Diversität von Wanzen in ökologischen Ausgleichsflächen und Weizenfeldern Masterarbeit zur Erlangung des akademischen Grades Diplom-Ingenieurin (Dipl.-Ing.) Masterstudium Agrarbiologie Eingereicht von Corinna Eggenreich, Bakk.rer.nat Universität für Bodenkultur Wien Betreuer: Univ. Prof. Mag. Dr. Thomas Frank Department für Integrative Biologie und Biodiversitätsforschung, Institut für Zoologie, Universität für Bodenkultur, Wien Wien, Juni 2012

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript
-
Diversität von Wanzen in ökologischen Ausgleichsflächen
und Weizenfeldern
Masterarbeit
zur Erlangung des akademischen Grades
Diplom-Ingenieurin (Dipl.-Ing.)
Masterstudium Agrarbiologie
Eingereicht von
Corinna Eggenreich, Bakk.rer.nat
Universität für Bodenkultur Wien
Betreuer:
Univ. Prof. Mag. Dr. Thomas Frank
Department für Integrative Biologie und Biodiversitätsforschung,
Institut für Zoologie, Universität für Bodenkultur, Wien
Wien, Juni 2012
-
I
Zusammenfassung
Eine Möglichkeit dem schnellen Verlust an biologischer Diversität in
Agrarökosystemen entgegenzuwirken, ist die Etablierung von Ackerbrachen, welche
seit den 1990 Jahren vermehrt in einigen europäischen Ländern angelegt wurden.
In dieser Masterarbeit wurden Artenreichtum und Abundanz von Wanzen
(Heteroptera) in drei verschiedenen Habitattypen in Wien Oberlaa untersucht,
nämlich in zwei naturnahen Habitaten (Ackerbrachen und Windschutzgürtel) und
Winterweizenfeldern.
Wanzen wurden ausgewählt, weil sie eine sehr gute Indikatorgruppe für die gesamte
Insektendiversität in Agrarökosystemen sind. Vorangegangene Arbeiten haben
gezeigt, dass der Reichtum der Wanzenfauna stark mit der gesamten
Insektendiversität korreliert. Es wurden Pflanzenartenzahl und Blütendeckung
aufgenommen um festzustellen, ob diese Faktoren einen Effekt auf die Wanzenfauna
ausüben.
Anhand einer einfaktoriellen ANOVA wurde festgestellt, dass die Gesamtzahl an
Individuen und Arten in den Ackerbrachen signifikant höher war als in den
Weizenfeldern und Windschutzgürteln. Auch die Individuen- und Artenzahl
phytophager Wanzenarten war in den Ackerbrachen signifikant höher als in den
beiden anderen Habitattypen. Die Hauptkoordinatenanalyse (PCO) zeigte, dass die
Ähnlichkeit der Wanzengemeinschaft und der Pflanzengemeinschaft in den
Ackerbrachen hoch war und sich deutlich von den Windschutzgürteln und
Weizenfeldern unterschied. Auch Windschutzgürtel und Weizenfelder wiesen
zueinander nur eine geringe Ähnlichkeit auf.
Arten- und Individuenzahl der Wanzen waren signifikant positiv mit der
Pflanzenartenzahl korreliert, was darauf schließen lässt, dass die Pflanzenartenzahl
ein Schlüsselfaktor für Wanzen ist.
Die Anlage von Ackerbrachen hatte einen positiven Effekt auf die Artenzahl von
Wanzen. Ferner wurden selten gewordene Arten durch die Anlage von Ackerbrachen
gefördert, weil Ackerbrachen diesen Arten neuen Lebensraum boten.
Schlagwörter: Ackerbrachen, Weizenfelder, Windschutzgürtel, Heteroptera,
Insektendiversität, Funktionelle Gruppen, Pflanzenartenzahl
-
II
Abstract
One opportunity to reduce the rapid decline in biological diversity in agricultural
landscapes is to establish fallows on arable land, which have been established since
the 1990s in several European countries. In this master thesis species richness and
abundance of heteropteran bugs in three different types of habitats were ascertained
in Vienna Oberlaa. The three types of habitats were two semi-natural habitats, i.e.
fallows and hedgerows, and winter wheat fields.
The true bugs (Heteroptera) were chosen as they are an appropriate indicator for
total insect diversity. This was based on previous work that had shown that the
richness of the bug fauna correlates strongly with total insect diversity. Plant species
richness and flower abundance were investigated to estimate their effects on the bug
fauna.
One-way ANOVA revealed that total species richness and individual numbers of bugs
were significantly higher in fallows than in winter wheat fields and hedgerows. Also
the numbers of phytophagous bugs were significantly higher in the fallows than in the
other two types of habitats. Principle Coordinate Analysis (PCO) showed that the bug
species composition and plant species composition in the fallows was very similar but
strongly separated from that in the hedgerows and the winter wheat fields. Bug and
plant assemblages between fallows and hedgerows were rather dissimilar.
The number of bug species and individuals were significantly positively associated
with the number of plant species, indicating that plant species richness is a key factor
for bugs.
Arable fallows had a favourable effect on the number of bug species. Moreover,
arable fallows favoured rare bug species as they found new habitat in the fallows.
Key words: Arable fallows, Winter wheat fields, Hedgerows, Heteroptera, Insect
diversity, Functional groups, Plant species
-
III
Inhaltverzeichnis
Zusammenfassung .................................................................................................... I
Abstract ..................................................................................................................... II
Abbildungsverzeichnis ........................................................................................... IV
Tabellenverzeichnis.................................................................................................. V
1 Einleitung ............................................................................................................... 1
2 Material und Methoden .......................................................................................... 4
2.1 Untersuchungsflächen ....................................................................................... 4
2.2 Fangmethode und Wanzenparameter ............................................................... 5
2.3 Vegetationsfaktoren .......................................................................................... 6
2.4 Datenanalyse .................................................................................................... 6
3 Ergebnisse ............................................................................................................. 9
3.1 Abundanz und Artenzahl von Wanzen in den Habitattypen............................... 9
3.2 Charakterisierung der Wanzengemeinschaft................................................... 14
4 Diskussion ........................................................................................................... 19
5 Literaturverzeichnis............................................................................................. 24
6 Danksagung ......................................................................................................... 28
Appendix ................................................................................................................. 29
-
IV
Abbildungsverzeichnis
Abb. 1: Die drei Untersuchungshabitate………………………………………………….5
Abb. 2: Verteilung der Wanzenindividuen und Wanzenarten ……………………….. 12
Abb. 3: Beziehung zwischen Pflanzenartenzahl und Wanzenindividuenzahl………13
Abb. 4: Beziehung zwischen Pflanzenartenzahl und Wanzenartenzahl…………….13
Abb. 5: Korrespondenzanalyse (CA)…………………………………………………….14
Abb. 6: Hauptkoordinatenanlyse (PCO) zur Darstellung der Ähnlichkeit der
Wanzengemeinschaft……………………………………………………………………..15
Abb. 7: Hauptkoordinatenanlyse (PCO) zur Darstellung der Ähnlichkeit der
Pflanzengemeinschaft…………………………………………………………………….16
-
V
Tabellenverzeichnis
Tab. 1: Artenliste…………………………………………………………………………11
Tab. 2: PERMANOVA: Paarvergleiche der Habitattypen für
Wanzengemeinschaft…………………………………………………………………...17
Tab. 3: PERMANOVA: Paarvergleiche der Habitattypen für
Pflanzengemeinschaft………………………………………………………………….17
-
1
1 Einleitung
Die moderne Landwirtschaft in Europa wird über intensive Landnutzung, Reduktion
und Fragmentierung von natürlichen und naturnahen Habitaten und den stetigen
Nutzungsanstieg von Chemikalien in der Landwirtschaft charakterisiert, dadurch
kommt es zu einem Verlust von Biodiversität auf landwirtschaftlichen Nutzflächen
(de Heer et al., 2005; Tscharntke et al., 2005).
Der rapide Verlust an der Vielfalt an Pflanzen- und Tierarten in der Landwirtschaft
kann durch zwei Gründe erklärt werden. Zum einem durch die steigende
Bewirtschaftung in der Landwirtschaft und zum anderem durch die Reduktion der
Habitatgröße durch Fragmentierung, was beides zu einem Verschwinden der Arten
führt (Tscharntke & Kruess, 1999; Steffan-Dewenter & Tscharntke, 2000). Eine hohe
Biodiversität kann nur erhalten bleiben, wenn in agrarisch genutzten Flächen eine
geringe Bewirtschaftungsintensität gewährleistet ist (Kőrösi et al., 2011).
Daher werden in europäischen Ländern seit den 1990 Jahren naturnahe Habitate,
wie etwa Ackerbrachen oder Windschutzgürtel, vermehrt etabliert (Ullrich & Edwards,
1999; Di Giulio et al. 2001; Collins et al., 2003; Haaland et al., 2011; Frank et al.,
2012). Trotzdem kürzte die europäische Agrarpolitik 2008 die Beihilfen für die
Stilllegung von Ackerflächen, welches zu einem weiteren Verlust von Habitaten und
Biodiversität in der europäischen Landwirtschaft führte (Tscharntke et al., 2011). In
Österreich gibt es Programme, welche heimische Landwirte finanziell unterstützen,
wenn diese ehemalige Ackerflächen stilllegen. In diesen Programmen werden
gemeinsam mit den Landwirten Pflegemaßnahmen für Ackerbrachen festgelegt.
Dazu gehört ein Programm der Stadt Wien (MA 22, Abteilung für Umweltschutz),
welches hinsichtlich seiner Auswirkungen auf Wanzen in vorliegender Arbeit
untersucht wird.
Die vermehrte Stilllegung von Ackerflächen hätte nicht nur einen ökonomischen
Aspekt, sondern würde vor allem einen wichtigen Beitrag für die Erhaltung natürlicher
Agrarökosysteme leisten (Corbet, 1995; Öckinger & Smith, 2007; Schmidt-Entling &
Döbeli, 2009; Aviron et al. 2011). Stillgelegte Ackerflächen können in der modernen
Landwirtschaft neue Lebensräume für nützliche Arthropoden schaffen. Diese
Habitate können als Überwinterungsorte dienen oder bieten Schutz während der
-
2
Bewirtschaftung von Ackerland. Ferner dienen Brachen als Nahrungsressource, weil
auf Ackerbrachen ein abwechselndes Beutetierspektrum zur Verfügung steht
(Zurbrügg & Frank, 2006; Schmidt-Entling & Döbeli, 2009).
In dieser Studie wurden drei verschiedene Habitattypen (Weizenfelder, Ackerbrachen
und Windschutzgürtel) untersucht. Bei den Weizenfeldern handelt es sich
ausschließlich um Winterweizenfelder (Triticum aestivium L.). Ackerbrachen und
Windschutzgürtel sind naturnahe Habitattypen, welche die Biodiversität in
Agrarregionen erhöhen. In einer stark agrarisch genutzten Landschaft sind
Ackerbrachen ein wichtiges Refugium für den Erhalt der Artenvielfalt (Marshall &
Moonen, 2002).
Die Anlage von Ackerbrachen wird in Österreich seit etwa 20 Jahren praktiziert, um
der negativen Entwicklung der Landschaft in den letzten Jahrzehnten
entgegenzuwirken. Ackerbrachen werden meist mit einer Wildblumenmischung in
einem Ackerfeld oder am Rand eines Ackers angesät. Diese müssen über sechs
Jahre aufrechterhalten bleiben, um eine sekundäre Sukzession zu ermöglichen
(Frank et al., 2009). Auch Windschutzgürtel haben einen wichtigen Stellenwert und
können die Biodiversität in Agrarlandschaften positiv beeinflussen (Marshall &
Moonen, 2002).
In dieser Studie wurden Abundanz, Artenreichtum, Diversität und Evenness von
Wanzen (Heteroptera) untersucht. Ebenfalls wurde der Einfluss der
Pflanzenartenzahl auf die Wanzendichte in den Ackerbrachen, Windschutzgürteln
und Weizenfeldern untersucht. In vorangegangen Arbeiten wurde beobachtet, dass
Abundanz und Artenreichtum von Wanzen in Ackerbrachen positiv mit der Zahl der
Pflanzenarten korrelierten (Ullrich, 2001; Frank & Künzle, 2006; Zurbrügg & Frank,
2006). Je höher die Pflanzenvielfalt am jeweiligen Standort, desto höher ist in der
Regel auch die Wanzenvielfalt.
Wanzen (Heteroptera) wurden aus mehreren Gründen als Indikatorgruppe
ausgewählt. Wanzen enthalten sowohl zoophage wie auch phytophage Arten
(Dolling, 1991). Einige Arten sind Generalisten, während andere zu den Spezialisten
gezählt werden. Nymphen und Adulte besiedeln dieselben Habitattyen und beide
reagieren sensibel auf Umweltveränderungen (Otto, 1996). Zudem haben
vorangegangene Studien gezeigt, dass die Vielfalt der Wanzenfauna stark mit der
gesamten Insektendiversität korreliert (Duelli & Obrist, 1998).
-
3
Die untersuchten Ackerbrachen sind Vertragsnaturschutzflächen, welche von der
Stadt Wien gefördert werden. Diese Studie soll zeigen, ob durch die Anlage von
Ackerbrachen Diversität, Abundanz und Artenreichtum von Wanzen gegenüber
Windschutzgürteln und Weizenfeldern erhöht werden können.
Es werden folgende Hypothesen formuliert:
(i) In Ackerbrachen sind Diversität, Abundanz und Artenreichtum
höher als in den Weizenfeldern und Windschutzgürteln, weil es in
Ackerbrachen eine höhere Vielfalt an Pflanzenarten gibt und somit
für phytophage Wanzen ein größeres Nahrungsangebot zur
Verfügung steht. Zudem wirken sich landwirtschaftliche Aktivitäten
in Weizenfeldern negativ auf die Wanzendiversität aus.
(ii) Abundanz und Artenreichtum der Wanzen korrelieren positiv mit
der Pflanzenartenzahl.
(iii) Die Artengemeinschaften von Wanzen und Pflanzen in den
Weizenfeldern sind ähnlich und unterscheiden sich deutlich von
Ackerbrachen und Windschutzgürteln. Auch Ackerbrachen und
Windschutzgürtel weisen zueinander nur eine geringe Ähnlichkeit
auf.
-
4
2 Material und Methoden
2.1 Untersuchungsflächen
In dieser Arbeit wurden drei verschiedene Habitattypen (Brachen, Windschutzgürtel
und Winterweizen; Abb. 1) untersucht. Das Untersuchungsgebiet liegt in Oberlaa, im
Süden Wiens.
Die Untersuchungsflächen sind geprägt durch intensive Landwirtschaft mit
vorwiegend konventioneller Wirtschaftsweise. Bei den untersuchten Ackerbrachen
handelte es sich um Vertragsnaturschutzflächen. Im Rahmen der Umweltschutz-
Abteilung der Stadt Wien (MA 22) werden Felder oder Feldstücke von Wiener
LandwirtInnen aus der Produktion genommen und gegen eine vertragliche
Entschädigung als Naturschutzflächen gepflegt. Auf den Naturschutzbrachen wurde
eine „Voitsauer Wildblumenmischung“ angesät. Hier finden seltene und gefährdete
Arten der Wiener Kulturlandschaft einen neuen Lebensraum (Bioforschung Austria,
2012). Windschutzgürtel wurden untersucht, weil sie neben den Ackerbrachen der
einzige weitere naturnahe Habitattyp im Untersuchungsgebiet waren.
Es wurden je vier Ackerbrachen und Windschutzgürtel sowie zwölf
Winterweizenfelder untersucht, weil Winterweizen die häufigste Ackerkultur im
Untersuchungsgebiet war. Je vier Winterweizenfelder grenzten an die untersuchten
Ackerbrachen und Windschutzgürtel, weitere vier Winterweizenfelder dienten als
Kontrollflächen, die etwa 300 m von der nächsten naturnahen Fläche entfernt waren.
Von den zwölf Weizenfeldern wurden neun konventionell und drei biologisch
bewirtschaftet.
Es wurden nur zehnjährige Ackerbrachen und über 15-jährige Windschutzgürtel
ausgewählt. Auch ein Abstand von 20 m in den Winterweizenfeldern musste
gewährleistet sein, damit kein Einfluss von den Feldrändern ausging.
Die Ackerbrachen wurden von den Pflanzenarten Arrhenatherum elatius, Dactylis
glomerata, Cirsium arvense und Centaurea scabiosa dominiert. In den
Winterweizenfleldern waren die dominanten Ackerbeikräuter Anthemis austriaca,
Veronica hederifolia, Papaver rhoeas und Viola arvensis. In den Windschutzgürteln
-
5
wurden der Unterwuchs vor allem von Alliaria petiolata, Stellaria media und Bromus
sterilis geprägt.
Die Größe der Untersuchungsflächen reichte von 3.302 m² bis 57.477 m².
Die Jahresdurchschnittstemperatur von 2003 bis 2012 liegt bei 10,95°C. Während
der Vegetationsperioden von April bis Oktober schwanken die Temperaturen
zwischen 9,98°C und 12,67°C, wobei das Maximum im Juli mit 21,52°C erreicht wird.
Der mittlere Jahresniederschlag beträgt 504,9l/m2 (Wetterstation-Oberlaa, 2012).
Abb. 1. Die drei Untersuchungshabitate: Linkes Bild zeigt einen Windschutzgürtel; Bild in
der Mitte eine 10-jährige Ackerbrache; Rechtes Bild zeigt ein Winterweizenfeld (©
Eggenreich; Lichtenegger)
2.2 Fangmethode und Wanzenparameter
Zwischen Ende Mai bis Mitte Juni 2011 wurden drei Fangtermine mit einem Abstand
von einer Woche durchgeführt. Die Ausdehnung des Untersuchungszeitpunkts
konnte nicht verlängert werden, weil die Weizenfelder einmal im Jahr (Ende Juni-
Anfang Juli) geerntet wurden. Wanzen wurden nur bei Lufttemperatur über 17°C,
Sonnenschein, geringem gleichmäßigem Wind und trockener Vegetation gefangen.
Die Wanzen wurden in einem Zeitraum von 10 Uhr vormittags bis 15:30 Uhr
nachmittags mittels einer standardisierten Keschermethode gefangen (Otto, 1996).
Ab zwölf Uhr mittags wurden zwei Stunden Pause gemacht, da die
Sonneneinstrahlung sehr stark war und die Wanzen sich vornehmlich in Bodennähe
aufhielten.
Der Kescher hatte einen Durchmesser von 35 cm und eine Maschenweite von 1 mm
und eine Stablänge von 1 m, damit der Abstand zur Vegetation gewährleistet war.
-
6
In jeder Fläche wurden 100 Kescherschläge durchgeführt, nach 50 Kescherschlägen
wurde das Netz geleert und die Wanzen, aber auch alle anderen gefangenen
Tiergruppen, sofort mit Essigsäureethylester (C4H8O2) abgetötet.
Für die Datenanalyse wurden die Abundanz der Wanzen (die Gesamtzahl der
Adulten und Larven pro Habitattyp) und der Artenreichtum der Wanzen (die
Gesamtheit der adulten Wanzenarten pro Habitattyp) verwendet.
Ferner wurde zwischen trophischen Ebenen unterschieden: zoophage (schließt
zoophtyophag ein) und phytophage Arten.
Die adulten Tiere wurden mittels entomologischer Bücher (Wagner 1952, 1966,
1967; Wachmann et.al. 2004, 2006, 2007, 2008) und mit der CD „CORISA“ (Strauß,
2010) bestimmt. Die Larven blieben unberücksichtigt.
2.3 Vegetationsfaktoren
Um den Einfluss der Vegetation auf die Wanzen abzuschätzen, wurden
Blütendeckung und Pflanzenartenzahl erhoben. Die Blütendeckung wurde zweimal
innerhalb der Wanzenfangperiode erhoben. Hierzu wurde ein Gitter mit einer Größe
von 20 x 20 cm mit 10 x 10 Feldern à 2 x 2 cm verwendet. Das Gitter wurde pro
Erhebungstermin 24 Mal zufällig in jede Untersuchungsfläche ausgeworfen. Danach
wurde die Anzahl der Felder ermittelt, in denen sich Blüten befanden. Die
Blütendeckung wurde mittels folgender Skala ermittelt: 0=0 Blüten, 1=1-20%, 2=21-
40%, 3=41-60%, 4=61-80%, 5=81-100% Blüten/40cm².
Mittels einer Vegetationsaufnahme wurde die Pflanzenartenzahl erhoben. In den
Weizenfeldern und Windschutzgürteln wurde auf 10 m2 ein Ausschnitt ausgezählt,
bei den Brachen waren die Aufnahmefläche 50 m².
2.4 Datenanalyse
Die statistische Auswertung erfolgte mittels SPSS Version 18 (einfaktorielle ANOVA,
t-Test unabhäniger Stichproben und Regression), CANOCO for Windows Version
4.5 (Detrended Correspondence Analysis (DCA) und Korrespondenzanalyse (CA))
und PRIMER 6 (Hauptkoordinatenanalyse (PCO), Permutational MANOVA
(PERMANOVA) und Similarity Percentages (SIMPER)).
-
7
Die Originaldaten zeigten eine schlechte Normalverteilung, daher wurden die
Originaldaten logarithmisch transformiert (log10-Transformation): X‘=log(X+1),
X=Originalwert und X‘=Wert nach der Transformation.
Prüfung auf Normalverteilung erfolgte mittels Kolmogorov-Smirnov-Tests und Q-Q-
Plots. Die transformierten Daten zeigten eine bessere Normalverteilung als die nicht
transformierten Daten.
Einfaktorielle ANOVA wurde verwendet, um die Individuen- und Artenzahl, Shannon-
Diversitätsindex, Evenness und die trophischen Gruppen zwischen den
verschiedenen Habitattypen zu vergleichen. Mit Hilfe eines Tukey-Tests wurden
multiple Vergleiche zwischen den Mittelwerten jener Gruppen angezeigt, die sich
signifikant voneinander unterschieden (Sachs, 1997). Als Ungleichverteilungsmaß
drückt die Evenness aus, wie häufig bzw. selten Individuen einer Art in einem
bestimmten Lebensraum vorkommen. Je höher die Evenness (erreicht Werte von 0-
1), desto ausgewogener sind die Individuenzahlen zwischen den Arten verteilt.
Um die drei biologisch bewirtschafteten Weizenfelder mit den neun konventionellen
Weizenfeldern zu vergleichen, wurde ein t-Test mit log10-tranfsformierten Daten
verwendet. Dabei wurde kein signifikanter Unterschied zwischen den biologisch und
konventionell bewirtschafteten Weizenfeldern festgestellt (Signifikanz für die Anzahl
der Individuen: p = 0.196; Signifikanz für die Anzahl der Arten: p = 0.399). Die
Beziehung zwischen der Blütendeckung und Arten- und Individuenzahl, Shannon-
Index und Evenness wurde anhand einer Regression festgestellt. Bei einer weiteren
Regression wurde die Beziehung zwischen der Pflanzenartenzahl und Arten- und
Individuenzahl bzw. Shannon-Index und Evenness untersucht. Die Regression mit
der Blütendeckung wurde allerdings verworfen, da die meisten Habitattypen
Nullwerte hatten und somit wenig repräsentativ waren.
Für die Charakterisierung der Artgemeinschaften wurde zuerst eine DCA
durchgeführt, um die lengths of gradients zu ermitteln. Mittels der DCA kann
festgestellt werden, ob eine CA oder eine Hauptkomponentenanalyse (PCA)
eingesetzt wird. Bei einem Gradientwert 4 eine CA
verwendet. Liegt ein Wert zwischen 2 und 4, können beide Analysemethoden
angewendet werden. Der Wert der lengths of gradients betrug 3.983, somit wurde
eine CA zur Charakterisierung der Artengemeinschaft verwendet (Ter Braak &
-
8
Šmilauer, 2002). Bei einer CA wird die gesamte Wanzengesellschaft über die
Individuenzahl pro Art und Untersuchungsfläche charakterisiert.
Für die Charakterisierung der Artengemeinschaft wurde nicht nur eine CA verwendet,
sondern auch eine Hauptkoordinationsanalyse (PCO) mittels PRIMER. PCO wurde
verwendet, weil die Wanzendaten viele Nullwerte aufwiesen. Eine PCO ist ein
Ordinationsverfahren. Mit diesem wird versucht, eine Vielzahl von Variablen eines
vieldimensionalen Koordinatensystems in einem neuen Koordinatensystem so
darzustellen, dass entlang der neuen synthetischen Koordinatenachsen, den
sogenannten Hauptkomponenten, soviel Variabilität wie möglich abgebildet wird.
Dabei bildet die erste Hauptkomponente den größten Anteil der Variabilität ab. Die
zwei oder drei Hauptkomponenten, welche den Großteil der Variabilität abbilden,
werden dann grafisch dargestellt (Leyer & Wesche, 2008). Die Originaldaten wurden
Wurzel transformiert, um eine hohe Abundanz abgewichten zu können. Bei den
Pflanzenartendaten wurde „Presence/Absence“ verwendet. Als
Ähnlichkeitskoeffizient wurde das Bray-Curtis-Ähnlichkeitsmaß verwendet, da dieses
Maß vor allem für Artgemeinschaften günstig ist. Dieses bildet Gradienten in der
Artenzusammensetzung oft hinreichend gut ab und ist daher von fundamentaler
Bedeutung für ökologische Fragestellungen (Leyer & Wesche, 2008).
Mit PERMANOVA, einem der Varianzanalyse (ANOVA) analogen Verfahren, kann
getestet werden, ob Faktoren (hier: Habitat) einen signifikanten Einfluss haben.
Mittels der PERMANOVA wurde untersucht, ob sich die fünf Habitattypen bezüglich
ihrer Artengemeinschaft (Wanzenarten und Pflanzenarten) unterscheiden. Es wurde
der Monte-Carlo-Test verwendet. Der Monte-Carlo-Test wird eingesetzt, wenn es zu
wenige Wiederholungen gibt. Zuerst wurde ein „Main-Test“ durchgeführt um
festzustellen, ob sich die Daten signifikant voneinander unterscheiden. Gibt es
signifikante Unterschiede wird als zweiter Schritt ein „Pairwise Test“ durchgeführt.
SIMPER berechnet die mittlere Ähnlichkeit einer Gruppe. Dabei werden jene
Wanzenarten aufgezeigt, die an der Ähnlichkeit innerhalb der Habitattypen beteiligt
sind. Ferner wird auch die mittlere Unähnlichkeit zwischen zwei Gruppen berechnet.
-
9
3 Ergebnisse
3.1 Abundanz und Artenzahl von Wanzen in den Habitattypen
Insgesamt wurden 2017 Individuen gefangen, davon waren 783 Adulte und 1234
Nymphen aus 31 Wanzenarten. In den Ackerbrachen waren 53 Arten und 1565
Individuen (543 Adulte, 1022 Nymphen) und in den Weizenfeldern waren 64 Arten
und 434 Individuen (237 Adulte, 197 Nymphen), wobei die meisten Individuen in den
Weizenfeldern angrenzend an die Windschutzgürtel gefangen wurden. In den
Windschutzgürteln wurden 3 Arten und 18 Individuen nachgewiesen (3 Adulte, 15
Nymphen).
Die dominierende Familie in allen Habitattypen war jene der Miridae (Tabelle 1). Bei
den funktionellen Gruppen lag eindeutig der Großteil bei den phytophagen Arten,
sechs Arten (Deraeocoris ruber, Deraeocoris morio, Brachycoleus decolor, Nabis
ferus, Nabis punctatus und Campylomma verbasci) zeigten eine zoophage
Ernährungsweise auf.
Die Anzahl der Wanzenarten und Individuen war in den Ackerbrachen signifikant
höher als in den anderen Untersuchungsflächen (Abb. 2a-b). In den
Windschutzgürteln war die Anzahl der Individuen und Arten signifikant niedriger als in
den anderen Habitattypen. Die Anzahl der Individuen in den Weizenkontrollflächen
war signifikant niedriger als in den beiden anderen Weizenfeldern (Abb. 2a). Es
zeigten sich für phytophage Individuen und die Gesamtzahl der Individuen (Abb. 1a,
c), sowie für phytophage Arten und die Gesamtzahl der Arten (Abb. 2b, d) genau
dieselbe Siginfikanzen. Die Anzahl der zoophagen Individuen war signifikant höher in
den Ackerbrachen als in den Weizenkontrollflächen und den Windschutzgürteln. Die
Weizenfelder angrenzend an die Windschutzgürtel, sowie die Weizenfelder
angrenzend an die Ackerbrachen unterschieden sich von keinem anderen Habitattyp
signifikant (Abb. 2e). In den Ackerbrachen und den Weizenfeldern angrenzend an die
Ackerbrachen gab es signifikant mehr zoophage Arten als in den Windschutzgürteln
(Abb. 2f). Es konnten keine signifikanten Unterschiede zwischen den Habitattypen
beim Shannon-Index festgestellt werden (F4,15 = 1.224, p = 0.343). Die Evenness der
-
10
Ackerbrachen war signifikant höher als jene in den Weizenkontrollflächen (F4,15 =
6.110, p = 0.009).
Bei einer einfachen linearen Regression (y = a + bx) wurde festgestellt, dass mit
steigender Pflanzenartenzahl die Zahl der Wanzenindividuen stieg (y = 4.839 +
4.236 x; R² = 0.853; p < 0.001; Abb. 3). Ebenfalls wurde eine signifikante positive
Beziehung zwischen Pflanzenartenzahl und Wanzenartenzahl festgestellt (y = 3.528
+ 0.305 x; R² = 0.607; p < 0.001; Abb. 4). Dieselbe Beziehung wurde für
perennierende Pflanzenarten und Wanzenarten festgestellt (y = 14.022 + 4.370 x; R²
= 0.860; p < 0.001), allerdings wurde für annuelle Pflanzenarten und Wanzenarten
kein signifikanter Bezug festgestellt (y = 16.998 + 8.687 x; R² = 0.097; p = 0.181).
-
11
Tabelle 1. Die 31 nachgewiesenen Arten und ihre Gesamtzahl an Individuen in den Brachen (B),
Weizenfeldern angrenzend an Brachen (WB), Weizenkontrollflächen (WF), Weizenfeldern
angrenzend an Windschutzgürteln (WWG) und Windschutzgürteln (WG).
Art Familie B WB WK WWG WG Gesamt
Adelphocoris lineolatus Goeze Miridae 10 4 1 1 1 17 Apolygus lucorum Meyer-Dür Miridae 6 0 0 1 0 7 Brachycoleus decolor Reuter Miridae 28 0 0 0 0 28 Campylomma verbacsi Meyer-Dür Miridae 0 1 0 0 0 1 Capsodes gothicus Linnaeus Miridae 0 0 0 1 0 1 Capsus ater Linnaeus Miridae 1 0 0 1 0 2 Closterotomus norwegicus Gmelin Miridae 29 35 2 30 0 96 Criocoris crassicornis Hahn Miridae 6 0 0 0 0 6 Deraeocoris morio Boheman Miridae 1 0 0 0 0 1 Deraeocoris ruber Linnaeus Miridae 0 0 0 2 0 2 Dolycoris baccarum Linnaeus Pentatomidae 1 0 0 0 0 1 Eurygaster maura Linnaeus Scutelleridae 3 8 4 0 0 15 Graphosmoa lineatum Linnaeus Pentatomidae 2 0 0 0 0 2 Leptopterna dolobrata Linnaeus Miridae 42 2 0 1 0 45 Lygus rugulipennis Popp. Miridae 0 12 9 16 0 37 Megaloceraea reticornis Geoffroy Miridae 122 0 0 1 1 124 Nabis ferus Linnaeus Nabidae 0 1 0 0 0 1 Nabis punctatus A. Costa Nabidae 0 4 2 2 0 8 Notostira elongata Geoffroy Miridae 5 3 1 1 0 10 Oncotylus punctipes Reuter Miridae 43 0 0 0 0 43 Oncotylus viridiflavus Goeze Miridae 18 0 0 0 0 18 Orthocephalus coriaceus Fabricius Miridae 1 0 0 0 0 1 Orthocephalus vittipennisHerrich-Schäffer Miridae 1 0 0 0 0 1 Plagiognathus chrysanthemi Wolff Miridae 132 18 6 44 1 201 Platyplax salviae Schilling Lygaeidae 10 0 0 0 0 10 Polymerus holoricerus Hahn Miridae 1 0 0 0 0 1 Polymerus unifasciatus Fabricius Miridae 36 0 0 0 0 36 Stenotus binotatus Fabricius Miridae 44 0 0 0 0 44 Trigonotylus caelestialium Kirkaldy Miridae 0 3 9 10 0 22 Trigonotylus ruficornis Geffroy Miridae 0 0 0 1 0 1 Tritomegas sexmaculatus Linnaeus Cydnidae 1 0 0 0 0 1
Individuen 543 91 34 112 3 783 Arten 23 11 8 14 3 31
-
12
(a) Gesamtzahl Log_Individuen (b) Gesamtzahl Log_Arten
(c) Log Zahl phytophager (d) Log Zahl phytophager Arten Individuen
(e) Log Zahl zoophager Indivduen (f) Log Zahl zoophager Arten
Abb. 2. Verteilung der Wanzenindividuen und Wanzenarten (Mittelwert ± Standartabweichung) in den Brachen (B),
Weizenfeldern angrenzend an Brachen (WB), Weizenkontrollflächen (WK), Weizenfeldern angrenzend an Windschutzgürtel
(WWG) und Windschutzgürtel (WG). Unterschiedliche Buchstaben über den Balken zeigen signifikante Unterschiede
zwischen den verschiedenen Habitattypen (Tukey test p < 0.05). Alle Daten wurden logarithmisch transformiert. (a)
Gesamtzahl Individuen: ANOVA, F4,15 = 56.767, p < 0.001. (b) Gesamtzahl Arten: F4,15 = 27.311, p < 0.001. (c) Zahl
phytophager Individuen: F4,15 = 55.490, p < 0.001. (d) Zahl phytophager Arten: F4,15 = 20.936, p < 0.001. (e) Zahl zoophager
Individuen: F4,15 = 5.396, p = 0.007. (f) Zahl zoophager Arten: F4,15 = 5.401, p = 0.008.
-
13
Abb. 3. Beziehung zwischen Pflanzenartenzahl und Wanzenindividuenzahl ( y = 4.839 +
4.236x; R² = 0.853; p < 0.001). Bedeutung der Symbole: ○ (Ackerbrachen), (Weizenfelder
angrenzend an Ackerbrachen), ◊ (Windschutzgürtel), ∆ (Weizenfelder angrenzend an
Windschutzgürteln) und □ (Weizenkontrollfelder).
Abb. 4. Beziehung zwischen Pflanzenartenzahl und Wanzenarten ( y = 3.2528 + 0.305x; R² =
0.607; p < 0.001). Bedeutung der Symbole: ○ (Ackerbrachen), (Weizenfelder angrenzend an
Ackerbrachen), ◊ (Windschutzgürtel), ∆ ( Weizenfelder angrenzend an Windschutzgürteln) und
□ (Weizenkontrollfelder).
Wa
nze
nin
div
idu
en
Pflanzenartenzahl
Pflanzenartenzahl
Wa
nze
nart
en
-
14
3.2 Charakterisierung der Wanzengemeinschaft
Eine Korrespondenzanalyse (CA) zeigt wie ähnlich die Artengemeinschaften in den
verschiedenen Habitattypen sind. Die beiden ersten Achsen erklärten 44,2% der
Varianz.
Die Weizenfelder zeigten eine hohe Ähnlichkeit der Artengemeinschaft und waren
getrennt von Ackerbrachen und Windschutzgürteln, die ihrerseits wiederum eine
hohe Ähnlichkeit aufwiesen. Die drei Weizenfelder waren einander sehr ähnlich, aber
sie grenzten nicht an die Achse 2 an, wie die Ackerbrachen und Windschutzgürtel
(Abb. 5).
Zusätzlich zu der CA wurde mit dem Programm PRIMER eine PCO durchgeführt.
Grund dafür war, dass die Wanzendaten zu viele Nullwerte en Zusätzlich zu der CA
-1.0 2.5
-1.0
2.0
WG2
WG3
WG4
WWG1
WWG2
WWG3
WWG4
B1
B2
B3
B4
WB1
WB2
WB3
WB4
WK1
WK2 WK3
WK4
Achse1
Ach
se
2
Abb. 5. Korrespondenzanalyse (CA) zur Darstellung der Ähnlichkeit der Wanzengemeinschaften zwischen Brachen
(B), Windschutzgürteln (WG), Weizenfeldern angrenzend an Brachen (WB), Weizenfeldern angrenzend an
Windschutzgürtel (WWG) und Weizenkontrollfeldern (WK). Um die Ähnlichkeiten zu verdeutlichen, wurden die vier
Wiederholungen von jedem Habitat miteinander verbunden. Beachte: In einem Windschutzgürtel wurden keine
Individuen gefangen, daher hier nur drei Punkte.
-
15
Zusätzlich zu der CA wurde mit dem Programm PRIMER eine PCO durchgeführt.
Grund dafür war, dass die Wanzendaten zu viele Nullwerte enthielten und die CA
somit kein repräsentatives Ergebnis hervorbrachte. Bei der Hauptkoordinatenanalyse
(PCO) erklärten die beiden ersten Achsen 45,6 % der Varianz. Deutlich wurde, dass
Windschutzgürtel und Ackerbrachen keine hohe Ähnlichkeit aufwiesen, im
Gegensatz zu dem Ergebnis der CA, wo Windschutzgürtel und Ackerbrachen sehr
hohe Ähnlichkeit aufwiesen. Die Ackerbrachen sind zueinander ähnlicher in ihrer
Artengemeinschaft als die Windschutzgürtel, da die Punkte der vier Ackerbrachen
enger zueinander lagen. Wie erwartet waren die drei Typen an Weizenfeldern auch
in der PCO sehr ähnlich zueinander, weshalb es keinen Unterschied zu der CA gab
(Abb. 6). Auch für die Pflanzenartenzahl wurde eine PCO durchgeführt. 66,8 % der
Varianz wurde von den beiden ersten Achsen erklärt. Bei der PCO für die
Pflanzenarten wurde noch deutlicher wie ähnlich die Ackerbrachen zueinander sind,
aber auch die Windschutzgürtel und Weizenfelder bildeten deutlich separate
Gruppen (Abb. 7).
Abb. 6. Hauptkoordinatenanalyse (PCO) zur Darstellung der Ähnlichkeit der Wanzengemeinschaft zwischen
Ackerbrachen, Weizenfeldern angrenzend an Ackerbrachen, Windschutzgürteln, Weizenfeldern angrenzend
an Windschutzgürtel und Weizenkontrollflächen. Alle vier Ackerbrachen liegen eng beieinander, daher
kann man auf eine hohe Ähnlichkeit der Artengemeinschaft schließen. Auch die Weizenfelder enthalten
ähnliche Artgemeinschaften und die Windschutzgürtel bilden ebenso eine separate Gruppe, die allerdings
innerhalb der Windschutzgürtel nur eine sehr geringe Ähnlichkeit aufweist. Beachte: In einem
Windschutzgürtel wurden keine Individuen gefangen, daher nur drei Sterne. Beachte: Hecke steht für
Windschutzgürtel und Hecke-Weizen steht für Weizenfelder angrenzend an Windschutzgürtel.
-
16
Die PERMANOVA zeigte, dass es einen signifikanten Unterschied in der
Wanzengemeinschaft zwischen den Habitaten gab (p < 0.001). Der anschließende
„Pairwise Test“ zeigte, dass sich die Ackerbrachen von den Weizenfeldern und
Windschutzgürteln signifikant unterschieden. Allerdings gibt es Unterschiede
zwischen permutierenden p-Werten (P(perm)) und Monte-Carlo-p-Werten (P(MC)).
Bei den permutierenden p-Werten wurde festgestellt, dass sich die Windschutzgürtel
signifikant von den Ackerbrachen und Weizenfeldern unterschieden. Der Monte-
Carlo-Test zeigte hingegen, dass sich die Windschutzgürtel nur von den
Ackerbrachen signifikant unterschieden, aber nicht signifikant von den Weizenfeldern
(Tabelle 2). Die Wanzengemeinschaft war in den Weizenfeldern ähnlich und bei
beiden p-Werten wurde innerhalb der Weizenfelder kein signifikanter Unterschied
festgestellt.
Abb. 7. Hauptkoordinatenanalyse (PCO) zur Darstellung der Ähnlichkeit der Pflanzengemeinschaft zwischen
Ackerbrachen, Weizenfeldern angrenzend an Ackerbrachen, Windschutzgürteln, Weizenfeldern angrenzend
an Windschutzgürtel und Weizenkontrollflächen. Alle vier Ackerbrachen liegen eng beieinander, daher kann
man auf eine hohe Ähnlichkeit der Artengemeinschaft schließen. Auch die Weizenfelder enthalten ähnliche
Artgemeinschaften und die Windschutzgürtel bilden ebenso eine separate Gruppe. Beachte: In zwei
Windschutzgürteln war kein Unterwuchs vorhanden, daher nur zwei Sterne. Beachte: Hecke steht für
Windschutzgürtel und Hecke-Weizen steht für Weizenfelder angrenzend an Windschutzgürtel.
-
17
Gruppen t P(perm) perms P(MC)
Hecke,Hecke-Weizen 1.4893 0.0278 35 0.1039
Hecke,Brache 1.7244 0.0271 25 0.0443
Hecke,Brache-Weizen 1.5644 0.0263 25 0.0764
Hecke,Kontrollweizen 1.4435 0.0286 35 0.1397
Hecke-Weizen,Brache 2.4348 0.0296 35 0.0046
Hecke-Weizen,Brache-Weizen 1.3851 0.0581 35 0.1438
Hecke-Weizen,Kontrollweizen 1.372 0.1718 35 0.1683
Brache,Brache-Weizen 2.5482 0.0272 35 0.0069
Brache, Kontrollweizen 2.7822 0.0265 35 0.0034
Brache-Weizen,Kontrollweizen 1.4412 0.1442 35 0.1386
Die PERMANOVA zeigte auch, dass es einen signifikanten Unterschied in der
Pflanzengemeinschaft zwischen den Habitaten gab (p < 0.001). Der „Pairwise Test“
ergab, dass sich Windschutzgürtel und Ackerbrachen signifikant in ihrer
Pflanzengemeinschaft unterschieden. Der permutierende p-Wert zeigt, dass sich nur
die Ackerbrachen signifikant von den beiden anderen Habitattypen unterschieden.
Allerdings ergaben die p-Werte des Monte-Carlo-Tests, dass sich nicht nur die
Ackerbrachen signifikant von den Weizenfeldern und Windschutzgürteln
unterschieden, sondern auch die Windschutzgürtel einen signifikante Unterschied
hinsichtlich ihrer Pflanzengemeinschaft aufwiesen und sich somit von den beiden
anderen Habitattypen signifikant unterschieden (Tabelle 3).
Gruppen t P(perm) perms P(MC)
Hecke,Hecke-Weizen 2.9412 0.0691 11 0.0101
Hecke,Brache 2.7078 0.0702 15 0.0146
Hecke,Brache-Weizen 4.0612 0.069 8 0.0032
Hecke,Kontrollweizen 2.81 0.0694 15 0.0146
Hecke-Weizen,Brache 3.0417 0.0276 35 0.0018
Hecke-Weizen,Brache-Weizen 0.92075 0.6282 15 0.4629
Hecke-Weizen,Kontrollweizen 0.91056 0.5753 18 0.4779
Brache,Brache-Weizen 3.6647 0.0297 19 0.0009
Brache,Kontrollweizen 3.1837 0.0297 26 0.0016
Brache-Weizen,Kontrollweizen 1.313 0.2529 15 0.2217
Tabelle 3. PERMANOVA: Paarvergleiche der Pflanzengemeinschaften zwischen den
Habitattypen. MC steht für Monte-Carlo-Test.
Tabelle 2. PERMANOVA: Paarvergleiche der Wanzengemeinschaft zwischen den Habitattypen.
MC steht für Monte-Carlo-Test.
-
18
Bei SIMPER zeigten die vier Ackerbrachen zueinander eine mittlere Ähnlichkeit von
51.7 auf, die ersten vier Wanzenarten Megaloceraea reticornis, Plagiognathus
chrysanthemi, Leptopterna dolobrata und Polymerus unifasciatus erklärten 62.6% der
Ähnlichkeitsvarianz (Appendix 4c).
Der Gruppenvergleich Ackerbrache und Windschutzgürtel erbrachte eine mittlere
Unähnlichkeit von 95.9, davon erklärten Megaloceraea reticornis, Plagiognathus
chrysanthemi, Stenotus binotatus und Leptopterna dolobrata 48.6% dieser
Unähnlichkeitsvarianz (Appendix 4g).
-
19
4 Diskussion
Die Dominanz der Miridae in der Studie ist typisch für naturnahe Habitate in einer
landwirtschaftlich genutzten Landschaft (Frank & Künzle, 2006). Die Zahl der
Individuen und Arten, sowie phytophage Wanzen waren vor allem in den
Ackerbrachen signifikant höher als in den Windschutzgürteln und Weizenfeldern.
Somit wird die erste Hypothese bestätigt, dass in Ackerbrachen Abundanz und
Artenreichtum höher sind als in beiden anderen Habitattypen, weil es in
Ackerbrachen eine höhere Vielfalt an Pflanzenarten gibt und phytophagen Wanzen
ein größeres Nahrungsangebot zur Verfügung steht.
In Habitattypen wie Ackerbrachen, die eine hohe Vielfalt an Pflanzenarten haben,
finden phytophage sowie zoophage Wanzenarten ein breites Angebot an Nahrung.
Dementsprechend war in dieser Arbeit die Zahl phytophager Wanzenarten in den
Ackerbrachen signifikant höher als in den Weizenfeldern und Windschutzgürteln, weil
in den untersuchten Ackerbrachen eine hohe Vielfalt an Pflanzenarten auftrat. In den
Windschutzgürteln wurde kaum bzw. wenig Unterwuchs festgestellt, daher wurden
auch nur drei Wanzenarten gefangen.
In den Weizenfeldern wurden hauptsächlich ubiquitäre Arten wie Closterotomus
norwegicus, Lygus rugulipennis, Trigonotylus caelestialium und Eurygaster maura
gefunden. Closterotomus norwegicus war die dominante Art in den Weizenfeldern
und häufig in den Ackerbrachen. Sie hat eine Präferenz für verschiedene
Asteraceae, kann aber auf landwirtschaftlichen Kulturen wie Weizen als Schädling
auftreten. Dabei werden beim Weizen in erster Linie die Reproduktionsorgane
besaugt. Eiablage findet nicht am Weizen statt, sondern an den Asteraceae, die sich
im Weizenfeld befinden (Wachmann et al., 2004).
Eine weitere dominante Art, die nur in den Weizenfeldern vorkam, war Lygus
rugulipennis. Sie kann ebenfalls an Weizen vorkommen und diesen besaugen
(Wachmann et al., 2004). T. calestialium und E. maura sind Arten, die ebenfalls die
Weizenpflanze als Wirtspflanze nutzen (Wachmann et al., 2004). In den
Weizenfeldern wurden hauptsächlich Arten gefunden, die an Weizen saugen und die
vorkommenden Ackerbreikräuter zur Eiablage nutzen können. Die in intensiv
-
20
genutzten Weizenfeldern auftretenden Arten gelten als sehr mobil und wenig auf
bestimmte Pflanzenarten spezialisiert (Di Giulio et al., 2001).
Megaloceraea reticornis (132 Individuen) und Plagiognathus chrysanthemi (122
Individuen) waren die dominanten Arten in den Ackerbrachen. Beide Arten besiedeln
vor allem trockene bis mäßig feuchte Offenlandstandorte mit artenreicher
Krautvegetation. M. reticornis kommt vor allem in ungemähten Grasstandorten vor,
auf denen Gräser wie Arrhenatherum, Festuca und Brachypodium wachsen
(Wachmann et al., 2004). In den untersuchten Ackerbrachen wurden diese Gräser
festgestellt, vor allem A. elatius dominierte in den Ackerbrachen. Daher ist es auch
nicht ungewöhnlich, dass M. reticornis die dominante Art in den Ackerbrachen war.
Leptopterna dolobrata war zwar keine dominante Art in den Ackerbrachen, aber
dennoch war die Art fast nur in den Ackerbrachen vorzufinden. Dies ist wohl darauf
zurück zuführen, dass diese Art grasige Standorte mit viel A. elatius bevorzugt
(Wachmann et al., 2004), was für die Ackerbrachen zutraf.
Oncotylus punctipes, Brachycleus decolor, Polymerus unifasciatus und Stenotus
binotatus wurden ausschließlich in den Ackerbrachen gefangen. Diese Wanzenarten
benötigen ungestörte Habitate ohne landwirtschaftliche Bewirtschaftung. O.
punctipes hält sich bevorzugt im Bereich der Blütenstände ihrer Nahrungspflanze
Tanacetum vulgare auf (Wachmann et al., 2004). Diese Pflanzenart wuchs in drei der
vier Ackerbrachen, allerdings wurde O. punctipes nur in jenen zwei Ackerbrachen
gefunden, in welchen die Dichte von T. vulgare am höchsten war. P. unifasciatus lebt
hauptsächlich auf Galium-Arten. Diese Wanzenart wurde in allen vier Ackerbrachen
gefangen, weil auch die Wirtspflanze häufig in diesem Habitattyp gefunden wurde.
In den Ackerbrachen wurden drei seltene Arten gefangen, nämlich Oncotylus
viridiflavus, Triconotylus ruficornis und Deraecoris morio. Alle drei Arten stehen in der
Roten Liste gefährdeter Arten Niederösterreichs (Rabitsch, 2007). O. viridiflavus und
D. morio zählen zur Kategorie 5 „Gefährdungsgrad nicht genau bekannt“. Kategorie 5
umfasst Arten, die entweder „vom Aussterben bedroht“, „stark gefährdet“ oder
„gefährdet“ sind, über die jedoch nicht genügend Information vorhanden ist. T.
ruficornis zählt zur Kategorie 6 „Nicht genügend bekannt“. Kategorie 5 umfasst Arten,
die wegen zu geringen Kenntnisstandes nicht sicher den Kategorien 0-5 zugeordnet
werden können (Rabitsch, 2007). Ackerbrachen bieten einen wichtigen Lebensraum
für gefährdete Arten, wobei O. viridiflavus in einer der vier Ackerbrachen häufig
-
21
vorkam. Diese Art kommt bevorzugt auf Centaurea-Arten (z.B.: C. scabiosa, C. nigra)
vor. Centaurea-Arten wurden gleich häufig in allen vier Ackerbrachen gefunden.
Warum O. viridiflavus nur in einer der vier Ackerbrachen vorkam ist unklar. Weitere
seltene Arten waren D. morio und T. ruficornis, welche allerdings Einzelfänge waren.
In den Windschutzgürteln wurden nur drei Individuen der drei Arten M. reticornis, P.
chrysanthemi und Adelphocoris lineolatus gefangen. Es wurden nur so wenige
Individuen gefangen, weil in den Windschutzgürteln kaum Unterwuchs vorhanden
war. P. chrysanthemi, welche häufig in dem an den entsprechenden
Windschutzgürtel angrenzenden Weizenfeld gefangen wurde, ist vermutlich von dort
zufällig in den Windschutzgürtel eingewandert. P. chrysanthemi kommt vor allem an
sonnigen und vegetationsreichen Standorten vor. Daher die Vermutung, dass diese
Art zufällig vom angrenzenden Weizenfeld in den Windschutzgürtel einwanderte. Die
beiden anderen Wanzenarten traten in allen Weizenfeldern nur selten auf und waren
daher Zufallsfunde in den Windschutzgürteln.
Entsprechend der Literatur (Frank & Künzle, 2006; Frank et al., 2009) war auch in
dieser Arbeit die Zahl zoophager Wanzen relativ gering im Verhältnis zur Zahl
phytophager Wanzen. Die zoophagen Wanzen umfassten etwa 5% der Gesamtzahl
der Wanzen, den Großteil bildeten phytophage Wanzen mit 95%. Die phytophagen
Individuen und Arten in den fünf Habitattypen zeigten dasselbe Signifikanzmuster wie
die Gesamtzahl der gefangenen Individuen und Arten. Die meisten zoophagen und
phytophagen Individuen wurden in den Ackerbrachen gefangen. In den
Ackerbrachen finden zoophage sowie phytophage Wanzen ein breiteres
Nahrungsspektrum als in den Weizenfeldern und Windschutzgürteln. Habitattypen
mit einer hohen Dichte und Anzahl von Pflanzenarten werden schneller von
Beutetieren kolonisiert (White & Hassal, 1994), denn die Beutetierpopulationen wie
Thysanoptera, Aphidoidea und Psylloidea bevorzugen vor allem Habitattypen mit
einer hohen Pflanzenartenzahl (Fauvel, 1999), wie sie in den Ackerbrachen
vorhanden ist. Zoophage Wanzen, die sich hauptsächlich von Aphidoidea und
Psylloidea ernähren, wären gute natürliche Antagonisten von potentiellen
Schädlingen. Laut Denys & Tscharntke (2002) hängt der Reichtum zoophager Arten
nicht nur vom Alter einer Ackerbrache ab, sondern auch von deren Größe. Es wurde
bestätigt, dass das Räuber-Beute-Verhältnis in größeren Ackerbrachen deutlich
-
22
höher war als in kleineren Ackerbrachen. Die Ackerbrachen in dieser Arbeit waren
relativ klein und möglicherweise wurde das Vorkommen zoophager Wanzenarten
beeinflusst.
Die Wanzenindividuen und Wanzenarten wurden am besten erklärt durch die Anzahl
aller Pflanzenarten. Auch in vorangegangenen Arbeiten wurde bereits gezeigt, dass
in naturnahen Habitaten im Agrarland mit steigender Pflanzenartenzahl die
Wanzenartenzahl und -individuenzahl steigt (Brändle et al., 2001; Frank & Künzle,
2006; Zurbrügg & Frank, 2006). Somit wurde die zweite Hypothese bestätigt, wonach
Abundanz und Artenreichtum der Wanzen positiv mit der Pflanzenartenzahl
korrelieren. Zudem wurde in dieser Arbeit eine signifikante positive Beziehung
zwischen Anzahl an Wanzenarten und perennierenden Pflanzenarten festgestellt.
Dies kann damit erklärt werden, dass in vorliegender Arbeit 10-jährige Ackerbrachen
untersucht wurden, in denen vor allem perennierende Pflanzenarten wuchsen. Daher
wurde wenig überraschend für annuelle Pflanzenarten keine signifikante Beziehung
mit den Wanzenarten festgestellt, weil die meisten Pflanzenarten in den
Ackerbrachen perennierend waren. Mit einer höheren Vielfalt an Pflanzenarten
verbessern sich die Lebensbedingungen für Wanzen. Wanzen finden ein größeres
Nahrungsangebot sowie auch spezielle Pflanzenarten vor, die den Wanzen als
Wirtspflanze dienen. Laut Ullrich (2001) ernähren sich die meisten spezialisierten
Wanzenarten von perennierenden Wirtspflanzen. Dies trifft für vorliegende Arbeit zu.
Oncotylus punctipes und Polymerus unifasciatus wurden nur in den Ackerbrachen
gefangen, weil nur in diesem Habitattyp die jeweiligen Wirtspflanzen auftraten,
nämlich Tanacetum vulgare für O. punctipes und Galium-Arten (z.B.: G. verum, G.
album) für P. unifasciatus.
Durch die Hauptkoordinatenanalyse (PCO) wurde die dritte Hypothese bestätigt, weil
die Wanzen- und Pflanzengemeinschaft eine deutliche Trennung der Ackerbrachen
von den beiden anderen Habitattypen anzeigten, was auf die Eigenständigkeit der
Ackerbrachengemeinschaften hinweist. Die Eigenständigkeit lässt sich dadurch
erklären, dass in Ackerbrachen fast nur perennierende Pflanzenarten gefunden
wurden und die Pflanzenartenzahl in den Ackerbrachen wesentlich höher war als in
den Weizenfeldern und Windschutzgürten. Somit konnten sich Wanzenarten
-
23
ansiedeln, die bevorzugt perennierende Pflanzenarten als Wirtspflanzen benötigen,
sowie Habitate benötigen, in denen kaum Störung vorherrscht, was für Ackerbrachen
zutrifft.
Meine Resultate belegten, dass die Anlage von Ackerbrachen einen positiven Effekt
auf die Artenzahl der Wanzen hatte. Da sich Wanzen in Agrarökosystemen als sehr
gute Indikatoren für die Gesamtdiversität von Insekten erwiesen (Duelli & Obrist,
1998), könnten die Ackerbrachen die generelle Insektendiversität im
Untersuchungsgebiet erhöhen. Ackerbrachen wurden schon von Thomas und
Marshall (1999) als geeignet zur Förderung der Insektendiversität eingestuft. Ein
wichtiger Aspekt ist, dass selten gewordene Arten durch die Anlage von
Ackerbrachen in landwirtschaftlich intensiv genutzten Regionen erhalten werden
können, weil Ackerbrachen ihnen neuen Lebensraum bieten (Haaland et al., 2011).
Ferner sind Ackerbrachen leicht handhabbar, weil dieser Habitattyp relativ einfach zu
etablieren ist und die Sukzession jederzeit wieder neu beginnen kann, indem diese
gepflügt werden (Thomas & Marshall, 1999). Aufgrund der hohen Artenzahl und des
Nachweises von drei gefährdeten Wanzenarten in Ackerbrachen leisten die hier
untersuchten Vertragsnaturschutzflächen der Stadt Wien einen wichtigen Beitrag zur
Förderung der Biodiversität in Agrarökosystemen.
-
24
5 Literaturverzeichnis
AVIRON S., HERZOG I., KLAUS I., SCHÜPBACH B. & JEANNERET P. 2011.
Effects of wildflower strip quality, quantity and connectivity on butterfly diversity in a
Swiss arable landscape. Restoration Ecology 19: 500-508
BRÄNDLE M., AMARELL U., AUGE H., KLOTZ S. & BRANDL R. 2001. Plant and
insect diversity along a pollution gradient: understanding species richness across
trophic levels. Biodiversity and Conservation 10: 1497-1511
COLLINS K.L., BOATMAN N.D., WILCOX A. & HOLLAND J. M. 2003. A 5-year
comparison of overwintering phytophagous predator densities within a beetle bank
and two conventional hedgebanks. Annals of Applied Biology 143: 63-71
CORBET S.A. 1995: Inscects, plants and succession: advantages of long-term set-
aside. Agriculture, Ecosystems and Environment 53: 201-217
DE HEER M., KAPOS V. & ten BRINK B.J.E. 2005. Biodiversity trends in Europe:
development and testing of a species trend indicator for evaluating progress towards
the 2010 target. Philosophical Transactions of the Royal Society B-Biological
Sciences 360: 297-308
DENYS C. & TSCHARNTKE T. 2002. Plant-insect communities and predator-prey
ratios in field margin strips, adjacent crop fields, and fallows. Oecologia 130: 315-
324
DI GIULIO M., EDWARDS P. J. & MEISTER E. 2001. Enhancing insect diversity in
agricultural grasslands: the roles of management and landscape structure. Journal of
Applied Ecology 38: 310-319
DOLLING W.R. 1991. The Hemiptera. Oxford Uni. Press. Oxford. 274 pp
DUELLI P. & OBRIST M. K. 1998. In search of the best correlates for local
organismal biodiverity in cultivated areas. Biodiveristy and Conservation 7: 297-309
FRANK T. & KÜNZLE I. 2006. Effect of early succession in wildflower areas on bug
assemblages (Insecta: Heteroptera). European Journal of Entomology 103: 61-70
FRANK T., KEHRLI P. & GERMANN C. 2007. Density and nutritional condition of
carabid beetles in wildflower areas of different age. Agriculture, Ecosystems and
Environment 120: 377-383
-
25
FRANK T., AESCHBACHER S., BARONE M., KÜNZLE I., LETHMAYER C. &
MOSIMANN C. 2009. Beneficial arthropods respond differentially to wildflower areas
of different age. Annales Zoologici Fennici 46: 465-480
FRANK T., AESCHBACHER S. & ZALLER J. G. 2012. Habitat age affects beetle
diversity in wildflower areas. Agriculture, Ecosystems and Environment 152: 21-26
HAALAND C., NAISBIT R. E. & BERSIER L.-F. 2011. Sown wildflower strips for
insect conservation: a review. Insect Conservation and Diversity 4: 60-80
KŐRÖSI A., BATARY P., OROSZ A., REDEI D. & BALDI A. 2011. Effects of grazing,
vegetation structure and landscape complexity on grassland leafhoppers (Hemiptera:
Auchenorrhyncha) and true bugs (Hemiptera: Heteroptera) in Hungary. Insect
Conservation and Diversity 5: 57-66
LEYER I. & WESCHE K. 2008. Multivariate Statistik in der Ökologie. Eine
Einführung. Springer-Lehrbuch, Berlin
MARSHALL E.J.P. & MOONEN A.C. 2002. Field margins in northern Europe: their
functions and interactions with agriculture. Agriculture, Ecosystems and Environment
89: 5-21
OTTO A. 1996. Die Wanzenfauna montaner Magerwiesen und Grünbrachen im
Kanton Tessin (Insecta, Heteroptera). Ph.D.thesis, ETH, Zürich.
ÖCKINGER E. & SMITH H. G. 2007. Semi-natural grasslands as population sources
for pollinating insects in agricultural landscapes. Journal of Applied Ecology 44: 50-
59
RABITSCH W. 2007. Rote Listen ausgewählter Tiergruppen Niederösterreichs.
Wanzen (Heteroptera). Amt der Niederösterreichischen Landesregierung, Abteilung
Naturschutz & Abteilung Kultur und Wissenschaft, St. Pölten, 280pp
SACHS L. 1997. Angewandte Statistik, Anwendung statistischer Methoden. Springer,
Berlin
SCHMIDT-ENTLING M.H. & DÖBELI J. 2009. Sown wildflower areas to enhance
spiders in arable fields. Agriculture, Ecosystems and Environment 133: 19-22
STEFFAN-DEWENTER I. & TSCHARNTKE T. 2000. Butterfly community structure in
fragmented habitats. Ecology Letters 3: 449-456
STRAUSS G. 2010. CORISA Wanzenabbildungen. Biberbach. www.corisa.de
-
26
TER BRAAK C.J.F. & ŠMILAUER P. 2002. Canoco Reference Manual and
CanoDraw for Windows User`s Guide. Software for Canonical Community Ordination
(version 4.5). Biometris, Wageningen and Česke Budějovice
THOMAS C. F. G. & MARSHALL E. J. P. 1999. Arthropod abundance and diversity in
differently vegetated margins of arable fields. Agriculture, Ecosystems and
Environment 72: 131-144
TSCHARNTKE T. & KRUESS A. 1999. Habitat fragmentation and biological control.
In: HAWKINS B.A. & CORNELL H.V. (eds). Theoretical Approaches to Biological
Control. Cambridge University Press, Cambridge, 190-20 pp
TSCHARNTKE T., KLEIN A.M., KRUESS A., STEFFAN-DEWENTER I. & THIES C.
2005. Landscape perspectives on agricultural intensification and biodiversity –
ecosystem service management. Ecology Letters 8: 857-874
TSCHARNTKE T., BATARY P. & DORMANN C.F. 2011. Set-aside management:
How do succession, sowing patterns and landscape context affect biodiversity?
Agriculture, Ecosystems and Environment 143: 37-44
ULLRICH K. S. & EDWARDS P. J. 1999.The colonization of wildflower-strips by
insects (Heteroptera). In MAUDSLEY M. J. & MARSHALL E. P. J. (eds),
Heterogeneity in Landscape Ecology. Proceedings of International Association for
Landscape Ecology, UK, 131-138 pp
ULLRICH K. S. 2001. The influence of wildflower-strips on plant and insect
(Heteroptera) diversity in an arable landscape. PhD thesis, ETH Zürich, 127pp
WACHMANN E., MELBER A. & DECKERT J. 2004. Die Tierwelt Deutschlands.
Vol.75. Wanzen Band 2 Cimicomorpha. Goecke & Evers, Kletern
WACHMANN E., MELBER A. & DECKERT J. 2006. Die Tierwelt Deutschlands.
Vol.77. Wanzen Band 1. Goecke & Evers, Keltern
WACHMANN E., MELBER A. & DECKERT J. 2007. Die Tierwelt Deutschlands.
Vol.78. Wanzen Band 3 Pentatomorpha I. Goecke & Evers, Keltern
WACHMANN E., MELBER A. & DECKERT J. 2008. Die Tierwelt Deutschlands.
Vol.81. Wanzen Band 4 Pentatomorpha II. Goecke & Evers, Keltern
WAGNER E. 1952. Blindwanzen und Miriden. Die Tierwelt Deutschlands und der
angrenzendenden Meeresteile 41. Gustav Fischer-Verlag, Jena, 218pp
-
27
WAGNER E. 1966. Die Tierwelt Deutschlands und der angrenzenden Meeresteile
nach ihren Merkmalen und nach ihrer Lebensweise. Vol. 54. Wanzen oder
Heteroptera I. Pentatomorpha. Fischer, Jena, 235pp
WAGNER E. 1967. Die Tierwelt Deutschlands und der angrenzenden Meeresteile
nach ihren Merkmalen und nach ihrer Lebensweise. Vol. 55. Wanzen oder
Heteroptera II. Cimicomorpha. Fischer, Jena, 235pp
WHITE P. C. L. & HASSAL M. 1994. Effects of management on spider communities
of headlands in cereal fields. Pedobiologia 38: 169-184
ZURBRÜGG C. & FRANK T. 2006. Factors influencing bug diversity (Insecta:
Heteroptera) in semi-natural habitats. Biodiversity and Conservation 15: 275-294
Internetquellen:
BIOFORSCHUNG AUSTRIA. 2012. http://www.bioforschung.at/Vertragsnaturschutz-
Lebensrau.244.0.html
WETTERSTATION-OBERLAA. 2012.
http://www.wetterstationoberlaa.at/index.php?option=com_content&view=article&id=2
46&Itemid=323
http://www.bioforschung.at/Vertragsnaturschutz-Lebensrau.244.0.htmlhttp://www.bioforschung.at/Vertragsnaturschutz-Lebensrau.244.0.html
-
28
6 Danksagung
An dieser Stelle möchte ich mich bei meinem Betreuer Prof. Dr. Thomas Frank für
die großartige Unterstützung und Hilfestellung bedanken. Dass er immer Zeit für
mich hatte, wenn Fragen oder Probleme auftraten.
Mein Dank richtet sich auch an Norbert Schuller, dass er immer Zeit für mich hatte,
wenn ich eine Vielzahl an Utensilien brauchte und der mich immer bei Laune hielt mit
seiner Kleintierhaltung.
Dem Institut für Zoologie danke ich für das Bereitstellen einer Räumlichkeit, sowie
der freundlichen Aufnahme im Institut.
Bei Rudi Schmid und Cornelia Heininger möchte ich mich bedanken, dass sie für
mich die Vegetationsaufnahmen in dem Untersuchungsgebiet durchgeführt haben.
Ferner bedanke ich mich bei Ronnie Walcher und Claudia Lichtenegger für die gute
Zusammenarbeit und den regen Gedankenaustausch hinsichtlich unserer
Masterarbeiten.
Janet Wissuwa, Jessica Cornils und Henriette Krebs haben mit mir unzählige
Stunden verbracht, um mir die Statistik näher zu bringen.
Der größte Dank gilt Rudolf Dobrounig und meinen Eltern, die während meines
Studiums immer an meiner Seite waren und mich immer mit Rat und Tat
unterstützen.
-
29
Appendix 1
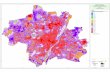
Karte zeigt einen Teil des Untersuchungsgebiets in Oberlaa. Die untersuchten
Habitattypen wurden durch verschiedene Farben ersichtlich gemacht.
Farbverzeichnis: Ackerbrachen (hellrot), Weizenfelder angrenzend an Ackerbrache
(dunkelrot), Windschutzgürtel (grün), Weizenfelder angrenzend an Windschutzgürtel
(gelb) und Kontrollweizenflächen (blau). (Quelle: http://maps.google.at)
-
30
Appendix 2
Karte zeigt einen Teil des Untersuchungsgebiets in Oberlaa. Die untersuchten
Habitattypen wurden durch verschiedene Farben ersichtlich gemacht.
Farbverzeichnis: Ackerbrachen (hellrot), Weizenfelder angrenzend an Ackerbrache
(dunkelrot), Windschutzgürtel (grün), Weizenfelder angrenzend an Windschutzgürtel
(gelb) und Kontrollweizenflächen (blau). (Quelle: http://maps.google.at)
-
31
Appendix 3
Karte zeigt einen Teil des Untersuchungsgebiets in Oberlaa. Die untersuchten
Habitattypen wurden durch verschiedene Farben ersichtlich gemacht.
Farbverzeichnis: Ackerbrachen (hellrot), Weizenfelder angrenzend an Ackerbrache
(dunkelrot), Windschutzgürtel (grün), Weizenfelder angrenzend an Windschutzgürtel
(gelb) und Kontrollweizenflächen (blau). (Quelle: http://maps.google.at)
-
32
Appendix 4
SIMPER (Similarity Percentages - species contributions)
SIMPER berechnet die Ähnlichkeit einer Gruppe und die mittleren Wiederholungen
zwischen allen Gruppen. Es werden jene Wanzenarten aufgezeigt, die an der
Ähnlichkeit innerhalb der Habitattypen beteiligt sind.
(a) Gruppe Hecke
Die Ähnlichkeit beträgt null
(b) Guppe Hecke-Weizen
Durchschnittliche Ähnlichkeit: 44.57
Wanzenarten Av.Abund Av.Sim Sim/SD Contrib% Cum.%
PLAGI 3.11 20.12 2.91 45.14 45.14
TRIGO 1.54 11.86 2.80 26.60 71.73
CLOST 2.27 9.32 0.83 20.91 92.64
LYGUS 1.37 3.28 0.41 7.36 100.00
(c) Gruppe Brachen
Durchschnittliche Ähnlichkeit: 51.66
Wanzenarten Av.Abund Av.Sim Sim/SD Contrib% Cum.%
MEGAL 5.32 12.84 2.69 24.86 24.86
PLAGI 5.15 9.44 1.36 18.28 43.14
LEPTO 2.55 5.19 1.61 10.05 53.19
POLYU 2.69 4.86 2.71 9.41 62.60
STENO 3.10 4.36 0.85 8.44 71.04
BRACH 2.37 4.26 2.51 8.24 79.28
CLOST 2.18 3.63 3.16 7.03 86.31
NOTOS 0.93 1.72 0.90 3.33 89.63
ONCOT 2.32 1.68 0.41 3.26 92.89
ADELP 1.30 1.61 0.89 3.12 96.01
-
33
(d) Gruppe Brache-Weizen
Durchschnittliche Ähnlichkeit: 48.18
Wanzenarten Av.Abund Av.Sim Sim/SD Contrib% Cum.%
CLOST 2.92 25.16 5.66 52.22 52.22
PLAGI 1.68 6.03 0.83 12.51 64.72
EURYG 1.16 5.91 0.81 12.27 76.99
ADELP 0.85 4.42 0.90 9.17 86.15
NABIS 0.85 4.42 0.90 9.17 95.32
(e) Gruppe Kontrollweizen
Durchschnittliche Ähnlichkeit: 44.39
Wanzenarten Av.Abund Av.Sim Sim/SD Contrib% Cum.%
LYGUS 1.30 14.25 0.87 32.11 32.11
TRIGO 1.24 10.05 0.85 22.63 54.75
EURYG 0.85 7.73 0.88 17.41 72.15
PLAGI 1.00 7.73 0.88 17.41 89.56
CLOST 0.50 2.32 0.41 5.22 94.78
NABIS 0.50 2.32 0.41 5.22 100.00
(f) Gruppe Hecke & Hecke-Weizen
Durchschnittliche Unähnlichkeit = 93.17
Wanzenarten Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
PLAGI 0.25 3.11 23.85 2.56 25.60 25.60
CLOST 0.00 2.27 17.20 1.47 18.46 44.07
TRIGO 0.00 1.54 14.17 2.40 15.21 59.28
LYGUS 0.00 1.37 12.58 0.97 13.50 72.78
MEGAL 0.25 0.25 3.83 0.71 4.11 76.89
TRIGR 0.00 0.25 3.18 0.56 3.41 80.30
ADELP 0.25 0.25 3.15 0.72 3.38 83.68
DERAR 0.00 0.35 2.68 0.56 2.88 86.56
NABIS 0.00 0.35 2.54 0.56 2.73 89.29
CAPSO 0.00 0.25 2.09 0.56 2.25 91.54
-
34
CAPSU 0.00 0.25 2.09 0.56 2.25 93.79
LEPTO 0.00 0.25 2.09 0.56 2.25 96.03
(g) Gruppe Hecke & Brache
Durchschnittliche Unähnlichkeit = 95.92
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
MEGAL 0.25 5.32 16.37 1.95 17.07 17.07
PLAGI 0.25 5.15 13.39 1.93 13.96 31.03
STENO 0.00 3.10 9.22 1.14 9.61 40.64
LEPTO 0.00 2.55 7.83 1.73 8.17 48.81
POLYU 0.00 2.69 7.50 2.41 7.82 56.63
CLOST 0.00 2.18 7.40 1.09 7.72 64.34
BRACH 0.00 2.37 6.30 3.54 6.56 70.90
ONCOT 0.00 2.32 5.25 0.94 5.47 76.38
NOTOS 0.00 0.93 3.28 1.33 3.42 79.80
ADELP 0.25 1.30 3.09 1.40 3.22 83.02
CRIOC 0.00 0.85 2.40 0.94 2.51 85.53
PLATY 0.00 1.06 2.29 0.93 2.39 87.92
ONCOV 0.00 1.06 2.09 0.56 2.18 90.10
EURYG 0.00 0.60 1.70 0.94 1.77 91.87
GRAPH 0.00 0.50 1.44 0.88 1.50 93.38
APOLY 0.00 0.61 1.21 0.56 1.26 94.64
POLYH 0.00 0.25 0.95 0.56 0.99 95.63
(h) Gruppe Hecke-Weizen & Brachen
Durchschnittliche Unähnlichkeit = 78.15
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
MEGAL 0.25 5.32 12.13 2.17 15.52 15.52
STENO 0.00 3.10 6.97 1.22 8.92 24.44
PLAGI 3.11 5.15 6.76 2.08 8.64 33.08
POLYU 0.00 2.69 5.78 2.28 7.39 40.48
LEPTO 0.25 2.55 5.33 1.59 6.81 47.29
-
35
BRACH 0.00 2.37 4.90 2.95 6.27 53.56
CLOST 2.27 2.18 4.49 1.27 5.74 59.30
ONCOT 0.00 2.32 4.26 0.95 5.45 64.75
TRIGO 1.54 0.00 3.59 2.72 4.59 69.34
LYGUS 1.37 0.00 3.17 0.89 4.06 73.39
ADELP 0.25 1.30 2.42 1.37 3.09 76.49
NOTOS 0.25 0.93 2.05 1.16 2.62 79.11
PLATY 0.00 1.06 1.88 0.92 2.40 81.51
CRIOC 0.00 0.85 1.83 0.96 2.34 83.85
ONCOV 0.00 1.06 1.75 0.56 2.23 86.09
APOLY 0.25 0.61 1.36 0.80 1.74 87.82
EURYG 0.00 0.60 1.29 0.96 1.66 89.48
GRAPH 0.00 0.50 1.10 0.91 1.40 90.88
CAPSU 0.25 0.25 0.83 0.73 1.06 91.94
DERAR 0.35 0.00 0.79 0.54 1.01 92.95
NABIS 0.35 0.00 0.78 0.54 0.99 93.94
POLYH 0.00 0.25 0.69 0.56 0.88 94.82
TRITO 0.00 0.25 0.69 0.56 0.88 95.70
(i) Gruppe Hecke & Brache-Weizen
Durchschnittliche Unähnlichkeit = 93.75
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
CLOST 0.00 2.92 27.59 3.95 29.43 29.43
PLAGI 0.25 1.68 12.94 1.22 13.81 43.24
EURYG 0.00 1.16 12.50 1.12 13.33 56.57
LYGUS 0.00 1.18 8.58 0.95 9.15 65.72
NABIS 0.00 0.85 7.07 1.65 7.54 73.26
ADELP 0.25 0.85 6.70 1.18 7.15 80.41
TRIGO 0.00 0.43 4.55 0.56 4.85 85.26
CAMPY 0.00 0.25 3.21 0.56 3.42 88.68
NABIF 0.00 0.25 3.21 0.56 3.42 92.10
NOTOS 0.00 0.43 2.79 0.56 2.97 95.08
-
36
(j) Gruppe Hecke-Weizen & Brache-Weizen
Durchschnittliche Unähnlichkeit = 57.86
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
PLAGI 3.11 1.68 9.29 1.35 16.05 16.05
CLOS 2.27 2.92 7.37 1.17 12.75 28.80
LYGUS 1.37 1.18 7.02 1.15 12.13 40.93
EURYG 0.00 1.16 5.95 1.21 10.28 51.21
TRIGO 1.54 0.43 5.93 1.67 10.24 61.45
NABIS 0.35 0.85 3.63 1.41 6.27 67.73
ADELP 0.25 0.85 3.48 1.21 6.02 73.75
NOTOS 0.25 0.43 2.35 0.79 4.06 77.80
LEPTO 0.25 0.35 2.08 0.79 3.60 81.40
DERAR 0.35 0.00 1.57 0.55 2.72 84.12
MEGAL 0.25 0.00 1.46 0.55 2.53 86.65
TRIGR 0.25 0.00 1.46 0.55 2.53 89.19
CAMPY 0.00 0.25 1.41 0.55 2.44 91.62
NABIF 0.00 0.25 1.41 0.55 2.44 94.06
CAPSO 0.25 0.00 1.18 0.55 2.04 96.10
(k) Gruppe Brache & Brache-Weizen
Durchschnittliche Unähnlichkeit = 78.20
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
MEGAL 5.32 0.00 12.99 2.19 16.61 16.61
PLAGI 5.15 1.68 8.57 1.82 10.96 27.57
STENO 3.10 0.00 7.08 1.21 9.06 36.63
POLYU 2.69 0.00 5.87 2.28 7.50 44.13
LEPTO 2.55 0.35 5.38 1.56 6.88 51.01
BRACH 2.37 0.00 4.97 2.97 6.36 57.36
ONCOT 2.32 0.00 4.31 0.95 5.52 62.88
CLOST 2.18 2.92 4.08 2.41 5.21 68.09
LYGUS 0.00 1.18 2.59 0.89 3.31 71.40
NOTOS 0.93 0.43 2.28 1.29 2.91 74.31
EURYG 0.60 1.16 2.23 1.11 2.85 77.16
-
37
ADELP 1.30 0.85 2.07 1.31 2.64 79.81
NABIS 0.00 0.85 1.94 1.51 2.48 82.28
PLATY 1.06 0.00 1.90 0.92 2.43 84.71
CRIOC 0.85 0.00 1.86 0.95 2.38 87.09
ONCOV 1.06 0.00 1.76 0.56 2.26 89.35
GRAPH 0.50 0.00 1.12 0.91 1.43 90.77
TRIGO 0.00 0.43 1.06 0.54 1.35 92.12
APOLY 0.61 0.00 1.02 0.56 1.30 93.43
CAMPY 0.25 0.25 0.83 0.70 1.07 94.49
POLYH 0.25 0.00 0.70 0.56 0.89 95.39
(l) Gruppe Hecke & Kontrollweizen
Durchschnittliche Unähnlichkeit = 92.84
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
LYGUS 0.00 1.30 21.16 1.33 22.79 22.79
TRIGO 0.00 1.24 19.50 1.25 21.00 43.80
EURYG 0.00 0.85 13.11 1.25 14.12 57.92
PLAGI 0.25 1.00 12.17 1.35 13.11 71.03
CLOST 0.00 0.50 7.22 0.86 7.77 78.81
NABIS 0.00 0.50 7.22 0.86 7.77 86.58
ADELP 0.25 0.25 6.07 0.71 6.54 93.12
MEGAL 0.25 0.00 4.05 0.53 4.37 97.48
(m) Gruppe Hecke-Weizen & Kontrollweizen
Durchschnittliche Unähnlichkeit = 62.18
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
PLAGI 3.11 1.00 13.23 1.62 21.28 21.28
CLOST 2.27 0.50 11.50 1.53 18.50 39.78
LYGUS 1.37 1.30 8.04 1.40 12.93 52.70
EURYG 0.00 0.85 5.10 1.40 8.20 60.90
TRIGO 1.54 1.24 4.81 1.13 7.74 68.64
NABIS 0.35 0.50 3.51 1.05 5.64 74.28
-
38
ADELP 0.25 0.25 2.23 0.73 3.58 77.87
NOTOS 0.25 0.25 2.04 0.74 3.28 81.14
MEGAL 0.25 0.00 1.98 0.55 3.18 84.33
TRIGR 0.25 0.00 1.98 0.55 3.18 87.51
DERAR 0.35 0.00 1.96 0.55 3.15 90.67
CAPSO 0.25 0.00 1.49 0.55 2.40 93.06
CAPSU 0.25 0.00 1.49 0.55 2.40 95.46
(p) Gruppe Brache & Kontrollweizen
Durchschnittliche Unähnlichkeit = 88.89
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
MEGAL 5.32 0.00 14.65 2.11 16.48 16.48
PLAGI 5.15 1.00 10.17 1.73 11.44 27.93
STENO 3.10 0.00 7.94 1.18 8.94 36.86
LEPTO 2.55 0.00 6.74 1.78 7.58 44.44
POLYU 2.69 0.00 6.53 2.32 7.35 51.79
BRACH 2.37 0.00 5.51 3.16 6.20 57.99
CLOST 2.18 0.50 5.01 0.87 5.64 63.63
ONCOT 2.32 0.00 4.71 0.95 5.30 68.93
LYGUS 0.00 1.30 3.39 1.50 3.82 72.74
TRIGO 0.00 1.24 3.21 1.35 3.61 76.35
ADELP 1.30 0.25 2.71 1.39 3.05 79.40
NOTOS 0.93 0.25 2.40 1.14 2.70 82.10
CRIOC 0.85 0.00 2.08 0.95 2.34 84.44
PLATY 1.06 0.00 2.07 0.92 2.32 86.76
ONCOV 1.06 0.00 1.91 0.56 2.14 88.91
EURYG 0.60 0.85 1.72 1.10 1.93 90.84
NABIS 0.00 0.50 1.28 0.91 1.44 92.27
GRAPH 0.50 0.00 1.25 0.90 1.40 93.68
APOLY 0.61 0.00 1.10 0.56 1.24 94.92
POLYH 0.25 0.00 0.80 0.56 0.90 95.81
-
39
(q) Gruppe Brache-Weizen & Kontrollweizen
Durchschnittliche Unähnlichkeit = 60.90
Wanzenarten Av.Abun Av.Abund Av.Diss Diss/SD Contrib% Cum.%
CLOST 2.92 0.50 15.72 2.60 25.81 25.81
PLAGI 1.68 1.00 8.11 1.22 13.33 39.14
LYGUS 1.18 1.30 7.54 1.23 12.38 51.52
TRIGO 0.43 1.24 6.91 1.28 11.35 62.87
EURYG 1.16 0.85 5.61 1.00 9.21 72.07
ADELP 0.85 0.25 4.60 1.19 7.55 79.62
NABIS 0.85 0.50 3.82 1.07 6.28 85.90
NOTOS 0.43 0.25 2.87 0.78 4.71 90.61
CAMPY 0.25 0.00 1.99 0.55 3.27 93.88
NABIF 0.25 0.00 1.99 0.55 3.27 97.16
-
40
Appendix 5
Liste der vorgefunden Pflanzenarten in den vier Ackerbrachen
(Vegetationsaufnahmen 2011: durchgeführt von Cornelia Heininger von der
Universität Wien)
Achillea millefolium Hypericum perforatum
Anthemis tinctoria Isatis tinctoria
Arrhenatherum elatius Knautia arvensis
Barbarea vulgaris Lamium maculatum
Bromus inermis Lathyrus tuberosus
Bromus sterilis Leonurus cardiac
Carduus acanthoides Malva moschata
Centaurea jacea Oenothera biennis
Centaurea scabiosa Phleum pratense
Centaurea stoebe Plantago lanceolata
Chenopodium album Poa angustifolia
Cichorium intybus Poa pratensis
Cirsium arvense Reseda lutea
Dactylis glomerata Rosa canina
Daucus carota Salvia nemorosa
Dipsacus fullonum Salvia sclarea
Dipsacus laciniatus Salvia verticillata
Echinops sphaerocephalus Saponaria officinalis
Echium vulgare Seseli libanotis
Elymus repens Silene vulgaris
Falcaria vulgaris Tanacetum vulgare
Erigeron annuus Tripleurospermum inodorum
Galium album Trisetum flavescens
Galium verum Verbascum thapsus
-
41
Appendix 6
Liste der vorgefunden Pflanzenarten in den zwölf Winterweizenfeldern
Anthemis austriaca Papaver rhoeas
Camelina microcarpa Stellaria media
Consolida regalis Tripleurospermum inodorum
Descurainia sophia Veronica hederifola
Galium aparine Veronica polita
Lactua serriola Viola arvensis
Liste der vorgefunden Pflanzenarten in den vier Windschutzgürteln (ohne Sträucher
und Bäume)
Alliaria petiolata Geum urbanum
Arrhenatherum elatius Stellaria media
Ballota nigra Urtica dioica
Bromus sterilis
(Vegetationsaufnahmen 2011: durchgeführt von Rudi Schmid von Bioforschung
Austria)
Diplomarbeit - Corinna Eggenreich - Diversität von Wanzen in ökologischen Ausgleichsflächen und WeizenfeldernZusammenfassung Abstract Inhaltverzeichnis Abbildungsverzeichnis Tabellenverzeichnis 1 Einleitung 2 Material und Methoden 2.1 Untersuchungsflächen 2.2 Fangmethode und Wanzenparameter 2.3 Vegetationsfaktoren 2.4 Datenanalyse
3 Ergebnisse 3.1 Abundanz und Artenzahl von Wanzen in3.2 Charakterisierung der Wanzengemeinsc
4 Diskussion 5 Literaturverzeichnis 6 Danksagung Appendix 1 Appendix 3 Appendix 4 Appendix 5 Appendix 6
Related Documents