ORIGINAL PAPER Dinitrogen fixation by biological soil crusts in an Inner Mongolian steppe Jirko Holst & Klaus Butterbach-Bahl & Chunyan Liu & Xunhua Zheng & Andreas J. Kaiser & Jörg-Peter Schnitzler & Sophie Zechmeister-Boltenstern & Nicolas Brüggemann Received: 7 October 2008 / Revised: 3 April 2009 / Accepted: 7 April 2009 / Published online: 6 May 2009 # Springer-Verlag 2009 Abstract Eurasian steppe ecosystems are nitrogen-limited and suffer additionally from high grazing intensities in many areas. Soil surface-bound cyanobacteria are able to fix nitrogen and can be the major source of plant available nitrogen in such ecosystems. In this study, the abundance and dinitrogen fixation capacity of the most common soil surface-bound microbial and lichen species were deter- mined at an ungrazed, a winter-grazed, and a heavily grazed steppe site in the Xilin River catchment, Inner Mongolia, People’ s Republic of China. The microorganisms were identified as Nostoc spec. and the lichen species as Xanthoparmelia camtschadalis (Ach.) Hale by a combina- tion of classical light microscopy, confocal laser scanning microscopy and molecular analysis of the internal tran- scribed spacer (ITS1) region of ribosomal RNA. Both species were found exclusively at grazed steppe sites, with a clear difference in abundance depending on the grazing intensity. At the winter-grazed site, Nostoc was more abundant than Xanthoparmelia; for the heavily grazed site, the opposite was found. N 2 fixation was quantified with both the acetylene reduction method and 15 N 2 incubation. Cyanobacterial colonies of Nostoc fixed N 2 vigorously, whereas X. camtschadalis did not at all. The fraction of nitrogen derived from the fixation of molecular nitrogen in Nostoc was 73%, calculated from 15 N natural abundance measurements of Nostoc with X. camtschadalis as refer- ence. The conservatively calculated N 2 uptake by Nostoc was 0.030–0.033 kg N ha −1 for the heavily grazed site and 0.080–0.087 kg N ha −1 for the winter-grazed site for the growing seasons of 2004 and 2005, respectively. Together with previous findings, this study demonstrates that N 2 fixation by Nostoc can potentially replace significant amounts, if not all, of the nitrogen lost in the form of N 2 O and NO soil emissions in this steppe ecosystem. Keywords Cyanobacteria . Lichen . Semi-arid grassland . Nostoc . Xanthoparmelia camtschadalis . Grazing . MAGIM Introduction In semi-arid and arid ecosystems, in which plant growth is usually not only water-limited, but also nitrogen-limited, biological soil crusts play an important ecological role in nitrogen fixation, soil stability, and water infiltration (Belnap and Lange 2001). Typical members of these soil surface- bound communities are cyanobacteria, lichens, algae, fungi, and bryophytes (Belnap et al. 2001). Especially, cyanobac- teria and cyanolichens (lichens associated with cyanobacteria as photobionts) have received great attention, since they are able to fix atmospheric N 2 , thereby delivering nitrogen to N- limited terrestrial ecosystems via leaching or decomposition (Evans and Lange 2001). The cyanobacterial N 2 fixation is controlled by environmental factors, such as moisture (hydration status), temperature, and light intensity. Photo- Biol Fertil Soils (2009) 45:679–690 DOI 10.1007/s00374-009-0378-7 J. Holst : K. Butterbach-Bahl : A. J. Kaiser : J.-P. Schnitzler : N. Brüggemann (*) Forschungszentrum Karlsruhe, Institute for Meteorology and Climate Research, Atmospheric Environmental Research (IMK-IFU), Kreuzeckbahnstrasse 19, 82467 Garmisch-Partenkirchen, Germany e-mail: [email protected] C. Liu : X. Zheng Institute of Atmospheric Physics, Chinese Academy of Sciences, Beijing 100029, People’ s Republic of China S. Zechmeister-Boltenstern Department of Soil Biology, Institute of Forest Ecology and Soil, Seckendorff-Gudent-Weg 8, Vienna 1131, Austria

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Dinitrogen fixation by biological soil crusts in an InnerMongolian steppe
Jirko Holst & Klaus Butterbach-Bahl & Chunyan Liu & Xunhua Zheng &
Andreas J. Kaiser & Jörg-Peter Schnitzler & Sophie Zechmeister-Boltenstern &
Nicolas Brüggemann
Received: 7 October 2008 /Revised: 3 April 2009 /Accepted: 7 April 2009 /Published online: 6 May 2009# Springer-Verlag 2009
Abstract Eurasian steppe ecosystems are nitrogen-limitedand suffer additionally from high grazing intensities inmany areas. Soil surface-bound cyanobacteria are able tofix nitrogen and can be the major source of plant availablenitrogen in such ecosystems. In this study, the abundanceand dinitrogen fixation capacity of the most common soilsurface-bound microbial and lichen species were deter-mined at an ungrazed, a winter-grazed, and a heavily grazedsteppe site in the Xilin River catchment, Inner Mongolia,People’s Republic of China. The microorganisms wereidentified as Nostoc spec. and the lichen species asXanthoparmelia camtschadalis (Ach.) Hale by a combina-tion of classical light microscopy, confocal laser scanningmicroscopy and molecular analysis of the internal tran-scribed spacer (ITS1) region of ribosomal RNA. Bothspecies were found exclusively at grazed steppe sites, witha clear difference in abundance depending on the grazingintensity. At the winter-grazed site, Nostoc was moreabundant than Xanthoparmelia; for the heavily grazed site,the opposite was found. N2 fixation was quantified with
both the acetylene reduction method and 15N2 incubation.Cyanobacterial colonies of Nostoc fixed N2 vigorously,whereas X. camtschadalis did not at all. The fraction ofnitrogen derived from the fixation of molecular nitrogen inNostoc was 73%, calculated from 15N natural abundancemeasurements of Nostoc with X. camtschadalis as refer-ence. The conservatively calculated N2 uptake by Nostocwas 0.030–0.033 kg N ha−1 for the heavily grazed site and0.080–0.087 kg N ha−1 for the winter-grazed site for thegrowing seasons of 2004 and 2005, respectively. Togetherwith previous findings, this study demonstrates that N2
fixation by Nostoc can potentially replace significantamounts, if not all, of the nitrogen lost in the form ofN2O and NO soil emissions in this steppe ecosystem.
Keywords Cyanobacteria . Lichen . Semi-arid grassland .
Nostoc .Xanthoparmelia camtschadalis . Grazing .MAGIM
Introduction
In semi-arid and arid ecosystems, in which plant growth isusually not only water-limited, but also nitrogen-limited,biological soil crusts play an important ecological role innitrogen fixation, soil stability, and water infiltration (Belnapand Lange 2001). Typical members of these soil surface-bound communities are cyanobacteria, lichens, algae, fungi,and bryophytes (Belnap et al. 2001). Especially, cyanobac-teria and cyanolichens (lichens associated with cyanobacteriaas photobionts) have received great attention, since they areable to fix atmospheric N2, thereby delivering nitrogen to N-limited terrestrial ecosystems via leaching or decomposition(Evans and Lange 2001). The cyanobacterial N2 fixation iscontrolled by environmental factors, such as moisture(hydration status), temperature, and light intensity. Photo-
Biol Fertil Soils (2009) 45:679–690DOI 10.1007/s00374-009-0378-7
J. Holst :K. Butterbach-Bahl :A. J. Kaiser : J.-P. Schnitzler :N. Brüggemann (*)Forschungszentrum Karlsruhe,Institute for Meteorology and Climate Research,Atmospheric Environmental Research (IMK-IFU),Kreuzeckbahnstrasse 19,82467 Garmisch-Partenkirchen, Germanye-mail: [email protected]
C. Liu :X. ZhengInstitute of Atmospheric Physics, Chinese Academy of Sciences,Beijing 100029, People’s Republic of China
S. Zechmeister-BoltensternDepartment of Soil Biology, Institute of Forest Ecology and Soil,Seckendorff-Gudent-Weg 8,Vienna 1131, Austria

synthetic production of energy and carbon compounds(electron donors) is a prerequisite for N2 fixation, but evenduring darkness endogenous carbon reservoirs allow for N2
fixation to a limited extent (Belnap 2001).Nostoc commune is distributed worldwide and can, like
in Inner Mongolia, form macroscopic colonies in terrestrialecosystems (Doods et al. 1995). It is a popular object ofinvestigation since its resilience to drought, freezing, andUV stress, the production of biologically active com-pounds, its N2 fixation ability, and the speed of metabolicrecovery after rehydration raise many physiological andecological questions (Scherer et al. 1984; Scherer andZhong 1991; Doods et al. 1995; Kajiyama et al. 1998;Shirkey et al. 2003; Tamaru et al. 2005; Wright et al. 2005).The lichen species Xanthoparmelia camtschadalis is aconspicuous and vagrant lichen with a white maculatesurface (Hale 1990). It is distributed over vast desert,steppe, and high plain areas in North America, Eurasia,Australia, and South Africa (Hale 1990). In Mongolia andChina, this lichen occurs frequently in steppes and half-deserts (Huneck et al. 1992). The photobionts of species ofthe genera Xanthoparmelia are considered to be green algae(Hale 1990; Büdel 2001).
Many studies have shown that grazing livestock canphysically damage biological soil crusts by trampling,especially under dry conditions (Warren and Eldridge 2001).Thus, grazing as a common land-use type in (semi-)aridregions can reduce soil crust cover, abundance, biomass, andbiodiversity, often resulting in enhanced soil erosion andnutrient losses, especially where well-developed soil crustsare affected. However, several studies reported also apromotion of certain soil crust species by a grazing-inducedreduction of vascular plants and increased radiation at thesoil surface (e.g., Warren and Eldridge 2001).
The standard procedures to measure N2 fixation are theacetylene reduction assay (e.g., Hardy et al. 1973;Zechmeister-Boltenstern and Kinzel 1990) and 15N2 incor-poration (Hüser 1966). The main obstacle of the acetylenereduction assay is the necessity of a cross-calibration with15N2 incorporation to derive a factor for the conversion ofthe amount of ethylene (C2H4) formed by acetylenereduction into the amount of N2 fixed, since reportedC2H4/N2 ratios vary widely. For N. commune, a relativelylow average ratio of 0.31 was recently found (Belnap2001). Besides this, a novel type of nitrogenase was foundthat does not reduce acetylene (Ribbe et al. 1997). Thisfurther limits the applicability of the acetylene reductionassay. A third way to estimate N2 fixation of plants andcyanobacteria/cyanolichens is the 15N natural abundancetechnique (Shearer et al. 1983; Boddey et al. 2000; Russowet al. 2005). This method utilizes the fact that the 15Nabundance of N2-fixing species is very similar to that ofatmospheric dinitrogen, whereas the 15N abundance of non-
N2-fixing organisms is significantly depleted in 15N(Russow et al. 2005), reflecting the differences in 15/14Ncomposition of atmospheric N2 and ecosystem N sources.In contrast to the acetylene reduction assay and 15N2
incorporation, a great advantage of this method is that italso records seasonal changes in N assimilation (Boddey etal. 2000) and integrates over longer time periods.
Knowledge about soil surface-bound organisms in InnerMongolia is scarce. Biological soil crusts investigated inNorthern China and Mongolia typically comprise lichens,cyanobacteria (e.g., Nostoc) and few algae and mosses(Büdel 2001; Chen et al. 2005). Due to the strong increasein livestock numbers in Inner Mongolia since the 1960s,vast steppe areas are already degraded (Tong et al. 2004).Thus, also the abundance and cover of, e.g., cyanobacteriaand, hence, nitrogen input by natural N2 fixation, are verylikely negatively affected. The main objectives of this studywere: (a) to estimate the abundance of soil surface-boundmicrobial and lichen species in a typical steppe ecosystemin Inner Mongolia, People’s Republic of China, independence on grazing intensity; (b) to identify the twomost abundant macroscopically visible soil surface-boundspecies with classical light and confocal laser scanningmicroscopy as well as molecular biological methods; (c) toquantify N2 fixation rates with three experimentalapproaches, i.e., acetylene reduction, 15N2 incorporation,and the 15N natural abundance method to make the estimateof N2 fixation rates as reliable as possible; and (d) toestimate seasonal N2 fixation budgets for two steppe areaswith different grazing intensities.
Materials and methods
Study area
The study was performed at a site ungrazed since 1999(UG), a winter-grazed (WG) site, and a heavily grazed site(HG) in the Xilin River catchment, Inner Mongolia,People’s Republic of China. This area is characterized bycontinental, semi-arid climate. The long-term (1982–2003)mean annual air temperature is 0.7°C, with July as thewarmest month of the year, and the long-term annual meanprecipitation is 343 mm, with high intra- and interannualvariation and a maximum mainly between June and August.Wind directions change from NW in winter to SE insummer (Ripley 1992). The field sites were situated in aLeymus chinensis steppe, which is a common vegetationtype in Northern China (Jiang and Meurer 2001). The UGand WG sites (43°33.0′ N, 116°40.0′ E) covered an area ofapproximately 35 and 40 ha, respectively, and wereestablished by the Inner Mongolia Grassland Ecosystem
680 Biol Fertil Soils (2009) 45:679–690

Research Station (IMGERS, 43°38′ N, 116°42′ E) in 1999and 2001, respectively. The winter-grazed site is grazed inthe period from October to April by 1.5–2 sheep ha−1.Before 2001, this area had been grazed year-round by lessthan 2 sheep ha−1 for more than 20 years. The HG site (43°34.5′ N, 116°40.6′ E) was situated approximately 2 kmnortheast of the WG site and was managed by a farmer.The high grazing intensity of 6–8 sheep ha−1 during thegrowing season (May–September) had led already tochanges in the floristic composition at this site. Dominantplant species at the HG site were Potentilla acaulis L.,Artemisia frigida Willd., and the C4 grass Cleistogenessquarrosa (Trin.) Keng, whereas the UG and WG sites weremainly covered by Stipa grandis P. Smirn. and L. chinensis(Trin.) Tzvel., accompanied by a variety of other grasses. Formore details on species composition and growth dynamics,see Bai et al. (2004). The maximum vegetation height atpeak biomass was considerably lower at the HG site (<5 cm)than at WG (12–20 cm) and UG (>30 cm); vegetationcoverage was 5–50% at HG, 25–95% at WG, and almost100% at UG (Hoffmann et al. 2008). In June, higheramounts of litter were found at the UG and WG sites(~90 g dry weight (dw) m−2) as compared to the HG site(25 g dw m−2; personal communication Y. Gao, Institute ofPlant Nutrition and Soil Science, University of Kiel,Germany). The soil at all sites was a calcic chernozem.The pH was slightly below 7. At the UG and WG sites, thesoil texture could be identified as a sandy clay loam with asoil organic carbon (SOC) content of 2.6%, whereas a sandyloam with a SOC content of 1.7% was found at the HG site.The UG and WG sites were characterized by an undulatingrelief, whereas the HG site was flat. The elevation of the UGand WG sites was approximately 1,270 m above sea level(a.s.l.), whereas the HG site was approx. 1,220 m a.s.l. Forfurther site characteristics, see Holst et al. (2007), Liu et al.(2007), and Steffens et al. (2008).
Sampling
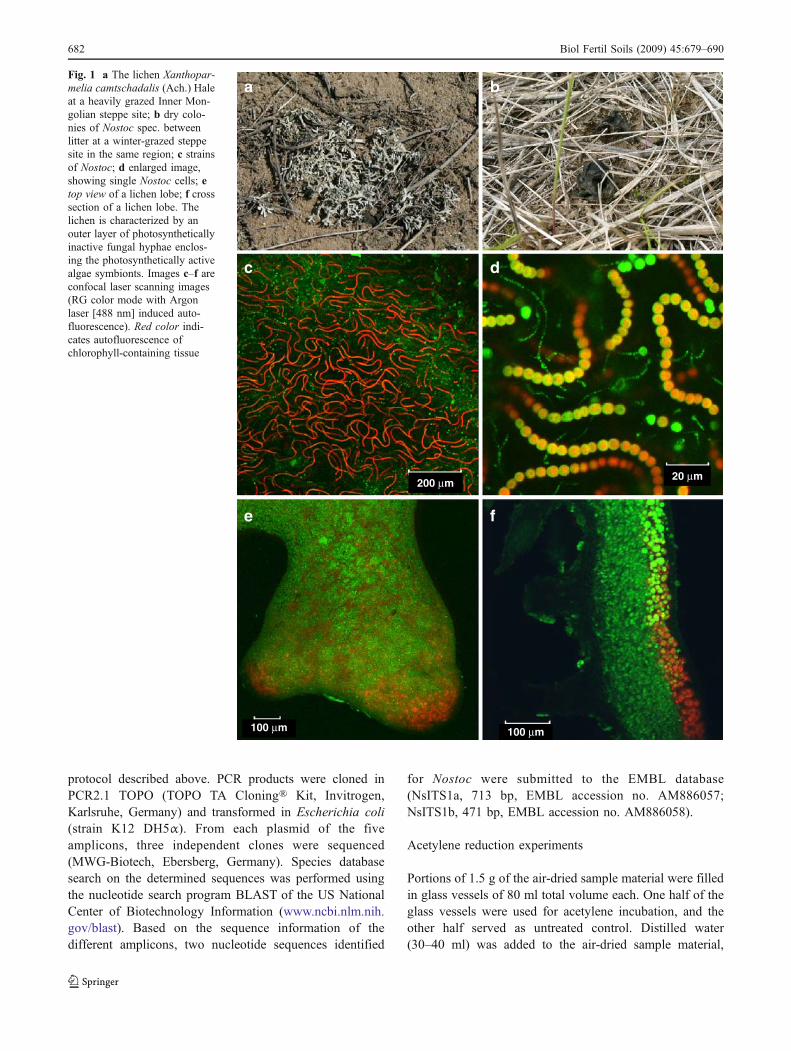
Samples of the two macroscopically visible soil surface-bound species that could be found in the study area, i.e., thelichen species X. camtschadalis (Ach.) Hale (Fig. 1a)and―at the time of collection undetermined―microbialcolonies (Fig. 1b) showing size changes in dependency ofhydration status, were collected in late summer 2005.Biomass density of both species was determined bypositioning frames (0.5 m2) at 28 positions, distributedrandomly over the entire area of each site, and collectingthe present lichens and microbial colonies. The samplematerial was air-dried and stored at room temperature untilthe start of the quantification of N2 fixation using theacetylene reduction technique and 15N2 incubation, respec-tively, in the following autumn and winter.
Species identification
For investigation of both the lichen photobiont of Xantho-parmelia and the microbial colonies, classical light andconfocal laser scanning microscopy (cLSM) was per-formed, following the key of Wirth (1995) in the case ofthe lichen photobiont, and the keys and descriptions ofCastenholz (2001) and Rippka et al. (2001) for the putativecyanobacteria. Fresh hand cuttings of the sample materialwere analyzed with a laser scanning microscope 510 (CarlZeiss Microimaging GmbH, Göttingen, Germany, equippedwith a Plan Neofluar ×10/0.3 objective or a C-Apochromat×40/1.2 water objective). Autofluorescence was visualizedin serial optical sections. A single line excitation (488 nm)and multiple channel emission technique was used. Forvisualization of multispectral image data, the differentchannels were associated with a color value on the displaysystem (RGB color mode). The two simultaneouslymeasured fluorescence channels were assigned to theprimary display colors red (long-pass filter >560 nm) andgreen (band-pass filter 505–530 nm).
To our knowledge, the photobiont species of X.camtschadalis is still unknown. The color of the collectedlichen material was pale green-bluish (Fig. 1a) and, thus,the occurrence of cyanobacteria as secondary symbionts(Rikkinen 2002) seemed to be possible. Therefore, molec-ular biological techniques were applied to analyze both thelichen and the microbial soil crust colonies for the presenceof cyanobacteria. For this purpose, DNA was extractedfrom air-dried and finely ground microbial and lichenmaterial (QIAprep Spin Miniprep Kit, Qiagen, Hilden,Germany). Quality and content of DNA were analyzedspectrophotometrically (NanoDrop ND-1000 spectropho-tometer, peqlab, Erlangen, Germany). In all cases, theabsorption ratio 260 over 280 nm was above 1.6. Forpolymerasee chain reaction (PCR) amplification (15 µl ofdistilled water, 2 µl of 10 mM Tris–HCl at pH 8.3, 0.4 µl of200 µM dNTP, 0.3 µl of 1.5 mM MgCl2, 0.4 µl of each0.5 µM primer, 1 µl of DNA, containing 1 µg DNA,0.24 µl of 2.5 U Taq polymerase), we used three sets ofoligonucleotide primers (MWG-Biotech AG, Ebersberg,Germany) to obtain three cyanobacterial 16S to 23Sribosomal RNA internally transcribed spacers (rRNA-ITS): the forward primer CSIF (without GC clamp) incombination with 373R, ITS3R, and ULR, respectively (forprimer sequences, see Janse et al. 2003). The cycling wasperformed by stepwise decrease of annealing temperaturesfollowing Janse et al. (2003). PCR products were separatedon 2% agarose gel, and bands stained with ethidiumbromide were analyzed with a UV imaging system (IntrasScience Imaging Instruments GmbH, Göttingen, Germany).The observed amplicons were cut out, purified (Qiagen Kit,Hilden, Germany), and re-amplified with the same PCR
Biol Fertil Soils (2009) 45:679–690 681
protocol described above. PCR products were cloned inPCR2.1 TOPO (TOPO TA Cloning® Kit, Invitrogen,Karlsruhe, Germany) and transformed in Escherichia coli(strain K12 DH5α). From each plasmid of the fiveamplicons, three independent clones were sequenced(MWG-Biotech, Ebersberg, Germany). Species databasesearch on the determined sequences was performed usingthe nucleotide search program BLAST of the US NationalCenter of Biotechnology Information (www.ncbi.nlm.nih.gov/blast). Based on the sequence information of thedifferent amplicons, two nucleotide sequences identified
for Nostoc were submitted to the EMBL database(NsITS1a, 713 bp, EMBL accession no. AM886057;NsITS1b, 471 bp, EMBL accession no. AM886058).
Acetylene reduction experiments
Portions of 1.5 g of the air-dried sample material were filledin glass vessels of 80 ml total volume each. One half of theglass vessels were used for acetylene incubation, and theother half served as untreated control. Distilled water(30–40 ml) was added to the air-dried sample material,
a
c
200 µm20 µm
100 µm 100 µm
fe
b
d
Fig. 1 a The lichen Xanthopar-melia camtschadalis (Ach.) Haleat a heavily grazed Inner Mon-golian steppe site; b dry colo-nies of Nostoc spec. betweenlitter at a winter-grazed steppesite in the same region; c strainsof Nostoc; d enlarged image,showing single Nostoc cells; etop view of a lichen lobe; f crosssection of a lichen lobe. Thelichen is characterized by anouter layer of photosyntheticallyinactive fungal hyphae enclos-ing the photosynthetically activealgae symbionts. Images c–f areconfocal laser scanning images(RG color mode with Argonlaser [488 nm] induced auto-fluorescence). Red color indi-cates autofluorescence ofchlorophyll-containing tissue
682 Biol Fertil Soils (2009) 45:679–690

and the wetted samples were pre-incubated for 1 h in openvessels under a cultivation lamp (Osram HQI-T 1,000 W,Osram GmbH, Munich, Germany). Photosynthetic photonflux density (PPFD) was approximately 270 µmol m−2 s−1
(determined with an LI-189 sensor, LI-COR Biosciences,Bad Homburg, Germany). After removing excess water, thevessels were closed gas tight with a screw cap equippedwith a septum. The remaining headspace was estimated tobe approximately 60 ml. To start incubations, 6 ml ofheadspace air was exchanged with acetylene (99.6 volume% (vol.%) C2H2, basi Schöberl, Rastatt, Germany), using agas-tight syringe. The acetylene was passed through 98%H2SO4 prior to application. Immediately after air–acetyleneexchange, a sample of 1 ml headspace air was taken, and allvessels were exposed to the cultivation lamp. Subsequently,four more gas samples of 1 ml each were taken in intervals of1–3 h. All gas samples were analyzed immediately for theirethylene concentration with a gas chromatograph (GC 8A,Shimadzu Deutschland GmbH, Duisburg, Germany),equipped with a flame ionization detector (FID). Syntheticair (21 vol.% O2 4.5, 79 vol.% N2 5.0, basi Schöberl) servedas carrier gas, and the temperatures of the FID and the columnoven were 250°C and 50°C, respectively. Calibration wasperformed using a range of different ethylene gas concen-trations made by mixing pure ethylene (C2H4, 99.95% purity;Air Liquide, Düsseldorf, Germany) with synthetic air.
15N2 fixation experiments
15N2 incubation experiments were conducted with bothspecies at two different temperatures (5°C and 25°C). Glassbowls filled either with 1.5 g air-dried cyanobacterialmaterial or with 2.0 g lichen material were incubated ingas-tight desiccators (~18 l total volume). Prior to the startof the experiment, the sample material was wetted withdistilled water and pre-incubated under the cultivation lampfor 1 h. Thereafter, excess water was removed, and thesample glass plates were put onto a perforated porcelainplate within the desiccator. Two liters of distilled water wasadded to each desiccator to keep air and the sample materialmoist. The desiccators were closed gas tight with lidscontaining a septum, placed in refrigerated open-bathcirculators, and exposed to the cultivation lamp. The light/dark regime was 15:9 h, which is comparable to the naturaldaylight period in summer in Inner Mongolia.
At the beginning of the incubations, 90 ml of 15N2 gas(99.87% Purity, Eurisotop, Saarbrücken, Germany) wasinjected via the septum in the lids, increasing the 15Nabundance in the headspace N2 to 1.707 atom percent (at.%), corresponding to a δ15N value of approx. 3,700‰. Theincubations lasted 14 days. The desiccator headspaceoxygen concentration was measured simultaneously usingan oxygen sensor (GOX100, Greisinger Electronic, Regen-
stauf, Germany). Whenever oxygen concentrations de-creased below 19 vol.% (especially during the lichenincubations at 25°C), we additionally injected oxygen(99.95% purity, basi Schöberl) to a final concentration ofapprox. 21 vol.% to avoid oxygen deficiency.
After the end of the incubation, all samples wereimmediately dried in a drying oven (105°C) for 24 h andthen ground in a ball mill (MM301, Retsch GmbH & Co.,Haan, Germany) for 3 min (25 rps). Approximately 2 mg ofthe sample material was filled into tin capsules (5×9 mmfor solids, IVA Analysentechnik, Meerbusch, Germany),with ten replicates for each sample. The δ15N and total Nwas analyzed with an elemental analyzer (Flash EA 1112,Thermo Fisher Scientific, Milan, Italy) coupled to a massspectrometer (DeltaPlusXP, Thermo Fisher Scientific,Bremen, Germany). Tin capsules containing 2 mg certifiedisotopic wheat flour standard (IVA Analysentechnik) wereused as reference. The N2 fixation rates were calculatedfrom the difference of the amount of 15N in labeled andunlabeled sample material and related to nitrogen contentand incubation time according to Eq. 1:
Nfix ¼15NNe �15NNs½ �15NN2 �15NNs½ �
Nc
$tincð1Þ
where Nfix is the N2 fixation rate (milligram nitrogen perkilogram dry weight per hour), 15NNe and 15NNs the 15Ncontent of Nostoc at the end and at the beginning of the15N2 fixation experiment, respectively, 15NN2 the 15Ncontent of the N2 fixed, Nc the nitrogen content of theNostoc samples (milligram per kilogram dry weight), andΔtinc the incubation time (hour).
15N natural abundance method
Samples of both species were ground using a ball mill(MM301, Retsch; 3 min at 25 rps). Two milligrams of thesample was transferred into tin capsules (5×9 mm tincapsules for solids, IVA Analysentechnik), and the δ15Nand total N content were determined using the elementalanalyzer and mass spectrometer described above.
Assumptions for extrapolation of N2 fixation
Nitrogen uptake rates were calculated on an area basis forthe WG and HG sites during the growing season (1 May to30 September), comparing a normally moist (data of 2004)with a relatively dry year (data of 2005). For extrapolationof our experimental results, we assumed that the N2-fixingspecies are only active during days with precipitation, sincethe surface drying in Inner Mongolia proceeds very quickly.However, the active phase of the N2-fixing species mighthave lasted longer, e.g., several days after the last rain event
Biol Fertil Soils (2009) 45:679–690 683

depending on the conditions following the rainfall, but aswe did not conduct specific experiments on this question,we kept our assumptions of N2-fixing activity conservative.According to the meteorological data from the IMGERS,there were 45 days with rainfall in the growing season 2004(total precipitation 288 mm) and 39 days in the growingseason of 2005 (141.5 mm). The experimentally determinedN2 fixation rates were related to three air temperatureranges: (a) 0–10°C, using the average of the fixation ratesdetermined with the 15N2 incubation at 5°C, (b) 20–30°C,using the average of the fixation rates determined with the15N2 incubation at 25°C, and (c) 10–20°C, using the meanof the fixation rates determined with 15N2 incubation at 5°Cand 25°C. In 2004, there were seven precipitation days withtemperatures between 0°C and 10°C, 37 precipitation dayswith temperatures between 10°C and 20°C, and one precipi-tation day with a temperature between 20°C and 30°C. In2005, there were six precipitation days with 0–10°C, 28precipitation days with 10–20°C, and five precipitation dayswith 20–30°C daily mean air temperature.
Results
Species determination
Confocal laser scanning microscopy images of the cyano-bacterial colonies showed embedded cell trichomes(Fig. 1c, d), which could be identified as formed by aspecies of the genus Nostoc Vaucher (Herdman et al. 2001).Confocal laser scanning microscopy and classical lightmicroscopy of X. camtschadalis revealed the typical lichenstructure with coccal green algae as photosymbiontsenclosed by fungal hyphae (Fig. 1e, f).
Cyanobacterial PCR products were found only forcyanobacterial colonies, but not for the lichen. In total,five PCR amplicons were obtained, one for the primercombination CSIF/373R (277 bp), two for CSIF/ITS3R(341 and 583 bp) and two for CSIF/ULR (471 and 713 bp),respectively. All five fragments were sequenced and usedfor analysis of homology with known DNA sequences ofthe NCBI database. The five sequences represent two 16S–23S ribosomal RNA intergenic transcribed spacer regions(NsITS1a, 713 bp; NsITS1b, 471 bp) with a sequenceidentity of 95% (Fig. 2). Compared to NsITS1a, NsITS1bshowed six deletions (Fig. 2), resulting in a reduced overalllength of 238 bp. Sequence analysis of the ITS1 region ofboth genes revealed highest similarity to ITS regions of thegenus Nostoc. Both partial 16S-ITS1-23S rRNA genesshowed the highest correlation with Nostoc desertorum(Table 1). Other matches showed similarities to Nostocindistinguenda, N. lichenoides, and N. commune. However,
a clear identification of the species or subspecies present inour cyanobacterial samples was not possible.
Species abundance
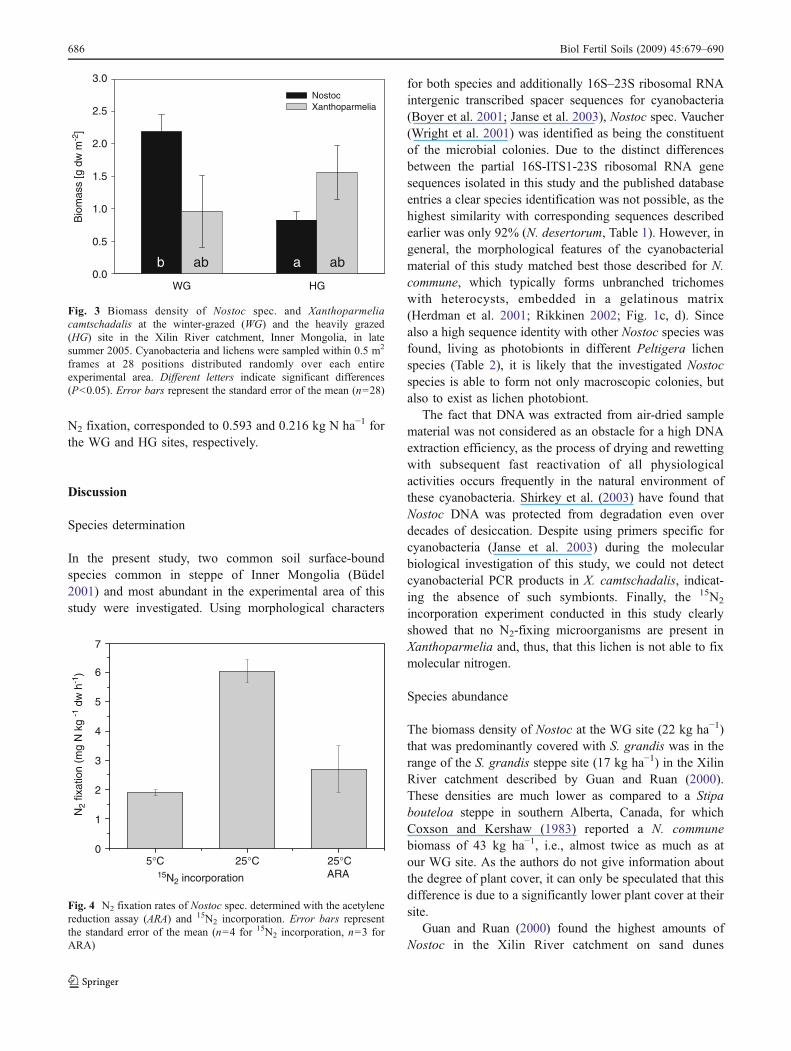
Despite intensive screening, we did not find any macroscopi-cally visible soil surface-bound species at UG, neither Nostocnor Xanthoparmelia. At WG and HG, we found only X.camtschadalis and Nostoc colonies. Nostoc was often foundtogether with small amounts of ground-covering litter. Theaverage Nostoc biomass density was significantly higher atthe WG site (22.0±2.7 kg dw ha−1) than at the HG site (8.3±1.3 kg dw ha−1; Fig. 3). For Xanthoparmelia, the inversepattern was found, with 9.6±5.6 kg dw ha−1 at WG and15.6±4.2 kg dw ha−1 at HG. However, the difference wasnot significant. At both sites, less than 1% of the total areawas covered with macroscopically visible soil crust species.
Acetylene reduction assay
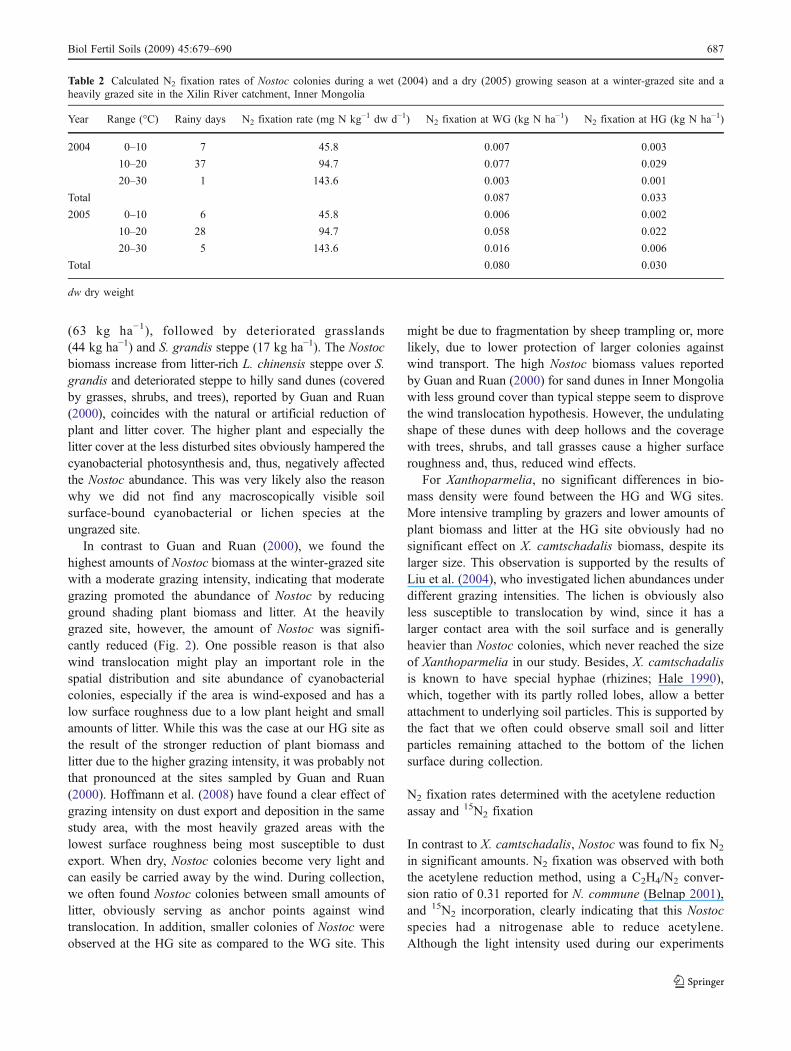
A substantial production of ethylene was observed inNostoc samples incubated with acetylene. No ethyleneproduction was found for Xanthoparmelia. However,Xanthoparmelia showed a color change from gray-greento reddish-orange during the acetylene incubation. Using aC2H4/N2 conversion factor of 0.31 (Belnap 2001), a meanN2 fixation rate of 2.7 mg N kg−1 dw h−1 was calculated forNostoc at 25°C (Fig. 4).
15N2 fixation
At both temperature levels (5°C and 25°C), 15N2 was fixed byNostoc. The mean 15N abundance was 0.3658±0.0001 at.%15N (n=4, ±SEM) before incubation, 0.4143±0.0019 at.%15N after 14 days (336 h) of incubation at 5°C, and 0.4876±0.0224 at.% 15N after 14 days of incubation at 25°C,indicating that 3.62% and 9.08% of the total nitrogen werederived from N2 (1.707 at.% 15N) fixed during theincubation at 5°C and 25°C, respectively. Xanthoparmeliadid not fix N2, the 15N abundance remained unchanged(0.3642±0.0001 at.% 15N before incubation; 0.3642±0.0002 at.% 15N after 14 days of incubation at 5°C;0.3639±0.0001 at.% 15N after 14 days of incubation at25°C incubation). The Nostoc samples incubated at 5°C and25°C had a mean N content of 1.77±0.03% and 2.24±0.04%, respectively. Thus, according to Eq. 1, the mean N2
fixation rates of Nostoc were 1.91 and 6.05 mg N kg−1 dwh−1 for the incubation at 5°C and 25°C, respectively (Fig. 4).On the basis of our upscaling assumptions, these ratescorrespond to a growing season N2 fixation of 0.087 and0.080 kg N ha−1 at the WG site and to 0.033 and 0.030 g Nha−1 at the HG site for the growing periods of 2004 and2005, respectively (Table 2).
684 Biol Fertil Soils (2009) 45:679–690

15N natural abundance
The mean δ15N value was −6.25±0.21‰ for Xanthopar-melia and −1.71±0.35‰ for Nostoc (n=12, ±SEM). Usingthe formulas given by Russow et al. (2005), the fraction of
N in Nostoc derived from atmospheric N2 was calculated as73% (with Xanthoparmelia as reference). The total amountof N stored in Nostoc was calculated as 0.812 and 0.295 kgN ha−1 for the WG and HG sites, respectively. Therefore,the total amount of N stored in Nostoc colonies, gained by
Table 1 Similarity of NsITS1a and NsITS1b internal transcribed spacer regions (NsITS1a: bp 133–665; NsITS1b: bp 133–423, for sequence seeFig. 2) to published cyanobacterial ITS sequences
Sequence Species compared Accession no. Query coverage (%) Identity (%)
NsITSa Nostoc desertorum CM1-VF14 AY579899.1 100 92
Nostoc indistinguenda CM1-VF10 AY579896.1 98 88
Nostoc sp. “Peltigera britannica cyanobiont” AY560646.1 96 83
Nostoc lichenoides CNP-AK1 AY579894.1 91 91
NsITSb Nostoc desertorum CM1-VF14 AY579901.1 100 92
Nostoc lichenoides CNP-AK1 AY579895.1 100 85
Nostoc commune EV1-KK1 AY579904.1 97 91
Nostoc indistinguenda CM1-VF10 AY579897.1 97 88
Fig. 2 Alignment of two partial16S-ITS1-23S ribosomal RNAgene sequences of Nostoc spec.isolated from samples collectedat an experimental field site inthe Inner Mongolia steppe. Grayboxes indicate 16S and 23SRNA transitions compared toEscherichia coli (dark gray;EMBL accession no. EF527444)and Phormidium inundatum(light gray; EMBL accession no.AM398954). Sequence identitybetween NsITSa and NsITSb is95%
Biol Fertil Soils (2009) 45:679–690 685
N2 fixation, corresponded to 0.593 and 0.216 kg N ha−1 forthe WG and HG sites, respectively.
Discussion
Species determination
In the present study, two common soil surface-boundspecies common in steppe of Inner Mongolia (Büdel2001) and most abundant in the experimental area of thisstudy were investigated. Using morphological characters
for both species and additionally 16S–23S ribosomal RNAintergenic transcribed spacer sequences for cyanobacteria(Boyer et al. 2001; Janse et al. 2003), Nostoc spec. Vaucher(Wright et al. 2001) was identified as being the constituentof the microbial colonies. Due to the distinct differencesbetween the partial 16S-ITS1-23S ribosomal RNA genesequences isolated in this study and the published databaseentries a clear species identification was not possible, as thehighest similarity with corresponding sequences describedearlier was only 92% (N. desertorum, Table 1). However, ingeneral, the morphological features of the cyanobacterialmaterial of this study matched best those described for N.commune, which typically forms unbranched trichomeswith heterocysts, embedded in a gelatinous matrix(Herdman et al. 2001; Rikkinen 2002; Fig. 1c, d). Sincealso a high sequence identity with other Nostoc species wasfound, living as photobionts in different Peltigera lichenspecies (Table 2), it is likely that the investigated Nostocspecies is able to form not only macroscopic colonies, butalso to exist as lichen photobiont.
The fact that DNA was extracted from air-dried samplematerial was not considered as an obstacle for a high DNAextraction efficiency, as the process of drying and rewettingwith subsequent fast reactivation of all physiologicalactivities occurs frequently in the natural environment ofthese cyanobacteria. Shirkey et al. (2003) have found thatNostoc DNA was protected from degradation even overdecades of desiccation. Despite using primers specific forcyanobacteria (Janse et al. 2003) during the molecularbiological investigation of this study, we could not detectcyanobacterial PCR products in X. camtschadalis, indicat-ing the absence of such symbionts. Finally, the 15N2
incorporation experiment conducted in this study clearlyshowed that no N2-fixing microorganisms are present inXanthoparmelia and, thus, that this lichen is not able to fixmolecular nitrogen.
Species abundance
The biomass density of Nostoc at the WG site (22 kg ha−1)that was predominantly covered with S. grandis was in therange of the S. grandis steppe site (17 kg ha−1) in the XilinRiver catchment described by Guan and Ruan (2000).These densities are much lower as compared to a Stipabouteloa steppe in southern Alberta, Canada, for whichCoxson and Kershaw (1983) reported a N. communebiomass of 43 kg ha−1, i.e., almost twice as much as atour WG site. As the authors do not give information aboutthe degree of plant cover, it can only be speculated that thisdifference is due to a significantly lower plant cover at theirsite.
Guan and Ruan (2000) found the highest amounts ofNostoc in the Xilin River catchment on sand dunes
5°C 25°C 25°C0
1
2
3
4
5
6
7
N2
fixat
ion
(mg
N k
g -1
dw
h-1
)
15N2 incorporation ARA
Fig. 4 N2 fixation rates of Nostoc spec. determined with the acetylenereduction assay (ARA) and 15N2 incorporation. Error bars representthe standard error of the mean (n=4 for 15N2 incorporation, n=3 forARA)
WG HG
Bio
mas
s [g
dw
m-2
]
0.0
0.5
1.0
1.5
2.0
2.5
3.0
NostocXanthoparmelia
a ababb
Fig. 3 Biomass density of Nostoc spec. and Xanthoparmeliacamtschadalis at the winter-grazed (WG) and the heavily grazed(HG) site in the Xilin River catchment, Inner Mongolia, in latesummer 2005. Cyanobacteria and lichens were sampled within 0.5 m2
frames at 28 positions distributed randomly over each entireexperimental area. Different letters indicate significant differences(P<0.05). Error bars represent the standard error of the mean (n=28)
686 Biol Fertil Soils (2009) 45:679–690
(63 kg ha−1), followed by deteriorated grasslands(44 kg ha−1) and S. grandis steppe (17 kg ha−1). The Nostocbiomass increase from litter-rich L. chinensis steppe over S.grandis and deteriorated steppe to hilly sand dunes (coveredby grasses, shrubs, and trees), reported by Guan and Ruan(2000), coincides with the natural or artificial reduction ofplant and litter cover. The higher plant and especially thelitter cover at the less disturbed sites obviously hampered thecyanobacterial photosynthesis and, thus, negatively affectedthe Nostoc abundance. This was very likely also the reasonwhy we did not find any macroscopically visible soilsurface-bound cyanobacterial or lichen species at theungrazed site.
In contrast to Guan and Ruan (2000), we found thehighest amounts of Nostoc biomass at the winter-grazed sitewith a moderate grazing intensity, indicating that moderategrazing promoted the abundance of Nostoc by reducingground shading plant biomass and litter. At the heavilygrazed site, however, the amount of Nostoc was signifi-cantly reduced (Fig. 2). One possible reason is that alsowind translocation might play an important role in thespatial distribution and site abundance of cyanobacterialcolonies, especially if the area is wind-exposed and has alow surface roughness due to a low plant height and smallamounts of litter. While this was the case at our HG site asthe result of the stronger reduction of plant biomass andlitter due to the higher grazing intensity, it was probably notthat pronounced at the sites sampled by Guan and Ruan(2000). Hoffmann et al. (2008) have found a clear effect ofgrazing intensity on dust export and deposition in the samestudy area, with the most heavily grazed areas with thelowest surface roughness being most susceptible to dustexport. When dry, Nostoc colonies become very light andcan easily be carried away by the wind. During collection,we often found Nostoc colonies between small amounts oflitter, obviously serving as anchor points against windtranslocation. In addition, smaller colonies of Nostoc wereobserved at the HG site as compared to the WG site. This
might be due to fragmentation by sheep trampling or, morelikely, due to lower protection of larger colonies againstwind transport. The high Nostoc biomass values reportedby Guan and Ruan (2000) for sand dunes in Inner Mongoliawith less ground cover than typical steppe seem to disprovethe wind translocation hypothesis. However, the undulatingshape of these dunes with deep hollows and the coveragewith trees, shrubs, and tall grasses cause a higher surfaceroughness and, thus, reduced wind effects.
For Xanthoparmelia, no significant differences in bio-mass density were found between the HG and WG sites.More intensive trampling by grazers and lower amounts ofplant biomass and litter at the HG site obviously had nosignificant effect on X. camtschadalis biomass, despite itslarger size. This observation is supported by the results ofLiu et al. (2004), who investigated lichen abundances underdifferent grazing intensities. The lichen is obviously alsoless susceptible to translocation by wind, since it has alarger contact area with the soil surface and is generallyheavier than Nostoc colonies, which never reached the sizeof Xanthoparmelia in our study. Besides, X. camtschadalisis known to have special hyphae (rhizines; Hale 1990),which, together with its partly rolled lobes, allow a betterattachment to underlying soil particles. This is supported bythe fact that we often could observe small soil and litterparticles remaining attached to the bottom of the lichensurface during collection.
N2 fixation rates determined with the acetylene reductionassay and 15N2 fixation
In contrast to X. camtschadalis, Nostoc was found to fix N2
in significant amounts. N2 fixation was observed with boththe acetylene reduction method, using a C2H4/N2 conver-sion ratio of 0.31 reported for N. commune (Belnap 2001),and 15N2 incorporation, clearly indicating that this Nostocspecies had a nitrogenase able to reduce acetylene.Although the light intensity used during our experiments
Table 2 Calculated N2 fixation rates of Nostoc colonies during a wet (2004) and a dry (2005) growing season at a winter-grazed site and aheavily grazed site in the Xilin River catchment, Inner Mongolia
Year Range (°C) Rainy days N2 fixation rate (mg N kg−1 dw d−1) N2 fixation at WG (kg N ha−1) N2 fixation at HG (kg N ha−1)
2004 0–10 7 45.8 0.007 0.003
10–20 37 94.7 0.077 0.029
20–30 1 143.6 0.003 0.001
Total 0.087 0.033
2005 0–10 6 45.8 0.006 0.002
10–20 28 94.7 0.058 0.022
20–30 5 143.6 0.016 0.006
Total 0.080 0.030
dw dry weight
Biol Fertil Soils (2009) 45:679–690 687

was considerably lower as compared to the mean daytimePPFD of ~480 µmol m−2 s−1 during summertime in InnerMongolia (personal communication B. Ketzer, TechnicalUniversity of Dresden, Germany), this was not consideredto be an obstacle for the quantification of N2 fixation rates,since soil cyanobacteria and cyanolichens were found toreach the maximum nitrogenase activity already under lowlight intensities (Belnap 2001). The fact that N2 fixationrates determined with acetylene reduction at 25°C werelower than those determined with 15N2 incorporation at thesame temperature could be attributed on the one hand to thefact that the C2H4/N2 conversion ratio of 0.31 might be toolow for the Nostoc species studied here. On the other hand,different types of nitrogenases can be involved in cyano-bacterial N2 fixation (Tsygankov 2007), and not all of themcan reduce acetylene (Ribbe et al. 1997). Thus, the higherobserved N2 fixation rates might be also attributed to theactivity of nitrogenases that cannot be detected by theacetylene reduction method.
The N2 fixation rates were considerably lower than thoseestimated by Guan and Ruan (2000) based on acetylenereduction measurements. Nitrogen fixation rates of N.commune or Nostoc spec. reported by Vlassak et al.(1973), Coxson and Kershaw (1983), and Scherer andZhong (1991) were also higher by at least a factor of 10than those in our study. However, the N2 fixation ratescalculated in the present study were of the same order ofmagnitude as nitrogen losses in the form of N2O (0.03–0.14 kg N2O–N ha−1 year−1) and NO (0.01–0.04 kg NO–Nha−1 year−1) in the same study area as previously reportedby Holst et al. (2007). The differences between N2 fixationrates in this and in other studies might have been caused byseveral factors. First of all, our assumption that thecyanobacteria were active only during days with precipita-tion is extremely conservative. Their active phase mighthave lasted longer, i.e., several days after the last rain eventdepending on the conditions following the rainfall, but wedid not conduct specific experiments on this question. Alsomoistening by dew during nighttime might have sustainedthe cyanobacterial activity. As another potential reason fordifferent results, Belnap (2001) stressed the influence ofpre-experimental conditions on the determination of N2
fixation rates of cyanobacteria. Factors like initial N and Ccontent (relevant for dark fixation), storage time andconditions after sampling, and also the chosen experimentaltemperature, light and hydration conditions play the mostimportant role. Continuous light during incubation, forinstance, was found to increase N2 fixation rates signifi-cantly over time during acetylene reduction as compared toa light–dark incubation (Vlassak et al. 1973), therebypotentially overestimating N2 fixation rates. Also, a tooshort incubation time can lead to an overestimation of N2
fixation rates, since N losses via denitrification and NH3
volatilization can be considerable during longer incubationtimes (Coxson and Kershaw 1983; Evans and Johanson1999; Evans and Lange 2001). The fact that we appliedboth a short-term (acetylene reduction) and a long-term(15N2) incubation, resulting in comparable N2 fixation rates,strengthens the reliability of our results. The diurnal light/dark pattern and the incubation time of 2 weeks should haveintegrated physiological variations after rewetting and dueto light changes as well as N losses.
15N natural abundance
Russow et al. (2005) observed δ15N values close to zero forN2-fixing lichens (≥ −2‰ for a cyanobacteria-containingCollema lichen) as compared to non-fixing lichens (−11‰for a Squamarina lichen). Furthermore, they estimated thefraction of N derived from atmospheric N2 to be 88% of thetotal Collema-N. Also in our study, the N2-fixing specieshad a higher δ15N value than the non-fixing species(−1.71‰ versus −6.25‰). We calculated a rate of Nderived from atmospheric N2 of 73% for Nostoc, compa-rable to the value reported by Russow et al. (2005). Thus,despite their N2-fixing capability, the cyanolichen Collemaand the Nostoc colonies derive a non-negligible part of theirN from other sources, like rainwater, the soil matrix, and/ordecomposing litter.
To calculate the total ecosystem N input by cyanobac-teria according to Russow et al. (2005), growth rates ofNostoc spec. would have been required, which were notavailable in this study. Therefore, we were only able torelate the estimated amounts of fixed N2, based on ourexperimentally determined fixation rates, to the total N poolof Nostoc. The estimated amounts of N2 fixed during thegrowing season corresponded to approximately 10% of thepresent Nostoc N stock (Table 2). However, this is incontrast to the observations of Gao and Yu (2000), whodetermined a considerable biomass increase between 12%and 20% for daily wetted Nostoc flagelliforme during a 15-day incubation period. N. flagelliforme also forms cyano-bacterial colonies and is distributed in arid and semi-aridareas of China. This can only be explained by differences inphysiological characteristics and growth rates betweendifferent Nostoc species, since we also ensured a sufficientwetness of the cyanobacterial colonies and a day/nightcycle during incubation, like reported by Gao and Yu(2000).
Conclusions
More than interannual variations in precipitation, grazingintensity had a major influence on the abundance of soilsurface-bound N2-fixing Nostoc colonies in the investigated
688 Biol Fertil Soils (2009) 45:679–690

L. chinensis steppe in Inner Mongolia, whereas theabundance of the non-N2-fixing lichen X. camtschadaliswas less affected. Moderate grazing intensity promoted theoccurrence of Nostoc in the investigated L. chinensis steppeas compared to ungrazed steppe, where no macroscopicallyvisible cyanobacteria and lichens were found, by improvingthe growth conditions for the cyanobacterial coloniesthrough reduction of plant biomass and litter. A furtherincrease in grazing pressure led to a significant decrease inNostoc biomass. However, even under such conditions, theabundance of the investigated species was higher ascompared to an ungrazed site. Thus, moderate grazing willsupport cyanobacterial N2 fixation in this N-limitedecosystem.
Acknowledgments The work has been supported by the GermanResearch Foundation (DFG, Research Unit No. 536, “Matter fluxes ingrasslands of Inner Mongolia as influenced by stocking rate”,MAGIM) and by the National Natural Science Foundation of China(NSFC, project no. 40331014). Special thanks go to Zhihong Yu andShubin Yu for their assistance during the collection of the samplematerial, to Ina Zimmer for DNA isolation, and to Rudolf Meier forhis support during mass spectrometric analysis.
References
Bai YF, Han XG, Wu JG, Chen ZZ, Li LH (2004) Ecosystem stabilityand compensatory effects in the Inner Mongolia grassland.Nature 431:181–184. doi:10.1038/nature02850
Belnap J (2001) Factors influencing nitrogen fixation and nitrogenrelease in biological soil crusts. In: Belnap J, Lange OL (eds)Biological soil crusts: structure, function and management.Springer, Berlin, pp 241–261
Belnap J, Lange OL (2001) Structure and functioning of biologicalsoil crusts: a synthesis. In: Belnap J, Lange OL (eds) Biologicalsoil crusts: structure, function and management. Springer, Berlin,pp 471–479
Belnap J, Büdel B, Lange OL (2001) Biological soil crusts:characteristics and distribution. In: Belnap J, Lange OL (eds)Biological soil crusts: structure, function and management.Springer, Berlin, pp 3–30
Boddey RM, Peoples MB, Palmer B, Dart PJ (2000) Use of the 15Nnatural abundance technique to quantify biological nitrogenfixation by woody perennials. Nutr Cycl Agroecosyst 57:235–270. doi:10.1023/A:1009890514844
Boyer SL, Flechtner VR, Johansen JR (2001) Is the 16S–23S rRNAinternal transcribed spacer region a good tool for use inmolecular systematics and population genetics? A case study incyanobacteria. Mol Biol Evol 18:1057–1069
Büdel B (2001) Biological soil crusts of Asia including the Don andVolga region. In: Belnap J, Lange OL (eds) Biological soil crusts:structure, function and management. Springer, Berlin, pp 87–94
Castenholz RW (2001) General characteristics of the cyanobacteria. In:Boone DR, Castenholz RW (eds) Bergey’s manual of systematicbacteriology. Vol. 1: The archaea and the deeply branching andphototrophic bacteria. Springer, Berlin, pp 474–487
Chen J, Zhang MY, Wang L, Shimazaki H, Tamura M (2005) A newindex for mapping lichen-dominated biological soil crusts indesert areas. Remote Sens Environ 96:165–175. doi:10.1016/j.rse.2005.02.011
Coxson DS, Kershaw KA (1983) The pattern of in situ summernitrogenase activity in terrestrial Nostoc commune from Stipa-Bouteloa grassland, southern Alberta. Can J Bot 61:2686–2693.doi:10.1139/b83-295
DoodsWK,Gudder DA,Mollenhauer D (1995) The ecology ofNostoc. JPhycol 31:2–18. doi:10.1111/j.0022-3646.1995.00002.x
Evans RD, Lange OL (2001) Biological soil crusts and ecosystemnitrogen and carbon dynamics. In: Belnap J, Lange OL (eds)Biological soil crusts: structure, function and management.Springer, Berlin, pp 263–279
Evans RD, Johanson JR (1999) Microbiotic crusts and ecosystemprocesses. Crit Rev Plant Sci 18:183–225. doi:10.1016/S0735-2689(99)00384-6
Gao K, Yu A (2000) Influence of CO2, light and watering on growthof Nostoc flagelliforme mats. J Appl Phycol 12:185–189.doi:10.1023/A:1008123203409
Guan X, Ruan X (2000) The characters of bionitrogen fixation ofNostoc commune and its function in nitrogen cycle in InnerMongolia grassland. Acta Agrestia Sin 8:13–17
Hale ME (1990) A synopsis of the lichen genus Xanthoparmelia(Vainio) Hale (Ascomycotina, Parmeliaceae). Smithson ContribBot 74:1–250
Hardy RW, Burns RC, Holsten RD (1973) Applications of the acetylene–ethylene assay for measurement of nitrogen fixation. Soil BiolBiochem 5:47–81. doi:10.1016/0038-0717(73)90093-X
Herdman M, Castenholz RW, Rippka R (2001) Form-genus VIII.Nostoc Vaucher 1803. In: Boone DR, Castenholz RW (eds)Bergey’s manual of systematic bacteriology. Vol. 1: The archaeaand the deeply branching and phototrophic bacteria. Springer,Berlin, pp 575–580
Holst J, Liu C, Brüggemann N, Butterbach-Bahl K, Zheng X, Wang Y,Han S, Yao Z, Han X (2007) Microbial N turnover and N-oxide(N2O/NO/NO2) fluxes in semi-arid grassland of Inner Mongolia.Ecosystems (N Y, Print) 10:623–634. doi:10.1007/s10021-007-9043-x
Hoffmann C, Funk R, Li Y, Sommer M (2008) Effect of grazing onwind driven carbon and nitrogen ratios in the grasslands of InnerMongolia. Catena, doi:10.1016/j.catena.2008.06.003
Huneck S, Ahti T, Cogt U, Poelt J, Sipman H (1992) Zur Verbreitungund Chemie von Flechten der Mongolei. III. Ergebnisse derMongolisch-Deutschen Biologischen Expedition seit 1962. NovaHedwig 54:277–308
Hüser R (1966) Experiences with the nitrogen-15 tracer technique inestimating the microbial fixation of elementary nitrogen in theorganic matter of forest soils. J Appl Radiat Isot (Spec.)(Suppl.):457–469
Janse I, Meima M, Edwin W, Kardinaal A, Zwart G (2003) High-resolution differentiation of cyanobacteria by using rRNA-internal transcribed spacer denaturing gradient gel electrophore-sis. Appl Environ Microbiol 69:6634–6643. doi:10.1128/AEM.69.11.6634-6643.2003
Jiang Y, Meurer M (2001) Die Steppen Nordchinas und ihre Belastungdurch weide- und landwirtschaftliche Landnutzung. GeogrRundsch 10:48–52
Kajiyama S, Kanzaki H, Kawazu K, Kobayashi A (1998) Nostofun-gicidine, an antifungal lipopeptide from the field grown terrestrialblue-green alga Nostoc commune. Tetrahedron Lett 39:3737–3740. doi:10.1016/S0040-4039(98)00573-5
Liu Z, Wang S, Han J, Wang Y, Chen Z (2004) Distribution of lichensbiomass and its affecting factors during restoration process ofInner Mongolia steppe. Chin J Appl Ecol 15:1294–1296
Liu C, Holst J, Brüggemann N, Butterbach-Bahl K, Yao Z, Jin Y, HanS, Han X, Krümmelbein J, Horn R, Zheng X (2007) Grazingreduces methane uptake by soils in a semi-arid steppe in InnerMongolia, China. Atmos Environ 41:5948–5958. doi:10.1016/j.atmosenv.2007.03.017
Biol Fertil Soils (2009) 45:679–690 689

Ribbe M, Gadkari D, Meyer O (1997) N2 fixation by Streptomycesthermoautotrophicus involves a molybdenum-dinitrogenase anda manganese-superoxide oxidoreductase that couple N2 reductionto the oxidation of superoxide produced from O2 by amolybdenum-CO dehydrogenase. J Biol Chem 272:26627–26633. doi:10.1074/jbc.272.42.26627
Rikkinen J (2002) Cyanolichens: an evolutionary overview. In: RaiAN, Bergman B, Rasmussen R (eds) Cyanobacteria in symbiosis.Kluwer, Dordrecht, pp 31–72
Ripley EA (1992) Grassland Climate. In: Coupland RT (ed)Ecosystems of the world. Vol 8A: Natural grasslands. Introduc-tion and western hemisphere. Elsevier, Amsterdam, pp 7–24
Rippka R, Castenholz RW, Herdman M (2001) Subsection IV.(Formerly Nostocales Castenholz 1989b sensu Rippka, Der-uelles, Waterbury, Herdman and Stanier 1979). In: Boone DR,Castenholz RW (eds) Bergey’s manual of systematic bacteriol-ogy. Vol. 1: The archaea and the deeply branching and photo-trophic bacteria. Springer, Berlin, pp 563–566
Russow R, Veste M, Böhme F (2005) A natural 15N approach todetermine the biological fixation of atmospheric nitrogen bybiological soil crusts of Negev desert. Rapid Commun MassSpectrom 19:3451–3456. doi:10.1002/rcm.2214
Scherer S, Ernst A, Chen TW, Böger P (1984) Rewetting of drought-resistant blue-green algae: time course of water uptake andreappearance of repiration, photosynthesis, and nitrogen fixation.Oecologia 62:418–423. doi:10.1007/BF00384277
Scherer S, Zhong ZP (1991) Desiccation independence of terrestrialNostoc commune ecotypes (cyanobacteria). Microb Ecol 22:271–283. doi:10.1007/BF02540229
Shearer G, Kohl HD, Virhinia RA, Bryan BA, Skeeters JL, Nilsen ET,Sharifi MR, Rundel PW (1983) Estimates of N2-fixation fromvariation in the natural abundance of 15N in Sonoran desertecosystem. Oecologia 56:365–373. doi:10.1007/BF00379714
Shirkey B, McMaster NJ, Smith SC, Wright DJ, Rodriguez H, JarugaP, Birincioglu M, Helm RF, Potts M (2003) Genomic DNA ofNostoc commune (Cyanobacteria) becomes covalently modifiedduring long-term (decades) desiccation but is protected from
oxidative damage and degradation. Nucleic Acids Res 31:2995–3005. doi:10.1093/nar/gkg404
Steffens M, Kölbl A, Totsche KU, Kögel-Knabner I (2008) Grazingeffects on soil chemical and physical properties in a semiaridsteppe of Inner Mongolia (P.R. China). Geoderma 143:63–72.doi:10.1016/j.geoderma.2007.09.004
Tamaru Y, Takami Y, Yoshida T, Sakamoto T (2005) Crucial role ofextracellular polysaccharides in desiccation and freezing tolerance inthe terrestrial cyanobacterium Nostoc commune. Appl EnvironMicrobiol 71:7327–7333. doi:10.1128/AEM.71.11.7327-7333.2005
Tong C, Wu J, Yong S, Yang J, Yong W (2004) A landscape-scaleassessment of steppe degradation in the Xilin River Basin, InnerMongolia, China. J Arid Environ 59:133–149. doi:10.1016/j.jaridenv.2004.01.004
Tsygankov AA (2007) Nitrogen-fixing cyanobacteria: a review. ApplBiochemMicrobiol 43:279–288. doi:10.1134/S0003683807030040
Vlassak K, Paul EA, Harris RE (1973) Assessment of biologicalnitrogen fixation in grasslands and associated sites. Plant Soil38:637–649. doi:10.1007/BF00010702
Warren SD, Eldridge DJ (2001) Biological soil crusts and livestock inarid ecosystems: are they compatible? In: Belnap J, Lange OL(eds) Biological soil crusts: structure, function and management.Springer, Berlin, pp 401–415
Wirth V (1995) Flechtenflora. Ulmer, StuttgartWright D, Prickett T, Helm RF, Potts M (2001) Form species Nostoc
commune (Cyanobacteria). Int J Syst Evol Microbiol 51:1839–1852
Wright DJ, Smith SC, Joardar V, Scherer S, Jervis J, Warren A, HelmRF, Potts M (2005) UV irradiation and desiccation modulate thethree-dimensional extracellular matrix Nostoc commune (Cyano-bacteria). J Biol Chem 280:40271–40281. doi:10.1074/jbc.M505961200
Zechmeister-Boltenstern S, Kinzel H (1990) Non-symbiotic nitrogenfixation associated with temperate soils in relation to soilproperties and vegetation. Soil Biol Biochem 22:1075–1084.doi:10.1016/0038-0717(90)90032-U
690 Biol Fertil Soils (2009) 45:679–690
Related Documents











