Dino£agellate cysts, freshwater algae and fungal spores as salinity indicators in Late Quaternary cores from Marmara and Black seas Peta J. Mudie a; , Andre ¤ Rochon a , Ali E. Aksu b , Helen Gillespie b a Geological Survey of Canada-Atlantic, Bedford Institute of Oceanography, 1 Challenger Drive, Box 1006, Dartmouth, NS, Canada B2Y 4A2 b Department of Earth Sciences, Centre for Earth Resources Research, Memorial University of Newfoundland, St. John’s, NF, Canada A1B 3X5 Received 15 September 2001; accepted 19 February 2002 Abstract Seismic profiles and mollusks have been used to suggest that from V12 500 to 7000 yr BP, the Black Sea was an isolated freshwater lake containing potable water and implying a surface salinity of 6 1. According to Ryan and Pitman (1999), these circumstances encouraged Neolithic settlement and farming on the shore of the Black Sea. This model conflicts with previous studies of dinoflagellate cysts and seismic profiles from the Marmara Sea. Here we investigate Ryan and Pittman’s model using palynological studies of organic-walled dinoflagellate cysts, acritarchs, freshwater algae, microforaminifera, and fungal remains as tracers of changes in surface salinity for seven cores of pleniglacial to Holocene sediments from the Marmara and Black seas. Core-top data from 16 sites along a salinity gradient from 39.5 in the Aegean Sea to V17 in the Black Sea show that the dinocyst species Impagidinium aculeatum, Impagidinium patulum, Operculodinium israelianum, Polysphaeridium zoharyi and Nematosphaeropsis labyrinthus are markers of Mediterranean Sea water and summer surface salinity s 24, and that Spiniferites cruciformis, Spiniferites inaequalis, Peridinium ponticum, Polykrikos spp. and Quinquecuspis concreta characterize the lower salinity of the Marmara and/or Black seas. The core-top data and correlatable down-core assemblage changes in time-equivalent sapropelic and brown muds show that there is no evidence for differential aerobic decay of dinocysts in the study area. The main acritarchs are Sigmopollis psilatum, Concentricystes cf. C. rubinus and cf. Acritarch-8 of Traverse (1978), all of which are absent from the Aegean Sea and decrease in abundance with increasing salinity; the first two taxa have been reported previously as freshwater species. Fungal remains show a similar distribution pattern to the freshwater acritarchs, indicating their origin from terrestrial environments. Freshwater Chlorococcales are almost confined to the Black Sea but they have rare occurrences in the Aegean, indicating long-distance transport. Microforaminiferal linings are abundant in the Marmara Sea but are absent in deep water of the Black Sea. In the Marmara Sea, mid^late Holocene assemblages ( 6 7 ka) are dominated by Lingulodinium machaerophorum, Operculodinium centrocarpum and halophilic Mediterranean Spiniferites spp. (S. mirabilis, S. hyperacanthus, S. bentorii), and a diversity of heterotrophic protoperidinioid and Polykrikos species are present in both sapropels and brown lutites. In contrast, the early Holocene interval (including sapropels) has fewer halophilic Spiniferites spp. and other Mediterranean taxa (O. israelianum, Polysphaeridium zoharyi) and more low salinity indicators (P. ponticum, 0025-3227 / 02 / $ ^ see front matter Crown Copyright ȣ 2002 Elsevier Science B.V. All rights reserved. PII:S0025-3227(02)00348-1 * Corresponding author. Fax: +1-902-426-4104. E-mail address: [email protected] (P.J. Mudie). MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart Marine Geology 190 (2002) 203^231 www.elsevier.com/locate/margeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Dino£agellate cysts, freshwater algae and fungal spores assalinity indicators in Late Quaternary cores
from Marmara and Black seas
Peta J. Mudie a;�, Andre¤ Rochon a, Ali E. Aksu b, Helen Gillespie b
a Geological Survey of Canada-Atlantic, Bedford Institute of Oceanography, 1 Challenger Drive, Box 1006,Dartmouth, NS, Canada B2Y 4A2
b Department of Earth Sciences, Centre for Earth Resources Research, Memorial University of Newfoundland,St. John’s, NF, Canada A1B 3X5
Received 15 September 2001; accepted 19 February 2002
Abstract
Seismic profiles and mollusks have been used to suggest that from V12 500 to 7000 yr BP, the Black Sea was anisolated freshwater lake containing potable water and implying a surface salinity of 6 1. According to Ryan andPitman (1999), these circumstances encouraged Neolithic settlement and farming on the shore of the Black Sea. Thismodel conflicts with previous studies of dinoflagellate cysts and seismic profiles from the Marmara Sea. Here weinvestigate Ryan and Pittman’s model using palynological studies of organic-walled dinoflagellate cysts, acritarchs,freshwater algae, microforaminifera, and fungal remains as tracers of changes in surface salinity for seven cores ofpleniglacial to Holocene sediments from the Marmara and Black seas. Core-top data from 16 sites along a salinitygradient from 39.5 in the Aegean Sea to V17 in the Black Sea show that the dinocyst species Impagidinium aculeatum,Impagidinium patulum, Operculodinium israelianum, Polysphaeridium zoharyi and Nematosphaeropsis labyrinthus aremarkers of Mediterranean Sea water and summer surface salinity s 24, and that Spiniferites cruciformis, Spiniferitesinaequalis, Peridinium ponticum, Polykrikos spp. and Quinquecuspis concreta characterize the lower salinity of theMarmara and/or Black seas. The core-top data and correlatable down-core assemblage changes in time-equivalentsapropelic and brown muds show that there is no evidence for differential aerobic decay of dinocysts in the studyarea. The main acritarchs are Sigmopollis psilatum, Concentricystes cf. C. rubinus and cf. Acritarch-8 of Traverse(1978), all of which are absent from the Aegean Sea and decrease in abundance with increasing salinity; the first twotaxa have been reported previously as freshwater species. Fungal remains show a similar distribution pattern to thefreshwater acritarchs, indicating their origin from terrestrial environments. Freshwater Chlorococcales are almostconfined to the Black Sea but they have rare occurrences in the Aegean, indicating long-distance transport.Microforaminiferal linings are abundant in the Marmara Sea but are absent in deep water of the Black Sea. In theMarmara Sea, mid^late Holocene assemblages (6 7 ka) are dominated by Lingulodinium machaerophorum,Operculodinium centrocarpum and halophilic Mediterranean Spiniferites spp. (S. mirabilis, S. hyperacanthus, S.bentorii), and a diversity of heterotrophic protoperidinioid and Polykrikos species are present in both sapropels andbrown lutites. In contrast, the early Holocene interval (including sapropels) has fewer halophilic Spiniferites spp. andother Mediterranean taxa (O. israelianum, Polysphaeridium zoharyi) and more low salinity indicators (P. ponticum,
0025-3227 / 02 / $ ^ see front matter Crown Copyright B 2002 Elsevier Science B.V. All rights reserved.PII: S 0 0 2 5 - 3 2 2 7 ( 0 2 ) 0 0 3 4 8 - 1
* Corresponding author. Fax: +1-902-426-4104.E-mail address: [email protected] (P.J. Mudie).
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
Marine Geology 190 (2002) 203^231
www.elsevier.com/locate/margeo

Pyxidinopsis psilata and S. cruciformis), suggesting that the overflow of Black Sea water was greater than now. Thetime-equivalent early Holocene unit in the Black Sea has a higher percentage of taxa found in freshwaterenvironments (including S. cruciformis forms 1^3; Gonyaulax apiculata, fungal remains and freshwater acritarchs) butthe continued presence of Spiniferites morphotypes and other taxa associated with brackish (V7^18) to saline surfacewater indicates that there was some Mediterranean water present at this time. The late glacial and Pleniglacialsediments in the Marmara Sea contain a lower diversity of dinoflagellates, dominated by S. cruciformis and P. psilata,as in early Holocene sediments of the Black Sea but including the short-spine morphotypes of S. cruciformis which arenot found in freshwater lakes. We conclude that although the glacial stages were marked by much lower sea-surfacetemperatures, there was either some periodic marine influence or the marine dinocysts were living in a brackish waterenvironment (V7^18) with the freshwater species being transported from glacial lakes. Overall, there is nopalynological evidence that the surface salinity of the Black or Marmara seas was ever as low as a freshwater lake.This finding is consistent with models that estimate the time required to desalinate the Black Sea after closing of theMarmara^Bosphorus gateway.Crown Copyright B 2002 Elsevier Science B.V. All rights reserved.
Keywords: dino£agellate cysts; acritarchs; fungi; freshwater algae; Marmara Sea; Black Sea; paleosalinity; Quaternary palynology;aerobic decay
1. Introduction
Previous studies of Late Quaternary sedimentsfrom the Aegean, Black and Marmara seas haveshown that dino£agellate cyst (dinocyst) assem-blages are important tracers of changes in surfacewater salinity in these epicontinental seas (Walland Dale, 1973; Aksu et al., 1995a; Dale, 1996;Mudie et al., 2001). In the Mediterranean andBlack seas, both calcareous and organic-walleddinocysts are found in the Holocene sediments.Of these groups, the organic-walled dinocystsare especially useful for paleoceanographic studiesbecause they are highly resistant to dissolutionwhich can alter the taphonomic representationof calcareous and siliceous microplankton. Earlystudies of Plio^Pleistocene sediments in the BlackSea by Traverse (1974) also used organic-walleddino£agellate cysts, together with acritarchs (algalspores of unknown or uncertain biological a⁄n-ity), as an index of marine versus freshwater con-ditions. Other freshwater colonial algae, such asPediastrum, were noted as potential markers ofincreased runo¡ to the Black Sea (e.g. Roman,1974; Traverse, 1974).
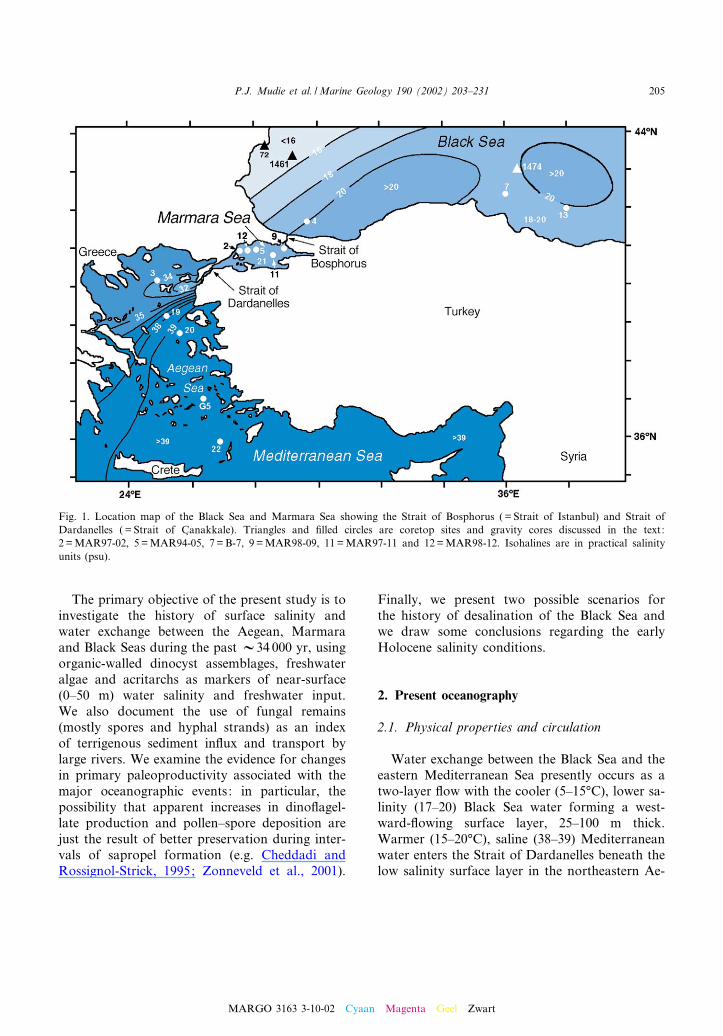
The Marmara Sea is a critical gateway for ex-change of water between the Black Sea, a perma-nently anoxic basin, and the Aegean Sea, which isa branch of the eastern Mediterranean Sea(Fig. 1). The Marmara Sea is presently linked to
the Black Sea through the V40-m-deep Strait ofBosphorus (Strait of Istanbul) and with the Ae-gean Sea through the V70-m-deep Strait of Dar-danelles (Strait of CManakkale). The importance ofthis gateway in the Quaternary paleoceanographicevolution of the eastern Mediterranean has longbeen recognized, particularly with regard to therole of freshwater runo¡ in promoting the devel-opment of sapropel layers (e.g. Vergnaud-Grazzi-ni et al., 1977; Thunell and Williams, 1989; Aksuet al., 1995a,b). Previous studies have suggestedthat during glacial periods, the levels of the Ae-gean and Black seas were below the Bosphorusand Dardanelles sills so that the inland Marmaraand Black seas became isolated basins, cut o¡from the Mediterranean Sea. Following the risein glacio-eustatic sea level from a lowstand of3110 m at V11 000 yr BP (Aksu et al., 1999b,2002a), the Bosphorus sill was breached andBlack Sea water was exported through the Mar-mara gateway to the Aegean Sea. However, thetiming of this event is controversial. Aksu et al.(1995a,b) correlated this £ooding event with de-position of Aegean sapropel S1 from 9600 to6400 yr BP, while Ryan et al. (1997) and Ryanand Pitman (1999) believe that the Bosphorus wasnot breached until 7000 yr BP when the MarmaraSea cascaded into the Black Sea catastrophicallyre-¢lling the basin and contributing to the Noah’sFlood myth.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231204
The primary objective of the present study is toinvestigate the history of surface salinity andwater exchange between the Aegean, Marmaraand Black Seas during the past V34 000 yr, usingorganic-walled dinocyst assemblages, freshwateralgae and acritarchs as markers of near-surface(0^50 m) water salinity and freshwater input.We also document the use of fungal remains(mostly spores and hyphal strands) as an indexof terrigenous sediment in£ux and transport bylarge rivers. We examine the evidence for changesin primary paleoproductivity associated with themajor oceanographic events: in particular, thepossibility that apparent increases in dino£agel-late production and pollen^spore deposition arejust the result of better preservation during inter-vals of sapropel formation (e.g. Cheddadi andRossignol-Strick, 1995; Zonneveld et al., 2001).
Finally, we present two possible scenarios forthe history of desalination of the Black Sea andwe draw some conclusions regarding the earlyHolocene salinity conditions.
2. Present oceanography
2.1. Physical properties and circulation
Water exchange between the Black Sea and theeastern Mediterranean Sea presently occurs as atwo-layer £ow with the cooler (5^15‡C), lower sa-linity (17^20) Black Sea water forming a west-ward-£owing surface layer, 25^100 m thick.Warmer (15^20‡C), saline (38^39) Mediterraneanwater enters the Strait of Dardanelles beneath thelow salinity surface layer in the northeastern Ae-
Fig. 1. Location map of the Black Sea and Marmara Sea showing the Strait of Bosphorus ( = Strait of Istanbul) and Strait ofDardanelles ( = Strait of CManakkale). Triangles and ¢lled circles are coretop sites and gravity cores discussed in the text:2 =MAR97-02, 5 =MAR94-05, 7= B-7, 9 =MAR98-09, 11=MAR97-11 and 12=MAR98-12. Isohalines are in practical salinityunits (psu).
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 205

gean Sea and £ows northeast (Oº zsoy et al., 1995;Polat and Tug›rul, 1996) to occupy the entire Mar-mara basin below the low salinity surface layer.After crossing the Strait of Bosphorus, the salinewater penetrates the Black Sea and forms the bot-tom water mass below the 100^200-m-thick sur-face layer.
At present, there is a net export of V300 km3
yr31 of water from the Black Sea into the AegeanSea across the Straits of Bosphorus and Darda-nelles (Oº zsoy et al., 1995). This out£ow resultsfrom the excess of precipitation and river dis-charge to the Black Sea (V650 km3 yr31) relativeto the evaporation (V350 km3 yr31). The Dan-ube, Dniester, Dnieper, Southern Bug and Donrivers are the major sources of freshwater intothe Black Sea, from a drainage area of V2 mil-lion km2.
Surface water circulation in the Black Sea isdominated by two large central cyclonic gyresand several smaller, anticyclonic coastal eddies(Og›uz et al., 1993). A narrow (6 75-km-wide)cyclonic peripheral ‘rim current’ separates thecentral basinal gyres from the coastal eddies(Aksu et al., 2002b). Weak anticyclonic eddiesare also situated east and west of the Strait ofBosphorus. Surface circulation in the MarmaraSea is dominated by the out£ow of low salinityBlack Sea water (BesMiktepe et al., 1994), which£ows as a narrow current for nearly the entirewidth of the basin, but broadens westward, form-ing large meander loops with three weak anticy-clonic gyres as it approaches the Strait of Darda-nelles (Aksu et al., 2002a). The bottom watercirculation in the Black Sea is controlled by therate of in£ux of denser Mediterranean waterthrough the straits of Bosphorus and Darda-nelles. This dense water £ows eastward and coolsover the shelves, then forms turbulent plumesthat sink beneath the halocline and slowly travelseastward into the deep basins between 100 and500 m water depth (Oº zsoy et al., 1995). Mixingof the surface water by winds and winter convec-tion occurs down to V200 m. Mixing of thebottom water layer (V400 m thick), however,is driven only by geothermal heat £ux, with avery slow turnover rate on the order of V2500years.
Table 1List of all the plankton-stage dino£agellates reported fromthe Black Sea (see text for references) and their correspond-ing cyst forms
Thecal stage name Cyst stage name
Ceratium candelabrum no cystCeratium furca no cystCeratium fusus no cystCeratium longipes no cystCeratium longirostrum no cystCeratium hexacanthum no cystCeratium pentagonum no cystCeratium tripos no cystDinophysis acuminata no cystDinophysis acuta no cystDinophysis caudata no cystDinophysis hastata no cystDinophysis homunculus no cystDinophysis ovum no cystDinophysis rotundata no cystDinophysis sacculus no cystDinophysis sphaerica no cystDinophysis sp. no cystExuviaella compressa no cystExuviaella cordata no cystGonyaulax polygramma no cystGoniodoma polyedricum no cystGymnodinium rotundatum no cystGyrodinium fusiforme no cystGyrodinium pellucidum no cystGyrodinium pusillum no cystLingulodinium polyedrum Lingulodinium
machaerophorumNoctiluca scintillans no cystOxytoxum milneri no cystProtoperidinium cinctum no cystProtoperidinium crassipes no cystProtoperidinium de¢ciens no cystProtoperidinium depressum no cystProtoperidinium diabolus no cystProtoperidinium divergens Peridinium ponticumProtoperidinium granii no cystProtoperidinium pentagonum Trinovantedinium applanatumProtoperidinium solidicorne no cystProtoperidinium steinii no cystProtoperidinium sp. no cystPhalacroma parvulum no cystPhalacroma sp. no cystPolykrikos schwartzii Polykrikos schwartziiPronoctiluca pelagica no cystPronoctiluca spinifera no cystProrocentrum micans brown spherical cystProtoceratium reticulatum Operculodinium centrocarpumProrocentrum compressum no cystScrippsiella trochoidea Scrippsiella trochoidea
(calcareous cyst)
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231206

2.2. Biological oceanography
Few data are available which describe the com-position of plankton and benthos in the Marmaraand Aegean seas (see Aubert et al., 1990). How-ever, there are detailed reports for the surfacelayer (0^50 m) of the Black Sea that comprisesmost of the water mass in which the dino£agellatepopulations would be growing. Table 1 lists allthe plankton-stage dino£agellates reported fromthe Black Sea and their corresponding cyst forms.It should be noted that the plankton ecology ofthe Black Sea has changed signi¢cantly since 1970because of NO3, PO4 and organic matter enrich-ment and SiO2 reduction as a consequence of damconstruction (Bodeneau, 1993; Bologa et al.,1995). In the northwest Black Sea, these changeshave resulted in an eight-fold increase in non-dia-tom algae relative to diatoms, including frequentred tide blooms of the dino£agellates Prorocen-trum cordatum, Scrippsiella trochoidea and Hete-rocapsa triquetra, as well as the coccolithophoridEmiliania huxleyi and the diatoms Nitschia, Ske-letonema and Chaetoceros. Summer blooms of thedino£agellate Noctiluca scintillans occur along thecoasts of Ukraine, Georgia and Turkey. Benli(1987) also reports blooms of Protoceratium reticu-latum (cyst form =Operculodinium centrocarpum)
and Prorocentrum micans in the southern BlackSea. In addition to these photosynthetic dino£a-gellate blooms, Bologa et al. (1995) note thatthere has been an increase of V19% in heterotro-phic dino£agellates since 1990, probably mainlyProtoperidinium pentagonum (cyst form =Trino-vantedinium applanatum) which blooms in thesouthern Black Sea (Benli, 1987). Aubert et al.(1990) also report blooms of Peridinium steiniiand Gymnodinium simplex in the Black Sea, andthey note that there is a large decrease in dino-£agellates in the Strait of Bosphorus and the Mar-mara Sea where only E. huxleyi forms blooms. Inthe Aegean Sea, however, there are blooms ofGymnodinium in addition to E. huxleyi and Coc-colithus pelagicus. It is also notable that the diver-sity and production of diatoms in the Aegean Seaare relatively sparse compared to the dino£agel-lates (Aubert et al., 1990).
3. Methods
Cores used in this study (Fig. 1, Table 2) werecollected from the RV Koca Piri Reis of the In-stitute of Marine Sciences and Technology, Do-kuz Eylu« l University, using a 4-m-long corer, with10 cm internal diameter and 400 kg weight. All
Table 2Geographic co-ordinates and water depths (in meters) for core-top samples used in this study
Sample Locality Latitude Longitude Depth(‡N) (‡E) (m)
22 Cretan Trough 35.8 26.8 1050G5 Central Aegean Sea 37.4 26.2 51020 Central Aegean Sea 38.5 24.5 70019 North Aegean Sea 39.1 24.9 5703 NE Aegean Sea 40.2 24.9 5602 W Marmara Sea 40.9 27.6 108012 W Marmara Sea 40.8 27.8 5495 Central Marmara Sea 40.9 28.1 100811 SE Marmara Sea 40.7 28.4 1119 E Marmara Sea 40.9 28.9 644 SW Black Sea 41.5 29.3 1121474 W Black Sea 42.2 37.8 211772 W Black Sea 43.5 28.4 151461 W Black Sea 43.0 30.0 19907 SE Black Sea 42.5 37.0 212013 SE Black Sea 42.0 37.9 2050
Locations of core tops in Fig. 1.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 207
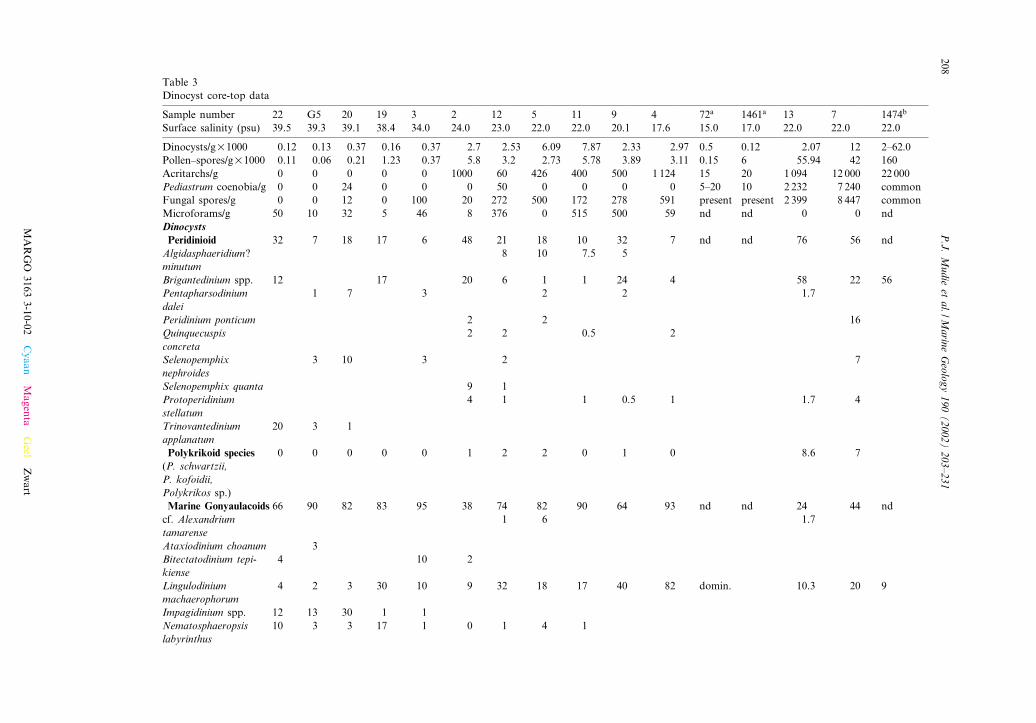
Table 3Dinocyst core-top data
Sample number 22 G5 20 19 3 2 12 5 11 9 4 72a 1461a 13 7 1474b
Surface salinity (psu) 39.5 39.3 39.1 38.4 34.0 24.0 23.0 22.0 22.0 20.1 17.6 15.0 17.0 22.0 22.0 22.0
Dinocysts/gU1000 0.12 0.13 0.37 0.16 0.37 2.7 2.53 6.09 7.87 2.33 2.97 0.5 0.12 2.07 12 2^62.0Pollen^spores/gU1000 0.11 0.06 0.21 1.23 0.37 5.8 3.2 2.73 5.78 3.89 3.11 0.15 6 55.94 42 160Acritarchs/g 0 0 0 0 0 1000 60 426 400 500 1 124 15 20 1 094 12 000 22 000Pediastrum coenobia/g 0 0 24 0 0 0 50 0 0 0 0 5^20 10 2 232 7 240 commonFungal spores/g 0 0 12 0 100 20 272 500 172 278 591 present present 2 399 8 447 commonMicroforams/g 50 10 32 5 46 8 376 0 515 500 59 nd nd 0 0 ndDinocystsPeridinioid 32 7 18 17 6 48 21 18 10 32 7 nd nd 76 56 ndAlgidasphaeridium?minutum
8 10 7.5 5
Brigantedinium spp. 12 17 20 6 1 1 24 4 58 22 56Pentapharsodiniumdalei
1 7 3 2 2 1.7
Peridinium ponticum 2 2 16Quinquecuspisconcreta
2 2 0.5 2
Selenopemphixnephroides
3 10 3 2 7
Selenopemphix quanta 9 1Protoperidiniumstellatum
4 1 1 0.5 1 1.7 4
Trinovantediniumapplanatum
20 3 1
Polykrikoid species 0 0 0 0 0 1 2 2 0 1 0 8.6 7(P. schwartzii,P. kofoidii,Polykrikos sp.)Marine Gonyaulacoids 66 90 82 83 95 38 74 82 90 64 93 nd nd 24 44 nd
cf. Alexandriumtamarense
1 6 1.7
Ataxiodinium choanum 3Bitectatodinium tepi-kiense
4 10 2
Lingulodiniummachaerophorum
4 2 3 30 10 9 32 18 17 40 82 domin. 10.3 20 9
Impagidinium spp. 12 13 30 1 1Nematosphaeropsislabyrinthus
10 3 3 17 1 0 1 4 1
MA
RG
O3163
3-10-02C
yaanM
agentaG
eelZw
art
P.J.
Mudie
etal./M
arineGeology
190(2002)
203^231208
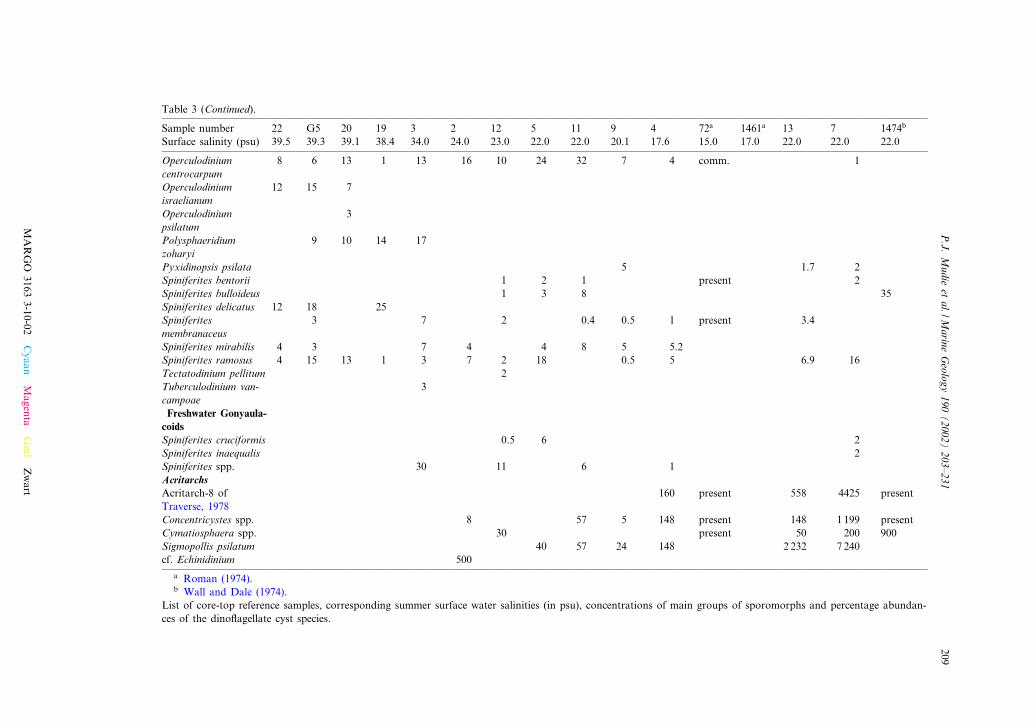
Table 3 (Continued).
Sample number 22 G5 20 19 3 2 12 5 11 9 4 72a 1461a 13 7 1474b
Surface salinity (psu) 39.5 39.3 39.1 38.4 34.0 24.0 23.0 22.0 22.0 20.1 17.6 15.0 17.0 22.0 22.0 22.0
Operculodiniumcentrocarpum
8 6 13 1 13 16 10 24 32 7 4 comm. 1
Operculodiniumisraelianum
12 15 7
Operculodiniumpsilatum
3
Polysphaeridiumzoharyi
9 10 14 17
Pyxidinopsis psilata 5 1.7 2Spiniferites bentorii 1 2 1 present 2Spiniferites bulloideus 1 3 8 35Spiniferites delicatus 12 18 25Spiniferitesmembranaceus
3 7 2 0.4 0.5 1 present 3.4
Spiniferites mirabilis 4 3 7 4 4 8 5 5.2Spiniferites ramosus 4 15 13 1 3 7 2 18 0.5 5 6.9 16Tectatodinium pellitum 2Tuberculodinium van-campoae
3
Freshwater Gonyaula-coidsSpiniferites cruciformis 0.5 6 2Spiniferites inaequalis 2Spiniferites spp. 30 11 6 1AcritarchsAcritarch-8 ofTraverse, 1978
160 present 558 4425 present
Concentricystes spp. 8 57 5 148 present 148 1 199 presentCymatiosphaera spp. 30 present 50 200 900Sigmopollis psilatum 40 57 24 148 2 232 7 240cf. Echinidinium 500
a Roman (1974).b Wall and Dale (1974).
List of core-top reference samples, corresponding summer surface water salinities (in psu), concentrations of main groups of sporomorphs and percentage abundan-ces of the dino£agellate cyst species.
MA
RG
O3163
3-10-02C
yaanM
agentaG
eelZw
art
P.J.
Mudie
etal./M
arineGeology
190(2002)
203^231209

MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231210

cores were stored upright on board ship and wereshipped to Memorial University of Newfoundland(MUN), where they were split, described and pho-tographed, except Core BLA90-07 ( = Core B-7),which was split, described and sampled in Turkey.Stratigraphic studies were made on six cores fromthe Marmara Sea and Black Sea, the sedimentol-ogy of which is described in detail by Hiscott andAksu (2002). One box core was studied from thesoutheastern Black Sea, the sedimentology ofwhich is described by Duman (1992), and forwhich initial dino£agellate cyst studies weremade by Mudie et al. (2001). Additional core-top data for interpretation of the dinocyst eco-stratigraphies and associated sporomorph assem-blages were obtained from six Aegean Sea sites(see Aksu et al., 1995a) and from ¢ve Black Seasites, including published data of Roman (1974),Wall and Dale (1974) and Traverse (1974).
Palynomorphs were extracted from samples of2^5 cm3 volume, using standard methods forQuaternary marine sediments (Rochon et al.,1999): sieving at 20- and 125-Wm mesh sizes, di-gestion in HCl and HF, and adding exotic sporesto obtain estimates of palynomorph concentrationper cm3. Palynomorph preservation is good inmost sections of the cores, but sandy intervalsoften contain many oxidized and abraded grains,while some levels also contain common reworkedpre-Quaternary (mainly Pliocene) pollen or sporesand more rarely, dino£agellate cysts. These re-worked palynomorphs were primarily recognizedby their distinctive morphology, but £attenedand/or yellowish grains of extant pollen typeswere also scored as reworked.
One or two slides of each processed samplewere counted at U25 magni¢cation, until a mini-mum of 300 exotic spores was reached. Thisyielded counts of 100^200 for total dinocysts inthe middle to upper Holocene samples; however,in the late glacial to early Holocene oxidized del-
taic intervals, counts were as low as 25^50 for twoslides and s 1000 exotic spores. Nomenclature ofthe dinocysts follows that used by Williams et al.(1998) and Rochon et al. (1999) except wherenoted. Pollen and spore concentrations reportedhere are from Mudie et al. (2002). During thecounting of dinocysts, pollen and fern+mossspores, records were kept of the number of micro-foraminiferal linings, acritarchs, fungal remainsand freshwater algae, including coenobia of Pe-diastrum and Botryococcus. Identi¢cation of fresh-water algal coenobia and fungal remains followsthe treatments of Matthiessen and Brenner (1996),Kohlmeyer and Kohlmeyer (1979) and Jarzen andElsik (1986), respectively. Counts were also madeof the ‘baggy cysts’ (saccocysts) of Traverse (1974,1988) because of their abundance in the Black Seasediments and their apparent association with lowsalinity.
4. Results
4.1. Core-top distributions of dinocysts and otherpalynomorphs
Table 3 shows the abundances of dinocysts andother sporomorphs in the core-top samples (Fig. 1)from a south^north transect of the Aegean Sea(GC22-KC3), from a transect of the MarmaraSea (MAR97-2 to MAR98-9) and from the west-ern (MAR98-4 to 72) and southeastern Black Sea(BS-13 to 1474). There is a general decrease in theconcentration of most sporomorphs with distanceaway from the relatively low salinity (6 23), nu-trient-rich surface water of the Marmara andBlack Seas, although low values of dinocysts arealso found o¡ the Danube River (sites 72, 1461).The main exception is in the microforaminiferallinings of foraminifera, the shapes of whichmostly resemble benthic species, but large trocho-
Fig. 2. Distribution of dino£agellate cysts and other sporomorphs in MAR97-02. (A) Summary diagram showing lithofacies, var-iations in organic-walled palynomorphs and total organic carbon (OC). Arrows show levels with uncorrected 14C ages in yr BP.Units and OC data are from Hiscott and Aksu (2002) and Abrajano et al. (2002), respectively. (B) Dinocyst diagram showingdown-core variations in abundance and in relative abundances of major species.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 211
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231212

spiroidal forms with the shapes of planktonic spe-cies are also sometimes present (see Aksu et al.,2002a and references therein). These marine paly-nomorphs show maximum abundances in theMarmara Sea and are well represented in the Ae-gean Sea but are absent in the anoxic Black Seabasins. Dinocyst abundances reach their maxi-mum abundance in the southeastern Black Seaand parts of the Marmara Sea. Unfortunately,our palynological data do not include the calcar-eous cysts of Scrippsiella trochoidea that producesred tide blooms in this area (Bodeneau, 1993),and are also common to abundant in the easternMediterranean Sea (Zonneveld et al., 2001). How-ever, scanning electron micrographs of Ross andDegens (1974) show abundant cysts of S. trochoi-dea in the sapropelic unit of the southeasternBlack Sea, in the path of the saline Mediterraneanwater in£ow. In contrast, a search for calcareouscysts in Marmara Sea cores did not reveal anyspecimens although coccoliths and diatoms arepresent (H. Gillespie, unpublished data).
The core-top distributions of chlorococcaleanalgae, acritarch and fungal spore concentrationsshow an order of magnitude decrease from theBlack Sea to Marmara Sea, and are absent atmost sites in the Aegean Sea. The chlorococca-leans are mostly (90%) coenobia of the freshwateralga Pediastrum boryanum, with minor amountsof Botryococcus, which lives in both fresh (6 1)and brackish (V1^10) waters (Matthiessen andBrenner, 1996; Kunz-Pirrung, 1998). Accordingto Batten (1996b), most fungal sporomorphs inpalynological preparations are usually derivedfrom soils in the watersheds and fringing marsh-lands because there are no known aquatic or ma-rine fungi that produce fossilizable spores or otherremains. In the core-top samples, most of the fun-gal remains are dark brown unicellular (Lycoper-don- and Tilletia-type) or dicellular (Puccinellia-type) spores, but in the Black Sea, short, un-
branched hyphal strands are common, alongwith occasional pluricellular spores.
The acritarch Concentricystes sp. appears to bethe same as C. rubinus Rossignol (1961) whichwas reported as being of freshwater origin; Ro-man (1974) also notes that it is also found in soilsof Romania. The acritarchs Cymatiosphaera andSigmopollis psilatum are commonly found in estu-arine environments (Wall and Dale, 1974; Mudie,1992; Solomon et al., 2000). The ‘baggy spores’(cf. Acritarch-8) of Traverse (1978) are thin-walled, pale brown leiospheres like his ‘baggycysts’ (Traverse, 1978) and saccocysts (Traverse,1988), but with no discernible archeopyle. Boththese taxa are common to abundant in the mod-ern sediments of the Black Sea and are presum-ably from fresh or low salinity environments be-cause they are not present in the core tops wheresummer surface salinity is greater than 18.
The modern dinocyst assemblages are mostlydominated by gonyaulacoid species, particularlyLingulodinium machaerophorum, Operculodiniumcentrocarpum and Spiniferites ramosus. There isa large increase in the abundance of protoperidi-nioid species (mostly Brigantedinium spp.) in theBlack Sea. This is in accord with the higher pri-mary production of this sea, but may partly re-£ect better preservation of these oxygen-sensitivecysts (Zonneveld et al., 1997, 2001) in the highlyanaerobic/anoxic sediments of the deep basins. Onthe other hand, the maximum abundances of themoderately oxygen-sensitive species Spiniferitesdelicatus, S. ramosus, S. membranaceus and S.mirabilis are highest in the Aegean Sea wherethe surface sediments are slightly oxidized (Aksuet al., 1995b). However, relatively high percen-tages of protoperidinioids may be found in partsof the Marmara and Aegean where surface sedi-ments are oxidized (e.g. sites 97-2, GC22), and thesensitive protoperidinioid cyst Trinovantediniumapplanatum is also con¢ned to the Aegean Sea,
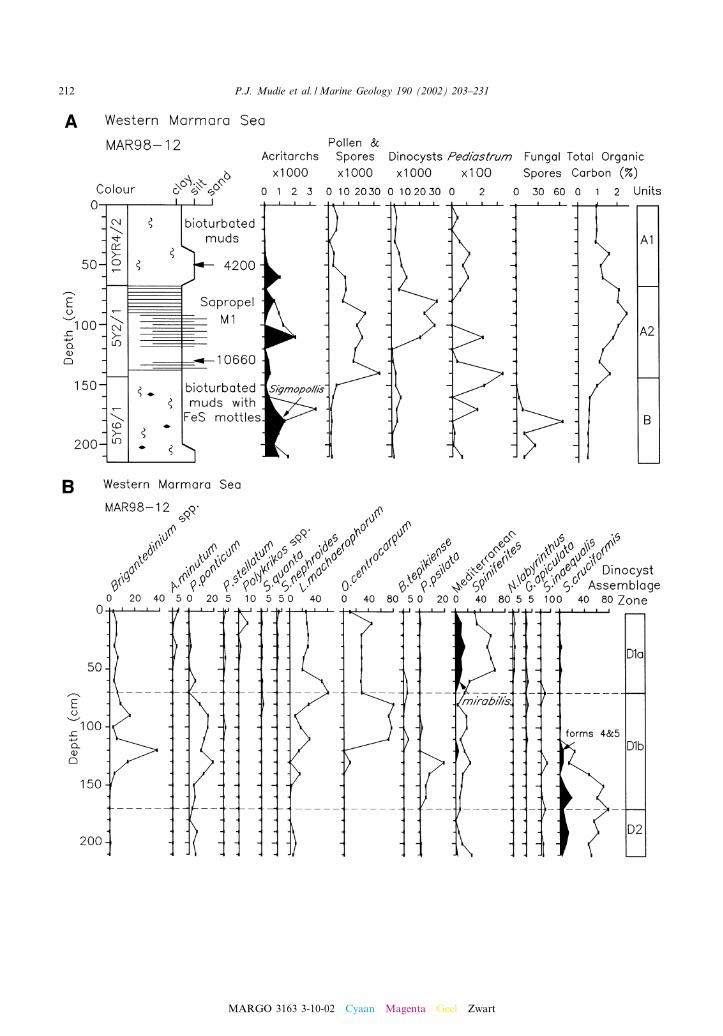
Fig. 3. Distribution of dino£agellate cysts and other sporomorphs in MAR98-12. (A) Summary diagram showing lithofacies, var-iations in organic-walled palynomorphs and total organic carbon (OC). Arrows show levels with uncorrected 14C ages in yr BP.Units and OC data are from Hiscott and Aksu (2002) and Abrajano et al. (2002), respectively. (B) Dinocyst diagram showingdown-core variations in abundance and in relative abundances of major species.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 213

although its thecate form, Protoperidinium penta-gonum, is abundant in the southern Black Sea.
In order to distinguish between assemblage dif-ferences resulting from potential aerobic decayversus salinity preferences, we can consider thesalinity ranges of only the most oxidation-resis-tant cyst species of the Mediterranean Sea region,viz. Impagidinium aculeatum, Operculodinium isra-elianum, Polysphaeridium zoharyi and Nemato-sphaeropsis labyrinthus (Zonneveld et al., 2001).The ¢rst three of these oxidation-resistant speciesare common in the Mediterranean, Adriatic andAlboran seas (see Mudie et al., 2001 and referen-ces therein) and they are absent from the surfacesediments of the Marmara or Black seas. There-fore, it appears that these taxa are reliable indica-tors of Mediterranean Sea water and summer sal-inities s 24. It is also notable that two otherspecies of Spiniferites (S. cruciformis and S. inae-qualis) are found only in the low salinity Blackand Marmara seas, while Peridinium ponticumand Polykrikos (mostly P. kofoidii) have a similardistribution in the inland seas. Quinquecuspis con-creta is present in the intermediate salinities of theMarmara Sea, in keeping with its maximum abun-dance in the strati¢ed relatively low salinitywaters at the head of the Persian Gulf (Bradfordand Wall, 1984).
4.2. Marmara Sea ecostratigraphy
In the western Marmara Sea, two cores werestudied from basinal sediments (MAR97-02 andMAR94-05) and one (MAR98-12) from the sad-dle between the basins. In the eastern MarmaraSea, one core (MAR97-11) was studied from thesouthern shelf edge and one (MAR98-09) fromnear the entrance to the Bosphorus Strait(Fig. 1). Multiple radiocarbon ages for molluskshells (see Aksu et al., 2002a,b) in the westerncores show that the sediments cover an age rangefrom 1990 to 29 540 yr BP (Figs. 2^4), while theeastern cores have ages from 4500 to 15 590 yr BP(Figs. 5 and 6).
The Holocene sediments, Unit A, in all thecores except the condensed sequence of MAR94-05, show a two- to three-fold increase in dinocystsdown-core, followed by a return to modern con-
centrations or lower numbers in the early Holo-cene. Dinocyst assemblages throughout Unit Aare dominated by the euryhaline species Lingulo-dinium machaerophorum, Operculodinium centro-carpum and Spiniferites ramosus, with variableamounts of Brigantedinium and other protoperidi-nioid cysts. Two Holocene dinocyst sub-zones arerecognized. The youngest sub-zone, D1a (0^V7000 yr BP) is dominated by L. machaeropho-rum or, near the Aegean Sea, by O. centrocarpum,together with a relatively high percentage of Spi-niferites mirabilis (here grouped with Spiniferiteshyperacanthus), rare occurrences of Operculodi-nium israelianum and/or Bitectatodinium tepikiense(MAR98-12, MAR97-11 and MAR98-09), and adiversity of heterotrophic taxa, including Algida-sphaeridium? minutum, Polykrikos kofoidii, Sele-nopemphix quanta, Selenopemphix nephroides andProtoperidinium (Stelladinium) stellatum. Theearly Holocene (V7000^10 000 yr BP) sub-zoneD1b is dominated by O. centrocarpum, with var-iable amounts of the low salinity indicators Peri-dinium ponticum, Pyxidinopsis psilata and Spini-ferites cruciformis, absence of Polykrikos and alower diversity of euryhaline Spiniferites and pro-toperidinioid species.
Concentrations of pollen and spores in Unit Ashow approximately the same trend as the dino-cyst concentrations, but there are clear di¡erencesin the duration of the early^mid Holocene max-ima, with the pollen^spore peak beginning severalthousand years earlier than the dinocyst peak inCores MAR97-11 and MAR98-09. All other spo-romorphs in Unit A are marked by two or threelarge peaks and troughs that roughly correspondto intervals of about 2000 years. The only clearpattern is the co-occurrence of peaks of fresh-water indicators, including Pediastrum, acritarchsand multicellular fungal spores at about 9000^10 000 yr BP. These peaks are concurrent withthe maxima found in the early Holocene Unit 3of Black Sea Core B-7 (see 4.3. Black Sea eco-stratigraphy).
The late glacial maximum (LGM) to early post-glacial Unit B (V25 000^11 000 yr BP) in CoresMAR98-12, MAR94-05 and MAR97-11 is distin-guished throughout by low concentrations of di-nocysts dominated (40^80%) by Spiniferites cruci-
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231214
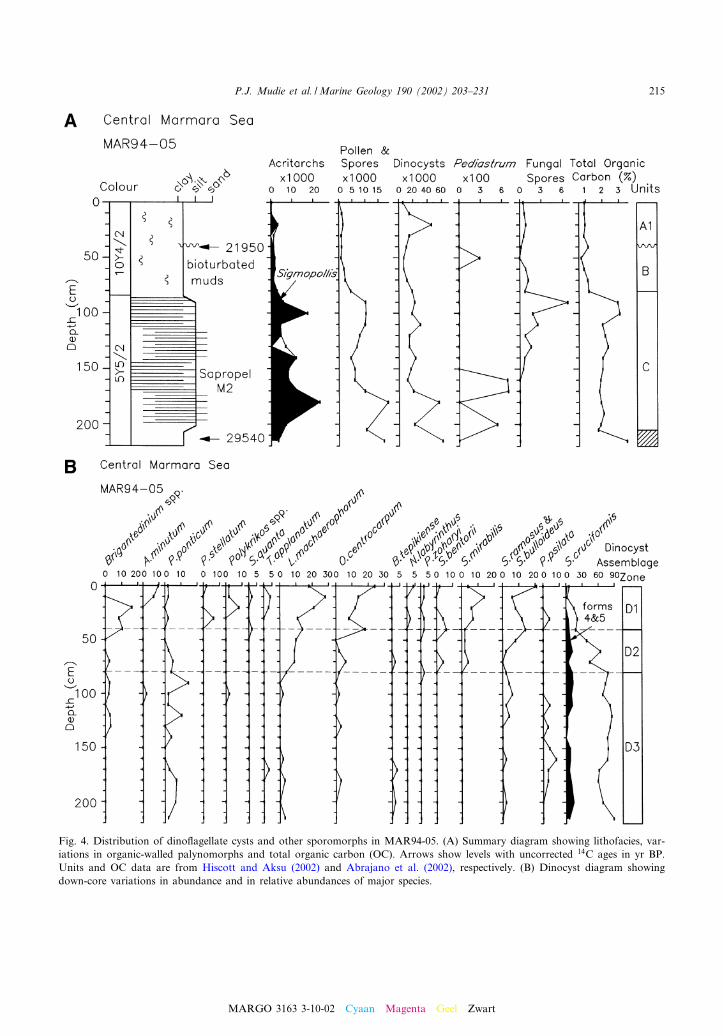
Fig. 4. Distribution of dino£agellate cysts and other sporomorphs in MAR94-05. (A) Summary diagram showing lithofacies, var-iations in organic-walled palynomorphs and total organic carbon (OC). Arrows show levels with uncorrected 14C ages in yr BP.Units and OC data are from Hiscott and Aksu (2002) and Abrajano et al. (2002), respectively. (B) Dinocyst diagram showingdown-core variations in abundance and in relative abundances of major species.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 215
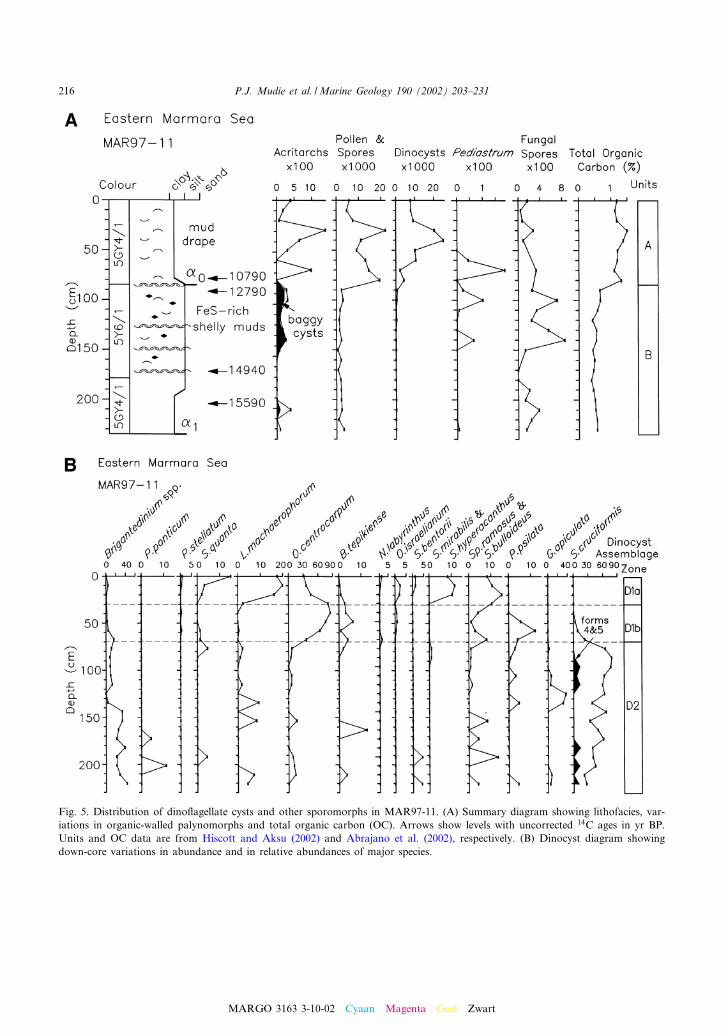
Fig. 5. Distribution of dino£agellate cysts and other sporomorphs in MAR97-11. (A) Summary diagram showing lithofacies, var-iations in organic-walled palynomorphs and total organic carbon (OC). Arrows show levels with uncorrected 14C ages in yr BP.Units and OC data are from Hiscott and Aksu (2002) and Abrajano et al. (2002), respectively. (B) Dinocyst diagram showingdown-core variations in abundance and in relative abundances of major species.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231216

formis (predominantly forms 1 and 2 of Mudie etal., 2001), together with minor amounts of Peri-dinium ponticum, Pyxidinopsis psilata and thefreshwater dino£agellate, Gonyaulax apiculata. InCores MAR98-12, small amounts of Spiniferitesinaequalis are also present: this species has previ-ously only been reported from the low salinityUnit 3 of Core B7 and other Late Quaternarycores from the Black Sea (Wall et al., 1973).The halophilic Mediterranean species Spiniferitesmirabilis, Spiniferites hyperacanthus, Spiniferitesbentorii and the hypersaline species Operculodi-nium israelianum and Polysphaeridium zoharyiare absent, but there are minor amounts of eu-ryhaline species including Spiniferites ramosus,Spiniferites bulloideus, Lingulodinium machaero-phorum and Operculodinium centrocarpum. Bri-gantedinium is also common in the lower interval(LGM s 15 000 yr BP) of the deltaic sequence inCore MAR97-11, despite the relatively low totalorganic carbon (TOC) content. There are alsorare occurrences of cf. Lophocysta cruciformisthat marks the cold, low salinity Pleniglacial-YDunit in Core MAR98-04 (see Fig. 1) of the south-western Black Sea (Rochon, unpublished data;Abrajano et al., 2002).
Pollen and spore concentrations are also low inthe late glacial, but concentrations of acritarchsand fungal spores are high, and there are sporadicpeaks of Pediastrum. In MAR98-12, Sigmopollispsilatum is the only acritarch present, while fungalspores are dominated by an unknown cf. Tetra-ploa sp. with branched processes. Elsewhere, thisunusual palynomorph is only known from theCanadian Arctic channels (Mudie and Rochon,2002).
The dino£agellate cysts in the Late Glacial^Pleniglacial Units C and D (V25 000^33 000 yrBP) of Core MAR94-05 (Fig. 4B) are markedby high concentrations of a low diversity Spinife-rites cruciformis^Pyxidinopsis psilata assemblage,as described in detail by Mudie et al. (2001). Pres-ence of the euryhaline species, Lingulodinium ma-chaerophorum, Operculodinium centrocarpum, andbrackish water morphotypes of S. cruciformis(forms 4 and 5) suggests that although salinitywas very low, the surface water was not entirelyfresh (6 1). The freshwater dinocyst Gonyaulax
apiculata (Kouli et al., 2001) is also not presentin this Pleniglacial unit. Pollen^spore concentra-tions are high in these units, particularly in UnitC that is a sapropel (M2 of Aksu et al., 2002a).
4.3. Black Sea ecostratigraphy
Core B-7 is from 2120 m water depth in thedeep basin of the southeastern Black Sea, about100 km from the location of Core 1474 studied byWall and Dale (1973). Core B-7 contains threelithofacies (Fig. 7A) which correspond to facies1^3 at Site 1474 (see Ross and Degens, 1974;Mudie et al., 2001). Facies I is a highly organic(2.5^5% TOC) coccolith-rich mud containingvarved intervals ; Facies II is a sapropel with 5^18% TOC; and Facies III is a silty mud (0.5%TOC) with occasional carbonate beds containingup to 2.5% TOC (Ross and Degens, 1974; De-gens, 1974). Correlation with dinocyst assemblagezones in the 14C-dated cores studied by Wall andDale (1974) and with regional lake pollen zones(Mudie et al., 2002) show that the zone 1/2 and 2/3boundaries have ages of about 2900 and 7140 yrBP, respectively. The bottom of the core has aprobable age of ca. 9 ka based on land^sea pollencorrelation (Mudie et al., 2001) and supported byan age of 8600 yr BP for a depth of 120 cm inother cores from this basin (Ross and Degens,1974).
In contrast to the Marmara Sea cores, the di-nocyst concentrations in the Holocene sedimentsof the Black Sea (Fig. 7A) show major increasesin the middle Holocene unit II and in sections ofthe early Holocene Unit III. Assemblages in UnitsI and II are dominated by Lingulodinium machaero-phorum, Operculodinium centrocarpum and Spini-ferites spp. (mostly S. mirabilis, S. ramosus and S.bentorii), with variable amounts of Brigantediniumand other protoperidinioid cysts (Fig. 7B). Unit I(0^20 cm) has assemblages (A in Fig. 7B) almostidentical to those of sub-zone D1a in the Mar-mara Sea, being dominated by L. machaeropho-rum and Spiniferites species that are common inthe Mediterranean^Aegean seas (S. mirabilis, S.hyperacanthus, S. bulloideus, S. delicatus and S.bentorii) and a diversity of heterotrophic taxa, in-cluding Brigantedinium spp., Algidasphaeridium?
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 217

Fig. 6. Distribution of dino£agellate cysts and other sporomorphs in MAR98-09. (A) Summary diagram showing lithofacies, var-iations in organic-walled palynomorphs and total organic carbon (OC). Arrows show levels with uncorrected 14C ages in yr BP.Units and OC data are from Hiscott and Aksu (2002) and Abrajano et al. (2002), respectively. (B) Dinocyst diagram showingdown-core variations in abundance and in relative abundances of major species.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231218
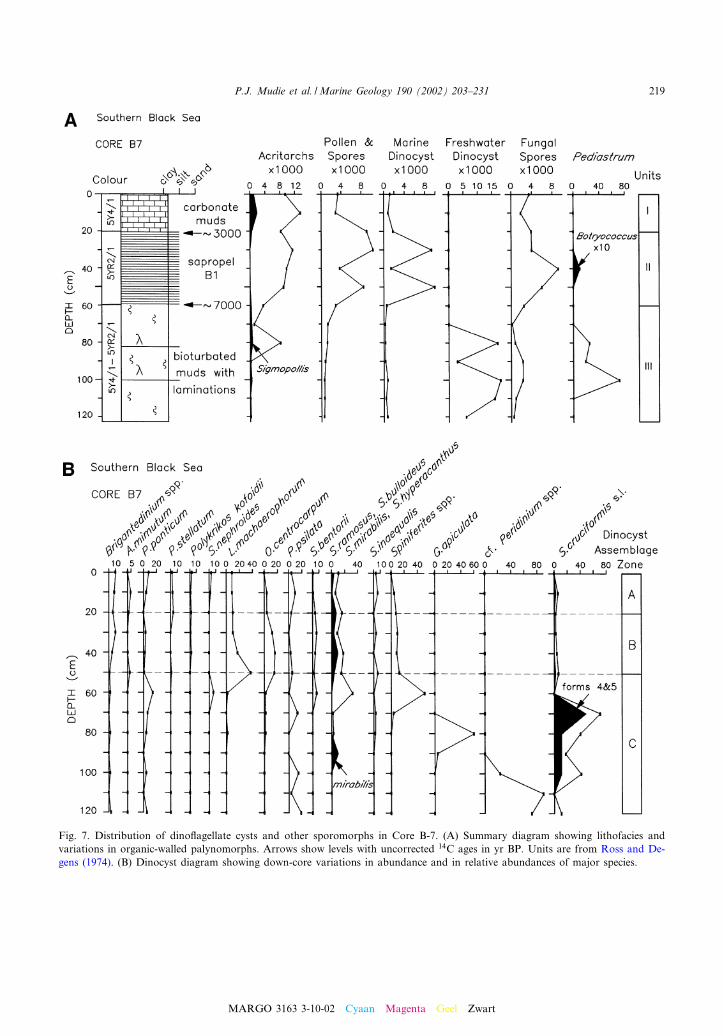
Fig. 7. Distribution of dino£agellate cysts and other sporomorphs in Core B-7. (A) Summary diagram showing lithofacies andvariations in organic-walled palynomorphs. Arrows show levels with uncorrected 14C ages in yr BP. Units are from Ross and De-gens (1974). (B) Dinocyst diagram showing down-core variations in abundance and in relative abundances of major species.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 219

minutum, Polykrikos kofoidii, Selenopemphixquanta, Selenopemphix nephroides and Protoperi-dinium (Stelladinium) stellatum. The main di¡er-ence between the Black and Marmara sea lateHolocene dinocyst assemblages is the presence inthe lower salinity Black Sea of Peridinium ponti-cum, Pyxidinopsis psilata, Spiniferites inaequalisand Spiniferites cruciformis. Furthermore, thereis an absence of common Mediterranean Sea spe-cies (see Aksu et al., 1995a; Mudie et al., 2001and references therein) such as Operculodiniumisraelianum, Bitectatodinium tepikiense and Nema-tosphaeropsis labyrinthus.
Dinocyst assemblages (B in Fig. 7B) in the mid-Holocene sapropelic unit 2 (20^60 cm) of Core B-7 di¡er from the late Holocene assemblages inhaving higher percentages of Operculodinium cen-trocarpum, a lower diversity of organic-walledprotoperidinioid species although abundant cal-careous cysts of Scrippsiella trochoidea are present(e.g. Ross and Degens, 1974). There is also anincrease in Spiniferites spp. with very short pro-cesses. The latter may be morphotypes of Spini-ferites ramosus as found in seen in low salinitywaters of the Baltic Sea (Ellegaard, 2000) buttheir speci¢c identity is presently uncertain.
Dinocyst assemblages (C in Fig. 7B) in theearly Holocene Unit III (60^120 cm) of the BlackSea are almost identical to those of dinocyst zoneB in the Marmara Sea, being distinguished by lowconcentrations dominated (40^80%) by Spinifer-ites cruciformis (including all ¢ve forms of Mudieet al., 2001), together with minor amounts ofPeridinium ponticum, Pyxidinopsis psilata, Spini-ferites inaequalis, Spiniferites ramosus and Spini-ferites mirabilis, and with sporadic occurrences ofLingulodinium machaerophorum, Operculodiniumcentrocarpum and Brigantedinium spp. The maindi¡erence in the Black Sea dinocyst assemblagesare: (1) higher percentages of S. cruciformis forms4 and 5; and (2) greater abundance of the fresh-water dino£agellate, Gonyaulax apiculata (seeKouli et al., 2001), and the ‘baggy dino£agellatecysts’ of Traverse (1974, 1988) which we tenta-tively ascribe to the genus Peridinium on the basisof the similarity of some specimens to the fresh-water taxon Type D cysts described by Norris andMcAndrews (1970).
As in the Marmara Sea cores, pollen^sporeconcentrations in Core B-7 (Fig. 7A) show almostthe same trend as the marine dinocyst concentra-tions, but with the mid-Holocene rise beginningless abruptly and prior to the sapropel deposition.In the Black Sea, the upper pollen concentrationpeak also occurs higher in the late Holocene Unit1. In contrast to the Marmara cores, however,most of the other sporomorph groups in the Ho-locene sediments of the Black Sea show sustainedmid^upper Holocene high abundances. The onlyexception is the restricted occurrence of chloro-coccalean coenobia in the earlier Holocene, witha notable peak in Pediastrum corresponding topeaks in concentrations of fungal spores andfreshwater dinocysts around V9 ka. Fungal re-mains in Units 2 and 3 are also much more abun-dant than in the Marmara cores, being dominatedby various pluricellular taxa, including Tetraploa.Acritarchs in Units 1 and 2 are dominated byCymatiosphaera spp., with occasional occurrencesof cf. Acritarch-8 of Traverse (1978).
5. Interpretation and discussion
5.1. Variations in palynomorph concentrations andassemblage composition as a function of post-depositional oxidation
Before valid interpretations can be made of thedown-core and inter-basin variations in palyno-morph assemblages, it is necessary to examinepossible e¡ects of aerobic decay (oxidative diagen-esis) on the assemblages in the Marmara andBlack sea cores.
The earliest interpretations of changes in BlackSea palynomorph concentrations and assemblagecomposition (e.g. Traverse, 1974; Wall and Dale,1974) did not investigate possible e¡ects of oxida-tive diagenetic overprinting, thereby leaving unan-swered the question of how much assemblages canchange as a function of preservation in sapropelicvs. non-sapropelic units. In contrast, recent workon the palynology of sapropel S1 from the Ban-nock Basin area of the southeastern Mediterra-nean Sea (Cheddadi et al., 1991; Cheddadi andRossignol-Strick, 1995; Zonneveld et al., 2001)
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231220

has shown that pollen^spore concentrations anddinocyst assemblage composition are di¡erentiallya¡ected by changes in the redox potential of thesediments, with all palynomorph groups beingwell represented in anaerobic sapropel layers butwith subsequent di¡erential loss of oxidation-sen-sitive species during post-depositional oxidativediagenesis of the sapropel. Of particular impor-tance is the evidence for aerobic decay of proto-peridinioid dino£agellates in the upper sapropelicunit (Zonneveld et al., 2001) which led to theconclusion that changes in protoperidinioid con-centrations and percentages are not always reli-able indicators of changes in paleoproduction orpaleoceanographic conditions.
In contrast to these southeast Mediterraneanstudies, however, Aksu et al. (1995a) studied thedinocysts, pollen and other acid-resistant particu-late organic matter in cores on a north^southtransect of the Aegean Sea in order to test varioushypotheses about the origin of the early Holocenesapropel S1, particularly the debate regarding itsorigin from increased marine primary productionvs. increased input of TOC during conditions ofhigh freshwater runo¡ and strong strati¢cation.The Aegean Sea cores show maximum dinocystabundances in the late glacial non-sapropelicmud unit (s 9600 yr BP), when low diversity as-semblages were dominated by euryhaline, moder-ately oxidation resistant species of Operculodiniumcentrocarpum and Nematosphaeropsis labyrinthus.S1 deposition (9600^6400 yr BP) is marked by alarge increases in terrigenous sporomorphs andrefractive humic compounds, but only small in-creases in dinocysts, mainly from larger amountsof Brigantedinium simplex and Bitectatodinium te-pikiense which are species associated with cold,relatively low salinity (V30^34) or strongly strati-¢ed waters in the North Atlantic (Rochon et al.,1999).
In these Aegean cores, the sapropel boundariesare marked by peaks in Ba and silt-size pyrite andthey do not show a ‘£oating’ Mn marker abovethe visible sapropel which indicates post-deposi-tional oxidation of the sapropelic layer in theSE Mediterranean core studied by Zonneveld etal. (2001). Furthermore, organic geochemistrystudies of low molecular weight n-alkanes in one
Aegean Sea core showed that the onset of sapro-pel formation was marked by a decline in marineproductivity (Aksu et al., 1999a), while an in-crease of high molecular weight n-alkanes indi-cated higher terrigenous carbon input. In the Ae-gean Sea sapropel S1, therefore, it is clear that thelow abundance of organic-walled dinocysts andhigh pollen^spore concentrations are a primaryfunction of increased runo¡ and terrigenous or-ganic carbon deposition and not the result of ox-idation-related diagenesis of organic matter.
The new data on palynomorph concentrationsand dinocyst composition in sapropels M1, M2and B1 of the Marmara and Black sea cores(see Figs. 3A, 4A and 7A) provide further oppor-tunity to examine the question of how to distin-guish between changes in paleoproductivity andchanges arising from diagenesis of oxidation-sen-sitive palynomorphs. Our results show that in allthe cores, there are increases in pollen^spore con-centrations associated with the organic-rich sap-ropels, but the increases begin prior to and endafter the termination of sapropel deposition. Thesapropel boundaries are sharply delimited, there-fore mixing is not likely to be the cause of theextended pollen^spore maxima. Furthermore,there are large changes in abundance of oxida-tion-sensitive pollen species, e.g. Quercus, withinthe non-sapropelic units (see Mudie et al., 2002).Therefore, it is clear that that amelioration ofclimate conditions is the primary reason for thepollen^spore increases, while the concomitant in-creases in river discharge merely enhance the in-£ux of terrigenous organic matter (including pol-len, spores and humic compounds), during thetime of sapropel deposition (see Aksu et al.,1999a).
Likewise, it is evident that the same patterns ofdown-core changes in total dinocyst concentra-tions occur in the Holocene sediments of the Mar-mara Sea cores regardless of the presence or ab-sence of a mid-Holocene sapropel. As in theAegean Sea Core 20 (Aksu et al., 1999a), thebase of the Holocene sapropel M1 is marked byan increase in oxidation-sensitive Brigantediniumspecies although %TOC is not much higher thanin the underlying mud unit (Fig. 3A). In contrast,the mid-Holocene B1 sapropel, with up to 18%
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 221

TOC (Ross and Degens, 1974), and the glacial-stage M2 sapropel, with up to 3% TOC, showno increases in Brigantedinium or other sensitiveprotoperidinioid species. In all the cores there is adown-core decrease in diversity of oxidation sen-sitive protoperidinioids and Polykrikos spp. re-gardless of the presence or absence of sapropelicunits. Overall, then, for our study area we do notsee any clear evidence of major changes in dino-cyst concentrations or assemblage compositionchanges that can be attributed to oxidative dia-genesis.
5.2. Concentrations of acritarchs, fungal sporesand freshwater sporomorphs
5.2.1. AcritarchsAll previous palynological studies of Holocene
sediments in the low salinity Black Sea havedrawn attention to the abundance of small, simplesphaeromorphic acritarchs, primarily Cymatio-sphaera spp. (Wall and Dale, 1974), cf. Micrhy-stridium spp. (echinate spheres of Acritarch-7 Tra-verse, 1978) and leiosphaerids (wrinkled bag-likeAcritarch-8 of Traverse, 1978). Variable numbersof these taxa are also present in the Marmara Seacores and in Core B-7. In addition, we foundlarge amounts of Sigmopollis spp., primarily S.psilatum, in glacial-stage sections of the MarmaraSea cores, as well as variable lesser amounts in theHolocene sediments.
The biological origin of the acritarchs in ourcores is not known with certainty but somesphaeroidal acritarchs, including Leiosphaeridiaspp., are believed to be prasinophycean green al-gae (Batten, 1996a). Although some modern pra-sinophytes like Halosphaera are restricted to ma-rine environments, very low N
13C values of fossildeposits indicate that other taxa are associatedwith low salinities and temperatures (Guy-Ohlson,1996).
Traverse (1978) assumed that all the acritarchsand dinocysts were indicators of marine water in-£ow to the Black Sea. He therefore added thetotal acritarch and dinocyst concentrations to ob-tain a ‘marine in£uence index’ in which high andlow values apparently corresponded to sea levelhighstands and intervals of periodic isolation
from the Aegean Sea, respectively. Our newcore-top data, however, show that the acritarchsare most abundant in modern sediments beneaththe low salinity waters (V22) of the Black Seaand they are absent from the Aegean Sea. Fur-thermore, as reported in 4.1. Core-top distribu-tions of dinocysts and other palynomorphs, theacritarch Concentricystes rubinus is believed tobe of freshwater or soil origin. Therefore, it isprobably not appropriate to group these fresh/brackish water indicators with well-known marinedino£agellate species in compiling an index of ma-rine in£uence.
It has previously been noted that sphaeromor-phic acritarchs are most abundant and diverse infresh to brackish water estuarine environments ofthe circum-arctic shelves (Solomon et al., 2000;Kunz-Pirrung, 1998), and that high numbers ofSigmopollis psilatum are usually associated withriver discharge (e.g. Mudie, 1992; Head, 1993).In general, then, the down-core increases in acri-tarchs and S. psilatum seen in most of the Mar-mara cores suggest periodic intervals of increasedruno¡, and the pleniglacial interval in CoreMAR94-05 shows a sustained increase in theselow salinity indicators.
5.2.2. Freshwater algaeThe Marmara Sea and Black Sea cores contain
fossilizable remains of two groups of freshwateralgae: coenobia of the colonial Chlorococcalesgenera, Pediastrum and (rarely) Botryococcus,and cysts of the freshwater dino£agellates Go-nyaulax apiculata and Spiniferites cruciformisforms 1^3 of Mudie et al., 2001). It is also likelythat the ‘baggy dino£agellate cysts’ of Traverse(1978) are fresh or brackish water taxa becausethey have not been recorded for marine sedi-ments.
The best known freshwater markers are the colo-nial chlorococcalean algae, Pediastrum boryanumvar. boryanum and Botryococcus cf. B. braunii.Pediastrum boryanum is a stenohaline species as-sociated with a wide range of temperature andtrophic conditions, while Botryococcus is a moresalt-tolerant taxon, sometimes living in brackishwater (Batten, 1996a; Matthiessen and Brenner,1996). Most modern species of Pediastrum live
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231222

in freshwater ponds, lakes and slow-moving riversand their occurrence in marine environments in-dicates £uvial transport (Batten, 1996a; Matthies-sen et al., 2000). In modern sediments of thesouthern Baltic Sea where salinity averages 5.95^7.71, Pediastrum and Botryococcus comprise morethan 75% of the aquatic palynomorphs, with theremainder being the euryhaline dinocyst speciesOperculodinium centrocarpum, a short-spinedform of Spiniferites bulloideus and the brackishwater species Pyxidinopsis psilata (Matthiessenand Brenner, 1996).
At present, Pediastrum boryanum is absent orvery rare (6 50/g) in core-top samples outside theBlack Sea where concentrations range from 5 to72/g. This concentration is much lower than inrecent lake sediments where in£uxes of Pedias-trum coenobia are about 1000^2000 cm2 yr31
(e.g. Burden et al., 1986). The present Black Seaconcentrations are also much lower than in Arcticestuaries where concentrations of coenobia rangefrom 6 500 to 3500/g, suggesting in£ux rates ofthe order of 50^350 cm2 yr31 (Matthiessen et al.,2000). In our cores, there is a general increase inPediastrum concentrations in the late glacial^earlyHolocene intervals of both the Marmara andBlack sea cores (Figs. 3A, 5A and 7A), withpeak abundances of V400^700 coenobia/g occur-ring in during late Pleniglacial to LGM time inCore MAR94-05. However, these chlorococcaleanfreshwater indicators are not consistently abun-dant in any of our cores and their relative abun-dance never exceeds that of the dinocysts.
Gonyaulax apiculata is a fairly common dino-£agellate in European lakes (Kouli et al., 2001).However, cysts of G. apiculata are known onlyfrom modern sediments in Lake Zurich in Swit-zerland and from late glacial^early Holocene sedi-ments of Lake Kastoria in northern Greece (Kou-li et al., 2001). It is therefore believed that highconcentrations of G. apiculata cysts (e.g. 500 000cysts/ml in Lake Kastoria) indicate alpine lacus-trine environments. Cysts of G. apiculata are ab-sent from our core-top samples, the late Holocenesediments of MAR97-2 and the Pleniglacial^LGM sediments of MAR94-5. Very low concen-trations occur sporadically in the late glacial andHolocene sediments of most other Marmara Sea
cores (Figs. 3B, 5B and 6B), suggesting long-dis-tance transport during periods of increased fresh-water runo¡ from rivers draining alpine lakes.The largest concentration (1000^2000 cysts/g) oc-curs as a single peak during the early Holoceneand corresponds to a peak in Pediastrum coeno-bia. This spiky distribution also suggests periodiccatastrophic £ooding from alpine or periglaciallakes rather than the existence of a freshwaterlake in the Black Sea basin.
In Lake Kastoria, abundant morphotypes ofSpiniferites cruciformis with expanded peri-phragms and well-developed spines (forms 1^3of Mudie et al., 2001) co-occur with cysts of Go-nyaulax apiculata. It has therefore been suggestedthat the occurrence of S. cruciformis in the BlackSea may indicate freshwater lake conditions(Kouli et al., 2001). In the Greek lake, there is alate glacial succession from dominance of Botryo-coccus followed by Pediastrum, then abundant S.cruciformis and G. apiculata, together with an in-crease in fungal remains. In contrast, no suchsuccession is observed in the early Holocene ofCore B-7 during the time that it is purported tohave been a freshwater lake (cf. Ryan and Pit-man, 1999). Furthermore, it is notable that inthe Black and Marmara seas, S. cruciformis mor-photypes with reduced or no processes (forms 4and 5 of Mudie et al., 2001) are always present.These morphotypes are not found in Lake Kasto-ria and it is likely that the reduced process lengthre£ects the stress of variable brackish surfacewater conditions in the Black Sea (Mudie et al.,2001).
It is also likely that the ‘baggy dino£agellatecysts’ of Traverse (1978) are fresh or brackishwater taxa because they have not been recordedfor marine sediments. The most common mor-photypes in our samples have a thin, pale browntwo-layered wall, discontinuous paracingulumand a ¢ve-sided archeopyle, resembling some ofthe cysts ascribed to Tectatodinium psilatum byWall et al. (1973), and listed as a brackish^fresh-water taxon (6 7). Other morphotypes closely re-semble Peridinium cinctum cysts and Cyst type Dof Norris and McAndrews (1970) that were ¢rstdescribed from sediments of inland lakes. Thesecysts are not present in our core-top samples and
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 223

they are only common in the early Holocene sedi-ments of the Black Sea where they co-occur withGonyaulax apiculata.
5.2.3. Fungal remainsIn general, relatively little is known about fungi
in marine environments and there is some ques-tion regarding the production of fossilizable re-mains by aquatic marine fungi (e.g. Kohlmeyerand Kohlmeyer, 1979). However, Cross et al.(1966) and Traverse (1974) have argued that thecorrelation between dinocyst and fungal sporeconcentrations in some nearshore environmentsindicates a common origin in nutrient-rich marinewaters. Yet there are no records of extant marinefungi apart from a few taxa that grow on salt-marsh and mangrove plants (Kohlmeyer andKohlmeyer, 1979; Batten, 1996b) and it is gener-ally believed that most fungal spores in marineenvironments are of freshwater or terrigenous ori-gin.
In our core-top samples, from the Aegean andMarmara seas, most of the fungal remains aredark brown unicellular (Lycoperdon- and Tille-tia-type) or dicellular (Puccinellia-type) spores.These are also the only types of fungal sporesfound in the Holocene Unit A of the Marmaracores. In Units I and II of the Black Sea Core B-7, however, there is a greater diversity of fungalremains, including common short, unbranchedhyphal strands and occasional pluricellular spores(e.g. Geoglossum sphagnophilum Type 77A of VanGeel, 1986), in addition to simple sordariaceousspores.
According to Kohlmeyer and Kohlmeyer(1979), only two kinds of smooth-walled aseptatebrown spores are produced by marine fungi andthere are no marine taxa that produce complexbrown spores or septate hyphae. The aseptatebrown spores originate from fungi growing onsaltmarsh grasses or mangrove leaves. Therefore,it is likely that the brown aseptate spores presentin the late Holocene units of all our cores comefrom saltmarshes in the estuaries and deltas ofrivers entering the Marmara and Black seas. Inthe Black Sea, the presence of pluricellular sporesand hyphae in the late Holocene Units I and II ofthe Black Sea suggests greater transport from
freshwater peat deposits where various fungalspore types and fruiting bodies are common(e.g. Van Geel, 1978 and references therein;Head, 1993).
Simple brown ascospores are rare or absent inthe late glacial and Pleniglacial Units B and C ofthe Marmara cores. The most common fungalremains (V50%) in these units are annulate cir-cular spores with psilate or granulate outer surfa-ces which split in half to form saucer-shaped discswith psilate inner walls. These fungal remainsclosely resemble spores of the Exesisporites sp.of Jarzen and Elsik (1986) which characterizethe high water zone of rivers in Zambia. Othercommon fungal remains in these older units in-clude Tetraploa, Dyadosporites, Polyadosporitesand Pluricellaesporites species, most of which arealso common in the high water river zone (Jarzenand Elsik, 1986). Head (1993) notes that Tetra-ploa is saprophytic on grasses, while other speciesare found in soils and coastal ponds.
More work is required to determine the preciserelationship between the fossil fungal remains inour cores and their fungal sources. However, it ishighly likely that all of the fungal remains in thelate Holocene sections of our cores indicate ter-rigenous particulate organic carbon input, includ-ing leafy debris from saltmarshes. The change infungal assemblage composition down-core indi-cates reduction or die-out of saltmarshes and in-creased in£ow of organic material during £oodingof riverbanks and peatmarshes. It is notable thatsimilar assemblages of fungal remains are markersof river £ooding in the modern Mackenzie Delta(Solomon et al., 2000).
5.2.4. Microforaminiferal liningsMicroforaminiferal linings of planktonic and
benthic foraminifera are abundant in the Mar-mara Sea and are well represented in the AegeanSea but are absent in the anoxic Black Sea basins.These ¢ndings are consistent with the distribu-tions of foraminiferal tests (see Aksu et al.,2002a and references therein) for the low oxygenbottom sediments of the Aegean and Marmaraseas and the high benthic productivity of the Mar-mara gateway. However, these workers did notexamine cores from the lower salinity waters of
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231224

the Black Sea so it remains to be shown whetherthe absence of microforaminiferal linings denotesabsence of foraminifera or a lack of preservationof these calcareous and arenaceous microfossils indeep water of the Black Sea.
5.3. Dinocyst assemblages and surface watersalinity and implications for glacial^interglacialpaleoceanography
In the Marmara Sea cores, two Holocene sub-assemblages are recognized. The youngest sub-as-semblage, D1a (0^V7000 yr BP) is dominated byLingulodinium machaerophorum or, near the Ae-gean Sea, by Operculodinium centrocarpum, to-gether with a relatively high percentage of Spini-ferites mirabilis (here grouped with Spiniferiteshyperacanthus), rare occurrences of Operculodi-nium israelianum and/or Bitectatodinium tepikiense(MAR98-12, MAR97-11 and MAR98-9), and adiversity of heterotrophic taxa, including Algida-sphaeridium? minutum, Polykrikos kofoidii, Sele-nopemphix quanta, Selenopemphix nephroides andProtoperidinium (Stelladinium) stellatum. Theearly Holocene (V7000^10 000 yr BP) sub-assem-blage D1b is dominated by O. centrocarpum, withvariable amounts of the low salinity indicatorsPeridinium ponticum, Pyxidinopsis psilata and Spi-niferites cruciformis, absence of Polykrikos and alower diversity of salt-tolerant euryhaline Spinifer-ites and protoperidinioid species.
The dominant species in these assemblages,Operculodinium centrocarpum and Lingulodiniummachaerophorum, are well-known euryhaline spe-cies with a wide range of temperature tolerance(Dale, 1996; Rochon et al., 1999). However,O. centrocarpum appears to be more tolerant oflow surface temperatures (Mudie, 1992; Dale,1996; Rochon et al., 1999), possibly accountingfor its greater dominance in the early Holocenesub-assemblage D1b. Disappearance of the halo-philic Mediterranean species, Spiniferites mirabilisand Operculodinium israelianum, in D1b and re-placement by species associated with low salinities(V3^17 psu) also clearly indicate that more freshwater was £owing into Marmara Sea at this time.
In the Black Sea, the late Holocene assemblagesof Unit I are almost identical to those of sub-zone
D1a in the Marmara Sea. The main di¡erence isthe presence of the low salinity indicator Peridi-nium ponticum, Pyxidinopsis psilata, Spiniferitesinaequalis, Spiniferites cruciformis and the absenceof the halophilic Mediterranean/Atlantic speciesOperculodinium israelianum, Bitectatodinium tepi-kiense and Nematosphaeropsis labyrinthus. The as-semblages in Unit 2 (V3000^6000 yr BP) are vir-tually the same as those in sub-zone D1b of theMarmara Sea cores. Assemblages in the early Ho-locene Unit III of the Black Sea di¡er in beingdominated by S. cruciformis, including both fresh-water forms 1^3 and the brackish water forms 4and 5. The predominance of S. cruciformis clearlyindicates that the surface salinity in the Black Seawas lower than present during the early Holocene.However, the presence of the brackish waterforms of S. cruciformis and other species notknown from modern freshwater environments(P. ponticum, S. inaequalis, Spiniferites ramosus,Spiniferites mirabilis, Lingulodinium machaeropho-rum, Operculodinium centrocarpum and Brigante-dinium spp.) makes it unlikely that the Black Seawas a freshwater lake.
In the Marmara Sea, the LGM^late glacial di-nocyst assemblages B and C are also dominatedby Spiniferites cruciformis and other ‘Black Sea’species and they are similar in composition to thelow salinity early Holocene assemblages in UnitIII. It is therefore clear that although the glacialstages were marked by much lower sea-surfacetemperatures, there was either some periodic ma-rine in£uence or the marine dinocysts were livingin a brackish water environment with the fresh-water species being transported from glacial lakes.The latter scenario may explain the relative spar-sity of chlorococcaleans and the relatively lowconcentrations of the freshwater dinocysts com-pared to fungal remains.
5.4. Implications of Aegean Sea^Marmara Sea^Black Sea linkages
The framework for the history of salinitychanges in the Marmara gateway begins in thelate Miocene^early Pliocene when the Mediterra-nean, Black and Caspian seas were linked to forma large marine basin, known as the Paratethys
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 225

(Ross, 1978; Schrader, 1978). During the Plio-cene, the Black Sea became semi-isolated, havinga restricted connection with the MediterraneanSea through a network of straits and inlandseas. Microfossils from DSDP Leg 42 (Schrader,1978; Gheorghian, 1978) suggest that the Plio^Pleistocene Black Sea was a low salinity (6 5)to brackish (5^15) lake, with intervals of strongermarine in£uence (15^20) during sea level high-stands in the youngest marine isotopic stages(MIS) 9, 7, 5 and 1.
Recently, however, some studies have postu-lated that after the last glacial maximumV20 000 yr BP, the Black Sea became a fresh-water lake and that from V12 500 to 7000 yrBP, it was isolated from the Marmara Sea(Ryan and Pitman, 1999). During this time oflowered water level, it has been suggested thatthe Black Sea contained potable water which en-couraged the settlement of humans and their live-stock, and which led to farming and crop irriga-tion on the emergent deltas (e.g. Ryan andPitman, 1999, pp. 230^233). However, for drink-ing water and sustained agricultural usage, watersalinity must be 6 1 (see Mudie, 1974). We havealso shown that marine or brackish water-tolerantdino£agellates persist in most sections of ourcores and that there is no palynological evidencefor early forest clearance or agriculture in thesouthern Black Sea (Mudie et al., 2002). Wetherefore consider it necessary to examine the pro-cesses controlling salination and desalination ofthe Black Sea in order to evaluate the likelihoodof reaching truly freshwater (i.e. 6 1) lake condi-tions in the time-frame of the Late Quaternaryglacial lowstands. If desalination during theselowstands is impossible, then the postulated earlyagriculture must have depended on ‘the constant,if sluggish £ow of water’ in the valleys and deltasof the reduced rivers which Ryan and Pitman(1999, pp. 230^231) described for the dry YDand 8-ka European cold intervals.
At present, the Straits of Dardanelles and Bos-phorus, and the intervening land-locked MarmaraSea constitute the gateway linking the Mediterra-nean and Black seas and controlling the £ux ofsaline water to and from the Black Sea. However,the Strait of Bosphorus may have formed only
during the last V100 000 years (Oktay et al.,2002) and before this, the Black and Marmaraseas were connected via the Sakarya Bosphorusstrait (Pfannenstiel, 1944) in the lower reach ofthe present Sakarya River and the Bolay|r Chan-nel linked the Marmara and Aegean seas. Duringthe Quaternary, there were periods of completeisolation of the Black Sea from the MediterraneanSea during glacial maxima when the straits weresubaerially exposed because of glacio-eustatic var-iations in global sea level and local tectonic fac-tors. These glacial-stage intervals of isolation al-ternated with interglacial periods of connectionand water exchange between the two seas.
The ¢rst major post-Pliocene incursion of salineMediterranean waters into the Black Sea occurredat ca. 300 000 yr BP (Schrader, 1978; Ross, 1978)when three indigenous coccolith species appeared(Percival, 1978), concurrently with in£uxes of ma-rine diatoms and silico£agellates (Schrader, 1978).This event marks the start of water exchange be-tween the Black and Marmara seas (initiallythrough the Sakarya Bosphorus channel) andcommencement of the Quaternary salination^de-salination cycles. During this early £ooding inter-val, surface salinity values exceeded 12 (Schrader,1978). By the mid^late Quaternary, benthic fora-minifera were dominated by brackish water taxa,with sporadic occurrences of euryhaline and sten-ohaline species (Gheorghian, 1978). The longtime-frame of these micropaleontological changessuggests that replacement of the brackish watersin deep Black Sea basins with saline Mediterra-nean waters took many thousands of years, sim-ilar to the processes modelled for the last glacial^Holocene transition (e.g. Boudreau and Leblond,1989).
The salinity history of Black Sea for the last300 000 years is controlled primarily by the depthsof the connecting channels and global sea level,but it is also further modulated by climate, thevolume of water exchange and the rate of upwardmixing of saltwater within the Black Sea. Theclimate controls the amount of freshwater £uvialinput to the Black Sea basin as well as the balancebetween precipitation and evaporation (P^E),whereas the water exchange via the Marmaragateway controls the amount of salt water input.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231226

Of critical importance is the fact that a salinelayer of Mediterranean water only penetratesinto the Black Sea (here called a salination phase)when thick enough to ¢ll the bottom of the en-trance strait. Any thinner developing salt wedgewill be entrained by the out£owing surface cur-rent, with salt returning to the Marmara Sea, aspart of the desalination phase. Further desalina-tion of the Black Sea requires overturn of therelict Mediterranean bottom water and its remov-al by out£ow through the Marmara Sea. Atpresent, this process is poorly understood (Oº zsoyet al., 1996) but in the deep basins, it is partlydriven by the geothermal gradient which is as-sumed to have been constant during the past300 000 years. Two scenarios will examine the ex-tremes of a range of possible Black Sea salinitycycle models, as outlined below.
5.4.1. Scenario 1: Black Sea LGM surface watersalinity was higher than today
Seismic re£ection and sediment core data fromthe Black Sea shelves show that during the tran-sition from the LGM to the Holocene, the BlackSea water level was V110 m below present (Aksuet al., 2002a,b). The sea level started to fall be-tween MIS 3 and 2, at which time the Black Seawas isolated from the Marmara Sea. When thelevel of the Black Sea fell below the breach depthof Bosphorus Strait (V340 m), desalination ofthe Black Sea ceased because there was no out-£ow across the Bosphorus; therefore the BlackSea salinity must have progressively increased asthe water level dropped to its minimum,V20 000 yr BP. Once isolated from global sealevel, the continued water level lowering from340 to 3110 m must have been caused by a re-duction in freshwater in£ow into the basin duringa period of PIE. The climate in central Europeremained very cold and dry from the peak of theLGM at V18 ka until approximately 9500 yr BP(e.g. Harrison et al., 1993, 1996) and the dis-charges of the largest rivers with headwaters incentral-eastern Europe must have been substan-tially reduced. Hence, although pollen evidence(Prentice et al., 1992; Mudie et al., 2002) indicatesthat P:E remained relatively high in northernAnatolia and around the Marmara Sea, the total
water balance for the Black Sea must have beentilted toward P6E in order to maintain a low-ered sea level in the Black Sea basin. This scenariorequires that salinity would increase in the iso-lated inland sea, and is in total disagreementwith the freshwater conditions suggested byRyan and Pitman (1999, pp. 230^231) for thistime interval. The closest approximation to the‘freshwater lake’ scenario would be a lake withlow salinity surface water and saline bottomwater, surrounded by extensive salt £ats. Thesecircumstances would provide neither potablelake water for humans nor favorable pastoralconditions (see Mudie, 1974) and they are notcompatible with the algal and fungal evidencefor high £uvial runo¡ presented in this paper.
5.4.2. Scenario 2: Black Sea LGM surface watersalinity was lower than today
Desalination of the Black Sea can take placeonly if the level of the Black Sea is above thatof the Strait of Bosphorus but the water is notdeep enough to allow full penetration of the Med-iterranean salt wedge along the length of the Bos-phorus Strait (e.g. Scholten, 1974). Under theseconditions the Black Sea would spill over intothe Marmara Sea, and through time would pro-gressively deplete the salt reservoir in the BlackSea basin. The time required to achieve suchfreshening can be estimated if the freshwater inputto the Black Sea basin and the mixing of thewater masses are known. The present volume ofthe Black Sea is V534 000 km3 of which 650 km3
is from freshwater runo¡. The average salinities ofthe surface and bottom water masses are 18 and22, respectively (Oº zsoy et al., 1991), and the ra-diocarbon age of the bottom water mass isV2000 yr BP. For our minimum salinity scenar-io, we shall set the level of the Black Sea at theend of MIS 3 (V34 000 yr BP), at the breachdepth of Bosphorus Strait, and maintain thefreshwater out£ow during the European colddry period at V10% of its present value (i.e.65 km3). With no penetration of saline Mediter-ranean water, it would require V13 000, V25 000and V53 000 years for the surface salinity valuesto reach 10, 5 and 1, respectively. Thus the lowestsalinity expected at V9000 yr BP would be closer
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 227

to 5 than to the 6 1 required for potable waterand sustained agriculture. If there was a largethrough£ow of glacial meltwater from V20 to15 ka, as postulated by Ryan and Pitman(1999), there may have been a 5000-yr intervalof somewhat more rapid desalination. However,with a colder, thicker layer of surface water, mix-ing of the fresh surface and saline bottom waterlayers is more likely to have been slower, notfaster than the present day because of the reducedsalt gradient within the isolated basin.
It is clear from these models that the desalina-tion of the Black Sea, although not impossible,requires a considerably longer period of timethan assumed by Ryan and Pitman (1999), in or-der to reach the level of potable surface water thatcould be used for irrigation. Because the level ofthe Black Sea dropped during the last glacial,reaching its lowstand of 3110 m during theLGM, the salinity of the Black Sea basin actuallyprobably increased slightly during this period.This leaves the only source of potable and irriga-tion water as the rivers discharging into theslightly brackish lake. It seems that it wouldalso have been necessary to desalinate the emer-gent delta and terrace soils, which is a dauntingtask even in modern times. Therefore, it is ques-tionable that the early farmers would have settledon the shores of the Black Sea. Furthermore, ifthere was an early settlement, then attempts atgrowing crops and fruit trees would have beenlimited to the most salt-tolerant species, e.g. bar-ley, beets and pomegranates. This scenario of alow salinity/brackish water lake with very little orno early agriculture is the one most compatiblewith the Black Sea pollen data of Mudie et al.(2002) and with the micro£oral evidence pre-sented in this report.
6. Conclusions
In the epicontinental seas adjoining the easternMediterranean, pollen and dinocyst assemblagesdo not show any clear evidence of major changesin concentration or assemblage composition thatcan be attributed to aerobic decay. Pollen^sporeassemblages also show no evidence of aerobic de-
cay in the Marmara gateway cores. This ¢ndingallows us to interpret down-core assemblagechanges in terms of £uctuations in paleoproduc-tion and surface water conditions, primarily asrelated to changes in freshwater runo¡ and cli-mate (P^E).
Modern sediment samples from transects in theAegean and Marmara seas and parts of the BlackSea show that the concentrations of dinocysts,acritarchs, chlorococcalean algae, pollen and fun-gi generally decrease with distance from the lowsalinity surface water of the Black Sea. Microfo-raminiferal linings of planktonic and benthic fo-raminifera show the opposite trend and they areabsent in the southeastern Black Sea basin. Thedinocyst species Impagidinium aculeatum, Opercu-lodinium israelianum, Polysphaeridium zoharyi arepresently restricted to the Mediterranean and Ae-gean seas and indicate summer salinities s 24. Incontrast, Spiniferites cruciformis, Spiniferites inae-qualis, Peridinium ponticum and Polykrikos spp.(mostly P. kofoidii) are found only in the low sa-linity Black and Marmara seas. Quinquecuspisconcreta is only present in the intermediate salin-ities of the Marmara Sea.
Late Quaternary dinocyst assemblages in theMarmara Sea show a succession from relativelylow diversity Pleniglacial and late glacial assem-blages dominated by Spiniferites cruciformis andother ‘Black Sea’ species associated with salinitiesof V3^18 to late Holocene assemblages similar tothose presently found in the Black Sea. Early Ho-locene assemblages in the Black Sea indicate low-er surface salinity than present but there is noclear evidence that the surface salinity was everas low as a freshwater lake.
Fungal remains (mostly spores and hyphalstrands) are useful markers of terrigenous sedi-ment in£ux and transport by large rivers. Thesesporomorphs, together with freshwater dinocystsand Pediastrum coenobia, clearly show that incontrast to the Ryan and Pitman model, the earlyHolocene Black Sea received a large volume ofriver runo¡ and was undoubtedly over£owinginto the Marmara Sea during the early Holocene.
Of two possible desalination models examinedfor the late glacial^early Holocene interval in theBlack Sea, only Scenario 2 provides surface water
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231228

conditions compatible with our data and trendingtowards the freshwater conditions postulated byRyan and Pitman (1999). This model requires asustained (V25 000 year) out£ow of fresh waterfrom the Black Sea during the late glacial andearly postglacial in order to achieve surface salin-ity as low as 5. The model also indicates that theisolated Black Sea would not have totally freshsurface water suitable for agricultural settlement.
Acknowledgements
We thank Prof. Dr. Erol Izdar, the Director ofthe Piri Reis Foundation for Maritime and Ma-rine Resources Development and Education, andProf. Dr. Orhan Uslu, the Director of the Insti-tute of Marine Sciences and Technology, for theirsupport and encouragement. Special thanks go tothe o⁄cers and crew of the RV Koca Piri Reis fortheir assistance in data collection, in particularCaptains Mehmet Oº zsayg|l| (MAR97 andMAR98) and the Chief Engineer Oº mer CMubuk,and we are grateful to Hu«seyin Avni Benli formaking available unpublished phytoplanktondata. We acknowledge research and ship timefunds from the Natural Sciences and EngineeringResearch Council of Canada, travel funds fromthe Dean of Science, Memorial University ofNewfoundland, a grant from the Piri Reis Foun-dation for Maritime and Marine Resources De-velopment and Education, and funding of Project920063 by the Geological Survey of Canada. Thispaper was also improved by the helpful reviews ofDrs. F. Marret (Cardi¡ University) and Jens Mat-thiessen (Alfred Wegener Institute, Bremerhaven).
References
Abrajano, T., Aksu, A.E., Hiscott, R.N. and Mudie, P.J.,2002. Aspects of carbon isotope biogeochemistry of LateQuaternary sediments from the Marmara Sea and BlackSea. Mar. Geol. 190, S0025-3227(02)00346-8.
Aksu, A.E., YasMar, D., Mudie, P.J., Gillespie, H., 1995a. Lateglacial^Holocene paleoclimatological and paleoceanographicevolution of the Aegean Sea: micropaleontological and sta-ble isotopic evidence. Mar. Micropaleontol. 25, 1^28.
Aksu, A.E., YasMar, D., Mudie, P.J., 1995b. Paleoclimatic and
paleoceanographic circumstances leading to the developmentof sapropel layer S1 in the Aegean Sea Basins. Palaeogeogr.Palaeoclimatol. Palaeoecol. 116, 71^101.
Aksu, A.E., Abrajano, J., Mudie, P.J., YasMar, D., 1999a. Or-ganic geochemical and palynological evidence for terrige-nous origin of the organic matter in Aegean Sea sapropelS1. Mar. Geol. 153, 303^318.
Aksu, A.E., Hiscott, R.N., YasMar, D., 1999b. Oscillating Qua-ternary water levels of the Marmara Sea and vigorous out-£ow into the Aegean Sea from the Marmara Sea-Black Seadrainage corridor. Mar. Geol. 153, 275^302.
Aksu, A.E., Hiscott, R.N., Kaminski, M.A., Mudie, P.J., Gil-lespie, H., Abrajano, T., YasMar, D., 2002a. Last glacial^Ho-locene paleoceanography of the Black Sea and MarmaraSea: stable isotopic, foraminiferal and coccolith evidence.Mar. Geol. 190, S0025-3227(02)00345-6.
Aksu, A.E., Hiscott, R.N., YasMar, D., IsMler, F.I., Marsh, S.,2002b. Seismic stratigraphy of Late Quaternary depositsfrom the southwestern Black Sea shelf: evidence for non-catastrophic variations in sea-level during the last 10,000years. Mar. Geol. 190, S0025-3227(02)00343-2.
Aubert, M., Revillon, P., Aubert, J., Leger, G., Drai, C., Ar-noux, A., Diana, C. et al., 1990. Mers d’Europe: EŁ tudeshydrobiologiques, chimiques et biologiques. Transfert depolluants entre la Mer Noire, la Mer de Marmara et laMer Ege¤e. Campagne Oce¤anographique du C.E.R.B.O.M.(Nice). Revue Internationale d’Oceanographie Me¤dicaleTome 100.
Batten, D.J., 1996a. Green and blue-green algae. 7C. ColonialChlorococcales. In: Jansonius, J., McGregor, D.C. (Eds.),Palynology: Principles and Applications, Volume 3. Ameri-can Association of Stratigraphic Palynologists, Salt LakeCity, UT, pp. 191^203.
Batten, D.J., 1996b. Palynofacies and palaeoenvironmental in-terpretation. In: Jansonius, J., McGregor, D.C. (Eds.), Paly-nology: Principles and Applications, Volume 3. AmericanAssociation of Stratigraphic Palynologists, Salt Lake City,UT, pp. 1011^1064.
Benli, H.A., 1987. Seasonal Distribution of Plankton in South-ern Black Sea and the E¡ects of Particulate Matter in Set-tling. Unpublished PhD Thesis, Dokuz Eylu«l University ^Institute of Marine Sciences and Technology, 86 pp. (inTurkish).
BesMiktepe, S., Sur, H.I., Oº zsoy, E., Latif, M.A., Og›uz, T.,Uº nlu«ata, Uº ., 1994. The circulation and hydrography of theMarmara Sea. Prog. Oceanogr. 34, 285^334.
Bodeneau, N., 1993. Microalgal blooms in the Romanian areaof the Black Sea and contemporary eutrophication condi-tions. In: Smayda, J., Shimizu, Y. (Eds.), Toxic Phytoplank-ton Blooms in the Sea. Elsevier, Amsterdam, pp. 203^209.
Bologa, A.S., Bodeanu, N., Petran, A., Tiganus, V., Zaitsev,Y.P., 1995. Major modi¢cations of the Black Sea benthicand planktonic biota in the last three decades. Bull. Inst.Oce¤anogr. Monaco Spec. No. 15, 85^110.
Boudreau, B.P., Leblond, P.H., 1989. A simple evolutionarymodel for water and salt in the Black Sea. Paleoceanography4, 157^166.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 229

Bradford, M.R., Wall, D.A., 1984. The distribution of Recentorganic-walled dino£agellate cysts in the Persian Gulf, Gulfof Oman and northwest Arabian Sea. PalaeontographicaAbt. B 192, 16^84.
Burden, E.T., McAndrews, J.H., Norris, G., 1986. Palynologyof Indian and European forest clearance and farming in lakesediment cores from Awenda Provincial Park, Ontario. Can.J. Earth Sci. 23, 43^54.
Cheddadi, R., Rossignol-Strick, M., 1995. Improved preserva-tion of organic matter and pollen in eastern Mediterraneansapropels. Paleoceanography 10, 301^309.
Cheddadi, R., Rossignol-Strick, M., Fontugne, M., 1991. East-ern Mediterranean paleoclimates from 26 to 5 ka B.P. docu-mented by pollen and isotopic analysis of a core in theanoxic Bannock Basin. Mar. Geol. 100, 53^66.
Cross, A.T., Thompson, G.G., Zaitze¡, J.P., 1966. Source anddistribution of palynomorphs in bottom sediments, southernpart of Gulf of California. Mar. Geol. 4, 467^524.
Dale, B., 1996. Dino£agellate cyst ecology: modelling and geo-logical applications. In: Jansonius, J., McGregor, D.C.(Eds.), Palynology: Principles and Applications, Volume 3.American Association of Stratigraphic Palynologists, SaltLake City, UT, pp. 1249^1276.
Degens, E.T., 1974. Cellular processes in Black Sea sediments.In: Degens, E.T., Ross, D.A. (Eds.), The Black Sea: Geol-ogy, Chemistry and Biology. Am. Assoc. Pet. Geol. Mem.20, 183^199.
Duman, M., 1992. Late Quaternary Geological Evolution ofthe Southern Black Sea Basin. Unpublished PhD Thesis,Dokuz Eylu« l University ^ Institute of Marine Sciences andTechnology, 101 pp. (in Turkish).
Ellegaard, M., 2000. Variations in dino£agellate cyst morphol-ogy under conditions of changing salinity during the last2000 years in the Limfjord, Denmark. Rev. Palaeobot. Paly-nol. 109, 65^81.
Gheorghian, M., 1978. Micropaleontological investigations ofsediments from Sites 379, 380 and 381 of Leg 42B. In: Ross,D.A., Neprochnov, Y.P. et al. (Eds.), Init. Rep. DSDP 42,783^787.
Guy-Ohlson, D., 1996. Green and Blue-green algae. 7B - Pra-sinophycean algae. In: Jansonius, J., McGregor, D.C.(Eds.), Palynology: Principles and Applications, Volume 3.American Association of Stratigraphic Palynologists, SaltLake City, UT, pp. 181^188.
Harrison, S.P., Prentice, I.C., Guiot, J., 1993. Climatic con-trols of Holocene lake-level changes in Europe. Clim. Dyn.8, 189^200.
Harrison, S.P., Yu, G., Tarasov, P., 1996. Late Quaternarylake-level record from northern Eurasia. Quat. Res. 45,138^159.
Head, M.J., 1993. Dino£agellates, sporomorphs, and otherpalynomorphs from the upper Pliocene St. Erth beds ofCornwall, Southwestern England. J. Paleontol. 67, 1^62.
Hiscott, R.N., Aksu, A.E., 2002. Late Quaternary history ofthe Marmara Sea and Black Sea from high-resolution seis-mic and gravity-core studies.
Jarzen, D.M., Elsik, W.C., 1986. Fungal palynomorphs recov-
ered from recent river deposits, Luangwa Valley, Zambia.Palynology 10, 35^60.
Kouli, K., Brinkhuis, H., Dale, B., 2001. Spiniferites crucifor-mis : a fresh water dino£agellate cyst? Rev. Palaeobot. Paly-nol. 113, 273^286.
Kohlmeyer, J., Kohlmeyer, E., 1979. Marine Mycology. Aca-demic Press, New York, 690 pp.
Kunz-Pirrung, M., 1998. Rekonstruktion der Ober£achenwas-sermassen der ostliche Laptevsee im Holoza«n anhand vonaquatischen Palynomorphen. Ber. Polarforsch. 281, 1^117.
Matthiessen, J., Brenner, W., 1996. Chlorococcalalgen undDino£agellaten-Zysten in rezenten Sedimenten des Greif-walder Bodens (suu«dliche Ostsee). Senckenb. Mar. 27, 33^48.
Matthiessen, J., Kunz-Pirrig, M., Mudie, P.J., 2000. Fresh-water chlorophycean algae in recent marine sediments ofthe Beaufort, Laptev and Kara Seas (Arctic Ocean). Int.J. Earth Sci. 89, 470^485.
Mudie, P.J., 1974. The potential economic uses of halophytes.In: Reimold, R.J., Queen, W.H. (Eds.), Ecology of Halo-phytes. Academic Press, New York, pp. 565^598.
Mudie, P.J., 1992. Circum-arctic Quaternary and Neogene ma-rine palyno£oras: paleoecolgy and statistical analysis. In:Head, M.J., Wrenn, J.H. (Eds.), Neogene and QuaternaryDino£agellate Cysts and Acritarchs. American Associationof Stratigraphic Palynology Foundation, College Station,TX, pp. 347^390.
Mudie, P.J., Rochon, A., 2002. The distribution of marinedino£agellate cysts in the Canadian Arctic. J. Quat. Sci.16, 603^620.
Mudie, P.J., Aksu, A.E., YasMar, D., 2001. Late Quaternarydinocysts from the Black, Marmara and Aegean Seas: var-iations in assemblages, morphology and paleosalinity. Mar.Micropaleontol. 43, 155^178.
Mudie, P.J., Rochon, A., Aksu, A.E., 2002. Pollen stratigra-phy of Late Quaternary cores from Marmara Sea: land^seacorrelation and paleoclimatic history. Mar. Geol. 190,S0025-3227(02)00349-3.
Norris, G., McAndrews, J.H., 1970. Dino£agellate cysts frompost-glacial lake muds, Minnesota (USA). Rev. Palaeobot.Palynol. 10, 131^156.
Og›uz, T., Latun, V.S., Latif, M.A., Vladimirov, V.V., Sur,H.I., Markov, A.A., Oº zsoy, E., Kotovshchikov, B.B., Ere-meev, V.V., Uº nlu«ata, Uº ., 1993. Circulation in the surfaceand intermediate layers of the Black Sea. Deep-Sea Res.40, 1597^1612.
Oktay, F.Y., Go«kasMan, E., Sak|ncM, M., Yalt|rak, C., Imren, C.,Demirbag›, E., 2002. The e¡ects of the North AnatolianFault Zone to the latest connection between Black Seaand Sea of Marmara. Mar. Geol. 190, S0025-3227(01)00246-8.
Oº zsoy, E., Top, Z., White, G., Murray, J.W., 1991. Doubledi¡usive intrusions, mixing and deep sea convection process-es in the Black Sea. In: Izdar, E., Murray, J.W. (Eds.),Black Sea Oceanography. NATO ASI Ser. C Math. Phys.Sci. 351, 17^42.
Oº zsoy, E., Latif, M.A., Tug›rul, S., Uº nlu«ata, Uº ., 1995. Ex-
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231230

changes with the Mediterranean £uxes, and boundary mix-ing processes in the Black Sea. Bull. Inst. Oce¤anogr. Mon-aco Spec. Vol. 15, 1^26.
Oº zsoy, E., Latif, M.A., Sur, H.I., Goryachkin, Y., 1996. Areview of the exchange £ow regimes and mixing in the Bos-phorus Strait. In: Briand, F. (Ed.), Mediterranean TributarySeas. Bull. Inst. Oce¤anogr. Monaco Spec. No. 17, CIESMESci. Ser. 2, pp. 187^204.
Percival, S.F., 1978. Indigenous and reworked coccoliths fromthe Black Sea. In: Ross, D.A., Neprochnov, Y.P. et al.(Eds.), Init. Rep. DSDP 42, 773^781.
Pfannenstiel, M., 1944. Diluviale Geologie des Mittelmeerge-bietes, die Diluvialen Entwicklundgstadien und die Urge-schichte von Dardanellen, Marmara Meer und Bosphorus.Geol. Rundsch. 34, 334^342.
Polat, CM., Tug›rul, S., 1996. Chemical exchange between theMediterranean and Black Sea via the Turkish Straits. In:Briand, F. (Ed.), Dynamics of Mediterranean Straits andChannels. Bull. Inst. Oce¤anogr. Monaco Spec. No. 17,CIESME Sci. Ser. 2, pp. 167^186.
Prentice, I.C., Guiot, J., Harrison, S.P., 1992. Mediterraneanvegetation, lake-level and paleoclimate at the Last GlacialMaximum. Nature 360, 658^670.
Rochon, A., de Vernal, A., Turon, J.L., Matthiessen, J., Head,M.J., 1999. Distribution of recent dino£agellate cysts in sur-face sediments from the North Atantic Ocean and adjacentseas in relation to sea-surface parameters. Contributions Se-ries No. 35, American Association of Stratigraphic Palynol-ogists Foundation, College Station, TX, 152 pp.
Roman, S., 1974. Palynoplanktologic analysis of some BlackSea cores. In: Degens, E.T., Ross, D.A. (Eds.), The BlackSea: Geology, Chemistry, and Biology. Am. Assoc. Pet.Geol. Mem. 20, 396^410.
Ross, D.A., 1978. Summary of results of Black Sea drilling.In: Ross, D.A., Neprochnov, Y.P. et al. (Eds.), Init. Rep.DSDP 42, 1149^1178.
Ross, D.A., Degens, E.T., 1974. Recent sediments of the BlackSea. In: Degens, E.T., Ross, D.A. (Eds.), The Black Sea:Geology, Chemistry, and Biology. Am. Assoc. Pet. Geol.Mem. 20, 183^199.
Rossignol, M., 1961. Analyse pollinique de se¤diments marinsquaternaires en Israel. II. Se¤diments pleistocenes. PollenSpores IV, 131^136.
Ryan, W.B.F., Pitman, III, W.C., 1999. Noah’s Flood: TheNew Scienti¢c Discoveries about Events that Changed His-tory. Touchstone Book, Simion and Schuster, New York,319 pp.
Ryan, W.B.F., Pittman, W.C., III, Major, C.O., Shimkus, K.,Maskalenko, V., Jones, G.A., Dimitrov, P., Go«ru«r, N.,Sak|ncM, M., Yu«ce, H., 1997. An abrupt drowning of theBlack Sea shelf. Mar. Geol. 138, 119^126.
Scholten, R., 1974. Role of the Bosphorus in Black Sea chem-istry and sedimentation. In: Degens, E.T., Ross, D.A.
(Eds.), The Black Sea: Geology, Chemistry, and Biology.Am. Assoc. Pet. Geol. Mem. 20, 115^126.
Schrader, H.J., 1978. Quaternary through Neogene history ofthe Black Sea, deduced from the paleoecology of diatoms,silico£agellates, ebridians, and chrysomonads. In: Ross,D.A., Neprochnov, Y.P. et al. (Eds.), Init. Rep. DSDP 42,789^901.
Solomon, S., Mudie, P.J., Cranston, R., Hamilton, T., Thibau-deau, S.A., Collins, E.S., 2000. Characterisation of marineand lacustrine sediments in a drowned thermokarst embay-ment, Richards Island, Beaufort Sea, Canada. Int. J. EarthSci. 89, 503^521.
Thunell, R.C., Williams, D.F., 1989. Glacial^Holocene salinitychanges in the Mediterranean Sea: hydrographic and depo-sitional e¡ects. Nature 338, 493^496.
Traverse, A., 1974. Palynological investigation of two BlackSea cores. In: Degens, E.T., Ross, D.A. (Eds.), The BlackSea: Geology, Chemistry, and Biology. Am. Assoc. Pet.Geol. Mem. 20, 381^388.
Traverse, A., 1978. Palynological analysis of DSDP Leg 42B(1975) cores from the Black Sea. In: Ross, D.A. et al. (Eds.),Init. Rep. DSDP 42, 993^1015.
Traverse, A., 1988. Paleopalynology. Unwin Hyman, Boston,MA, 600 pp.
Van Geel, B., 1978. A palaeoecological study of Holocene peatbog sections in Germany and the Netherlands. Rev. Palaeo-bot. Palynol. 25, 1^20.
Vergnaud-Grazzini, C., Ryan, W.B.F., Cita, M.B., 1977. Sta-ble isotopic fractionation, climate change and episodic stag-nation in the eastern Mediterranean during the Late Quater-nary. Mar. Micropaleontol. 2, 353^370.
Wall, D., Dale, B., 1973. Paleosalinity relationships of dino-£agellates in the late Quaternary of the Black Sea ^ a sum-mary. Geosci. Man VII, 95^102.
Wall, D., Dale, B., 1974. Dino£agellates in Late Quaternarydeep-water sediments of Black Sea. In: Degens, E.T., Ross,D.A. (Eds.), The Black Sea: Geology, Chemistry, and Biol-ogy. Am. Assoc. Pet. Geol. Mem. 20, 364^380.
Wall, D., Dale, B., Harada, K., 1973. Descriptions of newfossil dino£agellates from the Late Quaternary of the BlackSea. Micropaleontology 19, 18^31.
Williams, G.L., Lentin, J.K., Fensome, R.A., 1998. The Lentinand Williams index of fossil dino£agellates. American Asso-ciation of Stratigraphic Palynologists Foundation Contribu-tion Series 34, College Station, TX, 817 pp.
Zonneveld, K.A.F., Versteegh, G.J.M., de Lange, G.J., 1997.Preservation of organic-walled dino£agellate cysts in di¡er-ent oxygen regimes; a 10,000 year natural experiment. Mar.Micropaleontol. 29, 393^405.
Zonneveld, K.A.F., Versteegh, G.J.M., de Lange, G.J., 2001.Paleoproductivity and post-depositional organic matter de-cay re£ected by dino£agellate cyst assemblages of the East-ern Mediterranean S1 sapropel. Mar. Geol. 172, 181^195.
MARGO 3163 3-10-02 Cyaan Magenta Geel Zwart
P.J. Mudie et al. /Marine Geology 190 (2002) 203^231 231
Related Documents