© New Phytologist (2004) 163: 381–392 www.newphytologist.org 381 Research Blackwell Publishing, Ltd. Distinct roles of Lotus japonicus SYMRK and SYM15 in root colonization and arbuscule formation Kirill Demchenko 1,2 , Thilo Winzer 3 , Jens Stougaard 4 , Martin Parniske 3 and Katharina Pawlowski 1 1 Albrecht von Haller Institute for Plant Sciences, Plant Biochemistry, Göttingen University, Justus-von-Liebig-Weg 11, 37077 Göttingen, Germany; 2 Komarov Botanical Institute RAS, Prof. Popov St. 2, 197376, St.-Petersburg, Russia; 3 The Sainsbury Laboratory, John Innes Centre, Colney Lane, Norwich NR4 7UH, UK; 4 Department of Molecular Biology, University of Aarhus, Gustav Wieds Vej 10C, 8000 Aarhus C, Denmark Summary • The colonization of Lotus japonicus roots by the arbuscular mycorrhizal fungus Glomus intraradices was analysed in plant mutants affected in the symbiosis genes, SYM15 or SYMRK. SYMRK encodes an LRR receptor-like kinase that is, like the SYM15 gene, essential for both mycorrhizal and rhizobial symbioses. • Different colonization patterns were observed in growing vs meristematically arrested roots. • Three steps in the interaction were differentially impaired in the mutants: surface opening, where the anticlinal cell walls of two adjacent epidermal cells separate from each other in the vicinity of fungal hyphae; intracellular passage of hyphae through an exodermal cell and an adjacent cell of the outermost cortical layer; and arbuscule formation in cells of the two innermost cortical layers. • The combined results indicate that LjSYMRK is required for the intracellular passage through exodermis and outermost cortical cell layer whereas LjSYM15 is required for surface opening and arbuscule formation. Key words: Lotus japonicus , Glomus intraradices , arbuscular mycorrhiza, arbuscule, root meristem, signal transduction, saprophytic growth. © New Phytologist (2004) 163 : 381–392 Author for correspondence: Martin Parniske Tel. + 44 1603 450249 Fax: + 44 1603 450111 Email: [email protected] Received: 12 January 2004 Accepted: 19 March 2004 doi: 10.1111/j.1469-8137.2004.01123.x Introduction Arbuscular mycorrhizae (AM) are symbioses formed between fungi of the order Glomales and the roots of > 80% of the extant terrestrial plant taxa (Newman & Reddell, 1987). The associations are considered mutually beneficial in that the plant supplies the fungus with carbon sources, while the fungus assists the plant with the uptake of phosphate and other mineral nutrients (Smith & Smith, 1997). Plant mutants unable to form AM represent useful tools for the identification of steps in the development of the symbiosis, and a subset of nodulation mutants, affected in their root nodule symbiosis with rhizobia, are also affected in the AM symbiosis (Duc et al., 1989). A large number of Nod mutants from the model legume Lotus japonicus (Handberg & Stougaard, 1992) has been analysed for their AM phenotype and six common SYM genes have been identified, mutation of which affects both rhizobial and AM symbioses. The phenotype of common SYM mutants indicates that the two symbiotic interactions overlap in their early steps (Kistner & Parniske, 2002). One of the common SYM genes is SYMRK whose product is involved in rhizobial Nod factor-dependent signal transduction as well as being required for signal exchange with AM fungi (Stracke et al ., 2002) . However, accumulating evidence also suggests that signalling independent of the common SYM genes contributes to the communication between the symbionts (Kosuta et al ., 2003). Another common SYM gene is SYM15 (Stougaard, 2001); represented in this study by two alleles; the weak Ljsym15-1 (Schauser et al ., 1998) and the strong Ljsym15-2 ( J. Stougaard, unpublished data). In mutants affected in the common SYM genes SYMRK , SYM3 and SYM4 , intraradical colonization of root patches by the fungal symbiont are observed at a low frequency (Wegel et al., 1998). The analysis of such events in Ljsym4 mutants by Novero et al . (2002) has identified three successive steps of

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

©
New Phytologist
(2004)
163
: 381–392
www.newphytologist.org
381
Research
Blackwell Publishing, Ltd.
Distinct roles of
Lotus japonicus SYMRK and SYM15
in
root colonization and arbuscule formation
Kirill Demchenko
1,2
, Thilo Winzer
3
, Jens Stougaard
4
, Martin Parniske
3
and Katharina Pawlowski
1
1
Albrecht von Haller Institute for Plant Sciences, Plant Biochemistry, Göttingen University, Justus-von-Liebig-Weg 11, 37077 Göttingen, Germany;
2
Komarov
Botanical Institute RAS, Prof. Popov St. 2, 197376, St.-Petersburg, Russia;
3
The Sainsbury Laboratory, John Innes Centre, Colney Lane, Norwich NR4 7UH,
UK;
4
Department of Molecular Biology, University of Aarhus, Gustav Wieds Vej 10C, 8000 Aarhus C, Denmark
Summary
• The colonization of
Lotus japonicus
roots by the arbuscular mycorrhizal fungus
Glomus intraradices
was analysed in plant mutants affected in the symbiosisgenes,
SYM15
or
SYMRK. SYMRK
encodes an LRR receptor-like kinase that is, likethe
SYM15
gene, essential for both mycorrhizal and rhizobial symbioses.• Different colonization patterns were observed in growing vs meristematicallyarrested roots.• Three steps in the interaction were differentially impaired in the mutants: surfaceopening, where the anticlinal cell walls of two adjacent epidermal cells separate fromeach other in the vicinity of fungal hyphae; intracellular passage of hyphae throughan exodermal cell and an adjacent cell of the outermost cortical layer; and arbusculeformation in cells of the two innermost cortical layers.• The combined results indicate that LjSYMRK is required for the intracellularpassage through exodermis and outermost cortical cell layer whereas LjSYM15 isrequired for surface opening and arbuscule formation.
Key words:
Lotus japonicus
,
Glomus intraradices
, arbuscular mycorrhiza,arbuscule, root meristem, signal transduction, saprophytic growth.
©
New Phytologist
(2004)
163
: 381–392
Author for correspondence:
Martin ParniskeTel. +44 1603 450249Fax: +44 1603 450111Email: [email protected]
Received:
12 January 2004
Accepted:
19 March 2004
doi: 10.1111/j.1469-8137.2004.01123.x
Introduction
Arbuscular mycorrhizae (AM) are symbioses formed betweenfungi of the order Glomales and the roots of > 80% of theextant terrestrial plant taxa (Newman & Reddell, 1987). Theassociations are considered mutually beneficial in that the plantsupplies the fungus with carbon sources, while the fungusassists the plant with the uptake of phosphate and other mineralnutrients (Smith & Smith, 1997). Plant mutants unable toform AM represent useful tools for the identification ofsteps in the development of the symbiosis, and a subset ofnodulation mutants, affected in their root nodule symbiosiswith rhizobia, are also affected in the AM symbiosis (Duc
et al
., 1989).A large number of Nod mutants from the model legume
Lotus japonicus
(Handberg & Stougaard, 1992) has beenanalysed for their AM phenotype and six common
SYM
geneshave been identified, mutation of which affects both rhizobial
and AM symbioses. The phenotype of common
SYM
mutantsindicates that the two symbiotic interactions overlap in theirearly steps (Kistner & Parniske, 2002). One of the common
SYM
genes is
SYMRK
whose product is involved in rhizobialNod factor-dependent signal transduction as well as beingrequired for signal exchange with AM fungi (Stracke
et al
., 2002)
.
However, accumulating evidence also suggests that signallingindependent of the common
SYM
genes contributes to thecommunication between the symbionts (Kosuta
et al
., 2003).Another common
SYM
gene is SYM15 (Stougaard, 2001);represented in this study by two alleles; the weak
Ljsym15-1
(Schauser
et al
., 1998) and the strong
Ljsym15-2
( J. Stougaard,unpublished data).
In mutants affected in the common
SYM
genes
SYMRK
,
SYM3
and
SYM4
, intraradical colonization of root patches bythe fungal symbiont are observed at a low frequency (Wegel
et al
., 1998). The analysis of such events in
Ljsym4
mutantsby Novero
et al
. (2002) has identified three successive steps of

www.newphytologist.org
©
New Phytologist
(2004)
163
: 381–392
Research382
fungal colonization of the plant root. They have shown thatthe colonization of
Lotus
roots by AM fungi is controlled atthree points: (i) at the intercellular penetration betweenepidermal cells; (ii) at the intracellular passage through theroot exodermis (see also Senoo
et al
., 2000); and (iii) at theformation of arbuscules in the two innermost cortical celllayers. Novero
et al
. (2002) concluded that
LjSYM4
is necessaryfor the accommodation of AM fungi in both epidermaland cortical cells. We have used a detailed cytological analysisto study the position of two common
SYM
genes,
LjSYMRK
(Stracke
et al
., 2002) and
LjSYM15
(Schauser
et al
., 1998, J.Stougaard, unpublished data) in this conceptual frameworkand identified significant differences between the symbioticperformance of the corresponding mutants. We conclude thatthe
SYMRK
and
SYM15
genes have distinct functions duringthe establishment of the arbuscular mycorrhizal symbiosis.
Materials and Methods
Plant growth conditions
To maximize colonization pressure,
L. japonicus
seedlingswere grown in a chive nurse plant system with
G. intraradices
as microsymbiont. For this, seeds of
Allium schoenoprassum
(chive) were surface-sterilised in 1.2% NaOCl for 3–5 min,washed with sterile dd H
2
O, and placed in a pot containingAbsodan (Damolin, Fur, Denmark) in 1/2 strength Hoagland’ssolution (Hoagland & Arnon, 1938) around a chive nurseplant with roots colonized by
G. intraradices
at 22
°
C in 16 hlight, 8 h dark. After 4–6 week, mycorrhization was analysedafter ink staining of single roots, and plants with mycorrhizedroots were transferred to individual pots and grown for4–6 wk.
L. japonicus
seeds were scarified for 10 min inconcentrated H
2
SO
4
, washed extensively in sterile dd H
2
Oand germinated on 1% water agar in Petri dishes, first forone day at 24
°
C in the dark, then for two days at 16 h lowlight intensity, 8 h dark. Ca. 20 seedlings were planted arounda chive nurse plant, grown at 22
°
C and 16 h light, 8 h darkand watered once per week with 1/2 strength Hoagland’ssolution.
Cytological methods
Ink staining for visualization of intra- and extraradical fungalstructures was performed according to Vierheilig
et al
. (1998).Root systems, or parts of roots, were incubated for 7 min in10% KOH at 95
°
C in small Petri dishes on a hot plate,washed 3
×
with dd H
2
O and 2
×
in 1% acetic acid, stainedfor 3 min at 96
°
C with 5% ink (Sheaffer Jet Black) in 8%acetic acid. To remove excessive dye, roots were rinsed with1% acetic acid once for 5 and once for 20 min. To remove airbubbles, roots were infiltrated in vacuum for 15 min. Rootswere mounted on microscope slides in glycerol and observedunder a microscope.
For Trypan Blue staining, roots were either autoclaved for10–15 min or incubated at 90
°
C in 10% KOH for up to2 h. After rinsing with water, they were soaked in 1% HCl atroom temperature for 30 min. Then, HCl was replaced witha Trypan Blue solution (0.05% (w/v) trypan blue, 50% (v/v)glycerol, 0.2% (v/v) HCl) and the roots were incubated at60
°
C for at least 1 h. After rinsing with water, excessive dyewas removed by repeated incubations in acidified glycerol(50% (v/v) glycerol, 0.5% (v/v) (HCl) at room temperature.
To visualize fungal structures in root systems before embed-ding for microscopy, root systems were fixed with Navashin’sfixative (1% chromic acid/40% formalin/100% acetic acid;10 : 4 : 1 (v/v/v); Sass, 1958), or with 4% paraformaldehydeand 0.25% glutaraldehyde overnight at 4
°
C. Roots were washedwith dd H
2
O for 3–4 h. For Feulgen staining, roots wereincubated in 5 N HCl for 40 min and stained with Schiff ’sreagent for 1 h. Schiff ’s reagent was prepared by dissolving5 g of basic Fuchsin (Fluka, Buchs, Switzerland) in 900 mlboiling dd H
2
O, letting the solution cool down, and adding20 ml of conc. HCl at 50
°
C and 20 g of K
2
S
2
O
5
dissolvedin 80 ml H
2
O at 25
°
C. Afterwards, root systems were washed4
×
for 10 min in SO
3
-water (20 g K
2
S
2
O
5
and 20 ml conc.HCl in 1 l dd H
2
O) and for 2
×
20 min in dd H
2
O and observedunder a stereo microscope. Colonized root sections were cutin
c
. 0.5 cm long pieces, dehydrated in a graded ethanol seriesand embedded in Paraplast plus (Sigma, St. Louis, MO, USA).Serial sections (8 µm) were cut with a rotation microtome(RM2125RT; Leica Microsystems, Wetzlar, Germany), attachedto SuperFrost® Plus slides (Roth, Karlsruhe, Germany) anddeparaffinized in xylene. For Feulgen/Alcian Blue staining,sections on slides were rehydrated in a graded ethanol series,equilibrated in dd H
2
O, and incubated in Schiff ’s reagentfor 1–2 h. Then, sections were washed for 3
×
15 min withSO
3
-water and for 2
×
5 min in dd H
2
O before beingincubated for 1–2 h in 1% Alcian Blue 8GX (Sigma) in 3%acetic acid. Sections were washed in dd H
2
O for 3
×
5 min,dehydrated in a graded ethanol series, equilibrated in xyleneand mounted with Eukitt™ (Kindler, Freiburg, Germany).
Microscopy
Microscope BX51 and stereo microscope SZX12 (OlympusOptical GmbH, Hamburg, Germany) were used. Results weredocumented using a ColorView II digital camera set and SISanalySIS® software (Soft Imaging System GmbH, Münster,Germany).
Results
Colonization of
Lotus japonicus
roots by
Glomus intraradices
The cytology of
L. japonicus
ecotype Gifu roots infected by
G. intraradices
was examined in detail. Roots of
L. japonicus
©
New Phytologist
(2004)
163: 381–392 www.newphytologist.org
Research 383
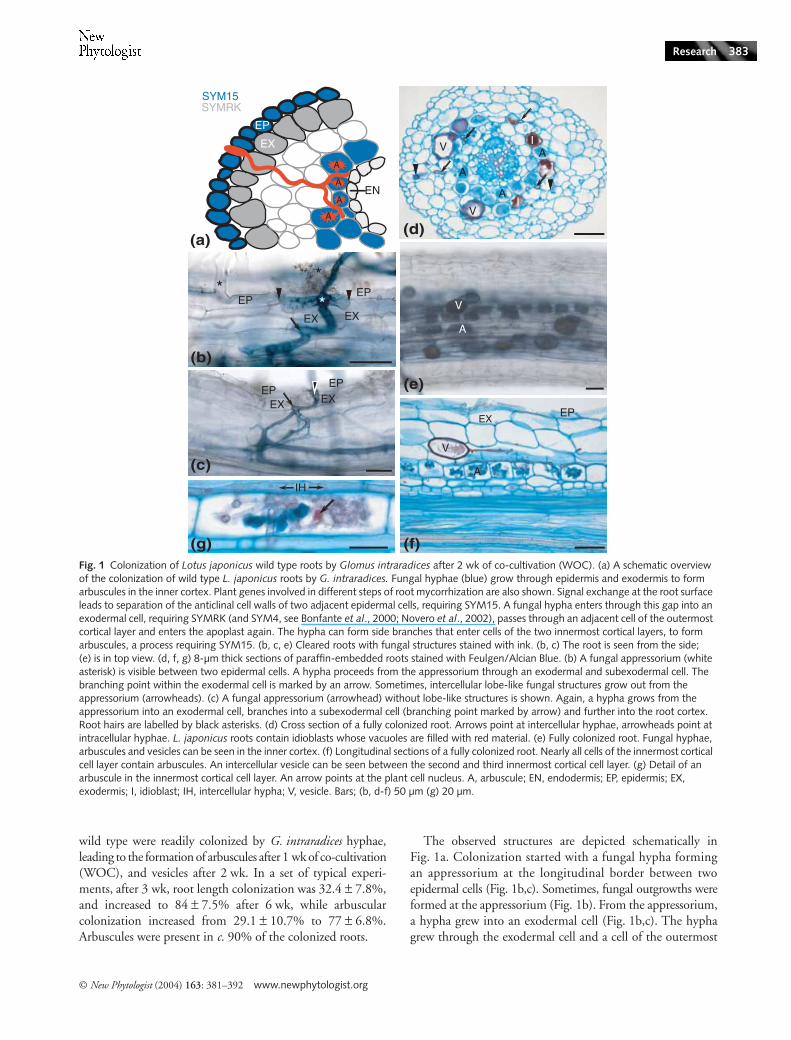
wild type were readily colonized by G. intraradices hyphae,leading to the formation of arbuscules after 1 wk of co-cultivation(WOC), and vesicles after 2 wk. In a set of typical experi-ments, after 3 wk, root length colonization was 32.4 ± 7.8%,and increased to 84 ± 7.5% after 6 wk, while arbuscularcolonization increased from 29.1 ± 10.7% to 77 ± 6.8%.Arbuscules were present in c. 90% of the colonized roots.
The observed structures are depicted schematically inFig. 1a. Colonization started with a fungal hypha formingan appressorium at the longitudinal border between twoepidermal cells (Fig. 1b,c). Sometimes, fungal outgrowths wereformed at the appressorium (Fig. 1b). From the appressorium,a hypha grew into an exodermal cell (Fig. 1b,c). The hyphagrew through the exodermal cell and a cell of the outermost
Fig. 1 Colonization of Lotus japonicus wild type roots by Glomus intraradices after 2 wk of co-cultivation (WOC). (a) A schematic overview of the colonization of wild type L. japonicus roots by G. intraradices. Fungal hyphae (blue) grow through epidermis and exodermis to form arbuscules in the inner cortex. Plant genes involved in different steps of root mycorrhization are also shown. Signal exchange at the root surface leads to separation of the anticlinal cell walls of two adjacent epidermal cells, requiring SYM15. A fungal hypha enters through this gap into an exodermal cell, requiring SYMRK (and SYM4, see Bonfante et al., 2000; Novero et al., 2002), passes through an adjacent cell of the outermost cortical layer and enters the apoplast again. The hypha can form side branches that enter cells of the two innermost cortical layers, to form arbuscules, a process requiring SYM15. (b, c, e) Cleared roots with fungal structures stained with ink. (b, c) The root is seen from the side; (e) is in top view. (d, f, g) 8-µm thick sections of paraffin-embedded roots stained with Feulgen/Alcian Blue. (b) A fungal appressorium (white asterisk) is visible between two epidermal cells. A hypha proceeds from the appressorium through an exodermal and subexodermal cell. The branching point within the exodermal cell is marked by an arrow. Sometimes, intercellular lobe-like fungal structures grow out from the appressorium (arrowheads). (c) A fungal appressorium (arrowhead) without lobe-like structures is shown. Again, a hypha grows from the appressorium into an exodermal cell, branches into a subexodermal cell (branching point marked by arrow) and further into the root cortex. Root hairs are labelled by black asterisks. (d) Cross section of a fully colonized root. Arrows point at intercellular hyphae, arrowheads point at intracellular hyphae. L. japonicus roots contain idioblasts whose vacuoles are filled with red material. (e) Fully colonized root. Fungal hyphae, arbuscules and vesicles can be seen in the inner cortex. (f) Longitudinal sections of a fully colonized root. Nearly all cells of the innermost cortical cell layer contain arbuscules. An intercellular vesicle can be seen between the second and third innermost cortical cell layer. (g) Detail of an arbuscule in the innermost cortical cell layer. An arrow points at the plant cell nucleus. A, arbuscule; EN, endodermis; EP, epidermis; EX, exodermis; I, idioblast; IH, intercellular hypha; V, vesicle. Bars; (b, d-f) 50 µm (g) 20 µm.

www.newphytologist.org © New Phytologist (2004) 163: 381–392
Research384
cortical layer and re-enters the root apoplast (Fig. 1d,e). Ahyphal network was established in the apoplast of the innerroot cortex. Typically, intercellular hyphae grew longitudinallyalong the root, forming branches that entered the inner cor-tical cells leading to arbuscule formation (Fig. 1g,f ), first inthe innermost cortical cell layer (Fig. 1f ) and later in thepenultimate cortical cell layer (Fig. 1d). After arbuscules haddeveloped, vesicles are formed intercellularly between cellsof the three innermost cortical layers (Fig. 1d–f ). The intra-cellular passage occurs through living plant cells that preventcontact between fungi and plant cell cytoplasm by the synthesisof a perifungal membrane that is in continuum with the plantcell plasma membrane (Parniske, 2000).
Early colonization of LjsymRK mutant roots by Glomus intraradices
With LjsymRK mutant lines, even after 4–6 WOC, extensivegrowth of extraradical hyphae but no fungal growth beyondthe exodermis could be observed (Fig. 2a). Typically, hyphaeexhibited balloon-like cell expansions after formation ofappressoria (Fig. 2b,c) and such swollen hyphae were oftenoriented in longitudinal direction of the root, between twofiles of root epidermal cells, over a distance of several cells(Fig. 2b). Occasionally, swollen hyphae were located between
epidermal and exodermal cells (Fig. 2b,c,e,f ). In rare cases,swollen hyphae were found within epi- or exodermal cells(Fig. 2d–f ). Numbers of appressoria were c. 4–10 timeshigher in all LjsymRK mutant lines compared to the wild type(Fig. 3a), but 90–98% of colonization attempts were abortedwithin the epidermis/outer cortex. The number of colonizationattempts that successfully grew longitudinally 1–3 cells pastthe entry point (Fig. 2e–f ) increased substantially betweenthe third and the sixth WOC but these remained confined tothe epidermis/outer cortex and the formation of arbuscules orvesicles was never observed. We could not establish qualitativeor quantitative differences in colonization structures betweenthe tested LjsymRK alleles. This is consistent with all of thembeing probably loss-of-function alleles (Stracke et al., 2002).
LjSYM15 is required for the epidermal opening response
Ljsym15-1 is a weak allele that leads to delayed nodulation(Schauser et al., 1998), whereas plants carrying Ljsym15-2 donot nodulate (data not shown). This difference in allelestrength is also reflected by their mycorrhiza phenotype.On plants carrying the strong allele Ljsym15-2, no successfulcolonizations were observed after up to 8 WOC. Extensivehyphal growth was observed around the roots, typically on the
Fig. 2 Colonization of LjSYMRK mutant roots by Glomus intraradices after 4–6 wk of co-cultivation (WOC) with the chive nurse plant. (a–c, e–g) cleared roots with fungal structures stained with ink. (d) An 8-µm thick cross section of a root embedded in paraffin, stained with Feulgen/Alcian Blue. (a, b, e–g) are in top view, in (c) the root is seen from the side. (a) After 4 WOC, extensive hyphal growth was observed around the roots. (b–g) Structures found after 6 WOC. (b) Hyphae on the root surface, precisely at the border between two files of epidermal cells (arrow). Fungal hypha entered the roots between two epidermal cells (asterisks in b, c) and form swellings between epidermal and exodermal cells (arrowhead in (c)) or within epidermal cells (arrowheads in (b)). (d) An arrow points at a fungal hypha that has entered an exodermal cell but does not proceed further. (e, f) Two different focal levels of a fungal appressorium (arrowhead) formed between two epidermal cells (arrows) that has grown an extension between epidermal and exodermal cells which has expanded into an adjacent epidermal cell (asterisk). (g) A fungal hypha has entered between two epidermal cells (arrows), grown between epidermis and endodermis and entered and filled an epidermal cell forming an intracellular vesicle. EP, epidermis; EX, exodermis; IV, intracellular vesicle. Bars; (a) 100 µm (b–d) 50 µm (e–g) 12.5 µm.
© New Phytologist (2004) 163: 381–392 www.newphytologist.org
Research 385
root surface in a longitudinal direction, precisely at the borderbetween two files of epidermal cells (Fig. 4a). By contrast toLjsymRK mutants, longitudinal growth between epidermalfiles or slight swelling of hyphae was only exceptionally observed(Fig. 4a). This is an important difference, which suggestsSYM15 is required for the entry of fungal hyphae betweentwo epidermal cells, whereas SYMRK is not.
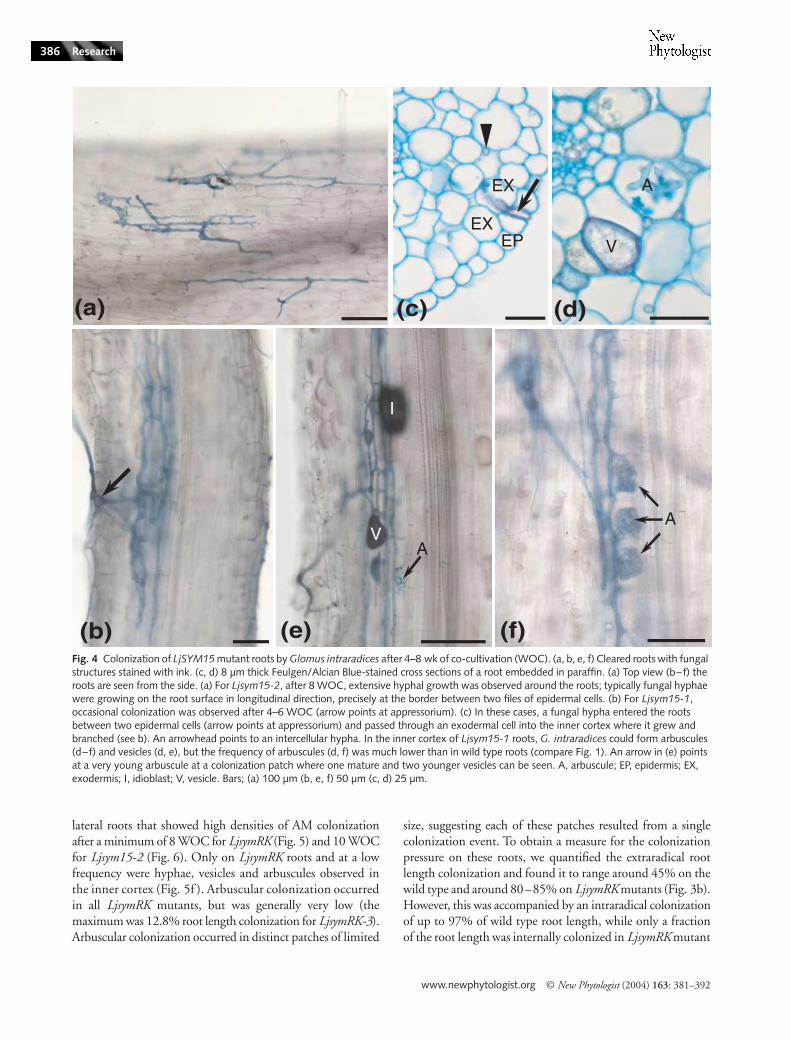
LjSYM15 is required for arbuscule formation
On roots of plants carrying the weaker allele Ljsym15-1,occasional colonization events were observed after 4–6 WOC(Fig. 4e–f ). In these cases, a fungal hypha entered the rootsbetween epidermal cells, penetrated an exodermal cell andpassed into the inner cortex (Fig. 4b–c) where it grew andbranched. Often, a slight swelling of the appressorium wasobserved, but never balloon-like structures like on LjsymRKroots (Fig. 2g). In the inner cortex, arbuscules and intercellularvesicles were formed (Fig. 4d–f ). However, in contrast withwild type roots, in which arbuscule formation preceded vesicleformation, vesicles were observed before or simultaneouslywith arbuscule formation in Ljsym15-1 roots (Fig. 4e) andextensive intraradical, extracellular hyphal growth beforearbuscule formation was always observed (Fig. 4b), indicatingan impairment in arbuscule formation in this mutant. In crosssections, most cells of the inner cortical layer of wild type rootscontained arbuscules (Fig. 1d), while in Ljsym15-1 sections,typically no or only one to three arbuscule-containing cellscould be seen (Fig. 4d). In longitudinal sections of wild typeroots, arbuscules were found in adjacent cells, forminglong files (Fig. 1e,f ). In Ljsym15-1 roots, only short files ofarbuscule-containing cells could be observed (Fig. 4f ). Thisevidence points to a role of LjSYM15 in arbuscule formation,which was further supported by observations on late colonizationevents on plants carrying the stronger Ljsym15-2 allele.
Late colonization of mutant roots
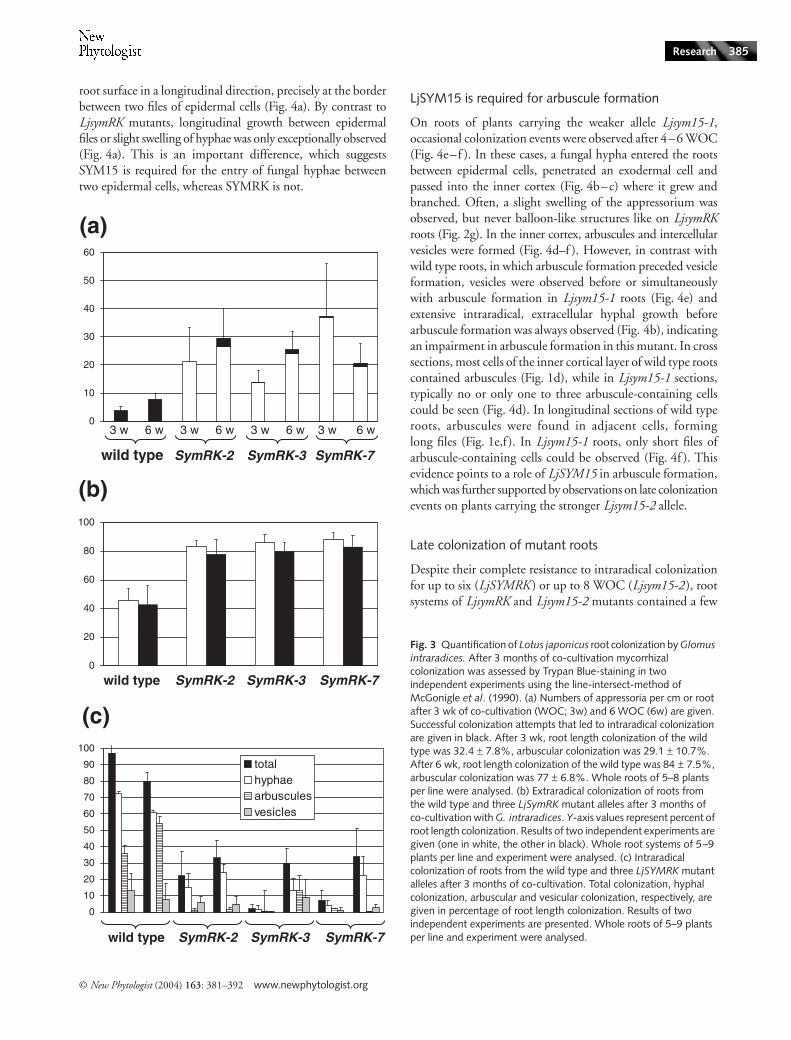
Despite their complete resistance to intraradical colonizationfor up to six (LjSYMRK ) or up to 8 WOC (Ljsym15-2 ), rootsystems of LjsymRK and Ljsym15-2 mutants contained a few
Fig. 3 Quantification of Lotus japonicus root colonization by Glomus intraradices. After 3 months of co-cultivation mycorrhizal colonization was assessed by Trypan Blue-staining in two independent experiments using the line-intersect-method of McGonigle et al. (1990). (a) Numbers of appressoria per cm or root after 3 wk of co-cultivation (WOC; 3w) and 6 WOC (6w) are given. Successful colonization attempts that led to intraradical colonization are given in black. After 3 wk, root length colonization of the wild type was 32.4 ± 7.8%, arbuscular colonization was 29.1 ± 10.7%. After 6 wk, root length colonization of the wild type was 84 ± 7.5%, arbuscular colonization was 77 ± 6.8%. Whole roots of 5–8 plants per line were analysed. (b) Extraradical colonization of roots from the wild type and three LjSymRK mutant alleles after 3 months of co-cultivation with G. intraradices. Y-axis values represent percent of root length colonization. Results of two independent experiments are given (one in white, the other in black). Whole root systems of 5–9 plants per line and experiment were analysed. (c) Intraradical colonization of roots from the wild type and three LjSYMRK mutant alleles after 3 months of co-cultivation. Total colonization, hyphal colonization, arbuscular and vesicular colonization, respectively, are given in percentage of root length colonization. Results of two independent experiments are presented. Whole roots of 5–9 plants per line and experiment were analysed.
www.newphytologist.org © New Phytologist (2004) 163: 381–392
Research386
lateral roots that showed high densities of AM colonizationafter a minimum of 8 WOC for LjsymRK (Fig. 5) and 10 WOCfor Ljsym15-2 (Fig. 6). Only on LjsymRK roots and at a lowfrequency were hyphae, vesicles and arbuscules observed inthe inner cortex (Fig. 5f ). Arbuscular colonization occurredin all LjsymRK mutants, but was generally very low (themaximum was 12.8% root length colonization for LjsymRK-3).Arbuscular colonization occurred in distinct patches of limited
size, suggesting each of these patches resulted from a singlecolonization event. To obtain a measure for the colonizationpressure on these roots, we quantified the extraradical rootlength colonization and found it to range around 45% on thewild type and around 80–85% on LjsymRK mutants (Fig. 3b).However, this was accompanied by an intraradical colonizationof up to 97% of wild type root length, while only a fractionof the root length was internally colonized in LjsymRK mutant
Fig. 4 Colonization of LjSYM15 mutant roots by Glomus intraradices after 4–8 wk of co-cultivation (WOC). (a, b, e, f) Cleared roots with fungal structures stained with ink. (c, d) 8 µm thick Feulgen/Alcian Blue-stained cross sections of a root embedded in paraffin. (a) Top view (b–f) the roots are seen from the side. (a) For Ljsym15-2, after 8 WOC, extensive hyphal growth was observed around the roots; typically fungal hyphae were growing on the root surface in longitudinal direction, precisely at the border between two files of epidermal cells. (b) For Ljsym15-1, occasional colonization was observed after 4–6 WOC (arrow points at appressorium). (c) In these cases, a fungal hypha entered the roots between two epidermal cells (arrow points at appressorium) and passed through an exodermal cell into the inner cortex where it grew and branched (see b). An arrowhead points to an intercellular hypha. In the inner cortex of Ljsym15-1 roots, G. intraradices could form arbuscules (d–f) and vesicles (d, e), but the frequency of arbuscules (d, f) was much lower than in wild type roots (compare Fig. 1). An arrow in (e) points at a very young arbuscule at a colonization patch where one mature and two younger vesicles can be seen. A, arbuscule; EP, epidermis; EX, exodermis; I, idioblast; V, vesicle. Bars; (a) 100 µm (b, e, f) 50 µm (c, d) 25 µm.
© New Phytologist (2004) 163: 381–392 www.newphytologist.org
Research 387
lines (Fig. 3c). By far the greatest proportion of overall rootlength colonization in the mutants was colonization by hyphaeand vesicles, not arbuscules. Hence, an effect of SYMRK onarbuscule initiation cannot be excluded.
Using light microscopy on paraffin-embedded plantmaterial stained with Feulgen/Alcian Blue, no difference could
be seen between arbuscules formed in growing wild type rootsand roots of LjsymRK mutant lines (Fig. 1d,f,g vs Fig. 5f,k).Like in the wild type, vesicles were formed near arbuscules inroots of LjsymRK mutant lines (Fig. 5f ). On roots of Ljsym15-2plants, however, no arbuscules were ever found. We concludethat SYMRK is not required for the formation of structurally
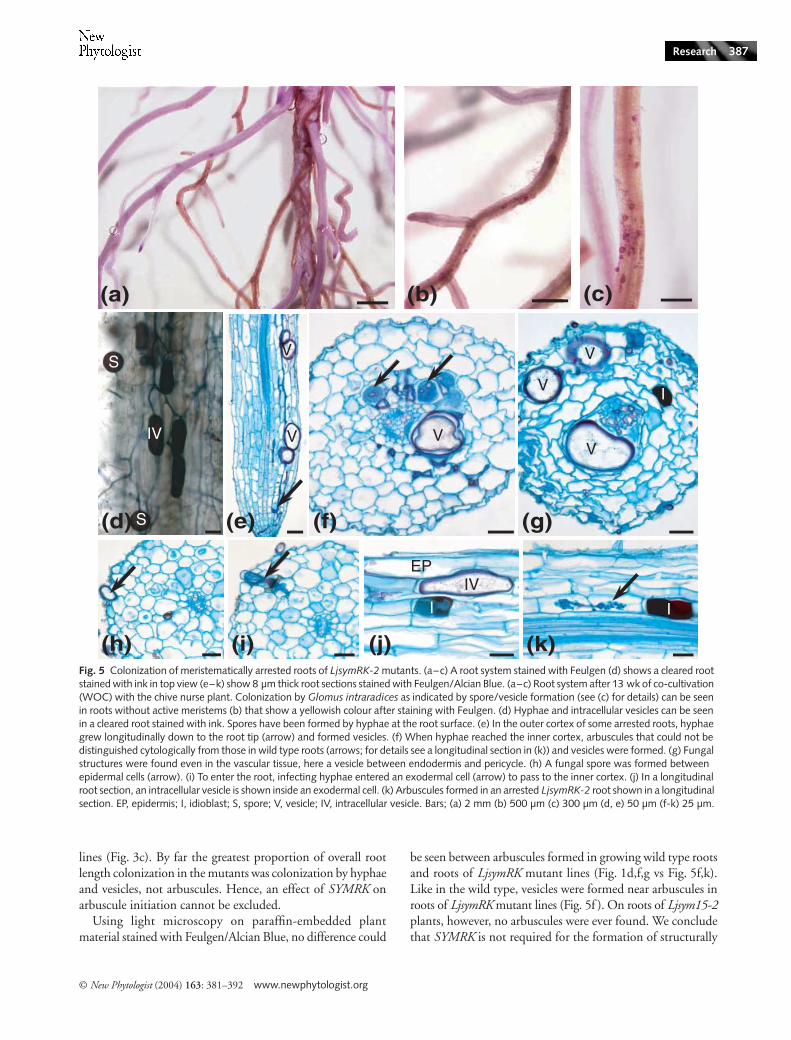
Fig. 5 Colonization of meristematically arrested roots of LjsymRK-2 mutants. (a–c) A root system stained with Feulgen (d) shows a cleared root stained with ink in top view (e–k) show 8 µm thick root sections stained with Feulgen/Alcian Blue. (a–c) Root system after 13 wk of co-cultivation (WOC) with the chive nurse plant. Colonization by Glomus intraradices as indicated by spore/vesicle formation (see (c) for details) can be seen in roots without active meristems (b) that show a yellowish colour after staining with Feulgen. (d) Hyphae and intracellular vesicles can be seen in a cleared root stained with ink. Spores have been formed by hyphae at the root surface. (e) In the outer cortex of some arrested roots, hyphae grew longitudinally down to the root tip (arrow) and formed vesicles. (f) When hyphae reached the inner cortex, arbuscules that could not be distinguished cytologically from those in wild type roots (arrows; for details see a longitudinal section in (k)) and vesicles were formed. (g) Fungal structures were found even in the vascular tissue, here a vesicle between endodermis and pericycle. (h) A fungal spore was formed between epidermal cells (arrow). (i) To enter the root, infecting hyphae entered an exodermal cell (arrow) to pass to the inner cortex. (j) In a longitudinal root section, an intracellular vesicle is shown inside an exodermal cell. (k) Arbuscules formed in an arrested LjsymRK-2 root shown in a longitudinal section. EP, epidermis; I, idioblast; S, spore; V, vesicle; IV, intracellular vesicle. Bars; (a) 2 mm (b) 500 µm (c) 300 µm (d, e) 50 µm (f-k) 25 µm.
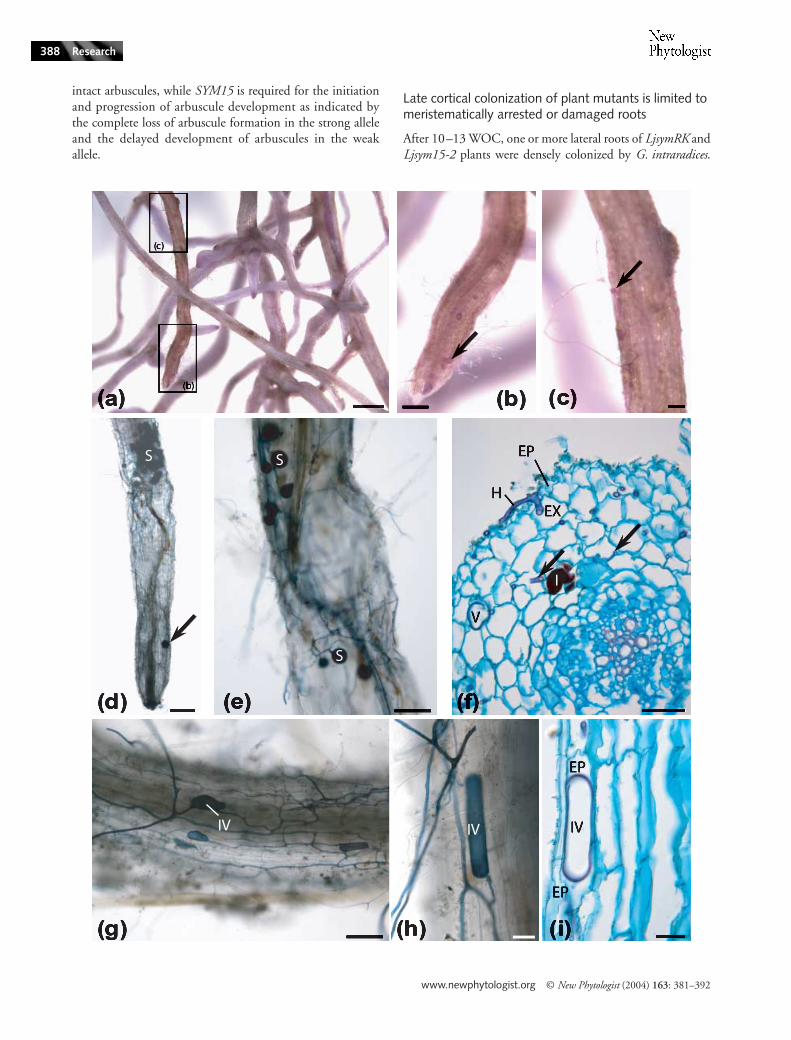
www.newphytologist.org © New Phytologist (2004) 163: 381–392
Research388
intact arbuscules, while SYM15 is required for the initiationand progression of arbuscule development as indicated bythe complete loss of arbuscule formation in the strong alleleand the delayed development of arbuscules in the weakallele.
Late cortical colonization of plant mutants is limited to meristematically arrested or damaged roots
After 10–13 WOC, one or more lateral roots of LjsymRK andLjsym15-2 plants were densely colonized by G. intraradices.

© New Phytologist (2004) 163: 381–392 www.newphytologist.org
Research 389
By using the Feulgen stain applied to entire root systems, wediscovered that late successful colonization patches wereconfined to meristematically arrested roots as evidenced byfull cell differentiation down to the root cap, or to damaged rootsthat had their tips torn off. Meristem arrest was confirmed byFeulgen/Alcian Blue staining of longitudinal sections (Figs 5eand 6d). Most lateral roots with arrested meristems, andall roots whose younger parts had been torn off, showed ayellowish colour after staining with Feulgen (Figs 5a–c and6a–c). We suspect that the yellow Feulgen stain is a functionof root ageing which occurs after meristem arrest. Aging wouldalso provide an explanation for the brittle nature of these rootsthat leads to frequent loss of the tip during uprooting. Onlyyellowish lateral roots, but not all of them, were colonized byG. intraradices. These observations suggest that meristem arrestand the physiological alteration of the root that leads to yellowFeulgen stain preceded late successful fungal colonizationevents. Many meristematically arrested LjsymRK and Ljsym15roots showed extensive hyphal and vesicular colonization(Figs 5e–f and 6b–e), also in the outer cortex (Figs 5e,g and6f; data not shown for Ljsym15-1) proceeding as far as into theroot tip (Figs 5e and 6d) and in LjsymRK roots occasionallyentering the vascular tissue (Fig. 5e,g).
Vesicles were often formed ectopically during late colonizationof mutant roots, e.g. in the outer cortex of LjsymRK and Ljsym15roots (Figs 5e,g and 6f; data not shown for Ljsym15-1) and pro-ceeding as far as into the root tip (Figs 5e and 6d). In LjsymRKroots fungi occasionally entered the vascular tissue (Fig, 5e,g).By contrast, in wild type roots, vesicles were only found betweenthe cells of the three innermost cortical cell layers (Fig. 1e,f ).
A mutant-specific feature was the occurrence of intracellularhyphal swellings and vesicles; arrested LjsymRK-2, -7 andLjsym15-2 roots harboured vesicles within epidermal or exo-dermal cells. We suspect that they developed in already deadplant cell wall ‘ghosts’. In sections, intracellular vesicles dis-played a similar cytoplasmic structure but seemed to possessthinner cell walls than extracellular vesicles (Figs 5d,j and6g–i). By contrast, in wild type roots, vesicles were alwaysintercellular and only found between the cells of the threeinnermost cortical cell layers (Fig. 1e–f ). Intracellular swellingsof hyphae were found exclusively in mutant roots, while extra-cellular swellings occurred at increased frequency comparedto the wild type (Fig. 2b,e–f ). Swollen hyphae were notnecessarily terminal organs, and if intracellular, never filled
the entire cell. In sections, their cytoplasmic structure resembledthat of hyphae, not of vesicles (data not shown).
Fungal entry into meristematically arrested lateral roots
Fungal hyphae entered LjsymRK roots between epidermalcells and reached the inner cortex of LjsymRK roots via passagethrough an exodermal cell (Fig. 5i). While the entry throughan exodermal cell superficially resembled the process of normalwild type colonization (Fig. 1a–c), fungal hyphae entered theapoplast again below the exodermis instead of proceedingintracellularly through the next cell layer. Furthermore, theexodermal cells used for passage always showed very strongstaining with Alcian Blue and collapsed cytoplasm, suggestingcell death (Fig. 5i), but it is unclear whether this was cause orconsequence of fungal penetration.
In Ljsym15-2 roots, entry of fungal hyphae was different fromall other interactions analysed. Entry of fungal hyphae wasfound mostly in parts of the root system where the cortex wasexposed (Fig. 6e). It is not clear whether this damage of epi- andexodermis occurred during root harvesting. However, the strik-ing correlation between entry point and absence of outer celllayers suggests that this damage occurred during co-cultivation.Very few cases were found where entry occurred intercellularlythrough what based on suberization of walls of adjacent cells,appeared to be dead regions of the outer cell layers (Fig. 6f; dataon suberization not shown). Hyphae proceeded from suchdamaged parts of the root, and grew longitudinally betweencell files into nondamaged parts of the root (Fig. 6b,d–f).
The effect of SYM15 on the intracellular passage throughthe epi-/exodermis could not be determined with certainty,since in plants carrying the strong allele, Ljsym15-2,fungal hyphae never entered an exodermal cell. The unper-turbed intracellular passage in roots carrying the weak allele,Ljsym15-1, suggests that SYM15 plays little or no role in thisstep.
Discussion
Different functions of SYMRK and SYM15 in root symbiosis
The common SYM genes are required for both nodulationand arbuscular mycorrhiza, and probably act in a pathway
Fig. 6 Colonization of meristematically arrested roots or dead roots of Lotus japonicus sym15–2 mutants. (a–c) Root systems stained with Feulgen (d, e, g, h) show cleared root systems stained with ink (f, i) show 8 µm thick root sections stained with Feulgen/Alcian Blue. (d, e, g, h) are in top view. (a–c) Ljsym15-2 root system after 13 wk of co-cultivation (WOC) with the chive nurse plant. Colonization by Glomus intraradices as indicated by vesicle/spore formation (b, c; arrows) can be seen. These roots showed a yellowish colour after staining with Feulgen. (d) In some infected roots, hyphae and spores (arrow) were found in the outer cortex down to the root tip. The root shows full differentiation down to the root cap. (e) Sometimes, colonization sites were found at points where root epidermis and outer cortex were destroyed. (f) Fungal hyphae have entered the root and reached the inner cortex (arrows). A vesicle has been formed in the outer cortex. (g) An infected root section shows extensive hyphal growth, as well as intracellular vesicles. (h, i) Intracellular vesicles were formed inside cells of the epidermis. A, arbuscule; EP, epidermis; EX, exodermis; H, hypha; S, spore; V, vesicle; IV, intracellular vesicle. Bars; (a) 1 mm (b, c) 250 µm (d, e, g) 100 µm (f) 50 µm (h, i) 25 µm.

www.newphytologist.org © New Phytologist (2004) 163: 381–392
Research390
leading from the perception of microbial signalling moleculesto the activation of symbiotic plant responses, includingsymbiosis-related gene activation and the induction of anintracellular accommodation programme ( Kistner & Parniske,2002). However, up to now, positioning of these genes relativeto each other was hampered by the lack of differentiatingphenotypic symptoms. In the interaction with rhizobia, theonly phenotypic marker that discriminates between theirfunction, is the calcium spiking response of root hair cellsto the application of Nod factors (Oldroyd et al., 2001a,b).Here we show that the steps involved in the development ofthe arbuscular mycorrhiza symbiosis are differentially impairedin mutants affected in the SYMRK or SYM15 genes.
Three steps for root colonization by AM fungi
Arbuscular mycorrhiza development involves cell type-specificresponses to fungal invasion. Because fungal colonizationcan at low but detectable frequencies bypass symbiotic blocksin outer cell layers, the effects of a symbiotic mutation on innercell layers can be studied. This potential to break through outerblocks makes fungal colonization a sensitive assay system tostudy the role of common SYM genes in cell types other thanthe epidermis. A synthesis between the results of Novero et al.(2002) and those presented here points to three successiveresponses of different cell layers that are required for fungalcolonization (Fig. 1a). First, the epidermis response allowsfungal penetration between two epidermal cells. In the vicinityof the fungus, the anticlinal cell walls of two adjacent epidermalcells separate from each other and a fungal hypha entersthrough this gap. The second and the third step both requirethe activation of an intracellular accommodation programmein different cell types and with different outcomes. The secondresponse is by epidermal (Wegel et al., 1998; Novero et al.,2002) or exodermal (this study) cells, and allows intracellularpassage through these layers. The third step, arbuscule formation,requires a more elaborate response of the host inner corticalcells to host a more complex fungal structure.
Infection of meristematically arrested roots
Ageing led to changes of LjsymRK and Ljsym15-2 roots thatmade them accessible for fungal hyphae as fungal colonizationonly occurred in meristematically arrested roots that stainedyellow with Feulgen. The responsible physiological changes seemto affect epidermal and exodermal cells. However, meristematicarrest did not alleviate the requirement of SYM15 for arbusculeformation by cortical cells. Older roots of nonmycorrhizalplants can be colonized by arbuscular-mycorrhizal fungiwith intercellular hyphae and inter or intracellular vesicles(reviewed by Brundrett, 1991; Giovannetti & Sbrana, 1998).These interactions lack appressoria and arbuscules and areconsidered as saprophytic growth of the fungus on the plantroots (Brundrett, 1991). The question arises whether the
infection of meristematically arrested roots represents a bonafide symbiotic interaction, or should be regarded as saprophyticfungal growth. On LjsymRK roots, arbuscules could be formed,indicating the late formation of a structurally intact symbiosis.For Ljsym15-2 roots where no arbuscules were formed, it wasnot possible to distinguish between late symbiosis and saprophyticgrowth.
The role of common SYM genes at different steps of AM development
The effect on LjSYMRK and LjSYM15 on the three steps inroot colonization by G. intraradices as found in this study issummarized in Fig. 1a.
Surface opening In the vicinity of the fungus, the anticlinalcell walls of two adjacent epidermal cells separate from eachother. This response is impaired in Ljsym15-2 mutants. Aconsequence of this is that the fungus never proceeds beyondthe cleft between epidermal cells, in contrast to symRK mutantswhere hyphae grow further and form balloon-like structureswhich is never observed in Ljsym15-2 mutants. We concludethat it is an active host response that requires the activity ofthe SYM15 but not of the SYMRK gene. The cytologicalanalysis of the infection process of Ljsym4 mutants by Noveroet al. (2002) showed that surface opening also occurs there,hence like SYMRK, SYM4 is not required for this process.
Intracellular passage through outer cell layers The next stepis the intracellular passage of fungal hypha through plant cellsinto the inner cortex. Here, two different ports of intracellularentry appear to be possible. Novero et al. (2002) and Wegelet al. (1998) presented evidence for fungal entry into epidermalcells, and subsequent progression of the hypha into theadjacent exodermal cell. In this study, we regularly observedfungal passage between two epidermal cells and entry into anexodermal cell. After intracellular passage through exodermisand the outermost cortical cell layer, fungal hyphae grewmostly intercellularly to the two inner cortical cell layerswhere arbuscule formation can take place.
Our observations of wild type roots suggest that a minimumof two cell layers have to be crossed intracellularly before releaseof hyphae and intercellular colonization of the cortex (see e.g.Fig. 1d). Both epidermis and exodermis seem to have thepotential to initialise the intracellular phase of hyphal growth,and both exodermis and the outermost cortical cell layer areable to support their further intracellular passage. Intracellularfungal accommodation by the outer cell layers was affectedin roots of LjsymRK (this study) and Ljsym4 plants (Bonfanteet al., 2000; Novero et al., 2002).
Hyphae were observed within epidermal and exodermalcells during failed colonization attempts of LjsymRK mutantroots, very similar to the situation observed in Ljsym4 rootsby Bonfante et al. (2000). For the passage through plant cell

© New Phytologist (2004) 163: 381–392 www.newphytologist.org
Research 391
walls fungal turgor pressure has been shown to be sufficient(Talbot, 2003). The swelling of fungal hyphae frequently observedin such cases of forced entry may be a result of changes infungal cell wall composition that are induced in course of theinteraction. It has been shown for Ljsym4-1, the cytoplasm ofboth interacting cells collapses, if the plant does not activelysupport the internalization (Bonfante et al., 2000).
Arbuscule formation During arbuscule formation, a sidebranch of an intercellular hypha enters a cell of one of thetwo innermost cortical layers and branches profusely. Inthe wild type, hyphae grow longitudinally along the rootand arbuscules form in every cell that is passed. This processwas disturbed in Ljsym15 mutants. Ljsym15-1 plants showeda delay and reduction in arbuscule formation (Fig. 4e,f ), andin Ljsym15-2 roots arbuscule formation was never observed.Novero et al. (2002) reported a change in the ratio of hyphal toarbuscular colonization of Ljsym4-1 roots and concludedthat SYM4 has a role in arbuscule initiation. SYM4 is thereforedistinct from both SYMRK and SYM15: like SYMRK it is notrequired for epidermal opening, but like SYM15 it has a rolein arbuscule formation.
Signalling during mycorrhizal colonization of plant roots
LjSYMRK, encoding an LRR receptor-like kinase (Strackeet al., 2002), seemed to affect only the intracellular passagethrough exodermis and outermost cortical cell layer. The resultspresented by Wegel et al. (1998), Bonfante et al. (2000) andNovero et al. (2002) indicate that LjSYM4 is probably essentialfor the same step. LjSYM15 is required for full surface openingand for arbuscule formation, both processes independent ofLjSYMRK. In congruence with these results, Ljsym4 (Harriset al., 2003) and by deduction from results obtained in Medicagotruncatula and pea (Endre et al., 2002), LjsymRK mutants areaffected in the calcium spiking response to rhizobial Nodfactors. LjSYMRK and LjSYM4 have therefore been placedat a similar hierarchical level in a genetic pathway ( Kistner &Parniske, 2002). LjSYM15, on the other hand, may be part ofa signal transduction pathway independent of LjSYMRK andLjSYM4.
The most parsimonious interpretations of the geneticdata are linear signalling pathways in plant-microbe symbioses(Stougaard, 2001). However, refined analysis of common SYMmutants showed that signal transduction networks are likelyto be involved (Kosuta et al., 2003). Our results clearly showthat it is crucial to consider the structural context whendrawing such models. Different plant cell types are involvedin the AM interaction, and these produce cell type-specificresponses. Our results suggest the existence of a signallingnetwork, of which individual components are required foran overlapping but noncongruent set of cell-type specificresponses. The detailed cytological analysis of the colonization
process in plant mutants carrying weak alleles, as well as theanalysis of late colonization of arrested roots, will continue toprove valuable in revealing the function of genes in symbioticsignalling in different cell types.
Acknowledgements
We would like to thank Anke Sirrenberg and Catherine Kistnerfor helpful discussions and Uwe Wedemeyer for taking careof the plants in Göttingen. This work was supported bythe German Research Council (Focus Program Mycorrhiza,SP1084). KD acknowledges additional support by RussianScience Support Foundation, the Russian Foundation forBasic Research and the Focus Program of the Russian Academyof Sciences. Research at The Sainsbury Laboratory is fundedby the Gatsby Charitable Foundation.
References
Bonfante P, Genre A, Faccio A, Martini I, Schauser L, Stougaard J, Webb J, Parniske M. 2000. The Lotus japonicus LjSym4 gene is required for the successful symbiotic infection of root epidermal cells. Molecular Plant–Microbe Interactions 13: 1109–1120.
Brundrett MC. 1991. Micorrhizas in natural ecosystems. Advances in Ecological Research 21: 171–313.
Duc G, Trouvelot A, Gianinazzi-Pearson V, Gianinazzi S. 1989. First report of non-mycorrhizal plant mutants Myc-negative obtained in pea Pisum sativum L. & faba bean Vicia faba L. Plant Science 60: 215–222.
Endre G, Kereszt A, Kevei Z, Mihacea S, Kalo P, Kiss GB. 2002. A receptor kinase gene regulating symbiotic nodule development. Nature 417: 962–966.
Giovannetti M, Sbrana C. 1998. Meeting a non-host: the behaviour of AM fungi. Mycorrhiza 8: 123–130.
Handberg K, Stougaard J. 1992. Lotus japonicus, an autogamous, diploid legume species for classical and molecular genetics. Plant Journal 2: 487–496.
Harris JM, Wais R, Long SR. 2003. Rhizobium-induced calcium spiking in Lotus japonicus. Molecular Plant–Microbe Interactions 16: 335–341.
Hoagland DR, Arnon DT. 1938. The water-culture method for growing plants without soil. In: California Agriculture Experiment Station Circular 347. Berkeley, USA: University of California.
Kistner C, Parniske M. 2002. Evolution of signal transduction in intercellular symbiosis. Trends in Plant Sciences 7: 511–518.
Kosuta S, Chabaud M, Lougnon G, Gough C, Denarie J, Barker DG, Becard G. 2003. A diffusible factor from arbuscular mycorrhizal fungi induces symbiosis-specific MtENOD11 expression in roots of Medicago truncatula. Plant Physiology 131: 952–962.
McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA. 1990. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytologist 115: 495–501.
Newman EI, Reddell P. 1987. The distribution of mycorrhizas among families of vascular plants. New Phytologist 106: 745–751.
Novero M, Faccio F, Genre A, Stougaard J, Webb KJ, Mulder L, Parniske M, Bonfante P. 2002. Dual requirement of the LjSym4 gene for mycorrhizal development in epidermal and cortical cells of Lotus japonicus roots. New Phytologist 154: 741–749.
Oldroyd GED, Engstrom EM, Long SR. 2001a. Ethylene inhibits the Nod factor signal transduction pathway of Medicago truncatula. Plant Cell 13: 1835–1849.

www.newphytologist.org © New Phytologist (2004) 163: 381–392
Research392
Oldroyd GED, Mitra RM, Wais RJ, Long SR. 2001b. Evidence for structurally specific negative feedback in the Nod factor signal transduction pathway. Plant Journal 28: 191–199.
Parniske M. 2000. Intracellular accommodation of microbes by plants: a common developmental program for symbiosis and disease? Current Opinion in Plant Biology 3: 320–328.
Sass JE. 1958. Botanical microtechnique. Iowa, USA: The Iowa State College Press.
Schauser L, Handberg K, Sandal N, Stiller J, Thykjaer T, Pajuelo E, Nielsen A, Stougaard J. 1998. Symbiotic mutants deficient in nodule establishment identified after T-DNA transformation of Lotus japonicus. Molecular and General Genetics 259: 414–423.
Senoo K, Solaiman MZ, Kawaguchi M, Imaizumi-Anraku H, Akao S, Tanaka A, Obata H. 2000. Isolation of two different phenotypes of mycorrhizal mutants in the model legume plant Lotus japonicus after EMS-treatment. Plant and Cell Physiology 41: 726–732.
Smith FA, Smith SE. 1997. Structural diversity in (vesicular)-arbuscular mycorrhizal symbioses. New Phytologist 137: 373–388.
Stracke S, Kistner C, Yoshida S, Mulder L, Sato S, Kaneko T, Tabata S, Sandal N, Stougaard J, Szczyglowski K, Parniske M. 2002. A plant receptor-like kinase required for both bacterial and fungal symbiosis. Nature 417: 959–962.
Stougaard J. 2001. Genetics and genomics of root symbiosis. Current Opinion in Plant Biology 4: 328–335.
Talbot NJ. 2003. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea. Annual Review of Microbiology 57: 177–202.
Vierheilig H, Coughlan AP, Wyss U, Piche Y. 1998. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Applied and Environmental Microbiology 64: 5004–5007.
Wegel E, Schauser L, Sandal N, Stougaard J, Parniske M. 1998. Mycorrhiza mutants of Lotus japonicus define genetically independent steps during symbiotic infection. Molecular Plant–Microbe Interactions 11: 933–936.
About New Phytologist
• New Phytologist is owned by a non-profit-making charitable trust dedicated to the promotion of plant science, facilitating projectsfrom symposia to open access for our Tansley reviews. Complete information is available at www.newphytologist.org
• Regular papers, Letters, Research reviews, Rapid reports and Methods papers are encouraged. We are committed to rapidprocessing, from online submission through to publication ‘as-ready’ via OnlineEarly – average first decisions are just 5–6 weeks.Essential colour costs are free, and we provide 25 offprints as well as a PDF (i.e. an electronic version) for each article.
• For online summaries and ToC alerts, go to the website and click on ‘Journal online’. You can take out a personal subscription tothe journal for a fraction of the institutional price. Rates start at £108 in Europe/$193 in the USA & Canada for the online edition(click on ‘Subscribe’ at the website)
• If you have any questions, do get in touch with Central Office ([email protected]; tel +44 1524 592918) or, for a localcontact in North America, the USA Office ([email protected]; tel 865 576 5261)
Related Documents











