Copyright Ó 2006 by the Genetics Society of America DOI: 10.1534/genetics.106.058115 Discovery and Mapping of Wheat Ph1 Suppressors J. Dvorak, 1 K. R. Deal and M.-C. Luo Department of Plant Sciences, University of California, Davis, California 95616 Manuscript received March 10, 2006 Accepted for publication May 5, 2006 ABSTRACT Pairing between wheat (Triticum turgidum and T. aestivum) homeologous chromosomes is prevented by the expression of the Ph1 locus on the long arm of chromosome 5B. The genome of Aegilops speltoides suppresses Ph1 expression in wheat 3 Ae. speltoides hybrids. Suppressors with major effects were mapped as Mendelian loci on the long arms of Ae. speltoides chromosomes 3S and 7S. The chromosome 3S locus was designated Su1-Ph1 and the chromosome 7S locus was designated Su2-Ph1. A QTL with a minor effect was mapped on the short arm of chromosome 5S and was designated QPh.ucd-5S. The expression of Su1-Ph1 and Su2-Ph1 increased homeologous chromosome pairing in T. aestivum 3 Ae. speltoides hybrids by 8.4 and 5.8 chiasmata/cell, respectively. Su1-Ph1 was completely epistatic to Su2-Ph1, and the two genes acting together increased homeologous chromosome pairing in T. aestivum 3 Ae. speltoides hybrids to the same level as Su1-Ph1 acting alone. QPh.ucd-5S expression increased homeologous chromosome pairing by 1.6 chiasmata/cell in T. aestivum 3 Ae. speltoides hybrids and was additive to the expression of Su2-Ph1. It is hypothesized that the products of Su1-Ph1 and Su2-Ph1 affect pairing between homeologous chromosomes by regulating the expression of Ph1 but the product of QPh.ucd-5S may primarily regulate recombination between homologous chromosomes. C ROSSING over between homeologous chromo- somes in allotetraploid wheat (Triticum turgidum, genomes AABB) and allohexaploid wheat (T. aestivum AABBDD) is prevented by the expression of the Ph1 locus on the long arm of chromosome 5B (Okamoto 1957; Riley and Chapman 1958; Sears and Okamoto 1958; Riley 1960). This locus allows crossing over to take place only between homologous chromosomes. Because the Ph1 locus is in the B genome, it was logical to expect that it was contributed to polyploid wheats by the diploid source of the B genome, which was most likely an extinct close relative of Aegilops speltoides (2n ¼ 2x ¼ 14, genomes SS) (Sarkar and Stebbins 1956; Dvorak and Zhang 1990; Dvorak 1998). The Ae. speltoides genome was therefore expected to compen- sate for the absence of Ph1 in T. aestivum 3 Ae. speltoides hybrids nullisomic for chromosome 5B. Surprisingly, high levels of homeologous chromosome pairing rather than a suppression were observed in the hybrids, not only in the absence of chromosome 5B but also in its presence (Riley 1960). Ae. speltoides is naturally vari- able for the ability to elicit homeologous chromosome pairing in hybrids with wheat (Dvorak 1972; Kimber and Athwal 1972). A virtual continuum of chromosome- pairing levels, ranging from a few to 16 chiasmata/ metaphase I (MI) cell, has been observed in various T. aestivum 3 Ae. speltoides hybrids, strongly suggesting that the trait is under complex genetic control in the Ae. speltoides genome (Dvorak 1972). Segregation analyses of crosses between various Ae. speltoides strains suggested that the trait is controlled by a minimum of two unlinked major loci interacting in a duplicate manner and an unknown number of minor genes (Chen and Dvorak 1984). Feldman and Mello-Sampayo (1967) suggested that Ae. speltoides affects homeologous chromosome pairing in wheat 3 Ae. speltoides hybrids by promoting homeo- logous chromosome pairing per se. Dover and Riley (1977) pointed out that variation in homeologous chromosome pairing in T. aestivum 3 Ae. speltoides hybrids occurs only if Ph1 is present and therefore pro- posed that the Ae. speltoides genome affects homeolo- gous chromosome pairing by suppressing the activity of Ph1. If the Ae. speltoides genome affects homeologous chro- mosome pairing by suppressing the expression of Ph1, then Ae. speltoides genotypes that bring about different levels of homeologous chromosome pairing in wheat 3 Ae. speltoides hybrids would bring about the same level of homeologous pairing in hybrids with species that do not show a Ph1-like activity. To test this hypothesis, Ae. speltoides lines differing greatly in the ability to elicit homeologous chromosome pairing in wheat 3 Ae. speltoides hybrids were crossed with diploid Ae. tauschii and Ae. caudata, which do not show a Ph1-like activity. No meaningful differences between the hybrids were found, indicating that Ae. speltoides genes act on the Ph1 locus (Chen and Dvorak 1984). 1 Corresponding author: Department of Plant Sciences, University of California, Davis, CA 95616. E-mail: [email protected] Genetics 174: 17–27 (September 2006)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Copyright � 2006 by the Genetics Society of AmericaDOI: 10.1534/genetics.106.058115
Discovery and Mapping of Wheat Ph1 Suppressors
J. Dvorak,1 K. R. Deal and M.-C. Luo
Department of Plant Sciences, University of California, Davis, California 95616
Manuscript received March 10, 2006Accepted for publication May 5, 2006
ABSTRACT
Pairing between wheat (Triticum turgidum and T. aestivum) homeologous chromosomes is prevented bythe expression of the Ph1 locus on the long arm of chromosome 5B. The genome of Aegilops speltoidessuppresses Ph1 expression in wheat 3 Ae. speltoides hybrids. Suppressors with major effects were mapped asMendelian loci on the long arms of Ae. speltoides chromosomes 3S and 7S. The chromosome 3S locus wasdesignated Su1-Ph1 and the chromosome 7S locus was designated Su2-Ph1. A QTL with a minor effect wasmapped on the short arm of chromosome 5S and was designated QPh.ucd-5S. The expression of Su1-Ph1and Su2-Ph1 increased homeologous chromosome pairing in T. aestivum 3 Ae. speltoides hybrids by 8.4 and5.8 chiasmata/cell, respectively. Su1-Ph1 was completely epistatic to Su2-Ph1, and the two genes actingtogether increased homeologous chromosome pairing in T. aestivum 3 Ae. speltoides hybrids to the samelevel as Su1-Ph1 acting alone. QPh.ucd-5S expression increased homeologous chromosome pairing by1.6 chiasmata/cell in T. aestivum 3 Ae. speltoides hybrids and was additive to the expression of Su2-Ph1.It is hypothesized that the products of Su1-Ph1 and Su2-Ph1 affect pairing between homeologouschromosomes by regulating the expression of Ph1 but the product of QPh.ucd-5S may primarily regulaterecombination between homologous chromosomes.
CROSSING over between homeologous chromo-somes in allotetraploid wheat (Triticum turgidum,
genomes AABB) and allohexaploid wheat (T. aestivumAABBDD) is prevented by the expression of the Ph1locus on the long arm of chromosome 5B (Okamoto
1957; Riley and Chapman 1958; Sears and Okamoto
1958; Riley 1960). This locus allows crossing over totake place only between homologous chromosomes.Because the Ph1 locus is in the B genome, it was logicalto expect that it was contributed to polyploid wheats bythe diploid source of the B genome, which was mostlikely an extinct close relative of Aegilops speltoides (2n ¼2x ¼ 14, genomes SS) (Sarkar and Stebbins 1956;Dvorak and Zhang 1990; Dvorak 1998). The Ae.speltoides genome was therefore expected to compen-sate for the absence of Ph1 in T. aestivum 3 Ae. speltoideshybrids nullisomic for chromosome 5B. Surprisingly,high levels of homeologous chromosome pairing ratherthan a suppression were observed in the hybrids, notonly in the absence of chromosome 5B but also in itspresence (Riley 1960). Ae. speltoides is naturally vari-able for the ability to elicit homeologous chromosomepairing in hybrids with wheat (Dvorak 1972; Kimber andAthwal 1972). A virtual continuum of chromosome-pairing levels, ranging from a few to �16 chiasmata/metaphase I (MI) cell, has been observed in various T.aestivum 3 Ae. speltoides hybrids, strongly suggesting that
the trait is under complex genetic control in the Ae.speltoides genome (Dvorak 1972). Segregation analysesof crosses between various Ae. speltoides strains suggestedthat the trait is controlled by a minimum of twounlinked major loci interacting in a duplicate mannerand an unknown number of minor genes (Chen andDvorak 1984).
Feldman and Mello-Sampayo (1967) suggested thatAe. speltoides affects homeologous chromosome pairingin wheat 3 Ae. speltoides hybrids by promoting homeo-logous chromosome pairing per se. Dover and Riley
(1977) pointed out that variation in homeologouschromosome pairing in T. aestivum 3 Ae. speltoideshybrids occurs only if Ph1 is present and therefore pro-posed that the Ae. speltoides genome affects homeolo-gous chromosome pairing by suppressing the activity ofPh1.
If the Ae. speltoides genome affects homeologous chro-mosome pairing by suppressing the expression of Ph1,then Ae. speltoides genotypes that bring about differentlevels of homeologous chromosome pairing in wheat 3
Ae. speltoides hybrids would bring about the same levelof homeologous pairing in hybrids with species that donot show a Ph1-like activity. To test this hypothesis, Ae.speltoides lines differing greatly in the ability to elicithomeologous chromosome pairing in wheat 3 Ae.speltoides hybrids were crossed with diploid Ae. tauschiiand Ae. caudata, which do not show a Ph1-like activity. Nomeaningful differences between the hybrids were found,indicating that Ae. speltoides genes act on the Ph1 locus(Chen and Dvorak 1984).
1Corresponding author: Department of Plant Sciences, University ofCalifornia, Davis, CA 95616. E-mail: [email protected]
Genetics 174: 17–27 (September 2006)

A great deal of work has been done on the character-ization of the Ph1 locus and its expression, culminatingwith the recently reported comparative sequencingof .2 Mbp of the Ph1 region (Griffiths et al. 2006).A conclusion was reached that Ph1 activity is synony-mous with the insertion of heterochromatin fromchromosome 3A into the cdc2 multigene locus in thePh1 region on chromosome 5B (Griffiths et al. 2006).
Following the rationale that the isolation and molec-ular characterization of Ae. speltoides Ph1 suppressors willassist with illuminating the molecular basis of thisimportant meiotic mechanism, we embarked on geneticcharacterization and mapping of Ae. speltoides Ph1suppressors, with the ultimate goal of isolating thesegenes. This work was made possible by the developmentof a genetic map of the Ae. speltoides genome (Luo et al.2005). The Ae. speltoides map revealed that recombina-tion is localized in very short distal regions of the Ae.speltoides chromosomes. As a result, most markers clusterinto centromeric blocks with little or no recombinationwhile the distances between distal markers are expanded,often generating separate linkage groups (Luo et al.2005). These characteristics of Ae. speltoides meiosismake it difficult to use molecular markers effectivelyin traditional QTL mapping. We therefore employedseveral strategies with progressively increasing precisionto detect and map loci that interact with the Ph1 locus inthe Ae. speltoides genome.
MATERIALS AND METHODS
Plants and mapping populations: Ae. speltoides population2-12-4, eliciting low homeologous chromosome pairing inhybrids with T. aestivum cv ‘‘Chinese Spring,’’ and populationPI369609-12-II, eliciting high homeologous chromosomepairing in hybrids with Chinese Spring, were used. T. aestivumis a self-pollinating species but Ae. speltoides is a cross-pollinatingspecies. Repeated self-pollination of Ae. speltoides plants re-sulted in sterility of the plants. The original open-pollinatedAe. speltoides accessions were therefore self-pollinated for twogenerations and then maintained as populations of inter-breeding plants. Hybrids of Chinese Spring with the Ae.speltoides population 2-12-4 had �7–9 chiasmata/cell at MIand those with the PI369609-12-II population had �15–16chiasmata/cell at MI (Chen and Dvorak 1984). Haplotypescontributed by the 2-12-4 population are designated asa-haplotypes and those contributed by the PI369609-12-IIpopulation are designated as b-haplotypes.
A total of 50 Ae. speltoides F2 plants were crossed with ChineseSpring and from 1 to 29 hybrids were produced per each of the50 F2 plants and grown in a greenhouse. The mean number ofchiasmata per cell was estimated in 15–30 meiocytes in anthersfrom one or two spikes. Mean chiasma number per cell wascomputed for each hybrid and each family of hybrids. These50 F2 plants were also among the 86 F2 plants used for theconstruction of the Ae. speltoides genetic map (Luo et al. 2005).
An additional 30 F2 plants were grown and crossed withChinese Spring. From 2 to 154 hybrids/family were grown inthe greenhouse. The mean number of chiasmata per cell wasdetermined in 15–30 meiocytes for each hybrid. DNAs wereisolated from 590 hybrids and Southern blots were prepared
and hybridized with 35 cloned DNA fragments. The associa-tion of QTL with marker loci detected by these DNA fragmentswas tested by partitioning the population into a- and b-hap-lotype subpopulations and testing the difference betweenmean phenotypic scores of the subpopulations with one-wayanalyses of variance (ANOVA). These analyses indicated theapproximate locations of the QTL. For further analyses, 317hybrids derived from 19 F2 plants heterozygous at loci as-sociated with each QTL were selected. This population wasused to estimate the effect of the substitution of the b- for thea-haplotype at each of the 35 marker loci in the backgroundof low-homeologous-chromosome-pairing haplotypes at QTLloci detected in the previous analyses.
Mapping of major loci: To locate major loci suppressing Ph1on the Ae. speltoides genetic map, progeny of the Ae. speltoides F2
plant 125, which was heterozygous at RFLP markers associatedwith all QTL detected, were grown in isolation. DNAs wereisolated from F3 plants, desired genotypes were selected on thebasis of RFLP, and plants were grown in isolation. The sameprocess was repeated in the F4 generation. Plants segregatingat markers associated with a single QTL, but homozygous forlow-pairing haplotypes at loci associated with the remainingQTL, were crossed with Chinese Spring. Numbers of chias-mata per cell were determined using from two to five spikesper hybrid as replicates. DNAs were isolated from the hybridsand used in RFLP mapping of Ph1 suppressors relative tomolecular markers.
One of the Ph1 suppressing loci was mapped near Xpsr1205on chromosome 3S. After the first round of mapping, thelocus was flanked by wheat EST markers XBF497740 andXBE488620. To map this Ph1 suppressor relative to other mo-lecular markers on the Ae. speltoides genetic map, five Ae.speltoides F4 plants heterozygous at the Xpsr1205, XBF497740,and XBE488620 loci, but homozygous for low-pairing haplo-types at QTL on other chromosomes, were crossed withChinese Spring and 269 hybrids were grown. DNAs wereisolated from each plant using the rapid DNA isolationtechnique described below. XBF497740 and XBE488620 gen-otypes were determined using dominant PCR assays. Xpsr1205genotypes were determined with a TaqMan assay. To detect Ae.speltoides DNA fragments in Chinese Spring 3 Ae. speltoideshybrids, PCR primer pairs that amplified only Ae. speltoidestarget DNA from genomic DNA of Chinese Spring 3 Ae.speltoides hybrids were designed (see below). Hybrids thatharbored chromosomes that had undergone recombinationin the XBF497740–XBE488620 interval were kept. At least fiveimmature spikes were harvested for meiotic analyses fromthese plants and DNAs were isolated from the remaining plantmaterial for RFLP.
Interspecific hybridization: Chinese Spring and Ae. speltoidesplants were grown in clay pots in the greenhouse. ChineseSpring flowers were manually emasculated and hand polli-nated with Ae. speltoides pollen 2 days after emasculation andthen repollinated the next day. Virtually all hybrid seeds wereshriveled. They were sterilized with a 0.5 dilution of commer-cial bleach in distilled water for 15 min, thoroughly rinsed indistilled water, and dried for 1 min on sterile filter paper. Theywere then placed on sterile filter paper moistened with a sterilesolution containing 100 mg/liter of ampicillin, 60 g/litersucrose, and 400 mg/liter glutamine. Petri dishes were kept at5� in a refrigerator for 5 days. Seeds were transferred into freshpetri dishes with filter papers moistened with the abovesolution and germinated at 26�. Seedlings were transplanteddirectly into Jiffy peat pots or clay pots in the greenhouse.
Meiotic analyses: Immature spikes were collected fromgreenhouse-grown plants and fixed in a freshly preparedsolution of 6 ethanol:3 chloroform:1 glacial acetic acid, v/v,for 24 hr at room temperature. Spikes were rinsed in 70%
18 J. Dvorak, K. R. Deal and M.-C. Luo

ethanol and stored in 70% ethanol in refrigerator untilanalyzed. Squashes of meiocytes were prepared with thestandard aceto-carmine method. Slides were systematicallyscanned under the microscope and the numbers of chiasmatawere determined in 10–30 meiocytes per spike. Only anthersin which the majority of meiocytes were at MI were used.
DNA isolation and DNA hybridization: A rapid DNAisolation technique was employed for PCR-based assays (J. L.Halverson and A. Van Deynze, personal communication). A6-cm2 segment of seedling leaf was placed into each well of a96-well flat-bottom block (QIAGEN, Valencia, CA, catalog no.19579). Two 8-mm stainless-steel grinding lapidary ball/coneswere inserted into each well, and the plate was heat sealed andfrozen at�80 for at least 3 hr. A hole was punched through theseal of each well with a 12-gauge needle, and the whole platewas lyophilized overnight. The plate was sealed with a platetape (QIAGEN, catalog no. 19570) immediately after lyophi-lization, and the samples were ground by shaking on acommercial paint shaker in two orientations for 30 sec each.Holes were punched through the film covering each well witha 12-gauge needle and widened with the conical bottoms of aPCR plate. One milliliter of fresh lysis buffer containing 500mm NaCl, 100 mm Tris pH 7.2, 50 mm EDTA, 0.5% sodiumbisulfite, 0.1% diethyldithiocarbamic acid, 0.1% ascorbic acid,and 2% polyvinylpolypyrillidone was added to each well whilehand agitating the solution. The plate was sealed with platetape and inverted several times until the plant material wasuniformly wetted. A hole was punched into each well using an18-gauge needle and the plate was incubated at 65� for 45 min.The tape and foil were carefully removed from the plate. Atotal of 800 ml of solution was transferred with a 1000-ml multi-channel pipette from each well into a round-bottom 96-wellblock and sedimented at 2000 3 g for 10 min. A total of 400 mlof supernatant was transferred from each well into a newround-bottom 96-well block, making sure that no particulatematter was transferred, 400 ml isopropanol was added, thewells were sealed with tape, and the plate was inverted fourtimes. DNAs were sedimented at 3000 rpm for 10 min,supernatant was poured off, and the pellet was rinsed with70% ethanol, air dried for 30 min, and dissolved in 200 ml of 10mm TRIS and 1 mm EDTA. A total of 3 ml of the DNA solutionwas used per PCR reaction.
For RFLP, nuclear DNA was isolated as described by Dvorak
et al. (1988). To prepare Southern blots, DNAs were digestedwith restriction endonucleases and Southern blots werehybridized as described earlier (Dvorak et al. 2004).
Design of Ae. speltoides genome-specific primers for ESTloci and TaqMan assay:
Sequence information for wheat ESTs BE497740 andBE442875 was obtained from the wheat EST database(http://wheat.pw.usda.gov/NSF/). Homology was searchedwith these sequences in the rice genomic sequence usingBLASTN at the NCBI (http://www.ncbi.nlm.nih.gov/) site.Intron/exon junctions were identified in the rice genomicsequence and primers were designed using the wheat ESTsequence to flank an intron. Genomic DNAs of Ae. speltoides (F2
family no. 134), Ae. tauschii (AL8/78 and AS75), T. urartu(G1812 and DV867), T. dicoccoides (PI428094), and ‘‘Langdon’’durum wheat were used as templates in PCR. Amplicons fromT. dicoccoides and Langdon were TA cloned (pGEM-T vectorsystem, Promega, Madison, WI). Amplicons were sequencedin both directions using PCR primers as DNA-sequencingprimers. Sequences were aligned and compared. Nucleotidepositions at which the Ae. speltoides nucleotide sequencediffered from the T. urartu, Ae. tauschii, and wheat A- andB-genome nucleotide sequences were used to design Ae.speltoides genome-specific primers. Their specificity was testedusing genomic DNAs of wheat/Ae. speltoides disomic additionlines (Friebe et al. 2000) and 10 Chinese Spring 3 Ae. speltoideshybrids, which had shown contrasting RFLP haplotypes at theXBE497740, XBE442875, and Xpsr1205 loci. Ae. speltoidesgenome-specific PCR primers were developed for all threemarkers (Table 1). Those for the XBE497740 and XBE442875loci fortuitously also differentiated between the Ae. speltoidesa- and b-haplotypes, providing an amplicon presence vs. ab-sence PCR-genotyping test. PCR primers for Xpsr1205 (Table1) showed no such differentiation. PCR amplicons of Ae.speltoides Xpsr1205a and Xpsr1205b RFLP haplotypes were se-quenced and sequences were compared. The differencesbetween the Xpsr1205a and Xpsr1205b sequences were ex-ploited for the design of TaqMan primers and probes (Table2). PCR primers and TaqMan probes were designed by AppliedBioSystems (Foster City, CA).
PCR conditions: The following conditions were used for PCRamplification of DNA from the Xpsr1205 and XBE442875 loci:96� for 5 min, 10 cycles of 94� for 30 sec, 59� for 30 sec, and 72�for 2 min, followed by 10 cycles of 94� for 30 sec, 57� for 30 sec,and 72� for 2 min, followed by 20 cycles of 94� for 30 sec, 56� for30 sec, and 72� for 2 min, with a final extension at 72� for 5 min.
The conditions for PCR amplification of DNA from theXBE497740 locus were 96� for 5 min, followed by 10 cycles of94� for 30 sec, 57� for 30 sec, and 72� for 2 min, 10 cycles of 94�for 30 sec, 55� for 30 sec, and 72� for 2 min, and 20 cycles of 94�for 30 sec, 54� for 30 sec, and 72� for 2 min, followed byextension at 72� for 5 min.
For the XBE497740 locus, PCR with Ae. speltoides genome-specific primers (Table 1) generated amplicons only for theXBE497740 b-haplotype, which was 150 bp long. For the
TABLE 1
Ae. speltoides genome-specific PCR primers for amplification of DNA from the indicated loci in wheat 3 Ae.speltoides hybrids
Locus Forward primer Reverse primer
Xpsr1205 CGGCAATGATGAGTGTGTCATT CAACTCCCAGTTTGCTGACAXbe442875 TTAGGTAGTTTTTGTTTTGTTTCATT TGCCCCGAAACAGAGATGXbe497740 CCTGATTACCTCCGTATAAAAAAT TCAGGAGCAATAGTTTTC TTGTCA
TABLE 2
Xpsr1205 PCR primers and TaqMan probe for detecting A/Gsingle nucleotide polymorphism at the Xpsr1205 locus
Primer or probe Dye Nucleotide sequence
PCR forward — GCACCCAGCTTGTATTGATTTCCTPCR reverse — GTGCCCTTCACGCTAGCATaqMan probe 1 VIC CATGACGGAAAGATATTaqMan probe 2 FAM CATGACGGAGAGATAT
FAM, fluorescein phosphoramide.
Wheat Ph1 Suppressors 19

XBE442875 locus, PCR with Ae. speltoides genome-specific prim-ers (Table 1) generated amplicons only for the XBE442875a-haplotype, which was �250 bp long.
Using the Ae. speltoides genome-specific PSR1205 primers(Table 1), an amplicon of �400 bp was produced for bothhaplotypes. The PCR reaction was diluted 1:100 with distilledwater and 1 ml was used as a template for the TaqMan assay.Since the TaqMan forward and reverse primers (Table 2) werenot Ae. speltoides-genome specific, it was important that theamount of genomic DNA present was minimized. The TaqManassay was carried out according to the manufacturer’s (AppliedBioSystems) specifications. ABI Prism 7000 Sequence DetectionSystem (Applied BioSystems) was used for real-time PCR.
Statistical analyses: Restriction fragment length polymor-phism at 137 loci detected by hybridization of 99 cDNA or PstIclones (Luo et al. 2005) in 50 F2 plants was employed in theconstruction of a genetic map using MapMaker 3.0 (Lincoln
et al. 1992) and in correcting segregation data with theKosambi mapping function (Kosambi 1943). MapMakerQTL function was used to compute LOD scores of associationsbetween molecular markers and mean numbers of chiasmatacomputed for each of the 50 families of Chinese Spring 3 Ae.speltoides hybrids.
The statistical significance of differences in the size ofintervals between markers on different genetic maps wastested as follows. Interval lengths in centimorgans wereconverted into recombination fractions. Variances of theestimates were computed according to Allard (1956). Thedifferences in the interval lengths between maps were testedby z-test.
The association between molecular markers and the level ofhomeologous chromosome pairing was estimated in thepopulation of 590 Chinese Spring 3 Ae. speltoides hybrids.Southern blots of DNAs of the 590 hybrids were hybridizedwith clones detecting 35 strategically placed RFLP loci. Allelicvariation at each locus was determined. For each locus, hybridswere grouped into two subpopulations, one containing thea-haplotype and the other the b-haplotype. The differencebetween mean chiasma numbers per cell between the twosubpopulations, F-value, and probability were computed withone-way analysis of variance. Similar analyses of variance wereperformed for groups of selected hybrids as described inresults.
To assess the statistical significance of interactions betweenand among QTL, 3 3 2 factorial ANOVA was performed usingthe mean numbers of chiasmata per cell as variables with theGLM procedure of SAS 9.1.
RESULTS
QTL analysis: Because parental stocks 2-12-4 andPI369609-12-II were not homozygous, only a single Ae.speltoides F1 plant from the cross 2-12-4 3 PI369609-12-IIwas used to produce all mapping populations. Fifty F2
plants were crossed with Chinese Spring, producing 50families of hybrids. The mean numbers of chiasmatawere scored in one to two spikes per hybrid, and familymeans were computed. DNAs were isolated from the 50F2 plants, a genetic map based on 137 RFLP loci wasconstructed, and QTL analysis was performed using thefamily mean numbers of chiasmata per cell as thephenotypic scores. The limited number of families, lowrecombination rates in proximal chromosome regions,and high recombination rates in the distal regions
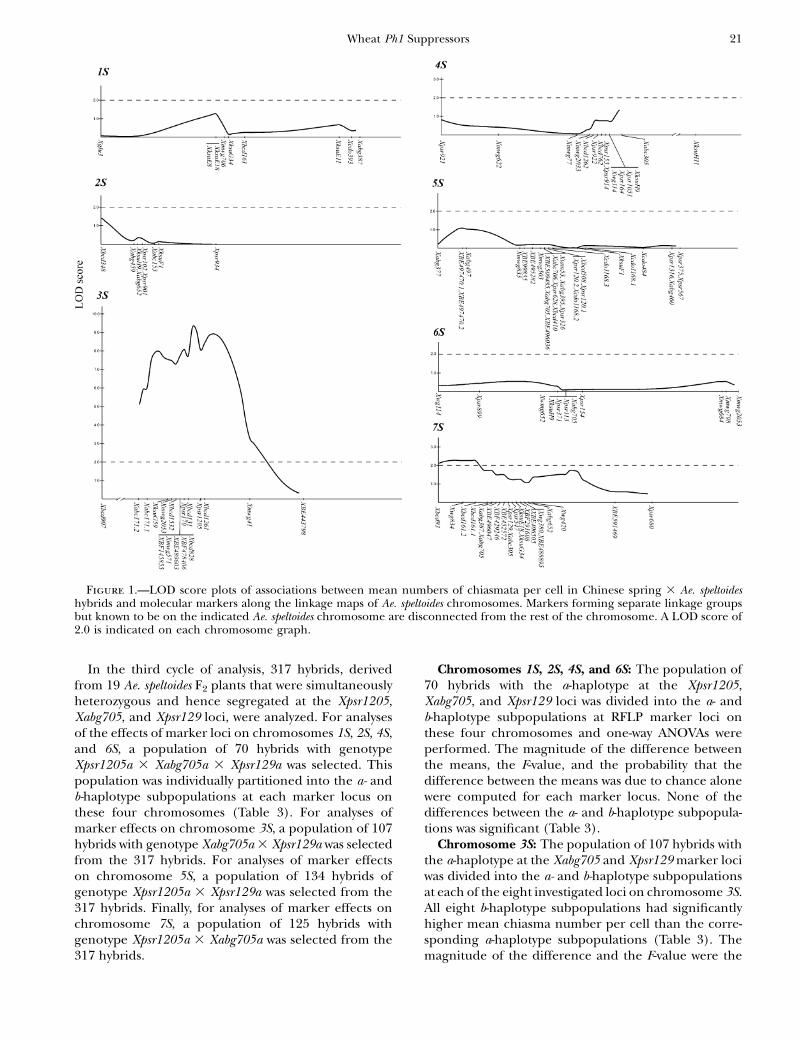
limited the resolution of the analysis. Using a LODscore of 2.0 as the significance boundary, QTL were de-tected on chromosomes 3S and 7S but their locations rel-ative to molecular marker loci were uncertain (Figure 1).
The high-pairing QTL haplotype on chromosome 3Scosegregated with the recessive lig gene for awnlesslateral spikelets that is in the centromeric region of the3S genetic map (Luo et al. 2005). Awnless lig/lig plantswere excluded from being parents in further crosseswith Chinese Spring to minimize the likelihood of in-cluding Ae. speltoides plants homozygous for the high-pairing QTL haplotype in the crosses.
A total of 30 additional F2 Ae. speltoides plants withawned lateral spikelets (Lig/lig or Lig/Lig) were selectedand crossed with Chinese Spring. Mean chiasma num-bers per cell were estimated in 590 hybrids and DNAswere isolated from them. Southern blots were preparedand hybridized with 35 PstI or cDNA clones, eachdetecting a single locus in the Ae. speltoides genome.The positions of the loci on the Ae. speltoides genetic mapwere determined earlier (Luo et al. 2005). The analysisof QTL effects and their location was performed inseveral cycles.
The objective of the first cycle of analysis (not shown)was to identify marker loci associated with major QTL.The population of 590 hybrids was partitioned into thea- and b-haplotype subpopulations at each marker locusand the mean numbers of chiasmata per cell werecomputed for each subpopulation. The largest differ-ences in chiasma number per cell between the a- andb-haplotype subpopulations were observed for themarkers on chromosomes 3S and 7S. On chromosome3S, the largest difference was at the Xpsr1205 locus onthe long arm. On chromosome 7S, the largest differencewas at the Xpsr129 locus on the long arm. High pairingwas associated with the b-haplotype at both marker loci.A total of 23 families comprising 468 hybrids derivedfrom Ae. speltoides parents heterozygous at the Xpsr1205and Xpsr129 loci were used for further analyses.
The objective of the second cycle of analysis was tosearch for minor QTL on the remaining five chromo-somes. A subpopulation of 177 Chinese Spring 3 Ae.speltoides hybrids of genotype Xpsr1205a 3 Xpsr129a(low-pairing haplotype at both loci) was selected andused to search for QTL on Ae. speltoides chromosomes1S, 2S, 4S, 5S, and 6S. For each investigated locus onthese five chromosomes, the population was subdividedinto the a- and b-haplotypes and the probability that thedifference in mean chiasma number per cell between apair of subpopulations was due to chance was estimatedwith one-way ANOVA. Several iterations of this cyclewere performed, each resulting in associating a QTLwith a marker locus with greater precision. In addi-tion to substantiating the association of major QTL withXpsr1205 and Xpsr129 loci, a minor QTL was associ-ated with marker locus Xabg705 in the short arm ofchromosome 5S.
20 J. Dvorak, K. R. Deal and M.-C. Luo
In the third cycle of analysis, 317 hybrids, derivedfrom 19 Ae. speltoides F2 plants that were simultaneouslyheterozygous and hence segregated at the Xpsr1205,Xabg705, and Xpsr129 loci, were analyzed. For analysesof the effects of marker loci on chromosomes 1S, 2S, 4S,and 6S, a population of 70 hybrids with genotypeXpsr1205a 3 Xabg705a 3 Xpsr129a was selected. Thispopulation was individually partitioned into the a- andb-haplotype subpopulations at each marker locus onthese four chromosomes (Table 3). For analyses ofmarker effects on chromosome 3S, a population of 107hybrids with genotype Xabg705a 3 Xpsr129a was selectedfrom the 317 hybrids. For analyses of marker effectson chromosome 5S, a population of 134 hybrids ofgenotype Xpsr1205a 3 Xpsr129a was selected from the317 hybrids. Finally, for analyses of marker effects onchromosome 7S, a population of 125 hybrids withgenotype Xpsr1205a 3 Xabg705a was selected from the317 hybrids.
Chromosomes 1S, 2S, 4S, and 6S: The population of70 hybrids with the a-haplotype at the Xpsr1205,Xabg705, and Xpsr129 loci was divided into the a- andb-haplotype subpopulations at RFLP marker loci onthese four chromosomes and one-way ANOVAs wereperformed. The magnitude of the difference betweenthe means, the F-value, and the probability that thedifference between the means was due to chance alonewere computed for each marker locus. None of thedifferences between the a- and b-haplotype subpopula-tions was significant (Table 3).
Chromosome 3S: The population of 107 hybrids withthe a-haplotype at the Xabg705 and Xpsr129 marker lociwas divided into the a- and b-haplotype subpopulationsat each of the eight investigated loci on chromosome 3S.All eight b-haplotype subpopulations had significantlyhigher mean chiasma number per cell than the corre-sponding a-haplotype subpopulations (Table 3). Themagnitude of the difference and the F-value were the
Figure 1.—LOD score plots of associations between mean numbers of chiasmata per cell in Chinese spring 3 Ae. speltoideshybrids and molecular markers along the linkage maps of Ae. speltoides chromosomes. Markers forming separate linkage groupsbut known to be on the indicated Ae. speltoides chromosome are disconnected from the rest of the chromosome. A LOD score of2.0 is indicated on each chromosome graph.
Wheat Ph1 Suppressors 21

largest at the Xpsr1205 locus in the distal portion of thelong arm. The substitution of the Xpsr1205b haplotypefor the Xpsr1205a haplotype increased homeologouschromosome pairing in Chinese Spring 3 Ae. speltoideshybrids by 8.4 chiasmata/cell.
Chromosome 5S: The population of 134 hybrids withthe a-haplotype at marker loci Xpsr1205 and Xpsr129 waspartitioned into the a- and b-haplotype subpopulationsfor each of the six investigated loci on chromosome 5S.The difference between the a- and b-haplotype subpop-ulations and the F-value reached the largest magnitudeat the Xabg705 and Xpsr628 loci, suggesting that a QTL islocated in the short arm of the chromosome in intervalXabg705–Xpsr628. The substitution of the Xabg705bhaplotype for the Xabg705a haplotype increased homeo-
logous chromosome pairing by 1.6 chiasmata/cell inChinese Spring 3 Ae. speltoides hybrids.
Chromosome 7S: The population of 125 hybrids withgenotype Xpsr1205a and Xpsr129a was divided into thea- and b-haplotype subpopulations at each of the eightinvestigated loci on this chromosome. At all marker loci,except for Xpsr680, the most distal locus on the longarm, the b-haplotype subpopulation had significantlyhigher homeologous chromosome pairing than thecorresponding a-haplotype subpopulation. The magni-tude of the difference between a pair of subpopulationmeans and the F-value were the greatest at Xpsr129. Thesubstitution of the Xpsr129b haplotype for the Xpsr129ahaplotype increased homeologous chromosome pair-ing by 5.8 chiasmata/cell in hybrids.
TABLE 3
The effects of the substitution of the b-haplotype for the a-haplotype at indicated loci on the mean chiasma number per cell insubpopulations of Chinese Spring 3 Ae. speltoides hybrids and one-way ANOVA of the difference between the
means of a-haplotype and b-haplotype subpopulations
Chromosome LocusNo. of hybrids in smaller
subpopulation
b-Haplotype subpopulationmean minus a-haplotype
subpopulation mean F Probability
1 XksuE18 13 �0.5 0.4 0.5131 Xbcd161 20 �1.0 2.7 0.1061 Xcdo393 37 0.6 0.6 0.3082 Xbcd348 32 �0.1 ,0.1 0.8182 Xpsr102 31 �0.1 ,0.1 0.9372 Xbcd111 30 �0.1 ,0.1 0.9112 Xbcd1069 31 �0.1 ,0.1 0.9383 Xabc171 49 5.3 61.3 ,0.0013 XksuG59 35 7.0 131.0 ,0.0013 Xbcd1532 36 6.6 111.8 ,0.0013 Xbcd828 35 7.0 137.8 ,0.0013 Xbcd131 35 7.3 163.9 ,0.0013 Xpsr1205 34 8.4 390.8 ,0.0013 Xbcd1261 35 8.1 315.5 ,0.0013 Xmwg41 41 4.6 37.7 ,0.0014 Xwg622 14 �1.2 1.6 0.2104 Xmwg2033 42 �0.6 1.0 0.3264 Xbcd1262 40 �0.4 0.4 0.5444 XksuH11 33 2.3 1.2 0.7355 XBE497470 33 0.3 0.2 0.6655 Xabg705 62 1.6 6.9 0.0095 Xpsr628 63 1.5 6.9 0.0105 Xbcd508 61 1.3 4.5 0.0355 XksuF1 36 0.7 1.1 0.2955 Xcdo484 13 0.5 0.3 0.6156 Xpsr899 13 1.0 2.7 0.1056 Xpsr113 30 0.8 2.1 0.1537 Xbcd93 17 3.5 25.7 ,0.0017 XBF429246 46 5.4 138.6 ,0.0017 XBF442572 47 5.4 139.2 ,0.0017 Xpsr129 52 5.8 215.0 ,0.0017 Xpsr547 51 5.6 183.5 ,0.0017 Xwg380 48 5.6 168.8 ,0.0017 XBE406505 46 4.8 89.6 ,0.0017 Xpsr680 17 0.2 0.7 0.717
22 J. Dvorak, K. R. Deal and M.-C. Luo
Gene interactions: Interactions between the three QTLwere examined by factorial ANOVA in which Xpsr1205,Xabg705, and Xpsr129 were factors, each having twohaplotype levels. Mean chiasma numbers per cell in 317hybrids belonging to 19 families segregating simulta-neously at the Xpsr1205, Xabg705, and Xpsr129 loci wereused as variables. An R 2 of 0.78 indicated that the effects ofthe three marker loci explained most of the variation inthis population. The effect of each marker locus was highlysignificant (Table 4). Also highly significant were inter-actions between the QTL associated with the Xpsr1205marker and the QTL associated with markers Xabg705 andXpsr129. No interaction was detected between QTLassociated with the latter two markers. Nor did ANOVAindicate a three-way interaction among the loci (Table 4).
The effects of these three QTL on mean chiasmanumber in Chinese Spring 3 Ae. speltoides hybrids aresummarized in Table 5. The background level of pair-ing in a hybrid with the a-haplotype at the Xpsr1205,Xabg705, and Xpsr129 loci was 7.0 chiasmata/cell. Theaverage pairing in 12 hybrids (360 cells) with meannumbers of chiasmata per cell ranging from 6.8 to 7.3was 15.7 univalents 1 5.4 bivalents 1 0.44 trivalents 1
0.05 quadrivalents. The number of trivalents per cellranged from 0 to 4 and the number of quadrivalents percell ranged from 0 to 2.
The chromosome 3S QTL raised the mean number ofchiasmata from the background of 7.0 to 15.4. Combi-nations of the 3S QTL with 5S or 7S QTL had noadditional effects, which is consistent with the factorialANOVA output, indicating a strong interaction betweenthe 3S QTL and those on 5S and 7S. In contrast, theeffects of the 7S and 5S QTL were entirely additive; themean number of chiasmata per cell increased from 12.4to 14. 2. The 5S QTL also showed a borderline additiveeffect in combinations with the 3S QTL and 7S QTL,raising the mean number of chiasmata from 15.4 to16.4/cell (Table 5).
Genetic mapping: Ae. speltoides F4 plant 125-80-13,homozygous for the low-pairing haplotypes on chromo-somes 5S and 7S but heterozygous for marker loci onchromosome 3S, was crossed with Chinese Spring andthe mean numbers of chiasmata per cell were deter-mined in 67 hybrids, analyzing from two to five spikesper hybrid. The frequency distribution of mean chiasmanumbers per cell was clearly bimodal among the
TABLE 4
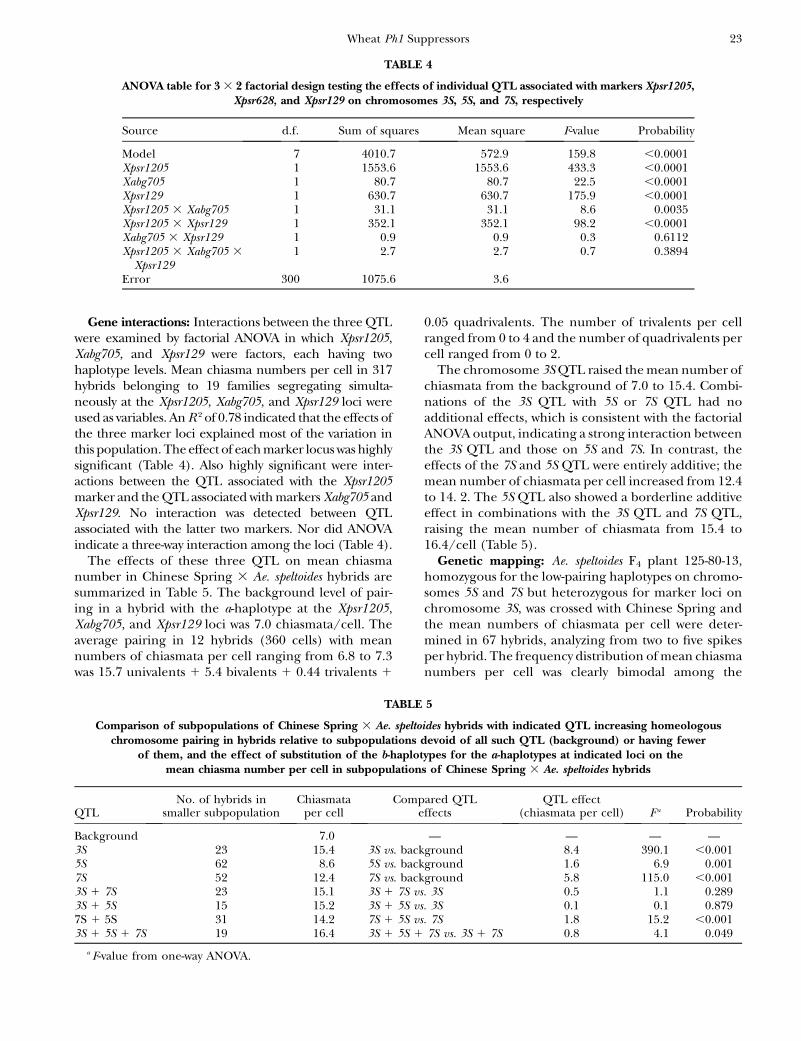
ANOVA table for 3 3 2 factorial design testing the effects of individual QTL associated with markers Xpsr1205,Xpsr628, and Xpsr129 on chromosomes 3S, 5S, and 7S, respectively
Source d.f. Sum of squares Mean square F-value Probability
Model 7 4010.7 572.9 159.8 ,0.0001Xpsr1205 1 1553.6 1553.6 433.3 ,0.0001Xabg705 1 80.7 80.7 22.5 ,0.0001Xpsr129 1 630.7 630.7 175.9 ,0.0001Xpsr1205 3 Xabg705 1 31.1 31.1 8.6 0.0035Xpsr1205 3 Xpsr129 1 352.1 352.1 98.2 ,0.0001Xabg705 3 Xpsr129 1 0.9 0.9 0.3 0.6112Xpsr1205 3 Xabg705 3
Xpsr1291 2.7 2.7 0.7 0.3894
Error 300 1075.6 3.6
TABLE 5
Comparison of subpopulations of Chinese Spring 3 Ae. speltoides hybrids with indicated QTL increasing homeologouschromosome pairing in hybrids relative to subpopulations devoid of all such QTL (background) or having fewer
of them, and the effect of substitution of the b-haplotypes for the a-haplotypes at indicated loci on themean chiasma number per cell in subpopulations of Chinese Spring 3 Ae. speltoides hybrids
QTLNo. of hybrids in
smaller subpopulationChiasmata
per cellCompared QTL
effectsQTL effect
(chiasmata per cell) F a Probability
Background 7.0 — — — —3S 23 15.4 3S vs. background 8.4 390.1 ,0.0015S 62 8.6 5S vs. background 1.6 6.9 0.0017S 52 12.4 7S vs. background 5.8 115.0 ,0.0013S 1 7S 23 15.1 3S 1 7S vs. 3S 0.5 1.1 0.2893S 1 5S 15 15.2 3S 1 5S vs. 3S 0.1 0.1 0.8797S 1 5S 31 14.2 7S 1 5S vs. 7S 1.8 15.2 ,0.0013S 1 5S 1 7S 19 16.4 3S 1 5S 1 7S vs. 3S 1 7S 0.8 4.1 0.049
a F-value from one-way ANOVA.
Wheat Ph1 Suppressors 23

hybrids; subpopulation means were 5.7 and 15.5 chias-mata/cell (Figure 2). While the latter mean was almostidentical to the expected 15.4 chaismata (Table 5), theformer mean was 1.3 chiasmata lower than the expected7.0 chiasmata/cell for the background. This differencewas attributed to the environment: the population ofhybrids derived from the F4 plant 125-80-13 was grownin the summer whereas the previous population wasgrown in the winter. The mean phenotypic difference of9.8 chiasmata/cell between the classes (Figure 3) madeit possible to allocate each of the 67 hybrids into eitherthe high- or the low-pairing class.
DNAs were isolated from the 67 hybrids, and South-ern blots were hybridized with 23 wheat ESTs (http://wheat.pw.usda.gov/cgi-bin/westsql/map_locus.cgi) andMWG41, BCD1261, and PSR1205 clones. A genetic mapof chromosome 3S was constructed. The map was 70.7cM long and was entirely collinear with the map of theclosely related Ae. tauschii chromosome 3D (not shown).The QTL completely cosegregated with Xpsr1205. Sincethe QTL behaved in all respects as a Mendelian locus, itwas designated as Su1-Ph1 (suppressor number 1 of thePh1 gene). Su1-Ph1b was the high-pairing haplotype.
To determine the position of the Su1-Ph1 locusrelative to Xpsr1205 and flanking markers, five F4
siblings of the Ae. speltoides plant 125-80-13, with thesame genotype as the Ae. speltoides plant 125-80-13, werecrossed with Chinese Spring and DNAs were isolated.The 266 hybrids produced were screened for recombina-tion in the XBF497740–XBE442875 interval harboringXpsr1205 (Figure 4) using Ae. speltoides genome-specificprimers. The average number of chiasmata per cell wasdetermined in hybrids harboring chromosomes recom-bined in the XBF497740–XBE442875 interval. DNAs werereisolated from the hybrids for Southern analyses. ThePCR- and TaqMan-based analysis and Southern-hybrid-ization-based analysis consistently showed that 1 of the266 hybrids harbored a chromosome with a crossoverbetween Su1-Ph1 and Xpsr1205. The crossover placedthe Su1-Ph1 locus 0.4 cM proximal to Xpsr1205 andXBE488620 (Figure 4).
To map 7S QTL, the Ae. speltoides F4 plant 125-112-16was crossed with Chinese Spring. The plant was homo-zygous for Xpsr1205a and Xabg705a and markers on theshort arm of chromosome 7S but heterozygous formarkers on the long arm of chromosome 7S. A total of74 hybrids were analyzed. The frequency distribution ofmean chiasma number per cell was clearly bimodalamong the 74 hybrids (Figure 5). The populationsegregated with 31 high-pairing haplotypes to 43 low-pairing haplotypes, which is close to a 2:1 ratio ratherthan the expected 1:1 ratio. The most likely cause of thisanomaly was segregation distortion operating againstthe high-pairing QTL haplotype. Segregation of theXpsr129b and Xpsr129a haplotypes among the 315hybrids analyzed earlier was 130:185, which does notdiffer from the observed segregation of the previous74 hybrids (P ¼ 0.98, Fisher’s exact test). Taking thesegregation distortion into account, the frequencydistribution of pairing scores among the 74 hybrids isconsistent with the segregation of a single Mendeliangene pair. We designate the locus as Su2-Ph1. Su2-Ph1b isthe high-pairing haplotype. The mean chiasma numberof hybrids in the low-homeologous-chromosome-pairing class was 6.6 chiasmata/cell and that of hybridsin the high-homeologous-chromosome-pairing class was11.9 (Figure 5), both numbers being close to the ex-pected 7.0 and 12.4 chiasmata/cell (Table 5). Recom-bination relative to seven molecular marker loci fromthe long arm of wheat chromosomes of homeologousgroup 7 (Dubcovsky et al. 1996; Hossain et al. 2004)
Figure 2.—Frequency distribution of the mean number ofchiasmata per cell among 67 hybrids from the cross ChineseSpring 3 F4 Ae. speltoides plant 125-80-13. The means of thelow-homeologous-pairing and high-homeologous-pairing sub-populations are indicated.
Figure 3.—Chromosome pairing in pollenmother cells of hybrids between Chinese Spring(CS) and Ae. speltoides differing by the haplotypeat the Su1-Ph1 locus.
24 J. Dvorak, K. R. Deal and M.-C. Luo

was used to estimate the position of Su2-Ph1 on theAe. speltoides genetic map. The Su2-Ph1 locus was be-tween the EST marker loci XBE496047 and XBG274334(Figure 4).
The interval Xwg420–XBE496047 mapped here waspreviously mapped on the Ae. speltoides map based on 86F2 plants from the 2-12-4 3 PI369609-12-II cross (Luo
et al. 2005). While the interval was 8.4 cM on that map, itwas 27.3 cM on the map constructed here (P , 0.01).The expansion of the map constructed here occurred inall intervals that could be compared between the twomaps.
DISCUSSION
Three Ae. speltoides loci controlling homeologouschromosome pairing were identified in T. aestivum 3
Ae. speltoides hybrids. Either individually or in variouscombinations, variation at these loci could elicit virtually
any level of homeologous chromosome pairing between7.0 chiasmata/cell and 16.4 chiasmata/cell in ChineseSpring 3 Ae. speltoides hybrids, thus accounting for thevariation originally described by Dvorak (1972). Twounlinked major suppressors of Ph1 were mapped onchromosomes 3S (Su1-Ph1) and 7S (Su2-Ph1) and shownto strongly interact with each other. Su1-Ph1b was fullyepistatic to Su2-Ph1b. However, without knowledge ofthe dominance relationships between the a- and b-hap-lotypes, it cannot be decided what type of epistaticinteraction is involved. The expression of Su1-Ph1 andSu2-Ph1 is not equivalent: Su1-Ph1 is a stronger suppres-sor than Su2-Ph1. Therefore, the Su1-Ph1 and Su2-Ph1dihybrid would produce three hybrid classes rather thanthe two expected for duplicate gene interaction. Never-theless, it is likely that the action of the two genes isclosely related at the molecular level and that the genesmay even have originated by duplication of a singleancestral locus.
The discovery of a QTL on chromosome 5S with aminor effect provides evidence for the existence of asystem of minor genes affecting homeologous chromo-some pairing in the Chinese Spring 3 Ae. speltoideshybrids predicted by Chen and Dvorak (1984). Follow-ing the rules of wheat QTL nomenclature, we designatethis QTL as QPh.ucd-5S (chromosome 5S QTL affectingpairing of homeologous chromosomes). QPh.ucd-5Sb isthe high-pairing haplotype of the QTL. QPh.ucd-5S islocated in the low-recombination region of chromo-some 5S. The lack of recombination of molecularmarkers in that region makes the location of QPh.ucd-5S uncertain. The effects of the QTL peaked near theXabg705 and Xpsr628 loci, both in the short arm of wheatchromosomes of homeologous group 5 (Gale et al. 1993;Dubcovsky et al. 1996). The expression of QPh.ucd-5Sis additive to Su2-Ph1 and under some circumstances
Figure 4.—Linkage maps of Ae. speltoides chromosome 3Sshowing the position of the Su1-Ph1 locus (boxed) relativeto EST markers (designated as BE or BF) and anonymousRFLP markers based on 67 (a) and 266 (b) Chinese Spring3 Ae. speltoides hybrids and the linkage map of Ae. speltoideschromosome 7S showing the position of the Su2-Ph1 locus(boxed) relative to EST markers (designated as BE, BF, andBG) and anonymous RFLP markers. Genetic distances be-tween markers are in centimorgans and are indicated to theleft of each chromosome.
Figure 5.—Frequency distribution of the mean number ofchiasmata per cell among 74 hybrids from the cross ChineseSpring 3 F4 Ae. speltoides plant 125-112-16. The means of thelow-homeologous-pairing and high-homeologous-pairing sub-populations are indicated.
Wheat Ph1 Suppressors 25

also to Su1-Ph1. The latter is observed in the enhance-ment of homeologous pairing from 15.1 to 16.4 chias-mata/cell in Chinese Spring 3 Ae. speltoides hybridsdue to the addition of QPh.ucd-5Sb to Su1-Ph1b and Su2-Ph1b.
The QPh.ucd-5S could correspond to the homeolo-gous chromosome pairing promoting locus on the shortarms of wheat chromosomes 5A, 5B, and 5D (Riley andChapman 1967; Dvorak 1976; Cuadrado et al. 1991)and on the Lophopyrum elongatum chromosome 5E(Dvorak 1987). Promotion of homeologous chromo-some pairing by these loci was suggested by theobservation that the level of homeologous chromosomepairing was reduced if the short arm of 5A, 5B, or 5D wasabsent from hybrids or haploids. However, the geneswere shown to be essential for crossing over betweenhomologous chromosomes and their primary role isvery likely controlling the recombination betweenhomologous chromosomes (Kota and Dvorak 1986).They show additive effects and an increase in their doseresults in an increase in homeologous chromosomepairing and partial suppression of Ph1 (Dvorak 1987).This expression pattern parallels the additive effects ofQPh.ucd-5S. On the basis of interactions among thethree loci discovered here, we hypothesize that Su1-Ph1and Su2-Ph1 act on Ph1 whereas QPh.ucd-5S affectsrecombination between homologous chromosomesand acts independently of the Ph1 mechanism.
Comparative sequencing of the wheat Ph1 region ledGriffiths et al. (2006) to the hypothesis that an in-sertion of heterochromatic sequence from chromo-some 3A into the cdc2-4 gene in the cdc2 multigenelocus on chromosome 5B is synonymous with Ph1. ThePh1 gene is completely dominant. Its deletion results inthe restoration of the default state, which is the presenceof homeologous chromosome pairing. That makes itunlikely that the genetic event that produced Ph1 was aloss-of-function mutation and poses difficulties with thehypothesis that the cdc2-4 pseudogene is synonymouswith Ph1. The cdc2 multigene locus is almost certainlypresent on chromosome 5S since no translocation in-volving the long arm of Ae. speltoides chromosome 5Swas detected in comparative mapping of the Ae. speltoidesgenome (Luo et al. 2005). Yet the long arm of chromo-some 5S had no effect on Ph1 activity. These argumentssuggest that Ph1 is an active gene rather than apseudogene.
In addition to Ae. speltoides, Ae. mutica is the only otherspecies possessing genes that fully suppress the Ph1activity (Dover and Riley 1972). The Ph1 suppressionin Ae. mutica is also controlled by two major genes(Dover and Riley 1972). Although the chromosomallocation of the Ae. mutica suppressors is unknown, it ispossible that they correspond to Su1-Ph1 and Su2-Ph1.
The Ph1 locus prevents recombination betweenhomeologous chromosomes in wheat and is thus apowerful barrier to introgression of alien genes from
wheat relatives into wheat and their exploitation forwheat improvement. Incorporation of a Ph1 suppressorinto wheat would greatly facilitate introgression of aliengenes into wheat chromosomes via recombinationbetween homeologous chromosomes. To accomplishthis goal, Chen et al. (1994) twice backcrossed Ae.speltoides to wheat and developed a wheat line showinga partial suppression of Ph1. The gene was named PhI
(inhibitor of Ph). It was suggested that it was intro-gressed into wheat chromosome 4D, presumably via acrossover between homeologous chromosomes 4D and4S. Weak expression of PhI and our failure to map any ofthe major Ph1 suppressors on chromosome 4S suggeststhat PhI corresponds to neither Su1-Ph1 nor Su2-Ph1.The presence of multivalents in hybrids with the back-ground level of chromosome pairing strongly suggeststhat Ph1 was partially suppressed even at the back-ground level in our study and that the PhI could possiblybe a locus that we failed to detect. Chromosomal map-ping of the PhI gene was based on indirect evidence, andthe possibility that the gene actually corresponds to aweaker allele at either the Su1-Ph1 or the Su2-Ph1 locusshould not be entirely dismissed.
Of the three loci discovered here, Su1-Ph1 is thelogical target for introgression into wheat and manipu-lation of homeologous recombination in wheat. Sincethe gene is epistatic to Su2-Ph1, no advantage would begained by combining Su1-Ph1 with Su2-Ph1. The Su1-Ph1gene is located in the distal region of chromosome 3Sthat is collinear with wheat chromosome 3B (Luo et al.2005). The identification of molecular markers tightlylinked to Su1-Ph1 and the development of Ae. speltoidesgenome-specific PCR primers for the detection of Ae.speltoides markers in the wheat genetic backgroundaccomplished here will greatly facilitate the detectionof Su1-Ph1 during the backcrossing of Ae. speltoides intowheat.
The mechanism of the localization of crossovers inshort distal regions of Ae. speltoides chromosomes (Luo
et al. 2005) is currently unknown. The observation that afamily of hybrids derived from an Ae. speltoides F4 plantshowed a dramatic increase in recombination in theproximal region compared to that observed in the F2
generation from the same cross suggests that geneticvariation may exist in material developed in this project,which could be exploited in genetic characterization ofthe mechanism controlling crossover localization in Ae.speltoides.
We thank O. D. Anderson, M. D. Gale, A. Graner, B. S. Gill, G. E. Hart,A. Kleinhofs, and M. E. Sorrells for sharing clones with us and the U. S.Department of Agriculture/Cooperative State Research, Educationand Extension Service/National Research Initiative for financialsupport by grant 2001-35301-10594 to J. Dvorak and M.-C. Luo.
LITERATURE CITED
Allard, R. W., 1956 Formulas and tables to facilitate the calculationof recombination values in heredity. Hilgardia 24: 235–278.
26 J. Dvorak, K. R. Deal and M.-C. Luo

Chen, K. C., and J. Dvorak, 1984 The inheritance of genetic varia-tion in Triticum speltoides affecting heterogenetic chromosomepairing in hybrids with Triticum aestivum. Can. J. Genet. Cytol.26: 279–287.
Chen, P. D., H. Tsujimoto and B. S. Gill, 1994 Transfer of PhIgenes promoting homoeologous pairing from Triticum speltoidesto common wheat. Theor. Appl. Genet. 88: 97–101.
Cuadrado, C., C. Romero and J. R. Lacadena, 1991 Meiotic pair-ing control in wheat-rye hybrids.1. Effect of different wheat chro-mosome arms of homoeologous group-3 and group-5. Genome34: 72–75.
Dover, G. A., and R. Riley, 1972 Variation at two loci affecting ho-meologous meiotic chromosome pairing in Triticum aestivum 3Aegilops mutica hybrids. Nature New Biol. 235: 1–6.
Dover, G., and R. Riley, 1977 Inferences from genetical evidenceon the course of meiotic chromosome pairing in plants. Philos.Trans. R. Soc. Lond. Ser. B 277: 313–326.
Dubcovsky, J., M. C. Luo, G. Y. Zhong, R. Bransteitter, A. Desai
et al., 1996 Genetic map of diploid wheat, Triticum monococcumL., and its comparison with maps of Hordeum vulgare L. Genetics143: 983–999.
Dvorak, J., 1972 Genetic variability in Aegilops speltoides affectinghomoeologous pairing in wheat. Can. J. Genet. Cytol. 14:371–380.
Dvorak, J., 1976 The relationship between the genome of Triticumurartu and the A and B genomes of T. aestivum. Can. J. Genet.Cytol. 18: 371–377.
Dvorak, J., 1987 Chromosomal distribution of genes in Elytrigiaelongata which promote or suppress pairing of wheat homoeolo-gous chromosomes. Genome 29: 34–40.
Dvorak, J., 1998 Genome analysis in the Triticum-Aegilops alliance,pp. 8–11 in 9th International Wheat Genetics Symposium, edited byA. E. Slinkard. University Extension Press, University of Sas-katchewan, Saskatoon, Saskatchewan, Canada.
Dvorak, J., and H. B. Zhang, 1990 Variation in repeated nucleotidesequences sheds light on the phylogeny of the wheat B and Ggenomes. Proc. Natl. Acad. Sci. USA 87: 9640–9644.
Dvorak, J., P. E. McGuire and B. Cassidy, 1988 Apparent sourcesof the A genomes of wheats inferred from the polymorphism inabundance and restriction fragment length of repeated nucleo-tide sequences. Genome 30: 680–689.
Dvorak, J., Z.-L. Yang, F. M. You and M. C. Luo, 2004 Deletion poly-morphism in wheat chromosome regions with contrasting re-combination rates. Genetics 168: 1665–1675.
Feldman, M., and T. Mello-Sampayo, 1967 Suppression of homeol-ogous pairing in hybrids of polyploid wheats 3 Triticum speltoides.Can. J. Genet. Cytol. 9: 307–313.
Friebe, B., L. L. Qi, S. Nasuda, P. Zhang, N. A. Tuleen et al.,2000 Development of a complete set of Triticum aestivum-Aegilops speltoides chromosome addition lines. Theor. Appl.Genet. 101: 51–58.
Gale, M. D., M. D. Atkinson, C. N. Chinoy, R. L. Harcourt, J. Jiaet al., 1993 Genetic maps of hexaploid wheat, pp. 29–40 in 8thInternational Genetic Symposium, edited by Z. S. Li and Z. Y. Xin.China Agricultural Scientech Press, Beijing.
Griffiths, S., R. Sharp, T. N. Foote, I. Bertin, M. Wanous et al.,2006 Molecular characterization of Ph1 as a major chromosomepairing locus in polyploid wheat. Nature 439: 749–752.
Hossain, K. G., V. Kalavacharla, G. R. Lazo, J. Hegstad, M. J.Wentz et al., 2004 A chromosome bin map of 2148 expressedsequence tag loci of wheat homoeologous group 7. Genetics 168:687–699.
Kimber, G., and R. S. Athwal, 1972 A reassessment of the course ofevolution of wheat. Proc. Natl. Acad. Sci. USA 69: 912–915.
Kosambi, D. D., 1943 The estimation of map distances from recom-bination values. Ann. Eugen. 12: 172–175.
Kota, R. S., and J. Dvorak, 1986 Mapping of a chromosome pairinggene and 5SrRNA genes in Triticum aestivum L. by a spontaneousdeletion in chromosome arm 5Bp. Can. J. Genet. Cytol. 28: 266–271.
Lincoln, S., M. Daly and E. Lander, 1992 Whitehead Institute Tech-nical Report, Ed. 3. Whitehead Institute, Cambridge, MA.
Luo, M. C., K. R. Deal, Z. L. Young and J. Dvorak,2005 Comparative genetic maps reveal extreme crossover local-ization in the Aegilops speltoides chromosomes. Theor. Appl.Genet. 111: 1098–1106.
Okamoto, M., 1957 Asynaptic effect of chromosome V. Wheat Inf.Serv. 5: 6.
Riley, R., 1960 The diploidization of polyploid wheat. Heredity 15:407–429.
Riley, R., and V. Chapman, 1958 Genetic control of the cytologi-cally diploid behaviour of hexaploid wheat. Nature 182: 713–715.
Riley, R., and V. Chapman, 1967 Effects of 5Bs in suppressing theexpression of altered dosage of 5BL on meiotic chromosome pair-ing in Triticum aestivum. Nature 216: 60–62.
Sarkar, P., and G. L. Stebbins, 1956 Morphological evidence concern-ing the origin of the B genome in wheat. Am. J. Bot. 43: 297–304.
Sears, E. R., and M. Okamoto, 1958 Intergenomic chromosome re-lationships in hexaploid wheat. Proceedings of the X Interna-tional Congress of Genetics, McGill University, Montreal, pp.258–259.
Communicating editor: J. A. Birchler
Wheat Ph1 Suppressors 27
Related Documents











