Disruption of the gene encoding 3b-hydroxysterol D 14 -reductase (Tm7sf2) in mice does not impair cholesterol biosynthesis Anna M. Bennati 1 , Gianluca Schiavoni 1 , Sebastian Franken 2 , Danilo Piobbico 3 , Maria A. Della Fazia 3 , Donatella Caruso 4 , Emma De Fabiani 4 , Laura Benedetti 5 , Maria G. Cusella De Angelis 5 , Volkmar Gieselmann 2 , Giuseppe Servillo 3 , Tommaso Beccari 1 and Rita Roberti 1 1 Department of Internal Medicine, University of Perugia, Italy 2 Institut fu ¨ r Physiologische Chemie, Rheinische Friedrich-Wilhelms-Universita ¨t, Bonn, Germany 3 Department of Clinical and Experimental Medicine, University of Perugia, Italy 4 Department of Pharmacological Sciences, University of Milan, Italy 5 Department of Experimental Medicine, University of Pavia, Italy In cholesterol biosynthesis, lanosterol undergoes removal of the methyl group at C14, leading to the formation of C14–C15 unsaturated sterol intermedi- ates. The enzymatic activity responsible for the reduc- tion of the introduced double-bond, 3b-hydroxysterol D 14 -reductase (EC 1.3.1.70), is carried out by the endoplasmic reticulum (ER) protein delta14-sterol reductase (C14SR) encoded by the TM7SF2 gene Keywords 3beta-hydroxysterol delta14-reductase; cholesterol biosynthesis; gene expression; lamin B receptor; Tm7sf2 Correspondence R. Roberti, Department of Internal Medicine, Laboratory of Biochemistry, University of Perugia, Via del Giochetto, 06122 Perugia, Italy Fax: +39 075 585 7428 Tel: +39 075 585 7426 E-mail: [email protected] (Received 24 May 2008, accepted 11 August 2008) doi:10.1111/j.1742-4658.2008.06637.x Tm7sf2 gene encodes 3b-hydroxysterol D 14 -reductase (C14SR, DHCR14), an endoplasmic reticulum enzyme acting on D 14 -unsaturated sterol interme- diates during the conversion of lanosterol to cholesterol. The C-terminal domain of lamin B receptor, a protein of the inner nuclear membrane mainly involved in heterochromatin organization, also possesses sterol D 14 -reductase activity. The subcellular localization suggests a primary role of C14SR in cholesterol biosynthesis. To investigate the role of C14SR and lamin B receptor as 3b-hydroxysterol D 14 -reductases, Tm7sf2 knockout mice were generated and their biochemical characterization was performed. No Tm7sf2 mRNA was detected in the liver of knockout mice. Neither C14SR protein nor 3b-hydroxysterol D 14 -reductase activity were detectable in liver microsomes of Tm7sf2 () ⁄ )) mice, confirming the effectiveness of gene inactivation. C14SR protein and its enzymatic activity were about half of control levels in the liver of heterozygous mice. Normal cholesterol levels in liver membranes and in plasma indicated that, despite the lack of C14SR, Tm7sf2 () ⁄ )) mice are able to perform cholesterol biosynthesis. Lamin B receptor 3b-hydroxysterol D 14 -reductase activity determined in liver nuclei showed comparable values in wild-type and knockout mice. These results suggest that lamin B receptor, although residing in nuclear membranes, may contribute to cholesterol biosynthesis in Tm7sf2 () ⁄ )) mice. Affymetrix microarray analysis of gene expression revealed that several genes involved in cell-cycle progression are downregulated in the liver of Tm7sf2 () ⁄ )) mice, whereas genes involved in xenobiotic metabolism are upregulated. Abbreviations C14SR ⁄ DHCR14, 3b-hydroxysterol D 14 -reductase; C27D 8 ,5a-cholesta-8(9)-en-3b-ol; C27D 8,14 ,5a-cholesta-8(9),14-dien-3b-ol; C29D 8 , 4,4-dimethyl-5a-cholesta-8(9)-en-3b-ol; C29D 8,14 , 4,4-dimethyl-5a-cholesta-8(9),14-dien-3b-ol; ER, endoplasmic reticulum; HEM, Hydrops- Ectopic calcification-Moth-eaten skeletal dysplasia; LBR, lamin B receptor. 5034 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Disruption of the gene encoding 3b-hydroxysterolD14-reductase (Tm7sf2) in mice does not impaircholesterol biosynthesisAnna M. Bennati1, Gianluca Schiavoni1, Sebastian Franken2, Danilo Piobbico3, Maria A. DellaFazia3, Donatella Caruso4, Emma De Fabiani4, Laura Benedetti5, Maria G. Cusella De Angelis5,Volkmar Gieselmann2, Giuseppe Servillo3, Tommaso Beccari1 and Rita Roberti1
1 Department of Internal Medicine, University of Perugia, Italy
2 Institut fur Physiologische Chemie, Rheinische Friedrich-Wilhelms-Universitat, Bonn, Germany
3 Department of Clinical and Experimental Medicine, University of Perugia, Italy
4 Department of Pharmacological Sciences, University of Milan, Italy
5 Department of Experimental Medicine, University of Pavia, Italy
In cholesterol biosynthesis, lanosterol undergoes
removal of the methyl group at C14, leading to the
formation of C14–C15 unsaturated sterol intermedi-
ates. The enzymatic activity responsible for the reduc-
tion of the introduced double-bond, 3b-hydroxysterolD14-reductase (EC 1.3.1.70), is carried out by the
endoplasmic reticulum (ER) protein delta14-sterol
reductase (C14SR) encoded by the TM7SF2 gene
Keywords
3beta-hydroxysterol delta14-reductase;
cholesterol biosynthesis; gene expression;
lamin B receptor; Tm7sf2
Correspondence
R. Roberti, Department of Internal Medicine,
Laboratory of Biochemistry, University of
Perugia, Via del Giochetto, 06122 Perugia,
Italy
Fax: +39 075 585 7428
Tel: +39 075 585 7426
E-mail: [email protected]
(Received 24 May 2008, accepted 11
August 2008)
doi:10.1111/j.1742-4658.2008.06637.x
Tm7sf2 gene encodes 3b-hydroxysterol D14-reductase (C14SR, DHCR14),
an endoplasmic reticulum enzyme acting on D14-unsaturated sterol interme-
diates during the conversion of lanosterol to cholesterol. The C-terminal
domain of lamin B receptor, a protein of the inner nuclear membrane
mainly involved in heterochromatin organization, also possesses sterol
D14-reductase activity. The subcellular localization suggests a primary role
of C14SR in cholesterol biosynthesis. To investigate the role of C14SR and
lamin B receptor as 3b-hydroxysterol D14-reductases, Tm7sf2 knockout
mice were generated and their biochemical characterization was performed.
No Tm7sf2 mRNA was detected in the liver of knockout mice. Neither
C14SR protein nor 3b-hydroxysterol D14-reductase activity were detectable
in liver microsomes of Tm7sf2() ⁄ )) mice, confirming the effectiveness of
gene inactivation. C14SR protein and its enzymatic activity were about half
of control levels in the liver of heterozygous mice. Normal cholesterol
levels in liver membranes and in plasma indicated that, despite the lack of
C14SR, Tm7sf2() ⁄ )) mice are able to perform cholesterol biosynthesis.
Lamin B receptor 3b-hydroxysterol D14-reductase activity determined in
liver nuclei showed comparable values in wild-type and knockout mice.
These results suggest that lamin B receptor, although residing in nuclear
membranes, may contribute to cholesterol biosynthesis in Tm7sf2() ⁄ ))
mice. Affymetrix microarray analysis of gene expression revealed that
several genes involved in cell-cycle progression are downregulated in the
liver of Tm7sf2() ⁄ )) mice, whereas genes involved in xenobiotic metabolism
are upregulated.
Abbreviations
C14SR ⁄ DHCR14, 3b-hydroxysterol D14-reductase; C27D8, 5a-cholesta-8(9)-en-3b-ol; C27D8,14, 5a-cholesta-8(9),14-dien-3b-ol; C29D8,
4,4-dimethyl-5a-cholesta-8(9)-en-3b-ol; C29D8,14, 4,4-dimethyl-5a-cholesta-8(9),14-dien-3b-ol; ER, endoplasmic reticulum; HEM, Hydrops-
Ectopic calcification-Moth-eaten skeletal dysplasia; LBR, lamin B receptor.
5034 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

[1,2]. A second protein of the inner nuclear membrane,
the lamin B receptor (LBR), catalyzes the 3b-hydroxys-terol D14-reductase reaction, as demonstrated by its
ability to complement C14SR-deficient yeast strains
[3,4] and by enzymatic assay of the protein overexpres-
sed in transfected COS-1 cells [5]. Recently, the mouse
gene encoding 3b-hydroxysterol D14-reductase has been
termed Dhcr14 [6]; in this study the former gene name
Tm7sf2 will be used. Tm7sf2 is located on chromosome
19A.
The expression of cholesterol biosynthesis genes is
regulated by cell sterol levels through the action of the
transcription factor SREBP-2 [7,8]. In HepG2 hepa-
toma cells, sterol starvation results in induction of the
TM7SF2 gene, C14SR protein and 3b-hydroxysterolD14-reductase activity. In addition, human TM7SF2
promoter is regulated by SREBP-2 [5]. Therefore, the
adaptability of TM7SF2 gene to the needs of choles-
terol biosynthesis appears well established. On the con-
trary, LBR gene expression is not responsive to sterol
starvation conditions [5] and its importance in choles-
terol biosynthesis remains unravelled.
The lack of 3b-hydroxysterol D14-reductase caused
by mutations of the LBR gene was previously indi-
cated as responsible for Hydrops-Ectopic calcification-
Moth-eaten skeletal dysplasia (HEM or Greenberg
dysplasia) [9–11]. The severe phenotype of natural
Lbr() ⁄ )) mutants, ichthyosis mice, has been described,
but no information on cholesterol levels and ⁄or sterol
intermediate accumulation has been reported [12].
Therefore, inactivation of Tm7sf2 gene in mice would
provide insights into the role of both genes encoding
3b-hydroxysterol D14-reductase, Tm7sf2 and Lbr, in
cholesterol biosynthesis.
While Tm7sf2() ⁄ )) mice generated in our laboratory
were under characterization, a paper was published
describing mice defective for Lbr (Lbr() ⁄ )), ichthyosis
mice), defective for Dhcr14 ⁄Tm7sf2 (Dhcr14(D4-7 ⁄ D4-7))
or defective for Dhcr14 ⁄Tm7sf2 and heterozygous for
Lbr (Lbr(+ ⁄ )):Dhcr14(D4-7 ⁄ D4-7)) [6]. The paper states
that HEM dysplasia is a laminopathy not caused by
3b-hydroxysterol D14-reductase deficiency. Mutants
have distinct physical and biochemical phenotypes, but
no sterol abnormalities were detected in liver, whereas
marked elevations of D14-sterols were seen in brain of
Lbr(+ ⁄ )):Dhcr14(D4-7 ⁄ D4-7) mice.
Here, we describe the generation of Tm7sf2() ⁄ ))
mice and their biochemical characterization. Determi-
nation of Tm7sf2 and Lbr mRNA expression in differ-
ent mouse tissues, expression of C14SR and LBR
proteins in liver and a measure of their 3b-hydroxys-terol D14-reductase activity are reported. Despite the
lack of C14SR, Tm7sf2() ⁄ )) mice are apparently
healthy and have normal cholesterol levels in liver
membranes and in plasma, suggesting that LBR can
function as 3b-hydroxysterol D14-reductase in vivo.
Furthermore, microarray analysis of gene expression in
liver comparing wild-type and Tm7sf2() ⁄ )) mice has
been performed.
Results
Identification of mouse Tm7sf2 gene
Genomic clones of Tm7sf2 were isolated by screening
a mouse 129 ⁄SvJ genomic library, subcloned and
sequenced. Comparison of the mouse genomic and the
cDNA sequences revealed that the exon–intron organi-
zation of the mouse Tm7sf2 gene is highly similar to
the homologous human gene [1]. The gene spans
� 5 kb and consists of 10 exons and 9 introns. Table 1
shows the size of the exons and introns and the
sequence of the exon–intron junctions. All 5¢ donor
and 3¢ acceptor splice sites conformed to the consensus
Table 1. Exon–intron organization of mouse Tm7sf2 gene. Exon and intron length is reported in parentheses.
Exon Intron
Sequence of exon–intron junction
5¢ splice donor–3¢ splice acceptor
1 (143 bp) 1 (349 bp) GGGCCGTTGG gtaaatggga–––ctctttccag GCGTCGCGGC
2 (197 bp) 2 (100 bp) CTGCACGAAG gtgtgtgatc–––gtacccgcag GTGGCCGAAG
3 (55 bp) 3 (274 bp) CCTATTAATG gtgactgggg–––tgtggttcag GCTTCCAGGC
4 (195 bp) 4 (84 bp) GGAAACTCAG gtgagaaggg–––ttgttcccag GAAATTCCAT
5 (104 bp) 5 (2112 bp) CATTGGCTGG gtatgctgac–––acttctttag GTTTTCATTA
6 (120 bp) 6 (88 bp) CTGGTATGAG gtgagactgg–––gttcctgcag GAGTCTGTCC
7 (169 bp) 7 (214 bp) CTCCTTAAGG gtcagtagga–––cttccctcag TTATTGGTTA
8 (81 bp) 8 (80 bp) AGCGTGGCTG gtaagctggg–––gtatttctag GTCTTGAGAC
9 (123 bp) 9 (250 bp) TTGCCCTGTG gtgagtgggt–––ttccctccag GGCTATCCCA
10 (253 bp) CTATCCCATC–––
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5035

GT–AG rule. The transcription initiation site, deter-
mined by RACE, was located at )91 bp upstream the
ATG start codon. A polyadenylation signal
(AATAAA) is present 49 bp downstream of the stop
codon. The genomic sequence has been submitted to
GenBank under accession number EU672836.
Tm7sf2 and Lbr expression in mouse tissues
Tm7sf2 and Lbr relative mRNA expression was mea-
sured in adrenal, brain, heart, kidney, liver, lung,
ovary and testis of 8-week-old mice using qRT-PCR.
The highest Tm7sf2 mRNA abundance was found in
liver, followed by ovary, testis, kidney and brain
(Fig. 1A). Testis and lung showed the highest Lbr gene
expression, followed by heart, ovary, kidney and liver
(Fig. 1B).
Tm7sf2 versus Lbr expression was determined by
using Lbr as the internal calibrator for each tissue.
Table 2 shows comparable expression of the two genes
in ovary, kidney and adrenal gland. Compared with
Lbr, � 8- and 2.5-fold higher Tm7sf2 expression was
found in liver and brain, respectively. By contrast, Lbr
versus Tm7sf2 expression was � 5-, 12- and 16-fold
higher in testis, heart and lung, respectively.
Generation of Tm7sf2-null mice
Tm7sf2 gene was disrupted in HM1 mouse embryonic
stem cells using a targeting vector in which exon 5 was
interrupted by a neomycin resistance cassette (Fig. 2B).
The homologous recombination between the targeting
vector and the endogenous Tm7sf2 allele resulted in
insertion of the neomycin phosphotransferase gene into
exon 5. Of 96 ES cell clones surviving G418 selection,
four targeted clones were identified by PCR analysis
and by Southern blot of EcoRI-digested genomic
DNA and two of them were injected into blastocysts.
Only clone E-53 generated germline transmitting chi-
meric male founders, which were intercrossed with
C57 ⁄B6 females to generate outbred strains. Offspring
were genotyped by PCR, which produced the expected
230 and 388 bp fragments from wild-type and dis-
rupted allele, respectively (Fig. 2B,C). Genotyping was
confirmed by Southern blot of EcoRI-digested DNA.
Labelled 8.9 and 7.8 kb fragments were obtained from
wild-type and mutated allele, respectively (Fig. 2B,D).
Offspring from heterozygous intercrosses showed the
typical Mendelian distribution of wild-type, heterozy-
gous and homozygous.
Tm7sf2() ⁄ )) mice develop normally, appear healthy
and are fertile. Histopathological analysis of liver, kid-
ney, adrenal and brain did not reveal differences
between control and mutant mice (data not shown).
Followed over a 3-month period, Tm7sf2() ⁄ )) mice
grow at the same rate as littermate control mice.
Groups of control and Tm7sf2() ⁄ )) female weighed at
14 months of age were 27.7 ± 1.9 g (n = 9) and
30.9 ± 1.9 g (n = 7), respectively. No apparent age-
dependent problems were observed in females or males
over a 14-month period. These results confirm previ-
ously reported data [6].
0.0
0.2
0.4
0.6
0.8
1.0
1.2A
B
Tm
7sf2
rel
ativ
e m
RN
A e
xpre
ssio
n
Adrenal
Brain
Heart
Kidney
LiverLung
Ovary
Testis
Adrenal
Brain
Heart
Kidney
LiverLung
Ovary
Testis
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Lb
r re
lati
ve m
RN
A e
xpre
ssio
n
Fig. 1. Expression of Tm7sf2 and Lbr mRNA in mouse tissues.
RNA was extracted from pooled tissues of three 8-week-old male
mice (females were used for ovary). Relative mRNA expression
was determined by qRT-PCR using liver and testis as internal cali-
brators for Tm7sf2 (A) and Lbr (B) genes, respectively. Experiments
were performed in triplicate and repeated twice with different RNA
preparations. Reported data are mean ± SD.
Tm7sf2 knockout mice A. M. Bennati et al.
5036 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

To test whether the mutation abolishes the expres-
sion of C14SR, RNA was extracted from liver and the
cDNA was synthesized by RT-PCR using the forward
and reverse primers that amplify the entire cDNA.
Figure 3A shows that the cDNA is absent in
Tm7sf2() ⁄ )) mice, whereas the expected 1.3 kb frag-
ment is obtained in control mice. Although no quanti-
tative PCR was performed, the cDNA was about half
of the control in heterozygous mice. Gapdh was ampli-
fied in parallel as housekeeping gene.
Western blot analysis of microsomes prepared from
liver was performed using anti-(bovine C14SR) serum,
which cross-reacts with the mouse protein. Figure 3B
shows that C14SR protein is absent in Tm7sf2() ⁄ ))
mice, whereas the band intensity is about half of
control (0.55 ± 0.09) in heterozygous mice, thus con-
firming the results obtained with the cDNA.
To investigate whether disruption of the Tm7sf2
gene modifies Lbr mRNA expression, qRT-PCR was
performed in tissues of 8-week-old wild-type and
Tm7sf2() ⁄ )) mice, using the wild-type as internal cali-
brator for each tissue. No significant differences of Lbr
mRNA expression in Tm7sf2() ⁄ )) mice, compared
with that of wild-type mice, were found in the exam-
ined tissues (adrenal, brain, heart, kidney, liver, lung,
ovary and testis) (data not shown).
Western blot analysis of nuclear membranes from
liver confirmed that LBR is expressed at about the
same level of control in Tm7sf2() ⁄ )) mice (Fig. 3C).
A
B
C
D
Fig. 2. Structure and targeted disruption of
Tm7sf2 gene. (A) The conversion of 4,4-
dimethyl-5a-cholesta-8(9),14-dien-3b-ol
(C29D8,14) to 4,4-dimethyl-5a-cholesta-8(9)-
en-3b-ol (C29D8) is catalysed by C14SR and
LBR. (B) Tm7sf2 genomic structure, target-
ing vector, and disrupted allele. Exon (filled
box) and intron lengths are approximately to
scale. PCR primers a, b, and neo are indi-
cated by arrows. The 3¢-probe used in
Southern blot experiments spans exon 10.
The size of EcoRI-digested wild-type and
disrupted allele is reported. (C) PCR geno-
typing of heterozygous Tm7sf2 intercross.
Primers a and b (B) amplify a 230-bp frag-
ment from the wild-type allele. Primers a
and neo (B) amplify a 388-bp fragment from
the disrupted allele. (D) Southern blot analy-
sis of mouse tail DNA isolated from the
progeny of a mating between heterozygous
parents. DNAs were digested with EcoRI
and hybridized with the 3¢-probe indicated in
(B).
Table 2. Tm7sf2 and Lbr expression in mouse tissues. Total RNA
was extracted from tissues of 8-week-old mice and retrotranscribed
as reported in Experimental procedures. Tm7sf2 and Lbr mRNA
expression was measured by qRT-PCR using the specific primers
(see Experimental procedures) and Lbr as internal calibrator for
each tissue. Hprt was used as the reference gene for sample nor-
malization. Data are mean ± SD of two experiments performed in
triplicate.
Tissue Tm7sf2 ⁄ Lbr ratio
Adrenal 0.68 ± 0.06
Brain 2.40 ± 0.51
Heart 0.09 ± 0.03
Kidney 0.90 ± 0.09
Liver 7.95 ± 0.71
Lung 0.06 ± 0.01
Ovary 1.06 ± 0.13
Testis 0.19 ± 0.05
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5037

3b-Hydroxysterol D14-reductase activity
3b-Hydroxysterol D14-reductase activity in microsomes
or in intact nuclei prepared from liver of 6-week-old
mice was determined in vitro by incubating the enzyme
source in the presence of the C27D8,14 sterol substrate.
Activity was measured on the basis of C27D8 forma-
tion and C27D8,14 disappearance, evaluated as the peak
area ratio between the individual sterol and cholestane,
the internal standard.
No enzymatic activity was detected in liver micro-
somes of Tm7sf2() ⁄ )) mice, whereas reduced activity
was detected in heterozygous compared with wild-type
mice (Fig. 4). The data are in accordance with C14SR
expression detected by western blot. The enzymatic
activity measured in intact nuclei, which can be
referred as LBR activity, was comparable in
Tm7sf2() ⁄ )) and Tm7sf2(+ ⁄ +) mice (Fig. 4).
The contribution of C14SR and LBR to the 3b-hy-droxysterol D14-reductase reaction in liver from wild-
type mice can be evaluated. On the basis of the
amount of incubated proteins (see legend in Fig. 4) in
the experimental conditions used, C14SR-specific activ-
ity was approximately eightfold higher than that of
LBR. Because 4.4 and 1.5 mg proteinÆg)1 of fresh tis-
sue were recovered as microsomes and nuclei, respec-
tively, C14SR enzymatic activity is >20-fold higher
than LBR enzymatic activity. This result is in accor-
dance with the high Tm7sf2 mRNA expression in liver,
compared with Lbr (Table 2).
Sterol determinations
Cholesterol concentration in liver microsomal mem-
branes of 6-week-old mice was measured both by
GC-MS analysis and by densitometry analysis of mem-
brane lipids separated by TLC. Despite the lack of
C14SR activity in microsomes, normal cholesterol
levels were found in these membranes (Table 3). No
differences were found between male and female mice.
Cholesterol biosynthetic precursors, including
C29D8,14, were not detectable by GC-MS analysis
of microsomal sterols, indicating that C29D8,14 inter-
mediate was not accumulated in Tm7sf2() ⁄ )) mice.
kb +/+ +/– –/–
Gapdh
1.5
Tm7sf2 cDNAA B CkDa +/+ –/–
66
45
31
+/–
Anti-C14SR
Microsomes
66kDa +/+ –/–
Anti-LBR
Nuclear membranes
Fig. 3. C14SR and LBR expression in liver. (A) PCR analysis of Tm7sf2 mRNA in the liver of Tm7sf2() ⁄ )), heterozygous and wild-type mice.
The primers used (see Experimental procedures) amplify a 1285-bp fragment. (B) Western blot analysis of liver microsomes. The anti-(bovine
C14SR) serum recognises a protein band with an apparent molecular mass of � 40 kDa. Equal amounts of protein (30 lg) were loaded in
each lane and checked by Ponceau staining of poly(vinylidene difluoride) membranes after protein transfer (data not shown). (C) Western
blot analysis of liver nuclear membranes. The anti-(human LBR) serum recognizes a protein band with an apparent molecular mass of
� 66 kDa. Equal amounts of proteins (50 lg) were loaded in each lane and checked by Ponceau staining of poly(vinylidene difluoride) mem-
branes after protein transfer (data not shown). Experiments were repeated at least three times with different RNA (A) or membrane (B and
C) preparations. One representative experiment is reported.
0.0
0.5
1.0
1.5
2.0
2.5
Ste
rol/c
ho
lest
ane
(pea
k ar
ea r
atio
)
Nucleit0 +/+ +/– –/– +/+ –/–
Microsomes
Fig. 4. 3b-Hydroxysterol D14-reductase activity. Microsomes
(0.25 mg protein) and intact nuclei (0.5 mg protein) prepared from
liver of wild-type, heterozygous and Tm7sf2() ⁄ )) mice were
assayed for 3b-hydroxysterol D14-reductase activity by incubation
for 30 min with C27D8,14 in the conditions described in Experimen-
tal procedures. Enzymatic activity was evaluated on the basis of
the decrease of peak area ratio between m ⁄ z 426 and m ⁄ z 372
ions (C27D8,14 ⁄ 5a-cholestane, filled columns) and the increase of
peak area ratio between m ⁄ z 428 and m ⁄ z 372 ions (C27D8 ⁄ 5a-
cholestane, open columns) at the expected retention time. C27D8
was absent at zero incubation time both in the microsomes and in
nuclei. Data shown are mean ± SD (n = 3).
Tm7sf2 knockout mice A. M. Bennati et al.
5038 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

Plasma cholesterol was comparable between 6-week-
old Tm7sf2(+ ⁄ +) and Tm7sf2() ⁄ )) mice, both in males
and females (Table 3). Groups of 14-month-old female
mice were also analysed, and comparable plasma
cholesterol levels were seen in wild-type and knockout
mice (Table 3).
Affymetrix oligonucleotide array hybridization
and data analysis
The expression of 45 101 transcripts represented on
the Affymetrix murine GeneChipMOE430 2.0 was
quantified in liver from wild-type and Tm7sf2() ⁄ ))
mice. Duplicate hybridizations were performed for
each sample, using the wild-type condition as a control
to measure fold change of gene expression in knockout
mice. Expression values were calculated using the
Robust Multi-Array Analysis (RMA) plugin [13] of the
genespring 7.3 software (Agilent Technologies, Milan,
Italy). Using this analysis, 1119 transcripts with a
SD < 0.2 were selected in the comparison knock-
out ⁄wild-type mice. Volcano Plot analysis of this tran-
script list was performed to identify transcripts with a
defined minimal fold change and statistically significant
P-value for a t-test of differences between samples. By
selecting a fold change > 1.5 and a P-value < 0.01, 66
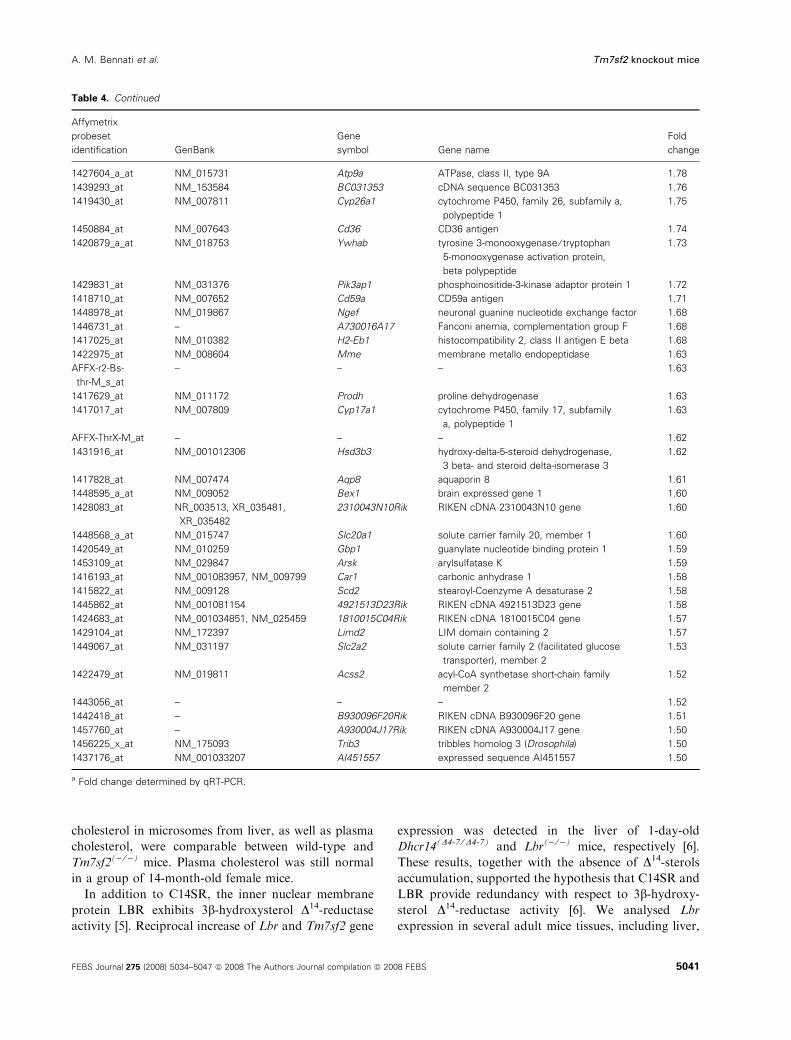
transcripts were identified as increased (Table 4) and 41
as decreased (Table 5) in livers of Tm7sf2() ⁄ )) mice.
Table 4 shows that several transcripts of oxidoreduc-
tases are increased, including members of cyt p450
families. Glutathione S-transferase, involved in xenobi-
otic metabolism, is also increased. Some genes involved
in cell proliferation and cell-cycle progression show
decreased transcripts (Table 5). qRT-PCR analysis was
applied to some of the genes that show the highest
up- or down expression comparing knockout to the
wild-type mice. Although with different fold changes,
the results obtained in the microarray experiment were
confirmed (Tables 4 and 5).
The complete panel of genes upstream and down-
stream Tm7sf2 in the cholesterol biosynthetic pathway
and Lbr were analysed carefully using less stringent
parameters (P-value for the t-test < 0.05). Neverthe-
less, no difference in their expression was found in the
liver of Tm7sf2() ⁄ )) mice.
Discussion
The discovery that inborn errors in cholesterol biosyn-
thesis are the cause of human malformation syndromes
characterized by severe developmental abnormalities
[14] highlights the role of cholesterol in mammalian
embryonic development. Indeed, cholesterol is a cova-
lent ligand of hedgehog morphogenic signalling pro-
teins [15]. Normal cholesterol biosynthesis is required
for cell membrane assembly and for the biosynthesis of
different biologically active sterol molecules. Further-
more, the cholesterol biosynthetic pathway furnishes a
variety of intermediate molecules involved in several
cell processes. In light of this, it is predictable that
mouse models bearing inactivation of any of the genes
of cholesterol biosynthesis are characterized by
impaired cell functions and developmental abnormali-
ties incompatible with life. Indeed, several mutant mice
lacking genes of the pre- or post-squalene segment of
the cholesterol biosynthetic pathway, that are lethal to
various degrees, have been described [16–23].
The mutant mouse model generated in our labora-
tory by inactivation of the Tm7sf2 gene lacks the ER
3b-hydroxysterol D14-reductase, C14SR. Nevertheless,
mice are viable and do not exhibit a pathological phe-
notype, as previously reported for Dhcr14(D4-7 ⁄ D4-7)
mice [6]. Using the antibody raised in our laboratory
against bovine liver C14SR, we were able to verify a
reduction and the complete absence of C14SR expres-
sion in liver microsomes of Tm7sf2(+ ⁄ )) and
Tm7sf2() ⁄ )) mice, respectively. In accordance with
these data, no 3b-hydroxysterol D14-reductase activity
is detectable in liver microsomes of Tm7sf2() ⁄ )) mice.
Despite the lack of 3b-hydroxysterol D14-reductase
activity of C14SR, normal cholesterol biosynthesis
occurs in Tm7sf2() ⁄ )) mice. Indeed, the level of
Table 3. Cholesterol in plasma and liver microsomal membranes. Total plasma cholesterol was measured using a commercial kit. For micro-
somal membrane cholesterol determination, samples were saponified and cholesterol was measured by GC-MS and by densitometric analy-
sis of lipids separated by TLC and stained as described in Experimental procedures.
Plasma cholesterol (mgÆdL)1)
Microsomal membrane cholesterol (nmolÆmg)1
protein)
Tm7sf2(+ ⁄ +) Tm7sf2() ⁄ )) Tm7sf2(+ ⁄ +) Tm7sf2() ⁄ ))
Male (6 weeks) 82.2 ± 10.3 (n = 8) 77.6 ± 14.5 (n = 12) 73.9 ± 5.8 (n = 6) 71.5 ± 6.5 (n = 6)
Female (6 weeks) 70.7 ± 12.9 (n = 15) 66.0 ± 11.8 (n = 10)
Female (14 months) 65.6 ± 21.1 (n = 9) 81.7 ± 20.2 (n = 7)
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5039

Table 4. Genes whose transcripts increase in liver of Tm7sf2() ⁄ )) mice. Total RNA from the pooled livers of three 6-week-old mice was pre-
pared for Affymetrix oligonucleotide hybridization as described in Experimental procedures. The relative mRNA expression of each gene in
livers of knockout mice compared with wild-type control is shown. Parameters for Volcano plot statistical analysis were fold change > 1.5
and P-value for the t-test < 0.01. Results from duplicate hybridizations are listed by fold change.
Affymetrix
probeset
identification GenBank
Gene
symbol Gene name
Fold
change
1438211_s_at NM_016974 Dbp D-site albumin promoter
binding protein
6.21 ⁄ 4.05a
1448793_a_at NM_011521 Sdc4 syndecan 4 3.43 ⁄ 4.80a
1420603_s_at NM_009016, NM_009017,
NM_009018, NM_020030,
NM_198193, XM_001006217
Raet1a, Raet1b,
Raet1c, Raet1d,
Raet1e
retinoic acid early transcript
1alpha, beta, gamma, delta, 1E
3.18
1449347_a_at NM_001081642, NM_021365,
NM_183094, XM_001471704,
XM_001471888, XM_001475552,
XM_001487778, XM_978371,
XR_035676, XR_035679
LOC100044048,
LOC100044049,
LOC100046087,
Xlr4a, Xlr4b,
Xlr4c, Xlr4e
X-linked lymphocyte-regulated
4A, 4B, 4C, 4E
3.02
1444438_at XM_356089, XM_904518 Cib3 calcium and integrin binding
family member 3
2.98 ⁄ 1.66a
1424853_s_at NM_010011, NM_201640,
XM_001471913
Cyp4a10, Cyp4a31,
LOC100044218
cytochrome P450, family 4,
subfamily a, polypeptides 10&31
2.68 ⁄ 1.83a
1416318_at NM_025429 Serpinb1a serine (or cysteine) peptidase
inhibitor, clade B, member 1a
2.65
1444810_at – – – 2.55
1421363_at NM_010003 Cyp2c39 cytochrome P450, family 2,
subfamily c, polypeptide 39
2.53 ⁄ 17.2a
1439560_x_at XM_488763, XM_489668 EG432995 predicted gene, EG432995 2.49
1419700_a_at NM_008935 Prom1 prominin 1 2.38
1453345_at NM_001081205 Npal1 NIPA-like domain containing 1 2.33
1421040_a_at NM_008182 Gsta2 glutathione S-transferase,
alpha 2 (Yc2)
2.32
1435459_at NM_018881 Fmo2 flavin containing monooxygenase 2 2.26
1435893_at NM_013703 Vldlr very low density lipoprotein
receptor
2.14
1423875_at NM_145505, XM_001473083 AI450540,
LOC100044843
expressed sequence AI450540 2.13
1431240_at NM_053165 Clec2h C-type lectin domain family 2,
member h
2.13
1421041_s_at NM_008181, NM_008182,
XM_001478795
Gsta1, Gsta2,
LOC100042295
glutathione S-transferase, alpha 1
(Ya); glutathione S-transferase,
alpha 2 (Yc2)
2.06
1438194_at – 2900019G14Rik RIKEN cDNA 2900019G14 gene 2.06
1456973_at – – – 2.05
1452501_at NM_010002 Cyp2c38 cytochrome P450, family 2,
subfamily c, polypeptide 38
1.98
1422903_at NM_010745 Ly86 lymphocyte antigen 86 1.93
1415932_x_at NM_015731 Atp9a ATPase, class II, type 9A 1.92
1418213_at NM_033373 Krt23 keratin 23 1.92
1444706_at – Nav2 neuron navigator 2 1.91
1423627_at NM_008706 Nqo1 NAD(P)H dehydrogenase, quinone 1 1.88
1450505_a_at NM_001034851, NM_025459 1810015C04Rik RIKEN cDNA 1810015C04 gene 1.85
1450648_s_at NM_207105 H2-Ab1 histocompatibility 2, class II
antigen A, beta1
1.84
1417900_a_at NM_013703 Vldlr very low density lipoprotein receptor 1.83
1455316_x_at XM_915804 ENSMUSG00000073624 predicted gene, ENSMUSG00000073624 1.81
1458585_at – – – 1.80
1447643_x_at NM_011415 Snai2 snail homolog 2 (Drosophila) 1.78
Tm7sf2 knockout mice A. M. Bennati et al.
5040 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS
cholesterol in microsomes from liver, as well as plasma
cholesterol, were comparable between wild-type and
Tm7sf2() ⁄ )) mice. Plasma cholesterol was still normal
in a group of 14-month-old female mice.
In addition to C14SR, the inner nuclear membrane
protein LBR exhibits 3b-hydroxysterol D14-reductase
activity [5]. Reciprocal increase of Lbr and Tm7sf2 gene
expression was detected in the liver of 1-day-old
Dhcr14(D4-7 ⁄ D4-7) and Lbr() ⁄ )) mice, respectively [6].
These results, together with the absence of D14-sterols
accumulation, supported the hypothesis that C14SR and
LBR provide redundancy with respect to 3b-hydroxy-sterol D14-reductase activity [6]. We analysed Lbr
expression in several adult mice tissues, including liver,
Table 4. Continued
Affymetrix
probeset
identification GenBank
Gene
symbol Gene name
Fold
change
1427604_a_at NM_015731 Atp9a ATPase, class II, type 9A 1.78
1439293_at NM_153584 BC031353 cDNA sequence BC031353 1.76
1419430_at NM_007811 Cyp26a1 cytochrome P450, family 26, subfamily a,
polypeptide 1
1.75
1450884_at NM_007643 Cd36 CD36 antigen 1.74
1420879_a_at NM_018753 Ywhab tyrosine 3-monooxygenase ⁄ tryptophan
5-monooxygenase activation protein,
beta polypeptide
1.73
1429831_at NM_031376 Pik3ap1 phosphoinositide-3-kinase adaptor protein 1 1.72
1418710_at NM_007652 Cd59a CD59a antigen 1.71
1448978_at NM_019867 Ngef neuronal guanine nucleotide exchange factor 1.68
1446731_at – A730016A17 Fanconi anemia, complementation group F 1.68
1417025_at NM_010382 H2-Eb1 histocompatibility 2, class II antigen E beta 1.68
1422975_at NM_008604 Mme membrane metallo endopeptidase 1.63
AFFX-r2-Bs-
thr-M_s_at
– – – 1.63
1417629_at NM_011172 Prodh proline dehydrogenase 1.63
1417017_at NM_007809 Cyp17a1 cytochrome P450, family 17, subfamily
a, polypeptide 1
1.63
AFFX-ThrX-M_at – – – 1.62
1431916_at NM_001012306 Hsd3b3 hydroxy-delta-5-steroid dehydrogenase,
3 beta- and steroid delta-isomerase 3
1.62
1417828_at NM_007474 Aqp8 aquaporin 8 1.61
1448595_a_at NM_009052 Bex1 brain expressed gene 1 1.60
1428083_at NR_003513, XR_035481,
XR_035482
2310043N10Rik RIKEN cDNA 2310043N10 gene 1.60
1448568_a_at NM_015747 Slc20a1 solute carrier family 20, member 1 1.60
1420549_at NM_010259 Gbp1 guanylate nucleotide binding protein 1 1.59
1453109_at NM_029847 Arsk arylsulfatase K 1.59
1416193_at NM_001083957, NM_009799 Car1 carbonic anhydrase 1 1.58
1415822_at NM_009128 Scd2 stearoyl-Coenzyme A desaturase 2 1.58
1445862_at NM_001081154 4921513D23Rik RIKEN cDNA 4921513D23 gene 1.58
1424683_at NM_001034851, NM_025459 1810015C04Rik RIKEN cDNA 1810015C04 gene 1.57
1429104_at NM_172397 Limd2 LIM domain containing 2 1.57
1449067_at NM_031197 Slc2a2 solute carrier family 2 (facilitated glucose
transporter), member 2
1.53
1422479_at NM_019811 Acss2 acyl-CoA synthetase short-chain family
member 2
1.52
1443056_at – – – 1.52
1442418_at – B930096F20Rik RIKEN cDNA B930096F20 gene 1.51
1457760_at – A930004J17Rik RIKEN cDNA A930004J17 gene 1.50
1456225_x_at NM_175093 Trib3 tribbles homolog 3 (Drosophila) 1.50
1437176_at NM_001033207 AI451557 expressed sequence AI451557 1.50
a Fold change determined by qRT-PCR.
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5041
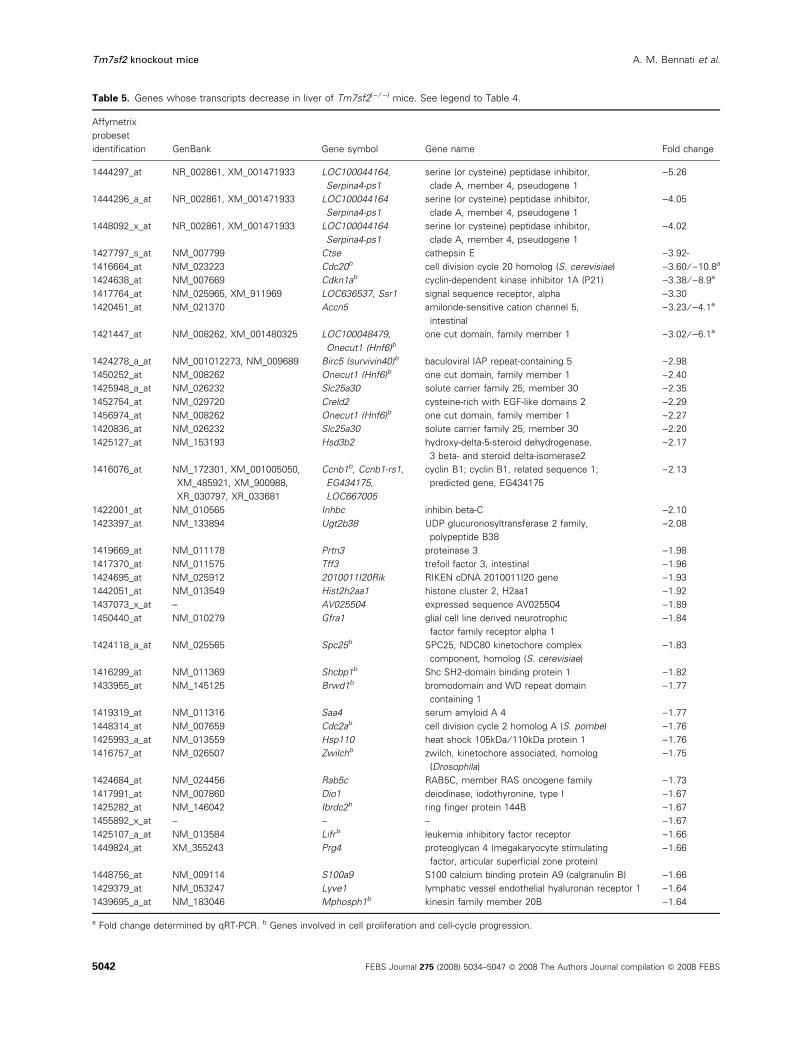
Table 5. Genes whose transcripts decrease in liver of Tm7sf2() ⁄ )) mice. See legend to Table 4.
Affymetrix
probeset
identification GenBank Gene symbol Gene name Fold change
1444297_at NR_002861, XM_001471933 LOC100044164,
Serpina4-ps1
serine (or cysteine) peptidase inhibitor,
clade A, member 4, pseudogene 1
)5.26
1444296_a_at NR_002861, XM_001471933 LOC100044164
Serpina4-ps1
serine (or cysteine) peptidase inhibitor,
clade A, member 4, pseudogene 1
)4.05
1448092_x_at NR_002861, XM_001471933 LOC100044164
Serpina4-ps1
serine (or cysteine) peptidase inhibitor,
clade A, member 4, pseudogene 1
)4.02
1427797_s_at NM_007799 Ctse cathepsin E )3.92-
1416664_at NM_023223 Cdc20b cell division cycle 20 homolog (S. cerevisiae) )3.60 ⁄ )10.8a
1424638_at NM_007669 Cdkn1ab cyclin-dependent kinase inhibitor 1A (P21) )3.38 ⁄ )8.9a
1417764_at NM_025965, XM_911969 LOC636537, Ssr1 signal sequence receptor, alpha )3.30
1420451_at NM_021370 Accn5 amiloride-sensitive cation channel 5,
intestinal
)3.23 ⁄ )4.1a
1421447_at NM_008262, XM_001480325 LOC100048479,
Onecut1 (Hnf6)bone cut domain, family member 1 )3.02 ⁄ )6.1a
1424278_a_at NM_001012273, NM_009689 Birc5 (survivin40)b baculoviral IAP repeat-containing 5 )2.98
1450252_at NM_008262 Onecut1 (Hnf6)b one cut domain, family member 1 )2.40
1425948_a_at NM_026232 Slc25a30 solute carrier family 25, member 30 )2.35
1452754_at NM_029720 Creld2 cysteine-rich with EGF-like domains 2 )2.29
1456974_at NM_008262 Onecut1 (Hnf6)b one cut domain, family member 1 )2.27
1420836_at NM_026232 Slc25a30 solute carrier family 25, member 30 )2.20
1425127_at NM_153193 Hsd3b2 hydroxy-delta-5-steroid dehydrogenase,
3 beta- and steroid delta-isomerase2
)2.17
1416076_at NM_172301, XM_001005050,
XM_485921, XM_900988,
XR_030797, XR_033681
Ccnb1b, Ccnb1-rs1,
EG434175,
LOC667005
cyclin B1; cyclin B1, related sequence 1;
predicted gene, EG434175
)2.13
1422001_at NM_010565 Inhbc inhibin beta-C )2.10
1423397_at NM_133894 Ugt2b38 UDP glucuronosyltransferase 2 family,
polypeptide B38
)2.08
1419669_at NM_011178 Prtn3 proteinase 3 )1.98
1417370_at NM_011575 Tff3 trefoil factor 3, intestinal )1.96
1424695_at NM_025912 2010011I20Rik RIKEN cDNA 2010011I20 gene )1.93
1442051_at NM_013549 Hist2h2aa1 histone cluster 2, H2aa1 )1.92
1437073_x_at – AV025504 expressed sequence AV025504 )1.89
1450440_at NM_010279 Gfra1 glial cell line derived neurotrophic
factor family receptor alpha 1
)1.84
1424118_a_at NM_025565 Spc25b SPC25, NDC80 kinetochore complex
component, homolog (S. cerevisiae)
)1.83
1416299_at NM_011369 Shcbp1b Shc SH2-domain binding protein 1 )1.82
1433955_at NM_145125 Brwd1b bromodomain and WD repeat domain
containing 1
)1.77
1419319_at NM_011316 Saa4 serum amyloid A 4 )1.77
1448314_at NM_007659 Cdc2ab cell division cycle 2 homolog A (S. pombe) )1.76
1425993_a_at NM_013559 Hsp110 heat shock 105kDa ⁄ 110kDa protein 1 )1.76
1416757_at NM_026507 Zwilchb zwilch, kinetochore associated, homolog
(Drosophila)
)1.75
1424684_at NM_024456 Rab5c RAB5C, member RAS oncogene family )1.73
1417991_at NM_007860 Dio1 deiodinase, iodothyronine, type I )1.67
1425282_at NM_146042 Ibrdc2b ring finger protein 144B )1.67
1455892_x_at – – – )1.67
1425107_a_at NM_013584 Lifrb leukemia inhibitory factor receptor )1.66
1449824_at XM_355243 Prg4 proteoglycan 4 (megakaryocyte stimulating
factor, articular superficial zone protein)
)1.66
1448756_at NM_009114 S100a9 S100 calcium binding protein A9 (calgranulin B) )1.66
1429379_at NM_053247 Lyve1 lymphatic vessel endothelial hyaluronan receptor 1 )1.64
1439695_a_at NM_183046 Mphosph1b kinesin family member 20B )1.64
a Fold change determined by qRT-PCR. b Genes involved in cell proliferation and cell-cycle progression.
Tm7sf2 knockout mice A. M. Bennati et al.
5042 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

and we did not find significant differences between wild-
type and Tm7sf2() ⁄ )) mice. In the liver, the result
obtained by qRT-PCR was further confirmed by the
Affymetrix oligonucleotide array hybridization experi-
ment. In addition, the expression of LBR protein in
nuclei from liver and its enzymatic activity were compa-
rable in wild-type and Tm7sf2() ⁄ )) mice. These results
indicate that, at least in adult Tm7sf2() ⁄ )) mice, LBR
can account for normal cholesterol biosynthesis without
increasing its expression. The discrepancy in Lbr gene
expression between 1-day-old Dhcr14(D4-7 ⁄ D4-7) and
adult Tm7sf2() ⁄ )) mice could be related to higher cho-
lesterol biosynthetic activity during development, com-
pared with adult mice. In vitro, C14SR exhibits higher
cholesterol biosynthetic capacity than LBR. Although it
cannot be excluded that this is due to the experimental
conditions used for enzymatic activity determination
(substrate and ⁄or cofactors), this result is in accordance
with higher Tm7sf2 gene expression in liver, compared
with Lbr. In vivo, the regulation of Tm7sf2 gene expres-
sion by cholesterol levels could represent a mechanism
to adapt 3b-hydroxysterol D14-reductase activity to
increased cholesterol needs.
The post-squalene segment of cholesterol biosynthe-
sis takes place in the ER. Among the enzymes involved
in this pathway, LBR is the unique residing in the
inner nuclear membranes. The N-terminal nucleoplas-
mic domain of LBR is involved in heterochromatin
organization [24], whereas the transmembrane domain
is responsible for 3b-hydroxysterol D14-reductase activ-
ity. Following its synthesis, LBR diffuses laterally from
the ER throughout the nuclear pore membranes and it
is retained in the inner nuclear membranes by its bind-
ing to the lamina [25]. It has been speculated whether
the ability of LBR to synthesize cholesterol in vivo is
restricted to its presence in the ER or in the nuclear
envelope [26]. Deficiency of LBR during granulopoiesis
results in hypolobulation of the mature neutrophil
nucleus, suggesting that LBR helps make the extra
nuclear membranes required during the nuclear lobula-
tion of granulopiesis [27]. Although LBR was not
detectable in liver microsomes, it cannot be excluded
that in vivo LBR supports cholesterol biosynthesis
when present in the ER. Alternatively, the mobility of
sterol intermediates in the lateral plane of the mem-
brane to reach enzymes differently compartmentalized
should occur.
The role of cholesterol and intermediates of its bio-
synthesis in cell growth and division is well known
[28,29]. The stringency of the requirement for choles-
terol during proliferation and cell-cycle progression
was investigated in promyelocytic HL-60 cells by com-
parison with other sterols of the biosynthetic pathway.
In the absence of exogenous cholesterol, accumulation
of intermediate sterols upstream 7-dehydrocholesterol,
including C29D8,14, resulted in the inhibition of cell
proliferation and cell cycle arrest in G2 ⁄M phase [30].
Affymetrix oligonucleotide array analysis showed that
several genes involved in cell proliferation and cell-
cycle progression have decreased expression in the liver
of Tm7sf2() ⁄ )) mice. Although no altered phenotype
has been observed in Tm7sf2() ⁄ )) mice so far, we
could speculate that an impaired response of liver cells
to proliferative stress is conceivable in these mice.
The evaluation of C14SR and LBR expression and
the determination of their enzymatic activity in the
liver of wild-type and Tm7sf2() ⁄ )) mice reinforce the
hypothesis that LBR and C14SR provide enzymatic
redundancy with respect to cholesterol synthesis [6].
Indeed, no sterol abnormalities were detected in
Tm7sf2() ⁄ )) mice. The significance of this enzymatic
redundancy is not clear. Even though it can be hypoth-
esized that multiple mechanisms have been developed
during evolution to ensure cholesterol biosynthesis, it
should be considered that the 3b-hydroxysterol D14-
reductase reaction is the only one in the post-squalene
pathway to be catalysed by two different enzymes. It is
worth noting that Drosophila LBR lacks sterol reduc-
tase activity, which could have been lost during evolu-
tion [31]. Different roles for Tm7sf2 and Lbr genes
during development or in tissues can be expected, rais-
ing the question of how 3b-hydroxysterol D14-reductase
activity can be switched between C14SR and LBR and
which regulatory mechanisms are involved.
Experimental procedures
Materials
Cholesterol, 5a-cholestane, and commercial antibodies were
purchased from Sigma (Milan, Italy). The polyclonal anti-
(bovine C14SR) serum was raised in our laboratory as
previously described [2]. The polyclonal anti-(human LBR)
serum was a kind gift of H. Hermann (German Cancer
Research Center, Heidelberg, Germany) [32]. Complete pro-
tease inhibitor cocktail tablets were from Roche Diagnos-
tics (Milan, Italy). RNAlater RNA Stabilization Reagent,
Qiazol Lysis Reagent, and RNeasy Mini Kit were from
Qiagen (Milan, Italy). QuickChange Site-Directed Muta-
genesis Kit, AffinityScript Multiple Temperature Reverse
Transcriptase, and Brilliant� SYBR� Green QPCR Master
Mix were purchased from Stratagene (La Jolla, CA, USA).
RiboLock RNase inhibitor, random hexamer primers, Taq
DNA polymerase, and restriction enzymes were from
Fermentas (St Leon-Rot, Germany). 5a-Cholesta-8,14-dien-3b-ol was synthesized as previously described [2].
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5043

Identification of mouse Tm7sf2 gene
Mouse Tm7sf2 mRNA was identified by NCBI database
comparison with the human mRNA (accession no.
AF096304). The mouse cDNA was synthesized by RT-PCR
using liver RNA as template and the following primers: for-
ward 5¢-ATGTCGACGATCATGACTTCTCGTGAGG-3¢and reverse 5¢-ATGTCGACTTCAACCTCTTAGGTG
GACC-3¢ (the SalI restriction site introduced for subclon-
ing in pBlueScript vector is given in italics). The sequenced
1285 bp cDNA (accession no. AF480070) encodes a puta-
tive protein 86% identical to human C14SR [1,2].
The cDNA was used to screen a 129 ⁄ SvJ mouse genomic
library in Lambda FIX II vector (Stratagene). Five library
clones that contained different chromosomal DNA frag-
ments were isolated and two contained the entire gene, as
demonstrated by Southern blot hybridization analysis.
Restriction endonuclease fragments from the Tm7sf2 geno-
mic clones were subcloned into pBlueScript vector and
sequenced.
Generation of Tm7sf2 knockout mice
A Tm7sf2 targeting vector was generated by subcloning
into pBluescript vector a 5380 bp KpnI ⁄XbaI fragment,
containing exons 1–5 of the gene (Fig. 2B). The neomycin
phosphotransferase gene (neo) was inserted in a SalI
restriction site generated by in vitro mutagenesis in
exon 5. The vector was linearized with NotI and electro-
porated into HM1 mouse embryonic stem cells (strain
129Sv ⁄ ola) [33]. After selection of G418-resistant embry-
onic stem cell colonies, homologous recombination was
established by PCR using a primer internal to the neo
gene (5¢-AGAACCTGCGTGCAATCCATCTTG-3¢) and a
3¢-primer external to the targeting construct (5¢-AAGCT
CTGCCTCCTGCATCAGC-3¢), which produced a 2560-
bp fragment. PCR cycling conditions were: 15 min
denaturation at 95 �C followed by 45 cycles of 30 s at
95 �C, 45 s at 64 �C, 3 min at 72 �C, and a final exten-
sion of 10 min at 72 �C. The positive clones were
subjected to Southern blotting after EcoRI genomic DNA
digestion, using a 450-bp probe located to the 3¢-end of
the gene and external to the targeting construct (Fig. 2B).
The probe detects an 8.9 kb EcoRI fragment in the wild-
type allele, and a 7.8 kb EcoRI fragment in the mutated
allele.
Targeted embryonic stem cell clone E-53 was injected
into C57 ⁄B6 blastocysts to generate chimeric mice. Chime-
ric males were mated with C57 ⁄B6 females and progenies
were analysed for germline transmission of the mutated
allele. Unless otherwise specified, the experiments were per-
formed with 129 ⁄Sv-C57 ⁄B6 hybrid descendants (F1) of
these animals. Backcross into C57 ⁄B6 was also carried out
for four generations to obtain mice with > 90% C57 ⁄B6genetic background. All experiments involving animals were
conducted according to protocols approved by the
Bioethics Committee of University of Perugia.
Mice were genotyped for the introduced Tm7sf2 mutated
gene by PCR analysis of tail genomic DNA using three spe-
cific primers (a, 5¢-AAGGCTTTGGTAGCTCCTGCCT-3¢;b, 5¢-TGAGGCCAGGTCTCAGCTCAC-3¢; neo, 5¢-GCT
ATCAGGACATAGCGTTGGC-3¢; see Fig. 2B). PCR
cycling conditions were: 2 min denaturation at 95 �C fol-
lowed by 35 cycles of 30 s at 95 �C, 45 s at 65 �C, 30 s at
72 �C, and a final extension of 5 min at 72 �C. Mouse
genotype was confirmed by Southern blotting analysis.
5¢ RACE
The transcription start site of the mouse Tm7sf2 gene was
determined by 5¢-RACE [34]. Mouse liver RNA was reverse
transcribed using the primer 5¢-AGGAGCTACCAAAGC
CTTCG-3¢ (nucleotides +468 to +449 from the ATG start
codon). A homopolymeric A-tail was then added to the
3¢-end using terminal transferase and dATP. The tailing
product was purified and amplified using the nested reverse
primer 5¢-CGACTCTTGTCCTTCAGTTCC-3¢ (nucleotides+284 to +264) and an oligo(dT) forward primer. The
product of amplification was amplified again using the
nested reverse primer 5¢-GTGCAGGCAGCAAATAGA
GC-3¢ (nucleotides +244 to +225) and an oligo(dT)
forward primer. The PCR product was analysed on 1.2%
agarose gel and sequenced in both strands.
Preparation of subcellular fractions from mouse
liver
Subcellular fractions were obtained from liver of 6-week-old
mice. Microsomes were prepared as previously described
[35,36] in the presence of complete protease inhibitor cock-
tail (Roche) and 0.03 mm phenylmethylsulfonyl fluoride.
Nuclei were isolated by the method of Kaufmann et al.
[37] and resuspended in 50 mm Tris ⁄HCl (pH 7.4), 0.25 m
sucrose, 5 mm MgSO4, containing 50% glycerol to avoid
freezing. For western blot analysis, the suspensions were
sonicated and the nuclear membrane fractions were recov-
ered by centrifuging at 100 000 g.
Preparations of microsomes and nuclei were not
cross-contaminated, as verified by western blotting with the
anti-(bovine C14SR) and anti-(human LBR) sera. Protein
concentration was determined by the method of Lowry [38].
Western blotting
Liver microsomal or nuclear membrane proteins were anal-
ysed by western blotting using polyclonal rabbit anti-
(bovine C14SR) serum or guinea-pig anti-(human LBR)
serum as previously described [5]. Equivalent protein load-
ing was checked by Ponceau staining of poly(vinylidene
Tm7sf2 knockout mice A. M. Bennati et al.
5044 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

difluoride) membranes after protein transfer. Labelled pro-
teins were detected by the enhanced chemiluminescence
assay, images were acquired using the VersaDoc Imaging
System, and signals were quantified using quantity one
software (Bio-Rad, Milan, Italy).
3b-Hydroxysterol D14-reductase activity
Determination of 3b-hydroxysterol D14-reductase activity
was performed by incubating the enzyme source for 30 min
at 37 �C with 5a-cholesta-8,14-dien-3b-ol (C27D8,14) [39]. To
measure the C27D8,14 substrate and the 5a-cholesta-8-en-3b-ol product (C27D8), sterols were purified and analysed by
GC-MS using 5a-cholestane internal standard as previously
described [2]. 3b-Hydroxysterol D14-reductase activity was
evaluated on the basis of peak area ratios between m ⁄ z 426
and m ⁄ z 372 ions (C27D8,14 ⁄ 5a-cholestane) or m ⁄ z 428 and
m ⁄ z 372 ions (C27D8 ⁄ 5a-cholestane) at the expected reten-
tion time. GC-MS analysis of sterols did not reveal reaction
products other than C27D8. In all conditions, C27D8 sterol
was undetectable at zero incubation time.
Sterol determinations
Sterols in liver microsomal membrane preparations were
evaluated by GC-MS. Lipids were saponified with methan-
olic KOH and sterols were extracted and analysed as
described previously [2]. Cholesterol was determined as
peak area ratio between m ⁄ z 368 and m ⁄ z 372 ions (choles-
terol ⁄ 5a-cholestane) using a calibration curve. Alterna-
tively, lipids were extracted by the method of Folch et al.
[40] and saponified. Cholesterol was separated by thin-layer
chromatography (n-hexane ⁄ diethyl ether ⁄ acetic acid,
70 : 30 : 1, v ⁄ v ⁄ v), visualized with Cu-acetate reagent [41],
and quantified as described for western blotting. Purified
cholesterol standard was run on the same plate as the sam-
ples to construct calibration curves.
Plasma cholesterol was determined enzymatically with a
commercial kit (Cholesterol Liquid, Sentinel, Milan, Italy).
RNA extraction and qRT-PCR analysis
Pooled tissues from three 8-week-old male mice (females
were used for ovary) in RNAlater RNA Stabilization
Reagent (Qiagen) were homogenized using Qiazol Lysis
Reagent (Qiagen). Total RNA was extracted according to
manufacture’s instructions and then subjected to clean up
on mini columns (RNeasy Mini Kit, Qiagen). RNA was
reverse transcribed using AffinityScript Multiple Tempera-
ture Reverse Transcriptase and random hexamer primers.
qRT-PCR amplifications were performed using Mx3000P�Real-Time PCR System with Brilliant� SYBR� Green
QPCR Master Mix (Stratagene) and ROX as reference dye.
Tm7sf2 specific primers were: forward, 5¢-GCCTCGGTTC
CTTTGACTTC-3¢; reverse, 5¢-CCATTGACCAGCCACAT
AGC-3¢. Lbr specific primers were: forward, 5¢-GTGCTCC
TGAGTGCTTAC-3¢; reverse, 5¢-GCCAATGAAGAAGT
CGTAC-3¢. Housekeeping control was mouse Hprt [42].
Experiments were performed in triplicate and repeated
twice with different RNA preparations. Results were
analysed using mx3000p�system software (Stratagene).
The specificity of the amplified products was assessed by
melting curve.
Affymetrix oligonucleotide array hybridization
and data analysis
Gene expression analysis was performed in liver of
6-week-old wild-type and knockout male mice after the
fourth backcross generation. Total RNA of pooled liver
tissue from three animals was extracted as described
above.
Probe synthesis, hybridization, and data analysis were
performed at the Affymetrix Microarray Unit (Campus
IFOM IEO, Milan, Italy). Biotin-labelled cRNA targets
were synthesized starting from 3 lg of total RNA. Double
stranded cDNA synthesis was performed with One-Cycle
cDNA Synthesis Kit, and biotin-labelled antisense RNA
was transcribed in vitro using IVT Labelling Kit. All steps
of the labelling protocol were performed with One-Cycle
Eukaryotic Target Labelling Assay as suggested by Affyme-
trix (http://www.affymetrix.com/support/technical/manual/
expression_manual.affx). Hybridization parameters were set
according to Affymetrix. Two copies of the complete gene-
chipmoe430 2.0 were hybridized with each biotin-labelled
target. Images were scanned using an Affymetrix GeneChip
Scanner3000 7G, using default parameters. The resulting
images were analysed using genechip operating software
v1.4 (GCOS1.4).
Some differentially expressed genes were analysed by
qRT-PCR as described above using the following primers.
Dbp: forward, 5¢-CTCGCCCTGTCAAGCATTCC-3¢;reverse, 5¢-TGATTGGTTGAGGCTTCAGTTCC-3¢. Sdc4:
forward, 5¢-GCGGCTCGGATGACTTTG-3¢; reverse, 5¢-AAGGGCTCAATCACTTCAGG-3¢. Cib3: forward, 5¢-ATGACTTCAACAATGACAACTAC-3¢; reverse, 5¢-ATC
CAGCACCTTCTCACAG-3¢. Cyp4a10 ⁄ 31: forward, 5¢-GC
CTCTGTGCTCGGTCTG-3¢; reverse 5¢-AGCCTTGAGTA
GCCATTGCC-3¢. Cyp2c39: forward 5¢-TGCTCTCCTAC
TCCTGATGAAG-3¢; reverse, 5¢-GGGCATGTGGTTCCT
GTCC-3¢. Cdc20: forward, 5¢-GCAACAGGAGGAGGA
ACCAG-3¢; reverse, 5¢-CATCCACAGCACTCAGACAG
G-3¢. Cdkn1a: forward, 5¢-AAAGTGTGCCGTTGTCTC-3¢;reverse, 5¢-AAAGTTCCACCGTTCTCG-3¢. Accn5: for-
ward, 5¢-GGTGACCATCCGCCAACTG-3¢; reverse, 5¢-CCGTAAGTGCTGTAGGTAATGAAG-3¢. Onecut1: forward,
5¢-CCTCTATGAATAACCTCTATACC-3¢; reverse, 5¢-TGCTGGGAGTTGTGAATG-3¢.
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5045

Acknowledgements
The financial support of Telethon–Italy (Grant no.
GGP030102) is gratefully acknowledged. We would
like to thank Aurelio Toia for assistance in GC-MS
analysis of sterols.
References
1 Holmer L, Pezhman A & Worman HJ (1998) The
human lamin B receptor ⁄ sterol reductase multigene
family. Genomics 54, 469–476.
2 Roberti R, Bennati AM, Galli G, Caruso D, Maras B,
Aisa C, Beccari T, Della Fazia MA & Servillo G (2002)
Cloning and expression of sterol delta14-reductase from
bovine liver. Eur J Biochem 269, 283–290.
3 Silve S, Dupuy P-H, Ferrara P & Loison G (1998)
Human lamin B receptor exhibits sterol C14-reductase
activity in Saccharomyces cerevisiae. Biochim Biophys
Acta 1392, 233–244.
4 Prakash A, Sengupta S, Aparna K & Kasbekar DP
(1999) The erg-3 (sterol D14,15-reductase) gene of
Neurospora crassa: generation of null mutants by
repeat-induced point mutation and complementation by
proteins chimeric for human lamin B receptor
sequences. Microbiology 145, 1443–1451.
5 Bennati AM, Castelli M, Della Fazia MA, Beccari T,
Caruso D, Servillo G & Roberti R (2006) Sterol depen-
dent regulation of human TM7SF2 gene expression:
role of the encoded 3b-hydroxysterol D14-reductase in
human cholesterol biosynthesis. Biochim Biophys Acta
1761, 677–685.
6 Wassif CA, Brownson KE, Sterner AL, Forlino A, Zer-
fas PM, Wilson WK, Starost MF & Porter FD (2007)
HEM dysplasia and ichthyosis are likely laminopathies
and not due to 3b-hydroxysterol D14-reductase defi-
ciency. Hum Mol Genet 16, 1176–1187.
7 Horton JD, Shah NA, Warrington JA, Anderson NN,
Park SW, Brown MS & Goldstein JL (2003) Com-
bined analysis of oligonucleotide microarray data from
transgenic and knockout mice identifies direct SREBP
target genes. Proc Natl Acad Sci USA 100, 12027–
12032.
8 Maxwell KN, Soccio RE, Duncan EM, Sehayek E &
Breslow JL (2003) Novel putative SREBP and LXR tar-
get genes identified by microarray analysis in liver of
cholesterol-fed mice. J Lipid Res 44, 2109–2119.
9 Greenberg CR, Rimoin DL, Gruber HE, DeSa DJ,
Reed M & Lachman RS (1988) A new autosomal reces-
sive lethal chondrodystrophy with congenital hydrops.
Am J Med Genet 29, 623–632.
10 Kelley RI & Herman GE (2001) Inborn errors of sterol
biosynthesis. Annu Rev Genomics Hum Genet 2, 299–
341.
11 Waterham HR, Koster J, Mooyer P, van Noort G, Kel-
ley RI, Wilcox WR, Wanders RJA, Hennekam RCM &
Oosterwijk JC (2003) Autosomal recessive HEM ⁄Greenberg skeletal dysplasia is caused by 3b-hydroxys-terol D14-reductase deficiency due to mutations in the
lamin B receptor gene. Am J Hum Genet 72, 1013–1017.
12 Schultz LD, Lyons BL, Burzenski LM, Gott B, Samuels
R, Schweitzer PA, Dreger C, Herrmann H, Kalscheuer
V, Olins AL et al. (2003) Mutations at the mouse ich-
thyosis locus are within the lamin B receptor gene: a
single gene model for human Pelger-Huet anomaly.
Hum Mol Genet 12, 61–69.
13 Irizarry RA, Bolstad BM, Collin F, Cope LM, Hobbs
B & Speed TP (2003) Summaries of Affymetrix Gene-
Chip probe level data. Nucleic Acids Res 31, e15.
14 Porter FD (2002) Malformation syndromes due to
inborn errors of cholesterol synthesis. J Clin Invest 110,
715–724.
15 Porter JA, Young KE & Beachy PA (1996) Cholesterol
modification of hedgehog signaling proteins in animal
development. Science 274, 255–259.
16 Tozawa R, Ishibashi S, Osuga J, Yagyu H, Oka T,
Chen Z, Ohashi K, Perrey S, Shionoiri F, Yahagi N
et al. (1999) Embryonic lethality and defective neural
tube closure in mice lacking squalene synthase. J Biol
Chem 274, 30843–30848.
17 Liu XY, Dangel AW, Kelley RI, Zhao W, Denny P,
Botcherby M, Cattanach B, Peters J, Hunsicker PR,
Mallon AM et al. (1999) The gene mutated in bare
patches and striated mice encodes a novel 3-beta-
hydroxysteroid dehydrogenase. Nat Genet 22, 182–
187.
18 Derry JM, Gormally E, Means GD, Zhao W, Meindl
A, Kelley RI, Boyd Y & Herman GE (1999) Mutations
in a D8-D7 sterol isomerase in the tattered mouse and
X-linked chondrodysplasys punctata. Nat Genet 22,
286–290.
19 Wassif CA, Zhu P, Kratz L, Krakowiak PA, Battaile
KP, Weight FF, Grinberg A, Steiner RD, Nwokoro
NA, Kelley RI et al. (2001) Biochemical, phenotypic
and neurophysiological characterization of a genetic
mouse model of RSH ⁄ Smith–Lemli–Opitz syndrome.
Hum Mol Genet 10, 555–564.
20 Fitzky BU, Moebius FF, Asaoka H, Waage-Baudet H,
Xu L, Xu G, Maeda N, Kluckman K, Hiller S, Yu H
et al. (2001) 7-Dehydrocholesterol-dependent proteolysis
of HMG-CoA reductase suppresses sterol biosynthesis
in a mouse model of Smith-Lemli-Opitz ⁄RSH syn-
drome. J Clin Invest 108, 905–915.
21 Ohashi K, Osuga J, Tozawa R, Kitamine T, Yagyu H,
Sekiya M, Tomita S, Okazaki H, Tamura Y, Yahagi N
et al. (2003) Early embryonic lethality caused by tar-
geted disruption of the HMG-CoA reductase gene.
J Biol Chem 278, 42936–42941.
Tm7sf2 knockout mice A. M. Bennati et al.
5046 FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS

22 Krakowiak PA, Wassif CA, Kratz L, Cozma D, Kavar-
ova M, Harris G, Grinberg A, Yang Y, Hunter AGW,
Tsokos M et al. (2003) Lathosterolosis, an inborn error
of human and murine cholesterol synthesis due to
lathosterol 5-desaturase deficiency. Hum Mol Genet
12, 1631–1641.
23 Mirza R, Hayasaka S, Takagishi Y, Cambe F, Ohmori
S, Maki K, Yamamoto M, Murakami K, Kaji T, Zad-
worny D et al. (2006) DHCR24 gene knockout mice
demonstrate lethal dermopathy with differentiation and
maturation defects in the epidermis. J Invest Dermatol
126, 638–647.
24 Holmer L & Worman HJ (2001) Inner nuclear mem-
brane proteins: functions and targeting. Cell Mol Life
Sci 58, 1741–1747.
25 Ellenberg J, Siggia ED, Moreira JE, Smith CL, Presley
JF, Worman HJ & Lippincot-Schwartz J (1997) Nuclear
membrane dynamics and reassembly in living cells: tar-
geting of an inner nuclear membrane protein in inter-
phase and mitosis. J Cell Biol 138, 1193–1206.
26 Hoffmann K, Sperling K, Olins AL & Olins DE (2007)
The granulocyte nucleus and lamin B receptor: avoiding
the ovoid. Chromosoma 116, 227–235.
27 Zwerger M, Herrmann H, Gaines P, Olins AL & Olins
DE (2008) Granulocytic nuclear differentiation of lam-
in B receptor-deficient mouse EPRO cells. Exp Hematol
36, 977–987.
28 Martınez-Botas J, Ferruelo AJ, Suarez Y, Fernandez C,
Gomez-Coronado D & Lasuncion MA (2001) Dose-
dependent effects of lovastatin on cell-cycle progression.
Distinct requirement of cholesterol and non-sterol meval-
onate derivatives. Biochim Biophys Acta 1532, 185–194.
29 Fernandez C, Lobo M, Gomez-Coronado D & Las-
uncion MA (2004) Cholesterol is essential for mitosis
progression and its deficiency induces polyploid cell for-
mation. Exp Cell Res 300, 109–120.
30 Fernandez C, Martın M, Gomez-Coronado D & Las-
uncion MA (2005) Effects of distal cholesterol biosyn-
thesis inhibitors on cell proliferation and cell-cycle
progression. J Lipid Res 46, 920–929.
31 Wagner N, Weber D, Seitz S & Krohne G (2004) The
lamin B receptor of Drosophila melanogaster. J Cell Sci
117, 2015–2028.
32 Dreger CK, Konig AR, Spring H, Lichter P & Herr-
mann H (2002) Investigation of nuclear architecture
with a domain-presenting expression system. J Struct
Biol 140, 100–115.
33 Magin TM, McWir J & Melton DW (1992) A new
mouse embryonic stem cell line with good germ line
contribution and gene targeting frequency. Nucleic
Acids Res 20, 3795–3796.
34 Frohman MA (1993) Rapid amplification of comple-
mentary DNA ends for generation of full-length com-
plementary DNAs: thermal RACE. Methods Enzymol
218, 340–356.
35 Mancini A, Del Rosso F, Roberti R, Orvietani P,
Coletti L & Binaglia L (1999) Purification of ethanol-
aminephosphotransferase from bovine liver microsomes.
Biochim Biophys Acta 1437, 80–92.
36 Roberti R, Mancini A, Freysz L & Binaglia L (1992)
Reversibility of the reactions catalyzed by cholinephos-
photransferase and ethanolaminephosphotransferase
solubilized from rat brain microsomes. Biochim Biophys
Acta 1165, 183–188.
37 Kaufmann SH, Gibson W & Shaper JH (1983) Charac-
terization of the major polypeptides of the rat liver
nuclear envelope. J Biol Chem 258, 2710–2719.
38 Lowry OH, Rosebrough NJ, Farr AL & Randall RJ
(1951) Protein measurement with the Folin phenol
reagent. J Biol Chem 193, 265–275.
39 Paik Y-K, Trzaskos JM, Shafiee A & Gaylor JL (1984)
Microsomal enzymes of cholesterol biosynthesis from
lanosterol. Characterization, solubilization, and partial
purification of NADPH-dependent D8,14-steroid14-reductase. J Biol Chem 259, 13413–13423.
40 Folch J, Lees M & Sloane Stanley GH (1957) A
simplified method for the isolation and purification of
total lipids from animal tissues. J Biol Chem 226, 497–
509.
41 Macala LJ, Yu RK & Ando S (1983) Analysis of brain
lipids by high performance thin-layer chromatography
and densitometry. J Lipid Res 24, 1243–1250.
42 Pieroni S, Della Fazia MA, Castelli M, Piobbico D,
Bartoli D, Brunacci C, Bellet MM, Viola-Magni M &
Servillo G (2008) HOPS is an essential constituent of
centrosome assembly. Cell Cycle 7, 1462–1466.
A. M. Bennati et al. Tm7sf2 knockout mice
FEBS Journal 275 (2008) 5034–5047 ª 2008 The Authors Journal compilation ª 2008 FEBS 5047
Related Documents











