Soil Biology & Biochemistry 38 (2006) 3083–3094 Distribution and abundance of fungi in the soils of Taylor Valley, Antarctica $ L. Connell a, , R. Redman c , S. Craig a , R. Rodriguez b,c a School of Marine Sciences, University of Maine, Orono, ME 04469, USA b US Geological Survey, WFRC, Seattle, WA 98115, USA c University of Washington, Biology Department, Seattle, WA 98195, USA Received 1 September 2005; received in revised form 16 February 2006; accepted 21 February 2006 Available online 4 April 2006 Abstract The occurrence and distribution of culturable fungi in Taylor Valley, Antarctica was assessed in terms of soil habitat. Soil transects throughout the valley revealed differential habitat utilization between filamentous and non-filamentous (yeast and yeast-like) fungi. In addition, there were significant differences in species distribution patterns with respect to soil pH, moisture, distance from marine coastline, carbon, chlorophyll a, salinity, elevation and solar inputs. Filamentous fungal abundance is most closely associated with habitats having higher pH, and soil moistures. These close associations were not found with yeast and yeast-like fungi demonstrating that yeast and yeast-like fungi utilize a broader range of habitat. An intensive survey of the Victoria Land is necessary to gain a better understanding of their role in soil functioning and nutrient cycling processes. r 2006 Elsevier Ltd. All rights reserved. Keywords: Fungi; Yeast; Dry Valleys; Victoria Land; Soil biology; Antarctica 1. Introduction Antarctica contains some of the most extreme habitats on earth, yet exposed soil areas in Victoria Land harbor a diversity of below ground biota. In less extreme habitats, soil microbiota are known to mediate most ecosystem processes. These communities control decomposition, soil respiration, the uptake and fixation of micro- and macro- nutrients, detoxification of heavy metals, and serve as major global carbon sinks (Schlesinger, 1991; Beare et al., 1995). The complexity of soil communities in temperate regions pose difficulties in studying the relationships between biotic and abiotic parameters of ecosystem function, and the factors controlling populations of soil microbiota remain poorly understood (Freckman and Virginia, 1998). The extreme climate and relatively simple community structure of the continental Antarctic desert regions offer a unique opportunity to define interactions between soil organisms that are central to many ecosystem processes. The soil community of the Antarctic polar desert is comprised of few species of bacteria, fungi, and invertebrates (Wynn-Williams, 1996). This ecosystem is primarily driven by abiotic rather than biotic factors (primary production is limited) with no vascular plants or large land animals. Taylor Valley in the McMurdo Dry Valley area of Victoria Land, has three permanently ice covered lakes (Fryxell, Hoare, and Bonney) and the valley sides are networked with glaciers and ephemeral streams. Liquid water is intermittently available to soil communities for a short period of time (1–2 months) during the austral summer season (Conovitz et al., 1998). The valley can be divided into three zones: coastal (next to the permanent ice cover of Explorers Cove); central (the Lake Fryxell and Lake Hoare basins); and interior (Lake Bonney basin and Taylor Glacier) (Marchant and Denton, 1996). The valley zones differ primarily in soil moisture, with the lowest ARTICLE IN PRESS www.elsevier.com/locate/soilbio 0038-0717/$ - see front matter r 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.soilbio.2006.02.016 $ This manuscript is submitted for inclusion in the special edition ‘‘Synthesis of Soil Biodiversity and Ecosystem Functioning in Victoria Land, Antarctica’’. Corresponding author. Tel.: +1 207 581 2470; fax: +1 207 581 2801. E-mail address: [email protected] (L. Connell).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
0038-0717/$ - se
doi:10.1016/j.so
$This manu
‘‘Synthesis of S
Land, Antarctic�CorrespondE-mail addr
Soil Biology & Biochemistry 38 (2006) 3083–3094
www.elsevier.com/locate/soilbio
Distribution and abundance of fungi in the soilsof Taylor Valley, Antarctica$
L. Connella,�, R. Redmanc, S. Craiga, R. Rodriguezb,c
aSchool of Marine Sciences, University of Maine, Orono, ME 04469, USAbUS Geological Survey, WFRC, Seattle, WA 98115, USA
cUniversity of Washington, Biology Department, Seattle, WA 98195, USA
Received 1 September 2005; received in revised form 16 February 2006; accepted 21 February 2006
Available online 4 April 2006
Abstract
The occurrence and distribution of culturable fungi in Taylor Valley, Antarctica was assessed in terms of soil habitat. Soil transects
throughout the valley revealed differential habitat utilization between filamentous and non-filamentous (yeast and yeast-like) fungi. In
addition, there were significant differences in species distribution patterns with respect to soil pH, moisture, distance from marine
coastline, carbon, chlorophyll a, salinity, elevation and solar inputs. Filamentous fungal abundance is most closely associated with
habitats having higher pH, and soil moistures. These close associations were not found with yeast and yeast-like fungi demonstrating that
yeast and yeast-like fungi utilize a broader range of habitat. An intensive survey of the Victoria Land is necessary to gain a better
understanding of their role in soil functioning and nutrient cycling processes.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Fungi; Yeast; Dry Valleys; Victoria Land; Soil biology; Antarctica
1. Introduction
Antarctica contains some of the most extreme habitatson earth, yet exposed soil areas in Victoria Land harbor adiversity of below ground biota. In less extreme habitats,soil microbiota are known to mediate most ecosystemprocesses. These communities control decomposition, soilrespiration, the uptake and fixation of micro- and macro-nutrients, detoxification of heavy metals, and serve asmajor global carbon sinks (Schlesinger, 1991; Beare et al.,1995). The complexity of soil communities in temperateregions pose difficulties in studying the relationshipsbetween biotic and abiotic parameters of ecosystemfunction, and the factors controlling populations of soilmicrobiota remain poorly understood (Freckman and
e front matter r 2006 Elsevier Ltd. All rights reserved.
ilbio.2006.02.016
script is submitted for inclusion in the special edition
oil Biodiversity and Ecosystem Functioning in Victoria
a’’.
ing author. Tel.: +1207 581 2470; fax: +1 207 581 2801.
ess: [email protected] (L. Connell).
Virginia, 1998). The extreme climate and relatively simplecommunity structure of the continental Antarctic desertregions offer a unique opportunity to define interactionsbetween soil organisms that are central to many ecosystemprocesses. The soil community of the Antarctic polar desertis comprised of few species of bacteria, fungi, andinvertebrates (Wynn-Williams, 1996). This ecosystem isprimarily driven by abiotic rather than biotic factors(primary production is limited) with no vascular plants orlarge land animals. Taylor Valley in the McMurdo DryValley area of Victoria Land, has three permanently icecovered lakes (Fryxell, Hoare, and Bonney) and the valleysides are networked with glaciers and ephemeral streams.Liquid water is intermittently available to soil communitiesfor a short period of time (1–2 months) during the australsummer season (Conovitz et al., 1998). The valley can bedivided into three zones: coastal (next to the permanent icecover of Explorers Cove); central (the Lake Fryxell andLake Hoare basins); and interior (Lake Bonney basin andTaylor Glacier) (Marchant and Denton, 1996). The valleyzones differ primarily in soil moisture, with the lowest

ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–30943084
moistures occurring in the interior zone and the highestmoisture in the coastal zone. The soils are poorlydeveloped, lacking both clay and organic materials, withvery weak buffering capacity (Campbell et al., 1998). Thesesoils are primarily unconsolidated alluvium consisting ofsand-sized particles imbedded in cobble and boulders,composed of gneiss, diorite and schist (Bockheim, 1997).Typic Anhyturbels are most abundant in the coastal regionof Taylor Valley (Bockheim, 2002) while the drier interiorregions are dominated by Typic Anhyorthels. The interiorsection has considerable salt crusts on the soils (Campbellet al., 1998; Bockheim, 2002) correlating to the age of theexposed surface.
Fungi (yeast, yeast-like, and filamentous species) arewidely dispersed and in low abundance in the Antarctic(Vishniac, 1996). Yeast and yeast-like fungi have beenidentified in virtually all of the Antarctic habitats rangingfrom cryptoendolithic communities at high elevationsin the Asgard Mountain Range in the McMurdo DryValleys region of Victoria Land (70 1S) to filling adominant role in some sub-Antarctic (below 50 1S) soils(Vishniac, 1996), as well as from marine communities (Fell,1974). One early study found as many as 27 filamentousspecies comprising 17 genera in air and soil samples takenfrom various locations around Antarctica (Sun et al.,1978), particularly those near established camps. Inmaritime and the sub-Antarctic, a number of mycorrhizalfungal species have been foundwith liverworts andmosses (Moller and Gams, 1993; Williams et al., 1994;Tosi et al., 2002) and endemic fungi were found in wood athistoric huts on Ross Island in Victoria Land (Blanchetteet al., 2004). Additionally, a diverse of cosmopolitanfungi have been isolated from the McMurdo Dry Valleylakes and in areas associated with human activity (Baubliset al., 1991).
Although filamentous and single cellular fungi havebeen isolated from a diversity of Antarctic soil types,only yeast are known to be endemic in the drier polardesert soils (Vishniac and Klinger, 1986; Vishniac, 1993).Beyond their role as decomposers, fungi play a function inearly site colonization, soil structure development,and transformation of soil nutrients (e.g. N, S, P, Fe)into bioavailable forms for movement into the foodweb (Carroll and Wicklow, 1992; Wicklow and Soder-strom, 1997). In addition, fungi appear to be oneof the principle taxa in the drier Antarctic soils capableof synthesizing sterols required by soil invertebrates(Weete, 1974; Nes and McKean, 1977). Yet, thedistribution, importance, and function of fungi in theMcMurdo Dry Valley soil ecosystem remains poorlydescribed.
The aim of this study was to determine the distributionand abundance of fungi in soils from the three landscapezones of Taylor Valley. Here, we present data on fungalspecies distribution patterns in relation to soil pH, salinity,moisture, carbon, chlorophyll-a (chl a), spatial locations,and solar inputs.
2. Methods and materials
2.1. Field sample sites
Three cross-valley transects were established in TaylorValley, Victoria Land, Antarctica, during the 2002–2003field season (November 2002–February 2003) (Fig. 1 andTable 1). The sites were chosen non-randomly to possess adiversity of geomorphic features that ranged from rela-tively moist near-marine influence, including sites nearlakes or ponds, as well as ephemeral streams to dry (LakeBonney basin). Sites were chosen to span a range ofexposed soil and glacial till ages as reconstructed byBurkins et al. (2000). These glacial tills are described asRoss I (12.4–23.8 ka), Taylor II/Bonney (74–98 ka), TaylorIII (200–210 ka) and Taylor IV (2100–3700 ka). Thesamples represent a range in soil moisture from 0.2 to7.7% (Table 1). Sample sites were in areas with soilclassification as sandy or sandy-skeletal, mixed Gelisols,dominated by Orthels (Bockheim, 2003). The sites spanneda horizontal distribution from the Taylor Valley floor toabove the predicted ancient Lake Washburn level (above350m) (Hall et al., 2000), which is a proposed ecologicallegacy consisting of ancient pools of organic nutrients fromcommunities in past climate regimes (Doran et al., 1994).These organics are potential relics, which may functionallylink ancient and modern communities (Moorhead et al.,1999). A range of solar input was also used to determinepotential sample sites. All sites were chosen to be free ofany visible algal mats or moss. Transect one (interior LakeBonney basin) had eight sample sites because of the need toresample locations higher in elevation than were collectedduring the first sampling trip. The two remaining transectshad six sample sites each, for a total of 20 sample sitesthrough-out the valley.At each sampling site a 10m square grid was laid out,
separated into quadrants (A, B, C and D) with a north/south and an east/west axis. From each quadrant two pitswere sampled, resulting in eight pits per sample site. Theselection of the sample pits within each quadrant wasdetermined by throwing a plastic ring (Aerobie, AerobieInc.). The internal diameter of the Aerobie ring (20 cm)determined the diameter of each sample pit. A photographwas taken using a fish-eye lens from the center of the gridfor use in determination of total solar input. Latitude andlongitude coordinates were taken from the center of eachgrid using a Trimble geodetic Real-Time Kinematic GPSreceiver (UNAVCO) unit with sub-meter accuracy. Soilwas aseptically removed from the top 10 cm of each samplepit (�1 kg), separated from larger material using a USStandard soil testing sieve, size No. 4 (pre-sterilized with70% ethanol), and collected in a sterile plastic bag.Additionally, a 50 g sub-sample of soil was collected forchl a analysis and covered immediately with aluminum foilto eliminate photo-bleaching of the chlorophyll. Allsamples were stored at 0 1C at field camps until transportto McMurdo Station based Crary Laboratory where they

ARTICLE IN PRESS
Fig. 1. Sample site locations in Taylor Valley, Antarctica (2002–2003). Sites are identified by labels, 300m elevation level is denoted with a yellow dotted
line. The red dot indicates the location of Taylor Valley in Antarctica.
L. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–3094 3085
were stored at 4 1C and processed within 12 h of arrival.Each sample was homogenized thoroughly in the plasticbag prior to sub-sampling for chemical and microbiologicalanalysis.
2.2. Soil chemistry
Soil pH, conductivity, moisture, and salinity: Soil salinitywas estimated from soil conductivity measurements(Rhodes, 1982). Soil moisture was determined using agravimetric method (Barrett et al., 2002). For soil pH a 1:2solution (soil:distilled deionized water) was allowed torehydrate for at least 2 h at 20 1C. Values were measureddirectly at 20 1C using a benchtop Orion 720A pH meter(Forester, 1998). Electrical conductivity was measuredusing a Corning 311 conductivity meter.
Original Carbon: A sub-sample of each soil wasanalyzed for original carbon using a Carlo Erba 1500element analyzer (Carlo Erba, Milan Italy) (Nelsonand Sommers, 1982) by Crary Laboratory staff. Totalcarbon was determined for one sample pit (A1) onlyfor each site.
Chlorophyll a analysis: The sub-sample taken for chl a
was kept frozen until ready for analysis by McMurdoStation Crary Laboratory staff using standard acetone:di-methyl sulfoxide extraction and flurometric quantification(Burkins et al., 2000).
2.3. Culturing and identification of microorganisms
Fungal cultures: A 100 g sub-sample of soil from eachsample pit was mixed with 100ml sterile water in a 250mlsterile flask and shaken at 150 rpm and 4 1C for 1 h. Theliquid slurry was decanted into 50ml centrifuge tubes andcentrifuged for 30m at 8500 rpm in a table-top IECClinical centrifuge, or until the supernatant became clear,at which time the supernatant was removed and the pelletwas resuspended in sterile water to a total volume of5ml. A total of six culture plates (100mm) were inoculatedwith 100 ml each representing 1 g soil per plate. Threereplicate sub-samples were analyzed per pit (18 platesper sample pit). Because the initial aim of this studywas the isolation of yeast and yeast-like taxa, onlyone medium was used, a standard yeast culture medium(YPD with 0.25mg/ml chloramphenicol and 0.1mg/mlampicillin). The plates were incubated at 15 1C for 4 weeksand colonies were assessed twice weekly. Colonies weredivided into broad morphological groups of filamentous(including molds) and non-filamentous (yeast and yeast-like taxa) and counted. Representative colonies werefurther isolated for identification to the genus level.Because of the similarity of many yeast colony morphol-ogies, not all of the representative isolates were identified,these cultures are currently being identified both morpho-logically and genetically.

ARTICLE IN PRESS
Table
1
SoilandSiteCharacteristics
Site
Coordinates
Sitetype
Elevation
(m)
pH
Salinityppt
Chl
a
(mg/kg)
Moisture
(%)
Carbon
(mg/gm)
Filamentous
(TotalCFU/site)
Non-filamentous
(TotalCFU/site)
02T10
162.53347E,�77.57735S
I,N,2,P
420
9.170.3
1.370.8
na
0.570.2
0.263
00
02T11
162.56153E,�77.68501S
I,N,4,F
240
8.270.1
3.171.9
61.371.4
0.441
10
02T12
162.57525E,�77.68645S
I,N,3,F
145
8.570.4
2.071.7
10
0.570.2
0.334
70
02T13
162.58082E,�77.68763S
I,V,2,F
95
8.870.5
0.070
125
3.072.3
0.231
134
3618
02T14
162.58549E,�77.68935S
I,S,2,F
100
9.270.1
0.470.2
205
0.2
70
0.284
51
33
02T15
162.60000E,�77.69152S
I,S,3.F
190
8.570.2
0.970.7
19
0.370.1
0.211
511
02T16
162.61019E,�77.69325S
I,S,3,F
255
8.470.3
2.572.7
13
0.670.6
0.262
14
0
02T17
162.63880E,�77.69669S
I,S,2,F
425
8.370.2
7.578
na
1.371
0.262
11
02T21
163.08927E,�77.58914S
C,N
,2,P
410
9.770.5
0.270.1
389
1.570.4
0.440
43809
0
02T22
163.12703E,�77.59268S
C,N
,2,F
145
9.770.5
0.771.4
50
1.771
0.593
9146
1006
02T23
163.17437E,�77.59744S
C,N
,1,F
25
9.570.4
0.670.5
258
1.670.9
1.694
651
9
02T24
163.11118E,�77.62396S
C,V,1,F
22
9.270.6
0.570.8
59
1.17
0.7
0.458
39
31
02T25
163.04220E,�77.64006S
C,S,3,F
165
8.370.6
2.271.6
30.770.2
0.348
10
23
02T26
162.88478E,�77.64649S
C,S,4,F
420
9.970.2
0.270.2
23
2.071.5
0.617
18903
0
02T31
163.43414E,�77.54323S
M,N
,2,P
345
9.770.2
0.370.1
270
1.470.5
0.693
17793
3
02T32
163.43224E,�77.55430S
M,N
,2,F
125
9.070.3
0.170.1
305
3.770.6
0.668
4500
255
02T33
163.49225E,�77.57515S
M,V,1,F
20
7.870.4
7.877.0
569
3.971.6
1.358
1377
1
02T34
163.59409E,�77.58408S
M,S,1.F
70
8.870.6
1.071.1
25
3.571.4
0.587
19292
32
02T35
163.51933E,�77.58880S
M,S,1,F
100
9.470.4
0.470.3
22
2.870.9
0.604
2244
12
02T36
163.53705E,�77.61191S
M,S,2,F
340
1070.5
0.770.6
230
1.370.4
0.805
725
14
Sitetypes
foreach
locationare
asfollows:I¼
interior,C¼
central,M¼
marine;
Glaicaltilltype,
1¼
Ross
I,2¼
TaylorII/Bonney,3¼
TaylorIII,4¼
TaylorIV
(from
Burkinset
all2000);Solar
Input,F¼
fulldailysolarinputP¼
partialdailysolarinput;Aspect,N¼
Northfacingaspect,S¼
South
facingaspect,V¼
valley
bottom.A
meanvariationofpH,salinity,moisture,andcarbonfor
the8pitsper
sample
site
with7standard
deviation.Totalcarbonandchlorophyll
avalues
are
forA1pitonly
per
sample
site.na¼
notavailable.
L. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–30943086

ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–3094 3087
DNA extraction: DNA was extracted from individualcolonies of representative cultures using DNEasy PlusDNA extraction kit (Qiagen Inc) following manufacturersdirections. The concentration of recovered DNA wasdetermined using a picogreen dsDNA quantification kit(Molecular Probes Inc) modified for use with a TurnerDesigns Picofluor handheld fluorometer following manu-facturers instructions.
DNA sequencing and analysis: The internal transcribedspacer (ITS) of the ribosomal genes were used foridentification of selected isolated colonies. ExtractedDNA from fungal isolates was used as template in PCRreactions. The ribosomal DNA for the ITS region betweenprimers ITS4 and ITS5 (White et al., 1990) were amplifiedas previously described (Connell, 2000). The resultantamplicons were purified and sequenced using previouslydescribed methods (Connell, 2000). The subsequent chro-matograms were edited using the Sequencher v 4.1program and compared (BLAST search) with sequencesfrom the international GenBank database for identifica-tion. ITS sequence were submitted to GenBank (Table 2).
2.4. Solar radiation
Solar radiation for each sample site was determined bytwo methodologies. Data inferred by Geographic Informa-tion System (GIS) coverages from previous studies (Danaet al., 1998) and direct measurement with hemisphericalphotography using HemiView analysis software version2.1(Delta-T Devices, Ltd.). HemiView software was used tocalculate solar input at site factors (described below).Hemispherical photography is particularly suited fordetermining potential solar input levels in areas withirregular terrain features. Photographs were taken inuniform sky conditions, looking upwards using a 1801fisheye lens and digital camera. The images were imported
Table 2
GenBank numbers for ITS rDNA sequences from fungal isolates collected in
Type Taxa (isolate #) GenBank BankIt nu
F1 Geomycetes sp. (0 1 0) DQ402527
F3 Chaunopycnis sp. (076) DQ402530
F3 Nematoctonus sp. (016) DQ402531
F5 Phaeosphaeria sp (004) DQ402538
F5 Phoma sp. (104) DQ402526
NF1 Thelebolus microsporus (009) DQ402525
NF1 Thelebolus sp. (068) DQ402532
NF2 Rhodotorula mucilaginosa (133) DQ402533
NF2 Dioszegia sp. 1 (101) DQ402528
NF3 Dioszegia sp 2 (116) DQ402529
NF3 Rhodosporidium kratochvilovae (053) DQ402534
NF4 Leucosporidium sp. (120) DQ402535
NF4 Cryptococcus nyarrowii (027) DQ402536
NF5 Cryptococcus saitoi (125) DQ402537
Genus and species identification, GenBank accession number, and the nearest a
indicates broad morphological group identification (F ¼ filamentous fungi, N
into HemiView to calculate solar input levels above andbelow terrain features, and hence direct, indirect (diffuse)and global site factors, as well as calculate the occurrenceof sunflecks and associated solar irradiance on any chosenday of the year. Site factors are indices of the proportion ofirradiance reaching a given location. Values range from 0to 1, with 0 being no solar input (complete sky obstruction)and 1 being the irradiance for an open location (completesky visibility ¼ no sky obstruction). Indirect site factor(ISF) is the proportion of diffuse solar input reaching agiven location relative to a location with no sky obstruc-tions. Direct site factor (DSF) is the proportion of directsolar input reaching a given location relative to that in alocation with no sky obstructions. Global site factor (GSF)is the proportion of potential global solar input (direct plusdiffuse) under a given location relative to that in the open(Anderson, 1964). Because many of the surroundingmountains were snow covered, and would therefore be aconfounding factor in image analysis, the area of the skywas delineated manually in digital images prior toimportation into HemiView for analysis.
2.5. Data analysis
Statistical analysis was performed using StatView ver-sion 5.0.1 (SAS). All regressions utilized log transformedvalues (log (1+total cfu)) for the dependent variables(fungal abundance). Stepwise regression utilized 13 inde-pendent variables (soil moisture, pH, carbon, chl a,salinity, distance from marine coast, distance from nearestglacier, distance from nearest lake or stream, elevation,solar radiation, and solar inputs [DSF, ISF, GSF]) tomodel significant variables. Statistical significance wasdetermined where Po0:10. Geo-referenced data wascompiled, organized, and mapped using ArcGIS version8 (ESRI).
Taylor Valley soil
mber Closest match Score (bits)
Geomyces sp. AY345347 1437
Chaunopycnis pustulata AF389193 825
Nematoctonus robustus U51978 733
Phaeosphaeria sp AY345346 1031
Phoma herbarum AY293788 601
Thelebolus microsporus DQ028268 908
Thelebolus caninus AY957550 904
Rhodotorula mucilaginosa AF444541 1088
Dioszegia changbaiensis AY242817 521
Dioszegia fristingensis AY562158 543
Rhodosporidium kratochvilovae AF444587 1086
Leucosporidium antarcticum AF444529 551
Cryptococcus nyarrowii AF400697 1001
Cryptococcus saitoi AF444372 1148
lignment from NCBI BLAST with score (bits) are shown for isolates. Type
F ¼ non-filamentous fungi).
ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–30943088
3. Results
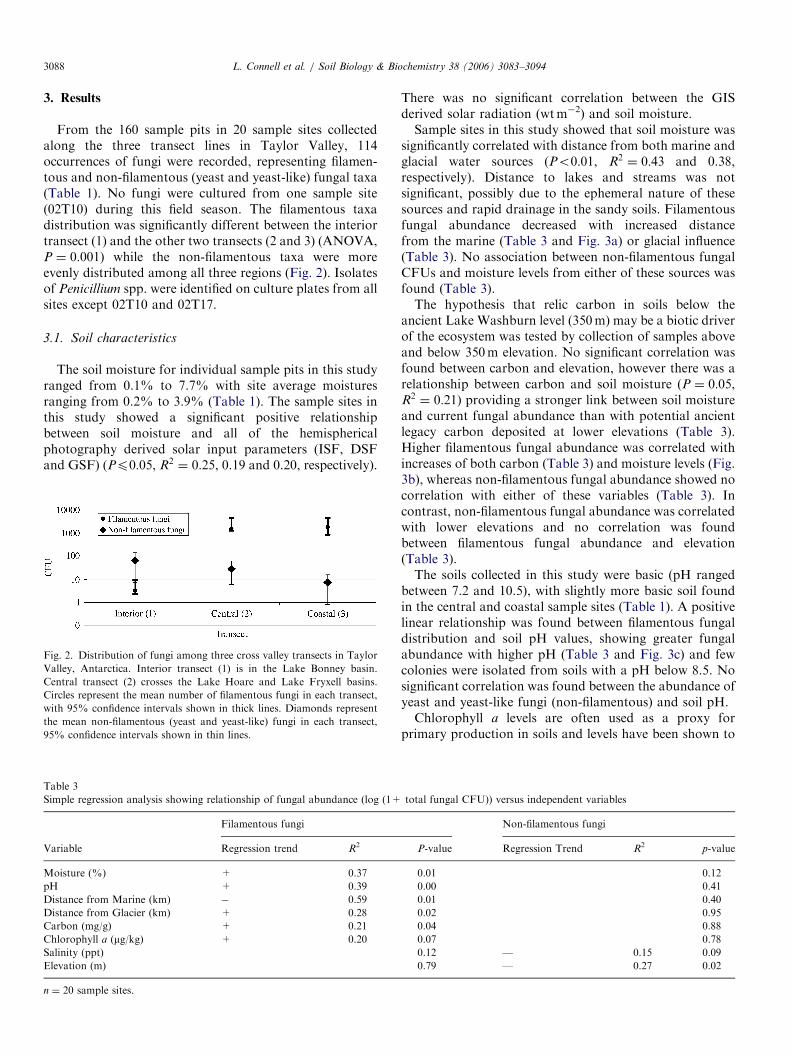
From the 160 sample pits in 20 sample sites collectedalong the three transect lines in Taylor Valley, 114occurrences of fungi were recorded, representing filamen-tous and non-filamentous (yeast and yeast-like) fungal taxa(Table 1). No fungi were cultured from one sample site(02T10) during this field season. The filamentous taxadistribution was significantly different between the interiortransect (1) and the other two transects (2 and 3) (ANOVA,P ¼ 0:001) while the non-filamentous taxa were moreevenly distributed among all three regions (Fig. 2). Isolatesof Penicillium spp. were identified on culture plates from allsites except 02T10 and 02T17.
3.1. Soil characteristics
The soil moisture for individual sample pits in this studyranged from 0.1% to 7.7% with site average moisturesranging from 0.2% to 3.9% (Table 1). The sample sites inthis study showed a significant positive relationshipbetween soil moisture and all of the hemisphericalphotography derived solar input parameters (ISF, DSFand GSF) (Pp0:05, R2 ¼ 0:25, 0.19 and 0.20, respectively).
Table 3
Simple regression analysis showing relationship of fungal abundance (log (1+
Filamentous fungi
Variable Regression trend R2
Moisture (%) + 0.37
pH + 0.39
Distance from Marine (km) � 0.59
Distance from Glacier (km) + 0.28
Carbon (mg/g) + 0.21
Chlorophyll a (mg/kg) + 0.20
Salinity (ppt)
Elevation (m)
n ¼ 20 sample sites.
Fig. 2. Distribution of fungi among three cross valley transects in Taylor
Valley, Antarctica. Interior transect (1) is in the Lake Bonney basin.
Central transect (2) crosses the Lake Hoare and Lake Fryxell basins.
Circles represent the mean number of filamentous fungi in each transect,
with 95% confidence intervals shown in thick lines. Diamonds represent
the mean non-filamentous (yeast and yeast-like) fungi in each transect,
95% confidence intervals shown in thin lines.
There was no significant correlation between the GISderived solar radiation (wtm�2) and soil moisture.Sample sites in this study showed that soil moisture was
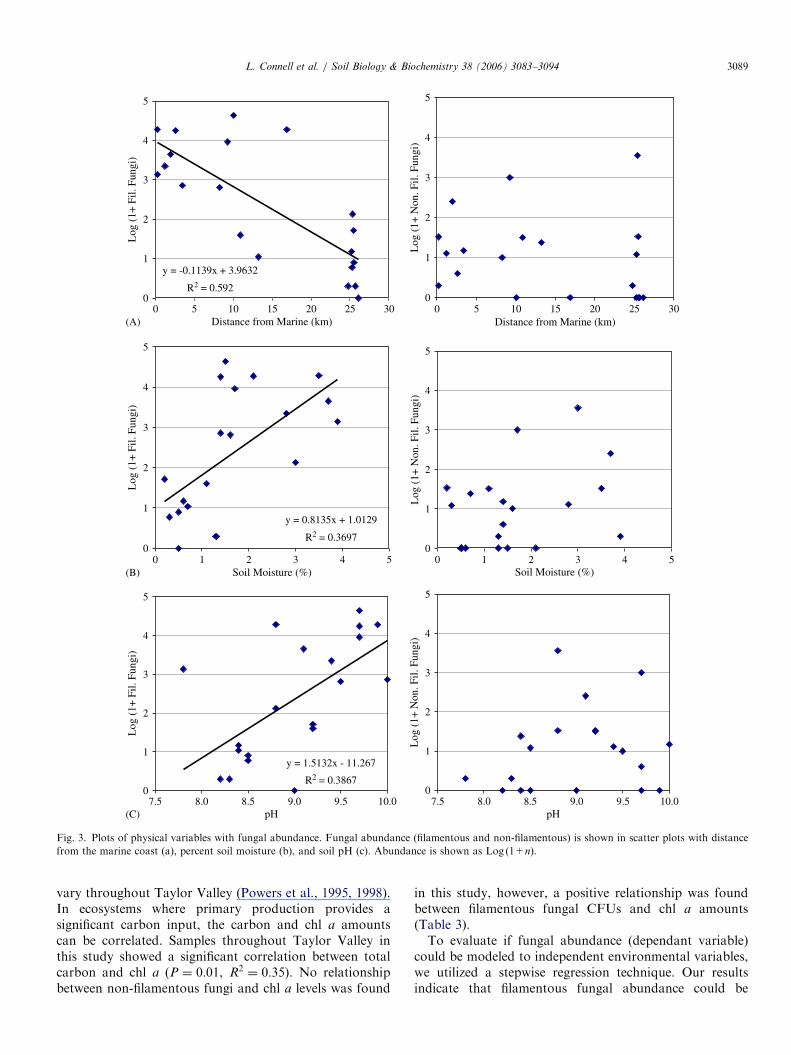
significantly correlated with distance from both marine andglacial water sources (Po0:01, R2 ¼ 0:43 and 0.38,respectively). Distance to lakes and streams was notsignificant, possibly due to the ephemeral nature of thesesources and rapid drainage in the sandy soils. Filamentousfungal abundance decreased with increased distancefrom the marine (Table 3 and Fig. 3a) or glacial influence(Table 3). No association between non-filamentous fungalCFUs and moisture levels from either of these sources wasfound (Table 3).The hypothesis that relic carbon in soils below the
ancient Lake Washburn level (350m) may be a biotic driverof the ecosystem was tested by collection of samples aboveand below 350m elevation. No significant correlation wasfound between carbon and elevation, however there was arelationship between carbon and soil moisture (P ¼ 0:05,R2 ¼ 0:21) providing a stronger link between soil moistureand current fungal abundance than with potential ancientlegacy carbon deposited at lower elevations (Table 3).Higher filamentous fungal abundance was correlated withincreases of both carbon (Table 3) and moisture levels (Fig.3b), whereas non-filamentous fungal abundance showed nocorrelation with either of these variables (Table 3). Incontrast, non-filamentous fungal abundance was correlatedwith lower elevations and no correlation was foundbetween filamentous fungal abundance and elevation(Table 3).The soils collected in this study were basic (pH ranged
between 7.2 and 10.5), with slightly more basic soil foundin the central and coastal sample sites (Table 1). A positivelinear relationship was found between filamentous fungaldistribution and soil pH values, showing greater fungalabundance with higher pH (Table 3 and Fig. 3c) and fewcolonies were isolated from soils with a pH below 8.5. Nosignificant correlation was found between the abundance ofyeast and yeast-like fungi (non-filamentous) and soil pH.Chlorophyll a levels are often used as a proxy for
primary production in soils and levels have been shown to
total fungal CFU)) versus independent variables
Non-filamentous fungi
P-value Regression Trend R2 p-value
0.01 0.12
0.00 0.41
0.01 0.40
0.02 0.95
0.04 0.88
0.07 0.78
0.12 — 0.15 0.09
0.79 — 0.27 0.02
ARTICLE IN PRESS
0
1
2
3
4
5
y = 1.5132x - 11.267
R2 = 0.3867
7.5 8.0 8.5 9.0 9.5 10.0pH
Log
(1+
Fil.
Fun
gi)
07.5 8.0 8.5 9.0 9.5 10.0
pH
1
2
3
4
5
Log
(1+
Non
. Fil.
Fun
gi)
0
1
2
3
4
5
Log
(1+
Fil.
Fun
gi)
y = -0.1139x + 3.9632
R2 = 0.5920
1
2
3
4
5
0 10 15 20 25 30Distance from Marine (km)
Log
(1+
Fil.
Fun
gi)
0 10 15 20 25 30Distance from Marine (km)
0
1
2
3
4
5
Log
(1+
Non
. Fil.
Fun
gi)
5 5
y = 0.8135x + 1.0129
R2 = 0.3697
0 3Soil Moisture (%)
1 2 4 50
1
2
3
4
5
Log
(1+
Non
. Fil.
Fun
gi)
Soil Moisture (%)0 31 2 4 5
(A)
(B)
(C)
Fig. 3. Plots of physical variables with fungal abundance. Fungal abundance (filamentous and non-filamentous) is shown in scatter plots with distance
from the marine coast (a), percent soil moisture (b), and soil pH (c). Abundance is shown as Log (1+n).
L. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–3094 3089
vary throughout Taylor Valley (Powers et al., 1995, 1998).In ecosystems where primary production provides asignificant carbon input, the carbon and chl a amountscan be correlated. Samples throughout Taylor Valley inthis study showed a significant correlation between totalcarbon and chl a (P ¼ 0:01, R2 ¼ 0:35). No relationshipbetween non-filamentous fungi and chl a levels was found
in this study, however, a positive relationship was foundbetween filamentous fungal CFUs and chl a amounts(Table 3).To evaluate if fungal abundance (dependant variable)
could be modeled to independent environmental variables,we utilized a stepwise regression technique. Our resultsindicate that filamentous fungal abundance could be
ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–30943090
attributed to three of all 13 variables measured for thisstudy, and these were soil moisture, pH and distance fromthe marine coastline (Po0:01, R2 ¼ 0:81). Only twoindependent variables could be utilized to significantlymodel non-filamentous fungi abundance, and these weresoil salinity and elevation (Po0:01, R2 ¼ 0:47).
3.2. Species diversity and richness
Colony morphologies were used to initially discerndifferent species and assess diversity in individual samplesand sites. DNA sequence analysis of ribosomal DNA wasused to identify the dominant species in each morphologicgroup. While each group contains a diversity of fungalspecies, the dominant species are listed in Table 4.Additional sequence analysis is underway to define speciesdiversity of each group. The NCBI data base containssequence information of a small percentage of the fungalspecies known to exist. This limited our ability to identifyfungi to species. A GenBank search using partial ITSsequence from several isolates did not always result inpositive identification. These isolates are currently being
Table 4
Distribution of fungi by colony morphology
F1 F2 F3 F4 F5 F6
02T10 — — — — — —
02T11 — — — — — X
02T12 X — — — — X
02T13 X — X — — X
02T14 X — X — — X
02T15 — — — — — X
02T16 X — — — — X
02T17 — — X — — —
02T21 X — X — — X
02T22 X — X — X X
02T23 X — — — X X
02T24 X — X — X X
02T25 X — X — — X
02T26 X — X — — X
02T31 X X X X X X
02T32 X — X X X X
02T33 X X X X X X
02T34 — — X — X X
02T35 X — X — X X
02T36 X — X — X X
The morphologies represent more than one taxa, however, the dominant spec
fungi. Fungal identifications are given below, also see Table 2.
F1—Geomycets sp.
F2—unknown species
F3—Chaunopycnis sp. and Nematoctonus robustus
F4—unknown species
F5—Phaeosphaeria sp.
F6—Penicillium ssp.
NF1—Hyphozyma and Thelebolus spp.
NF2—Rhodotorula sp. and Diozsega spp.
NF3—Rhodosporidium sp. and Diozsega spp. form 2.
NF4—Leucosporidium sp. and Cryptococcus spp.
NF5—Cryptococcus spp. and Trichosporon sp.
NF6—black yeast (undefined)
identified by taxonomic means in other laboratories.Trichosporon spp. were identified using genetic comparisonby Fell et al. (2006) (Trichosporon ovoides, Genbankaccession number DQ111748, T. domesticum, Genbankaccession number DQ111746, and T. louberii, Gen Bankaccession number DQ111745). Penicillium spp. wereidentified to genus by physical taxonomic means only.The black yeast isolated from 02T15 and two filamentousforms (F2 and F4—Table 4) isolated from 02T31, 02T32and 02T33, respectively, could not be maintained in cultureand were not identified further.There was significant spatial discontinuity with regard
to species distribution patterns. For example, in areaswhere large numbers of the same species were found, suchas site 02T21, the population was strongly dominatedby Chaunopycnis sp. while at site 02T13, in the floodplain of Priscu Stream, the population was dominatedby yeast (Trichosporon spp.). Greater fungal speciesrichness was found in other locations. At least six generaof yeast or yeast-like fungi, as well as four genera offilamentous fungi were isolated from soils at site 02T32 inthe coastal zone (Table 4).
NF1 NF2 NF3 NF4 NF5 NF6
— — — — — —
— — — — — —
— — — — — —
X — X — X —
X — — — — —
— — — — X X
— — — — — —
— — X — — —
— — — — — —
X X — X X —
X — X — — —
X X X X — —
— X — X — —
— — — — — —
— — — X X —
X X X X — —
X X — — — —
X — X — — —
X X — — — —
— X — — — —
ies in each is listed below. F ¼ filamentous fungi, NF ¼ non-filamentous

ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–3094 3091
Yeast were found to be relatively abundant in some ofthe driest samples (02T14, 02T15) and Penicillium spp.were the only filamentous fungi found in samples that wereequally as dry (02T11 and 02T12). Of the filamentousfungal isolates identified, two were abundant in the centraland coastal zones, Geomyces sp. and Phaeosphaeria sp.Geomyces is a cosmopolitan eurythermal fungi that hasbeen isolated numerous times from the Antarctic (Sunet al., 1978; Del Frate and Caretta, 1990; Onofri et al.,1994). Phaeosphaeria spp. have also been identified nearlyas often from the sub-Antarctic islands as well as fromVictoria Land (Sun et al., 1978; Del Frate and Caretta,1990; Onofri et al., 1994). These taxa were found in relativeabundance in both the central and coastal transects and aremost likely endemic to the Antarctic. Another fungi thatwas abundant in the coastal region was the nematophagousfungi, Nematoctonus sp. This work did not assess thepresence of nematodes in soil samples, so no correlation ofthe two taxa co-distribution can be made.
4. Discussion
This study represents the first season in a systematicfungal survey of Taylor Valley, Antarctica. The distribu-tion and abundance of fungi in Taylor Valley was greaterthan anticipated, with several undescribed species isolated.There was habitat utilization that limited the distributionof filamentous fungi to the central and coastal zones wheresoils contained more moisture and a higher pH, while non-filamentous taxa were more evenly distributed throughoutthe valley. Even though the absolute numbers of yeast andyeast-like taxa were lower than those of filamentous fungi(Table 1), these organisms appear to be able to utilize abroader range of habitats. The step-wise regression modelshowed a correlation of yeast and yeast-like fungalabundance with both lower elevation and lower salinity.This result may be skewed by the very high number ofTrichosporon spp. found in the flood plain of PriscuStream, 02T13 (Table 1) where salts from the soils arewashed during annual high stream flow periods.
Collectively, the fungi represented cosmopolitan andendemic species. This is best represented by the distributionof Penicillium spp. throughout the valley compared to thelimited distribution of an unidentified black yeast (NF6,Table 4). Penicillium spp. are cosmopolitan and as many asfive species have been found in other studies collected fromVictoria Land soil and air (Cameron and Morelli, 1974;Sun et al., 1978). This study found Penicillium spp. in lownumbers at many of the sample locations throughout theentire valley and not as abundant as Phaeosphaeria sp. orGeomyces sp. Although some Penicillium species associatedwith food are known to be xerotolerant (Dix and Webster,1995), it is possible that the broad distribution ofPenicillium spp. may reflect human activities throughoutthe valley and the spread of spores from contaminated foodsince active growth for many members of this generarequires at least 22% relative humidity (Pieckova and
Jesenska, 1999). Determining the origin of Penicillium spp.would require comparative studies in the areas that havenot had long-term field camps frequented by humans.The predominant water inputs into the Taylor Valley
soils are marine influenced precipitation, glacier melt run-off, as well as lakes and streams (Conovitz et al., 1998).Precipitation is very limited in this ecosystem, with anaverage of only 10 cmyr�1 (Bromley, 1985). Most of thismoisture is lost to sublimation very rapidly, making thesesoils among the driest in the world, having an averagemoisture level below 5% (Freckman and Virginia, 1998).Solar irradiance is an important source of energy input intoTaylor Valley associated with heating of surfaces, andthereby glacial and stream melting that is, in turn, a majorsource of water for soil moisture (Dana et al., 1996, 1998).There can be very short time frames when liquid moistureis available for biological activity, ranging from a fewhundred hours yr�1 in locations along the coast to havingonly one event over a three year period in a valley adjacentto Taylor Valley (Wright Valley) (Wall et al., 2004).A model utilizing moisture, pH, and distance from the
marine coast can be used as a predictor for filamentous, butnot for non-filamentous taxa abundance. This three-variable stepwise regression model for predicting filamen-tous fungal abundance accounted for 81% of the variation.The six linear regressions found in Table 3 also show thatthe filamentous fungal groups appear to be more influencedby environmental gradients than the yeast and yeast-liketaxa. Most of the filamentous fungi found in the areasdistant from the marine coast were Penicillium spp. that areknown to produce spores able to tolerate dry conditions(Corry, 1987). Non-filamentous fungi were not influencedby pH in a linear fashion, however, these taxa appeared tohave a optimal range centering on pH 9 (Fig. 3c).Information about the abundance and distribution of
other soil biota in Taylor Valley is limited. The only wellstudied group are nematodes. Three species of theseorganisms occur throughout the valley and their abun-dance can be correlated with both distance from the marinecoast as well as soil moisture (Courtright et al., 2001). Inthis way the distribution of nematodes is similar to that offilamentous fungi found in this study. However, Courtrightet al. (2001) found an additional parameter that limited thedistribution of nematodes, soil salinity. The nematodestudy found that sites with higher salinities restrictednematode abundance, whereas this study found low pHlimiting to the abundance of filamentous fungi. In this way,nematode distribution and filamentous fungal distributionare dissimilar in regard to soil salinity and pH levels. It willbe interesting to determine if specific fungi are associatedwith nematodes in future work.Moisture levels are key to, not only abundance of
filamentous fungi and nematodes (Courtright et al., 2001),but also other members of soil communities as indicated bycarbon and chl a concentration. Both carbon and chl a
levels used as indicators for primary production areassociated with moisture and independent of elevation in

ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–30943092
this study. This is most likely explained by existence ofpockets of moisture found at sites in higher elevation,where melt water is restricted from drainage by geo-morphic features. For example, site 02T21 is situated closeto a steep slope with coarse and rocky soil texture andsmall melt pools. In contrast, moisture from other potentialmajor water sources as measured by distance from themarine coast, glaciers, and streams or lakes, show differentassociations with filamentous fungal abundance (Table 3).Moisture from marine influence is positively correlatedwith CFU abundance of these taxa, but fungal abundanceis negatively associated with distance from glaciers. Thismay be due to lower annual soil temperatures near glaciers,a parameter not measured in this study. Proximity tostreams and lakes did not predict soil moisture levelssufficient to support a gradient of abundance of filamen-tous fungi. Campbell et al., 1998 has shown that these soilslose moisture rapidly with increased distance from thewater source, diminishing to levels equivalent to surround-ing soils by 8m. Only one site in this study was within 8mof a stream or lake (02T13, near Priscu stream) with amean soil moisture level of 3% (Table 1). This site wassampled during a time when the ephemeral stream wasflowing and annual soil moistures may be lower in highlysandy soils, a parameter not measured in this study.Soil moistures at nearby sample locations were o0.5%(Table 1) showing that the adjacent soils are dryerthan those utilized by most filamentous fungi found inthis study (Fig. 3b).
Solar input can increase the amount of melting and run-off, as well as thawing of soils (Dana et al., 1998), therebyincreasing potential soil moisture. GIS derived solarradiation (wtm�2) was not correlated with soil moisturein this study, thus this methodology was not a goodpredictor of soil moisture. In contrast, potential solar input(ISF, DSF, GSF), as determined through hemisphericalphotography analysis, was found to be a good indicator ofsoil moisture. It is likely that the geographic scale ofsampling used for the GIS derived model (Dana et al.,1998) was not fine enough to show a significant correlation.Our results demonstrate that utilizing GIS solar radiationto select sites for soil moisture levels may be of limitedusefulness. Therefore, the use of hemispherical derived datain studies with few, geographically disjunct sample sitesmay provide more robust information.
It is interesting that correlations between soil nutrientsand fungal distribution abundance is observed in temperateecosystems (Treseder, 2005; Zak, 2005), but only forfilamentous taxa in this study. Since only carbon wasmeasured, it is possible that other chemicals such asphosphorus, sulfur or micronutrients may be limiting thegrowth of the yeast and yeast-like fungi (Treseder, 2005).Alternatively, the lack of chemical versus yeast and yeast-like fungal correlation may be due to an inability to detectthe chemistry responsible for fungal growth. For example,it is possible that these fungi may be mineralizing soilparticles and consuming nutrients as they are generated, a
process that may not be visualized by standard approaches.Additionally, when all of the cultured isolates are fullyanalyzed and assessed separately, significant correlationsmay be found.One of the limitations to understanding biodiversity on
this planet is determining the species richness in complexsubstrates such as soil or sediment (Kurtzman and Fell,2005). The fact that we identified several undescribedspecies may not be a unique phenomenon. The same mayoccur in all soil environments if enough culture conditionsand DNA extractions (for non-culturable organisms) areconsidered. Assessing the abundance and diversity of fungiin soil is complicated by temporal changes in soil chemicaland climatic parameters (Lodge et al., 2005; Zak, 2005).These abiotic aspects of the soil environment may havesignificant affects on the dynamics of the fungal populationstructure. In this study, we assessed one point-in-timeduring the austral summer for each transect. Althoughthese data indicate that fungi are more abundant anddiverse than previously thought, the population structureof these organisms will need to be assessed over time,within and between seasons. Only then will we be able todetermine if these fungi represent non-reproducing relics ofthe past when climatic conditions were more conducive togrowth, or reproductively active populations.
Acknowledgements
The authors wish to thank M. Iszard, J. Perkins, K.Clegg and A. Barrett for laboratory assistance, our TEAparticipant B. Schulz for field work as well as RaytheonPolar Support Service, the LTER Crary AnalyticalLaboratory staff, UNAVCO, and PHI for logistical andlaboratory support while in Antarctica. This research wasfunded by the National Science Foundation (NSF) Officeof Polar Programs grant OPP-0125611 (LC & RR). Thispaper is part of a special issue resulting from NSF OPP-0406141 support to DH Wall for the Victoria LandSynthesis workshop.
References
Anderson, M.C., 1964. The photographic computation of light condition.
Journal of Ecology 52, 27–41.
Barrett, J.E., Virginia, R.A., Wall, D.H., 2002. Trends in resin and KCL-
extractable soil nitrogen across landscape gradients in Taylor Valley,
Antarctica. Ecosystems 5, 289–299.
Baublis, J.A., Wharton Jr., R.A., Volz, P.A., 1991. Diversity of micro-
fungi in an Antarctic dry valley. Journal of Basic Microbiology 31 (1),
3–12.
Beare, M., Coleman, D.C., Crossley Jr., D.A., Hendrix, P.F., Odum, E.P.,
1995. A hierarchical approach to evaluating the significance of soil
biodiversity to biogeochemical cycling. Plant and Soil 170, 5–22.
Blanchette, R.A., Held, B.W., Jurgens, J.A., Mcnew, D.L., Harrington,
T.C., Duncan, S.M., Farrell, R.L., 2004. Wood-destroying soft rot
fungi in historic expedition huts of Antarctica. Applied and Environ-
mental Microbiology 70 (3), 1328–1335.
Bockheim, J., 1997. Properties and classification of cold desert soils from
Antarctica. Soil Science Society of America Journal 61, 224–231.

ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–3094 3093
Bockheim, J., 2003. University of Wisconsin Antarctic Soils Database.
Digital Media. National Snow and Ice Data Center/World Data
Center for Glaciology, Boulder, CO.
Bockheim, J.G., 2002. Landform and soil development in the McMurdo
Dry Valleys, Antarctica: a regional synthesis. Arctic, Antarctic, and
Alpine Research 34 (3), 308–317.
Bromley, A.M., 1985. Weather observations in Wright Valley, Antarctica.
Information Publication 11. New Zealand Meterolgical Service,
Wellington, New Zealand, p. 37
Burkins, M.B., Virginia, R.A., Chamberlain, C.P., Wall, D.H., 2000.
Origin and distribution of soil organic matter in Taylor Valley,
Antarctica. Ecology 81 (9), 2377–2391.
Cameron, R.E., Morelli, F.A., 1974. Viable microorganisms from
Antarctic Ross Island and Taylor Valley Drill cores. Antarctic Journal
of the United States 9, 113–115.
Campbell, D.I., Claridge, G.G.C., Campbell, D.I., Balks, M.R., 1998.
The soil environment of the McMurdo Dry Valleys, Antarctica.
In: Priscu, J.C. (Ed.), Ecosystems Dynamics in a Polar Desert,
vol. 72. American Geophysical Union, Washington DC, USA,
pp. 297–322.
Carroll, G.C., Wicklow, D.T., 1992. The Fungal Community, Its
Organization and Role, in the Ecosystem. Marcel Dekker, Inc.,
New York, USA, p. 976.
Connell, L.B., 2000. Nuclear ITS region of the alga Heterosigma akashiwo
(Chromophyta: Raphidophyceae) is identical in isolates from Atlantic
and Pacific basins. Marine Biology 136 (6), 953–960.
Conovitz, P.A., McKnight, D.M., MacDonald, L.H., Fountain, A.G.,
House, H.R., 1998. Hydrological processes influencing streamflow
variation in Fryxell basin, Antarctica. In: Priscu, J.C. (Ed.),
Ecosystems Dynamics in a Polar Desert: The McMurdo Dry Valleys,
Antarctica, vol. 72. American Geophysical Union, Washington DC,
USA, pp. 93–108.
Corry, J.E.L., 1987. Relationships of water activity to fungal growth. In:
Benchant, L.R. (Ed.), Food and Beverage Mycology. AVI Pub. Co.,
Philadelphia, pp. 51–99.
Courtright, E.M., Wall, D.H., Virginia, R.A., 2001. Determining habitat
suitability for soil invertebrates in an extream environment:
The McMurdo Dry Valleys, Antarctica. Antarctic Science 13 (1),
9–17.
Dana, G.L., Wetzel, M.A., Wharton, Jr. R.A., 1996. Satellite-derived
surface temperatures in the Dry Valleys, Antarctica. In: Smith, W.,
Stamnes, K. (Eds.), IRS ‘96: Current Problems in Atmospheric
Radiation. DEEPAK Publishing, pp. 530–533.
Dana, G.L., Wharton Jr., R.A., Dubayah, R., 1998. Solar radiation in
McMurdo dry valleys, Antarctica. In: Priscu, J.C. (Ed.), Ecosystems
Dynamics in a Polar Desert, vol. 72. American Geophysical Union,
Washington DC, USA, pp. 39–64.
Del Frate, G., Caretta, G., 1990. Fungi isolated from Antarctic material.
Polar Biology 11, 1–7.
Dix, N.J., Webster, J., 1995. Fungal Ecology. Chapman Hall, London,
UK, p. 549.
Doran, P.T., Wharton Jr., R.A., Lyons, W.B., 1994. Paleolimnology of
the McMurdo Dry Valleys, Antarctica. Journal of Paleolimnology 10
(2), 85–114.
Fell, J.W., 1974. Yeasts in Oceanic Regions. In: Jones, E.B.G. (Ed.),
Recent Advances in Aquatic Microbiology. Paul Elek Ltd., London,
pp. 93–124.
Fell, J.W., Scorzetti, G., Connell, L., Craig, S., 2006. Biodiversity of
micro-eukaryotes in Antarctic Dry Valley soils with less than 5% soil
moisture. Soil Biology & Biochemistry, this issue, doi:10.1016/
j.soilbio.2006.01.014.
Forester, J., 1998. Determination of soil pH. In: Alef, K., Nannipieri, P.
(Eds.), Methods in Applied Soil Microbiology and Biochemistry.
Academic Press, San Diego, CA, USA, pp. 55–56.
Freckman, D.H.W., Virginia, R.A., 1998. Soil biodiversity and commu-
nity structure in the McMurdo Dry Valleys, Antarctica. In: Priscu, J.C.
(Ed.), Ecosystems Dynamics in a Polar Desert, vol. 72. American
Geophysical Union, Washington DC, USA, pp. 323–335.
Hall, B.L., Denton, G.H., Hendy, C.H., 2000. Evidence from Taylor
Valley for a grounded ice sheet in the Ross Sea, Antarctica.
Geografiska Annaler 82A (2/3), 275–303.
Kurtzman, C.P., Fell, J.W., 2005. Yeasts. In: Muller, G.M., Bills, G.F.,
Foster, M.S. (Eds.), Biodiversity of Fungi: Inventory and
Monitoring Methods. Elsevier Academic Press, San Diego, CA,
USA, pp. 337–342.
Lodge, D.J., Ammirati, J.F., O’Dell, T.E., Muller, G.M., Huhndorf, S.M.,
Wang, C.J., Stokland, J.N., Schmit, J.P., Ryvarden, L., Leacock, P.R.,
Mata, M., Umana, L., Wu, Q.F., Czederpiltz, D.L., 2005. Terrestrial
and lignicolous macrofungi. In: Muller, G.M., Bills, G.F., Foster, M.S.
(Eds.), Biodiversity of Fungi: Inventory and Monitoring Methods.
Elsevier Academic Press, San Diego, CA, USA, pp. 127–158.
Marchant, D.R., Denton, G.H., 1996. Miocene and pliocene paleoclimate
of the Dry Valley region, Antarctica, southern Victoria Land; A
geomorphological approach. Marine Micropaleontology 27, 253–271.
Moller, C., Gams, W., 1993. Two new hyphomycetes isolated from
Antarctic lichens. Mycotaxon 58, 441–450.
Moorhead, D.L., Doran, P.T., Fountain, A.G., Lyons, W.B., McKnight,
D.M., Priscu, J.C., Virginia, R.A., Wall, D.H., 1999. Ecological
legacies: Impacts on the ecosystems of the McMurdo Dry Valleys.
BioScience 49 (12), 1009–1019.
Nelson, D.W., Sommers, L.E., 1982. Total carbon, organic carbon, and
organic matter. In: Page, A.L., Miller, D.R., Keeney, D.R. (Eds.),
Methods of Soil Analysis, Part 2, Chemical and Microbiological
Methods. SSSA, Madison, WI, ASA Agronomy 9, 539–594.
Nes, W.R., McKean, M.L., 1977. Biochemistry of Steroids and Other
Isopentenoids. University Park Press, Baltimore, MD.
Onofri, S., Tosi, S., Persiani, A.M., Maggi, O., Riess, S., Zucconni, L.,
1994. Mycological researches in Victoria Land Terresterial Ecosys-
tems. In: Battaglia, B., Bisol, P.M., Varotto, V. (Eds.), Proceedings of
the Second Meeting on ‘‘Antarctic Biology’’. University Patavine,
Padova, pp. 19–32.
Pieckova, E., Jesenska, Z., 1999. Microscopic fungi in dwellings and their
health implications in humans. Annual Agricultural Environmental
Medicine 6, 1–11.
Powers, L.E., Freckman, D.H.W., Virginia, R.A., 1995. Spatial distribu-
tion of nematodes in polar desert soils of Antarctica. Polar Biology 15,
325–333.
Powers, L.E., Ho, M., Freckman, D.H.W., Virginia, R.A., 1998.
Distribution, community structure, and microhabitats of soil inverte-
brates along an elevation gradient in the Taylor Valley, Antarctica.
Arctic and Alpine Research 30 (2), 133–141.
Rhodes, J.D., 1982. Soluble salts. In: Page, A.L., Miller, R.H., Keeney,
D.R. (Eds.), Methods of soil analysis Part 2 Chemical and
Microbiological Properties. American Society of Agronomy-Soil
Science of America, Madison, WI, pp. 167–179.
Schlesinger, W.H., 1991. Biogeochemistry. Academic Press, San Diego,
USA, p. 443.
Sun, S.H., Huppert, M., Cameron, R.E., 1978. Identification of some
fungi from soil and air of Antarctica. Antarctic Research Series
(Washington) 30, 1–26.
Tosi, S., Casado, B., Gerdol, R., Caretta, G., 2002. Fungi isolated from
Antarctic mosses. Polar Biology 25, 262–268.
Treseder, K.K., 2005. Nutrient acquisition strategies of fungi and their
relation to elevated atmospheric CO2. In: Dighton, J., White, J.F.,
Oudemans, P. (Eds.), The Fungal Community: Its Organization and
Role in the Ecosystem. Taylor & Francis, Boca Raton, Florida, USA,
pp. 713–731.
Vishniac, H.S., 1993. The microbiology of Antarctic soils. In: Friedmann,
E.I. (Ed.), Antarctic Microbiology. Wiley-Liss Inc., New York, USA,
pp. 297–342.
Vishniac, H.S., 1996. Biodiversity of yeasts and filamentous microfungi in
terrestrial Antarctic ecosystems. Biodiversity and Conservation 5,
1365–1378.
Vishniac, H.S., Klinger, J.M., 1986. Yeasts in the Antarctic deserts. In:
Megusar, F., Ganter, M. (Eds.), Perspectives in Microbial Ecology.
Slovene Society for Microbiology, Ljubljana, Yugoslavia, pp. 46–51.

ARTICLE IN PRESSL. Connell et al. / Soil Biology & Biochemistry 38 (2006) 3083–30943094
Wall, A., Balks, M.R., Campbell, D.I., Paetzold, R.F., 2004. Soil moisture
measurement in the Ross Sea region of Antarctica using Hydra soil
moisture probes. Supersoil 2004: Proceedings of the 3rd Australian
New Zealand Soils Conference, University of Sydney, Australia,
www.regional.org.au/au/asssi/supersoil2004.
Weete, J.D., 1974. Fungal Lipid Biochemistry: Distribution and Metabo-
lism. Plenum Press, New York.
White, T.J., Bruns, T., Lee, S., Taylor, J., 1990. Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In:
Innis, M., Gelfand, J., Sninsky, J., White, T.J. (Eds.), PCR Protocols:
A Guide to Methods and Applications. Academic Press, Orlando, FL,
pp. 315–322.
Wicklow, D.T., Soderstrom, B.E., 1997. Environmental and Microbial
Relationships. Springer, Berlin, pp. 99–114.
Williams, P.G., Roser, D.J., Seppelt, R.D., 1994. Mycorrhizas of
hepatics in continental Antarctica. Mycological Research 98 (1),
34–36.
Wynn-Williams, D.D., 1996. Antarctic microbial diversity: the basis of
polar ecosystem processes. Biodiversity and Conservation 5,
1271–1293.
Zak, J., 2005. Fungal communities of desert ecosystems: links to climate
change. In: Dighton, J., White, J.E., Oudemans, P. (Eds.), The Fungal
Community: Its Organization and Role in the Ecosystem. Taylor &
Francis, Boca Raton, Florida, USA, pp. 659–681.
Related Documents











