Molecular Microbiology (2003) 50(3), 751–762 doi:10.1046/j.1365-2958.2003.03712.x © 2003 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology 1365-2958Blackwell Publishing Ltd, 200350 3751762Original ArticleR. Singh et al.Role of mptpB in the virulence of mycobacteria Accepted 7 July, 2003. *For correspondence. E-mail akt1000@ hotmail.com; Tel. (+91) 11 26888434; Fax (+91) 11 26885270. Disruption of mptpB impairs the ability of Mycobacterium tuberculosis to survive in guinea pigs Ramandeep Singh, 1 Vivek Rao, 1 H. Shakila, 2 Radhika Gupta, 1 Aparna Khera, 1 Neeraj Dhar, 1 Amit Singh, 1 Anil Koul, 1,3 Yogendra Singh, 3 M. Naseema, 2 P. R. Narayanan, 2 C. N. Paramasivan, 2 V. D. Ramanathan 2 and Anil K. Tyagi 1 * 1 Department of Biochemistry, University of Delhi South campus, Benito Juarez Road, New Delhi-110021, India. 2 Tuberculosis Research Centre, Mayor V. R. Ramanathan Road, Chetput, Chennai-600031, India. 3 Institute of Genomics and Integrative Biology, Mall Road, Delhi-110007, India. Summary Protein tyrosine kinases and tyrosine phosphatases from several bacterial pathogens have been shown to act as virulence factors by modulating the phospho- rylation and dephosphorylation of host proteins. The identification and characterization of two tyrosine phosphatases namely MptpA and MptpB from Myco- bacterium tuberculosis has been reported earlier. MptpB is secreted by M. tuberculosis into extracellu- lar mileu and exhibits a pH optimum of 5.6, similar to the pH of the lysosomal compartment of the cell. To determine the role of MptpB in the pathogenesis of M. tuberculosis , we constructed a mptpB mutant strain by homologous recombination and compared the ability of parent and the mutant strain to survive intra- cellularly. We show that disruption of the mptpB gene impairs the ability of the mutant strain to survive in activated macrophages and guinea pigs but not in resting macrophages suggesting the importance of its role in the host–pathogen interaction. Infection of guinea pigs with the mutant strain resulted in a 70- fold reduction in the bacillary load of spleens in infected animals as compared with the bacillary load in animals infected with the parental strain. Upon rein- troduction of the mptpB gene into the mutant strain, the complemented strain was able to establish infec- tion and survive in guinea pigs at rates comparable to the parental strain. These observations demon- strate a role of MptpB in the pathogenesis of M. tuberculosis . Introduction Mycobacterium tuberculosis is the foremost cause of human deaths by any single infectious agent (Dolin et al ., 1994). This intracellular pathogen can successfully sur- vive inside the host macrophages in spite of the antimi- crobicidal effector functions of the macrophages (Lowrie, 1983; Sibille and Reynolds, 1990; Sathish and Shinnick, 1994). A number of studies have shown that mycobacteria are able to persist within phagocytes because phago- somes containing live organisms fail to fuse with lysos- omes and are depleted in proton ATPases responsible for acidification of phago-lysosomes (Armstrong and D’Arcy, 1971; Sturgill-Koszycki et al ., 1994). Several M. tubercu- losis genes viz. katG (catalase peroxidase), ahpC (alkylhydroperoxide reductase), sodA , sodC (superoxide dismutase) and noxR3 (nitric oxide reductase) have been implicated in protection against reactive oxygen and nitro- gen intermediates (Sherman et al ., 1996; Manca et al ., 1999; Ruan et al ., 1999; Dussurget et al ., 2001). Recently, it has also been shown that phagosomes containing live mycobacteria recruit and retain a tryptophan-aspartate- containing host protein, which prevents their delivery to lysosomes (Ferrari et al ., 1999). In order to evade the host immune machinery, many bacterial pathogens secrete effector molecules into host cells allowing a pathogen to modify host proteins that promote their survival in the host (Devinney et al ., 2000). Protein phosphorylation has been shown to play an impor- tant role in sensing of extracellular signals by a pathogen and co-ordinating intracellular events (Kennelly and Potts, 1996). Protein tyrosine kinases and phosphatases control the growth and differentiation of eukaryotic cells by mod- ulating the tyrosine phosphorylation of regulatory proteins (Stone and Dixon, 1994). Several protein phosphatases and kinases from bacterial pathogens have been shown to act as virulence factors (Guan and Dixon, 1990; Galyov et al ., 1993; Kaniga et al ., 1996). Protein tyrosine phos- phatases YopH and SptP from Yersinia pseudotuberculo- sis and Salmonella typhimurium , respectively, have been well characterized. Protein tyrosine dephosphorylation of macrophage proteins by YopH prevents phagocytosis- linked signalling pathways (Bliska et al ., 1991; Bliska and Black, 1995; Fallman et al ., 1995; Andersson et al ., 1996; Black and Bliska, 1997). Similarly, SptP of S. typhimurium has been shown to play an important role in cytoskeletal

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Microbiology (2003)
50
(3), 751–762 doi:10.1046/j.1365-2958.2003.03712.x
© 2003 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology 1365-2958Blackwell Publishing Ltd, 200350
3751762
Original Article
R. Singh et al.Role of mptpB in the virulence of mycobacteria
Accepted 7 July, 2003. *For correspondence. E-mail [email protected]; Tel. (
+
91) 11 26888434; Fax (
+
91) 11 26885270.
Disruption of
mptpB
impairs the ability of
Mycobacterium tuberculosis
to survive in guinea pigs
Ramandeep Singh,
1
Vivek Rao,
1
H. Shakila,
2
Radhika Gupta,
1
Aparna Khera,
1
Neeraj Dhar,
1
Amit Singh,
1
Anil Koul,
1,3
Yogendra Singh,
3
M. Naseema,
2
P. R. Narayanan,
2
C. N. Paramasivan,
2
V. D. Ramanathan
2
and Anil K. Tyagi
1
*
1
Department of Biochemistry, University of Delhi South campus, Benito Juarez Road, New Delhi-110021, India.
2
Tuberculosis Research Centre, Mayor V. R. Ramanathan Road, Chetput, Chennai-600031, India.
3
Institute of Genomics and Integrative Biology, Mall Road, Delhi-110007, India.
Summary
Protein tyrosine kinases and tyrosine phosphatasesfrom several bacterial pathogens have been shown toact as virulence factors by modulating the phospho-rylation and dephosphorylation of host proteins. Theidentification and characterization of two tyrosinephosphatases namely MptpA and MptpB from
Myco-bacterium tuberculosis
has been reported earlier.MptpB is secreted by
M. tuberculosis
into extracellu-lar mileu and exhibits a pH optimum of 5.6, similar tothe pH of the lysosomal compartment of the cell. Todetermine the role of MptpB in the pathogenesis of
M.tuberculosis
, we constructed a
mptpB
mutant strainby homologous recombination and compared theability of parent and the mutant strain to survive intra-cellularly. We show that disruption of the
mptpB
geneimpairs the ability of the mutant strain to survive inactivated macrophages and guinea pigs but not inresting macrophages suggesting the importance ofits role in the host–pathogen interaction. Infection ofguinea pigs with the mutant strain resulted in a 70-fold reduction in the bacillary load of spleens ininfected animals as compared with the bacillary loadin animals infected with the parental strain. Upon rein-troduction of the
mptpB
gene into the mutant strain,the complemented strain was able to establish infec-tion and survive in guinea pigs at rates comparableto the parental strain. These observations demon-strate a role of MptpB in the pathogenesis of
M. tuberculosis
.
Introduction
Mycobacterium tuberculosis
is the foremost cause ofhuman deaths by any single infectious agent (Dolin
et al
.,1994). This intracellular pathogen can successfully sur-vive inside the host macrophages in spite of the antimi-crobicidal effector functions of the macrophages (Lowrie,1983; Sibille and Reynolds, 1990; Sathish and Shinnick,1994). A number of studies have shown that mycobacteriaare able to persist within phagocytes because phago-somes containing live organisms fail to fuse with lysos-omes and are depleted in proton ATPases responsible foracidification of phago-lysosomes (Armstrong and D’Arcy,1971; Sturgill-Koszycki
et al
., 1994). Several
M. tubercu-losis
genes viz.
katG
(catalase peroxidase),
ahpC
(alkylhydroperoxide reductase),
sodA
,
sodC
(superoxidedismutase) and
noxR3
(nitric oxide reductase) have beenimplicated in protection against reactive oxygen and nitro-gen intermediates (Sherman
et al
., 1996; Manca
et al
.,1999; Ruan
et al
., 1999; Dussurget
et al
., 2001). Recently,it has also been shown that phagosomes containing livemycobacteria recruit and retain a tryptophan-aspartate-containing host protein, which prevents their delivery tolysosomes (Ferrari
et al
., 1999).In order to evade the host immune machinery, many
bacterial pathogens secrete effector molecules into hostcells allowing a pathogen to modify host proteins thatpromote their survival in the host (Devinney
et al
., 2000).Protein phosphorylation has been shown to play an impor-tant role in sensing of extracellular signals by a pathogenand co-ordinating intracellular events (Kennelly and Potts,1996). Protein tyrosine kinases and phosphatases controlthe growth and differentiation of eukaryotic cells by mod-ulating the tyrosine phosphorylation of regulatory proteins(Stone and Dixon, 1994). Several protein phosphatasesand kinases from bacterial pathogens have been shownto act as virulence factors (Guan and Dixon, 1990; Galyov
et al
., 1993; Kaniga
et al
., 1996). Protein tyrosine phos-phatases YopH and SptP from
Yersinia pseudotuberculo-sis
and
Salmonella typhimurium
, respectively, have beenwell characterized. Protein tyrosine dephosphorylation ofmacrophage proteins by YopH prevents phagocytosis-linked signalling pathways (Bliska
et al
., 1991; Bliska andBlack, 1995; Fallman
et al
., 1995; Andersson
et al
., 1996;Black and Bliska, 1997). Similarly, SptP of
S. typhimurium
has been shown to play an important role in cytoskeletal

752
R. Singh
et al.
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
50
, 751–762
rearrangements by interacting with Rac-1 and Cdc42leading to internalization of bacteria into non-phagocyticcells (Fu and Galan, 1999).
Sequence analysis of the
M. tuberculosis
genomerevealed the presence of 11 serine/threonine kinases,two tyrosine phosphatases and one serine/threoninephosphatase (Cole
et al
., 1998). A few serine/threoninekinases (PknA, PknB, PknD, PknF and PknG) andtyrosine phosphatases (MptpA and MptpB) of
M. tuber-culosis
have been studied for their biochemical proper-ties (Peirs
et al
., 1997; Av-Gay
et al
., 1999; Koul
et al
.,2000; 2001; Chaba
et al
., 2002). Both MptpA and MptpBare secreted by
M. tuberculosis
into extracellular fluid(Koul
et al
., 2000). Using Southern blot analysis, it wasearlier demonstrated that
mptpB
was present in the slow-growing species such as
M. tuberculosis
H
37
Rv,
M. tuber-culosis
H
37
Ra and
M. bovis
BCG but absent from thefast-growing species such as
M. smegmatis
(Koul
et al
.,2000). MptpB of
M. tuberculosis
was shown to dephos-phorylate the phosphotyrosine residue of myelin basicprotein (Koul
et al
., 2000). Because phosphorylation anddephosphorylation regulate several crucial processes ineukaryotic cells, secreted tyrosine phosphatases mayserve as key molecules that modify host proteins to itsadvantage and enable the tubercle bacilli to survivewithin the host. By virtue of its
in vitro
tyrosine phos-phatase activity, secretory nature and optimal pH of 5.6for activity, we speculated that MptpB might interfere withhost signal transduction pathways facilitating the survivalof the pathogen in the host.
In order to evaluate the role of MptpB in the pathogen-esis of
M. tuberculosis
, an
mptpB
mutant strain was con-structed by homologous recombination. The mutant strainlacking tyrosine phosphatase activity associated withMptpB was employed to understand the role of this proteinin the pathogenesis of
M. tuberculosis
. Using the guineapig model of experimental tuberculosis, we report that the
mptpB
mutant strain showed impaired ability to survive inthe host.
Results
Construction of the
mptpB
mutant and complemented strain of
M. tuberculosis
In order to establish whether MptpB plays a role in thepathogenesis of
M. tuberculosis
, an
mptpB
mutant strainof
M. tuberculosis
was constructed by using a non-repli-cative suicidal vector pBK
D
B (Fig. 1). The targeting vector,pBK
D
B, carried the coding region of
mptpB
along with1045 bp upstream and 1140 bp downstream flankingsequences. A portion of the coding region (108 bp) ofMptpB was deleted and replaced with the gene conferringresistance to hygromycin in pBK
D
B. The vector also car-
ried the gene conferring resistance to kanamycin in itsbackbone as a second antibiotic selection marker for neg-ative screening of allelic exchange events at the homolo-gous site (Fig. 1).
Electroporation of
M. tuberculosis
with pBK
D
B and UV-treated pBK
D
B resulted in 22 and 3 hygromycin-resistanttransformants respectively. PCR analysis revealed that allthe transformants contained a hygromycin cassette indi-cating that these colonies were not spontaneous resis-tance mutants and arose from integration of the suicidalvector into the mycobacterial genome (data not shown).An allelic exchange event by homologous recombinationshould result in the incorporation of a hygromycin resis-tance gene but not the vector backbone (having kanamy-cin resistance gene) into the mycobacterial genome.Thus, transformants resistant to hygromycin but sensitiveto kanamycin were selected to score for a homologousrecombination event. All the transformants obtained onelectroporation of untreated DNA were kanamycin resis-tant while the three transformants obtained on electropo-ration of UV-pre-treated DNA were sensitive tokanamycin. This suggested that an allelic exchange eventat the homologous site had taken place in the case ofthese three transformants obtained upon electroporationof UV-pre-treated DNA.
Fig. 1.
Schematic representation of the
mptpB
locus in the genome of parental (A) and mutant strain (C) of
Mycobacterium tuberculosis
. Details of construction of pBK
D
B (B), a non-replicative suicide vector used for disruption of
mptpB
is given in
Experimental procedures
.
Hyg
R
and
Kan
R
represent genes conferring resistance to hygromycin and kanamycin respectively. ML and MR represent
M. tuberculosis
DNA from the
mptpB
locus, oriT represents the origin of DNA repli-cation and
sacB
represents the gene encoding levansucrase.
Role of
mptpB
in the virulence of mycobacteria
753
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
50
, 751–762
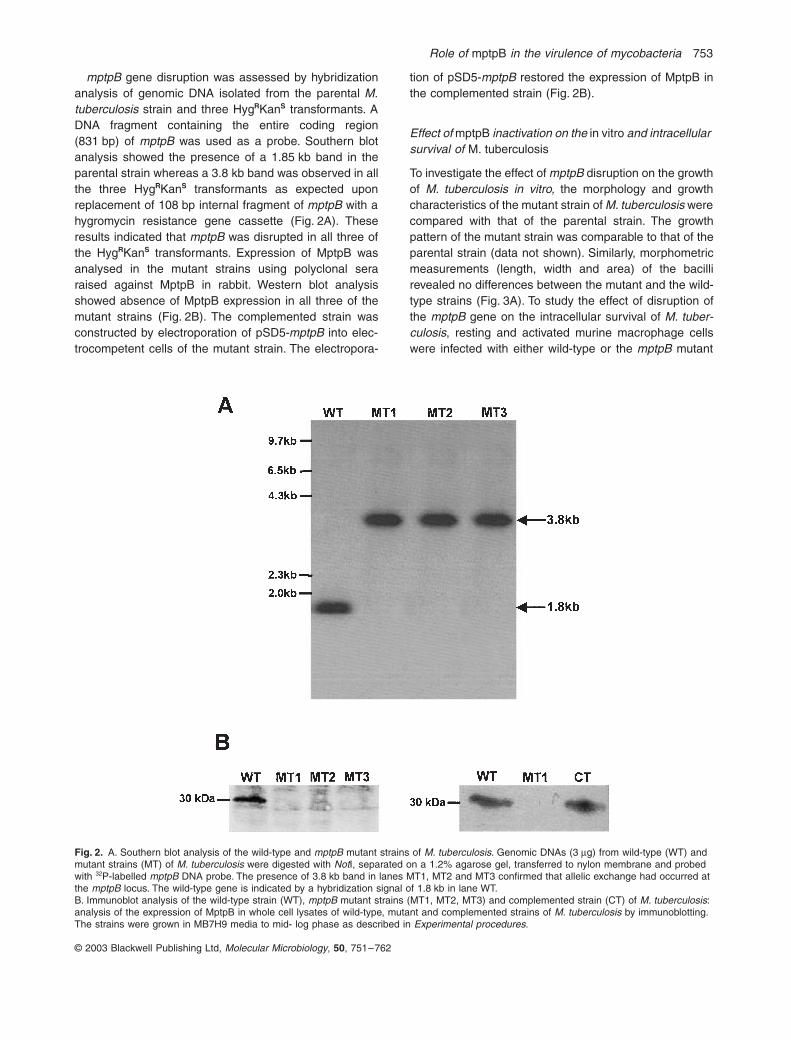
mptpB
gene disruption was assessed by hybridizationanalysis of genomic DNA isolated from the parental
M.tuberculosis
strain and three Hyg
R
Kan
S
transformants. ADNA fragment containing the entire coding region(831 bp) of
mptpB
was used as a probe. Southern blotanalysis showed the presence of a 1.85 kb band in theparental strain whereas a 3.8 kb band was observed in allthe three Hyg
R
Kan
S
transformants as expected uponreplacement of 108 bp internal fragment of
mptpB
with ahygromycin resistance gene cassette (Fig. 2A). Theseresults indicated that
mptpB
was disrupted in all three ofthe Hyg
R
Kan
S
transformants. Expression of MptpB wasanalysed in the mutant strains using polyclonal seraraised against MptpB in rabbit. Western blot analysisshowed absence of MptpB expression in all three of themutant strains (Fig. 2B). The complemented strain wasconstructed by electroporation of pSD5-
mptpB
into elec-trocompetent cells of the mutant strain. The electropora-
tion of pSD5-
mptpB
restored the expression of MptpB inthe complemented strain (Fig. 2B).
Effect of mptpB inactivation on the in vitro and intracellular survival of M. tuberculosis
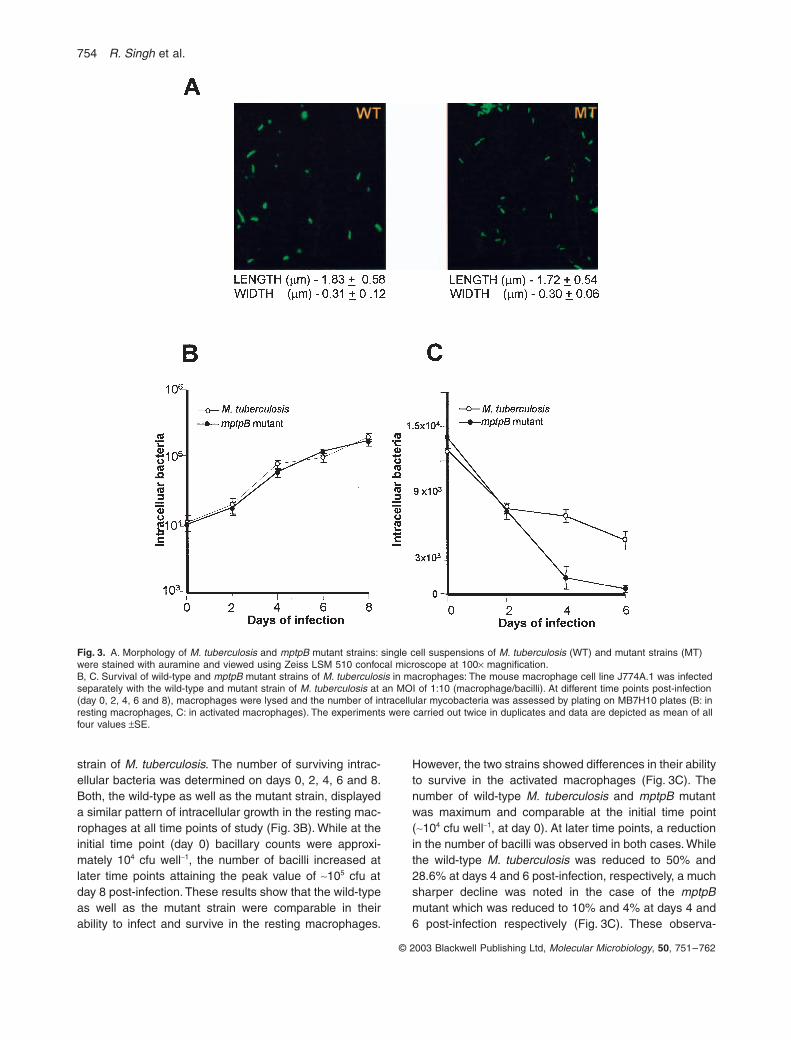
To investigate the effect of mptpB disruption on the growthof M. tuberculosis in vitro, the morphology and growthcharacteristics of the mutant strain of M. tuberculosis werecompared with that of the parental strain. The growthpattern of the mutant strain was comparable to that of theparental strain (data not shown). Similarly, morphometricmeasurements (length, width and area) of the bacillirevealed no differences between the mutant and the wild-type strains (Fig. 3A). To study the effect of disruption ofthe mptpB gene on the intracellular survival of M. tuber-culosis, resting and activated murine macrophage cellswere infected with either wild-type or the mptpB mutant
Fig. 2. A. Southern blot analysis of the wild-type and mptpB mutant strains of M. tuberculosis. Genomic DNAs (3 mg) from wild-type (WT) and mutant strains (MT) of M. tuberculosis were digested with NotI, separated on a 1.2% agarose gel, transferred to nylon membrane and probed with 32P-labelled mptpB DNA probe. The presence of 3.8 kb band in lanes MT1, MT2 and MT3 confirmed that allelic exchange had occurred at the mptpB locus. The wild-type gene is indicated by a hybridization signal of 1.8 kb in lane WT.B. Immunoblot analysis of the wild-type strain (WT), mptpB mutant strains (MT1, MT2, MT3) and complemented strain (CT) of M. tuberculosis: analysis of the expression of MptpB in whole cell lysates of wild-type, mutant and complemented strains of M. tuberculosis by immunoblotting. The strains were grown in MB7H9 media to mid- log phase as described in Experimental procedures.
754 R. Singh et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
strain of M. tuberculosis. The number of surviving intrac-ellular bacteria was determined on days 0, 2, 4, 6 and 8.Both, the wild-type as well as the mutant strain, displayeda similar pattern of intracellular growth in the resting mac-rophages at all time points of study (Fig. 3B). While at theinitial time point (day 0) bacillary counts were approxi-mately 104 cfu well-1, the number of bacilli increased atlater time points attaining the peak value of ~105 cfu atday 8 post-infection. These results show that the wild-typeas well as the mutant strain were comparable in theirability to infect and survive in the resting macrophages.
However, the two strains showed differences in their abilityto survive in the activated macrophages (Fig. 3C). Thenumber of wild-type M. tuberculosis and mptpB mutantwas maximum and comparable at the initial time point(~104 cfu well-1, at day 0). At later time points, a reductionin the number of bacilli was observed in both cases. Whilethe wild-type M. tuberculosis was reduced to 50% and28.6% at days 4 and 6 post-infection, respectively, a muchsharper decline was noted in the case of the mptpBmutant which was reduced to 10% and 4% at days 4 and6 post-infection respectively (Fig. 3C). These observa-
Fig. 3. A. Morphology of M. tuberculosis and mptpB mutant strains: single cell suspensions of M. tuberculosis (WT) and mutant strains (MT) were stained with auramine and viewed using Zeiss LSM 510 confocal microscope at 100¥ magnification.B, C. Survival of wild-type and mptpB mutant strains of M. tuberculosis in macrophages: The mouse macrophage cell line J774A.1 was infected separately with the wild-type and mutant strain of M. tuberculosis at an MOI of 1:10 (macrophage/bacilli). At different time points post-infection (day 0, 2, 4, 6 and 8), macrophages were lysed and the number of intracellular mycobacteria was assessed by plating on MB7H10 plates (B: in resting macrophages, C: in activated macrophages). The experiments were carried out twice in duplicates and data are depicted as mean of all four values ±SE.
Role of mptpB in the virulence of mycobacteria 755
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
tions indicated that disruption of the mptpB gene hadimpaired the ability of M. tuberculosis to survive in theactivated macrophages.
Virulence of the mptpB mutant strain of M. tuberculosis in guinea pig
To determine whether the disruption of mptpB geneswould have any effect on the survival of M. tuberculosisin vivo, guinea pigs in groups of eight animals wereinfected subcutaneously with 5 ¥ 105 cfu of parental,mutant or the complemented strain of M. tuberculosis.Animals were euthanized 3 and 6 weeks post-infection. Atboth time points of euthanasia, spleens were homoge-nized and viable bacilli were enumerated (represented aslog10 cfu for each group).
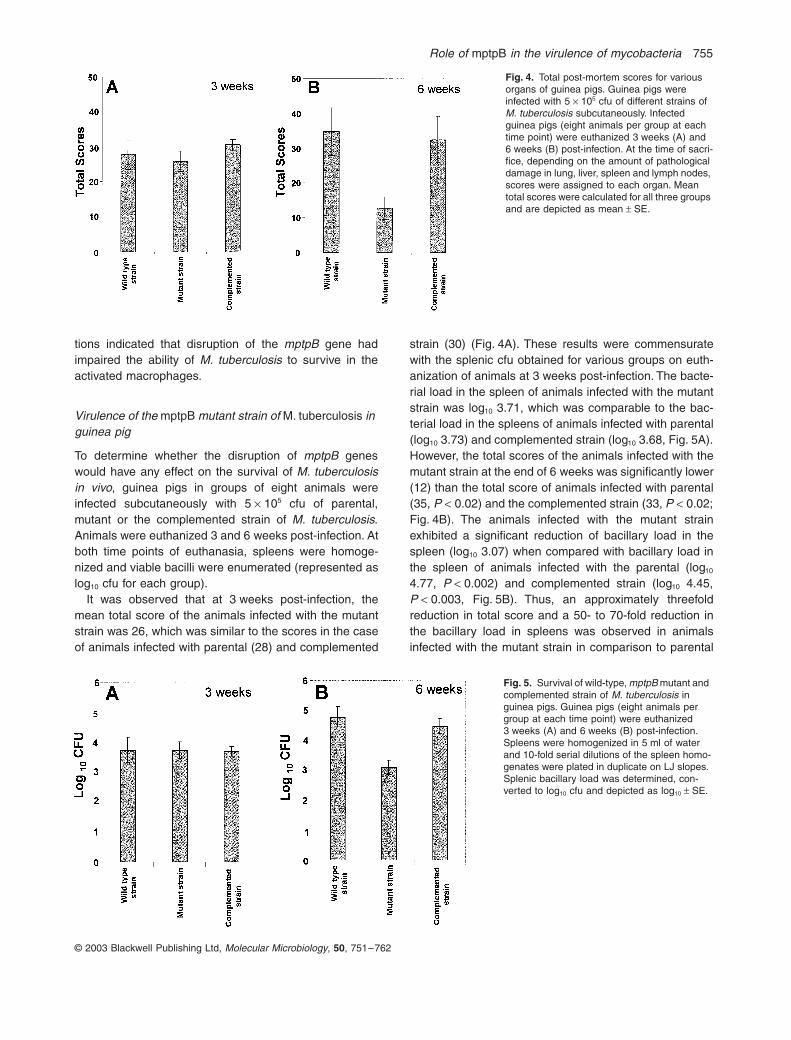
It was observed that at 3 weeks post-infection, themean total score of the animals infected with the mutantstrain was 26, which was similar to the scores in the caseof animals infected with parental (28) and complemented
strain (30) (Fig. 4A). These results were commensuratewith the splenic cfu obtained for various groups on euth-anization of animals at 3 weeks post-infection. The bacte-rial load in the spleen of animals infected with the mutantstrain was log10 3.71, which was comparable to the bac-terial load in the spleens of animals infected with parental(log10 3.73) and complemented strain (log10 3.68, Fig. 5A).However, the total scores of the animals infected with themutant strain at the end of 6 weeks was significantly lower(12) than the total score of animals infected with parental(35, P < 0.02) and the complemented strain (33, P < 0.02;Fig. 4B). The animals infected with the mutant strainexhibited a significant reduction of bacillary load in thespleen (log10 3.07) when compared with bacillary load inthe spleen of animals infected with the parental (log10
4.77, P < 0.002) and complemented strain (log10 4.45,P < 0.003, Fig. 5B). Thus, an approximately threefoldreduction in total score and a 50- to 70-fold reduction inthe bacillary load in spleens was observed in animalsinfected with the mutant strain in comparison to parental
Fig. 4. Total post-mortem scores for various organs of guinea pigs. Guinea pigs were infected with 5 ¥ 105 cfu of different strains of M. tuberculosis subcutaneously. Infected guinea pigs (eight animals per group at each time point) were euthanized 3 weeks (A) and 6 weeks (B) post-infection. At the time of sacri-fice, depending on the amount of pathological damage in lung, liver, spleen and lymph nodes, scores were assigned to each organ. Mean total scores were calculated for all three groups and are depicted as mean ± SE.
Fig. 5. Survival of wild-type, mptpB mutant and complemented strain of M. tuberculosis in guinea pigs. Guinea pigs (eight animals per group at each time point) were euthanized 3 weeks (A) and 6 weeks (B) post-infection. Spleens were homogenized in 5 ml of water and 10-fold serial dilutions of the spleen homo-genates were plated in duplicate on LJ slopes. Splenic bacillary load was determined, con-verted to log10 cfu and depicted as log10 ± SE.
756 R. Singh et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
or complemented strains. We have carried out this exper-iment three times obtaining similar results.
Histopathological analysis
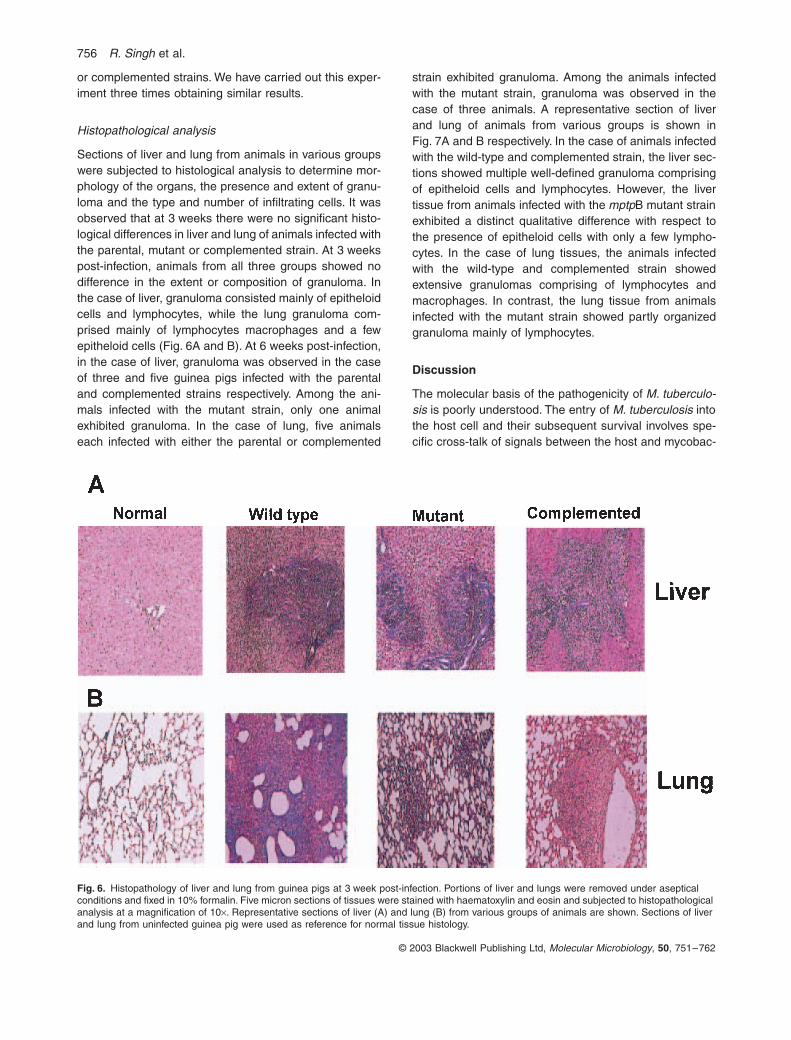
Sections of liver and lung from animals in various groupswere subjected to histological analysis to determine mor-phology of the organs, the presence and extent of granu-loma and the type and number of infiltrating cells. It wasobserved that at 3 weeks there were no significant histo-logical differences in liver and lung of animals infected withthe parental, mutant or complemented strain. At 3 weekspost-infection, animals from all three groups showed nodifference in the extent or composition of granuloma. Inthe case of liver, granuloma consisted mainly of epitheloidcells and lymphocytes, while the lung granuloma com-prised mainly of lymphocytes macrophages and a fewepitheloid cells (Fig. 6A and B). At 6 weeks post-infection,in the case of liver, granuloma was observed in the caseof three and five guinea pigs infected with the parentaland complemented strains respectively. Among the ani-mals infected with the mutant strain, only one animalexhibited granuloma. In the case of lung, five animalseach infected with either the parental or complemented
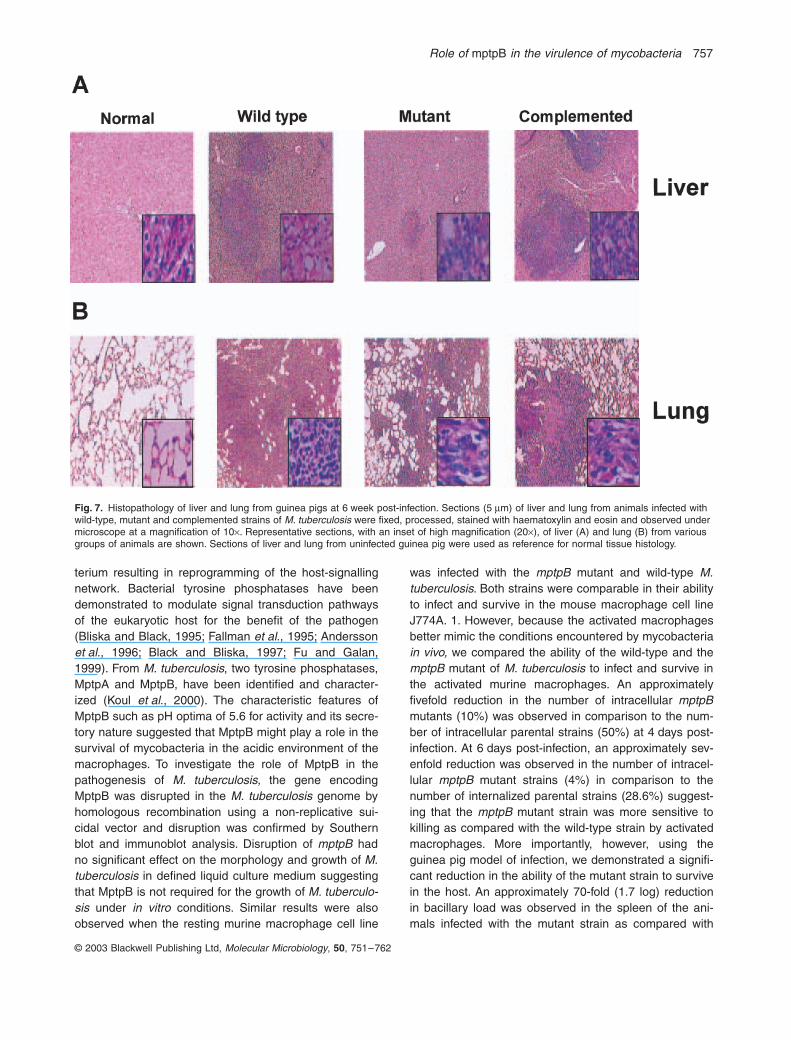
strain exhibited granuloma. Among the animals infectedwith the mutant strain, granuloma was observed in thecase of three animals. A representative section of liverand lung of animals from various groups is shown inFig. 7A and B respectively. In the case of animals infectedwith the wild-type and complemented strain, the liver sec-tions showed multiple well-defined granuloma comprisingof epitheloid cells and lymphocytes. However, the livertissue from animals infected with the mptpB mutant strainexhibited a distinct qualitative difference with respect tothe presence of epitheloid cells with only a few lympho-cytes. In the case of lung tissues, the animals infectedwith the wild-type and complemented strain showedextensive granulomas comprising of lymphocytes andmacrophages. In contrast, the lung tissue from animalsinfected with the mutant strain showed partly organizedgranuloma mainly of lymphocytes.
Discussion
The molecular basis of the pathogenicity of M. tuberculo-sis is poorly understood. The entry of M. tuberculosis intothe host cell and their subsequent survival involves spe-cific cross-talk of signals between the host and mycobac-
Fig. 6. Histopathology of liver and lung from guinea pigs at 3 week post-infection. Portions of liver and lungs were removed under aseptical conditions and fixed in 10% formalin. Five micron sections of tissues were stained with haematoxylin and eosin and subjected to histopathological analysis at a magnification of 10¥. Representative sections of liver (A) and lung (B) from various groups of animals are shown. Sections of liver and lung from uninfected guinea pig were used as reference for normal tissue histology.
Role of mptpB in the virulence of mycobacteria 757
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
terium resulting in reprogramming of the host-signallingnetwork. Bacterial tyrosine phosphatases have beendemonstrated to modulate signal transduction pathwaysof the eukaryotic host for the benefit of the pathogen(Bliska and Black, 1995; Fallman et al., 1995; Anderssonet al., 1996; Black and Bliska, 1997; Fu and Galan,1999). From M. tuberculosis, two tyrosine phosphatases,MptpA and MptpB, have been identified and character-ized (Koul et al., 2000). The characteristic features ofMptpB such as pH optima of 5.6 for activity and its secre-tory nature suggested that MptpB might play a role in thesurvival of mycobacteria in the acidic environment of themacrophages. To investigate the role of MptpB in thepathogenesis of M. tuberculosis, the gene encodingMptpB was disrupted in the M. tuberculosis genome byhomologous recombination using a non-replicative sui-cidal vector and disruption was confirmed by Southernblot and immunoblot analysis. Disruption of mptpB hadno significant effect on the morphology and growth of M.tuberculosis in defined liquid culture medium suggestingthat MptpB is not required for the growth of M. tuberculo-sis under in vitro conditions. Similar results were alsoobserved when the resting murine macrophage cell line
was infected with the mptpB mutant and wild-type M.tuberculosis. Both strains were comparable in their abilityto infect and survive in the mouse macrophage cell lineJ774A. 1. However, because the activated macrophagesbetter mimic the conditions encountered by mycobacteriain vivo, we compared the ability of the wild-type and themptpB mutant of M. tuberculosis to infect and survive inthe activated murine macrophages. An approximatelyfivefold reduction in the number of intracellular mptpBmutants (10%) was observed in comparison to the num-ber of intracellular parental strains (50%) at 4 days post-infection. At 6 days post-infection, an approximately sev-enfold reduction was observed in the number of intracel-lular mptpB mutant strains (4%) in comparison to thenumber of internalized parental strains (28.6%) suggest-ing that the mptpB mutant strain was more sensitive tokilling as compared with the wild-type strain by activatedmacrophages. More importantly, however, using theguinea pig model of infection, we demonstrated a signifi-cant reduction in the ability of the mutant strain to survivein the host. An approximately 70-fold (1.7 log) reductionin bacillary load was observed in the spleen of the ani-mals infected with the mutant strain as compared with
Fig. 7. Histopathology of liver and lung from guinea pigs at 6 week post-infection. Sections (5 mm) of liver and lung from animals infected with wild-type, mutant and complemented strains of M. tuberculosis were fixed, processed, stained with haematoxylin and eosin and observed under microscope at a magnification of 10¥. Representative sections, with an inset of high magnification (20¥), of liver (A) and lung (B) from various groups of animals are shown. Sections of liver and lung from uninfected guinea pig were used as reference for normal tissue histology.

758 R. Singh et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
the bacillary load from the animals infected with wild-typestrain at 6 week post-infection. This difference in thesplenic bacillary load in both groups of animals was notobserved earlier, at 3 weeks post-infection (log10 3.73 forthe wild-type strain and log10 3.71 for the mptpB mutantstrain) suggesting that the wild-type as well as the mutantstrain were capable of establishing the infection to a com-parable extent. However, at 6 weeks post-infection thebacillary load in the spleens of animals infected withparental strain increased from log10 3.73 to log10 4.77. Inthe case of animals infected with the mptpB mutantstrain, the splenic bacterial load decreased from log10
3.71 to log10 3.07 and this decrease was found to bestatistically significant. These observations suggest thatinitially both strains (mutant and wild type) of M. tubercu-losis are capable of establishing the infection to a similarextent. However, the mutant strain displayed reducedability to sustain infection in guinea pigs.
That the loss of virulence of M. tuberculosis was a directconsequence of disruption of mptpB, was shown by theability of the complemented strain to establish an infectionand survive in the host tissues at levels similar to thoseobserved in the case of wild-type M. tuberculosis. Theseobservations clearly suggest that MptpB plays an essen-tial role in the virulence of M. tuberculosis.
It is well established that phosphatases not only antag-onize kinases but can also act co-operatively to generateantagonistic signals (Sun and Tonks, 1994). Thus, it is notsurprising that pathogenic microorganisms secretetyrosine phosphatases to modulate host antimicrobialfunctions. YopH is required for full display of virulence ofYersinia (Guan and Dixon, 1990). A strain of S. typhimu-rium carrying a null mutation in sptP is defective in its
ability to survive and establish an infection in BALB/c micesuggesting the role of SptP in the pathogenesis of Salmo-nella (Kaniga et al., 1996). At present it is not clear howMptpB exerts its function. A number of host functions suchas antigen presentation, macrophage activation andmembrane trafficking that influence the course of M.tuberculosis pathogenesis are modulated by the levels oftyrosine phosphorylation (Adams and Hamilton, 1984).Because the mptpB mutant is impaired in its ability tosurvive in activated macrophages, IFN-g-mediated signaltransduction pathways may represent a plausible targetfor MptpB.
Experimental procedures
Bacterial strains and DNA manipulations
The bacterial strains and plasmids used in this study arelisted in Table 1. Plasmids were prepared in Escherichiacoli HB101 and DH5a strains using the Qiagen miniprep kitaccording to the manufacturer’s recommendations (QiagenInc.). All mycobacterial strains were grown in Middlebrook(MB) 7H9 medium (Difco) supplemented with 0.5% glyc-erol, 0.2% Tween-80 and 1¥ ADC (Difco) or MB 7H10medium (Difco) supplemented with 1¥ OADC (Difco). Whenrequired, the following antibiotics were added at the speci-fied concentration: ampicillin (50 mg ml-1 for E. coli ), kana-mycin A sulphate (25 mg ml-1 for both E. coli andmycobacteria) and hygromycin B (150 mg ml-1 for E. coliand 50 mg ml-1 for mycobacteria). All restriction endonu-cleases, DNA-modifying enzymes such as T4 DNA ligase,T4 polynucleotide kinase (New England Biolabs Inc.) andPfu DNA polymerase (Stratagene GmbH) were usedaccording to manufacturers’ recommendations. All recombi-nant DNA manipulations were carried out according tostandard protocols.
Table 1. Strains and plasmids used in this study.
Strain/plasmid Description Reference
Escherichia coli HB101 F– D (gpt-proA) 62 leuB6 glnV44 ara-14 galK2 lacY1 D(mcrC-mrr)rpsL20 (Strr) xyl-5 mtl-1 recA13
Life Technologies
E. coli DH5aF¢ F¢/endA1 hsdR17 (rk– mk
+) glnV44 thi-1 recA1 gyrA (Nalr) relA1 D(lacIZYA-argF) U169 deoR [f80dlacD(lacZ) M15]
Life Technologies
Mycobacterium tuberculosisErdman
Virulent strain of M. tuberculosis Kindly provided by Dr Jaya S. TyagiAIIMS, New Delhi, India.
mptpB mutant strain M. tuberculosis with its mptpB gene disrupted by the hygromycinresistance cassette
This study
Complemented strain mptpB mutant strain complemented with a wild-type copy of mptpB This studypLitmus 38 Cloning vector with colE1 origin of DNA replication New England BiolabspLit28 res-hyg-res pLitmus-28 derivative carrying an E. coli origin of DNA replication
and hygromycin resistance gene cassetteA kind gift from Dr Stoyan Bardarov,
Albert Einstein College of Medicine,New York, USA
pSD5 Mycobacteria – E. coli shuttle vector carrying the kanamycinresistance gene
Dasgupta et al. (1998)
pJQ200SK A non-replicative suicidal vector carrying genes for SacB andgentamicin resistance
A kind gift from Dr Brigitte Gicquel,Pasteur Institute, France
pJQDB Derivative of pJQ200SK carrying mptpDB::hygr cloned in the XbaI site This studypBKDB Derivative of pJQDB carrying the kanamycin resistance gene cloned
in the vector backboneThis study
pSD5-mptpB Mycobacteria-E. coli shuttle vector pSD5 carrying the mptpB gene This study

Role of mptpB in the virulence of mycobacteria 759
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
Construction of targeting vector pBKDB
For disruption of mptpB of M. tuberculosis, vector pBKDBwas constructed. For this, a DNA fragment containing1045 bp upstream of the open reading frame (ORF) ofmptpB along with the initial 356 bp of the ORF of mptpB wasPCR amplified using M. tuberculosis Erdman DNA as tem-plate and primers ptpB.1 having a BspHI site (5¢-cca tca tgactg tgg aac cta ttc ctg tcg gcc-3¢) and ptpB.2 having a NdeIsite (5¢-ggg cat atg ggc tgg att cgc cgg act cgc cg-3¢). Theamplicon was end-repaired and cloned into EcoRV digestedpLitmus-38 resulting in pLitB1. Another DNA fragment con-taining 367 bp of the mptpB ORF corresponding to the C-terminal region of MptpB along with 1140 bp downstream ofthe ORF of mptpB was PCR amplified using gene-specificprimers ptpB.3 carrying a NdeI site (5¢-ggg cat atg ggt gctcac cca ctg ctt cgc ggg-3¢) and ptpB.4 having a BspHI site(5¢-cca tca tga gtc ggt gac ccc cgt ata gcc cgg-3¢). Theamplicon was end-repaired and separately cloned intoEcoRV digested pLitmus-38 resulting in pLitB2. The vectorpLitB1 was digested with NdeI and ScaI and the larger DNAfragment containing the initial 356 bp of the mptpB ORFalong with 1045 bp upstream of the ORF was gel purifiedusing a Qiagen gel extraction kit. Similarly, pLitB2 wasdigested with NdeI and ScaI and the smaller DNA fragmentcontaining the 367 bp of the mptpB ORF corresponding tothe C-terminal region of MptpB along with 1140 bp down-stream of the ORF was gel purified. The larger fragmentobtained by the digestion of pLitB1 and the smaller fragmentobtained by the digestion of pLitB2 were then ligatedtogether resulting into pLitDB. The mptpB-specific primersptpB.2 and ptpB.3 were non-overlapping; as a result, thevector pLitDB contained the coding region of mptpB (with adeletion of 108 bp from the central region of ORF) and1045 bp of upstream and 1140 bp of downstream flankingsequences and a unique NdeI site in the ORF of mptpB atthe deletion site for the cloning of the hygromycin resistancegene cassette. The hygromycin resistance gene cassettewas excised out from pLit28res-hyg-res as a BamHI–XbaIfragment, end-repaired and cloned into NdeI digested, end-repaired pLitDB resulting in pLitDBH. A 4.9 kb DNA fragmentcontaining mptpDB::hygr was excised out from pLitDBH as aSpeI–NheI fragment and cloned into XbaI-digestedpJQ200SK (a non-replicative suicide vector, Pelicic et al.,1996) yielding pJQDB. The gene conferring resistance tokanamycin was excised out from pSD5 as a NheI–BstEIIfragment, end repaired and cloned into SmaI-digestedpJQDB resulting in pBKDB. The vector pBKDB provided 1.4and 1.5 kb homologous regions upstream and downstreamof the hygromycin-resistant gene, respectively, for recombi-nation to occur between targeting DNA and the mycobacte-rial genome.
Construction of the mptpB mutant andcomplementation strain
The targeting vector pBKDB was pre-treated with ultravio-let light (UV) as described (Hinds et al., 1999). In brief,UV irradiation of DNA was carried out in an UVstratalinker 1800 (Amersham Life Science) at 100 mJ cm-2.UV-treated DNA as well as untreated DNA (3 mg each)
was electroporated into electrocompetent cells of M. tuber-culosis. The cells were revived in 1 ml of MB7H9 mediafor 48 h, plated on MB7H10/OADC plates supplementedwith hygromycin (50 mg ml-1) and incubated at 37∞C for3 week. Colonies were screened for hygromycin resistanceand kanamycin sensitivity. Disruption of the mptpB gene ofM. tuberculosis was confirmed by Southern blot andimmunoblot analysis.
For complementation studies, the complete mptpB genewas PCR amplified using gene-specific primers and clonedinto pSD5 hsp60 (Dasgupta et al., 1998). The resulting plas-mid pSD5-mptpB was electroporated into the mptpB mutantstrain of M. tuberculosis.
Southern blot analysis
Genomic DNA from mycobacteria was extracted asdescribed earlier. DNA (3 mg) was digested with NotI andresolved on a 1.2% agarose gel. The gel was treated accord-ing to standard protocols. DNA fragments were transferredto Hybond-N membrane (Amersham Pharmacia Biotech.),cross-linked by UV irradiation and hybridized under standardconditions (Southern, 1975) with 32P-labelled fragment con-taining the complete coding region of mptpB, in 50% forma-mide at 42∞C for 16 h. Blots were washed once with 2¥ SSC/0.1% SDS at room temperature for 30 min followed by twowashes with 0.1¥ SSC/0.5% SDS at 65∞C for 30 min andsubjected to autoradiography.
Analysis of expression of MptpB in M. tuberculosis
To analyse the expression of MptpB in wild type, mutant andcomplemented strains of M. tuberculosis, the bacteria wasgrown in MB7H9 medium supplemented with 1¥ ADC enrich-ment, 0.5% glycerol and 0.2% Tween-80. All mycobacterialcultures were grown at 37∞C to mid-log phase. The cells weresolubilized in SDS sample buffer and approximately 40 mgwhole cell lysate was electrophoresed on 10% SDS–PAGEand transferred to nitrocellulose membrane (AmershamPharmacia Biotech.). The membrane was blocked for 2 h in3% BSA and then incubated for 2 h with polyclonal seraraised against MptpB (1:10 000) in rabbit. Thereafter themembrane was washed thrice with PBS supplemented with0.05% Tween-20 (PBST) and then incubated for 1 h withHRP-conjugated goat anti-rabbit IgG (1:2500, JacksonImmuno Research Laboratories, Inc.). The membrane wasthen washed thrice with PBST and the expression was anal-ysed using Di-amino Benzidene and H2O2.
Morphological studies of bacilli
Smears prepared from single cell suspensions of the cul-tured wild-type and mutant strains were stained withauramine (Blair et al., 1969). Confocal images of the bacilliwere obtained by scanning with a Zeiss 510 laser scanningmicroscope using 100¥ oil immersion lens. The imageswere then imported into Micro Image (Olympus Optical Co.,Version 4.0) and morphometric measurements of at least300 bacilli each of the mutant and wild-type strains weremade.

760 R. Singh et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
Preparation of strains for infection studies
Wild-type, mutant and complemented strains of M. tubercu-losis were grown to mid-log phase in MB7H9 media at 37∞C.The bacilli were harvested by centrifugation at 5000 g for10 min, washed twice with 1¥ PBS, resuspended in 1/10th ofthe culture volume and stored as 1 ml aliquots at -70∞C untiluse. Viable count of bacteria was calculated by plating vari-ous dilutions in duplicates on MB7H10 and incubating theplates at 37∞C for 3 weeks.
In vitro infection of the mouse macrophage cell line by wild-type and mutant M. tuberculosis strains
J774A.1 mouse macrophage cell line resting or activated withrIFN-g (50Uml-1, 16 h) was seeded in a six-well plate (NalgeNunc International) at a density of 2 ¥ 105 per well. Beforeinfection, the cells were washed once with 1¥ Hanks’ bal-anced salt solution (HBSS, GibcoTM Invitrogen Corporation)and medium was replaced with Dulbecco’s modified Eagle’smedium (DMEM, GibcoTM Invitrogen Corporation) supple-mented with 10% heat inactivated fetal calf serum (FCS,GibcoTM Invitrogen Corporation). The bacterial strains werewashed twice with DMEM and resuspended in DMEMsupplemented with 5% FCS. The cells were infected withwild-type and mutant strain at a MOI of 1:10 (macrophage/bacteria). The cells were incubated at 37∞C in a 5% CO2
atmosphere. After 6 h of infection, cells were washed twicewith 1¥ HBSS and then with 2 ml of DMEM supplementedwith FCS (10%), Antibiotic-antimycotic (1%, GibcoTM Invitro-gen Corporation) and amikacin (20 mg ml-1). On days 0, 2, 4,6 and 8, infected cells were lysed in 1 ml of 0.1% Triton X-100 for 15 min. The number of bacilli at different time pointswas determined by plating 10-fold serial dilutions in dupli-cates on MB 7H10 medium and incubating the plates at 37∞Cfor 3 weeks.
Virulence studies in guinea pigs
Guinea pigs (Hartley strain) weighing 200–400 g were pur-chased from National Center for Laboratory Animal Science(NCLAS), Hyderabad, India. Guinea pigs in groups of eightanimals were infected with 5 ¥ 105 cfu of either the wild-typeor the mptpB mutant strain or the complemented strain of M.tuberculosis subcutaneously. The animals (eight animals pergroup at each time point) were euthanized at 3 or 6 weekspost-infection. The visible lesions in liver, spleen, lung andlymph glands were scored according to Mitchison’s scoringsystem shown in Table 2 (Mitchison, 1964). To estimate theviable bacilli count, the spleens were homogenized in 5 ml ofsterile water. Bacteria in the infected spleens were enumer-ated by plating 10-fold serial dilutions of the homogenates induplicate on Lowenstein–Jensen (LJ) slants and incubatingthe slants at 37∞C for 4 weeks. The total organ score and cfudata are expressed as the mean ± standard error obtained ina group of eight animals.
For histopathological analysis, at each time point of sacri-fice, portions of liver and lung were removed aseptically andfixed in 10% formalin. The tissue was embedded in paraffinwax and cut into five micron sections using a microtome. The
sections were stained with haematoxylin and eosin and sub-jected to histopathological analysis. In infected tissues, thepercentage of granuloma and cellular composition of a gran-uloma were microscopically accessed as described earlier(Jayashankar and Ramanathan, 1999).
Statistical analysis
The Student’s unpaired t-test was applied for statistical eval-uation of the data assuming normal distribution. A P-value of<0.05 was considered as a statistically significant observa-tion for any parameter.
Acknowledgements
This work was supported by a grant from the Indian Councilof Medical Research, India. R.S., V.R. and N.D. are thankful
Table 2. Virulence scoring chart for infected organs.
Characteristics Score
SpleenHeavy involvement with numerous large tubercles and
areas of necrosis40
Moderate involvement with numerous small tuberclesor markedly enlarged spleen plus numerous small tubercles
30
Scanty involvement with few large tubercles or numer-ous small but easily visible tubercles
20
Minimal involvement with one or two large tubercles or moderate number of just visible tubercles
10
LiverHeavy involvement with numerous large tubercles and
areas of necrosis30
Moderate involvement with moderate number of large tubercles or numerous small tubercles, no areas of necrosis
23
Scanty involvement with scanty tubercles, easily visible 15Minimal involvement with just visible tubercles 8
LungHeavy involvement with numerous large tubercles
measuring 3–5 mm in diameter20
Moderate involvement with occasional large tubercles (3–5 mm) or more numerous small tubercles(1–2 mm)
15
Scanty involvement up to four large tubercles or number of small tubercles
10
Minimal involvement with small scanty tubercles 5Secondary lymph glandsAbscess at the site of injection plus enlargement plus
caseation of inguinal gland and sublumbar/coelic10
Abscess at the site of injection plus enlargement plus caseation of inguinal gland, no sublumbar or coelic involvement
8
Abscess at the site of injection plus enlargement but no caseation
OrNo abscess at the site of injection but caseation of
inguinal gland
6
Abscess at the site of injection, but no enlargement or caseation
5
Enlargement but no caseation or no abscess at the site of injection
2
The above virulence scoring chart is based on Mitchison’s scoringsystem (Mitchison, 1964).

Role of mptpB in the virulence of mycobacteria 761
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
to the University Grants Commission, India and A.K., A.S.are thankful to the Council of Scientific and IndustrialResearch, India for research fellowships. We thank Dr J. S.Tyagi for critically reading the manuscript. Bindu Nair isacknowledged for technical assistance. Rajiv Chawla isacknowledged for efficient preparation of the manuscript. Wethank Jaya Gopinath, S. Nambirajan, K. Chandran and M.Asokan for their technical assistance.
References
Adams, D.O., and Hamilton, T. A. (1984) The cell biologyof macrophage activation. Annu Rev Immunol 2: 283–318.
Andersson, K., Carballeira, N., Magnusson, K.E., Persson,C., Stendahl, O., Wolf-Watz, H., and Fallman, M. (1996)YopH of Yersinia pseudotuberculosis interrupts early phos-photyrosine signalling associated with phagocytosis. MolMicrobiol 20: 1057–1069.
Armstrong, J., and D’Arcy, H.P. (1971) Response of culturedmacrophages to Mycobacterium tuberculosis with obser-vations on fusion of lysosomes with phagosomes. J ExpMed 134: 713–740.
Av-Gay, Y., Jamil, S., and Drews, S.J. (1999) Expression andcharacterization of the Mycobacterium tuberculosis serine/ threonine protein kinase PknB. Infect Immun 67: 5676–5682.
Black, D.S., and Bliska, J.B. (1997) Identification of p130cas
as a substrate of Yersinia YopH (Yop51), a bacterial pro-tein tyrosine phosphatase that translocates into mamma-lian cells and targets focal adhesions. EMBO J 16: 2730–2744.
Blair, E.B., Weiser, O.L., and Tull, R.H. (1969) Microbacteri-ology Laboratory Methods – Laboratory Report no. 323.Denver: CO: US Army Medical Research and NutritionLaboratory, Fitzsimmoun General Hospital.
Bliska, J.B., and Black, D.S. (1995) Inhibition of the Fcreceptor-mediated oxidative burst in macrophages by theYersinia pseudotuberculosis tyrosine phosphatase. InfectImmun 63: 681–685.
Bliska, J.B., Guan, K., Dixon, J. E., and Falkow, S. (1991)Tyrosine phosphatase hydrolysis of host proteins by anessential Yersinia virulence determinant. Proc Natl AcadSci USA 88: 1187–1191.
Chaba, R., Raje, M., and Chakraborti, P.K. (2002) Evidencethat a eukaryotic-type serine/threonine protein kinase fromMycobacterium tuberculosis regulates morphologicalchanges associated with cell division. Eur J Biochem 269:1078–1085.
Cole, S.T., Brosch, R., Parkhill, J., Garnier, T., Churcher, C.,Harris, D., et al. (1998) Deciphering the biology ofMycobacterium tuberculosis from the complete genomesequence. Nature 393: 537–544.
Dasgupta, S.K., Jain, S., Kaushal, D., and Tyagi, A.K. (1998)Expression systems for study of mycobacterial gene regu-lation and development of recombinant BCG vaccines. Bio-chem Biophys Res Commun 246: 797–804.
Devinney, R., Mortimer, O.S., and Finlay, B.B. (2000) Phos-phatases and kinases delivered to the host cell by bacterialpathogens. Trends Microbiol 30: 29–34.
Dolin, P.J., Raviglione, M.C., and Kochi, A. (1994) Globaltuberculosis incidence and mortality during 1990–2000.Bull WHO 72: 213–220.
Dussurget, O., Stewart, G., Neyrolles, O., Pescher, P.,Young, D., and Marchal, G. (2001) Role of Mycobacteriumtuberculosis Copper-Zinc Superoxide dismutase. InfectImmun 69: 529–533.
Fallman, M., Andersson, K., Hakansson, S., Magnusson,K.E., Stendahl, O., and Wolf-Watz, H. (1995) Yersiniapseudotuberculosis inhibits Fc receptor mediated phago-cytosis in J774 cells. Infect Immun 63: 3117–3124.
Ferrari, G., Langen, H., Naito, M., and Pieters, J. (1999) Acoat protein on phagosomes involved in the intracellularsurvival of mycobacteria. Cell 97: 435–447.
Fu, Y., and Galan, J.E. (1999) A Salmonella protein antago-nizes Rac-1 and Cdc42 to mediate host-cell recovery afterbacterial invasion. Nature 401: 293–297.
Galyov, E.E., Hakansson, S., Forsberg, A., and Wolf-Watz,H. (1993) A secreted protein kinase of Yersinia pseudotu-berculosis is an indispensable virulence determinant.Nature 361: 730–732.
Guan, K., and Dixon, J.E. (1990) Protein tyrosine phos-phatase activity of an essential virulence determinant inYersinia. Science 249: 553–559.
Hinds, J., Mahenthiralingam, E., Kempsell, K.E., Duncan, K.,Stokes, R.W., Parish, T., and Stoker, N.G. (1999)Enhanced gene replacement in mycobacteria. Microbiol-ogy 145: 519–527.
Jayashankar, K., and Ramanathan, V.D. (1999) Biochemicaland histochemical changes relating to fibrosis followinginfection with Mycobacterium tuberculosis in the guineapig. Indian J Med Res 110: 91–97.
Kaniga, K., Uralil, J., Bliska, J.B., and Galan, J.E. (1996) Asecreted tyrosine phosphatase with modular effectordomains in bacterial pathogen Salmonella typhimurium.Mol Microbiol 21: 633–641.
Kennelly, P.J., and Potts, M. (1996) Fancy meeting you here!A fresh look at ‘prokaryotic’ protein phosphorylation. J Bac-teriol 178: 4759–4764.
Koul, A., Choidas, A., Treder, M., Tyagi, A.K., Drlica, K.,Singh, Y., and Ullrich, A. (2000) Cloning and characteriza-tion of secretory tyrosine phosphatase of Mycobacteriumtuberculosis. J Bacteriol 182: 5425–5432.
Koul, A., Choidas, A., Tyagi, A.K., Drlica, K., Singh, Y., andUllrich, A. (2001) Serine/threonine protein kinases PknFand PknG of Mycobacterium tuberculosis: characterizationand localization. Microbiology 147: 2307–2314.
Lowrie, D.B. (1983) How macrophages kill tubercle bacilli. JMed Microbiol 16: 1–14.
Manca, C., Paul, S., Barry, C.E., III, Freedman, V.H., andKaplan, G. (1999) M. tuberculosis catalase and peroxidaseactivities and resistance to oxidative killing in human mono-cytes in vitro. Infect Immun 67: 74–79.
Mitchison, D.A. (1964) The virulence of tubercle bacilli frompatients with pulmonary tuberculosis in India and othercountries. Bull Int Union Against Tuberculosis 35: 287.
Peirs, P., De Wit, L., Braibant, M., Huygen, K., and Con-tent, J. (1997) A serine/threonine protein kinase fromMycobacterium tuberculosis. Eur J Biochem 244: 604–612.
Pelicic, V., Reyrat, J.M., and Gicquel, B. (1996) Expression

762 R. Singh et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 751–762
of the Bacillus subtilus sacB gene confers sucrose sensi-tivity on mycobacteria. J Bacteriol 178: 1197–1199.
Ruan, J., John, G., Ehrt, S., Riley, L., and Nathan, C. (1999)noxR3, a novel gene from Mycobacterium tuberculosis pro-tects Salmonella typhimurium from nitrosative and oxida-tive stress. Infect Immun 67: 3276–3283.
Sathish, M., and Shinnick, T.M. (1994) Identification of genesinvolved in resistance of mycobacteria to killing by mac-rophages. Ann New York Acad Sci 730: 26–36.
Sherman, D.R., Mdluli, K., Hickey, M.J., Arain, T.M.,Morris, S.L., Barry, C.E., III, and Stover, C.K. (1996)Compensatory ahpC gene expression in isoniazid resis-tance Mycobacterium tuberculosis. Science 272: 1641–1643.
Sibille, Y., and Reynolds, H.Y. (1990) Macrophages and poly-
morphonuclear neutrophils in lung defense and injury. AmRev Respir Dis 141: 471–501.
Southern, E.M. (1975) Detection of specific sequencesamong DNA fragments separated by gel electrophoresis.J Mol Biol 98: 503.
Stone, R.L., and Dixon, J.E. (1994) Protein tyrosine phos-phatases. J Biol Chem 269: 31323–31326.
Sturgill-Koszycki, S., Schlesinger, P.H., Chakraborty, P.,Haddix, P.L., Collins, H.L., Fok, A.K., et al. (1994) Lack ofacidification in Mycobacterium phagosomes produced byexclusion of the vesicular proton-ATPase. Science 263:678–681.
Sun, H., and Tonks, N.K. (1994) The coordinated action ofprotein tyrosine phosphatases and kinases in cell signal-ling. Trends Biochem Sci 19: 480–485.
Related Documents











