa Yape and Palo Colorado pools, Salar de Llamara, Cejas lagoon at Salar de Atacama and Laguna Amarga, as well as from the saltworks at Piura and Ancash (Perti), San Francisco, California (USA), Laguna Grande de Hidalgo, La Pampa (Argentina), were used to raise laboratory populations up to adulthood. The same methodology was applied with viable nauplii obtained after cross-breeding experiments with some of the former popula- tions. These nauplii were cultured in 70 g L -1 brine at 24 ~ under a 12L: 12D photoperiod. A mixture of Dunaliella salina Teodoresco and Tetraselmis suecica Butcher (1:1) was used as food. The medium was renewed twice a week.
Once 50 per cent of the females attained adulthood and full ovisac devel- opment, a random sample of thirty ovigerous females was removed. They were anesthetized and in each specimen the morphological characters were measured under the dissection microscope (Hontoria and Amat, 1992a,b; Amat et al., 1995a,b). These morphological data were used to discriminate populations (Anderson, 1984), and the origin of the populations was used as the separation criterion. The calculations were performed using Statistical Package for Social Sciences.
Cross-breeding
As it was too complicated to mate all populations studied, and in order to avoid the production of encysted offspring, most interpopulational crosses were performed with the population from Cejas lagoon (Salar de Atacama) because of the exclusive ovoviviparous mode of reproduction of its females.
Matings were performed between Laguna Grande de Hidalgo (Argentina) vs. Cejas; Palo Colorado vs. Cejas, and Laguna Grande de Hidalgo vs. Palo Colorado, while only males from Pichilemu and Piura were mated to Cejas females. Females and males were isolated when sexually immature to ensure that females had not been inseminated before the start of mating. Crosses were attempted keeping isolated single pairs or matings in small beakers (60 ml) with 70 g L -1 brine at 24 ~ and fed the microalgae mixture as previously described. The culture medium was changed every three days, the offspring was counted and removed, and dead mates were replaced by isolated new ones.
Live nauplii obtained from Cahuil vs. Cejas and Piura vs. Cejas matings were cultured until adulthood under the conditions previously described in

28
order to obtain F1 populations for adult morphometrical studies. In case of encysted offspring production, these cysts were kept in 250 g L -1 brine prior to washing with distilled water and oven drying at 40 ~
In order to verify F1 and F2 viability, as well as the dominant type of offspring (oviparity and/or ovoviviparity), several males and females of the autochthonous Chilean populations were allowed to cross-breed in common vessels (500 ml): Palo Colorado vs. Pichilemu, and Pichilemu vs. Cejas.
Results
The ecosystems in which active Artemia populations were found are listed in Table 1, where more information on their geographical situation is given. No Artemia populations were found in the biotopes prospected in the provinces of Chafiaral and Copiap6. The presence of brine shrimp here can not be excluded absolutely, specially in the free waters of Maricunga and Pedernales, but the low temperatures and salinities recorded during this prospection (early spring in the southern hemisphere) were not suitable for this Anostracan.
There are important differences in the origin and characteristics of the biotopes presented in Table 1. The rock pools along the shores of Playa Yape, Pampilla and Palo Colorado, and the saltworks at Pichilemu are in connection with the sea. Rock pools are supralitoral, located at the highest level attainable by high tide, but are sporadically filled with rain water. The saltworks are exploited traditionally by evaporation of seawater in salt pans located in the estuary outlet to the Pacific Ocean, like Nilahue River for the Cahuil salterns.
In the case of the salars (Figure 2), artemia populations were only found where peripheral salt water ponds or little lagoons exist, receiving water from melting Andean ice or sub-surface brines. These ponds are seasonal or permanent.
The salars with Artemia populations are different in geographical loca- tion and hydrological regime. While the Salar de Llamara is situated in a saline plain ("pampa salitrosa"), with ponds receiving limited amounts of underground water, the Salar de Atacama and Salar de Surire are located in evaporitic basins with permanent inlet of underground waters coming from the Andes, produced by the exceptional meteorological phenomenon called "Invierno Boliviano", with plenty of rainfall and snow between December and March in the Altiplano zone. These lagoons in the Pre-Andean salars, like Cejas, Tebenquiche and Barros Negros, show discrete brine levels year- round, are usually deep and presumably intercommunicate under the surface of saline detritic crusts (Chong, 1988).
Finally, Laguna Amarga ("bitter") can be considered an exception to a previous statement (Campos, 1984) on the absence of saline lakes in Chile

29
Altiplano salars
Salar de West Atacama Volcanoes ~
Pampa salitrosa Range \., / ~ - ~ ~'J, JC~ / Shore pOOlS (saline plain) " ~ . " \ 2 f \ ~t "4000 m aria sa tworKs , -~ \ �9 \ \ "~' ", =: -3000
Salar de LLamara \ Ravines
�9 ~ "tO0O Pa c
-o
I . q - -COASTAL~ ~.l I',1-~ CENTRAL DEPRESSION . . . . . . . . . . ~ 4F. PREANDF_.AN-t. 14 ALTIPLANO - - P
R A N G E . . . . . . . . . . . A N D ~ N R A N G E . . . . . . . .
Figure 2. Cross section through the north of Chile showing morphostructural distribution of diverse hypersaline biotopes referred to in Table 1 (Modified from Chong, 1988, and TURISTEL, 1994).
south to latitude 28 ~ because this lagoon belongs to a series of saline lakes present in a semi-arid area within an endorrheic basin, with a common deficit between precipitation and evaporation, and strong winds during spring and summer, which cause its saline condition and explain the name of Laguna Amarga (Campos et al., 1994; Soto et al., 1994).
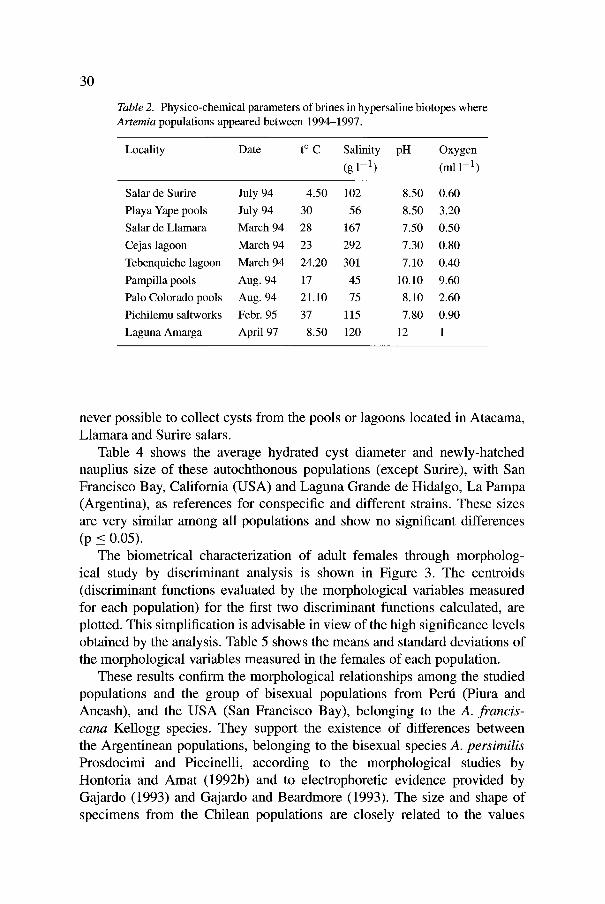
The most important physico-chemical features of the biotopes where Artemia populations were sampled in 1994-97 are summarized in Table 2. They show the usual differences between moderate (salt pans and shore pools) and hypersaline brines (lagoons in the salars), with high pH in the former, respectively close to neutral in the latter, and with low dissolved oxygen levels in lagoons.
More details on the physico-chemical cycles of the brines in Cejas lagoon (Salar de Atacama), the best studied and known in the Chilean Andes, are given in Zdfiiga et al. (1994).
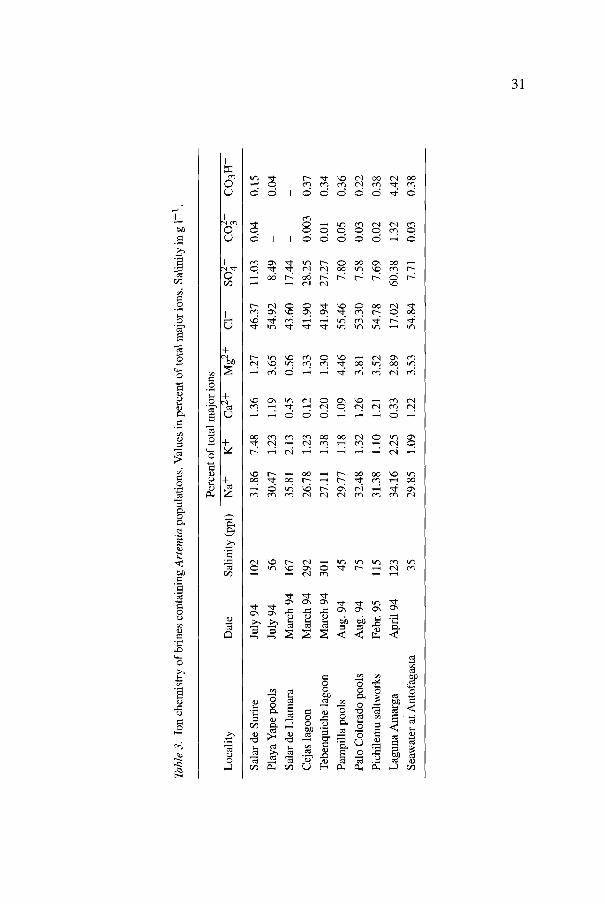
A summary of the patterns of cation and anion dominance according to the salinity in these brines is given in Table 3. Among cations Na + is invariably dominant, and Mg 2+ always exceeds Ca 2+ in brines from seawater origin as well as in endorrheic brines, especially at high salinities. Among anions, C1- is usually dominant at the lower salinities in seawater and salar brines, but SO~- increases substantially in endorrheic lagoons in the salares of high salinities, which is attributed (Zdfiiga et al., 1991) to the volcanic nature of the Andes in this area.
All Artemia populations found in these biotopes were bisexual with male/female ratio 1:1. They usually showed a low density, and reproductive females displayed different offspring output, with a dominant ovoviviparity in lagoons from salars, and different levels of oviparity in solar salt pans (Pichilemu) and shore pools (Yape, Palo Colorado, Pampilla). Cysts were only available in small quantities from the rocky pools at Playa Yape. Most cysts were obtained from the cultured populations in the laboratory. It was
30
Table2. Physico-chemical parameters of brines in hypersaline biotopes where Artemia populations appeared between 1994-1997.
Locality Date t ~ C Salinity pH Oxygen (g 1-1) (ml 1-1)
Salar de Surire July 94 4.50 102 8.50 0.60 Playa Yape pools July 94 30 56 8.50 3.20 Salar de Llamara March 94 28 167 7.50 0.50 Cejas lagoon March 94 23 292 7.30 0.80 Tebenquiche lagoon March 94 24.20 301 7.10 0.40 Pampilla pools Aug. 94 17 45 10.10 9.60 Palo Colorado pools Aug. 94 21.10 75 8.10 2.60 Pichilemu saltworks Febr. 95 37 115 7.80 0.90 Laguna Amarga April 97 8.50 120 12 1
never possible to collect cysts from the pools or lagoons located in Atacama, Llamara and Surire salars.
Table 4 shows the average hydrated cyst diameter and newly-hatched nauplius size of these autochthonous populations (except Surire), with San Francisco Bay, California (USA) and Laguna Grande de Hidalgo, La Pampa (Argentina), as references for conspecific and different strains. These sizes are very similar among all populations and show no significant differences (p _< 0.05).
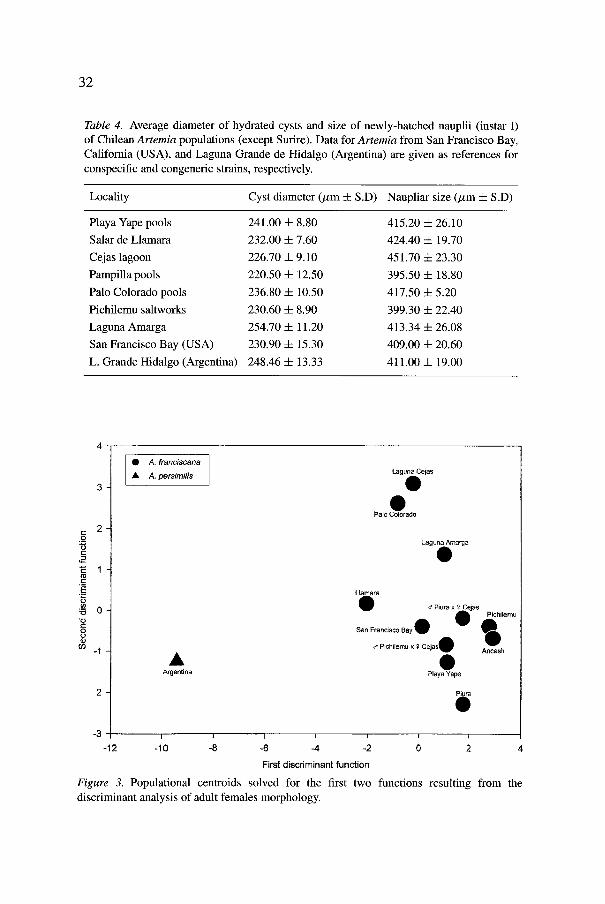
The biometrical characterization of adult females through morpholog- ical study by discriminant analysis is shown in Figure 3. The centroids (discriminant functions evaluated by the morphological variables measured for each population) for the first two discriminant functions calculated, are plotted. This simplification is advisable in view of the high significance levels obtained by the analysis. Table 5 shows the means and standard deviations of the morphological variables measured in the females of each population.
These results confirm the morphological relationships among the studied populations and the group of bisexual populations from Perti (Piura and Ancash), and the USA (San Francisco Bay), belonging to the A. francis- cana Kellogg species. They support the existence of differences between the Argentinean populations, belonging to the bisexual species A. persimilis Prosdocimi and Piccinelli, according to the morphological studies by Hontoria and Amat (1992b) and to electrophoretic evidence provided by Gajardo (1993) and Gajardo and Beardmore (1993). The size and shape of specimens from the Chilean populations are closely related to the values
31
0
C~
o
32
Table 4. Average diameter of hydrated cysts and size of newly-hatched nauplii (instar I) of Chilean Artemia populations (except Surire). Data for Artemia from San Francisco Bay, California (USA), and Laguna Grande de Hidalgo (Argentina) are given as references for conspecific and congeneric strains, respectively.
Locality Cyst diameter (/zm 4- S.D) Naupliar size (/zm 4- S.D)
Playa Yape pools 241.00 4- 8.80 415.20 4- 26.10
Salar de Llamara 232.00 -4- 7.60 424.40 • 19.70
Cejas lagoon 226.70 4- 9.10 451.70 -t- 23.30
Pampilla pools 220.50 4- 12.50 395.50 4- 18.80
Palo Colorado pools 236.80 4- 10.50 417.50 4- 5.20
Pichilemu saltworks 230.60 4- 8.90 399.30 4- 22.40
Laguna Amarga 254.70 4- 11.20 413.34 4- 26.08
San Francisco Bay (USA) 230.90 -4- 15.30 409.00 4- 20.60
L. Grande Hidalgo (Argentina) 248.46 -t- 13.33 411.00 4- 19.00
2
"E 1
E_
"o 0
8 03 -1
-2
0 A. franciscana
�9 A. persimilis Laguna Cejas
Palo Colorado
Laguna Amarga
Llarnara
o" Pichilemu x ~ Cejas O Ancash
Argentina Playa Yape
Piura
-3 i i i i i i i
- 12 - 10 -8 -6 -4 -2 0 2 4
First discriminant function
Figure 3. Populational centroids solved for the first two functions resulting from the discriminant analysis of adult females morphology.

33
~o ~ •

34
of the population from San Francisco Bay (California, USA) and of the two Peruvian populations (Piura and Ancash). The morphological data of the hybrid populations obtained from crosses between Chilean populations (Pichilemu x Cejas), as well as between Chilean and Peruvian ones (Piura x Cejas), are very conclusive to ascertain the relationships in this A.franciscana group.
The separation is very evident along the first discriminant function (Figure 3). The variables that mostly influence this function are the size of the caudal furca, the number of furcal setae, and the general appearance, related to the proportion of the abdominal length to the total length. The females of the Argentinean population display a much shorter and sometimes even rudi- mentary furca, with one or two setae, compared to the A. franciscana females that normally show more than ten furcal setae. The A. persimilis females appear to be longer and thinner because of their relatively long abdomen, compared to the Chilean populations.
The results obtained from cross-breeding trials and the reproductive performance tests (oviparity vs. ovoviviparity) are presented in Table 6. Data are arranged to observe possible or suspected reproductive isolation between Chilean populations from diverse origin (Cejas vs. Palo Colorado, Pichilemu vs. Cejas), between Chilean populations (Cejas and Palo Colorado) and the autochthonous population from Laguna Grande de Hidalgo - Argentina (A. persimilis). In general terms the ovoviviparous progeny produced by matings among Chilean Artemia populations, and between these and San Francisco Bay (USA) and Piura (Perti) was viable for the most part. These popula- tions can be considered conspecific, in spite of levels up to 11 per cent of complete abortive offspring observed in Cejas vs. Palo Colorado and Piura vs. Cejas, and an unexpected level of mortality of Cejas females when mated with Pichilemu males.
The pairings arranged between the Chilean populations from Cejas lagoon and Palo Colorado shore pools, with the Argentinean one from Laguna Grande de Hidalgo, predominantly produced ovoviviparous abortive offspring, especially when Hidalgo males were mated with Cejas females. These females used to die before producing their first offspring; to obtain 79 broods 98 females mated with 34 males, producing 73 per cent completely abortive ovoviviparous offspring, and a few nauplii that did not attain further development. The encysted offspring obtained did not hatch. Palo Colorado females mated to the Argentinean males produced encysted and ovovivi- parous abortive offspring in a similar proportion, with a few cases in which abortive nauplii appeared together with cysts. This could be explained by assuming that these cysts showed subitaneous hatching. Similarly, the Argen- tinean females mated to Chilean males (Cejas and Palo Colorado) produced
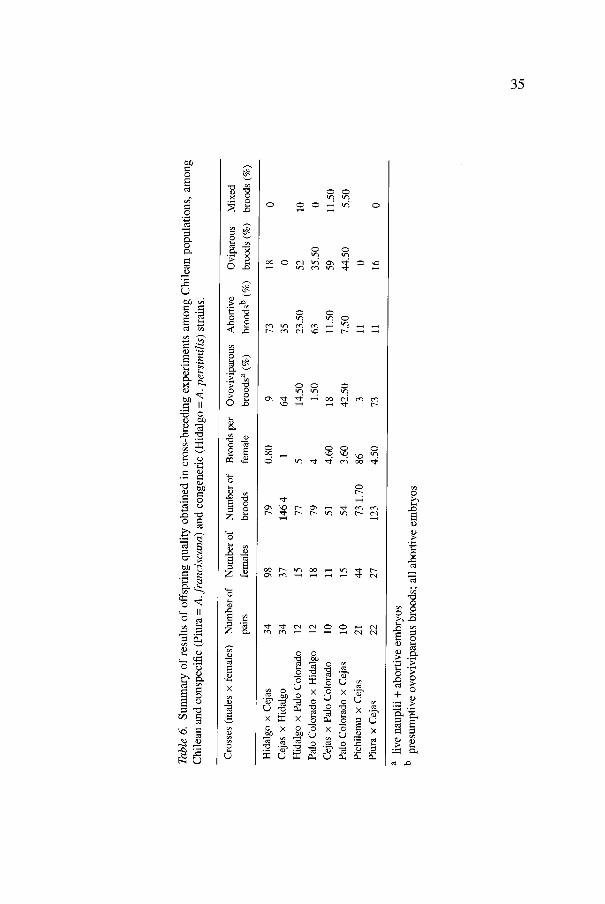
35
O
O
= ,m 0
r,.) ~ d
~'~
~ g
a'g o .,,-
~ ' ~
~.~
eg: ~ ' ~
~ e
,s �9
�9
.s >. �9
�9
s
o .o
= z
= z
,.o
z
N
•
N g m
m . ~ g ~ . .
�9 o
x ~ x S ~ S ~
�9
b~
0
r ,..o

36
the same offspring diversity, with a clear dominance of ovoviviparous abortive offspring.
The pairings among the Chilean populations from Palo Colorado, Cejas and Pichilemu gave predominantly viable ovoviviparous offspring. Nauplii obtained from Cejas females mated to Pichilemu and Piura males were grown up to adulthood and produced viable hybrid F1 and F2 populations. Finally, a similar situation to that described with Argentinean males mating to Cejas females could be observed when these females were mated to Pichilemu males, i.e. there was a certain level of female mortality and the mean value for offspring/female ratio was less than 2, while it was possible to get more than 4 broods per female in all cases.
Discussion
Artemia was found in nine of the twenty studied places. Four sites occur on the sea shore, and the other five locations belong to lacustrine systems evolved in Pre-Andean and Andean salars. While the coastal biotopes belong to diverse geographical regions, from North (Regi6n I) to South (Regi6n IV, VI), the endorrheic sites appear in the three Northern regions (I, II, liD, and in Patagonia (Regi6n XII), counted among the most arid areas in the world, due to a combination of geological, hydrological, volcanic and climatic factors that gave origin to vast saline plains with a salt composition consisting of sulphates, chlorides, nitrates, borates, carbonates and iodates (Chong, 1988; Gajardo et al., 1992).
Although a few more locations are cited in the available biogeographic data (Gajardo, 1993), this survey can be considered as fairly exhaustive, at least in the northern part of Chile.
The physico-chemical features of these biotopes are cyclically influenced by the climate, especially in the endorrheic lagoons and seashore pools, and by human activities in solar salterns. Our results, in agreement with Soto et al. (1994), Campos et al. (1996) and Saijo et al. (1995), show that the ionic composition in all these biotopes is dominated by sodium among the cations, with the sequence Na + K > Mg > Ca, common in saline lakes and hypersaline brines from seawater origin. Chloride and sulphate were the dominant anions. The sulphates represented 9-20 per cent of total anions in the seashore biotopes (C1 > SO4 > HCO3.CO3) but accounted for up to 60- 70 per cent of total anions in endorrheic systems (SO4 > C1 > HCO3.CO3). Similar sequences were found in the endorrheic lacustrine ecosystems of Regi6n III (lagoons of Santa Rosa, Verde, Negro Francisco and Salar de Pedernales), where high levels of borates were evidenced (1.3-3.3 per cent of total anions).

37
According to Bayly (1993), the absence of brine shrimp populations in lagoons at salars in the Chafiaral and Copiap6 provinces (Regi6n III) is related to low salinities; Daphniopsis sp. (Cladocera) and Boeckella poopoensis (Copepoda, Centropagidae) were observed in these habitats. B. poopoensis was so abundant in Santa Rosa lagoon (Salar de Maricunga) that it accu- mulated into thick blood-red patches in small puddles along the shoreline (Dejoux, 1993). This halobiont copepod is often found in saline waters up to 80 g L -1 while nearly all 14 species of Boeckella cited in South America (Bayly, 1992), are confined to freshwater. In the Central Andes no lakes have both Boeckella and Artemia (Hurlbert et al., 1984). Salinity is a fairly good predictor for the presence of these crustaceans, with Artemia occurring at the higher salinities.
The observed Artemia populations showed different female reproductive performance. Oviparous females were only found in the solar salt pans and pools near the sea shore, while in ponds and lagoons located in inland endorrheic ecosystems there was exclusive ovoviviparity. The unstable and stressful behaviour of coastal biotopes might explain this difference. The rocky pools at Playa Yape, Pampilla and Palo Colorado, as well as saltpan ponds, dry up periodically. Artemia populations living there must produce cysts as a major survival mechanism. On the contrary, the inland saline waters found in salars, hydrologically connected with underground water from the Andes, are permanent and only show variations in salinity and ionic composi- tion; this environmental stability should promote a permanent ovoviviparous reproduction. Similarly the decline in cyst production in commercial salt- erns in Macau (Rio Grande do Norte, Brazil) has been claimed to be due to the gradual elimination of the genotypes predisposed towards oviparity (Berthelemy-Okazaki and Hedgecock, 1987; Tackaert and Sorgeloos, 1991).
Phenotypic differentiation among Chilean Artemia populations, and between these and San Francisco (A. franciscana) or Laguna Grande de Hidalgo (A. persimilis), was not possible using cyst and nauplii biometrics, as they do not display significant differences.
The evidence provided by the morphometric characterization and multivariate discriminant analysis confirms the assumption that Chilean populations of Artemia belong to the species A. franciscana, and are not related to the neighbouring Artemia populations from Argentina (A. persimilis). This evidence is further strengthened through the cross-breeding results.
Our data generally agree with the widely accepted concept of Mayr (1942) of biological species as "reproductively isolated non-interbreeding popula- tions". This stresses the validity of the morphometric approach, supported as well by Gajardo and Beardmore (1993) and Gajardo (1993).

38
In conclusion, we confirmed the exclusive presence of A. franciscana in Chile, along its 4000 km coastline occurring in very different hypersaline ecosystems. It remains an open question how A. franciscana was dispersed in these diverse habitats and in the surrounding countries like Perti and, perhaps, Bolivia. Its presence in neighbouring and similar areas in Argen- tina is equally unknown, since there is an absolute lack of information on Artemia occurrence in hypersaline ecosystems in the N W Andean provinces of Argentina, and on their taxonomy. The verified presence of A. persimilis in the Central Provinces (Buenos Aires, La Pampa) is the only identification of Artemia made in this country. Further prospection and characterization of Artemia populations in the South Cone will contribute to the understanding of the zoogeographical distribution and the genetic differentiation process of Artemia species in the New World.
Acknowledgements
This work was supported by Programa FONDECYT (Chile) and Programa de Cooperaci6n Cientffica con Iberoamtr ica (Spain), and further coordination support through Programa CYTED (Spain).
References
Amat, F., Hontoria, E, Ztifiiga, O. and Wilson, R. 1994. Localizaci6n y caracterizaci6n de poblaciones chilenas del crust~ceo Artemia (branquitpodo anostr~iceo). Mem. VII Congreso Latinoamericano de Acuicultura, pp. 83-99. Bogota, Colombia.
Amat, E, Barata, C., Hontoria, E, Navarro, J.C. and Var6. I. 1995a. Biogeography of the genus Artemia (Crustacea, Branchiopoda, Anostraca) in Spain. International Journal of Salt Lake Research 3: 175-190.
Amat, E, Barata, C. and Hontoria, E 1995b. A Mediterranean origin for the Veldrif (South Africa) Artemia Leach population. Journal of Biogeography 22: 49-59.
Anderson, T.W. 1984. An Introduction to the Multivariate Statistical Analysis. John Wiley & Sons, New York, N.Y. (USA).
Bayly, I.A.E. 1992. Fusion of the genera Boeckella and Pseudoboeckella (Copepoda) and revision of their species from South America and sub-Antartic islands. Revista Chilena de Historia Natural 65" 17-63.
Bayly, I.A.E. 1993. The fauna of athalassic saline waters in Australia and the Altiplano of South America: Comparisons and historical perspectives. Hydrobiologia 267: 225-231.
Berthelemy-Okazaki, N.J and Hedgecock, D. 1987. Effect of environmental factors on cyst formation in the brine shrimp Artemia. In: E Sorgeloos, D.A. Bengtson, W. Decleir and E. Jaspers (eds)Artemia Research and Its Applications, Vol. 3, pp. 167-182. Universa Press, Wetteren, Belgium.
Campos, H. 1984. Limnological study of Araucanian lakes (Chile). Verhandlungen Interna- tionale Vereinigung Limnologie 22: 1319-1327.

39
Campos, H., Soto, D., Steffen, W., Parra, O., Agtiero, G. and Ztifiiga, L. 1994. Limnological study of Lake Sarmiento (Chile): A subsaline lake from Chilean patagonian of South America. Archiv ftir Hydrobiologie (Suppl. 99) (1/2): 217-234.
Campos, H., Soto, D., Parra, O., Steffen, W. and Agtiero, G. 1996. Limnological studies of Amarga Lagoon, Chile: A saline lake in Patagonian South America. International Journal of Salt Lake Research 4: 301-314.
Chong, G. 1984. Die Salare in Nordchile - Geologie, Strnktur und Geochimie. Geotekt, Forschung, 67, 146 p.
Chong, G. 1988. The Cenozoic saline deposits of the Chilean Andes between 18~ ~ and 27~ south latitude. In: H. Bahlburg, Ch. Breitkreuz and E Giese (eds) Lecture Notes in Eaerth Sciences, Vol. 17, pp. 137-151. The Southern Central Andes. Springer-Verlag, Berlin.
Dejoux, C. 1993. Benthic invertebrates of some saline lakes of the Sud Lipez region, Bolivia. Hydrobiologia 267: 257-267.
Drago, E. and Quir6s, R.E. 1996. The hydrochemistry of the inland waters of Argentina: A review. International Journal of Salt Lake Research 4:315-325.
Gajardo, G., Wilson, R. and Zfifiiga, O. 1992. Report on the occurrence of Artemia in a saline deposit of the Chilean Andes (Branchiopoda, Anostraca). Crustaceana 63(2): 169-174.
Gajardo, G. and Beardmore, J.A. 1993. Electrophoretic evidence suggests that the Artemia found in the Salar de Atacarna, Chile, is A.franciscana Kellog. Hydrobiologia 257: 65-71.
Gajardo, G. 1993. Genetical and phenotypical characterization of Artemia strains from South America: Relevance for aquaculture. Memorias II Congreso Ecuatoriano de Acuicultura, pp. 91-95. ESPOL, Ecuador.
Gajardo, G., da Concei~ao, M., Weber, L. and Beardmore, J.A. 1995. Genetic variability and interpopulational differentiation of Artemia strains from South America. Hydrobiologia. 302: 21-29.
Hammer, U.T., 1986. Saline Lake Ecosystems of the World. Junk Eds. Dordrecht. Hontoria, E 1990. Caracterizaci6n de tres poblaciones originarias del area levatina espafiola
del crust~iceo branqui6podo Artemia. Aplicaci6n en acuicultura. Tesis, Universidad Aut6noma de Barcelona, 326 pp.
Hontoria, E and Amat, F. 1992a. Morphological characterization of adult Artemia (Crustacea, Branchiopoda) from different geographical origin. Mediterranean populations. Journal of Plankton Research 14(7): 949-959.
Hontoria, E and Amat, E 1992b. Morphological characterization of adult Artemia (Crustacea, Branchiopoda) from different geographical origin. American populations. Journal of Plankton Research 14(10): 1461-1471.
Hurlbert, S.H., Lopez, M. and Keith, J.O. 1984. Wilson's phalarope in the Central Andes and its interaction with the Chilean flamingo. Revista Chilena de Historia Natural 57: 47-57.
Mayr, E. 1942. Systematics and the Origin of Species. Columbia University Press, New York. Prrez, M.L., Valverde, J.R., Batuecas, B., Amat, F., Marco, R. and Garesse, R. 1994. Speci-
ation in the Artemia genus: Mitochondrial DNA analysis of bisexual and parthenogenetic brine shrimps. Journal of Molecular Evolution 38: 156-168.
Pilla, E.J.S. and Beardmore, J.A. 1994. Genetic and morphometric differentiation in Old World bisexual species of Artemia (the brine shrimp). Heredity 73: 47-56.
Saijo, Y., Mitamura, O. and Tanaka, M. 1995. A note on the chemical composition of lake waters in Laguna Amarga, a saline lake in Patagonia, Chile. International Journal of Salt Lake Research 4(2): 165-167.

40
Servant-Vildary, S. and Mello e Sousa, S.H. 1993. Palaeohydrology of the quaternary saline lake Ballivian (southern Bolivian Altiplano) based on diatom studies. International Journal of Salt Lake Research 2(1): 69-95.
Soto, D., Campos, H., Steffen, W., Parra, O. and Ztifiiga, L. 1994. Linmology of the Torres del Paine lake district (Chilean Patagonia): A case of pristine N-limited lakes and ponds. Archiv ftir Hydrobiologie (Suppl. 99): 181-197.
Strickland, J.D. and Parsons, T.R. 1972. A Practical Handbook of Seawater Analysis. Fisheries Research Board of Canada, Bulletin 167, 310 pp.
Tackaert, W. and Sorgeloos, P. 1991. Semi-intensive culturing in fertilized ponds. In: R.A. Browne, P. Sorgeloos and C.N.A. Trotman (eds)Artemia Biology, pp. 287-315. CRC Press, Boca Raton, Florida.
Triantaphyllidis, G.V., Abatzopoulos, T.J. and Sorgeloos, P. 1998. Review of the biogeography of the genus Artemia (Crustacea, Anostraca). J. Biogeography 25: 213-226.
TURISTEL. 1994. Gufa turfstica del Norte de Chile. NORTE. Arica a La Ligua. TURISCON and CTC, Santiago de Chile, 216 pp.
Vanhaecke, P. and Sorgeloos, P. 1980. The biometrics of Artemia strains from different geographical origin. In: G. Persoone, P. Sorgeloos, O. Roels and E. Jaspers (eds) The Brine Shrimp Artemia, Vol. 3, pp. 393-405. Universa Press, Wetteren, Belgium, 456 p.
Vanhaecke, P., Tackaert, W. and Sorgeloos, P. 1987. The biogeography of Artemia: An updated review. In: P. Sorgeloos, D.A. Bengtson, W. Decleir and E. Jaspers (eds) Artemia Research and Its Applications, Vol. 1, pp. 129-155. Universa Press, Wetteren, Belgium. 380 pp.
Williams, W.D., Carrick, T.R., Bayly, I.A.E., Green, J. and Herbst, D.B. 1995. Invertebrates of salt lakes of the Bolivian Altiplano. International Journal of Salt Lake Research 4(1): 65-77.
Wilson, R.O., Ztifiiga, O. and Ramos, R. 1993. Estudio comparativo de la morfologfa del lobulo frontal de cuatro cepas chilenas de Artemia con microscopia electr6nica. Estudios Oceanol6gicos: 12.
Ztifiiga, L.R., Campos, V., Pinochet, H. and Prado, B. 1991. A limnological reconnaissance of Lake Tebenquiche, Salar de Atacama, Chile. Hydrobiologia 210: 19-24.
Zfifiiga, O., Wilson, R., Ramos, R., Retamales, E. and Tapia, L. 1994. Ecologfa de Artemia franciscana en la laguna Cejas, Salar de Atacama (Chile). Estudios Oceanol6gicos 13: 71-84.
Related Documents











