Diplomarbeit Mechanisms of nutrient dynamics in the canopy of a tropical lowland rain forest zur Erlangung des akademischen Grades Magistra der Naturwissenschaften an der Fakultät für Lebenswissenschaften der Universität Wien eingereicht von Nina Hinko Wien, Oktober 2007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Diplomarbeit
Mechanisms of nutrient dynamics in the canopy
of a tropical lowland rain forest
zur Erlangung des akademischen Grades
Magistra der Naturwissenschaften
an der
Fakultät für Lebenswissenschaften
der Universität Wien
eingereicht von
Nina Hinko
Wien, Oktober 2007

Index
1. General Introduction 1
Tropical rain forests 1
Nutrient cycling in tropical rain forests 1
Throughfall 4
Dry deposition 7
Canopy exchange processes 9
Study aims 14
References 15
2. Manuscript 20
Mechanisms of nutrient dynamics in the canopy of a tropical lowland rain forest
Abstract 20
Introduction 21
Materials and Methods 24
Results 31
Discussion 34
References 41
Figures and tables 45
3. Zusammenfassung 52
4. Appendix 55

- 1 -
1. General Introduction
1.1. Tropical rain forests
Tropical rain forests are distributed between the tropics of Cancer (23° 26′ 22″ N latitude) and
Capricorn (23° 26′ 22″ S latitude) and occur in all three possible tropical land areas: Africa,
South-East Asia and Australia (paleotropics) and America (neotropics) where the Americas
possess the most extensive rain forests. They exhibit an exorbitant extreme species richness
and it is estimated that approximately two thirds (170.000) of the world’s plant species occur
in tropical forests (Whitmore 1998). Further tropical rain forests are characterized by high solar
radiation because of the low latitude, a relatively stable climate throughout the year with
continuously high temperatures and high relative humidity with dry seasons being short or
negligible. Tropical rain forests therefore exhibit the highest annual rates of net primary
production (NPP) of all terrestrial ecosystems (Montagnini 2005; Jordan 1971).
Nonetheless tropical ecosystems are highly complex systems and considered to be fragile or
ecologically sensitive. Therefore effects of global change, implying not only climate change but
land use change and change in atmospheric composition and deposition can be critical for
rainforest functioning. Despite tropical rainforests exhibiting the highest biodiversity and
biomass production, most tropical soils are supposed to be highly leached and weathered and
therefore to have an extremely low fertility. Hence the major part of the nutrient pool is stored
in above-ground biomass and nutrient cycling and conservation mechanisms play a crucial role
in maintaining ecosystem productivity in tropical rain forests (Whitmore 1998).
1.2. Nutrient cycling in tropical rain forests
In addition to external nutrient inputs to and outputs from ecosystems nutrient cycling i.e.
internal fluxes of nutrients within ecosystems are thought to be crucial for sustained high
primary production of tropical rain forests. Nutrient inputs are supplied through chemical
weathering of parent material, atmospheric deposition and biological fixation of atmospheric
nitrogen. Nutrient losses occur through erosion and leaching of soils into stream- and
groundwater, emission of gases, wind-driven relocation and disturbances by natural events
such as fire. Nutrient cycling within an ecosystem is comprised of uptake of nutrients by
vegetation and microorganisms, following incorporation into organic material and finally the
release of nutrients via litterfall and throughfall and their decomposition by microorganisms. In
natural ecosystems, for instance tropical rain forests, the amount of nutrients which cycle
internally exceeds those of inputs and outputs by far and tropical rain forests therefore can be
considered as rather closed systems. In consideration of human activities, nutrient inputs and

- 2 -
outputs in ecosystems obtain an increasing significance because of additional entry of nutrients
due to fertilization, fossil fuel combustion and removal through harvest and fire (Chapin III
2002).
The productivity in terrestrial ecosystems is constrained by climate (rainfall, temperature,
seasonality, etc) and nutrient supplies mostly from the soil. Nutrients are made available for
uptake by plants through the processes of decomposition and mineralization of dead organic
matter which in turn comes from plants via litterfall, animals and microbes. Dissolved mineral
nutrients are absorbed by plants via roots and move upward in the xylem with the transpiration
water stream into leaves. There they are allocated to production of new tissue retranslocation,
or storage. Finally nutrients return to the soil via two main pathways: litterfall and throughfall
i.e. precipitation which is penetrating the canopy before it is reaching the ground. Additional
plants can trap dissolved organic nutrients e.g. nitrogen from decomposing material as soon as
they are released as well through direct transfer mediated mainly by mycorrhiza (Herrera
1978).
As mentioned above nutrient cycling is essential for continued plant growth in tropical forest
ecosystems and nutrient cycles therefore are generally found to be rather closed. Nevertheless
it must be pointed out that not all tropical forests grow on low fertility soils despite a great
proportion are oligotrophic ecosystems such as the Amazon Basin. In contrast to these nutrient
poor rain forests which receive nutrients solely from litter- and rainfall there also existing
nutrient rich tropical rain forest ecosystems especially in Central America where many soils
derived from volcanic rock. They consist of a substantial fraction of (mineral) nutrients in the
rooting zone and show nutrient cycles that are more open.
Factors that influence nutrient cycling in tropical rain forests are of biological (plant, bacterial
and fungal species), environmental (moisture and temperature) and chemical nature (nature of
soil nutrients, secondary plant components) (Attiwill 1993). Decomposition in tropical forests is
generally enhanced at high temperature, high relative moisture and availability of sufficient
oxygen which accelerate microbial activity. Hence tropical lowland rainforests exhibit a small
litter pool (Chapin III 2002; Whitmore 1998).
Litter consists of aboveground leaf litter and of belowground root litter. Soils in tropical rain
forests exhibit a remarkable amount of fine root biomass in the surface soil and litter layer
(Klinge 1973). Stark (1978) demonstrated with tracer-experiments that the fine root mat can
directly absorb dissolved nutrients released from decomposing litter and reaching the surface
soil via precipitation and throughfall. This nutrient conserving mechanism of the fine root mat
plus associated mycorrhizas enhances the uptake of nutrients, especially phosphorus (Herrera

- 3 -
1978). Nearly all tree species worldwide have ecto- or arbuscular mycorrhiza. Consequently,
the root mat plays an important role in the retention and recycling of nutrients on the forest
floor.
In tropical lowland rainforests internal recycling sustains the availability of N and P for plants
though phosphorus return via litterfall is very small compared to nitrogen (Vitousek 1984).
According to Vitousek (1982) it is assumed that P is cycled very efficient and N seems to have
low within-stand efficiency despite its rapid circulation. This supports that P availability and not
that of N limits productivity of most tropical lowland forests given that the most growth limiting
nutrient defines the cycling rate of all other nutrients (Chapin III 2002). It is not proven that N
is not used efficiently due to the fact that in those forests cycling occurs really fast and a unit of
N could circulate several times per year (Vitousek 1984). However, plant available phosphorous
is generally present at very small concentrations and is retained within the plants. Intra-plant
nutrient conservation (Jordan 1980) occurs via reabsorption and translocation of mobile
nutrients from senescing tissue through the phloem to other plant parts, i.e. to young leaves
where mineral nutrients are required to build up organic matter or to storage organs.
Nevertheless some nutrients such as Calcium are immobile in the phloem and therefore cannot
be resorbed.
Transfers of nutrients to the forest floor occur through litterfall (LF) and through precipitation in
the form of throughfall (TF) and stemflow (SF). The contribution of TF and SF as pathways for
solute (nutrient) inputs to the forest floor is important particularly in forests with abundant
rainfall and infertile soils, although amounts of nutrients in precipitation are generallly lower
than those recycled by litterfall (Tobón 2004). Nevertheless throughfall fluxes can be
ecologically significant for certain elements. In the case of potassium McDowell (1998)
demonstrated that K+ flux in TF is twice as much as in LF for a tropical wet rain forest in Puerto
Rico. In contrast nitrogen as well as calcium and phosphorus inputs to the soil derive largely
from litterfall. Calcium content in litterfall usually exceeds that of living foliage because Ca is
immobilized in cell wall components such as pectates (Johnson 1992) and can not be removed
from senescent leaves before shedding. Cavelier (1997) compared annual rates of N, P and K in
throughfall with rates in litterfall from Veneklaas (1991) for a tropical montane cloud forest. N
inputs in net throughfall NTF were significantly lower than mean inputs in litterfall while P
inputs were similar in both LF and NTF. In contrast K inputs were significantly higher in NTF
than in LF. The considerable increases of K in TF can be attributed to its high mobility and
abundance in leaves (Tobón 2004).

- 4 -
1.3. Throughfall
It is has been often demonstrated that precipitation is altered in its chemical composition
during interception through the forest canopy and that throughfall is predominantly enriched in
certain elements (Parker 1983; Lovett 1996; Veneklaas 1990; Whitmore 1998). Precipitation
which penetrates the forest canopy can reach the forest floor as throughfall (rainfall which
passed through the canopy) and as stemflow (rainfall which runs off the tree stem). Jordan
(1980) found that throughfall made up 80-90% of incident precipitation in a tropical rainforest
whereas stemflow accounted for only 1-2% of total solute fluxes. This agrees with results from
Chuyong (2004) who reported that throughfall and stemflow in a central African rain forest
accounted for 92.4-96.6% and 1.5-2.2%, respectively. Despite the very small contribution to
total nutrient fluxes stemflow is generally more enriched in nutrients than throughfall and
therefore displays a substantial yet localized nutrient input to forest floor (Parker 1983). The
difference (1.9-5.4%) between incident precipitation and TF (and SF) represents the
interception loss within the canopy. Throughfall, stemflow and interception measurements for a
tropical montane rain forest in Costa Rica resulted in 70, 2 and 28% of incident rainfall,
respectively (Hölscher 2004).
In consideration of ecosystem nutrient fluxes precipitation is a supplementary and small but
important nutrient input to forests by which nutrients from outside enter the ecosystem. For
certain elements throughfall is the major pathway for recycling and therefore the internal
nutrient dynamics of the forest (Parker 1983), particularly in wet tropical rainforests where
rainfall is high and nutrient cycling is known to be of considerable importance to meet the
nutrient demand by vegetation. Furthermore dissolved nutrients in throughfall are highly
available to organisms compared to those of litterfall, may be an important pulse at the
beginning of the rainy season and probably facilitate litter decomposition and mineralization
(Chuyong 2004) thereby reducing litter turnover times. Processes involved in chemical
alteration of precipitation are exchange processes between the canopy and the water on plant
surfaces such as leaching and uptake of dissolved material by canopy components, as well as
dissolution and wash off of dry deposited atmospheric materials such as aerosols, particulate
and gaseous substances (Lovett 1984; Hansen 1994; Parker 1983).
Parker (1983) reviewed that for K+, Na+, Mg2+ and S, the rates of NTF exceed those of litterfall
while N, P and Ca derive largely from litterfall. It is assumed that potassium derives mainly
from foliar or canopy leaching because of it is high mobility. Calcium and magnesium are also
found in higher concentrations in TF compared to BP. Calcium was shown to originate almost
equally from leaching of exchangeable pools in leaf apoplast and from wash off of particulate

- 5 -
dry deposition (Forti 1992). In contrast Mg2+, Na+ and Cl- are introduced mainly by sea salt
particles. H+ concentration (acidic deposition) in rainwater decreases during passage through
the canopy which implies that uptake (ion-exchange) processes occur. The absorption from H+
triggers increased leaching of other soluble or ion exchangeable cations such as K+, Ca2+ and
Mg2+, and weak organic acids from foliage. Numerous studies demonstrated a decline of
inorganic nitrogen concentration in precipitation during canopy penetration, especially for NH4+,
suggesting retention of N in the canopy. Apart from throughfall enrichment of mineral nutrients
it is well known that TF is additional loaded with organic compounds such as DOC and DON.
The origin of these sources is found within the canopy through leaching from canopy litter,
herbivore activities and leaching from bryophytes and exchangeable foliar pools (Coxson
1992).
Throughfall is defined as:
TF = BP + DD + CE
where TF is throughfall, BP is incident (bulk) precipitation, DD is dry deposition and CE
describes canopy exchange processes with uptake and leaching of nutrients within the canopy.
The net throughfall flux can then be calculated as:
NTF = TF – BP = DD + CE
referring to the sum of dry deposition and canopy exchange representing net throughfall
(Lovett 1996). Positive rates of NTF indicate a net enrichment of elements in throughfall due to
dry deposition and canopy leaching whereas negative rates present a net decrease in nutrient
concentration in throughfall due to canopy uptake or interception loss.
The separation between these processes, dry deposition and canopy exchange, and
quantification of their contribution to throughfall still remains a challenge. Moreover, it has
been suggested that both processes are highly dynamic (Hansen 1994) regarding to time,
space and the influence by several factors. Present data are often difficult to compares because
sampling design and analytical methods are highly variable. Collection and measurement of
bulk precipitation includes among wet deposition (elements dissolved in precipitation) also a
fraction of dry deposition such as dust and particles which enter the sampling gauge. Moreover,
dry deposition occurs to canopy surfaces causing an overestimation of canopy leaching rates
(Parker 1983). An essential part of dry deposition – deposition of gaseous substances – is
therefore not accounted for and moreover is complicated to measure. Though studies exist for
temperate forests in tropical rain forests the knowledge is even scare. Temperate forests
exhibit a less complex forest and canopy structure and are often even-sized and even-aged
compared to tropical rain forests and therefore allow an easier access for investigations.

- 6 -
The flux and chemistry of throughfall are affected by various factors and processes of
atmospheric, hydrological, chemical and biological nature, and as well human activities, which
themselves are highly temporally and spatially variable (Parker 1983; Potter 1991; Tobón
2004). Numerous studies reported that fluxes of dissolved nutrients are highly correlated with
the amount, duration and the intensity of precipitation penetrating the canopy (Reiners 1984;
Parker 1983; Lovett 1996; Tobón 2004; Potter 1991) that are much higher in tropical forests
than in temperate forests. A high intensity of rainfall leads to a decrease in ion concentration in
TF during the event while low intensities correspond to elevated solute concentrations.
Precipitation of low intensity prolongs the residence time for water within the canopy and hence
exchange processes have more time to take place (Lovett 1992; Hansen 1994). Throughfall
chemistry also strongly depends on the type of precipitation such as clouds, droplets, mist and
dew. Indirect horizontal precipitation such as clouds have elevated concentrations of elements
and can be important especially in tropical montane cloud forests where quantities of
throughfall can exceed those of bulk precipitation and therefore cause an extra input of water
(Cavelier 1997) and nutrients (Veneklaas 1990).
Factors such as altitude, forest type and canopy structure, stand age or successional state,
species composition, seasonality and nutrient supply are also important factors influencing
throughfall fluxes. The leaf area index (LAI) as an index for the canopy density also influences
the amount of throughfall or rather interception loss. During the drier season in a tropical
montane cloud forest the interception by the canopy was lower than during the rainy season
probably due to a decreased LAI because of less growth and an interception maximum at the
beginning of the wet season as a result of increased LAI from leaf flush (Cavelier 1997). Potter
(1991) reported also up to 12 times higher NTF during the growing season than during
dormant season in the Appalachian forests, indicating the considerable influence of foliage in
altering throughfall chemistry. The occurrence of epiphytes may increase also wet and dry
deposition through the enlargement of the surface area for impaction and sedimentation
(Reiners 1984).
Moreover Hansen (1994) reported that ion concentrations in TF during the initial part of a rain
event (the first 2 mm) were highest after long dry periods which indicated that inter-event
deposited (external source) and probably foliar material (internal source) are easily leached in
the early phase of wetting. In subsequent rain the ion concentrations generally decrease except
for NH4+ and K+ which showed a high variability. Atmospheric nutrient inputs (aerosols, dust or
gases) also vary with proximity to sources such as the ocean, volcanoes, industries and
agriculture (Attiwill 1993; Parker 1983). High concentrations of Na+, Cl- and Mg2+ are often
closely related to the sea nearby whereas enhanced N, P and S concentrations are linked to
deposition of air pollutants and hence derive from anthropogenic sources.

- 7 -
Nevertheless, canopy exchange processes are also related to the chemical composition in
incident precipitation. If the concentration of a certain nutrient is high, net fluxes are more
likely to be negative i.e. a net uptake by the canopy while low concentrations in rain lead to a
positive net flux from the canopy to the forest soil i.e. a net loss of nutrients (Veneklaas 1990).
Highly leachable elements, e.g. potassium, do not follow this pattern.
As mentioned above the nature of the canopy such as epiphytic colonization and abundance of
lichens and bryophytes affect the interception and alteration of rainfall. The greater the canopy
roughness which means more species with different crown structures and more gaps in the
upper canopy, the higher dry deposition rates may occur and the more precipitation is
intercepted by the canopy (Lovett 1984; Veneklaas 1990). Forti (1991) found out that ion
concentrations of wet deposition samples above the canopy compared to those from clearing
are overestimated due to the fact that the roughness of the upper canopy layer generates
vortices and turbulent mass flow at the top of the canopy which resulted in enrichment of
aerosols above the canopy and an increase of ions in incident precipitation. In contrast, Hansen
(1996) reported that due to these turbulent air movements less incident precipitation was
collected and that ion concentrations above the canopy and from an open field were similar.
Dry deposition and canopy exchange as well are expected to change with canopy depth
(Hansen 1994; Parker 1983; Lovett 1992). DD is generally assumed to be highest in the upper
canopy due to their greater exposure to wind and turbulent air whereas in lower canopy layers
the incident rainfall is already enriched in nutrients through their wash-off or leaching from the
upper layers.
Evaporation of intercepted rainwater in inter-event periods can cause an increase of ion
concentration on foliage surfaces previously dissolved in rainwater. Additionally
evapotranspiration exceeds incident rainfall in dry seasons and may provoke an increase in
transpiration and thereby in nutrient release from leaves through their stomata (Parker 1983).
The main processes affecting the variability in throughfall chemistry, dry deposition and canopy
exchange (Tobón 2004; Ragsdale 1992; Lovett 1984), will be discussed in further detail below.
1.4. Dry deposition
Atmospheric deposition
Atmospheric deposition represents an external input of elements or nutrients to ecosystems
and includes wet deposition and dry deposition. Wet deposition comprises incident precipitation
i.e. water droplets in which aerosols, gases or ions are dissolved. Solutes can originate from
other locations nearby or distant (e.g. the ocean) or dissolution took place shortly before

- 8 -
reaching the top of the canopy. Dry deposition combines all input processes between rain
events and is made up of physical processes such as sedimentation and impaction of particles
and aerosols and absorption of gaseous forms (Parker 1983). An additional crucial biotic
nutrient input within the canopy is the fixation of atmospheric nitrogen (N2) by free-living
cyanobacteria or symbiotic epiphyllous microorganisms (lichens) growing on leaf surfaces. A
considerable portion of this newly fixed N by epiphylls may be transferred to the host leaf
accounting for 10-25% of total leaf N content. Moreover N2 fixation rates are highly correlated
with forest and foliage age due to the time needed for colonization of epiphylls on leaf surfaces
(Bentley 1987; Bentley 1984).
Dry deposition
Dry deposited materials originate from natural sources such as soil processes through microbial
activity and anthropogenic sources: combustion of fossil fuel and coal, biomass burning,
industrial plants, agriculture and animal waste (Lovett 1992). Particulate dry deposits are
classified in dust, sand, volcanic ash, sea salts, air pollutants and fertilizer and related ions are
Ca2+, Mg2+, Na+, Cl-, NO3-, NH4
+, SO42-, PO4
3-. Human activities are also responsible for notable
emissions of gases and aerosols such as SO2, NOx (NO and NO2), NH3 and HNO3. Furthermore,
Veneklaas (1990) and Kellman (1982) reported sporadic nutrient inputs to tropical ecosystems
through proximate volcanic activity.
Despite N2 being the most abundant single element in the atmosphere with about 78% by
volume and its key role in plant nutrition, it is not available for higher plants since it occurs in
the non-reactive form N2. As mentioned above some bacteria or algae can fix N2 and
incorporate it into organic matter though its contribution to total N fluxes within a forest-
ecosystem is generally considered to be low. The less abundant but reactive N forms, mainly
oxides (NO, NO2, N2O, HNO3 vapour, NO3-) but also reduced forms (NH4
+ and NH3), are crucial
in atmospheric chemical reactions. Dry deposition of N primarily occurs through gaseous HNO3
and NH3 because they are more reactive and are easily absorbed by the plant surfaces (Lovett
1992; Hill 2005). Moreover, Balestrini (2001) found for alpine forests a close correlation
between nitrate/ sulphate and ammonium/ hydrogen deposition and presumed that these ions
were mostly deposited as ammonium sulphate and nitrate aerosols resulting from ammonia
and sulphuric and nitric acid reactions. At tropical latidues land use change e.g. conversion of
forest in agricultural lands and increasing biomass burning are assumed to be responsible for
increased N emissions to the atmosphere.
A large fraction (40-50%) of deposited inorganic N was captured by the canopy of temperate
forests because of uptake by plants and epiphytic lichens and of microbial immobilization

- 9 -
(Garten 1998). This basically agrees with investigations from Clark (1998) who reported that
80% of NO3--N and 61% of NH4
+-N from atmospheric N deposition were retained by the canopy
in a tropical montane rain forest and in turn epiphytic bryophytes and its aggregations with
vascular epiphytes accounted for 80% of this retention. Experiments from Wilson (1998)
showed that uptake of wet deposited inorganic N made a small but sizeable contribution to the
total N demand of the vegetation. Moreover Garten (1998) suggested that the canopy uptake
of dry deposited N is a major contributor to N fluxes within the canopy. Hence throughfall
measurements underestimate the atmospheric deposition of N compounds to a large degree
(Hansen 1996).
The dry deposited material within or on the top of the canopy can enter the nutrient cycle of
the forest-ecosystem by several pathways: washing off of particulate substances and aerosols
from foliage surfaces by incident precipitation or absorption of gases through stomata and
cuticlefree spaces on tissue surfaces (ectodesmata) as well dissolution in the water film on the
leaf surface. The dissolved material then can either drip down as throughfall to the forest floor
or can be taken up by the leaves or associated epiphytes and epiphylls by ion exchange
processes. Dry deposition rates are mainly influenced by the duration of inter-event periods
and further by wind speed and turbulence, deposition velocity and the canopy structure such as
foliage characteristics, surface roughness and wetness (Lovett 1992).
1.5. Canopy exchange processes
In the last decades numerous studies paid attention to canopy exchange processes, focusing on
the extent to which they contribute to solute fluxes from the atmosphere to the soil surface
(Tobón 2004; Filoso 1999; Draaijers 1997; Hansen 1994; Lovett 1984; Lovett 1996; Lovett
1992). It has been proposed that under nutrient limited conditions the forest canopy exhibits
nutrient conservation mechanisms to secure nutrients for forest growth and production (Jordan
1980) and that epiphytes may play an important regulatory role in nutrient conservation in
tropical rain forests (Coxson 1995). Moreover, canopy effects on nutrient flux change during
forest succession because epiphyte loading increases with progressing succession. Various
methods were designed to obtain a deeper insight in canopy exchange processes and to
separate dry deposition from canopy exchange processes. The determination of their particular
contribution to throughfall fluxes has been attempted by several approaches such as labelling
of leachable nutrients with radioisotopes and recently with stable isotopes; collection of
throughfall and incident precipitation to estimate dry deposition (Hansen 1996; Lovett 1984;
Potter 1991) and mathematical modelling of canopy exchange budgets (Draaijers 1997).
However, controls of these processes such as wash-off of dry deposited solutes, leaching and

- 10 -
uptake of solutes and their contribution to the nutrient cycle in tropical rain forests (Tobón
2004) are still not well known.
Canopy structure
The forest canopy represents the whole plant community plus dead organic matter above
ground and is highly stratified because of trees of many sizes and different ages. The canopy
can be divided into overstorey, made up by the crowns of dominant and fully illuminated trees,
and understorey which comprises all woody plants in the lowest, shaded layer. The outer
canopy is the canopy surface adjacent to the atmosphere (Parker 1995). Forest canopies in
tropical rain forests are unique and highly diverse subsystems and support a rich flora vascular
and cryptogamic plant species. Forest canopies exhibit a great spatial variability, e.g. through
huge trees (emergents) which widely exceed the average level of the canopy (Whitmore 1998)
and gaps through falling trees. Gaps allow the penetration of light to the forest floor, resulting
in marked differences in microclimate, structure, and nutrient and water availability.
Epiphytes
The plants within the canopy of tropical rain forests possess different life-forms including trees,
epiphytes, hemiepiphytes, woody vines and epiphylls. Tropical rain forest canopies therefore
exhibit an associated high diversity of animals mainly insects. Arboreal plants so-called
epiphytes live or grow upon or attached to trees (phorophytes) without parasitizing them.
Epiphytes do not have access to terrestrial soils and their roots serve as anchorage to support
them on the host tree, although they are nutritionally independent of their host tree. Epiphytes
are a substantial canopy component in tropical humid rain forests and, with the associated
dead canopy organic matter, they represent a remarkable part of above-ground biomass and
nutrient pools (Grubb 1977; Nadkarni 1992). The development and abundance of epiphytes is
strongly related to water supply and they therefore occur primarily in environments with
abundant rainfall and short or no dry period (Coxson 1995). The main part of epiphytic
vegetation within the canopy of a tropical rain forest is composed of non-vascular plants such
as mosses, liverworts and lichens. In tropical rain forests the phyllosphere (leave surface) is
also colonized by so-called epiphylls which consist of algae, bacteria, fungi, ferns and
bryophytes and is considered as an own microcosm. On the basis of model simulations
Hölscher (2004) suggested that of 28% interception loss of incident precipitation epiphytic
bryophytes contributed only 6%. Nevertheless, epiphytic bryophytes can contribute
significantly to the water storage capacity of forest canopies through withholding rainwater as
they can store several times more water than those of their own dry weight (Hölscher 2004;

- 11 -
Nadkarni 1984). Non-vascular epiphytes and epiphylls are poikilohydric which means that they
equilibrate with environmental moisture therefore adequate water supply is pivotal to sustain
their metabolic activity since they are very sensitive to desiccation because of their lack of
roots, cuticula and epicuticular waxes. (Coxson 1991) reported that desiccated bryophytes are
highly susceptible to leaching during the initial rewetting phase. This resulted in a pulse release
of nutrients and organic solutes because of accumulation of sugars and polyols in intracellular
pools to stabilize membrane function during desiccation (Gupta 1977).
Epiphytes generally meet a large fraction of their water and nutrient demand from atmospheric
deposition. They sequester nutrients from allochthonous sources such as precipitation and dry
deposition and from N2-fixing microorganisms. Nutrients are stored intermittently and may be
released by drying and rewetting or displacement to the forest floor. Moreover, they can obtain
notable amounts of nutrients from wash off of dry deposited material and from autochthonous
sources such as leaching of foliar solutes of higher plants (Wania 2002). Vascular epiphytes
have different adaptations to capture nutrients: development of a root mat and morphological
and physiological adapted leaves to trap water and nutrients or plant detritus from
allochthonous and autochthonous sources. Therefore nutrient pools of epiphytes can be high
according to Nadkarni (1984). The nutrient capital retained by canopy epiphytes was similar to
that contained in canopy foliage in a tropical cloud forest. Apart from that a notable fraction of
dead organic material (mainly from epiphytes) decomposes in the canopy before reaching the
forest floor, resulting in a significant enrichment of mineral nutrients in throughfall and
stemflow (Veneklaas 1990; Nadkarni 1991). Nutrients from live or dead epiphytes are also
released into the nutrient cycles of forest ecosystems as epiphytic material falls in large clumps
to the forest floor through wind, disruption by animals, and the falling of supporting branches
and whole host trees (Coxson 1995; Nadkarni 1992). Moreover Wanek (2003; 2005)
demonstrated extensive bidirectional transfer of nutrients between epiphylls and their host
leaves and suggested that the phyllosphere plays a key role in controlling N fluxes, particularly
in the lower wet canopy of ravine forests.
Due to their dependence on water and nutrient supplies from the atmosphere epiphytes
indicate global change and climate more sensitively than co-occuring soil-rooted vegetation
(Benzing 1995). Especially nonvascular epiphytes such as lichens are sensitive to pollutant
loading through the atmosphere (Coxson 1995). Hence the notion that epiphytes enhance the
nutrient and water storage capacity in living aboveground vegetation, suggests that logging
and land use change may have strong effects on soil fertility and ecosystem function (Nadkarni
1984).

- 12 -
Canopy uptake and leaching processes occur along the same pathways and are primarily based
on passive processes: (1) ion diffusion along concentration gradients, (2) ion exchange
between surface water films and plant tissues, and (3) the absorption of gases through
stomata or epidermal cells via ectosdesmata (leaf surfaces without cuticula) (Draaijers 1997;
Hambuckers 1993). However, active processes may also occur such as guttation and active
nutrient uptake by epiphytes and tree foliage, and may have been underestimated so far.
Canopy uptake
Uptake of nutrients can occur via the cuticula and via stomata. For instance the uptake of NH3
and gaseous HNO3 by leaf tissue occurs through stomata and cuticles (adsorption to
epicuticular waxes), or via dissolution in water films on leaf surfaces. The fate of adsorbed
gases and their major sink is not clear and difficult to trace. Gases can be retained by plants
permanently or be dissolved, leached or re-emitted to the atmosphere (Draaijers 1997).
Uptake rates differ strongly between higher plants and epiphytic or epiphyllous cryptogams
where absorption rates of bryophytes were more than 20-fold higher compared to foliage
(Wanek 2003). Jordan (1980) estimated that nutrient uptake by plants from atmospheric
sources (wet and dry deposition) represents a nutrient conserving mechanism to enable the
ecosystem to survive on low fertility soils. Other retention mechanisms prevent or minimize
nutrient loss from plants such as sclerophylly (Jordan 1981; Vitousek 1986) and retranslocation
of foliar nutrients from senescent leaves to other plant parts before abscission (Vitousek 1986).
Numerous studies demonstrated that concentrations of inorganic N decrease as incident rainfall
passes through the canopy. This suggests that N is taken up by canopy components (Parker
1983). Wilson (1992; 1998) reported that foliar uptake as well as release of inorganic N in
coniferous forests is a function of N concentration in the foliage and that needles prefer the
uptake of NH4+ compared to NO3
-. The authors explained the NH4+ preference on the basis that
the cuticle surfaces have a net negative charge which attracts cations. Moreover, they showed
that an increase in ammonium concentration in simulated rain solution increased influx of
ammonium to the canopy.
Canopy leaching
Leaching is of widespread occurrence in nature and represents a passive process where plants
loose nutrients from their tissue (mainly leaves) to rainwater. (Tukey 1962) defined leaching as
the loss of inorganic and organic metabolites from above-ground plant parts by the leaching
action of aqueous solutions including rain, mist and dew. Inorganic nutrients including all
essential minerals and organic metabolites such as sugars, amino acids and organic acids and

- 13 -
were found to leach from foliage (Tukey 1966). The loss of nutrients occurs primarily through
passive processes such as diffusion and ion exchange between the water film on the plant
surface and a pool of exchangeable ions in the intercellular free space of foliage (Parker 1983;
Tukey 1970; Hambuckers 1993).
There are numerous factors, both internal and external to the plant, which influence the
leaching process: tissue age and type, foliage nutrient content and plant species. Furthermore
nutrients exhibit differences in their leachability depending on their function in metabolic
processes (Tukey 1970) and chemical characteristics (charge, size, valence, lipophily).
The susceptibility to loss increases with increasing maturity of plant tissue and reaches a
maximum at senescence. In addition Tukey (1970) reported that nutrients which are
accumulated in excess of requirements are leached more easily and rapidly after initial wetting.
Mature leaves possess larger pools of such exchangeable nutrients than young foliage because
the latter require and utilize these nutrients quickly for growth. Hence they are less susceptible
to leaching than older leaves. Moreover, characteristics of the tissue (leaf) surface affect
leaching as well. Sclerophylly as a morphological adaptation to reduce water loss via
transpiration contributes also to nutrient retention. For instance, the wettability of smooth and
waxy leaf surfaces is less than those of pubescent and unwaxed surfaces. Herbivore (insect)
activities within the canopy or any kind of injury of plant foliage can cause increased leaching
through damaged leaves. Moreover the efficiency of leaching is influenced by the amount,
intensity and chemical composition of precipitation. Foliage which is continuously wetted
through light rainfall is leached more easily than through short intensive rain events (Tukey
1970). In past 20-years it has been shown that deposition of air pollutants such as acid
deposition and ozone may also accelerate canopy leaching through cation exchange between
H+ and base cations and damage of cell membranes, respectively (Lovett 1992).
Foliar leaching is of major importance in K+- cycling due to the fact that throughfall often
exceeds litterfall as a K+- return pathway significantly. This is based on high apoplastic K+
concentrations in plant tissue, representing a large exchangeable pool (Sattelmacher 2001).
Ca2+ and Mg2+ are tightly bound in structural tissues or enzyme complexes and therefore leach
less extensively from the smaller exchangeable apoplastic pools in leaf foliage. The major
portion of calcium which is leached from plants derives from recently absorbed calcium in
exchangeable forms located outside the cell wall (Mecklenburg 1966). Calcium deficiency
through leaching has been shown to accelerate root uptake of calcium which then is
translocated into foliage and again is susceptible to leaching until it is immobilized through
incorporation into cell wall compounds.

- 14 -
1.6. Study aims
The present study aims at investigating the controls of throughfall chemistry such as dry
deposition, canopy leaching and canopy uptake and at determining to what extent the
respective canopy components contribute to canopy exchange processes. We hypothesized that
epiphytes, primarily epiphytic bryophytes, contribute most to the alteration in nutrient
throughfall chemistry. Since investigations of throughfall and bulk precipitation concentrations
alone do not provide further information on canopy exchange processes or the origin of
nutrients we performed tracer experiments to follow the pathway of macro-nutrients, especially
potassium, calcium and nitrogen. We assumed that nitrogen derives from dry deposition as well
as from canopy leaching and applied 15N stem injection in situ techniques and controlled
surface wash experiments to determine the origin of nitrogen. The study further intended to
quantify the gross and net fluxes of dissolved nitrogen, and its inorganic (NO3- and NH4
+) and
organic (DON) species within the canopy. To investigate gross fluxes we adopted a 15N-pool
dilution technique.
We further expected that the base cations, potassium and calcium, in throughfall originate from
autochthonous and allochthonous sources, respectively. Potassium, a highly mobile element
and important cytoplasmic solute is assumed to derive from canopy leaching whereas calcium
is supposed to be immobilized in structural plant tissues and therefore to originate mainly from
dry deposition and its wash off from foliage surfaces. To examine the sources of both base
cations to throughfall we used tracer injection experiments with Rb+ for K+ and Sr2+ for Ca2+,
respectively, as the tracers exhibit similar chemical characteristics as the tracees.
The experiments were conducted in a tropical wet lowland rain forest in the Piedras Blancas
National Park, Costa Rica.

- 15 -
REFERENCES
ATTIWILL, P. M. and ADAMS, M. A. (1993). "Tansley Review No. 50: Nutrient cycling in
forests." New Phytologist 124: 561-582.
BALESTRINI, R. and TAGLIAFERRI, A. (2001). "Atmospheric deposition and canopy
exchange processes in alpine forest ecosystems (northern Italy)." Atmospheric
Environment 35(36): 6421-6433.
BENTLEY, B. L. (1987). "Nitrogen fixation by epiphylls in a tropical rainforest." Annals of the
Missouri Botanical Garden 74: 234-241.
BENTLEY, B. L. and CARPENTER, E. J. (1984). "Direct transfer of newly-fixed nitrogen from
free-living epiphyllous microorganisms to their host plant." Oecologia 63: 52-56.
BENZING, D. H. (1995). Vascular epiphytes. Forest Canopies. M. E. Lowman and N. M.
Nadkarni. San Diego, Academic Press: 225-254.
CAVELIER, J., JARAMILLO, M., et al. (1997). "Water balance and nutrient inputs in bulk
precipitation in tropical montane cloud forest in Panama." Journal of Hydrology 193(1-
4): 83-96.
CHAPIN III, F. S., MATSON, P.A., MOONEY, H.A. (2002). "Principles of Terrestrial
Ecosystem Ecology." Springer-Verlag New York.
CHUYONG, G. B., NEWBERY, D. M., et al. (2004). "Rainfall input, throughfall and stemflow
of nutrients in a central African rain forest dominated by ectomycorrhizal trees."
Biogeochemistry 67(1): 73-91.
CLARK, K. L., NADKARNI, N. M., et al. (1998). "Atmospheric deposition and net retention of
ions by the canopy in a tropical montane forest, Monteverde, Costa Rica." Journal of
Tropical Ecology 14: 27-45.
COXSON, D. S. (1991). "Nutrient Release from Epiphytic Bryophytes in Tropical Montane
Rain-Forest (Guadeloupe)." Canadian Journal of Botany-Revue Canadienne De
Botanique 69(10): 2122-2129.
COXSON, D. S., MCINTYRE, D. D., et al. (1992). "Pulse release of sugars and polyols from
canopy bryophytes in tropical montane rain forest (Guadeloupe, French West Indies)."
Biotropica 24(2a): 121-133.
COXSON, D. S. and NADKARNI, N. M. (1995). Ecological roles of epiphytes in nutrient
cycles of forest ecosystems. Forest Canopies. M. E. Lowman and N. M. Nadkarni. San
Diego, Academic Press: pp. 495-543.
DRAAIJERS, G. P. J., ERISMAN, J. W., et al. (1997). "The impact of canopy exchange on
differences observed between atmospheric deposition and throughfall fluxes."
Atmospheric Environment 31(3): 387-397.

- 16 -
FILOSO, S., WILLIAMS, M. R., et al. (1999). "Composition and deposition of throughfall in a
flooded forest archipelago (Negro River, Brazil)." Biogeochemistry 45(2): 169-195.
FORTI, M. C. and MOREIRA-NORDEMANN, L. M. (1991). "Rainwater and throughfall
chemistry in a "Terra Firme" rain forest: central Amazonia." Journal of Geophysical
Research 96(D4): 7415-7421.
FORTI, M. C. and NEAL, C. (1992). "Hydrochemical Cycles in Tropical Rain-Forests - an
Overview with Emphasis on Central Amazonia." Journal of Hydrology 134(1-4): 103-
115.
GARTEN, C. T., SCHWAB, A. B., et al. (1998). "Foliar retention of 15N tracers: implications
for net canopy exchange in low- and high-elevation forest ecosystems." Forest Ecology
and Management 103(2-3): 211-216.
GRUBB, P. J. (1977). "Control of forest growth and distribution on wet tropical mountains:
with special reference to mineral nutrition." Annual Review of Ecology and Systematics
8: 83-107.
GUPTA, R. K. (1977). "A study of photosynthesis and leakage of solutes in relation to the
desiccation effects in bryophytes." Canadian Journal of Botany 55: 1186-1194.
HAMBUCKERS, A. and REMACLE, J. (1993). "Relative Importance of Factors Controlling the
Leaching and Uptake of Inorganic-Ions in the Canopy of a Spruce Forest."
Biogeochemistry 23(2): 99-117.
HANSEN, K. (1996). "In-canopy throughfall measurements of ion fluxes in Norway spruce."
Atmospheric Environment 30(23): 4065-4076.
HANSEN, K., DRAAIJERS, G. P. J., et al. (1994). "Concentration Variations in Rain and
Canopy Throughfall Collected Sequentially During Individual Rain Events." Atmospheric
Environment 28(20): 3195-3205.
HERRERA, R. (1978). "Direct Phosphorous Transfer from Litter to Roots."
Naturwissenschaften 65: 208-209.
HILL, K. A., SHEPSON, P. B., et al. (2005). "Measurement of wet deposition of inorganic
and organic nitrogen in a forest environment." Journal of Geophysical Research-
Biogeosciences 110(G2).
HÖLSCHER, D., KÖHLER, L., et al. (2004). "The importance of epiphytes to total rainfall
interception by a tropical montane rainforest in Costa Rica." Journal of Hydrology 292(1-
4): 308-322.
JOHNSON, D. W. (1992). Base cations. Atmospheric Deposition and Forest Nutrient Cycling.
D. W. Johnson and S. E. Lindberg. New York, Springer-Verlag. Ecological Studies 91: pp.
233-235.
JORDAN, C. F. (1971). "World Pattern In Plant Energetics." American Scientist 59(4): 425-&.

- 17 -
JORDAN, C. F., GOLLEY, F., et al. (1980). "Nutrient Scavenging of Rainfall by the Canopy of
an Amazonian Rain-Forest." Biotropica 12(1): 61-66.
JORDAN, C. F. and HERRERA, R. (1981). "Tropical Rain Forests - Are Nutrients Really
Critical." American Naturalist 117(2): 167-180.
KELLMAN, M., HUDSON, J., et al. (1982). "Temporal variability in atmospheric nutrient
influx to a tropical ecosystem." Biotropica 14: 1-9.
KLINGE, H. (1973). "Root mass estimation in lowland tropical rainforests of Central
Amazonia, Brazil. I. Fine root masses of a pale yellow latosol and a giant humus
podzol." Tropical Ecology 14: 29-38.
LOVETT, G. M. (1992). Atmospheric Deposition and Canopy Interactions of Nitrogen.
Atmospheric Deposition and Forest Nutrient Cycling. D. W. Johnson and S. E. Lindberg,
Springer-Verlag, New York. Ecological Studies 91: pp. 152-166.
LOVETT, G. M. and LINDBERG, S. E. (1984). "Dry deposition and canopy exchange in a
mixed oak forest as determined by analysis of throughfall." Journal of Applied Ecology
21: 1013-1027.
LOVETT, G. M. and LINDBERG, S. E. (1992). "Concentration And Deposition Of Particles And
Vapors In A Vertical Profile Through A Forest Canopy." Atmospheric Environment Part A-
General Topics 26(8): 1469-1476.
LOVETT, G. M., NOLAN, S. S., et al. (1996). "Factors regulating throughfall flux in a new
New-Hampshire forested landscape." Canadian Journal of Forest Research-Revue
Canadienne De Recherche Forestiere 26(12): 2134-2144.
LOVETT, G. M. and SCHAEFER, D. A. (1992). Canopy interactions of Ca2+, Mg2+, and K+.
Atmospheric Deposition and Forest Nutrient Cycling. D. W. Johnson and S. E. Lindberg,
Springer-Verlag. Ecological Studies 91: pp. 253-275.
MCDOWELL, W. H. (1998). "Internal nutrient fluxes in a Puerto Rican rain forest." Journal of
Tropical Ecology 14: 521-536.
MECKLENBURG, R. A., TUKEY, H. B. J., et al. (1966). "A mechanism for the leaching of
calcium from foliage." Plant Physiology 41: 610-613.
MONTAGNINI, F. and JORDAN, C. F. (2005). "Tropical Forest Ecology." Springer-Verlag.
NADKARNI, N. M. (1984). "Epiphyte biomass and nutrient capital of a neotropical elfin
forest." Biotropica 16(4): 249-256.
NADKARNI, N. M. and MATELSON, T. J. (1991). "Fine litter dynamics within the tree
canopy of a tropical cloud forest." Ecology 72(6): 2071-2082.
NADKARNI, N. M. and MATELSON, T. J. (1992). "Biomass and nutrient dynamics of
Epiphytic litterfall in a neotropical montane forest, Costa Rica." Biotropica 24(1): 24-30.
PARKER, G. G. (1983). "Throughfall and stemflow in the forest nutrient cycle." Advances in
Ecological Research 13: pp. 57-133.

- 18 -
PARKER, G. G. (1995). Structure and microclimate of forest canopies. Forest Canopies. M. D.
Lowman and N. M. Nadkarni, Academic Press: pp. 73-106.
POTTER, C. S., RAGSDALE, H. L., et al. (1991). "Atmospheric Deposition and Foliar
Leaching in a Regenerating Southern Appalachian Forest Canopy." Journal of Ecology
79(1): 97-115.
RAGSDALE, H. L., LINDBERG, S. E., et al. (1992). Atmospheric Deposition and Throughfall
Fluxes of Base Cations. Atmospheric Deposition and Forest Nutrient Cycling. D. W.
Johnson and S. E. Lindberg. New York, Springer-Verlag. Ecological Studies Vol. 91: pp.
235-275.
REINERS, W. A. and OLSON, R. K. (1984). "Effects of canopy components on throughfall
chemistry: an experimental analysis." Oecologia 63: 320-330.
SATTELMACHER, B. (2001). "Tansley review no. 22 - The apoplast and its significance for
plant mineral nutrition." New Phytologist 149(2): 167-192.
STARK, N. M. and JORDAN, C. F. (1978). "Nutrient retention by the root mat of an
Amazonian rain forest." Ecology 59: 434-437.
TOBÓN, C., SEVINK, J., VERSTRATEN, J. M. (2004). "Solute fluxes in throughfall and
stemflow in four forest ecosystems in northwest Amazonia." Biogeochemistry 70: 1-25.
TUKEY, H. B. (1966). "Leaching of Metabolites from above-Ground Plant Parts and Its
Implications." Bulletin of the Torrey Botanical Club 93(6): 385-&.
TUKEY, H. B. (1970). "The leaching of substances from plants." Annual Reviews in Plant
Physiology 21: 305-324.
TUKEY, H. B. J. and TUKEY, H. B. (1962). The loss of organic and inorganic materials by
leaching from leaves and other above-ground plant parts. Radioisotopes in Soil-Plant
Nutrition Studies. Vienna, Intern. Atmoic Energy Agency: pp. 289-302.
VENEKLAAS, E. J. (1990). "Nutrient fluxes in bulk precipitation and throughfall in two
montane tropical rain forests, Colombia." Journal of Ecology 78: 974-992.
VENEKLAAS, E. J. (1991). "Litterfall and nutrient fluxes in two montane tropical rain forests,
Colombia." Journal of Tropical Ecology 7: 319-336.
VITOUSEK, P. M. (1982). "Nutrient cycling and nutrient use efficiency." The American
Naturalist 119(4): 553-572.
VITOUSEK, P. M. (1984). "Litterfall, nutrient cycling, and nutrient limitation in tropical
forests." Ecology 65(1): 285-298.
VITOUSEK, P. M. and SANFORD, R. L. J. (1986). "Nutrient cycling in moist tropical forest."
Annual Reviews in Ecology and Systematics 17: 137-167.
WANEK, W. and PÖRTL, K. (2005). "Phyllosphere nitrogen relations: reciprocal transfer of
nitrogen between epiphyllous liverworts and host plants in the understorey of a lowland
tropical wet forest in Costa Rica." New Phytologist 166(2): 577-588.

- 19 -
WANEK, W., PÖRTL, K., et al. (2003). Effects of epiphyll colonization on phyllosphere
nutrient relations in a tropical wet forest, Costa Rica. Proceedings of the 2nd Symposium
of the A.F.W. Schimper-Foundation est. by H. and E. Walter. S.-W. Breckle. Stuttgart,
Verlag Günter Heimbach: 129-144.
WANIA, R., HIETZ, P., et al. (2002). "Natural N-15 abundance of epiphytes depends on the
position within the forest canopy: source signals and isotope fractionation." Plant Cell
and Environment 25(4): 581-589.
WHITMORE, T. C. (1998). An Introduction to Tropical Rain Forests, 2nd Edition. New York,
Oxford University Press.
WILSON, E. J. (1992). "Foliar Uptake and Release of Inorganic Nitrogen-Compounds in Pinus-
Sylvestris L and Picea-Abies (L) Karst." New Phytologist 120(3): 407-416.
WILSON, E. J. and TILEY, C. (1998). "Foliar uptake of wet-deposited nitrogen by Norway
spruce: An experiment using 15N." Atmospheric Environment 32(3): 513-518.

- 20 -
2. Manuscript
Mechanisms of nutrient dynamics in the canopy of a tropical lowland rain forest
ABSTRACT
Throughfall represents an important pathway for internal recycling and external input of
nutrients in tropical rain forests. The objectives of this study were (1) to differentiate
between the main processes, dry deposition and canopy exchange, that control throughfall
chemistry, (2) to assess gross canopy N fluxes, and (3) to investigate the effect of canopy
components on canopy N exchange fluxes by in situ tracer applications in a tropical wet
lowland rainforest, Costa Rica. The relative contribution of canopy exchange and dry
deposition to throughfall fluxes of K+, Ca2+, and N were investigated by application of Rb+,
Sr2+, and 15N as respective tracers via stem injection of subcanopy trees, sampling of
throughfall above and below the canopy and additional surface wash experiments of labelled
leaves. We further developed a static model and dynamic model to demonstrate the relative
contribution as unidirectional and bidirectional fluxes, respectively. Gross canopy N fluxes
(influx and efflux) and net flux of N forms ammonium, nitrate and DON within canopy-
related branches were assessed by adoption of the 15N-Pool dilution method. Throughfall
below subcanopy trees was enriched in all major inorganic solute nutrients (K+, Na+, NH4+,
Mg2+, Ca2+, SO42-, PO4
3-,NO3-, and Cl-) and organic compounds (DOC and DON). Significant
amounts of tracer were found in all trees and leachates with the exception of 15N for the
latter one. On the basis of a two-source mixing model foliage leaching of K+ and Ca2+
contributed to throughfall enrichment to 14-32% and 32%, respectively. Relative
contributions of dry deposition and canopy exchange to net throughfall fluxes calculated by
the dynamic model were 25-60% and 40-75%, respectively, for Potassium and 25% and
75%, respectively, for Calcium. Gross canopy NH4+ fluxes were significantly higher than net
NH4+ flux. Influxes of NO3
- and DON were significantly different from effluxes, resulting in
net uptake by the canopy. The highest quantity of incorporated 15N was found in epiphytic
bryophytes (16.4%) and highest 15N uptake rate was shown by the epiphyll-fraction. The
study demonstrates that tracer in situ applications allow to investigate the highly dynamic
and complex throughfall processes and to dissect the fractional contribution of dry
deposition and canopy exchange. We further show that small net fluxes are the result of
much larger gross influxes and effluxes of N compounds and canopy exchange is therefore
controlled by bidirectional exchange fluxes. The results further demonstrate that epiphytic
communities play a major role in solute fluxes in canopies and therefore in nutrient
dynamics of tropical rain forests.

- 21 -
Keywords: dry deposition, canopy exchange, net throughfall, 15N, rubidium, strontium, stem
injection, tropical lowland rainforest, canopy leaching, canopy uptake, 15N-Pool dilution,
epiphytic bryophytes
INTRODUCTION
The chemical composition of precipitation is generally altered during interception by the
forest canopy where resulting throughfall becomes enriched in major inorganic and organic
soluble compounds. In consideration of total nutrient fluxes throughfall provides a small but
important supplementary nutrient input from outside into forest ecosystems. However, it
represents a major pathway in internal nutrient recycling through transfer of soluble
nutrients from the canopy to the forest floor (Parker 1983; Potter et al. 1991; Cavelier et al.
1997; Filoso et al. 1999; Chuyong et al. 2004; Tobón 2004). The significance of this
throughfall pathway is highlighted in tropical rain forests with continuous high rainfall and
low nutrient availability where nutrient cycling is a key for sustaining the productivity of
forest ecosystems.
Two main processes control throughfall chemistry, i.e. (1) dry deposition and subsequent
wash-off of dry deposited aerosols, gaseous and particulate materials from canopy surfaces
(Lovett 1992) and (2) canopy exchange (uptake and leaching) processes of dissolved
substances between external surface solution and the apoplast (exchangeable pool) of plant
tissues (Parker 1983; Lovett and Lindberg 1984). The balance between these processes is
assumed to be highly dynamic. Canopy uptake and leaching preferably occurs through the
cuticle and is primarily based on passive processes along concentration gradients (diffusion
of ions or molecules, and ion exchanges) and to an unknown portion by active processes
that are energy-dependent and can operate against concentration gradients (Tukey 1966;
Tukey 1970; Hambuckers and Remacle 1993).
Moreover, throughfall fluxes, throughfall chemistry and controls thereof exhibit a high
temporal and spatial variability (Hansen 1996; Lovett et al. 1996) which is determined by
complex interactions of factors and processes of atmospheric, hydrological, chemical and
biological nature and human activities as well. The amount, duration and intensity of
incident rainfall (Parker 1983; Potter et al. 1991; Hansen et al. 1994), the length of
antecedent dry periods (Lovett et al. 1996; Tobón 2004), the proximity to external sources
(Veneklaas 1990), the forest structure, age and canopy composition (Nadkarni 1984;
Reiners and Olson 1984; Hölscher et al. 2003; Hölscher et al. 2004; Clark 2005), the
texture of interacting surfaces (Tukey 1970; Reiners and Olson 1984) and the nutrient
status of the forest (Parker 1983) have been demonstrated to affect throughfall chemistry
and fluxes.
In tropical rain forests, canopy dwelling plants (vascular epiphytes, bryophytes and lichens),

- 22 -
which make up a remarkable portion of the canopy, acquire notable amounts of their
nutrients from atmospheric sources and from internal sources such as foliar leachates of
host plants and litter (Wania et al. 2002; Wanek and Pörtl 2005). Further, they exhibit a
nutrient pool equivalent to that in canopy foliage considering their low contribution to total
forest biomass (Nadkarni 1984). This leads to the suggestion that epiphytes may have an
important impact on the nutrient dynamics in tropical forest ecosystems.
To correct nutrient fluxes for wet (bulk) deposition net throughfall (NTF) has been defined
on an basis (Lovett et al. 1996):
NTF = TF – WD = DD + CE
where TF is throughfall, WD the incident precipitation, DD dry deposition and CE describes
the canopy exchange. Positive rates of NTF indicate a net increase of soluble compounds or
elements in throughfall through wash-off of dry deposition and canopy leaching. Negative
rates of NTF imply a decrease of solute concentration in TF due to canopy uptake or
interception loss.
However, while wet deposition is relatively easy to measure, the processes
underlying NTF such as dry deposition and canopy exchange are difficult to quantify. In
recent years attempts (Potter et al. 1991; Hansen et al. 1994; Lovett et al. 1996; Clark et
al. 1998) were made to separate these processes and to quantify their fractional
contribution to NTF. This becomes the more relevant in view of global atmospheric changes
e.g. increasing deposition of N and P and its impact on terrestrial ecosystem functions since
wet and dry deposition constitutes an external atmospheric nutrient input to the ecosystem
(allochthonous source) while canopy exchange represents an internal transfer of nutrients
(autochthonous source) (Lovett et al. 1996).
Lovett and Lindberg (1984) put forward a multiple regression approach on the basis
of event throughfall and precipitation measurements to estimate the contribution of dry
deposition and canopy exchange in net throughfall. This approach is based on the
assumption that dry deposition is solely correlated to the length of the antecedent dry
period and canopy exchange exclusively occurs during rain event and therefore scales with
amount of event precipitation. This approach has been successfully applied to temperate
and boreal ecosystems but less so in tropical rainforests and doubts have been raised on
the validity of this approach. Dry deposition can be underestimated since material
accumulated between rain events is not washed off completely and therefore is not included
in the NTF or dry deposition will be overestimated as it includes secreted substances from
tissue surfaces and material accumulated in throughfall samplers during dry periods. The
canopy budget model was developed by Ulrich (1983) and applied by Draaijers and Erisman
(1995) where Na+ in throughfall was assumed to entirely derive from dry deposition. Since
the input of Na+ from sea salts exceeds by far any internal circulation and/ or internal
sources or sinks were thought to be negligible and Na+ was proposed as a conservative
tracer of dry deposited elements in the canopy. However, Wanek et al. (2007)

- 23 -
demonstrated a large contribution of canopy leaching to Na+ fluxes in net throughfall of
mangroves, and Na+ uptake has also been observed in the Esquinas forest, Costa Rica
(Hofhansl, 2007, pers. commun.). Therefore, the use of other, more adequate tracers, e.g.
stable and radioisotopes in situ may provide an opportunity to gain deeper knowledge into
the mechanisms of net throughfall chemistry.
Earlier tracer studies with the stable isotope 15N solely focused on nitrogen deposition and
foliar uptake of 15N in temperate and boreal forests after spraying tracer solutions on the
canopy or parts thereof (Bowden et al. 1989; Boyce et al. 1996; Garten et al. 1998; Wilson
and Tiley 1998) but they did not consider foliar leaching. The one exception was published
by Draaijers et al. (1997) where a 35S tracer experiment was conducted in a Douglas-fir
forest to estimate the amount of sulfate in TF originating from root-derived sulfate and
leached from foliage. They labelled the surrounding soils of trees with radioactive
ammonium 35S-sulfate and calculated the relative contribution of foliar leached sulfate to
sulfate in TF using 35S specific activity. However, no such tracer studies have been
undertaken to investigate the mechanisms of solute changes in throughfall in tropical
rainforests, where the complexity and heterogeneity of forest canopies by far exceeds that
of higher latitude forests.
The present study therefore aimed to differentiate between dry deposition and
canopy exchange of potassium, calcium and nitrogen, respectively, using tracer in situ
applications via stem injection. We used 15N as well-established tracer for 14N, Rb+ as tracer
for K+ and Sr2+ as tracer for Ca2+ fluxes since the tracer have been shown to behave
chemically and biologically similar as their respective investigated element. To derive the
relative contribution of DD and CE to net throughfall fluxes in a tropical lowland forest we
developed a static model and a dynamic model of canopy fluxes. While the former model is
based on net fluxes only (deposition, uptake, leaching) the latter model is based on bi-
directional exchange fluxes (influx, efflux) in the canopy that occur simultaneously and
therefore represent gross fluxes in contrast to net fluxes. Potassium and calcium were
chosen since throughfall in tropical rain forests is generally enriched in those major cations.
However, the major process for K+ enrichment is assumed to be foliar leaching while for
Ca2+ dry deposition is thought to contribute significantly (Lovett and Lindberg 1984;
Chuyong et al. 2004). This is based on their different chemical behaviour and biotic
compartmentation: K+ is highly mobile and occurs in high concentrations in exchangeable
pools of plant tissues (Sattelmacher 2001) whereas Ca2+ is predominantly incorporated in
stable pools of structural plant tissues such as cell walls (Mecklenburg et al. 1966). Nitrogen
shows a much more complex pattern of changes in throughfall, owing to more diverse forms
of N such as ammonium, nitrate, and low- and high-molecular weight organic compounds
and an additional input process, i.e. biological fixation of atmospheric N2. N has been shown
to be taken up mostly in inorganic forms and to be released in organic forms. We therefore
hypothesized that the major source of K+ and Ca2+ are leaching and dry deposition,
respectively, and that inorganic N will be taken up by the canopy.

- 24 -
For the first time a 15N Pool dilution method was adopted to assess the gross N fluxes within
the canopy. As canopy exchange processes are assumed to be highly dynamic, particularly
in the case of N, we hypothesized that high rates of gross N fluxes (influx to and N efflux
from the canopy) occur behind rather small net N fluxes. We further expected that
epiphytes, predominantly epiphytic bryophytes, are the most important canopy component
affecting canopy exchange processes, due to their lack of a cuticle and large abundance.
The experiments were conducted in a humid tropical lowland rain forest in the Piedras
Blancas National Park, Costa Rica.
MATERIALS AND METHODS
Study site
This study was performed in the Esquinas forest of the Piedras Blancas National Park, which
is located close to the Golfo Dulce on the southern Pacific coast of Costa Rica (8°42’46’’ N,
83°12’90’’ W). The site is classified as tropical wet lowland rainforest (Holdridge 1967) with
altitudes from 0 to 597 m a.s.l and possesses narrow ridges, steep slopes with dense
drainage networks and valleys interspersed with meandering rivers. Average annual bulk
precipitation from 1998 to 2006 was 5720 mm (recorded at the Tropical Field Station La
Gamba) with a short, weak dry season from January to March (monthly mean precipitation
approx. 180 mm), and high rainfall from September to November (monthly mean about 800
mm). Throughfall (2005 to 2006) constituted 90.6% and 87.4% of total rainfall in the
primary ravine forest and secondary ravine forest, respectively (Hofhansl, 2007, pers.
commun.). Mean annual temperature between 1997 and 2001 was 27.4°C (monthly
average from 23.2°C to 31.5°C) at the Tropical Field Station La Gamba, which is next to the
Esquinas Rainforest, and 25.2°C (monthly average from 22.3°C to 28°C) inside the forest
(Weissenhofer and Huber 2001). According to Pamperl (2001) the prevailing soils are, in the
order of occurrence, ulitsols, inceptisols and entisols.
The experiments were conducted in an undisturbed primary ravine forest and a secondary
ravine forest (age 25 yrs) the latter one has been used as a cacao plantation.
Tracer techniques
Preliminary tests were carried out in February 2005; the main experiments were conducted
in September and October 2005. For the tracer experiments two different labelling
techniques were developed and performed: (1) stem injection of 15N, Rb+ and Sr2+ for
subcanopy trees (5-12 m height) along with a controlled leaf surface wash experiment and
(2) application of 15N-enriched simulated rain events (spraying on the canopy) to study
gross and net nitrogen fluxes on selected host tree branches that are densely covered with
vascular and cryptogamic epiphytes.

- 25 -
(1) Stem injection of 15N, Rb+ and Sr2+
Plant material and experimental setup
For the stem injection approach appropriate understorey tree species with easily reachable
and adequately developed crown, low or no epiphytic colonization and a sufficient number of
plans were selected at the two forest sites, namely Psychotria elata (Sw) Hammel
(Rubiaceae) in the primary ravine forest and Siparuna thecaphora (Poepp. & Endl.) A. DC.
(Monimiaceae) in the secondary ravine forest. Four trees of S. thecaphora were treated with 15N- and Rb+/Sr2+-solution and each four trees of P. elata were labelled either with 15N- or
Rb+/Sr2+-solution.
To inject the tracer solutions into the xylem channels stems were drilled with a borer (inner
diameter = 4 mm) to about 3 mm depth and a short metal tube was fitted into the hole.
The tube was connected by a flexible PVC tube to a 20 ml syringe reservoir (plastic) which
was attached above the hole. The reservoir and connecting tubes were filled with 20 mL
labelling solution (bubble free) and the reservoir was covered with parafilm and fixed to the
plant stem with tape. The 15N-labelling solution was made of Na15NO3, 15NH4Cl and
15NH415NO3 (equimolar) with a total N concentration of 60 mM and 98 at % 15N and the
Rb+/Sr2+-solution consisted of RbCl (100 mM) and SrCl2 (50 mM). Rb+ has been shown to
behave (bio-) chemically similar to K+ (Nyholm and Tyler 2000) and Rb-radioisotopes have
e.g. been used to study K+ uptake kinetics and fluxes in roots. Ca2+ and Sr2+ have been
applied to infer ecosystem Ca2+ sources and dynamics. We therefore used rubidium as
tracer for potassium and strontium as tracer for calcium fluxes within the canopy of
understorey trees. During the treatment period (two weeks) the solution was replenished
and the amount taken up by the plant was recorded daily.
Sampling
Sampling of rain water above and below the treated plants started after two weeks of tracer
amendment and included 3 rain events (< 10 mm to minimize dilution of ion concentrations
in throughfall) for each plant in short intervals. Water samples (throughfall above the
canopy and below the canopy) were collected in four replicates (plants), i.e. four trees
labelled by stem injection, per rain event. Each throughfall sampler was constructed from
two gutters (0.75 m length) fixed above and beneath the canopy of the sample tree on a
bamboo-pillar. Both gutters were provided with an outlet where the incident rainfall was
channelled through flexible PVC tubes down into plastic flasks (1.5 L). The flasks were
emptied, cleaned carefully and washed with deionized water after each rain event during
the sampling period. Aliquots of bulk precipitation samples from the open field at the Field
Station La Gamba were also taken. The collected rain water (approx. 100 mL each) was
poured into PE flasks and HgCl2 was added (0.1 mM final concentration) to avoid microbial
growth and alteration of nutrient content in throughfall. The water samples were kept frozen
until transport to Vienna.

- 26 -
Leaf samples of each tree were collected before starting the labelling and after the labelling
period, between the two throughfall samplers after every collected rain event. One half of
fresh leaves was treated in a microwave oven to stop enzymatic activities and further dried
at 60 °C for 72 h. The other half of labelled leaf samples was used for controlled leaching
experiments. For this three to five leaves of each leaf sample per plant and event were put
into a plastic bag with zip and a defined amount (100 mL to 200 mL, depending on leaf
size) of deionized water (>18.2 MΩ cm-1) was added. After two hours the leaching
experiment was stopped and water with leachates was collected in scintillation flasks and
treated similar as throughfall samples. The fresh leached leaves were dried superficially,
scanned to determine leaf area and finally treated in the microwave oven and dried as
described above.
(2) Application of 15N-enriched simulated rain events
Plant material and experimental setup
For the 15N-enriched rainwater application branches (1.0-1.5 m length) with a typical load of
vascular epiphytes, epiphytic bryophytes and epiphylls on host leaves from the primary
ravine forest were selected. Two horizontal branches of Alchornea cf.costaricensis Pax & K.
Hoffm. (Euphorbiaceae) and three horizontal branches of Ficus tonduzii Standl. (Moraceae)
were cut and hung horizontally under a tin roof at the forest edge in the garden of the
Tropical Field Station La Gamba to avoid uncontrolled contact with incident rainfall. The cut
surface of the host branch was covered with water-soaked tissue in a plastic bag taped to
the branch to minimize water stress of host leaves.
Rainwater was collected on an event basis in a big PVC barrel (200 L) in the open field, each
before a simulated rain event as far as it was raining. In intervening periods with no rainfall
the barrel was covered by a plastic foil and put into the shadow to minimize chemical
alteration. The first simulated rain event applied was enriched with the stable isotope 15N in
equimolar amounts of NO3-, NH4
+ and DON (0.55 µM 15NH415NO3/
15N-L-glutamic acid
equivalent to 10 at% 15N). The following simulated rain events were conducted with
unlabelled rainwater. Under each branch a plastic cover was fixed to ensure complete
throughfall collection. The dripped off throughfall was channelled and filtered through small
mesh nylon into polyethylene canisters. Before starting the experiment the branches were
sprayed with unlabelled rainwater to test water retention and the amount of water required
to penetrate the whole branches and its canopy components. The amount was depending on
the load size of epiphytes (1.5-2.5 L). To provide enough time for solute exchange every
single simulated rain event was applied in two aliquots of 6 to 14 min duration with an
interval of about one hour in between. The branches were sprayed evenly with a defined
amount of rainwater using a pressure sprayer to simulate drizzle. Simulated rain events
were conducted twice a day in constant intervals over a period of four to five days. This was

- 27 -
the maximal possible time before the host leaves started to loose turgor. Separate
equipment was used for 15N labelled rain event and normal rain events, respectively, and
the equipment was washed and cleaned carefully after each rain event.
Sampling
To quantify the amount of rainwater applied the remaining volume within the pressure
sprayer was weighed. After a drip off time (two hours) the volume of throughfall collected in
the polyethylene canisters was measured. Aliquots (250 mL) of rainwater and throughfall
filtered through fine mesh nylon were transferred into PE bottles after each rain event,
HgCl2 was added (0.1 mM final concentration) and samples were kept frozen until transport.
At the end of the experiment the branches were divided into different canopy components:
vascular epiphytes, epiphytic bryophytes, epiphytic lichens, epiphylls, host leaves, canopy
soil, canopy litter, bark and wood. Unlabelled reference material of canopy components and
host leaves were sampled from other branches of the same trees at the original place. The
fractions were treated in a microwave to stop microbial or enzymatic activities and the
material further dried at 60 °C for 72 h. Dry weight of each fraction was determined and
samples were weighed and packed airtight until transport to Vienna.
15N-Pool-Dilution approach
Pool dilution techniques are commonly used to estimate gross N transformation rates in
soils (Luxhoi 2004). For instance to measure gross N mineralization the NH4+-pool is
labelled by addition of 15NH4+. N mineralization produces NH4
+ from an unlabelled organic N
pool, therefore adds 14NH4+ to the pool and ultimately leads to dilution of the 15N:14N signal
in the NH4+-pool. In contrast, processes that consume NH4
+ such as microbial uptake
(immobilization), plant uptake or nitrification, do not change the 15N:14N signature of this
pool and utilize NH4+ at the 15N:14N ratio present in the NH4
+-pool.
This method was transferred to gross canopy fluxes of N. The N fractions (NO3-, NH4
+ and
DON) of rainwater before canopy contact represent the pools which are labelled with 15N.
During penetration through and interception by the canopy the pools are altered. Inputs of 14N may occur through canopy leaching or N2 fixation processes and 15N:14N losses may
happen due to canopy uptake processes. Determination of isotope signatures and N
concentrations enabled the quantification of gross and net N canopy fluxes. Furthermore, 15N dilution is studied along a time axis of e.g. 4 to 24 hours. In the approach transferred to
canopy processes the time axis is represented by rainwater before and after canopy
exchange and rates are therefore presented on an event basis and not per hour or day.

- 28 -
Fig.1: Scheme of the 15N-Pool-Dilution approach for (A) determination of gross N mineralization rates and (B)
transferred to gross canopy fluxes of N; WD: wet deposition, TF: throughfall
Chemical analyses
Chemical analyses were carried out in Vienna, Department of Chemical Ecology and
Ecosystem Research, University of Vienna.
Stable isotope analyses
Dried organic materials such as leaves, bark and epiphytes of both experiments were
redried at 80°C for 24h and ground to a fine powder in a ball mill (Retsch MM2 and MM200).
Aliquots of 1-2 mg were weighed into Sn-capsules and 15N and total N content were
analysed by continuous-flow IRMS (isotope ratio mass spectrometer). The system was
made up of an elemental analyser (EA 1110, CE Instruments, Milan, Italy) which was
connected to the IRMS (DeltaPLUS, Finnigan MAT, Bremen, Germany). Reference gas (high
purity N2, AGA, Vienna, Austria) was calibrated to the atmospheric N2 standard (at-air)
using IAEA-NO3, IAEA-N-1, and -2 reference material (International Atomic Energy Agency,
Vienna, Austria).
Aliquots (50 mL) of rain samples (throughfall above and below the canopy, and leachates)
of the stem injection experiment were therefore evaporated to dryness with a rotary
evaporator (R-114/ R-124, Büchi, Switzerland). Samples were redissolved in 0.5 mL
distilled water, dried again using a SpeedVac concentrator (SC 110, Savant, NY) and
redissolved in 100 µL distilled water. Aliquots (50 µL) were dried in Sn-capsules and
analysed for 15N and total N content by IRMS.
Water samples of the simulated rain application tracer experiment
For stable isotope analyses rain and throughfall samples of the 15N-enriched rain event (i.e. 15N-Pool dilution approach) and the consecutive two unlabelled rain events were used. Rain
and throughfall samples of the 15N-enriched rain event and the first unlabelled rain event
Mineralization
Consumption
+ 15N addition
+ 14N
NH4+ pool
in soil
- 15N:14N
15N:14NH4+
(A)
Canopy Leaching
Canopy Uptake
+ 15N addition
+ 14N
RainwaterWD & TF
- 15N:14N
15N:14NH4+
15N:14NO3-
15N:14DON
(B)

- 29 -
were fractionated into the N forms ammonium, nitrate and dissolved organic nitrogen
(DON). Water samples of the second unlabelled rain event were treated as mentioned
above. Small aliquots of original samples were analysed for anions and cations by HPLC (DX
500, Dionex, Vienna, Austria) and total dissolved N and DOC concentration by NPOC/TDN-
analyzer (Shimadzu, Japan) as well.
Isolation of N fractions
According to Hertenberger and Wanek (2004) the microdiffusion approach to isolate
ammonium can be combined with common cation-exchange chromatography to separate
the organic nitrogen fraction from nitrate. Sample volumes were determined, pH was
adjusted to 5-6 with 0.1 N HCL to avoid volatile loss of NH3 or NO2 during evaporation, and
then samples were concentrated under reduced pressure to about 10 to 15 ml with a rotary
evaporator and weighed again. The concentrates were poured into Schott-flasks (50 mL)
and acid traps were added. Acid traps were made of quartz fibre filter discs containing 12 µL
2N H2SO4 which were enclosed in a strip of Teflon band that is gas permeable but
waterproof. After addition of MgO (100 mg) to increase pH >9.0, the bottles were
immediately closed and NH3 released into the head space was captured by the floating acid
traps. The flasks were placed on a shaker for 5-7 days (75 rpm, 37°C). Then acid traps, still
enclosed in Teflon band, were removed and dried over conc. H2SO4 under vacuum
(24-48 h). The dry filter discs were transferred into Sn-capsules shortly before IRMS-
analysis to avoid corrosion of the capsules. The remaining concentrates containing nitrate
and DON were filtered to remove MgO and pH was adjusted to 5-6. Samples were then
applied onto cation-exchange resin columns (2-3 mL DOWEX 50Wx8, 50-100 mesh,
H+-form, Fluka). The columns were washed two times with 10 mL distilled water to collect
nitrate in the flow-through and the eluate was neutralized with 0.1 N KOH. The nitrate
fractions were evaporated to dryness redissolved in 0.5 mL distilled water, again dried by
SpeedVac and redissolved in 70 µL distilled water. Samples were quantitatively transferred
and dried in Sn-capsules and analysed for 15N and total N content by IRMS. The adsorbed
DON fractions were eluted from the resin with 25 mL 1 N HCL and treated as the nitrate
fractions after addition of 100 µL 1 N KOH.
Ion, NPOC and TDN analyses
Hot-water extracts were prepared by extracting aliquots of 20 mg ground leaf material with
1 mL deionized water at 95°C for 60 min. Inorganic cations (K+, Rb+, NH4+, Na+, Ca2+, Sr2+,
Mg2+) and anions (Cl-, NO3-, SO4
2-, PO43-) in rain samples (bulk precipitation, throughfall
above and below the canopy, and leachates) and hot-water extracts of the stem injection
experiment were analysed by HPLC (high pressure liquid chromatography, DX 500, Dionex,
Vienna, Austria) and conductivity detection. Anions were separated on an anion exchange
column (AS11, 4x250 mm, Dionex) using a linear KOH gradient (2 to 40 mM in 6 min, total
run time 12 min). Cations were separated on a cation exchange column (CS16, 5x250 mm,

- 30 -
Dionex) by an isocratic method with methanesulfonic acid as eluent (28 mM, 45 min,
60 °C). Rain water samples of the simulated rain events were analysed for anions (Cl-, NO3-,
SO42-, PO4
3-) as mentioned above and but method to analyse cations (K+, NH4+, Na+, Ca2+,
Mg2+) were modified (30 mM methanesulfonic acid for 26 min and 40 °C).
NPOC (non purchable organic carbon) and TDN (total dissolved nitrogen) of rain samples
(bulk precipitation, throughfall above and below the canopy from both experiments) were
determined using a TOC-VCPH/CPN/ TNM-1 analyzer (Shimadzu, Japan). Inorganic carbon was
automatically removed during measurement through addition of 2 N HCl and purging with
synthetic air (CO2 free). DON was calculated by subtracting DIN (dissolved nitrate and
ammonium) from measured TN.
Calculations
Enrichment factor for the stem injection experiment
The enrichment factor (EF) for TFb (throughfall below the experimental plant) was
calculated as:
(i) EF = c(TFb)/ c(TFa)
where c is the concentration of the respective solute in TFb and TFa (throughfall above the
experimental plant)
Two-source mixing model for the stem injection experiment
The fractional contribution of leaching from host tree foliage to throughfall below the canopy
(TFb) was estimated for potassium, calcium and nitrogen with a modified two-source mixing
model:
(ii) % PLEA = 100 x (RTFa - RTFb)/ (RTFa – RLEA)
where PLEA represents the relative fraction of potassium, calcium or nitrogen, leached from
foliage of labelled woody plants to throughfall below the canopy (TFb), R the ratios of
tracers e.g. Rb+:K+, Sr2+:Ca2+ or 15N:14N in TFa, TFb, and leachates from controlled
leaching-experiment (LEA). In consideration of the dynamic model CE (canopy exchange)
and DD (dry deposition) are estimated in absolute numbers:
(iii) CE = TFb x % PLEA = TFa x EF x % PLEA
For TFa an arbitrary concentration of 1 is assumed and CE represents the relative
contribution of canopy exchange to TFb. Consequential the relative contribution of DD to
TFb can be estimated as:

- 31 -
(iv) DD = TFb – (TFa + CE)
15N-Pool-Dilution equation
The equations to determine gross N mineralization or nitrification from Bengston (2006)
were adopted and modified to calculate gross N canopy fluxes:
ln ((fTF-k)/(fWD-k))ln (WTF/WWD)
(v) Efflux = x WWD - WTF
ln ((fTF-k)/(fWD-k))ln (WTF/WWD)
(1 + ) x WWD - WTF(vi) Influx =
where k represents the natural abundance of 15N in rainwater (at% 15Nunlabelled WD), (fWD-k)
the APE (atom percent excess) of 15N in applied rainwater (wet deposition, WD), (fTF-k) the
APE of 15N in throughfall (TF), WWD the initial N pool in wet deposition (in µg N), and WTF the
N pool in TF (in µg N) of the respective fractions i.e. ammonium, nitrate or DON. Therefore,
each flux was calculated separately for NH4+, NO3
- and DON.
Statistical analyses
Statistical analysis was performed with STATGRAPHICS PLUS 5.0. Software (Statistical
Graphics Corp., Rockville, MD, USA). Differences between three or more groups were tested
by one-way analysis of variance (ANOVA) (multiple range test, LSD test) (P <0.05).
RESULTS
Stem injection experiment
The stem injection experiment was performed to examine the potential contribution of
subcanopy trees to nutrient enrichments in throughfall through foliage leaching. In
preliminary tests in February 2007 we observed that stem injection was an effective
labelling method for the understorey trees P. elata (representative for secondary forest) and
S. thecaphora (representative for primary forest). Foliar, hot-water extractable
concentrations of rubidium and strontium increased 15-fold and 1.5-fold, respectively,
compared to unlabelled plants, and δ15N values ranged from 80 to 2000 (data not shown).
During the main experiment conducted in September and October 2007 a
considerable enrichment of rubidium, strontium, and 15N in leaves was reached by stem
injection (Fig.2, closed bars). Moreover controlled leaching experiments were carried out to
determine tracer leaching efficiency of labelled leaves. Similar ratios were detected for

- 32 -
Rb+:K+ and Sr2+:Ca2+ in leachates (LEA) compared to leaves (L) in S. thecaphora (Fig. 2D
and 2E). P. elata exhibited a markedly lower leaching efficiency for all tracers applied, as
also shown for 15N in S. thecaphora (Fig 2A, 2B, 2C, and 2F).
Ratios of tracers and their corresponding elements (Rb+:K+, Sr2+:Ca2+ and 15N:14N) in bulk
precipitation, collected in the open field (P), were slightly higher in comparison to those in
throughfall collected above the canopy of treated plants (TFa), though no significant
differences were observed.
Comparison of tracer:element ratios of TFa and TFb (throughfall below the canopy) showed
no statistically significant differences. Enrichment factors for solutes in throughfall (TFb) are
shown in Table 1. Concentration of all determined solutes were enriched in TFb with
greatest enrichment factors in PO43- (3.17) followed by K+ (2.52) and the other solutes
(1.18 - 1.96). Interestingly EF was lowest for all N solutes, Na+ and DOC (Table 1).
Based on concentration ratios of Rb+:K+, and Sr2+:Ca2+, respectively, and on a modified
two-source mixing model we estimated the fractional contribution of leaf leachates from P.
elata and S. thecaphora to throughfall (TFb) enrichment of K+ and Ca2+ (Fig. 2A, 2D, and
2E). The two sources of K+ (Rb+) and Ca2+ (Sr2+) to TFb are TFa and LEA. This approach
was only used where the two sources, TFa and LEA were significantly different i.e. Rb+:K+ in
P. elata and S. thecaphora and Sr2+:Ca2+ in S. thecaphora (Fig. 2A, 2D, and 2E). We
assumed that TFb was composed of TFa and LEA and from dry deposition (DD). The ratios
of Rb+:K+ and Sr2+:Ca2+, respectively, in LEA represented the maximum possible
contribution of foliage leaching of woody plants to TFb, while the respective ratios in TFa
represent the minimum contribution by LEA. The results showed that S. thecaphora
considerably contributed to throughfall enrichment of both potassium and calcium via
foliage leaching i.e. 14% and 32%, respectively. P. elata contributed 32% of K+ to TFb by
leaching.
Though 15N:14N ratios were significantly higher in bulk leaves than in all other fractions the
tracer (15N) did not show up in leachable pools at the time of experiment (Fig. 2C, 2F). The 15N:14N ratios therefore did not differ significantly between LEA, P, TFa, and TFb for both P.
elata and S. thecaphora and therefore hindered the application of the two source model to
evaluate N sources in throughfall (see also Sr2+:Ca2+ in P. elata, Fig, 2B).
Simulated rain experiment
Net fluxes
A total of five branches (A. cf. costaricensis plus F. tonduzzi) where treated with simulated
rain events twice a day over a period of 3 days where the first applied event was enriched
with the stable isotope 15N. Net fluxes were calculated as volume-weighted concentrations
of TF minus those of WD (wet deposition). Negative net fluxes indicate net leaching of

- 33 -
canopy components while positive net fluxes refer to net uptake. Time dependent course of
event based net fluxes of N fractions (NO3-, NH4
+, DON – dissolved organic nitrogen), base
cations (K+, Mg2+, Ca2+ and Na+), anions (SO42-, PO4
3-) and DOC (dissolved organic carbon)
showed a high variability throughout the experiment but no correlation among ions and
fractions were evident, respectively (data not shown). Thus we calculated the sum of net
fluxes of seven events conducted within three days (Fig. 3). To increase the comparability
between branches event based and integrated net fluxes were calculated on the basis of the
sum of dry weights of potentially canopy components contributing to net solute exchange
(bark, canopy litter, canopy soil, epiphylls, vascular epiphytes, leaves, lichens, and epiphytic
bryophytes) not including wood.
Net fluxes of DOC were more than ten-fold higher compared to all other solutes and showed
the highest increase in throughfall (102 ± 42 µmol g-1 DW 3 d-1) with a range from 262 to
29 µmol g-1 DW 3 d-1 and therefore was excluded from statistical analysis of the other
solutes.
Net fluxes of N fractions indicated a net uptake by the canopy and mean values differed not
significantly with 0.29 ± 0.28, 0.82 ± 0.15 and 0.33 ± 0.69 µmol g-1 DW 3 d-1 for nitrate,
ammonium and DON, respectively.
All base cations exhibited a net loss (leaching) from the canopy. Potassium concentration
significantly increased from precipitation to throughfall (-7.1 ± 2.1 µmol g-1 DW 3 d-1)
followed by calcium and magnesium (-2.02 ± 0.29, -0.79 ± 0.37 µmol g-1 DW 3 d-1,
respectively). Canopy exchange of sodium and chloride resulted in net uptake (1.22 ± 0.79
and 6.2 ± 1.2 µmol g-1 DW 3 d-1, respectively). In contrast anions such as sulfate and
phosphate showed a weak net canopy leaching (-0.21 ± 0.10 and -0.21 ± 0.06 µmol g-1 DW
3 d-1, respectively).
Gross N fluxes
A 15N-Pool dilution assay was performed to determine gross canopy N fluxes i.e. efflux and
influx of NH4+, NO3
-, and DON (Fig. 4) and their integral i.e. net fluxes were assessed as
well. Overall all three fractions showed significant higher influx than efflux which resulted in
net canopy uptake. Furthermore, influx of nitrate and DON did not differ significantly from
net fluxes with the exception of ammonium. Both rates of NH4+-influx to and -efflux from
the canopy were significantly higher than net flux and were of same magnitude (-0.160 ±
0.040 and 0.156 ± 0.048 µmol 15N g-1 DW, respectively). The resulting net ammonium
uptake rate was therefore almost zero (0.004 ± 0.008 µmol 15N g-1 DW). Net uptake rates
for NO3- and DON were 0.019 ± 0.011 and 0.081 ± 0.035 µmol 15N g-1 DW, respectively.
Gross fluxes of nitrate were markedly lower compared to those of ammonium and DON i.e.
6-fold lower for efflux and up to 15-fold lower for influx. Statistically differences of N-
fractions ammonium, nitrate and DON in gross canopy fluxes (influx, efflux) and net N

- 34 -
uptake are shown in Table 2. Efflux and net N uptake of NH4+ significantly differ from those
of NO3- and DON.
Canopy components
The mean total biomass of the five branches averaged 137 ± 17 g dry weight whereof wood
accounted for 64.4% but was excluded from further calculations through negligible
contribution to exchange processes of solutes: <0.1% of the initial 15N-amount was found in
wood. The partitioning of dry biomass into different canopy components is shown in Figure
5A. The greatest fraction was made up of leaves (L) and bryophytes (M) which exhibited
similar quantities with 19.3 ± 5.6 g and 17.2 ± 4.0 g, respectively. In addition they
significantly differed from all other fractions, which ranged from 0.5 ± 0.2 to 5.0 ± 3.6 g in
descending order: vascular epiphytes (Ev), epiphylls (Eph), canopy litter (CL), bark (Bk),
canopy soil (CS), and lichens (Lich).
The recovery of 15N, applied by the first simulated rain event, in throughfall and canopy
components, amounted to 69.8%. Throughfall, the first three events were measured for 15N, accounted for almost two thirds of the 15N detected (43.3%). About 96% of throughfall 15N was found in throughfall collected during the first 15N-enriched rain event. The
partitioning of 15N within the canopy is shown in Figure 5B in percent of initial 15N amount
applied in rainwater. The highest quantity of incorporated 15N was determined in epiphytic
bryophytes with 16.4% or 0.2 µmol 15N. All other canopy fractions did not differ significantly
although epiphylls and leaves exhibited somewhat higher 15N contents (3.3% and 3.2%,
respectively) than residual fractions whose proportions ranged from 1.3% to 0.4% in the
descending order: bark, canopy litter, canopy soil, vascular epiphytes and lichens. 15N uptake rates of canopy components were estimated (Fig. 5C) but in contrast to
distribution of 15N within the biomass, the epiphyll-fraction showed the highest uptake rates
(0.033 ± 0.013 µmol 15N g-1 DW after 3 days). Bark, canopy litter, vascular epiphytes,
lichens and mosses incorporated 15N from incident rainwater at a lower rate (0.016 ± 0.008
to 0.012 ± 0.005 µmol 15N g-1 DW) compared to epiphylls. Despite leaves made up the
highest proportion of dry mass their uptake rates were lowest (0.001 µmol 15N g-1 DW).
DISCUSSION
Models to demonstrate controls of throughfall chemistry
We developed two models to interpret alterations in throughfall chemistry and their control
mechanisms (Fig. 6). On the one hand we adopted a static model which is based on
unidirectional net fluxes (NTF) alone (Fig. 6, left plot) which can be either negative or
positive, or zero. A negative NTF of a solute corresponds to a decrease in its concentration

- 35 -
after penetration of the canopy (Fig. 6, A) and is generally caused through uptake
processes within the canopy. By contrast an increase in nutrient concentrations in
throughfall results in positive net throughfall fluxes and is caused by foliar leaching and/ or
dry deposition (Fig. 6, C). Apart from nutrient depletions or enrichments in throughfall
solute fluxes of incident rainfall (TFa) and throughfall (TFb) may be balanced and the
resulting net flux is zero (Fig. 6, B) although exchange processes take place within the
canopy. In this case, investigations of net nutrient fluxes are inadequate to explain the
nutrient dynamics within the canopy. Canopy exchange in situations of zero net fluxes can
only be demonstrated by tracer studies.
Thus, we additionally put forward a dynamic model (Fig. 6, right plot) which implies
alterations in throughfall chemistry and concentrations through bidirectional fluxes i.e.
foliar leaching and foliar uptake. For instance, the concentration of a solute may not
change during penetration of the canopy, therefore the net nutrient flux is zero. However,
the solute might be taken up by some canopy components and be released by others at
the same time. According to the extent of canopy exchange fluxes, the dynamic model is
illustrated by three different scenarios: scenario one (Fig. 6, sc1) indicates no contribution
of exchange processes to throughfall chemistry, in scenario two (Fig. 6, sc2) 50% of
solutes are exchanged and scenario three (Fig. 6, sc3) shows a complete exchange of a
nutrient in rainwater during canopy passage.
To investigate these exchange processes and given that net fluxes do not provide answers
to the origin of soluble nutrients in throughfall, we conducted tracer experiments for
selected nutrients, namely potassium, calcium, and nitrogen, to follow their pathways
within the canopy. Tracers used were Rb+, Sr2+, and 15N that behave (bio-) chemically
similar to the tracee. Stem injection tracer experiments along with controlled foliar
leaching experiments, may allow differentiating whether nutrient enrichment in throughfall
resulted from dry deposition or foliage exchange and to what extent (Draaijers et al.
1997).
We demonstrate four cases in the stem injection experiment (Fig. 7) where tracer:element
ratios of throughfall (TFb) are compared to the corresponding ratios of throughfall above
the labelled canopy (TFa) and foliar leachates (LEA). Case 1 does not allow any derivation
of the fractional contribution of dry deposition or foliage exchange to TFb enrichment since
no significant amounts of tracer were found in leachates. The remaining cases (Fig. 7, case
2-4) allow detecting potential foliage exchange effects on throughfall chemistry. In case 2
ratios of TFb and TFa do not differ although the tracer was found in LEA. This suggests that
foliar exchange does not contribute to TFb chemistry. Net positive fluxes of potassium,
calcium or nitrogen in throughfall therefore are controlled by dry deposition and for
nitrogen by N2-fixation. Further this case agrees with scenario one from the dynamic
model. In case 3, TFb is obviously tracer enriched (higher ratio) due to foliar exchange

- 36 -
contributing about half to TFa but the contribution of dry deposition can not be estimated
with this approach and fits with scenario 2 of the dynamic model. The model can be
expanded based on enrichment factors (EF) and tracer:element ratios to derive the fraction
of canopy exchange (CE) and dry deposition (DD) (see (i) and (ii)). Finally in case 4,
according to scenario three of the dynamic model, throughfall chemistry is replaced
completely by foliar uptake and -leaching.
Throughfall enrichment
Measurements of throughfall chemistry below the canopy of understorey trees (P. elata
and S. thecaphora) in a tropical lowland forest showed an increase of all examined solutes
and therefore positive net throughfall fluxes (Table 1). We hypothesized that TF
enrichment of potassium was controlled by foliar leaching, whereas calcium derived mainly
from dry deposition and nitrogen originated from both foliar leaching and dry deposition,
and from N2-fixation.
In three cases of the stem injection experiment (Fig. 2A, 2D and 2E) injected rubidium and
strontium was found in leachates therefore allowing to estimate the contribution of canopy
exchange (leaching) to TFb for potassium and calcium in the understorey tree S.
thecaphora (in the primary ravine forest) and for potassium in P. elata (in the secondary
forest). According to the static model the relative contribution of foliar leaching to
potassium and calcium in throughfall below S. thecaphora canopies was 14% and 32%,
respectively, and for potassium in throughfall under P. elata 32% (see equation (ii)).
Considering that net leaching of solutes to throughfall is the result of canopy exchange
processes their contribution was evaluated by the dynamic model (see equation (iii) and
(iv)). K+ in throughfall under P. elata seems to derive to almost equivalent parts from foliar
exchange (4/9) and dry deposition (5/9) whereas potassium and calcium concentrations in
throughfall under S. thecaphora were controlled by canopy exchange (3/4). In the case of
P. elata results of the tracer experiment are contradicting to our hypothesis as it is
generally agreed that potassium derives mainly from canopy leaching due to its extreme
mobility and high apoplastic K+ concentration in plant tissues (Parker 1983; Sattelmacher
2001). Comparative data for dry deposition fluxes of potassium were found for temperate
forests and tropical rain forests. For a mixed hardwood forest (Lindberg et al. 1986)
estimated that dry deposition contributed about 60% of potassium as coarse particles.
Lovett and Lindberg (1984) suggested that dry deposited K+ derived from suspended soil
or biological material. Moreover they observed highest K+-aerosol concentrations from
within the forest itself indicating an in-canopy source.
Calcium in throughfall is considered to almost equally originate from wash off of particulate
dry deposition (Lovett and Lindberg 1984; Lindberg et al. 1986; Forti and Neal 1992) and
foliar leaching (Tukey 1970; Parker 1983). However, we observed that a large portion

- 37 -
derives from foliage leaching. Calcium is generally tightly bound in structural plant tissue,
e.g. cell walls, and leaching of calcium mostly occurs from exchangeable pools in leaves,
where calcium has been recently accumulated. Moreover, mature tissues are more
susceptible to leaching than younger leaves since the latter accumulate calcium directly
within cell walls and have a smaller exchangeable pool for calcium (Mecklenburg et al.
1966). Though Hansen et al. (1994 and 1996) reported that dry deposition rather has an
impact on throughfall fluxes in the upper canopy layers, where trees are highly exposed to
turbulent air movements, our results indicate an effect of dry deposition on K+ and Ca2+ in
the understorey.
In the other three cases the stem injection experiment failed, including those for nitrogen
for both species (Fig. 2B, 2C and 2F). We therefore could not clarify whether increased
concentrations of nitrogen and calcium in TFb were caused by dry deposition, foliar
exchange or in the case of nitrogen by atmospheric nitrogen fixation as well. Although the
tracer injection was successful and tracer was allocated to foliage, this signal was not
found in foliar leachates. It is therefore likely that the injected tracers, 15N and strontium,
were incorporated and immobilized in stable pools within the plant, e.g. cell walls and
proteins, and therefore were removed from the exchangeable pool, the apoplast solution,
during the two weeks of injection.
Consequently this experiment showed that foliar leaching from understorey woody plants
contributes to throughfall enrichment of potassium and calcium by 40-75% in a lowland
tropical rain forest. Nevertheless, the experimental setup was not suitable to investigate N
exchange processes within the canopy and to determine whether canopy impacts are
controlled by unidirectional (static model) or bidirectional (exchange) fluxes (dynamic
model). Though we were able to estimate net canopy fluxes (DD, leaching) the underlying
gross fluxes i.e. canopy uptake and canopy leaching, could not be studied by this
approach. Therefore we adopted the 15N isotopic pool dilution approach, generally used to
determine gross N mineralization rates in soils (Bengston 2006), to detect gross canopy N
fluxes, or more precisely influxes to and effluxes from the canopy, for different N species,
namely ammonium, nitrate and DON (dissolved organic nitrogen). The terms efflux and
influx instead of uptake and leaching, respectively are more appropriate as we were not
able to determine dry deposition of nitrogen on canopy surfaces. In this approach
rainwater labelled with 15N the tracer is diluted by canopy leaching (efflux) and “consumed”
by canopy uptake processes (influx).
The 15N pool dilution experiment clearly demonstrated that high rates of gross N fluxes of
solutes occur across canopy components in tropical rainforests even though net fluxes are
minor or negligible (see Fig. 4, NH4+). Moreover, this also supports the dynamic model
therefore pointing to a greater importance of canopy exchange processes than evident

- 38 -
from net flux measurements alone. Canopies obviously are exchanging nutrients in a
bidirectional way (Wanek et al. 2003; Wanek and Pörtl 2005) and at higher rates than
previously anticipated which is analogy to foliar CO2- and H2O- exchanges that by far
exceed net CO2 assimilation and water losses (Heldt 1998; Helliker and Griffiths 2007).
Additionally the 15N-pool dilution approach revealed a net uptake of all examined N species,
ammonium, nitrate and DON (Fig. 3 and 4). This is in contrast to previous studies (Coxson
1991; Lovett and Lindberg 1993; Filoso et al. 1999; Wania et al. 2002) where throughfall
measurements in tropical forests showed that inorganic N, nitrate and ammonium, were
indeed taken up by the canopy, but DON leached from the canopy. Filoso et al. (1999)
suggested that large enrichments of DON in throughfall resulted from retention and
assimilation of deposited inorganic N on canopy surfaces, mainly by epiphytic algae, and
subsequent washing off or leaching as DON during later rain events. Further Coxson
(1991) reported that nutrients, mainly organic compounds, were released to a great extent
by epiphytic bryophytes. Due to the fact that bryophytes often provide a habitat for N2-
fixing bacteria and the occurrence of lichens with cyanobacterial symbionts (Bentley 1987)
release of N from atmospherically fixed nitrogen might provide another source of DON
within the canopy. On the basis of our data we cannot solve this discrepancy. DON such as
amino acids are readily taken up by plants as an N source which plants can immediately
utilize. Since in this experiment labelled DON was provided in the form of 15N-Glutamic
acid, canopy components such as cryptogamic epiphytes might utilize added DON in
rainwater. Although the N was supplied in a single pulse similar results were found for
three days of treatment with simulated rain events with ambient rainwater (Fig. 3) and for
a whole year period (Hofhansl, pers. commun.).
Even though net uptake of ammonium and nitrate did not differ (Table 2) gross influx of
ammonium (and DON) were about ten times higher than influx of nitrate. This may be due
to negatively charged sites in cell walls or cuticles e.g. carboxyl groups of pectins which
affect the passage of ions through the apoplast. The movement of ions in cell walls
(apoplast) is characterized by electrostatic interactions leading to preferred uptake of
cations (NH4+, positive charged amino acids) and repulsion of anions (NO3
-) (Clarkson
1993; Marschner 1995; Sattelmacher 2001).
At the first sight, the data with regard to net N fluxes from the second tracer experiment
disagree with results from the stem injection tracer experiment, since in the latter
experiment weak net leaching of ammonium, nitrate, and DON was found. However, it has
to be considered that the investigated canopies of the stem injection experiment only
consisted of foliage and small branches of woody plants and did not exhibit any visual
epiphytic or epiphyllous colonization whereas the branches of the second experiment were
richly covered with epiphytic bryophytes and vascular epiphytes and host tree leaves were

- 39 -
colonized by epiphylls. Thus, results of both experiments can not be directly compared
together but clearly demonstrate that epiphytic communities obviously play a major role in
solute fluxes in canopies and therefore in nutrient dynamics of tropical rain forests. This is
consistent with other studies (Nadkarni 1984; Chuyong et al. 2004) which observed higher
retention of N within the canopy with greater abundance of epiphytic bryophytes. Nadkarni
(1984) determined for a tropical cloud forest that the epiphyte biomass constituted less
than two percent of total forest dry mass but retained nutrients equivalent to 45% of that
in forest foliage and therefore might act as a “buffer” or an additional nutrient pool for
previous inaccessible sources and later release to other canopy components.
Net throughfall fluxes of DOC (dissolved organic carbon) were substantially higher in the
second experiment than in the first experiment. This might indicate that epiphytes also
have an important effect on DOC fluxes (Table 1 and Fig.3). According to other throughfall
studies in tropical wet forests (Coxson et al. 1992; Möller 2005; Schwendenmann 2005)
DOC generally shows the highest fluxes and concentrations in throughfall compared to
other solutes (Parker 1983) and implies leaching of organic compounds from both higher
and lower plant tissues such as sugars, polyols and organic acids (Filoso et al. 1999); pulse
release of these compounds were observed by Coxson et al. (1992) from desiccated
epiphytic bryophytes during initial rewetting events.
The 15N-pool dilution experiment also allowed evaluating net 15N of various canopy
components. Dry mass of the colonized branches mainly consisted of their leaves and
epiphytic bryophytes (mosses) (Fig. 5A). In fact, the main part of the epiphytic community
was made up of bryophytes which absorbed the greatest fraction of the applied 15N tracer
(16%, Fig. 5B) whereas host leaves did not retain more as other fractions (< 5%). Clark et
al. (1998) reported that epiphytic bryophytes and epiphytic assemblages accounted for
80% retention of atmospheric deposited inorganic N and Clark (2005) estimated that 33-
67% of wet deposited inorganic N was retained by these canopy components in a tropical
montane forest. 15N-uptake rates of canopy components were highest for epiphylls (mainly
comprised bryophytes), intermediate for other epiphytes (vascular epiphytes, lichens,
mosses), and lowest for tree leaves. Even bark, canopy soil and canopy litter showed net 15N retention. Wilson and Tiley (1998) determined in a Norway spruce forest that uptake of
N by bark was 2-10 times higher than in needles, due to more permeable barriers and
greater wettability. Bryophyte gametophytes are virtually “barrier-free”, due to the lack of
a cuticle and epicuticular waxes, which facilitates active and passive ion exchange
processes. Similar results were observed by Wanek and Pörtl (2005) in a 15N-pulse
labelling experiment where epiphyllous bryophytes quickly incorporated 15N. Nevertheless,
epiphytic and epiphyllous bryophytes exhibit rapid turnover times (Coxson 1991) and
therefore do not act as a long term storage for nutrients. Moreover, concerning their
poikilohydric character, desiccated bryophytes and lichens loose large quantities of

- 40 -
nutrients in the first minutes during rewetting (Gupta 1976; Coxson 1991). The lack of
epiphytes and epiphylls in canopies of the stem injection experiments might possible
explain why nitrogen was released in throughfall and emphasizes that different canopy
components may show very different characteristics with respect to solute exchange.
Wanek et al. (2003) reported that plant species may exhibit large differences in foliar
solute exchange with the environment as they showed extensive N exchange between
specific host plant species and epiphylls when plants were 15N labelled.
Net fluxes of other cations and anions of both experiments are consistently with other
throughfall studies (Forti and Neal 1992; Cavelier et al. 1997; Filoso et al. 1999; Chuyong
et al. 2004) with exception of chloride and sodium in the second experiment. Previously it
has been assumed that Na+ (and Cl-) behaves conservatively in tree canopies i.e. show not
net uptake or leaching and therefore can be used as a tracer for dry deposition (Hansen
1996; Draaijers et al. 1997). This has been the basis of the canopy budget model (Ulrich
1983) to estimate the relative contribution of dry deposition and canopy exchange to net
throughfall. Net uptake of Na+ on a short term and an annual basis (Fig. 3; and Hofhansl,
pers. commun) as well as significant leaching of Na+ in mangrove canopies cast significant
doubt on the applicability of the canopy budget model. However, the controls and
pathways of sodium and chloride remain obscure in this study.
In conclusion the study demonstrates that (1) in situ application of tracers allows to
investigate the fractional contribution of external sources (dry and wet deposition) and
internal sources (canopy exchange) of solutes in throughfall, (2) that canopy exchange is
controlled by bidirectional fluxes i.e. canopy efflux and canopy influx, and (3) epiphytic
communities play a major role in nutrient dynamics in canopies of tropical rain forests.

- 41 -
REFERENCES
BENGSTON, P., FALKENGREN-GRERUP, U., BENGTSSON, G. (2006). "Spatial
distributions of plants and gross N transformation rates in a forest soil." Journal of
Ecology 94: 754-764.
BENTLEY, B. L. (1987). "Nitrogen fixation by epiphylls in a tropical rainforest." Annals of
the Missouri Botanical Garden 74: 234-241.
BOWDEN, R. D., GEBALLE, G. T., et al. (1989). "Foliar uptake of 15N from simulated
cloud water by red spruce (Picea rubens) seedlings." Canadian Journal of Forest
Research 19(382-386).
BOYCE, R. L., FRIEDLAND, A. J., et al. (1996). "Direct canopy nitrogen uptake from
15N-labeled wet deposition by mature red spruce." Canadian Journal of Forest
Research 26: 1539-1547.
CAVELIER, J., JARAMILLO, M., et al. (1997). "Water balance and nutrient inputs in bulk
precipitation in tropical montane cloud forest in Panama." Journal of Hydrology
193(1-4): 83-96.
CHUYONG, G. B., NEWBERY, D. M., et al. (2004). "Rainfall input, throughfall and
stemflow of nutrients in a central African rain forest dominated by ectomycorrhizal
trees." Biogeochemistry 67(1): 73-91.
CLARK, K. L., NADKARNI, N. M., et al. (1998). "Atmospheric deposition and net
retention of ions by the canopy in a tropical montane forest, Monteverde, Costa
Rica." Journal of Tropical Ecology 14: 27-45.
CLARK, K. L., NADKARNI, N.M., GHOLZ, H.L. (2005). "Retention of Inorganic Nitrogen
by Epiphytic Bryophytes in a Tropical Montane Forest." Biotropica 37: 328-336.
CLARKSON, D. T. (1993). "Roots and the Delivery of Solutes to the Xylem." Philosophical
Transactions of the Royal Society of London Series B-Biological Sciences 341(1295):
5-17.
COXSON, D. S. (1991). "Nutrient Release from Epiphytic Bryophytes in Tropical Montane
Rain-Forest (Guadeloupe)." Canadian Journal of Botany-Revue Canadienne De
Botanique 69(10): 2122-2129.
COXSON, D. S., MCINTYRE, D. D., et al. (1992). "Pulse release of sugars and polyols
from canopy bryophytes in tropical montane rain forest (Guadeloupe, French West
Indies)." Biotropica 24(2a): 121-133.
DRAAIJERS, G. P. J. and ERISMAN, J. W. (1995). "A canopy budget model to assess
atmospheric deposition from throughfall measurements." Water, Air, and Soil
Pollution 85: 2253-2258.
DRAAIJERS, G. P. J., ERISMAN, J. W., et al. (1997). "The impact of canopy exchange
on differences observed between atmospheric deposition and throughfall fluxes."
Atmospheric Environment 31(3): 387-397.

- 42 -
FILOSO, S., WILLIAMS, M. R., et al. (1999). "Composition and deposition of throughfall
in a flooded forest archipelago (Negro River, Brazil)." Biogeochemistry 45(2): 169-
195.
FORTI, M. C. and NEAL, C. (1992). "Hydrochemical Cycles in Tropical Rain-Forests - an
Overview with Emphasis on Central Amazonia." Journal of Hydrology 134(1-4): 103-
115.
GARTEN, C. T., SCHWAB, A. B., et al. (1998). "Foliar retention of 15N tracers:
implications for net canopy exchange in low- and high-elevation forest ecosystems."
Forest Ecology and Management 103(2-3): 211-216.
GUPTA, R. K. (1976). "Physiology of Desiccation Resistance in Bryophytes - Intraspecific
Differences in Acrocladium-Cuspidatum (Hedw) Lindb." Indian Journal of
Experimental Biology 15(11): 1074-1075.
HAMBUCKERS, A. and REMACLE, J. (1993). "Relative Importance of Factors Controlling
the Leaching and Uptake of Inorganic-Ions in the Canopy of a Spruce Forest."
Biogeochemistry 23(2): 99-117.
HANSEN, K. (1996). "In-canopy throughfall measurements of ion fluxes in Norway
spruce." Atmospheric Environment 30(23): 4065-4076.
HANSEN, K., DRAAIJERS, G. P. J., et al. (1994). "Concentration Variations in Rain and
Canopy Throughfall Collected Sequentially During Individual Rain Events."
Atmospheric Environment 28(20): 3195-3205.
HELDT, H.-W. (1998). Pflanzenbiochemie. Heidelberg, Spektrum Akademischer Verlag.
HELLIKER, B. R. and GRIFFITHS, H. (2007). "Toward a plant-based proxy for the
isotope ratio of atmospheric water vapor." Global Change Biology 13(4): 723-733.
HERTENBERGER, G. and WANEK, W. (2004). "Evaluation of methods to measure
differential N-15 labeling of soil and root N pools for studies of root exudation." Rapid
Communications in Mass Spectrometry 18(20): 2415-2425.
HOLDRIDGE, L. R. (1967). Life zone ecology. San Jose, Costa Rica, Tropical Science
Center.
HÖLSCHER, D., KÖHLER, L., et al. (2003). "Nutrient fluxes in stemflow and throughfall in
three successional stages of a upper montane rain forest in Costa Rica." Journal of
Tropical Ecology 19: 557-565.
HÖLSCHER, D., KÖHLER, L., et al. (2004). "The importance of epiphytes to total rainfall
interception by a tropical montane rainforest in Costa Rica." Journal of Hydrology
292(1-4): 308-322.
LINDBERG, S. E., LOVETT, G. M., et al. (1986). "Atmospheric deposition and canopy
interactions of major ions in a forest." Science 231: 141-145.
LOVETT, G. M. (1992). Atmospheric Deposition and Canopy Interactions of Nitrogen.
Atmospheric Deposition and Forest Nutrient Cycling. D. W. Johnson and S. E.
Lindberg, Springer-Verlag, New York. Ecological Studies 91: pp. 152-166.

- 43 -
LOVETT, G. M. and LINDBERG, S. E. (1984). "Dry deposition and canopy exchange in a
mixed oak forest as determined by analysis of throughfall." Journal of Applied
Ecology 21: 1013-1027.
LOVETT, G. M. and LINDBERG, S. E. (1993). "Atmospheric deposition and canopy
interactions of nitrogen in forests." Canadian Journal of Forest Research-Revue
Canadienne De Recherche Forestiere 23: 1603-1616.
LOVETT, G. M., NOLAN, S. S., et al. (1996). "Factors regulating throughfall flux in a new
New-Hampshire forested landscape." Canadian Journal of Forest Research-Revue
Canadienne De Recherche Forestiere 26(12): 2134-2144.
LUXHOI, J., NIELSEN, N.E., JENSEN, L.S. (2004). "Effect of soil heterogeneity on gross
nitrogen mineralization measured by 15N-pool dilution techniques." Plant and Soil
262: 264-275.
MARSCHNER, H. (1995). Mineral Nutrition of Higher Plants. London, Academic Press.
MECKLENBURG, R. A., TUKEY, H. B. J., et al. (1966). "A mechanism for the leaching of
calcium from foliage." Plant Physiology 41: 610-613.
MÖLLER, A., KAISER, K., GUGGENBERGER, G. (2005). "Dissolved organic carbon and
nitrogen in precipitation, throughfall, soil solution, and stream water of the tropical
highlands in northern Thailand." Journal of Plant Nutrition and Soil Science-
Zeitschrift Fur Pflanzenernahrung Und Bodenkunde 168: 649-659.
NADKARNI, N. M. (1984). "Epiphyte biomass and nutrient capital of a neotropical elfin
forest." Biotropica 16(4): 249-256.
NYHOLM, N. E. I. and TYLER, G. (2000). "Rubidium content of plants, fungi and animals
closely reflects potassium and acidity conditions of forest soils." Forest Ecology And
Management 134(1-3): 89-96.
PAMPERL, S. (2001). Soils in the Golfo Dulce region. An Introductory Field Guide to the
Flowering Plants of the Golfo Dulce Rain Forests, Costa Rica. A. Weber, W. Huber, A.
Weissenhofer, N. Zamora and G. Zimmermann. Linz, Stapfia, Biologiezentrum des
OÖ Landesmuseum. 78: 28-33.
PARKER, G. G. (1983). "Throughfall and stemflow in the forest nutrient cycle." Advances
in Ecological Research 13: pp. 57-133.
POTTER, C. S., RAGSDALE, H. L., et al. (1991). "Atmospheric Deposition and Foliar
Leaching in a Regenerating Southern Appalachian Forest Canopy." Journal of Ecology
79(1): 97-115.
REINERS, W. A. and OLSON, R. K. (1984). "Effects of canopy components on throughfall
chemistry: an experimental analysis." Oecologia 63: 320-330.
SATTELMACHER, B. (2001). "Tansley review no. 22 - The apoplast and its significance for
plant mineral nutrition." New Phytologist 149(2): 167-192.
SCHWENDENMANN, L., VELDENKAMP E. (2005). "The Role of Dissolved Organic
Carbon, Dissolved Organic Nitrogen, and Dissolved Inorganic Nitrogen in a Tropical
Wet Forest Ecosystem." Ecosystems 8: 339-351.

- 44 -
TOBÓN, C., SEVINK, J., VERSTRATEN, J. M. (2004). "Solute fluxes in throughfall and
stemflow in four forest ecosystems in northwest Amazonia." Biogeochemistry 70: 1-
25.
TUKEY, H. B. (1966). "Leaching of Metabolites from above-Ground Plant Parts and Its
Implications." Bulletin of the Torrey Botanical Club 93(6): 385-&.
TUKEY, H. B. (1970). "The leaching of substances from plants." Annual Reviews in Plant
Physiology 21: 305-324.
ULRICH, B. (1983). Interaction of forest canopies with atmospheric constituents: SO2,
alkali and earth alkali cations and chloride. Effects of Accumulation of Air Pollutants
in Forest Ecosystems. B. Ulrich and J. Pankrath. Dordrecht, The Netherlands, Reidel:
33-45.
VENEKLAAS, E. J. (1990). "Nutrient fluxes in bulk precipitation and throughfall in two
montane tropical rain forests, Colombia." Journal of Ecology 78: 974-992.
WANEK, W., HOFMANN, J., et al. (2007). "Canopy interactions of rainfall in an off-shore
mangrove ecosystem dominated by Rhizophora mangle (Belize)." Journal of
Hydrology 345: 70-79.
WANEK, W. and PÖRTL, K. (2005). "Phyllosphere nitrogen relations: reciprocal transfer
of nitrogen between epiphyllous liverworts and host plants in the understorey of a
lowland tropical wet forest in Costa Rica." New Phytologist 166(2): 577-588.
WANEK, W., PÖRTL, K., et al. (2003). Effects of epiphyll colonization on phyllosphere
nutrient relations in a tropical wet forest, Costa Rica. Proceedings of the 2nd
Symposium of the A.F.W. Schimper-Foundation est. by H. and E. Walter. S.-W.
Breckle. Stuttgart, Verlag Günter Heimbach: 129-144.
WANIA, R., HIETZ, P., et al. (2002). "Natural N-15 abundance of epiphytes depends on
the position within the forest canopy: source signals and isotope fractionation." Plant
Cell and Environment 25(4): 581-589.
WEISSENHOFER, A. and HUBER, W. (2001). Basic geographical and climatic features of
the Golfo Dulce region. An Introductory Field Guide to the Flowering Plants of the
Golfo Dulce Rain Forests, Costa Rica. A. Weber, W. Huber, A. Weissenhofer, N.
Zamora and G. Zimmermann. Linz, Stapfia, Biologiezentrum des OÖ Landesmuseum.
78: 11-14.
WILSON, E. J. and TILEY, C. (1998). "Foliar uptake of wet-deposited nitrogen by Norway
spruce: An experiment using 15N." Atmospheric Environment 32(3): 513-518.
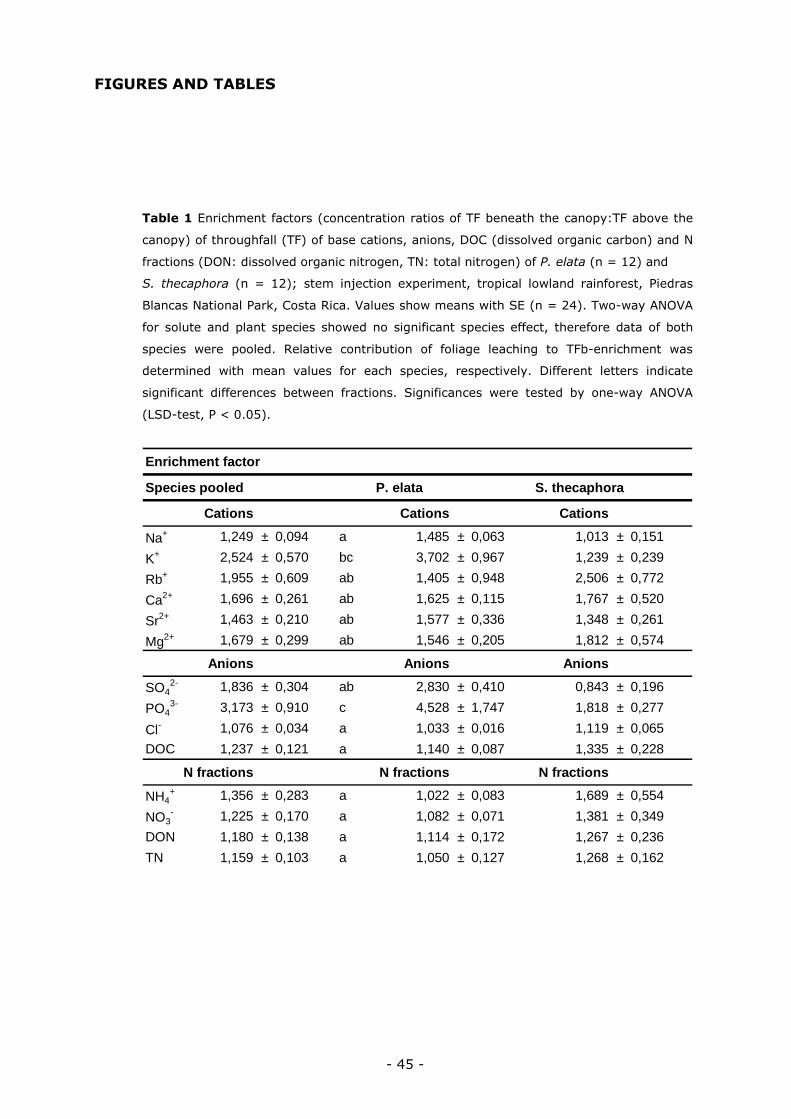
- 45 -
FIGURES AND TABLES
Table 1 Enrichment factors (concentration ratios of TF beneath the canopy:TF above the
canopy) of throughfall (TF) of base cations, anions, DOC (dissolved organic carbon) and N
fractions (DON: dissolved organic nitrogen, TN: total nitrogen) of P. elata (n = 12) and
S. thecaphora (n = 12); stem injection experiment, tropical lowland rainforest, Piedras
Blancas National Park, Costa Rica. Values show means with SE (n = 24). Two-way ANOVA
for solute and plant species showed no significant species effect, therefore data of both
species were pooled. Relative contribution of foliage leaching to TFb-enrichment was
determined with mean values for each species, respectively. Different letters indicate
significant differences between fractions. Significances were tested by one-way ANOVA
(LSD-test, P < 0.05).
Enrichment factor
Species pooled P. elata S. thecaphora
Cations Cations Cations
Na+ 1,249 ± 0,094 a 1,485 ± 0,063 1,013 ± 0,151
K+ 2,524 ± 0,570 bc 3,702 ± 0,967 1,239 ± 0,239
Rb+ 1,955 ± 0,609 ab 1,405 ± 0,948 2,506 ± 0,772
Ca2+ 1,696 ± 0,261 ab 1,625 ± 0,115 1,767 ± 0,520
Sr2+ 1,463 ± 0,210 ab 1,577 ± 0,336 1,348 ± 0,261
Mg2+ 1,679 ± 0,299 ab 1,546 ± 0,205 1,812 ± 0,574
Anions Anions Anions
SO42- 1,836 ± 0,304 ab 2,830 ± 0,410 0,843 ± 0,196
PO43- 3,173 ± 0,910 c 4,528 ± 1,747 1,818 ± 0,277
Cl- 1,076 ± 0,034 a 1,033 ± 0,016 1,119 ± 0,065
DOC 1,237 ± 0,121 a 1,140 ± 0,087 1,335 ± 0,228
N fractions N fractions N fractions
NH4+ 1,356 ± 0,283 a 1,022 ± 0,083 1,689 ± 0,554
NO3- 1,225 ± 0,170 a 1,082 ± 0,071 1,381 ± 0,349
DON 1,180 ± 0,138 a 1,114 ± 0,172 1,267 ± 0,236
TN 1,159 ± 0,103 a 1,050 ± 0,127 1,268 ± 0,162

- 46 -
Psychotria elataR
b/K
0,00
0,01
0,02
0,03
0,04
Sr/
Ca
0,00
0,05
0,10
0,15
0,20
0,25
Siparuna thecaphora
Rb
/K
0,00
0,01
0,02
0,03
0,04
Sr/
Ca
0,00
0,05
0,10
0,15
0,20
0,25
P TF a TF b LEA L
15N
/1
4N
0,002
0,003
0,004
0,005
0,006
0,007
0,008
P TF a TF b LEA L
15N
/1
4N
0,002
0,003
0,004
0,005
0,006
0,007
0,008
aa
a
b
c
a aa
bb
a a aa
b
abc
a
ab
bc c
* a aa
b
* aab
a
b
A
B
C
D
E
F
Fig. 2 Foliar concentration ratios of tracers and corresponding elements: Rb+:K+(A, D), Sr2+:Ca2+
(B, E) and 15N:14N (C, F) of different rainwater fractions, leachates and leaves of P. elata and
S. thecaphora; stem injection experiment, tropical lowland rainforest, Piedras Blancas National Park,
Costa Rica. Values are means with SE (n=12). Different letters indicate significant differences between
fractions. Significances were tested by one-way ANOVA (LSD-test, P < 0.05). TFa: Throughfall above
the canopy (n = 12), TFb: Throughfall below the canopy, LEA: Leachates of controlled leaching
experiment, L: Leaves. * P: Precipitation (n = 1) was excluded from statistical analysis.

- 47 -
Net
flu
x (
µm
ol.
g-1
DW
.3d
-1)
-10
-8
-6
-4
-2
0
2
4
6
8
10
Net
flu
x D
OC
(µ
mo
l. g
-1 D
W. 3
d-1
)
-160
-120
-80
-40
0
40
80
120
160
NO3- NH4
+ DON Cl- SO42- PO4
3- Na+ K+ Mg2+ Ca2+ DOC
a
b
bc c d
bcbcbcbc c
*
Fig. 3 Sum of net fluxes of N fractions (DON: dissolved organic nitrogen), anions, cations and DOC
(dissolved organic carbon) of branches from A. cf. costaricensis and F. tonduzzi treated with
simulated rain events over a period of 3 days, tropical lowland rainforest, Piedras Blancas National
Park, Costa Rica. Values show means with SE (n = 5). Different letters indicate significant
differences between fractions. Significances were tested by one-way ANOVA (LSD-test, P < 0.05).
* DOC fraction was excluded from the statistical analysis shown.

- 48 -
Ammonium
-0,4
-0,2
0,0
0,2
0,4
Nitrate
Flu
xes
(µm
ol
15N
. g
-1 D
W e
ven
t-1)
-0,06
-0,04
-0,02
0,00
0,02
0,04
0,06
DON
EFFLUX INFLUX NET UPTAKE-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
a
c
b
a
bab
a
b
b
Fig 4 Gross canopy N fluxes (efflux and influx) and net N
uptake of ammonium, nitrate and DON (dissolved
organic nitrogen) of the canopy components of branches
of A. cf. costaricensis and F. tonduzzi, determined by 15N-pool dilution experiment; tropical lowland rainforest,
Piedras Blancas National Park, Costa Rica. Values show
means with SE (n = 5). Different letters indicate
significant differences between fractions. Significances
were tested by one-way ANOVA (LSD-test, P < 0.05).

- 49 -
Table 2 Statistically differences of N fractions ammonium, nitrate
and DON (dissolved organic nitrogen) in gross canopy fluxes
(efflux and influx) and net N uptake of the canopy components of
branches of A. cf. costaricensis and F. tonduzzi (n = 5),
determined by 15N-pool dilution experiment; tropical lowland
rainforest, Piedras Blancas National Park, Costa Rica. Different
letters indicate significant differences between fractions.
Significances were tested by one-way ANOVA (LSD-test,
P < 0.05).
N species EFFLUX INFLUX NET UPTAKE
NH4+ a n.s. a
NO3- b n.s. ab
DON ab n.s. b

- 50 -
Fig. 5 Dry weights (A), allocation
of 15N in percent of initial 15N
amount applied in rain water (B),
and uptake rate of 15N (C) of
different canopy components from
branches of A. cf. costaricensis
and F. tonduzzi, simulated rain
application experiment, tropical
lowland rainforest, Piedras Blancas
National Park, Costa Rica Values
show means with SE (n = 5).
Different letters indicate significant
differences between fractions.
Significances were tested by one-
way ANOVA (LSD-test, P < 0.05).
Bk bark
CL canopy litter
CS canopy soil
Eph epiphylls
Ev vascular epiphytes
L leaves
Lich lichens
M mosses (bryophytes)
Allo
cati
on
(%
15N
of
RW
)
0
5
10
15
20
25
DW
(g
)
0
5
10
15
20
25
30
A
B
Bk CL CS Eph Ev L Lich M
Up
take r
ate
(µ
mo
l 1
5N
. g
-1 D
W.
even
t-1)
0,00
0,01
0,02
0,03
0,04
0,05
C
a a a a
a
a
b
b
aa a
a
a
a
b
a
ab
a
ab ab
a
ab
b
ab
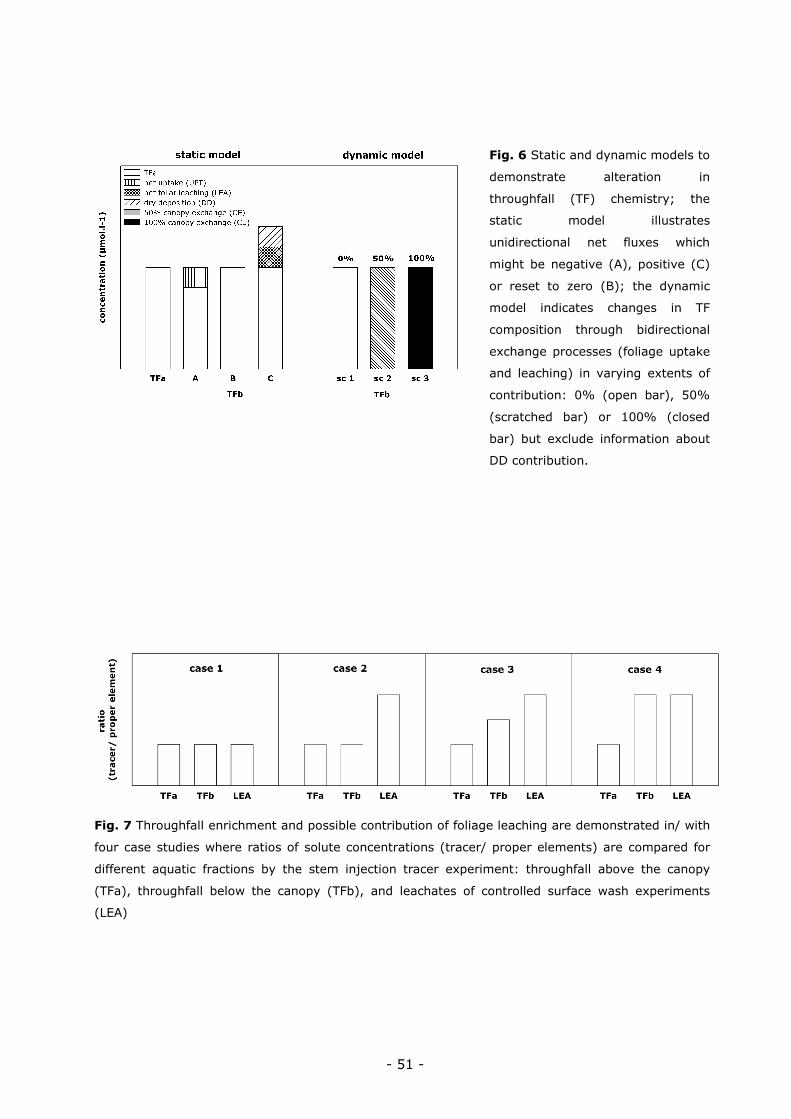
- 51 -
Fig. 6 Static and dynamic models to
demonstrate alteration in
throughfall (TF) chemistry; the
static model illustrates
unidirectional net fluxes which
might be negative (A), positive (C)
or reset to zero (B); the dynamic
model indicates changes in TF
composition through bidirectional
exchange processes (foliage uptake
and leaching) in varying extents of
contribution: 0% (open bar), 50%
(scratched bar) or 100% (closed
bar) but exclude information about
DD contribution.
Fig. 7 Throughfall enrichment and possible contribution of foliage leaching are demonstrated in/ with
four case studies where ratios of solute concentrations (tracer/ proper elements) are compared for
different aquatic fractions by the stem injection tracer experiment: throughfall above the canopy
(TFa), throughfall below the canopy (TFb), and leachates of controlled surface wash experiments
(LEA)

- 52 -
3. Zusammenfassung
Die chemische Zusammensetzung von Niederschlag ändert sich während der Durchdringung
des Kronenraumes von tropischen Regenwäldern. Generell kommt es zu einer
Nährstoffanreicherung bzw. Anreicherung an gelösten anorganischen und organischen
Substanzen im daraus resultierenden Kronentrauf. Die dabei entscheidenden regulierenden
Prozesse sind Trockendeposition und Austauschprozesse im Kronenraum. Trockendeposition
stellt, neben dem Niederschlag, einen externen atmosphärischen Nährstoffeintrag an
partikulärem und gasförmigen Substanzen, sowie Aerosolen in Waldökosysteme dar. Diese
werden an Oberflächen im Kronenraum deponiert und mit dem einfallenden Regenwasser
abgewaschen. Austauschprozesse im Kronenraum stellen einen internen Rezyklierungsweg
an Nährstoffen dar, die dadurch, neben Streufall und -Abbau, wieder pflanzenverfügbar
werden. Die Nährstoffdynamik des Kronentraufes ist insofern für tropische Waldökosysteme
von großer Bedeutung, da sie durch kontinuierlich hohe Niederschlagsmengen
gekennzeichnet sind und aufgrund geringer Nährstoffverfügbarkeit ihren Bedarf vorwiegend
durch Recycling decken. Austauschprozesse im Kronenraum inkludieren einerseits die
Aufnahme gelöster Substanzen aus dem Regenwasser und darin gelösten Verbindungen aus
Trockendeposition über Oberflächen pflanzlichen Gewebes (Cuticula, Borke) und
andererseits Auswaschung gelöster Nährstoffe aus labilen wässrigen Pools (Apoplast) des
Pflanzengewebes. Frühere Studien mit unterschiedlichen Versuchsansätzen zeigten die
Problematik, diese Prozesse zu trennen und zu quantifizieren, was auf ein komplexes und
dynamisches Interagieren von Trockendeposition und Kronenraum-Austauschprozessen
schließen lässt. Die Applikation von (konservativen) Tracern in situ, wie z.B. stabilen
Isotopen, stellt eine Möglichkeit dar, tiefere Erkenntnisse über Nährstoffflüsse im
Kronenraum zu erlangen.
Zielsetzung der vorliegenden Arbeit war es, Trockendeposition und Aufnahme und
Auswaschung von gelösten Nährstoffen im Kronenraum eines tropischen
Tieflandregenwaldes anhand verschiedener in situ Tracer-Experimenten zu unterscheiden.
Zum einen wurde der jeweilige relative Beitrag von Trockendeposition und Kronenaustausch
zu Kronentrauf-Nettoflüssen von Kalium, Calcium und Stickstoff via Stamminjektion der
entsprechenden Tracer (Rb+ für K+, Sr2+ für Ca2+ und 15N für 14N) in Bäume im Unterwuchs,
kontrollierten Auswaschungsversuchen und anschließender Auswertung über zwei
Modellansätze ermittelt. Rubidium und Strontium eignen sich gut als Tracer, da sie
chemisch und biologisch ähnliche Eigenschaften wie Kalium und Calcium besitzen. In einem
weiteren Experiment wurde erstmals die so genannte „15N Pool dilution“-Methode, welche
bisweilen zur Bestimmung von Brutto N-Mineralisierungsraten in Böden angewandt wurde,
adaptiert um Brutto N-Flüsse innerhalb des Kronenraumes zu untersuchen. Kronenraum-
Austauschprozesse, also Aufnahme (Influx) und Auswaschung (Efflux) von Nährstoffen im
Kronenraum, gelten als sehr dynamische Prozesse sind aber schwer zu quantifizieren,

- 53 -
besonders im Falle von N, der in anorganischer Form eher aufgenommen wird und in
organischer Form von Kronenraumkomponenten eher ausgewaschen wird. Während im
ersten Versuch durch Modellansätze der Netto-Austausch von N im Kronenraum und dessen
Anteil in Kronentrauf-Flüssen ermittelt wurde, diente der letztere Versuch dazu, die
tatsächlichen Aufnahme-Flüsse (Influx) und Abgabe-Flüsse (Efflux) von Stickstoff im
Kronenraum zu untersuchen. Hierzu wurde Regenwasser mit 15N (in Form von NH4+, NO3
+
und DON) angereichert und auf den Kronenraum repräsentierende Äste mit starkem
Bewuchs an Epiphyten, wie Bryophyten, Flechten und vaskulären Epiphyten, sowie
Epiphyllen appliziert, um gleichzeitig festzustellen, welche Kronenraumkomponente am
stärksten solche Austauschprozesse beeinflusst.
Die Applikation von Tracer via Stamminjektion und die Auswaschungsversuche verliefen für
Kalium und Calcium erfolgreich. Kalium- und Calcium-Flüsse weisen oft eine Netto-
Anreicherung im Kronentrauf auf, unterscheiden sich jedoch hinsichtlich ihrer Herkunft.
Obwohl die Anreicherung beider Kationen im Kronentrauf sowohl aus
Kronenaustauschprozessen als auch Trockendeposition resultieren können, weist Kalium
einen höheren Eintrag durch Auswaschungsprozesse im Kronenraum auf und Calcium wird
zu einem größeren Anteil als partikuläres Material von außen eingetragen. Kalium kommt in
pflanzlichem Gewebe in hohen Konzentrationen in wässrigen Lösungen vor und ist sehr
mobil, während Calcium vor allem in permanenten Strukturelementen in der Pflanze
gebunden ist. Mit den Tracer-Experimenten konnte nachgewiesen werden, dass Kalium wie
auch Calcium im Kronentrauf von Unterwuchspflanzen in einem tropischen
Tieflandregenwald zu einem beträchtlichen Teil aus Trockendeposition und Kronenraum-
Austauschprozessen stammten.
Die jeweiligen Beiträge von Kronenraum-Austauschprozessen und Trockendeposition im
Kronentrauf wurden einerseits anhand eines statischen Modells als unidirektionale Netto-
Zunahme oder –Abnahme demonstriert und andererseits mittels eines dynamischen
Modells, welches Konzentrationsänderungen im Kronentrauf anhand bidirektionaler
Austauschprozesse im Kronentrauf aufzeigt, ausgewertet. Die Brutto N-Flüsse (Influx and
Efflux) innerhalb der „Kronenraum“-Äste zeigten sehr hohe Influx- wie auch Efflux-Raten
von Ammonium, welche einen Netto NH4+-Fluss von nahezu Null ergaben. Brutto NO3
-- und
DON-Flüsse resultierten in einer Netto-Aufnahme, doch nichts desto trotz fanden auch
beträchtliche Effluxe statt. Der höchste Anteil an inkorporiertem 15N innerhalb der
verschiedenen Kronenraumkomponenten wurde in epiphythischen Bryophyten
nachgewiesen, während die höchsten 15N Aufnahmeraten die Epiphyllen-Fraktion, die fast
zur Gänze auch aus kryptogamen Pflanzen besteht, aufwies.
Aus den Ergebnissen dieser Arbeit lässt sich schlussfolgern, dass die Applikation von Tracern
in situ eine exzellente, wenn auch aufwendige und erweiterbare, Möglichkeit darstellt, um
hoch dynamische und komplexe Prozesse, wie Kronenraum-Austauschprozesse und

- 54 -
Trockendeposition, zu untersuchen. Weiters ermöglicht das dynamische Model die den
Kronentrauf kontrollierenden Prozesse besser abzubilden als das statische Modell, des
ausschließlich auf Nettoflüssen basiert. Die hohen Bruttoflüsse gelöster Substanzen im
Kronenraum zeigen, trotz geringer Nettoflüsse, dass im Kronenraum tropischer
Tieflandregenwälder sowohl Aufnahme als auch Abgabe von Nährstoffen zeitgleich
stattfinden können und zudem der Bewuchs im Kronenraum durch kryptogame Epiphyten
von großer Bedeutung in der Nährstoffdynamik tropischer Tieflandregenwälder sind.

- 55 -
4. Appendix:
Fig. 8 Concentrations of the base cations K+ and Ca2+, and nitrogen against concentrations of their
tracers Rb+, Sr2+ and 15N, respectively, and mean ratio values with SE (n = 12) in rainwater fractions
(TFa and TFb) of P. elata and S. thecaphora; stem injection experiment, tropical lowland rainforest,
Piedras Blancas National Park, Costa Rica.

- 56 -
Fig. 9 Correlations of concentrations of base cations, anions, DOC (dissolved organic carbon) and N
fractions (DON: dissolved organic nitrogen) between throughfall above the canopy (TFa) and
throughfall below the canopy (TFb) of P. elata (∆, n = 12) and S. thecaphora (o, n = 12) in logarithm-
diagrams; stem injection experiment, tropical lowland rainforest, Piedras Blancas National Park, Costa
Rica.
Related Documents











