Fabiana Alves Mourão Orientadora: Profa. Dra. Claudia Maria Jacobi Co-orientador: Prof. Dr. José Eugênio Côrtes Figueira Dinâmica do forrageamento da hemiparasita Struthanthus flexicaulis Mart.(Loranthaceae) e sua influência na estrutura da comunidade vegetal de campos rupestres ferruginosos – MG

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Fabiana Alves Mourão
Orientadora: Profa. Dra. Claudia Maria Jacobi
Co-orientador: Prof. Dr. José Eugênio Côrtes Figueira
Dinâmica do forrageamento da hemiparasita Struthanthus
flexicaulis Mart.(Loranthaceae) e sua influência na estrutura
da comunidade vegetal de campos rupestres ferruginosos –
MG

Universidade Federal de Minas Gerais
Instituto de Ciências Biológicas
Programa de Pós-graduação em Ecologia, Conservação e Manejo da Vida Silvestre
Tese de Doutorado
Dinâmica do forrageamento da hemiparasita Struthanthus flexicaulis
Mart.(Loranthaceae) e sua influência na estrutura da comunidade vegetal
de campos rupestres ferruginosos – MG
Fabiana Alves Mourão
Orientadora: Profa. Dra. Claudia Maria Jacobi
Co-orientador: Prof. Dr. José Eugênio Côrtes Figueira
Belo Horizonte, fevereiro de 2011
Tese apresentada à Universidade Federal de
Minas Gerais, como pré-requisito para a
obtenção do título de Doutora em Ecologia.

ii
Dedico este trabalho aos meus pais Heliana e
Ronaldo e ao Rick, pelo amor incondicional.

iii
Agradecimentos
A Claudia Maria Jacobi, minha orientadora querida e amiga, que me conduziu
tão bem e com tanto carinho durante todos estes anos. Obrigada Claudia por ser meu
exemplo, aprendi com você muito mais que teorias ecológicas e desenvolvimento de
projetos.
A José Eugênio Côrtes Figueira (o Zé), meu co-orientador, por ser brilhante,
ético, gentil e amigo. Obrigada Zé, por ter me acompanhado desde a graduação, me
apoiando sempre e me fazendo acreditar que eu podia chegar lá. Sua amizade, carinho e
dedicação foram essenciais durante todos esses anos.
Ao prof. Ronald Dickman, por compartilhar gentilmente seus avançados
conhecimentos sobre modelos matemáticos e nos permitir conhecer um pouquinho do
mundo da física. Obrigada Dickman por suas valiosas aulas particulares e por todo
auxilio que recebemos durante a elaboração e desenvolvimento do modelo de
simulação.
Ao Júlio Cesar Mansur Filho, meu amigo da física, inteligente, gentil, prestativo
e paciente. Sua ajuda, dedicação e infinitas explicações foram fundamentais para a
elaboração e conclusão do capítulo 2 desta tese.
Aos meus queridos estagiários e verdadeiros amigos que tornaram possível a
realização deste trabalho: Manuel Gontijo, Rafael Barros, Arthur Dias e Matheus
Scarpelli. Agradeço também aos estagiários de finais de semana: Cecília, Juliana, Luiza,
Fernando, que abdicaram de alguns sábados para nos ajudar com a coleta de dados.
Aos professores membros da banca: Rafael Arruda, Tadeu Guerra, Fernando
Vale, Frederico Neves, Maria Auxiliadora Drumond e Clemens Schlindwein, que
gentilmente aceitaram avaliar este trabalho.
Ao Programa ECMVS pelo apoio recebido nas saídas de campo. À secretaria do
curso por resolver pacientemente todas as minhas dificuldades. Aos professores e
colegas do curso pelos ensinamentos e amizade.
Ao IEF pela licença de coleta, aos funcionários do Parque Estadual da Serra do
Rola Moça pelo apoio logístico e ao CNPq pelo auxílio financeiro (Bolsa).

iv
Aos amigos da Biotrópicos e aos milhões de amigos do ICB e do Parque
Estadual da Serra do Rola Moça!!!
Aos meus pais Heliana e Ronaldo, e minha avó Terezinha, que não entendem
muito bem o que eu faço, mas sempre torceram pelo meu sucesso. Às minhas irmãs
Renata e Luciana, por serem motivos de muitas alegrias. A minha afilhadinha Ana
Luiza e meu sobrinho Gui, presentes de Deus. Ao meu primo Fernando (In memorian)
que sempre torceu por mim e estaria muito feliz por este dia ter chegado!
A Rick, minha razão de viver, meu porto seguro nos momentos de maior
dificuldade e para quem eu sempre quero voltar para compartilhar a alegria de uma
vitória!
A todos aqueles que de alguma forma contribuíram para meu crescimento
pessoal e profissional e a alguém que eventualmente eu tenha me esquecido de
agradecer.
OBRIGADA!!!

v
APOIO:

vi
SUMÁRIO
Resumo 1
Abstract 2
Introdução Geral 3
Relevância deste estudo 5
Referências Bibliográficas 8
Capítulo 1 – Forrageamento direcionado da parasita Struthanthus flexicaulis 13
Resumo 14
Introdução 15
Materiais e Métodos 17
Resultados 21
Discussão 26
Referências bibliográficas 29
Anexo 1 - Diferentes fenofases de Struthanthus flexicaulis (Loranthaceae) 33
Capítulo 2 - Modelagem espacial do espalhamento da parasita Struthanthus
flexicaulis em paisagens de afloramentos ferruginosos, Brasil.
34
Resumo 35
Introdução 36
Materiais e Métodos 38
Resultados 47
Discussão 51
Referências bibliográficas 54

vii
Capítulo 3 - Alocação de recursos da hemiparasita Struthanthus flexicaulis
(Loranthaceae) em condições de estresse hídrico do ambiente.
58
Resumo 59
Introdução 60
Materiais e Métodos 62
Resultados 65
Discussão 70
Referências bibliográficas 74
Capítulo 4 - Efeito top-down da hemiparasita Struthanthus flexicaulis
(Loranthaceae) e seu papel chave na estruturação de comunidades vegetais em
afloramentos ferruginosos do Quadrilátero Ferrífero, Brasil.
77
Resumo 78
Introdução 79
Materiais e Métodos 81
Resultados 85
Discussão 91
Referências bibliográficas 96
Anexo 2A - Lista das espécies vegetais - área de altas densidades da parasita 102
Anexo 2B - Lista das espécies vegetais - área de baixas densidades da parasita 103
Considerações Finais 104

1
Resumo
O objetivo principal do estudo foi investigar a importância da planta
hemiparasita Struthanthus flexicaulis (Loranthaceae) na estrutura de comunidades
vegetais em afloramentos ferruginosos, por meio de uma abordagem integrando dados
de alocação de recursos e de modelagem da sua dispersão. Para isso a tese foi
estruturada em quatro capítulos que contemplaram respectivamente os seguintes
objetivos específicos: 1) determinar os possíveis fatores que influenciam o
espalhamento de S. flexicaulis na comunidade vegetal e verificar se esta hemiparasita
apresenta comportamento de forrageamento direcionado; 2) modelar o espalhamento
desta hemiparasita usando o modelo de Gás em Rede em uma paisagem com densidade
da hospedeira Mimosa calodendron semelhante à encontrada na área de estudos, 3)
comparar a alocação de recursos de S. flexicaulis em diferentes condições de estresse
hídrico do ambiente; e 4) avaliar os efeitos do parasitismo na comunidade (Top-down e
Espécie-chave). A densidade de hospedeiras influenciou o espalhamento de S.
flexicaulis nesta comunidade. O sucesso de propagação vegetativa dependeu da altura
das hospedeiras, bem como o distanciamento entre suas copas. O forrageamento
direcionado dos ramos da hemiparasita foi comprovado, já que alteravam
significativamente suas trajetórias de crescimento indo de encontro à nova fonte de
recursos. Os resultados da simulação reforçaram a importância da distância entre
hospedeiras, e também da dispersão ornitocórica na velocidade de espalhamento da
parasita. S. flexicaulis alocou recursos de maneira diferenciada nas estações seca e
chuvosa: investiu mais no crescimento vegetativo nos meses secos, em detrimento de
estruturas reprodutivas, mais abundantes na chuva. Esta hemiparasita causou um efeito
top-down e foi considerada espécie-chave nesta comunidade, pois, embora não tenha
modificado a diversidade ou composição de espécies, provocou grande impacto na
cobertura vegetal de algumas espécies importantes no afloramento ferruginoso
estudado.

2
Abstract
The main objective of this study was to establish the importance of the
hemiparasitic plant Struthanthus flexicaulis (Loranthaceae) in the structure of plant
communities on ironstone outcrops, by means of an approach that integrated data on
resource allocation and dispersal modeling. To achieve this, the dissertation was
structured in four chapters with the following specific objectives: 1) to determine the
possible factors that influence the spread of S. flexicaulis in the plant community and
verify of the parasite has an oriented foraging behavior; 2) to model the spatial dispersal
of the parasite by means of a lattice gas automata model across a landscape of its main
host (Mimosa calodendron) similar to that found in the study area; 3) to compare
resource allocation of S. flexicaulis in different environmental water stress conditions,
and 4) to evaluate the effects of parasitism in the community (top-down and keystone
species). Host density influenced the spread of S. flexicaulis in the community. The
vegetative propagation depended on host height as well as distance among hosts.
Oriented branch foraging behavior was confirmed, since branches significantly altered
their growth trajectories towards new resource points. The simulation results reinforced
the importance of distance among hosts as well as ornitochory on the speed of parasite
spread. S. flexicaulis allocated resources differentially between the dry and wet seasons:
it invested more in vegetative growth during the dry months, in detriment of
reproductive structures, more abundant during the rainy period. This parasite caused a
negative top-down effect and was considered a keystone species in its community
because, although it did not modify diversity or species composition, it exerted a great
impact on the percent cover of some important hosts.

3
Introdução geral
As plantas parasitas constituem um interessante grupo para estudos ecológicos,
pois podem modificar a estrutura e dinâmica da comunidade onde estão inseridas,
reduzindo a biomassa e alterando a alocação de recursos das espécies parasitadas
(Tennakoon & Pate 1996; Lei 2001, Press & Phoenix 2005, Shen et al. 2006). Elas
podem afetar negativamente processos fisiológicos, reprodutivos e ecológicos de suas
hospedeiras (Press et al. 1999, Lei 1999, 2001) interferindo no balanço de água e
nutrientes, reduzindo as taxas de fotossíntese e respiração e, em algumas situações,
podendo levá-las à morte (Aukema 2003). Apesar do efeito negativo nas espécies
parasitadas, podem também desempenhar papel benéfico na comunidade por que são
utilizadas como fonte de recursos para uma variedade de animais vertebrados, entre eles
polinizadores e dispersores de sementes (Watson 2001, Aukema 2003, Kelly et al.
2004). As plantas parasitas podem exercer um efeito top down em suas hospedeiras e
permitir a existência de espécies competitivamente subordinadas (Press, 1998),
principalmente se as espécies mais parasitadas forem as dominantes.
Devido a estes efeitos, já foram consideradas espécies-chave das suas
comunidades (Press & Phoenix 2005) e até determinantes da diversidade alfa (Watson
2001). São também apontadas como importantes elementos na paisagem porque
aumentam a complexidade estrutural do ambiente (Bennets et al. 1996). Com exceção
destas referências recentes, o seu papel e sua conservação têm sido bastante ignorados
em diversos estudos de comunidades vegetais (Norton & Reid 1997), e a maioria dos
estudos ecológicos são realizados em laboratório ou com espécies consideradas pragas
agrícolas, em lugar de comunidades naturais (Pennings e Callaway 2002).
A especificidade por hospedeiras é um fator chave no estudo do grupo de plantas
parasitas e muitos padrões já foram observados (Norton & De Lange 1999, Arruda et al.
2009). Em ambientes homogêneos como florestas de coníferas (Shaw et al. 2004), a
especificidade pode ser vantajosa, pois aumentaria a eficiência na retirada de água e sais
minerais da hospedeira. Neste tipo de paisagem a planta parasita teria sua dispersão
vegetativa garantida por meio do espalhamento dos ramos, pois todas as hospedeiras
(única espécie) poderiam ser facilmente infectadas. Entretanto, ser especialista em
ambientes heterogêneos pode ser desvantajoso, pois aumenta a dificuldade – seja por
parte da planta ou do agente dispersor – de encontrar o hospedeiro específico, tornando
inviável a sobrevivência em um ambiente onde esta hospedeira ocorre em baixas

4
densidades (Norton & Carpenter 1998, Arruda et al. 2006). Foi sugerido que o grau de
especificidade está relacionado não somente à abundancia local como também ao grau
de constância das hospedeiras no tempo e no espaço (Norton & Carpenter 1998, Norton
& De Lange 1999), complementando a hipótese da fragmentação do recurso proposta
por Janzen (1981).
A facilidade de encontrar hospedeiras é somente um dos fatores que contribuem
para a especificidade das parasitas, junto com outros anatômicos e até químicos que
promovem o reconhecimento das plantas e a formação de haustórios, raízes altamente
especializadas capazes de retirar água e sais minerais de suas hospedeiras (Rodl & Ward
2002, Press & Phoenix 2005, Arruda et al. 2006, Shen et al. 2006). Assim, muitas
plantas parasitas são generalistas e atacam um grande número de espécies (Pennings &
Callaway 2002), e podem aumentar a infestação do parasitismo em hospedeiras que
oferecem maiores condições de sobrevivência e reprodução. As leguminosas são as
hospedeiras preferidas devido à capacidade de fixação de nitrogênio (Radomiljac et
al.1999, Bowie & Ward 2004).
Entre as plantas parasitas melhor conhecidas estão famílias como Loranthaceae e
Viscaceae que também ocorrem no Brasil. Os principais dispersores de sementes das
mais de suas 1200 espécies são aves, motivo pelo qual são vulgarmente conhecidas
como ervas-de-passarinho (Kujt 1969, Barboza 2000, Cazetta & Galetti 2003, Arruda et
al. 2012). Muitas espécies destas famílias são hemiparasitas, pois possuem folhas
clorofiladas capazes de realizar fotossíntese e retiram água e sais minerais de suas
hospedeiras, em lugar de produtos fotoassimilados (Calder & Bernardt 1983). Outras,
geralmente aclorofiladas, não são capazes de realizar fotossíntese e são chamadas de
holoparasitas.
A família Loranthaceae é constituída de 75 gêneros (Calvin & Wilson 2006),
distribuídos em todos os continentes, exceto nos pólos (Calder & Bernardt 1983). No
Brasil foram descritas cerca de 100 espécies, distribuídas em dez gêneros (Souza &
Lorenzi 2005). São consideradas epiparasitas porque infectam os ramos aéreos de suas
hospedeiras e a presença de raízes epicorticais que ocorre em todas as espécies do
gênero Struthanthus, crescem ao longo dos galhos das hospedeiras formando haustórios
espaçadamente, com potencial de crescimento clonal em decorrência da fragmentação
dos ramos (Ehleringer et al. 1985, Calvin & Wilson 2006). O interesse primário no
estudo de Loranthaceae no Brasil tem sido os aspectos anatômicos e embriológicos da
sua relação com hospedeiras (Venturelli 1981, Venturelli 1984, Venturelli & Kraus

5
1989, Mourão 2007, Amaral et al. 2011, Mourão et al.- submetido), enquanto que as
interações ecológicas hemiparasita-hospedeiras têm sido comparativamente pouco
estudadas. No entanto, sugere-se que representantes desta família sejam generalistas em
regiões tropicais, pois usam com sucesso uma grande variedade de espécies hospedeiras
(Arruda et al. 2006, Mourão et al. 2006, Mourão 2007, Fadini 2011).
Embora diversos aspectos da ecologia das ervas-de-passarinho têm sido estudados
em diferentes ecossistemas (Davidar 1983, Sargent 1995), especialmente a dispersão de
sementes (Sutton 1951, Davidar 1978, Overton 1994, del Rio et al. 1996, López-de-
Buen e Ornelas 1999, Aukema & del Rio 2002) e a especificidade de parasitismo (Kuijt
1963, Barlow & Wiens 1977, Thomson & Mahal 1983, Hoffman et al. 1986), ou sua
condição fisiológica (Dean et al. 1994, Bickford et al. 2005), ainda há poucos estudos
quantitativos sobre a dinâmica populacional das Loranthaceae a qual, obrigatoriamente,
está vinculada à distribuição espacial das suas hospedeiras (Shaw et al. 2004, Teodoro
et al., dados não publicados). Diante disso, torna necessária uma abordagem de pesquisa
em que a paisagem de hospedeiras esteja definida, tanto na composição de espécies
quanto nas suas qualidades como recurso para a parasita.
Comparados com campos quartzíticos do Quadrilátero Ferrífero (MG), e embora
sujeitos a um grau muito maior de degradação devido ao seu interesse comercial (Viana
& Lombardi 2007, Mourão & Stehmann 2007) os afloramentos ferruginosos (campos
rupestres sobre canga) têm sido muito pouco estudados, seja em levantamentos
florísticos ou em estudos ecológicos (Jacobi et al. 2007, Mourão 2007). Neste sentido, o
presente estudo fará uso de uma paisagem natural para descrever a densidade de
hospedeiras, sua configuração espacial (presença ou ausência de rochas expostas) e
frequência de parasitismo (que determina a mortalidade diferencial de hospedeiras) para
simular o espalhamento de S. flexicaulis nestes afloramentos ferruginosos (canga).
Relevância do estudo
O ecossistema escolhido para a realização deste trabalho foi o campo rupestre
sobre canga couraçada, também conhecido como afloramento ferruginoso. A canga é
um substrato de distribuição restrita, e no Brasil encontra-se principalmente concentrada
no Quadrilátero Ferrífero (QF), região de ecótone de dois hotspots brasileiros, o
Cerrado e a Mata Atlântica, que têm sido profundamente ameaçados e modificados
pelas atividades humanas. A canga vem sofrendo impactos constantes devido à

6
exploração do minério de ferro e à urbanização, com enormes perdas na biodiversidade
local e regional (Viana & Lombardi 2007, Jacobi & Carmo 2008). Ao contrário do que
se pensava até pouco tempo atrás, as comunidades vegetais dos campos rupestres sobre
canga são bastante ricas. Um recente estudo mostrou que em apenas 100 ha foram
encontradas 430 espécies de angiospermas pertencentes a 78 famílias, que
correspondem a 36% de todas as famílias de angiospermas brasileiras (Jacobi et al.
2007). Devido às ameaças relatadas acima e ao restrito conhecimento sobre os campos
rupestres sobre canga, as espécies vegetais ali localizadas são prioritariamente indicadas
para o desenvolvimento de estudos ecológicos, incluindo padrões de riqueza e de
distribuição das espécies (Jacobi & Carmo, 2008).
Este estudo apresenta uma nova abordagem tanto no que se refere à modelagem do
espalhamento de uma planta parasita quanto suas interações ecológicas, podendo ser o
primeiro a evidenciar o efeito top-down através do parasitismo em plantas como um
possível fator estruturador da comunidade vegetal. Estudos recentes têm demonstrado
poucas evidências do efeito top-down em comunidades terrestres (Schmitz et al. 2000).
A maioria destes estudos procura demonstrar este efeito através da predação (Atlegrim
1989, Chase 1998, Marquis & Whelan 1994, Fonseca 1994, Oliveira 1997, Beckerman et
al. 1997, Kneitel & Miller 2002) e alguns poucos através de parasitóides (Gomez &
Zamora 1994). O termo top-down surgiu na década de 60 quando Hairston e
colaboradores (1960) difundiam a ideia de cascata trófica onde um controle da cadeia
poderia ser realizado “de cima para baixo”, ou seja, herbívoros controlando a abundância
de plantas. Os conceitos top-down e bottom-up foram utilizados em diversos trabalhos
que consideravam dois, três ou mais níveis tróficos. Grande parte dos experimentos
envolvendo mais níveis tróficos tem sido realizada em ambientes aquáticos por
apresentarem uma composição menos complexa da comunidade ou uma melhor
definição da cadeia trófica (Strong 1992). Os experimentos no ambiente terrestre,
entretanto, usam cadeias de forma incompleta, dificilmente passam de três níveis
tróficos, e geralmente envolvem remoção ou adição de predadores.
Além disso, este estudo utiliza o modelo físico de difusão de partículas
(Autômatos Celulares) conhecido como de Gás em Rede, para estudar o espalhamento
de S. flexicaulis sobre a vegetação. Este modelo tem sido pouco explorado por biólogos,
mas alguns trabalhos utilizaram-no para investigar a dinâmica de predadores e suas
presas e para estudar a mobilidade de organismos (Satulovsky & Tome 1994, Chen et
al. 2006, Rodrigues & Tomé 2008). Este modelo é capaz de incorporar características

7
da paisagem natural e informações sobre os indivíduos, fornecendo padrões espaciais de
distribuição de espécies semelhante às observações de campo. Os dados sobre
interações em ambientes de afloramentos ferruginosos são escassos e aspectos como o
efeito de parasitas na estrutura e funcionamento de comunidades vegetais nesses
ecossistemas, por exemplo, são totalmente desconhecidos, dificultando qualquer
tentativa de conservação e manejo destas áreas. Neste sentido, através dos resultados
gerados neste trabalho, poderão ser elaboradas ações de manejo que visem a
conservação das espécies vegetais que vivem sobre a canga.

8
Referências bibliográficas
Amaral, M. M. D. & Ceccantini, G. C. T. 2011. The endoparasite Pilostyles ulei
(Apodanthaceae Cucurbitales) influences wood structure in three host species of
Mimosa. IAWA Journal, 32: 1-13.
Arruda, R.; Carvalho, L. N. & Del-Claro, K. 2006. Host specificity of a Brazilian
mistletoe, Struthanthus aff. polyanthus (Loranthaceae), in cerrado tropical savanna.
Flora, 201: 127-134.
Arruda, R.;Fadini, R. F.; Mourão, F. A. ; Jacobi, M. J. & Teodoro, G. S. 2009. Natural
History and Ecology of neotropical mistletoe. Encyclopedia of Life Support Systems,
UNESCO, 24p.
Arruda, R.;Fadini, R. F.; Carvalho, L.; Del-Claro, K.; Mourão, F. A.; Jacobi; C. M.;
Teodoro, G. S.; van den Berg, E.; Caires, C. S. & Dettke, G. A. 2012. Ecology of
Neotropical mistletoe: an important canopy-dwelling component of Brazilian forests
and savannas. Acta Brasília Botânica.
Aukema, J. E. 2003. Vectors, viscin, and Viscaceae: mistletoes as parasites, mutualists
and resources. Frontiers in Ecology and the Environment, 1: 212-219.
Aukema, J. E.; del Rio, C. M. 2002. Where does a fruit-eating bird deposit mistletoe
seeds? Seed deposition patterns and an experiment. Ecology, 83: 3489-3496.
Atlegrim, O. 1989. Exclusion of birds from bilberry stands: impact on insect larval
density and damage to bilberry. Oecologia, 79: 136-139.
Barboza, M. A. 2000. Loranthaceae e Viscaceae no bioma cerrado. Dissertação de
mestrado Universidade de Brasília, 76 pp.
Barlow, B. A. & Wiens, D. 1977. Host-parasite resemblance in Australian mistletoes:
the case for cryptic mimicry. Evolution, 31: 69-84
Bennets, R. E., White G. C., Hawksworth F.G. & Severs S. E. 1996. The influence of
dwarf mistletoe on bird communities in Colorado ponderosa pine forest. Ecological
Applications, 6: 899-909
Bickford, C. P.; Kolb, T. E.; Geils, B. W. 2005. Host physiological condition regulates
parasitic plant performance: Arceuthobium vaginatum subsp cryptopodum on Pinus
ponderosa. Oecologia, 146: 179-189.
Bowie, M. & Ward, D. 2004. Water and nutrient status of the mistletoe Plicosepalus
acaciae parasitic on isolated Negev Desert populations of Acacia raddiana differing
in level of mortality. Journal of Arid Environments, 56: 487-508.

9
Calder, M. & Bernardt, P. 1983. The biology of mistletoes. M. Calder & Bernardt (eds.)
Academic Press, Sydney, 333 pp.
Calvin, C. L. & Wilson C. A. 1995. Relationship of the mistletoe Phoradendron
macrophyllum (Viscaceae) to the wood of its host. Iawa Journal, 16: 33-45.
Calvin, C. L. & Wilson C. A. 2006. Comparative morphology of epicortical roots in Old
and New World Loranthaceae with reference to root types, origin, patterns of
longitudinal extension and potential for clonal growth. Flora, 201: 345-353.
Cazetta, E. & Galetti, M. 2003. Ecologia das ervas-de-passarinho. Ciência Hoje, 33: 73-
75.
Chase, J. 1998. Central place forager effects on the food chain dynamics and special
pattern in Northern California meadows. Ecology, 79: 1236-1245.
Chen, Q.; Mao, J. & Li, W. 2006. Stability analysis of harvesting strategies in a cellular
automata based predator-prey model. Lecture Notes in Computer Science 4173: 268-
276.
Davidar, P. 1978. Dispersal in some Loranthaceae of Nilgiris. Journal Bombay Natural
History Society, 75: 943-945.
Davidar, P. 1983. Birds and neotropical mistletoes: effects on seedling recruitment.
Oecologia, 60: 271-273.
Dean, W. R. J.; Midgley, J. J. & Stock, W. D. 1994. The distribution of mistletoes in
South-Africa- patterns of species richness and host choice. Journal of Biogeography,
21: 503-510.
Del Rio, C. M.; Silva, A.; Medel, R. & Hourdequin, M. 1996. Seed dispersers as
disease vectors: bird transmission of mistletoe seeds to plant hosts. Ecology, 77: 912-
921.
Dzerefos, C. M.; Witkowski, E. T. F. & Shackleton, C. M. 2003. Host-preference and
density of woodrose-forming mistletoes (Loranthaceae) on savanna vegetation,
South Africa. Plant Ecology, 167: 163-177.
Ehleringer, J. R., Schulze, E. D., Ziegler, H., Lange, O. L., Farquhar, G. D. & Cowar, I.
R. (1985). Xylem-tapping mistletoes: water or nutrients parasites? Science, 227:
1479-1481.
Fadini, R. F. 2011. Non-overlap of hosts used by three congeneric and sympatric
loranthaceous mistletoes in an Amazonian savanna: host generalization to extreme
specialization. Acta Botanica Brasílica, 25: 337-345.

10
Fonseca, C. R. 1994. Herbivory and the long-lived leaves of an Amazonia ant-tree.
Journal of Ecology, 82: 833-842.
Guerra, T. J. A. 2005. Componentes quantitativos e qualitativos da dispersão de
sementes de Struthanthus flexicaulis (Loranthaceae) em uma área de campo
rupestre do sudeste brasileiro. Dissertação de mestrado. Universidade Estadual
Paulista, 64p.
Gomez, J. M. & Zamora, R. 1994. Top-down effects in a tritrophic system: parasitoids
enhance plant fitness. Ecology, 75: 1023-1030.
Hoffman, A. J. Fuentes, E. R., Cortes, I. Liberona, F. & Costa, V. 1986 Tristerix
tetrandrus (Loranthaceae) and its hosts plants in the Chilean matorral: Patterns and
mechanisms. Oecologia, 69: 202-206.
Jacobi, C. M.; Carmo, F. F.; Vincent, R. C. & Stehmann, J. R. 2007. Plant communities
on ironstones outcrops: a diverse and endangered Brazilian ecosystem. Biodiversity
and Conservation, 16: 2185-2200.
Jacobi, C. M. & Carmo, F. F. 2008. The contribution of ironstone outcrops to plant
diversity in the Iron Quadrangle, a threatened Brazilian landscape. Ambio, 37:324–
326.
Janzen, D. H. 1981. The peak in North American Ichneumonid species richness lies
between 38˚ and 42˚ N. Ecology, 62: 532-537.
Kelly, D.; Ladley, J.J.; Robertson, A.W. 2004. Is dispersal easier than pollination?
Two tests in new Zealand Loranthaceae. New Zealand Journal of Botany, 42: 89-
103.
Kneitel, J. M. & Miller, T. E. 2002. Resource and top-predator regulation in the pitcher
plant (Sarracenia purpurea) inquiline community. Ecology, 83: 680-688.
Kuijt, J. 1963. On the ecology and parasites of the Costa Rica Tree Mistletoe,
Gaiadendron punctatum (Ruiz and Pavon). Canadian Journal of Botany, 41: 927-
938.
Kuijt, J. 1969. The biology of parasitic plants. University of California Press, Berkeley,
246pp.
Lei, S. A. 1999. Age, size and water status of Acacia gregii influencing the infection
and reproductive success of Phoradendron californicum. American Midland
Naturalist, 141: 358-365.
Lei, S. A. 2001. Survival and development of Phoradendron californicum and Acacia
greggii during a drought. Western North American Naturalist, 61: 78-84.

11
Marquis, R. J. &Whelan, C. J. 1994.Insectivorous birds increase growth of white oak
through consumption of leaf-chewing insects. Ecology, 75: 2007-2014.
Mourão, F. A.; Carmo, F. F; Ratton, P.; Jacobi, C. M. 2006. Hospedeiras da
hemiparasita Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) em campos
rupestres ferruginosos, Quadrilátero Ferrífero, MG. Lundiana, 7: 103-110.
Mourão, F.A. 2007. Padrão de parasitismo de Struthanthus flexicaulis (Mart.) Mart.
(Loranthaceae) e seus efeitos na comunidade vegetal de um campo rupestre sobre
canga do Quadrilátero Ferrífero (MG). Dissertação de mestrado, Universidade
Federal de Minas Gerais, Belo Horizonte. 64pp.
Mourão, F. A.; Vale, F. H. A; Pimenta, M & Jacobi, C. M. Caracterização anatômica e
histoquímica do hemiparasitismo de Struthanthus flexicaulis (Mart.) Mart.
(Loranthaceae) em diferentes hospedeiras em campos rupestres sobre canga no
Parque Estadual da Serra do Rola Moça – MG. Submetido à Revista Brasileira de
Botânica.
Mourão, A. & Stehmann, J. R. 2007. Levantamento da flora do campo rupestre sobre
canga hematítica couraçada remanescente na Mina do Brucutu, Barão de Cocais,
Minas Gerais, Brasil. Rodriguésia, 54: 775-786.
Norton, D. A. & Carpenter, M. A. 1998. Mistletoes as parasites: host specificity and
speciation. Trends in Ecology and Evolution, 13: 101-105.
Norton, D. A. & De Lange, P. J. 1999. Host specificity in parasitic mistletoes
(Loranthaceae) in New Zealand. Functional Ecology, 13:552-559.
Norton, D.A. & Reid, N. 1997. Lessons in ecosystem management from management of
threatened and pest loranthaceous mistletoes in New Zealand and Australia.
Conservation Biology, 11: 759-769.
Oliveira, P. S. 1997. The ecological function of extrafloral nectaries: herbivore
deterrence by visiting ants and reproductive output in Caryocar brasilienses
(Caryocaraceae). Functional Ecology, 11: 323-330.
Overton, J. M. 1994. Dispersal and infection in mistletoe metapopulations. Journal of
Ecology, 82: 711-723.
Overton, J. M. 1996. Spatial autocorrelation and dispersal in mistletoes: Field and
simulation results. Vegetatio, 125: 83-98.
Pennings, S. C.; Callaway, R. M. 2002. Parasitic plants: parallels and contrasts with
herbivores. Oecologia, 131: 479-489.

12
Press M. C.; Scholes, J. D. & Watling, J. R. 1999. Parasitic plants: physiological e
ecological interactions whit their hosts in Press, M. C.; Scholes, J. D. & Barker M.
G.; (eds.) Physiological plant Ecology. Blackwell Scientific Oxford, 175-197 pp.
Press, M. C. & Graves, J. D. 1995. Parasitic plants. M. C. Press & J. D. Graves (eds.)
Chapman & Hall, London, 292pp.
Press, M. C. & Phoenix, G. K. 2005. Impacts of parasitic plants on natural communities.
New Phytologist, 166: 737-751.
Radomiljac, A. M.; McComb, J. A. & Pate, J. S. 1999. Gas exchange and water
relations of the root hemiparasite Santalum album L. in association with legume
and non-legume hosts. Annals of Botany, 83: 215-224.
Rodrigues, A. L. & Tomé, T. 2008. Reaction-diffusion stochastic lattice model for a
predator-prey system. Brazilian Journal of Physics, 38: 87-93.
Rodl, T. & Ward, D. 2002. Host recognition in a desert mistletoe: early stages of
development are influenced by substrate and host origin. Functional Ecology, 16:
128-134.
Sargent, S. 1995. Seed fate in a tropical mistletoe: the importance of host twig size.
Funtional Ecology, 9: 197-204.
Satulovsky, J. E. & Tomé T. 1994. Stochastic lattice gas model for predator-prey
system. Physical Review Letters. 49: 50 -73.
Schmitz, O. J.; Hamback, P. A. & Beckerman, A. P. 2000. Trophic cascades in
terrestrial systems: a review of the effects of carnivore removals on plants. The
American Naturalist, 155: 141-153.
Shaw, D. C.; Watson, D. M. & Mathiasen, R. L. 2004. Comparison of dwarf mistletoes
(Arceuthobium spp., Viscaceae) in the Western United States with mistletoes
(Amyema spp., Loranthaceae) in Australia - ecological analogs and reciprocal
models for ecosystem management. Australian Journal of Botany, 52: 481-498.
Shen, H.; Ye, W.; Hong, L.; Huang, H.; Wang, Z.; Deng, X.; Yang, Q.; Xu, Z. 2006.
Progress in parasitic plant biology: Host selection and nutrient transfer. Plant
Biology, 8: 175-185.
Souza, V. C. & Lorenzi, H. 2005. Botânica sistemática: guia ilustrado para identificação
das famílias de Angiospermas da flora brasileira, baseado em APG II. Instituto
Plantarum.

13
Strong, D. R. 1992. Are trophic cascades all wet? Differentiation and donor-control in
speciose ecosystems. Ecology, 73: 747-754.
Sutton, G. M. 1951. Mistletoe dispersal by birds. Wilson Bulletin, 63:235-237.
Tennakoon, K. U.; Pate, J. S. 1996. Effects of parasitism by a mistletoe on the structure
and functioning of branches of its host. Plant Cell and Environment, 19: 517-528.
Thomson, V. E. and Mahal, B. E. 1983. Host specificity by a mistletoe, Phoradendron
villosum (Nutt.) Nutt. Subsp villosum, on three oak species in California. Botanical
Gazette, 144: 124-131.
Venturelli, M. 1981. Estudos sobre Struthanthus vulgaris Mart.: Anatomia do fruto e
semente e aspectos de germinação. Revista Brasileira de Botânica, 4: 131-147.
Venturelli, M. 1984. Estudos sobre Struthanthus vulgaris Mart.: aspectos anatômicos de
raiz adventícia, caule e folha. Revista Brasileira de Botânica, 7: 79-89.
Venturelli, M. & Kraus, J. E., 1989. Morphological and anatomical aspects of the
primary haustorium of Struthanthus vulgaris Mart. (Loranthaceae) in vitro. Revista
Brasileira de Botânica, 12: 17–22.
Viana, P. L. & Lombardi, J. A. 2007. Florística e caracterização dos campos rupestres
sobre a canga na Serra da Calçada, Minas Gerais, Brasil. Rodriguésia, 58: 159-177.
Watson. D. M. 2001. Mistletoe - A keystone resource in forests and woodlands
worldwide. Annual Review of Ecology and Systematics, 32: 219-249.

13
CAPÍTULO 1
Forrageamento direcionado da hemiparasita
Struthanthus flexicaulis
Fabiana Alves Mourão1, Claudia Maria Jacobi
1 & José Eugenio Cortes Figueira
1
1Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade
Federal de Minas Gerais.

14
Resumo
As plantas parasitas podem se dispersar vegetativamente e esse tipo de crescimento tem
como finalidade colonização de novas áreas ou captação de recursos (forrageamento).
Se a busca de uma hospedeira for direcionada, a parasita estaria forrageando. O objetivo
desse trabalho foi determinar os possíveis fatores que influenciam a propagação
vegetativa da parasita Struthanthus flexicaulis na comunidade vegetal e verificar se
apresenta forrageamento direcionado. Foram demarcadas três áreas para a realização
das observações. Nas duas menores que apresentavam diferentes densidades de Mimosa
calodendron - sua principal hospedeira em afloramentos ferruginosos, foram avaliados
os efeitos da densidade dessas plantas na propagação de ramos (contágio). Na maior
área foram avaliados: 1) a influência da distância entre copas de hospedeiras nas
passagens dos ramos; 2) o sucesso de propagação vegetativa dos ramos e 3) o
forrageamento (mudança na direção dos ramos). A área de maior densidade de
hospedeiras apresentou uma distância média de borda entre vizinhas mais próximas de
27,8 ± 38,2 cm e na área de menor densidade essa distância foi de 140,7 ±123,7 cm.
Quanto menor a distância maior foi a chance de transmissão da parasita. Esta é uma
relação exponencial negativa. A propagação vegetativa de ramos ocorreu mais
frequentemente na área com maior densidade de mimosas, em que 89% dos ramos da
parasita tiveram sucesso ao encontrar uma nova hospedeira. Já na área menos densa
apenas 21% conseguiram explorar novas copas. A propagação vegetativa para duas ou
três hospedeiras consecutivas foi mais comum na área de maior densidade (30%); na de
menor densidade somente cerca de 5% dos ramos passavam para mais de uma
hospedeira (χ 2=226,5; gl=1; p<0,001). O efeito combinado da altura de hospedeiras e
distância entre copas foi determinante para o sucesso de propagação vegetativa da
parasita. A parasita apresentou forrageamento direcionado, pois após o encontro de um
de seus ramos com uma nova hospedeira e a formação do primeiro haustório, tanto os
ramos laterais quanto os ramos vizinhos alteravam significativamente suas trajetórias de
crescimento, indo de encontro à nova fonte de recursos (t=8,788; n=30; p<0,0001). Os
ramos parecem receber estímulos que modificam o direcionamento de suas trajetórias
visando a captura de recursos.
Palavras-chave: busca por recursos, dispersão Loranthaceae, parasitismo.

15

16
Introdução
Aspectos importantes em estudos populacionais como recrutamento,
sobrevivência e reprodução dos organismos possibilitam compreender o destino de uma
espécie na comunidade (Crawley 2007). Estas informações se tornam ainda mais
completas se os mecanismos de dispersão dos indivíduos forem estudados. A dispersão
é fundamental para os organismos e é particularmente difícil para as plantas parasitas.
Esta dificuldade não se resume a apenas atrair o agente dispersor, que em muitos casos
são aves (Barlow 1983), mas também suas sementes precisam ser depositadas e fixadas
em galhos de hospedeiras compatíveis (Ladley & Kelly 1996). A compatibilidade existe
se as hospedeiras não apresentarem defesas efetivas contra os processos de fixação,
germinação e a penetração do haustório, que são raízes especializadas em captar
nutrientes (Yan 1993; Roxburgh & Nicolson 2005; Calvin & Wilson 2006).
As plantas parasitas, cujos haustórios são do tipo epicorticais, podem formar
clones por meio da fragmentação quando dispersam vegetativamente (Calvin & Wilson
2006). A dispersão vegetativa tem como finalidade a colonização de novas áreas, como
em plantas aquáticas cujos rametes se destacam (Grace 1993) ou a captura de recursos
através do forrageamento (de Kroon et al. 2009). Para muitos pesquisadores o
forrageamento é de fundamental importância para compreender as interações de uma
comunidade, pois pode determinar a dinâmica da competição entre as espécies (Pianka
1981), as relações entre predadores e suas presas (Fryxell & Lundberg 1994, Krivan &
Sikder 1999), bem como as interações indiretas entre as espécies (Wootton 1992).
O forrageamento em plantas tem sido discutido amplamente por diversos autores
(de Kroon & Schieving 1990; Karban 2008; Kembel et al. 2008; de Kroon et al, 2009;
McNickle et al. 2009). Segundo de Kroon e colaboradores (2009) o forrageamento em
plantas pode ser definido como plasticidade fisiológica ou alterações morfológicas que
direta ou indiretamente melhoram a captura de recursos essenciais. Em ambientes
heterogêneos as folhas e raízes das plantas são posicionadas de maneira não aleatória,
visando captar ativamente os nutrientes essenciais, a água e a luz (Kaban 2008). Esse
comportamento remete à teoria de forrageamento ótimo, em que os organismos
procuram maximizar a absorção de energia por unidade de tempo. Isso significa que
estes organismos devem gastar o mínimo de energia para encontrar, capturar, manipular
e consumir uma presa que possa maximizar a eficiência da alimentação. Para alguns
autores como Runyon e colaboradores (2006) e Kaban (2008) o forrageamento de

17
plantas parasitas seria semelhante ao de animais, pois enquanto elas localizam suas
fontes de alimento através do reconhecimento de compostos químicos liberados por
hospedeiras, os animais utilizam a visão para capturar suas presas. Os animais podem
assumir comportamentos distintos ora explorando ativamente um habitat em busca de
recursos, ora assumindo um comportamento mais passivo de senta e espera;
analogamente as plantas podem apresentar diferentes padrões de comportamento sob
diferentes condições ambientais devido a sua plasticidade morfológica (de Kroon &
Hutchings 1995).
Entretanto, os padrões de forrageamento em plantas parecem ser mais
complexos que em animais (de Kroon & Schieving 1990). Os animais podem se
deslocar de um local com baixa para outro com alta disponibilidade de recursos,
gastando certa quantidade energia (Norberg 1997). As plantas, por outro lado, precisam
investir em crescimento para alongar suas partes vegetativas na tentativa de escapar de
um local pobre em recursos, tais como nutrientes, água e luz (Kroon & Schieving 1990;
McNickle et al. 2009). As plantas parasitas, por exemplo, podem responder rapidamente
a mudanças na qualidade da hospedeira (Calvin & Wilson 2006), alterando seus padrões
de crescimento quando atacam preferencialmente algumas espécies de plantas na
tentativa de aumentar a eficiência na aquisição de recursos (Salzman 1985; Slade &
Hutchings 1987; Kelly 1990). A Convolvulaceae Cuscuta subinclusa emite ramos
exploratórios que se enrolam nos galhos das hospedeiras com a finalidade de reconhecê-
las antes de investir na formação de haustórios (Kelly 1990). Além disso, foi
comprovado experimentalmente que esta mesma parasita produziu mais haustórios em
hospedeiras que ofereciam melhores condições nutricionais (Kelly 1990).
Apesar do que foi exposto acima, os padrões de forrageamento de plantas
parasitas são pouco estudados e pouco se sabe sobre os mecanismos envolvidos na
localização e seleção de hospedeiras (Kelly 1992; Runyon et al. 2006). Como as plantas
parasitas dependem dos recursos de outras plantas para sua própria sobrevivência
(Runyon et al. 2006), podem afetar negativamente processos fisiológicos, reprodutivos
e ecológicos das hospedeiras, causando grande impacto na comunidade (Tennakoon &
Pate 1996, Press et al. 1999, Lei 1999, 2001, Press & Phoenix 2005, Shen et al. 2006).
Desta maneira, compreender seus padrões de forrageamento significa ter a capacidade
de prever a estratégia de busca por recursos, como também determinar possíveis efeitos
do parasitismo sobre a comunidade vegetal.

18
O objetivo desse trabalho foi determinar os possíveis fatores que influenciam a
propagação vegetativa Struthanthus flexicaulis na comunidade vegetal e verificar se esta
parasita apresenta forrageamento direcionado. Para isso as seguintes hipóteses foram
testadas: 1) A dispersão da parasita na paisagem depende da densidade de potenciais
hospedeiras e seus tamanhos; 2) O crescimento de ramos de S. flexicaulis é afetado pelo
encontro com uma nova fonte de recursos. As predições para estas hipóteses foram
respectivamente: 1) S. flexicaulis se espalhará mais rapidamente na comunidade vegetal
quanto maior a densidade e altura de potenciais hospedeiras e menor a distância entre
suas copas; 2) os ramos da parasita mudarão sua trajetória em busca de novas fontes de
recursos quando um dos ramos formar haustórios em outras hospedeiras. De acordo
com Campbell et al. (1991) o forrageamento em plantas pode ser definido como a
habilidade das espécies em proliferar suas raízes em manchas de solo mais ricas
nutricionalmente. Este conceito foi tomado como base para avaliar e discutir o
forrageamento de S. flexicaulis através do crescimento direcionado de seus ramos.
Metodologia
Local de estudo
O estudo foi desenvolvido numa área protegida do Parque Estadual da Serra do
Rola Moça, localizada na porção sul da Cadeia do Espinhaço- Brasil, uma longa cadeia
de montanhas paralela ao oceano Atlântico. Esta área é composta por afloramentos
ferruginosos e está localizada no Quadrilátero Ferrífero (20° 03’ 60”S, 44° 02’ 00” W,
1300 m). O clima da região é mesotérmico, do tipo Cwa de acordo com a classificação
de Köppen (Nimer & Brandão 1989). A precipitação anual é de 1000 a 1500 mm, com
uma pronunciada estação seca de abril a setembro, e temperatura média anual de 25°C
(Rizzini 1997). Os ventos constantes e altos índices de radiação ultravioleta contribuem
para a severidade do ambiente, agravada durante a estação seca. A vegetação é baixa,
composta por poucos indivíduos de porte arbóreo, dominada por eudicotiledôneas como
Asteraceae, Fabaceae e Myrtaceae, e monocotiledôneas como Poaceae, Cyperaceae e
Orchidaceae (Viana & Lombardi 2007, Jacobi et al. 2007). Na área de estudos M.
calodendron (Fabaceae) é uma das espécies arbustivas mais abundante (Jacobi et al.
2007) e frequentemente parasitada por Sthuthanthus flexicaulis (Mourão et al. 2009).
Espécie-alvo

19
Struthanthus flexicaulis Mart. (Loranthaceae) é uma das parasitas brasileiras
mais comuns, com ampla distribuição nas áreas de cerrado do Brasil Central (Rizzini
1980). A espécie generalista e na área de estudos pode parasitar 44 espécies
pertencentes a 19 famílias na área de estudos (Mourão et al. 2006). Possui ramos longos
e flageliformes. Cada ramo pode dar origem a outros laterais de até segunda ordem.
Estes ramos laterais podem algumas vezes crescer mais que o ramo principal. Esta
espécie possui folhas oblongo-abovadas, é dióica sendo composta por ramos com flores
pequenas (3-4 mm) exclusivamente masculinas e femininas e frutos pequenos (6-7
mm). O pico de frutificação ocorre ao final da estação chuvosa entre os meses de março
e abril (F. A. Mourão, obs. pess.). Os frutos são muito consumidos pela guaracava-de-
topete-uniforme Elaenia cristata (Tyrannidae), seu principal dispersor em algumas
áreas da Serra do Espinhaço (Guerra 2005).
Métodos
Foram demarcadas duas áreas mediram 900 m2
(30 x 30m) e diferiram quanto à
densidade de M. calodendron caracterizada pela distância entre copas (N = 281; Z = -
8,505; p < 0,0001). Nestas duas áreas foram realizadas observações do efeito da
densidade sobre a propagação vegetativa das parasitas. Outra área com
aproximadamente 1400m2 (40 x 35m) foi marcada aleatoriamente para observar a
influência da distância de hospedeiras no alcance dos ramos e o forrageamento da
parasita. As observações do efeito da distância e forrageamento foram realizadas
durante quatro dias na estação chuvosa, e as do efeito da densidade foram realizadas
durante dois dias na estação seca. Neste estudo, a referência às hospedeiras remete
exclusivamente a M. calodendron.
Efeito da densidade sobre a propagação vegetativa da parasita
Para avaliar o efeito da densidade de plantas na propagação vegetativa de ramos
(contágio), foram selecionadas duas áreas: A com alta densidade de M. calodendron e B
com baixa densidade. Nessas áreas, foram selecionados visualmente indivíduos de M.
calodendron cujas copas estavam totalmente cobertas pelos ramos da hemiparasita.
Como estes dados foram coletados durante a estação seca e estas hospedeiras
apresentavam sinais nítidos de perda de folhas, foi assumido que os ramos da parasita
teriam necessariamente que forragear até encontrar uma nova hospedeira para garantir
sua sobrevivência. Em cada indivíduo, os ramos foram acompanhados para verificar se

20
haviam se estabelecido (formação de haustórios) em novos indivíduos de M.
calodendron. Em caso afirmativo o número de conexões consecutivas (passagens) era
contado. Nas duas áreas, o número de passagens consecutivas de ramos foi comparado
pelo teste do Qui-quadrado.
Efeito da distância, sucesso de propagação vegetativa e forrageamento dos ramos da
parasita
Para avaliar a influência da distância entre hospedeiras nas passagens dos ramos
da parasita foram selecionados 28 grupos totalizando 259 indivíduos de M.
calodendron. Em cada agrupamento foram identificados os indivíduos infectados por S.
flexicaulis, cujos ramos poderiam partir das copas e parasitar outras hospedeiras. Estes
indivíduos de M. calodendron parasitados foram chamados de “fonte de ramos”. A
partir dos indivíduos fonte foram tomadas as seguintes medidas: distância entre as
bordas das vizinhas mais próximas e sucesso de propagação vegetativa de ramos
(formação de haustórios). O número de passagens bem sucedidas foi contado e
distribuído em 12 classes de distâncias que variaram de 10 em 10 cm. Foi feita uma
regressão linear após a transformação logarítmica para ambos os eixos.
Para avaliar a probabilidade de contágio de novas plantas foram selecionadas 30
M. calodendron parasitadas com alturas que variaram de 60 a 90 centímetros, pois a
altura média dessas hospedeiras na área de estudos é de 50 cm. Nessas mimosas, a
altura da saída dos ramos da parasita era medida, bem como a distância que estes ramos
percorriam até tocar o substrato. Essas medidas foram feitas em três pontos: altura
inicial (medida logo após a saída do ramo e onde a distância=0); altura média (onde a
metade da distância foi percorrida pelo ramo) e altura final (altura=0 e a distância é a
máxima percorrida pelo ramo). Curvas exponenciais negativas foram ajustadas aos
pontos de altura máxima e mínima relacionando as alturas às distâncias. As equações
dessas curvas permitem determinar uma região onde qualquer copa dentro dos limites
máximos e mínimos das curvas poderá ser parasitada, pois elas descrevem dispersão de
ramos saindo das alturas máximas e mínimas. Essas equações foram utilizadas para
determinar a probabilidade de contágio de outras mimosas situadas a diferentes
distâncias da hospedeira de onde partem ramos da parasita, através da relação:
(Y(X)máx. – Y(X) min)/79, onde Y(X) máx. = altura dos ramos mais altos; Y(X) min =
altura dos ramos mais baixos e 79 é a altura máxima prevista, pelo modelo de regressão,
para um ramo ao sair da copa das hospedeiras de 90 cm de altura. Um gráfico de

21
probabilidade foi calculado considerando variações de distância de 0 a 100 cm
(intervalos de 5 em 5 cm de distância).
Para avaliar forrageamento dos ramos foram selecionados outros 30 indivíduos de
M. calodendron parasitados. No centro de suas copas foi colocado um octógono (Fig. 1)
dividido em oito setores numerados, onde cada um correspondia a um ângulo de 45
graus. Em todas as copas o setor 1 foi voltado para o norte. Todos os ramos que saíam
das copas eram contados e separados por setor. Foi verificado se estes ramos passavam
para outras hospedeiras quando houvesse a formação de haustórios. O programa Systat
foi usado para representar graficamente a densidade de ramos por setores de 45 graus.
A concentração de ramos associados a setores onde a passagem de ramos foi bem
sucedidas foi comparada, por teste t pareado, com a concentração nos demais setores.
Para verificar se outras variáveis, tais como a posição do sol ou direção dos ventos da
região, poderiam interferir no direcionamento, comparou-se a saída do número de ramos
em cada setor por meio de análise de variância (ANOVA) cujas variáveis foram: setores
(posicionamento) e número de ramos.
Fig. 1- Octógono colocado sobre a copa das hospedeiras para quantificar a saída de
ramos da parasita. Cada parte colorida (setor) corresponde a um ângulo de 45 graus. As
setas brancas indicam os ramos fora das copas (exploratórios).
Resultados
A densidade de mimosas intensificou a transmissão de ramos (Fig. 2). A área de
maior densidade de hospedeiras (2,1 indivíduos/m2; N=201) apresentou uma distância

22
de borda média entre vizinhas mais próximas de 27,8 e desvio padrão de ± 38,2 cm e a
mediana foi de 5 cm (n=201). Já na área de menor densidade (0,8 indivíduos/m2; N=81)
essa distância foi de 140,7 e desvio padrão de ± 123,7 cm e a mediana de 131 cm
(n=81). A propagação vegetativa de ramos para outras hospedeiras ocorreu mais
frequentemente na área com maior densidade de mimosas sendo que 89% dos ramos da
parasita tiveram sucesso em encontrar uma nova planta e formar haustórios. Nessa área
a propagação para duas ou três hospedeiras consecutivas foi mais comum, totalizando
mais de 30% das observações. Em contrapartida, na área menos densa apenas 21%
conseguiram explorar novas copas e somente cerca de 5% dos ramos passavam para
mais de uma hospedeira. Houve diferença significativa no número de passagens de
ramos entre duas áreas (χ2=226,5; gl=1; p<0,001).
Fig. 2 – Influência da densidade de hospedeiras na propagação vegetativa consecutiva
de ramos. Área de maior densidade (distância média entre bordas de hospedeiras
vizinhas = 27,8 cm) e Área de menor densidade (distância média entre bordas de
hospedeiras vizinhas = 140 cm).
A propagação vegetativa de ramos variou negativamente em função da distância
entre as bordas das copas das hospedeiras. Dos 265 ramos presentes nos agrupamentos
de mimosas, 171 tiveram sucesso na propagação vegetativa para outras hospedeiras. As
distâncias entre bordas variaram de 0 a 121 cm. O número de propagação se manteve
alto quando as distâncias eram inferiores a 20 cm e diminuiu significativamente a partir
deste valor (r2=0,700; n=12; p=0,001) (Fig. 3).

23
20 40 60 80100
120140
Log Distância (cm)
20
40
6080
100
Lo
g P
rop
ag
ação
veg
eta
tiv
a b
em
su
ced
ida
Fig. 3- Relação entre a distância entre bordas das copas das mimosas e propagação
vegetativa de ramos da parasita. Esta relação é descrita pela equação da reta Y= 6,684 –
1,182 X.
A probabilidade de propagação vegetativa de ramos variou conforme a altura de
sua saída das copas das hospedeiras. As saídas de ramos das alturas máximas e mínimas
delimitaram uma região onde qualquer copa situada dentro destes limites tem chance de
ser parasitada (Fig. 4). Nesse modelo, a altura máxima de saída dos ramos das copas das
hospedeiras foi de 79 cm e a mínima foi de 36 cm. As curvas exponenciais negativas
foram descritas em seu limite superior pela equação Y = ln (4,371 – 0,065 * Distância)
e por Y = ln (3,585 – 0,059 * Distância), em seu limite inferior (Fig. 4). Estas equações
são parecidas, refletindo elasticidade semelhante dos ramos. A altura das hospedeiras
influenciou a probabilidade de propagação vegetativa de ramos, pois plantas que se
encontravam muito próximas das hospedeiras (distâncias entre bordas das copas
menores que 10 cm) a probabilidade de ser atingida variou de 36 a 70%. Estas
probabilidades caíram bastante quando a distância entre copas foi superior a 20 cm
variando entre 18 e 0,07%.

24
0 10 20 30 40 50 60 70
Distância (cm)
0
20
40
60
80
Alt
ura
(cm
)
0 10 20 30 40 50 60 700
20
40
60
80
0 20 40 60 80 100 120
Distância (cm)
0
10
20
30
40
50
60
70
80
Pro
bab
ilid
ad
e d
e p
ass
ag
em
Fig. 4 – Sucesso de propagação vegetativa de ramos. Em A estão representadas as
curvas descritas pelas equações que relacionam altura dos ramos com as distancias as
bordas da copa e em B a probabilidade de contágio de outras plantas com copas situadas
a diferentes alturas e em diferentes distancias.
As observações na área de 1400m2 constataram que
os ramos apresentaram
forrageamento direcionado, pois após o encontro de um ramo com uma nova hospedeira
e a formação do primeiro haustório, tanto os ramos laterais quanto os vizinhos
alteravam suas trajetórias de crescimento indo de encontro à nova fonte de recursos.
A
B

25
Este crescimento não teve nenhuma relação com outras variáveis tais como a posição do
sol ou direção dos ventos da região, uma vez que não houve diferenças significativas
entre os setores em relação ao número de ramos que saíam das copas das hospedeiras
(F=0,8941, gl=72, p=0,52). O estímulo parece ocorrer após a formação do haustório
(t=8,788; n=30; p<0,0001) (Fig. 5). Ramos principais conectados às novas hospedeiras
desenvolviam vários ramos laterais para o setor onde um dos ramos tinha feito a
primeira conexão. O mesmo direcionamento ocorreu com parte dos ramos vizinhos, que
parecem receber o mesmo estímulo e mudaram a trajetória de seu crescimento em
direção à nova hospedeira. É importante ressaltar que a maioria dos vetores menores,
que representam menor número de ramos no setor, indicaram casos em que não houve a
formação de haustório (H) (Fig. 5A e B). Entretanto, todos os maiores vetores,
representando o maior número de ramos no setor, tiveram a formação de pelo menos um
haustório (H) (Fig. 5A e B).
Fig. 5A - Crescimento direcionado dos ramos quando há a formação do primeiro
haustório (Teste t=8,788; n=30; p<0,0001).

26
Fig. 5B – Detalhe do crescimento direcionado dos ramos quando há a formação do
primeiro haustório (representado pela letra H). As letras H corresponderam aos setores
onde havia pelo menos uma passagem de ramos (formação de haustório em uma nova
hospedeira).

27
Discussão
Enquanto a deposição de sementes de plantas parasitas depende quase que
exclusivamente do comportamento e das escolhas por poleiros pelas aves dispersoras
(Aukema 2003) e está correlacionada à frequência de plantas na paisagem (del Rio et al.
1996), a propagação vegetativa é favorecida pela densidade de hospedeiras (Mourão
2007, Mourão et al. 2009). Isso foi corroborado neste trabalho uma vez que a
propagação vegetativa de ramos de mimosas parasitadas para vizinhas foi favorecida
pela proximidade entre suas copas. Na área de maior densidade, quase todas as parasitas
conseguiram se estender para outras plantas quando suas hospedeiras apresentavam
nítidos sinais de caducifolia, garantindo assim a sua sobrevivência. Além da distância
entre bordas, a altura das hospedeiras influenciou indiretamente as chances de contágio
por ramos da parasita. Os ramos que saiam de copas mais altas percorriam um caminho
mais longo até tocar o substrato influenciando o sucesso de sua propagação para outras
plantas. No entanto, na área de menor densidade de M. calodendron a propagação de
ramos ficou comprometida, e apenas uma pequena fração da população de parasitas
conseguiu atingir outras hospedeiras. Como a distância dificulta a propagação dos
ramos, os mal sucedidos tendem a permanecer na mesma planta e devem morrer junto
com suas hospedeiras. Em paisagens com baixas densidades de plantas, as copas estão
mais distantes e a forma de propagação mais eficaz seria a dispersão ornitocórica.
Hospedeiras mais altas são as preferidas das aves, pois estas são as mais atrativas como
poleiro (Aukema & Del Rio 2002).
O efeito combinado da distância e altura de hospedeiras foi determinante para o
sucesso de propagação da parasita. A emissão de ramos de S. flexicaulis desde alturas
maiores das copas das hospedeiras permite percorrer uma maior distância antes de tocar
no chão, favorecendo a propagação vegetativa desses ramos para outras plantas. O
tamanho da hospedeira é um ótimo preditor da probabilidade de parasitismo de uma
planta (Overton 1994), pois hospedeiras maiores são mais atrativas para as aves e
também favoreceriam propagação vegetativa dos ramos para outras plantas. Além disso,
copas maiores a seriam alvos mais fáceis para os ramos de S. flexicaulis. Estes
resultados remetem ao modelo de metapopulação, em que um conjunto de sub-
populações locais interage através da dinâmica do fluxo de indivíduos (Hanski 1991,
Teodoro 2010). Assim como no estudo de Overton (1994), que comparou a dispersão da
parasita Phrygilanthus sonorae em árvores dos gêneros Bursera e Cytocarpa, e de

28
Teodoro (2010) que comparou a de Psittacanthus robustos em Vochysia thyrsoidea , a
propagação de S. flexicaulis pode ser equiparada à dinâmica de uma metapopulação.
Essas parasitas são compostas por sub-populações isoladas espacialmente em
fragmentos de habitat (hospedeiras) e estão unidas por fluxos biológicos, seja por
dispersão vegetativa ou ornitocórica. A dinâmica de ambas depende da distribuição
espacial de suas plantas hospedeiras que podem estar isoladas espacialmente numa
matriz de habitat desfavorável. O fluxo de plantas parasitas entre as hospedeiras se daria
em função da altura e distância entre suas copas (dispersão vegetativa) ou seus
tamanhos/altura (especialmente na dispersão ornitocórica, veja Guerra 2005). Quando
as hospedeiras morrem, morrem também as suas parasitas, a não ser que seus ramos
tenham atingido outras plantas vizinhas por dispersão vegetativa, ou suas sementes
tenham sido levadas a plantas mais distantes por aves. A paisagem pode ser vista como
uma coleção de manchas temporariamente favoráveis ao parasitismo (hospedeiras vivas,
parasitadas ou não) e de manchas favoráveis à propagação da parasita, dispersas numa
matriz de solo inóspito (Overton 1994).
Apesar de algumas plantas poderem se movimentar e crescer em resposta à luz ou
outros estímulos ambientais (Kaban 2008), os ramos de S. flexicaulis não apresentaram
este tipo de comportamento, pelo menos no ambiente aberto da canga onde luz não
parece ser limitante. Os ramos apresentaram um padrão aleatório de crescimento quando
não havia a formação de haustório em uma nova hospedeira. Eles saíam das copas,
geralmente em várias direções, indicando uma tentativa de cobrir uma maior área de
busca. Como as plantas têm crescimento modular, cada série de unidades repetitivas
(módulos) pode simultaneamente ter um comportamento de forrageamento diferente,
como proposto por McNickle et al. (2009) para raízes. Este modo de forrageamento foi
comparado ao dos insetos sociais, onde a seleção deve favorecer comportamentos que
aumentem a aptidão total da colônia, ou, no caso de plantas, de todo o indivíduo, e não
de cada unidade modular (McNickle et al. 2009). Diferentemente de outras plantas, que
precisam manter uma conexão entre as raízes e o resto, as parasitas podem se
multiplicar por fragmentação, após a formação dos haustórios em novas hospedeiras,
rompendo sua ligação com o indivíduo que o originou.
Após o encontro com uma nova hospedeira e a formação de haustórios, ocorre a
proliferação de novos ramos laterais também em direção à hospedeira, apresentando um
crescimento direcionado. Esse crescimento aumenta a eficiência na obtenção de
recursos, como previsto pela Teoria do Forrageamento Ótimo. Outros ramos vizinhos

29
parecem receber estímulos que modificam o direcionamento de suas trajetórias visando
a captura de recursos, crescendo para o mesmo local da formação dos haustórios. Como
os haustórios das plantas parasitas respondem às substâncias químicas liberadas por
outras espécies, permitindo o reconhecimento e ataque a hospedeiras apropriadas
(Kaban 2008), um mecanismo semelhante poderia ser responsável por emitir e captar
estímulos de reconhecimento de novas hospedeiras e causar mudanças de trajetória e/ou
a proliferação de novos ramos. Com base em evidências empíricas da literatura, de
Kroon et al. (2005) argumentam que os comportamentos que envolvem decisões sobre
frequência de ramificação de raízes, crescimento dos entrenós e aumento no eixos de
raízes, ocorrem em níveis modulares, e as respostas são induzidas por sinais que são
percebidos localmente. Da mesma forma, é possível que os ramos de parasita, S.
flexicaulis se comuniquem e influenciem o direcionamento de ramos vizinhos. Esta
seria uma estratégia eficiente que otimizaria a busca por recursos. No caso de plantas
parasitas o encontro com uma nova hospedeira significa usufruir de uma nova fonte de
recursos e com isso ampliar a capacidade da sua população crescer, se reproduzir e
colonizar novos ambientes.

30
Referências Bibliográficas
Aukema, J. E.; del Rio, C. M. 2002. Variation in mistlestoe seed deposition: effects of
intra- and interspecific host characteristics. Ecography, 25: 139-144.
Aukema, J. E. 2003. Vectors, viscin, and Viscaceae: mistletoes as parasites, mutualists
and resources. Frontiers in Ecology and the Environment, 1: 212-219.
Barlow, B. A. 1983. Biogeography of Loranthaceae and Viscaceae. In: Calder, M.;
Bernhardt, P. (Editors), The Biology of Mistletoes, pp. 19-46. Sydney: Academic
Press.
Calvin, L. C. & Wilson, C. A. 2006. Comparative morphology of epicortical roots in
Old and New World Loranthaceae with reference to root type, origin, patterns of
longitudinal extension and potential for clonal growth. Flora, 201: 51-64
Campbell, B. D.; Grime, J. P. & Mackey, J. M. L. 1991. A trade-off between scale and
precision in resources foraging. Oecologia 87:532-538.
Crawley, M. J. 2007. Plant Ecology, 2 º Ed. 717 p. Ed. Blackwell Science, London.
de Kroon, H. & Hutchings, M. J. 1995. Morphological plasticity in clonal plants: The
foraging concept reconsidered. Journal of Ecology, 83:143-152.
de Kroon, H., Schieving, F. 1990. Resource partitioning in relation to clonal growth
strategy. In: Clonal Growth in Plants (eds. Van Groenendael J, de Kroon H).
Academic Publishing, The Hague, 426 p.
de Kroon, H. & Schieving , F. 1991. Resource allocation patterns as a function of clonal
morphology: a general model applied to a foraging clonal plant. Journal of Ecology,
79:519-530.
de Kroon, H.; Huber, H.; Stuefer, J. F.; & van Groenemdael, J. M. 2005. A modular
concept of phenotypic plasticity in plants. The New Phytologist, 166: 73-82.
de Kroon, H.; Visser, E. J. W.; Huber, H.; Mommer, & Hutchings, M. J. 2009. A
modular concept of plant foraging behavior: the interplay between local responses
and systemic control. Plant Cell and Environment, 32: 704-712.
Donohue, K. 1995. The spatial demography of mistletoe parasitism on a Yemeni
Acacia. International Journal of Plant Sciences, 156:816-823.
Dzerefos, C. M. Witkowski, E. T. F. & Shackleton, C. M. 2003. Host-preference and
density of woodrose-forming mistletoes (Loranthaceae) on savanna vegetation, South
Africa. Plant Ecology. 167:163-177.

31
Fryxell, J. M. & Lundberg, P. 1994. Diet choice and predator-prey dynamics.
Evolutionary Ecology, 8: 407-421.
Grace, J. B. 1993. The adaptive significance of clonal reproduction in angiosperms: an
aquatic perspective. Aquatic Botany, 44: 159-180.
Journal of Ecology, 82: 833-842.
Guerra, T. J. A. 2005. Componentes quantitativos e qualitativos da dispersão de
sementes de Struthanthus flexicaulis (Loranthaceae) em uma área de campo
rupestre do sudeste brasileiro. Dissertação de mestrado. Universidade Estadual
Paulista, 64p.
Hanski, I. 1991. Single species metapopulation dynamics: concepts. Models and
observations. Biological Journal of the Linnean Society, 42: 17-38.
Harper, J. L. 1977. Population Biology of plants. Academic Press, London
Harper, J. L. 1985. Modules, branches, and the capture of resources. In: Population
biology and evolution of clonal organisms (J.B.C. Jackson, L.W. Buss & R. Cook,
eds). Yale University Press, New Haven, p.1-34.
Jacobi, C. M., F. F. Carmo, R. C. Vincent, and J. R. Stehmann. 2007. Plant communities
on ironstones outcrops: a diverse and endangered Brazilian ecosystem. Biodiversity
and Conservation, 16: 2185-2200.
Kaban, R., 2008. Plant behavior and communication. Ecology Letters, 11:727-739.
Kelly, K. C. 1990. Plant foraging: a marginal value model and coiling response in
Cuscuta subinclusa. Ecology, 71: 1916-1925.
Kelly, K. C. 1992. Resource choice in Cuscuta europaea. Proceedings of National
Academy of Sciences, 89: 12194-12197.
Kembel SW, de Kroon H, Cahill JF Jr, Mommer L. 2008. Improving the scale and
precision of hypotheses to explain root foraging ability. Annals of Botany, 101:
1295–1301.
Krivan, V. & Sikder, A. 1999. Optimal foraging and predator-prey dynamics II.
Theoretical Population Biology, 55: 111-126.
Ladley, J. J. & Kelly, D. 1996. Dispersal, germination and survival of New Zealand
mistletoes (Loranthaceae): Dependence on birds. New Zealand Journal of Ecology,
20: 69-79.
Lei, S. A. 1999. Age, size and water status of Acacia gregii influencing the infection
and reproductive success of Phoradendron californicum. American Midland
Naturalist, 141: 358-365.

32
Lei, S. A. 2001. Survival and development of Phoradendron californicum and Acacia
greggii during a drought. Western North American Naturalist, 61: 78-84.
McNickle, G. G.; St. Clair, C. C & Cahill Jr, J. F. 2009. Focusing the metaphor: plant
root foraging behavior. Trends in Ecology and Evolution, 24:419-426.
Mourão, F. A.; Carmo, F. F; Ratton, P.; Jacobi, C. M. 2006. Hospedeiras da
hemiparasita Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) em campos
rupestres ferruginosos, Quadrilátero Ferrífero, MG. Lundiana, 7:103-110.
Mourão, F.A. 2007. Padrão de parasitismo de Struthanthus flexicaulis (Mart.) Mart.
(Loranthaceae) e seus efeitos na comunidade vegetal de um campo rupestre sobre
canga do Quadrilátero Ferrífero (MG). Dissertação de mestrado, Universidade
Federal de Minas Gerais, Belo Horizonte. 64pp.
Mourão, F. A., Jacobi, C. M. Figueira, J. E. C & Batista, E. K. L. 2009. Effects of the
parasitism of Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) on the fitness of
Mimosa calodendron Mart. (Fabaceae), an endemic shrub from rupestrian fields over
ironstone outcrops, Minas Gerais State, Brazil. Acta botanica brasilica 23: 820-825.
Nimer, E. & Brandão, A. M.. 1989. Balanço Hídrico e clima da região dos Cerrados.
Instituto Brasileiro de Geografia e Estatística (IBGE), Rio de Janeiro, Brazil.
Norberg, R. A. 1997. Na ecological theory on foraging time and energetics and choice
of optimal food-searching method. Journal of Animal Ecology 46:511-529.
Norton, D. A. & Carpenter, M. A. 1998. Mistletoes parasites: host specificity and
speciation. Trends in Ecology and Evolution, 13: 101-105.
Overton, J.M. 1994. Dispersal and infection in mistletoe metapopulations. Journal of
Ecology, 82: 711-723.
Pianka, E. R. 1981. Competition and niche theory. In: Theorical Ecology. 2ed p. 167-
196. Oxford: Blackwell Scientific.
Press, M. C. & Phoenix, G. K. 2005. Impacts of parasitic plants on natural communities.
New Phytologist, 166: 737-751.
Press M. C.; Scholes, J. D. & Watling, J. R. 1999. Parasitic plants: physiological e
ecological interactions whit their hosts in Press, M. C.; Scholes, J. D. & Barker M.
G.; (eds.) Physiological plant Ecology. Blackwell Scientific Oxford, 175-197 pp.
Rizzini, C. T. 1980. Loranthaceae of the central Brazil. Arquivos do Jardim Botânico do
Rio de Janeiro, 24: 19-50.
Rizzini, C. T. 1997. Tratado de Fitogeografia do Brasil: Aspectos Ecológicos,
Sociológicos e Florísticos. 2 º ed. Âmbito Cultural Edições Ltda, Brazil.

33
Roxburgh, L. & Nicolson, S.W. 2005. Patterns of host use in two African mistletoes:
the importance of mistletoe-host compatibility and avian disperser behaviour.
Functional Ecology, 19: 865-873.
Runyon, J. B. Mescher, M. C. & De Moraes, C. M. 2006. Volatile chemical cues guide
host location and host selection by parasitic plants. Science, 313: 1964-1967.
Salzman, A. G. 1985. Habitat selection in a clonal plants. Science, 228: 603-604.
Slade, A. J. & Hutchings, M. J. 1987. The effects of light intensity on foraging in the
clonal herb Glechoma hederacea. Journal of Ecology, 75: 639-650.
Shen, H.; Ye, W.; Hong, L.; Huang, H.; Wang, Z.; Deng, X.; Yang, Q.; Xu, Z. 2006.
Progress in parasitic plant biology: Host selection and nutrient transfer. Plant
Biology, 8: 175-185.
Tennakoon, K. U. & Pate, J. S. 1996. Effects of parasitism by a mistletoe on the
structure and functioning of branches of its host. Plant, Cell and Environment,
19:517-528.
Teodoro, G. S. 2010. Estrutura e dinâmica metapopulacional da erva-de-passarinho
Psittacamthus robustos Mart. (Loranthaceae) em manchas de habitat em três áreas de
Cerrado. Dissertação de mestrado. Universidade Federal de Lavras, 85 p.
Viana, P. L. & Lombardi, J. A. 2007. Florística e caracterização dos campos rupestres
sobre a canga na Serra da Calçada, Minas Gerais, Brasil. Rodriguésia, 58: 159-177.
Wootton, J. T. 1992. Indirect effects, prey susceptibility, and habitat selection: impacts
of birds on limpets and algae. Ecology, 73:981-991.
Yan, Z. G. 1993. Low water potentials as a deterrent to mistletoes. Haustorium, 27:2.

34
Anexo 1 – Diferentes fenofases de Struthanthus flexicaulis (Mart.) Mart.
(Loranthaceae)
Retirado de Mourão 2007.
Fig.1- Struthanthus flexicaulis (Mart.) Mart. A) Ramos longos e flageliformes com folhas verdes
oblongo-abovaladas de tamanhos variados; B) Botões e flores; C) Frutos; D) Germinação da
semente; E) Apressórios (indicados pela setas). Escala: Barra = 1cm.
A
C B
E D

34
CAPÍTULO 2
Modelagem espacial do espalhamento da planta
parasita Struthanthus flexicaulis (Loranthaceae) em
paisagens de afloramentos ferruginosos, Brasil.
Fabiana Alves Mourão1, Júlio César Mansur Filho
2, Claudia Maria Jacobi
1, José
Eugênio Côrtes Figueira1 e Ronald Dickman
2
1 Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade
Federal de Minas Gerais.
2 Departamento de Física, Instituto de Ciências Exatas, Universidade Federal de Minas
Gerais.

35
Resumo
A utilização de modelos matemáticos em estudos ecológicos auxilia na
compreensão dos padrões de dispersão de organismos na paisagem. Modelos de
simulação de espalhamento de plantas parasitas já têm sido propostos, principalmente
para avaliar a dispersão ornitocórica. Este estudo se propõe utilizar do modelo de Gás
em Rede na simulação da dinâmica do espalhamento de Struthanthus flexicaulis em
uma paisagem virtual com características semelhantes à da área de estudos. A dinâmica
da propagação da parasita foi modelada em duas etapas: 1) definição da paisagem para
seu espalhamento e 2) escolha dos parâmetros do modelo. Na construção da paisagem
foi considerado apenas um tipo de hospedeira. A paisagem gerada manteve as
características de densidades e distribuição espacial de hospedeiras semelhante às que
foram determinadas em campo. Os parâmetros avaliados foram a velocidade de
propagação da parasita e o número de hospedeiras infectadas após a saturação. Os
parâmetros demográficos da população de hospedeiras foram desconsiderados. A
dispersão ornitocórica foi a principal maneira de infestação na paisagem durante os
primeiros 12 meses. As áreas de maiores agrupamentos de hospedeiras foram as que
mais recebem ramos através da dispersão vegetativa. Ao final de 192 meses, mimosas
isoladas raramente foram parasitadas. Os resultados desta simulação demonstram a
importância da agregação de hospedeiras no espalhamento da parasita, sendo a
dispersão ornitocórica fundamental para acelerar o processo de espalhamento na
paisagem simulada.
Palavras-chave: autômatos celulares, densidade de hospedeiras, dispersão vegetativa e
ornitocórica, Gás em Rede, parasita.

36
Introdução
Os padrões de dispersão de organismos na paisagem possibilitam compreender os
aspectos ecológicos e evolutivos de uma espécie e estão relacionados à distribuição,
dinâmica e interações com outros organismos (Kareiva 1994). Esses padrões podem ser
modelados matematicamente através de duas abordagens: paisagens espacialmente
implícitas ou explícitas. Em uma abordagem espacialmente implícita considera-se que
todas as manchas (fragmentos de habitat) na paisagem são iguais e igualmente
acessíveis para todos os organismos, como no modelo de metapopulações inicialmente
proposto por Richard Levins (Levins 1970, Hanski & Simberloff 1997). Já na
abordagem espacialmente explícita, as manchas são localizadas em coordenadas
geográficas fixas permitindo a avaliação das distâncias de dispersão dos organismos e a
relação entre populações dentro de manchas de habitat e outras características da
paisagem (Dunning et al. 1995). Um exemplo dessa abordagem são os modelos
baseados em indivíduos (MBI) (Huston et al. 1988) que permitem acompanhar o
destino de cada indivíduo de uma população separadamente (Caswell & Etter 1993,
Mistro & Jacobi 1999, Cumming 2002; Cannas et al. 2003) e por isso adquirem um
grau de realismo maior, podendo incorporar características biológicas distintas
associadas à idade, tamanho ou estágio de desenvolvimento de cada indivíduo na
população.
Modelos de simulação de espalhamento de plantas parasitas em que a paisagem é
um mapa de diferentes hospedeiras já têm sido propostos, mas principalmente para
caracterizar o comportamento das aves que dispersam suas sementes (Overton 1996;
Vallauri 1998; Lavorel et al. 1999). Os modelos que consideram populações
estruturadas espacialmente ajudaram a entender mecanismos envolvidos na dispersão,
por aves, das sementes das Loranthaceae Phrygilanthus sonorae (Overton 1994) e
Amyema preissii (Lavorel et al. 1999) entre as árvores de uma floresta real cujos dados
coletados em campo foram utilizados para compor uma paisagem simulada. As espécies
vegetais não estão presentes no ambiente apenas em decorrência da qualidade do
habitat-matriz, mas sim devido a uma distribuição diferenciada de recursos, o que
resulta em aproveitamentos diferentes da paisagem. No estudo de estrutura de
metacomunidades em um ecossistema aquático, Cottenie e de Meester (2004)
destacaram a qualidade dos recursos como o principal fator influenciando a distribuição

37
de fitoplâncton, para justificar o comportamento de dispersão de populações entre
fragmentos.
De forma semelhante aos resultados encontrados por Cottenie e de Meester
(2004), a comunidade vegetal de afloramentos ferruginosos tem sua distribuição
limitada pela escassez de água e nutrientes, temperaturas elevadas e reduzida camada de
solo, e deve enfrentar o efeito combinado desses e outros fatores estressantes (Silva et
al. 1996, Klein 2000, Rosière & Chemale 2000, Vilela et al. 2004). A fina camada de
solo, quando existente, é constituída de metais pesados e minério de ferro (Benites et al.
2007) e se acumula em fissuras e reentrâncias da rocha. A vegetação adere-se a essas
fissuras (onde a água pode ficar algum tempo retida na estação chuvosa, Silva 1991),
crescendo entre as fendas e em pequenos acúmulos de matéria orgânica (Barth 1987).
Como as fissuras na rocha aparecem irregularmente na paisagem, permitem a formação
de um mosaico onde eventualmente estão presentes ilhas de vegetação (Barth 1987). O
solo influencia a sobrevivência das plantas em cada ilha contribuindo para a
heterogeneidade fisionômica da comunidade (Vincent 2004).
A distribuição de plantas parasitas é menos afetada por condições edáficas,
hidrológicas e nutricionais, devido à sua capacidade de retirar água e nutrientes das
hospedeiras. Entretanto, suas hospedeiras podem sofrer variações demográficas em
larga escala espacial e temporal provenientes da disponibilidade diferencial de recursos,
limitando por sua vez a distribuição das parasitas (Ehleringer & Marshall, 1995). Ainda
há poucos estudos quantitativos sobre sua dinâmica populacional, que são
obrigatoriamente vinculadas à distribuição espacial das suas hospedeiras (Shaw et al.
2004, Teodoro 2010, Arruda & van der Berg dados não publicados). A ecologia das
plantas parasitas têm sido estudada em diferentes ecossistemas (Arruda et al. in press.),
focando especialmente a dispersão de sementes (Sutton 1951, Davidar 1978, Overton
1994, del Rio et al. 1996, López-de-Buen e Ornelas 1999, Aukema & del Rio 2002,
Amico et al. 2009) e a especificidade de parasitismo (Kuijt 1963, Barlow & Wiens
1977, Thomson & Mahal 1983, Hoffman et al. 1986, Mourão et al. 2006, Arruda et al.
2006, Fadini 2011), ou sua condição fisiológica (Dean et al. 1994, Bickford et al. 2005).
Muitos modelos propostos na Física têm sido adaptados e usados em sistemas
biológicos, pois têm relativa simplicidade e procuram explicar os fenômenos de forma
objetiva com elevado rigor matemático (Giacomini 2007). Um exemplo é o modelo de
Gás em Rede. Este modelo faz parte de uma família de modelos conhecidos como
Autômatos Celulares, que lida com difusão de partículas que se movem em uma rede

38
discreta. Cada partícula pode se propagar para sítios vizinhos, movendo-se através de
saltos de um sítio para outro, em um espaço de tempo determinado (Wolf 2002). O
modelo de Gás em Rede tem sido utilizado para investigar a dinâmica de predadores e
suas presas (Satulovsky & Tome 1994, Chen et al. 2006), e para estudar a mobilidade
de organismos (Rodrigues & Tomé 2008). Este modelo é capaz de incorporar padrões
espaciais de distribuição de espécies, bem como outras características importantes do
sistema estudado (Rodrigues & Tomé 2008), utilizando regras e condições iniciais
muito simples, característica que o torna bastante interessante para simular sistemas
mais complexos. O objetivo deste trabalho foi construir e utilizar o modelo de Gás em
Rede na simulação da dinâmica do espalhamento de S. flexicaulis em uma paisagem
virtual com características semelhantes à da área natural, comparando sua dispersão
ornitocórica e vegetativa. A principal hipótese é que a dispersão ornitocórica,
responsável por novos focos de parasitismo, potencializará a velocidade de
espalhamento da parasita, enquanto a dispersão vegetativa será responsável pela
colonização dos agrupamentos, levando à saturação do sistema.
Material e Métodos
A construção do modelo da dinâmica da propagação da parasita S. flexicaulis na
paisagem foi feita em duas etapas. A primeira consistiu em definir a paisagem e a
segunda em estabelecer os parâmetros do modelo de espalhamento. Na construção da
paisagem foi considerado apenas um tipo de hospedeira, Mimosa calodendron
(Fabaceae), que é mais abundante e mais parasitada na área de estudo (Mourão et al.
2006; 2009). Esta espécie é endêmica dos afloramentos ferruginosos do Quadrilátero
Ferrífero (Barneby 1991; Jacobi & Carmo 2012), apresenta porte arbustivo e suas
populações são agregadas, aparecendo como grandes manchas verdes na paisagem
(Mourão et al. 2009). Por se tratar de uma leguminosa, M. calodendron possivelmente
contribui para o enriquecimento de nitrogênio do solo dos afloramentos que são pobres
em nutrientes. Sua copa densa ajuda a reduzir as altas temperaturas do substrato e a
aumentar significativamente a formação de solo, através da incorporação de matéria
orgânica (Jacobi et al. 2007).
Construção da Paisagem para a simulação
Para criar a paisagem da simulação foi tirada uma fotografia aérea de uma área de
aproximadamente 8200 m2, que depois de impressa foi subdividida em 16 partes onde

39
sobre cada uma foi colocada sob uma rede quadrada (L x M) de 10 x 10 cm2 dividida
em intervalos de 0,5 cm (quadrículas)(Fig. 1A). Foram determinados: 1) o número total
de quadrículas, representando todos os sítios possíveis para a localização de hospedeiras
(N constante = 400 por que todas as partes possuíam tamanhos de rede iguais) e 2) em
cada parte, o número das quadrículas que foram preenchidas por 50% ou mais pelas
hospedeiras (mimosas). Estas últimas representam os sítios que efetivamente foram
encontradas hospedeiras e, portanto são favoráveis à presença da parasita. Para
determinar a densidade de M. calodendron (ρ), o número de sítios favoráveis foi
dividido pelo total de sítios possíveis (favoráveis e não favoráveis).
Para descobrir a fração de pares vizinhos (o grau de agregação das hospedeiras na
paisagem) calculou-se o r1,1. Este parâmetro descreve a probabilidade de se ter vizinhos
ocupados por hospedeiras (quadrícula esquerda e/ou superior conforme a figura 1B).
Figura 1 – Representação esquemática da obtenção dos valores de densidade de
hospedeiras e de pares de vizinhos em: A) Rede de 6 x 6 cm colocada sobre a foto aérea
mostrando a localização de hospedeiras. A barra lateral em amarelo corresponde a 1 cm
na figura e o centro de uma quadrícula à outra corresponde a 1 m em escala real. Esta
imagem foi quadriculada para estimar sua densidade e probabilidade de se encontrar
vizinhas; B) representação esquemática da estratégia utilizada para a contagem do
número de hospedeiras vizinhas sem repeti-las. Os sentidos das setas indicam como os
pares de vizinhas foram determinados.
Cada quadrícula pode ter zero, um ou dois vizinhos ocupados (sítios favoráveis).
Após analisar cada quadrícula e determinar o número de vizinhos existentes, calculou-se
o número de pares de vizinhos possíveis que é dado por 2LM – (L+M); sendo L =
número de quadrículas na vertical e M = número de quadrículas na horizontal. A fração
A B
Quadrícula
Quadrícula

40
de pares vizinhos (r1,1) foi calculado dividindo-se o número de vizinhos encontrados
pelo número de pares de vizinhos possíveis (Fig. 1).
Uma vez calculada a densidade de partículas (hospedeiras) e a ocorrência de pares
vizinhos, foi utilizado o modelo de difusão de partículas de Gás em Rede, que na sua
forma mais simples, possui o número de partículas (hospedeiras) fixo e as interações
somente ocorrem entre pares de vizinhos. A paisagem gerada manteve as características
de densidades e distribuição espacial de hospedeiras de forma semelhante às que foram
determinadas em campo. Neste modelo a interação entre partículas (hospedeiras) pode
ser descrita por potenciais repulsivos de curto alcance (repulsão de partículas) com a
energia escrita da seguinte forma:
Onde σ i,j = 1 ou 0 caso o sítio esteja ocupado ou vazio e σ k,l = posição de cada sítio.
Devido a esse tipo de interação, cada sítio (quadrícula) pode conter apenas uma
partícula (hospedeira) e o espaçamento de rede representa o raio da mesma. Na
paisagem real, o centro de uma quadrícula à outra corresponde a aproximadamente 1 m.
Para fins de ilustração pode-se imaginar que o centro de cada hospedeira está situado
nos vértices da rede (Fig. 2). O valor da energia do sistema (E) corresponderá à
paisagem real, pois leva em consideração a densidade de hospedeiras e a probabilidade
de formação de agrupamento (número de pares vizinhos). Um paralelo pode ser traçado
entre a energia de repulsão de partículas e o substrato dos afloramentos ferruginoso, em
que maior energia de repulsão (valor absoluto) significaria substratos mais compactos
(sem fissuras nas rochas e solos, quando existentes, pouco profundos). Como as
hospedeiras se estabelecem em fissuras ou fendas no substrato, intuitivamente áreas de
menor densidade de hospedeiras e menores probabilidades de formação de
agrupamentos devem apresentar maior energia de repulsão de partículas.
Figura 2- Ilustração da posição espacial das hospedeiras em uma rede quadrada.

41
A energia de repulsão é influenciada pela Temperatura (T), uma grandeza física
que mede a energia cinética de cada uma das partículas de um sistema em equilíbrio. É
possível variar a paisagem modificando apenas os valores de temperatura e mantendo os
de densidade de partículas (hospedeiras) e número de pares vizinhos (Fig. 3). Este
procedimento é necessário para determinar em que temperatura o modelo irá rodar as
simulações, já que a paisagem gerada deve se assemelhar à paisagem real. De acordo
com o comportamento de partículas de gás na Física quando a temperatura é alta, menor
agrupamento de partículas é esperado. Neste estudo isso corresponderia a regiões cada
vez menos apropriadas para a propagação vegetativa da parasita, já que as mimosas
estariam dispostas de maneira mais isolada na paisagem.
Foi usado o teste de Monte Carlo para simular a dinâmica de interação de
partículas, e a paisagem foi aceita quando a energia de repulsão de partículas foi menor
ou igual à energia gerada com os dados de campo. Para encontrar este parâmetro foram
usadas diferentes Temperaturas (T). A implementação do algoritmo computacional do
processo foi feito pelo método Markoviano, que é um processo estocástico e tem como
propriedade a falta de memória, sendo sua evolução de um tempo tn a tn+1 dependente
apenas do seu valor no presente.
Neste modelo as N partículas foram distribuídas aleatoriamente numa rede
quadrada L x L sítios com condições de contorno periódicos. Isso significa que as
partículas localizadas em uma das bordas da rede fazem contato com as outras do lado
oposto, de modo que todas as partículas tenham iguais condições de se tornarem
contaminadas e de transmitir propágulos da parasita. A configuração final da paisagem
foi dada após o tempo de relaxação (estabilização), quando o sistema chega a um estado
de equilíbrio de menor energia (Fig. 3).
Depois de fixado o valor da densidade, os valores de r1,1 correspondentes ao da
paisagem real foram estimados a partir da variação de temperatura para definir qual ou
quais deveriam ser usadas no modelo (Fig. 4). Os resultados da variação dos valores de
r1,1 estão dentro dos limites da variação encontrada para a paisagem real (Tabela 1). A
partir destes resultados definiu-se o tamanho da rede L= 100 que foi usada no modelo e
a temperatura de 0,53 e L=50 e temperatura de 0,525 já que apresentaram agrupamentos
de partículas melhor distribuídas na paisagem, semelhante à paisagem rea

42
Figura 3 – Paisagens geradas pelo modelo de Gás em Rede em uma rede quadrada L x L
sítios com L = 400. A densidade de hospedeiras foi ρ = 0,2. Os padrões para cada
temperatura foram gerados após 150.000 permutações do teste de Monte Carlo. As
variações de temperatura permitem modificar a agregação das partículas (hospedeiras),
sendo mais agregadas em temperaturas menores. Estes padrões podem ser visualizados
na sequencia acima, que parte de agregados compactados com bordas suaves até uma
total desagregação.

43
Tabela 1 – Valores de r1,1 calculados a partir da variação de temperatura e que se
encontra dentro do erro esperado para a paisagem real, para os tamanhos de rede L = 50,
L = 100, L = 200 e L = 400.
Figura 4 – Estimativa dos valores de probabilidade de formação de pares de vizinhos
(r1,1) com a variação de temperatura no modelo (T). A legenda representa o tamanho das
redes discretas utilizadas. Em A a área destacada mostra a variação dos valores de r 1,1
em função das temperaturas e em B tem-se uma ampliação desta área. Note que os
valores de r1,1 escolhidos são próximos ao encontrado para a paisagem real e sua
variação está de acordo com o erro encontrado.
Figura 4 – Estimativa dos valores de probabilidade de formação de pares de vizinhos
(r1,1) com a variação de temperatura no modelo (T). A legenda representa o tamanho das
redes discretas utilizadas. Na figura da esquerda a área destacada mostra a variação dos
valores de r 1,1 em função das temperaturas e à direita tem-se uma ampliação desta área.
Note que os valores de r1,1 escolhidos são próximos ao encontrado para a paisagem real e
sua variação está de acordo com o erro encontrado.
Parâmetros do modelo de propagação da parasita
Após a germinação, as plântulas de S. flexicaulis emitem ramos em forma de
flagelos, com pequenas folhas e fixados à planta hospedeira espaçadamente por
A B

44
haustórios, e gradualmente ocupam a copa da hospedeira. A partir das bordas desta copa,
os ramos de S. flexicaulis podem alcançar a copa de outra hospedeira, da mesma ou de
outra espécie, situada em diferentes distâncias com probabilidades que variam de 70 a
0% (Cap. 1 – figura 4B). Na área de estudo há variações notáveis na densidade de
plantas e, portanto, nas chances de crescimento de S. flexicaulis. A chance de passar de
uma hospedeira para outra depende do distanciamento de entre copas e quando a
distância é superior a 1m pode interromper localmente o crescimento vegetativo
(Mourão dados não publicados).
Algumas características dos afloramentos ferruginosos e da parasita
supostamente influenciam diretamente o processo de expansão deverão obrigatoriamente
compor o algoritmo que simulará a expansão de S. flexicaulis. Nesta versão preliminar
do modelo foram incluídas as seguintes características 1) agrupamento de hospedeiras;
2) desenvolvimentos de ramos exploratórios; 3) contágio de novas hospedeiras
dependente da distância; 4) morte dos ramos ocorrendo quando em contato direto com a
superfície do afloramento ferruginoso que, durante o dia, pode atingir temperaturas
bastante elevadas; 5) rochas nuas. Futuramente poderão ser incluídos no modelo outros
parâmetros tais como: 1) preferências da ave por local de alimentação (altura e
agrupamento das hospedeiras); 2) diferentes regras para o contágio de novas hospedeiras
que depende: da altura, diâmetro, distância e tipo de hospedeira; 3) modificações na
qualidade da matriz através do acréscimo de diferentes porcentagens de rochas nuas e
presença de outras plantas; 4) variações na densidade de hospedeiras; 5) morte,
nascimento e crescimento de hospedeiras.
Uma vez definida a paisagem, o modelo considera sítios saudáveis (Ns) quando
há ocorrência de mimosas não infectadas e sítios infectados (Ninf). Em cada iteração, os
sítios poderão ser infectados até os terceiros vizinhos (Fig. 5).
Figura 5 – Desenho esquemático do número de vizinhos e seu posicionamento na rede.

45
A simulação do espalhamento de S. flexicaulis num mosaico ambiental
representando a variedade de hospedeiras de tamanhos iguais e solo nu típicos da área
ocorre quando uma hospedeira é sorteada como fonte da parasita (dispersão
ornitocórica) e a partir dela ocorrerá a expansão de ramos vegetativamente, iniciando a
simulação. O tempo de cada iteração no modelo correspondeu a 1 mês e neste mesmo
tempo os ramos da parasita podem atingir um crescimento médio de 20 cm (F. A.
Mourão, obs. pess.).
A infecção de uma hospedeira pode ocorrer de duas maneiras: 1) propagação por
aves, que independe da distância de uma mimosa infectada e assumiu-se que a taxa de
dispersão ornitocórica seria de µ=0,005/mês e 2) propagação vegetativa, onde
assumindo-se que a taxa seria de λ = 0,1/mês. A taxa de espalhamento (dispersão
ornitocórica + vegetativa) foi dada por: R= 4 ctλ Ninf + µ Ns , onde ct= 1 + 1/√2 + ½;
Ninf = número de sítios infectados (hospedeiras infectadas) e Ns = número de sítios
saudáveis (mimosas que podem ser infectadas). Após a escolha e infecção das
hospedeiras (sítios saudáveis), os ramos das hospedeiras infectadas poderão atingir
outras plantas (Fig. 6). Caso parte de um ou mais ramos se posicione dentro da copa de
outra hospedeira a transmissão terá sido bem sucedida e o processo se reiniciará.
Figura 6- Esquema do funcionamento do modelo de propagação.

46
A expansão pode ser interrompida caso a hospedeira já esteja infectada ou os
ramos não se posicionem dentro de sítios considerados bons (copas de mimosas). Nesse
caso, será investigado como alterações na estrutura da paisagem poderiam afetar a
velocidade de espalhamento e o número de plantas virtuais parasitadas.
A cada unidade de tempo, a propagação de ramos pode ocorrer na horizontal,
vertical e diagonal, e poderá atingir primeiros, segundos e terceiros vizinhos com
probabilidades diferenciadas. Essas probabilidades de infecção são maiores para os
primeiros vizinhos, sendo 0,7 vezes menor para segundos e 2 vezes menor para
terceiros.
As regras para a simulação foram:
1) Escolher um dos quatro primeiros vizinhos do sítio infectado e se estiver saudável
infecta-lo;
2) Escolher um dos quatro segundos vizinhos e se estiver saudável, infectá-lo;
3) Escolher um dos quatro terceiros vizinhos e se estiver saudável, infectá-lo;
4) Caso exista um sítio saudável entre o sítio central infectado e seu terceiro vizinho,
infectá-lo também.
5) Atualizar o tempo.
O modelo guardará o número de sítios (hospedeiras) infectados a cada simulação
para os padrões gerados usando uma paisagem de L=100, e as taxas µ=0,005/mês e
λ=0,1/mês. As simulações foram feitas para tempo t = 12, 24, 36, 48, 60 e 192 meses.
No tempo t=0, ocorre o sorteio das quadrículas que serão infectadas por dispersão
ornitocórica (deposição de sementes), numa taxa de µ* Ns. Em seguida, os ramos da
parasita começam seu desenvolvimento, levando 10 meses para atingir uma vizinha. A
cada iteração, novos focos de parasitismo são sorteados. Pelas regras do modelo
hospedeiras já infectadas não podem mais ser sorteadas. Esta regra superestima a
dispersão ornitocórica já que em uma situação real as aves podem depositar sementes
nas mesmas hospedeiras. Este algoritmo poderia ser melhorado se fossem coletados
dados de campo para calcular a probabilidade de deposição de sementes na mesma
hospedeira, em hospedeiras mortas, em agrupamentos e em plantas isoladas.
Para obter a velocidade de expansão foi calculada a média de hospedeiras
infectadas de 10 amostras de paisagens diferentes, iniciando a partir de cada uma das
280 mimosas na rede não infectada (ρ*L2) para uma rede quadrada L=50, t=0,525 e ρ =
0,112, mantendo as mesmas características da paisagem real. No final temos ρ(t)
calculado para 2800 amostras. As iterações foram até 1000 meses (aproximadamente 83

47
anos). O desvio padrão da média para esta densidade foi da ordem de 10-4
ou menores,
deixando a barra de erro insignificante. A taxa de λ = 0,1/mês para a dispersão
vegetativa foi mantida fixa durante a simulação, pois a propagação via passagem de
ramos deve se manter relativamente constante ao longo do tempo. Mas a dispersão
ornitocórica é incerta, pois depende da área de ocorrência das aves, o tamanho de sua
população, suas preferencias alimentares, seu comportamento, dentre outros fatores.
Assim, variou-se a taxa µ de 0,005/mês, em 10 (0,0005/mês) e em 100 (0,00005/mês)
vezes.
Nesta simulação foram desconsiderados a mortalidade de hospedeiras
infectadas, o recrutamento de novas hospedeiras, seu crescimento ao longo do tempo e
seus diferentes tamanhos. Tampouco foram incorporados dados sobre estocasticidade
demográfica e ambiental nos cálculos. Por exemplo, sabe-se que a morte da hospedeira
pode interromper localmente o crescimento vegetativo, provocando a morte da parasita
quando esta não consegue emitir ramos capazes de atingir outra hospedeira. A chance
de passar de uma hospedeira para outra depende também do tamanho e da arquitetura de
suas copas, já que copas maiores são “alvos” mais fáceis de serem alcançados pelos
ramos que copas menores. Entretanto, no modelo todas as mimosas tiveram chances
iguais de serem infectadas pelos ramos da parasita, pois não diferem quanto ao tamanho
e idade. A passagem dos ramos é interrompida quando a parasita se propaga para um
sítio inapropriado, ou seja, que não contém uma mimosa.
Resultados
Paisagem
A paisagem apresentou vários agrupamentos de hospedeiras muito semelhantes
aos encontrados na área de estudo. Em uma área de apenas 16 m2
foi possível encontrar
até 36 hospedeiras, demonstrando o alto padrão de agregação (Mourão 2007). A
densidade média de hospedeiras (ρ) encontrada para a paisagem real foi de 0,112 ±
0,005 e a probabilidade média de agregação (r1,1) foi de 0,061 ± 0,007. A escolha do
tamanho da rede de L = 50 e L = 100 foram baseadas no tempo de simulação e
disposição das hospedeiras na paisagem virtual (Fig. 7).

48
Figura 7 – Características da paisagem gerada pela temperatura escolhida (T=0,53). O
tamanho da rede de L=100 e r1,1 = 0,061±0,002. O agrupamento de hospedeiras (em
verde) foi semelhante ao da área de estudo.
Simulação
Na simulação, durante os primeiros 12 meses a dispersão ornitocórica foi a
principal maneira de infestação na paisagem, já que os ramos da parasita levariam 10
meses aproximadamente para atingir o centro de uma quadrícula susceptível à infecção
(Fig. 8a). Já após 24 meses ficou evidente que as áreas de maiores densidades de
hospedeiras foram as que mais recebem ramos através da dispersão vegetativa (Figs. 8b
e c). Essas manchas são completamente tomadas com a passagem do tempo (Figs. 8c-f).
Como as hospedeiras continuam recebendo sementes das aves, novos pontos de
infestação iniciam-se, elevando a velocidade de espalhamento da parasita e aumentando
sua área de atuação. É interessante notar que ao final de 192 meses mimosas isoladas
raramente foram infectadas (Fig. 8f). A distância acima de 2 m entre as hospedeiras
impediu a passagem de ramos, já que neste modelo foi desconsiderada a existência de
outras plantas. Neste caso, estas hospedeiras isoladas só poderiam ser infectadas apenas
via dispersão por aves.

49

50
Figura 8 – Paisagens simuladas com propagação da parasita para rede L= 100 e
intervalos de tempo t= 12, 24, 36, 48, 60 e 192 meses. Pontos verdes representam
mimosas saudáveis, os azuis as mimosas infectadas por pássaros (dispersão
ornitocórica) e os vermelhos as mimosas infectadas por passagem de ramos (dispersão
vegetativa).
Velocidade de espalhamento
As curvas de velocidade de espalhamento da parasita são do tipo sigmoide,
apresentando-se lento inicialmente, seguido por um rápido aumento exponencial e
finalmente estabilizando quando o sistema está saturado. O espalhamento lento ocorreu
quando plantas sadias foram infectadas basicamente por deposição de “sementes da
parasita” nos galhos das hospedeiras (dispersão ornitocórica), e a duração desta fase
dependeu da taxa de dispersão ornitocórica (Fig. 9). Quando essa taxa foi de 0,005/mês,
a fase exponencial teve seu fim próximo a 98 meses, mas quando foi reduzida em 100
vezes, teve seu fim em aproximadamente 500 meses. Durante o aumento exponencial as
duas dispersões atuaram simultaneamente, acelerando o espalhamento da parasita na
paisagem. A saturação do sistema ocorreu quando praticamente todos os agrupamentos
foram parasitados (Fig. 8f), limitando a passagem de ramos.
Fig. 9. Densidade média de mimosas infectadas, os valores de λ e µ correspondem às
taxas de propagação vegetativa e infecção espontânea (dispersão ornitocórica). As
linhas coloridas indicam variações em uma destas taxas.
(meses)

51
Mantendo as taxas de dispersão vegetativa e ornitocórica propostas para este
modelo, em aproximadamente 98 meses a densidade média de hospedeiras infectadas
seria de 80% e chegaria a 90% em 500 meses (Fig. 9). Quando a taxa da dispersão
ornitocórica foi reduzida em 10 vezes (linha azul marinho da Fig. 9), demorou 200
meses para que a população de mimosas se tornasse 70% infectada e a porcentagem
máxima ficou em torno de 78% em 1000 meses. Já quando foi reduzida em 100 vezes
(linha verde da Fig. 9), em 500 meses haveria cerca de 50% de mimosas infectadas.
Discussão
A dispersão ornitocórica foi fundamental para acelerar o processo de
espalhamento da parasita na paisagem simulada. O papel da ave dispersora tem grande
importância no ciclo de vida destas plantas, pois suas sementes necessitam da remoção
do seu exocarpo para germinar (Ladley & Kelly 1996). As aves contribuem para a
reprodução sexuada da parasita garantindo o aumento da diversidade gênica na espécie
através da propagação de sementes. Como recompensa as aves obtém energia
consumindo frutos de S. flexicaulis que são provavelmente ricos em lipídios (Watson
2001, Guerra 2005).
Na natureza, a agregação de hospedeiras pode facilitar a localização de manchas
na paisagem por aves dispersoras e aumentar as chances de deposição de sementes
(dispersão ornitocórica) (Aukema & Martinez del Rio 2002). O comportamento da ave
foi de certa maneira retratado no modelo, quando a agregação de partículas aumentou a
chance de novos focos de parasitismo iniciar em agrupamentos. A dispersão
ornitocórica, que depende exclusivamente do comportamento das aves, pode atingir
regiões mais distantes. Sabe-se que a ave Elaenia cristata (Tyrannidae), principal
dispersora de S. flexicaulis em um afloramento rupestre no sudeste do Brasil, foi capaz
de dispersar sementes desta parasita a uma distância de até 86 metros (Guerra 2005). A
atuação das aves depositando sementes a grandes distâncias na paisagem é uma
explicação plausível para o acelerado processo de dispersão. Este mesmo fenômeno foi
observado por Clark et al. (1998) quando descreveu a importância dos eventos raros de
dispersão de sementes quando são carregadas para muito longe da planta-mãe, iniciando
novas colonizações e acelerando o processo de migração de árvores após a era glacial.
De acordo com este autor, a distância percorrida por estas sementes pode ser descrita

52
por uma curva de cauda longa e estes raros eventos são os responsáveis pelo acelerado
avanço da população de plantas na paisagem.
O espalhamento de S. flexicaulis em curtas distâncias se deveu principalmente à
dispersão vegetativa. Como o crescimento de ramos é limitado, não ultrapassando 1,5
m/ano (F. A. Mourão obs. pes.), esse tipo de dispersão tem curto alcance. A dispersão
vegetativa é favorecida quando as copas das hospedeiras estão muito próximas, já que
os ramos da parasita podem passar de uma hospedeira à outra dentro de uma mesma
mancha. Quando as hospedeiras se encontravam isoladas, a dispersão vegetativa tendeu
a zero, sendo a dispersão ornitocórica praticamente o único modo de infecção de plantas
na paisagem.
Obviamente, pelas regras do modelo, manchas isoladas não transmitem parasitas
via dispersão vegetativa e não podem infectar novas hospedeiras localizadas em outras
manchas. Isso só foi possível quando as aves depositavam sementes em manchas não
infectadas, iniciando o espalhamento em outras áreas. Em uma comunidade real, a
permeabilidade da matriz é maior do que a considerada nesta simulação, já que outras
plantas fazem parte do ambiente e poderiam ser infectadas ou até mesmo servir como
uma “ponte” entre uma mimosa e outra. Os ramos da parasita teriam maior mobilidade
de espalhamento, minimizando o efeito do substrato inóspito (Overton 1994),
conectando as manchas espacialmente isoladas e acelerando a dispersão.
Sabe-se que a altura das hospedeiras tende a ser positivamente correlacionada
com a intensidade de infestação (Overton 1994; Aukema 2003; Mourão 2007). Na
dispersão vegetativa, hospedeiras mais altas tem maior sucesso na passagem de ramos,
pois estes podem atingir plantas vizinhas de diferentes tamanhos e os ramos podem se
crescer, tocar o substrato e morrer (F. A. Mourão, dados não publicados). Já na
dispersão ornitocórica, a propagação de sementes se dá em função da escolha do poleiro
pelas aves, sendo as maiores as preferidas (Aukema & Del Rio 2002). Nesta simulação
o modelo não considerou características individuais das hospedeiras como altura,
diâmetro e mortalidade. Tampouco variou a densidade e agregação de hospedeiras. Mas
por se tratar de um modelo flexível (Lee & Yang 1952, Chen et al. 1992, Romano
2002), tais parâmetros poderiam ser facilmente incorporados ao modelo, o que iria
torná-lo mais realístico, embora extremamente complexo. Futuros cenários serão
propostos para avaliar como a velocidade de expansão desta parasita pode variar em
função destes parâmetros.

53
A simulação utilizando o modelo de Gás em Rede sugere que tanto a dispersão
ornitocórica quanto a vegetativa desempenham papeis fundamentais e diferenciados no
espalhamento de S. flexicaulis na área estudada. Enquanto a dispersão ornitocórica
carrega as sementes da parasita e as deposita em para manchas mais distantes, a
dispersão vegetativa garante a colonização da nova área.
Referências bibliográficas
Amico G. C., Rodríguez-Cabal M. A., Aizen M. A. 2009. The potential key seed-
dispersing role of the arboreal marsupial Dromiciops gliroides. Acta Oecologica 35:
8–13.
Arruda, R.; Carvalho, L. N. & Del-Claro, K. 2006. Host specificity of a Brazilian
mistletoe, Struthanthus aff. polyanthus (Loranthaceae), in cerrado tropical savanna.
Flora, 201: 127-134.
Aukema, J. E.; del Rio, C. M. 2002. Where does a fruit-eating bird deposit mistletoe
seeds? Seed deposition patterns and an experiment. Ecology, 83: 3489-3496.
Barlow, B. A. & Wiens, D. 1977. Host-parasite resemblance in Australian mistletoes:
the case for cryptic mimicry. Evolution, 31: 69-84
Barneby, R. C. 1991. Sensitivae censitae: a description of the genus Mimosa Linneaus
(Mimosaceae) in the New World. The New York Bootanical Gardens, New York,
835p.
Barth, O. M. 1987. Importância apícola dos campos naturais e rupestres da Serra Norte
de Carajás (PA). Boletim do Museu Paraense Emilio Goeldi. Série Botânica, 3:
21-28.
Benites, V. M.; Shaefer, C. E. G. K.; Simas, F. N. B. & Santos, H. G. 2007. Soils
associated with rock outcrops in the Brazilian mountain ranges Mantiqueira and
Espinhaço. Revista Brasileira de Botânica. 4: 569-577.
Bickford, C. P.; Kolb, T. E.; Geils, B. W. 2005. Host physiological condition regulates
parasitic plant performance: Arceuthobium vaginatum subsp cryptopodum on Pinus
ponderosa. Oecologia, 146: 179-189.
Cannas, S. A.; Marco, D. E.; Paez, S. A. 2003. Modelling biological invasions: species
traits, species interactions, and habitat heterogeneity. Mathematical Biosciences,
183: 93-110.
Caswell, H.; Etter, R.J. 1993. Ecological interactions in patchy environments: from
patch-occupancy models to cellular automata. IN: Patch Dynamics, S.A. Levin, T.

54
Powell e J. H. Steele eds., Lecture Notes in Biomathematics 96:93-109, Springer-
Verlag, Berlin.
Chen, H.; Chen, S. & Mattheus, W. H. 1992. Recovery of the Navier-Stokes equations
using a lattice-gas Boltzmann method. Physical Review, 45: 5339-5342.
Chen, Q.; Mao, J. & Li, W. 2006. Stability analysis of harvesting strategies in a cellular
automata based predator-prey model. Lecture Notes in Computer Science 4173: 268-
276.
Clark, J. S.; Fastie, C.; Hurtt, G.; Jackson, S. T.; Johnson, C.; King, G. A.; Lewis, M.;
Lynch, J.; Pacala, S.; Prentice, C.; Schupp, E. W.; Webb III, T. & Wyckoff, P. 1998.
Reid’s paradox of rapid plant migration. BioScience, 48:13-24.
Cottenie, K.; De Meester, L. 2004. Metacommunity structure: Synergy of biotic
interactions as selective agents and dispersal as fuel. Ecology, 85:114-119.
Cumming, G. 2002. Habitat shape, species invasions, and reserve design: Insights from
simple models. Conservation Ecology, 6:245.
Davidar, P. 1978. Dispersal in some Loranthaceae of Nilgiris. Journal Bombay Natural
History Society, 75: 943-945.
Davidar, P. 1983. Birds and Neotropical mistletoes: effects on seedling recruitment.
Oecologia, 60: 271-273.
Dean, W. R. J.; Midgley, J. J. & Stock, W. D. 1994. The distribution of mistletoes in
South-Africa- patterns of species richness and host choice. Journal of Biogeography,
21: 503-510.
Del Rio, C. M.; Silva, A.; Medel, R. & Hourdequin, M. 1996. Seed dispersers as
disease vectors: bird transmission of mistletoe seeds to plant hosts. Ecology, 77: 912-
921.
Dunning, J. B., Stewart, D. J., Danielson, B. J., Noon, B. R., Root, T. L., Lamberson, R.
H. & Stevens, E. E. 1995. Spatially Explicit Population Models: Current forms and
future uses. Ecological Applications 5: 3-11.
Ehleringer, J. R. & Marshall, J. D. 1995. Water relations. In: Press, M. C. & Grave, J.
D. 1995. Eds. Parasitic plants. London: Chapman & Hall, pp.292.
Fadini, R. F. 2011. Non-overlap of host used by three congeneric and sympatric
loranthaceous mistletoe species in an Amazonian savanna: host generalization to
extreme specialization. Acta Botânica Brasílica, 25: 337-345.
Giacomini, H. C. 2007. Sete motivações teóricas para o uso de modelagem baseada no
indivíduo em ecologia. Acta Amazônica 37: 431-446.

55
Hanski, I. & Simberloff, D. 1997. The metapopulation approach, its history, conceptual
domain, and application to conservation. In: Metapopulation Biology. Eds: Hanski,
I. A. & Gilpin, M. E. p 5 -26. Academic Press, San Diego, California.
Hoffman, A. J. Fuentes, E. R., Cortes, I. Liberona, F. & Costa, V. 1986. Tristerix
tetrandrus (Loranthaceae) and its hosts plants in the Chilean matorral: patterns and
mechanisms. Oecologia, 69: 202-206.
Huston, M. A., DeAngelis, D. & Post, W. 1988. Size bimodality in monospecific
populations: a critical review of potential mechanisms. American Naturalist, 129:
678-707.
Jacobi, C. M. & Carmo, F. F. (orgs.) 2012. Diversidade florística nas cangas do
Quadrilátero Ferrífero (no prelo).
Jacobi, C. M.; Carmo, F. F.; Vincent, R. C. & Stehmann, J. R. 2007. Plant communities
on ironstones outcrops: a diverse and endangered Brazilian ecosystem. Biodiversity
and Conservation, 16: 2185-2200.
Kareiva, P. 1994. Special Feature: Space: The Final Frontier for Ecological Theory.
Ecology. 75:1.
Klein, C. 2000. Geochemistry and Petrology of some Proterozoic Banded Iron-
Formations of the Quadrilátero Ferrífero, Minas Gerais, Brazil. Economic Geology,
95: 405-428.
Kuijt, J. 1963. On the ecology and parasites of the Costa Rica Tree Mistletoe,
Gaiadendron punctatum (Ruiz and Pavon). Canadian Journal of Botany, 41: 927-
938.
Lavorel, S.; Smith, M.S.; Reid, N. 1999. Spread of mistletoes (Amyema preissii) in
fragmented Australian woodlands: a simulation study. Landscape Ecology, 14: 147-
160.
Lee, T. D. & Yang, C. N. 1952. Statistical theory of equations of state and phase
transitions lattice-gas and ising model. Physical Review 87: 410-419.
Levins, R. 1970.Extinction. Some Mathematical Problems in Biology. Ed. Gestenhaber,
M. p 77 -107. American Mathematical Society, Providence, Reino Unido.
López-de Buen, L. e Ornelas, J. F. 1999. Frugivorous birds, host selection and the
mistletoe Psittacanthus schiedeanus, in central Veracruz, Mexico. Journal of
Tropical Ecology, 15: 329-340.
Mistro, D. C.; Jacobi, C. M. 1999. Autômatos celulares na dispersão de pólen. IN: IX
Congresso Internacional de Biomatemática, Actas, v.1, p. 158-166.

56
Mourão, F. A.; Carmo, F. F; Ratton, P.; Jacobi, C. M. 2006. Hospedeiras da
hemiparasita Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) em campos
rupestres ferruginosos, Quadrilátero Ferrífero, MG. Lundiana, 7: 103-110.
Mourão, F. A., Jacobi, C. M. Figueira, J. E. C & Batista, E. K. L. 2009. Effects of the
parasitism of Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) on the fitness of
Mimosa calodendron Mart. (Fabaceae), an endemic shrub from rupestrian fields over
ironstone outcrops, Minas Gerais State, Brazil. Acta botanica brasilica 23: 820-825.
Overton, J. M. 1994. Dispersal and infection in mistletoe metapopulations. Journal of
Ecology, 82: 711-723.
Overton, J. M. 1996. Spatial autocorrelation and dispersal in mistletoes: Field and
simulation results. Vegetatio, 125: 83-98.
Rodrigues, A. L. & Tomé, T. 2008. Reaction-diffusion stochastic lattice model for a
predator-prey system. Brazilian Journal of Physics, 38: 87-93.
Romano, S. 2002. Computer simulation study of nematogenic lattice-gas model with
fourth-rank interactions. International Journal of Modern Physics B, 19:2901-2915.
Sargent, S. 1995. Seed fate in a tropical mistletoe: the importance of host twig size.
Functional Ecology, 9: 197-204.
Satulovsky, J. E. & Tomé T. 1994. Stochastic lattice gas model for predator-prey
system. Physical Review Letters. 49: 50 -73.
Shaw, D. C.; Watson, D. M. & Mathiasen, R. L. 2004. Comparison of dwarf mistletoes
(Arceuthobium spp., Viscaceae) in the Western United States with mistletoes
(Amyema spp., Loranthaceae) in Australia-ecological analogs and reciprocal models
for ecosystem management. Australian Journal of Botany, 52: 481-498.
Silva, M. F. F.; Secco, R. S. & Lobo, M. G. A. 1996. Aspectos ecológicos da vegetação
rupestre da serra dos Carajás, estado do Pará, Brasil. Acta Amazonica, 26: 17-44.
Sutton, G. M. 1951. Mistletoe dispersal by birds. Wilson Bulletin, 63:235-237.
Teodoro, G. S. 2010. Estrutura e dinâmica da erva-de-passarinho Psittacanthus
Robustus Mart. em manchas de habitat em três áreas de cerrado Tese Mestrado em
Ecologia Universidade Federal de Lavras, MG (Brazil), 74 p.
Thomson, V. E. and Mahal, B. E. 1983. Host specificity by a mistletoe, Phoradendron
villosum (Nutt.) Nutt. Subsp villosum, on three oak species in California. Botanical
Gazette, 144: 124-131.

57
Vallauri, D. 1998. Parasite dynamics of Viscum album L. in Austrian black pine stands
in the Saignon watershed (southwestern Alps). Annales des Sciences Forestieres,
55: 823-835.
Viana, P. L. & Lombardi, J. A. 2007. Florística e caracterização dos campos rupestres
sobre a canga na Serra da Calçada, Minas Gerais, Brasil. Rodriguésia, 58: 159-177.
Vincent, R. C. 2004. Florística, fitossociologia e relações entre a vegetação e o solo em
área de campos ferruginosos no Quadrilátero Ferrífero, Minas Gerais. Tese de
doutorado. Universidade de São Paulo, São Paulo, 145p.
Vilela, R. A.; Melo, R. J.; Costa, T. A. V.; Lagoeiro, L. E. & Varajão, C. A. C. 2004.
Petrografia do minério hematita compacta da Mina do Tamanduá (Quadrilátero
Ferrífero, MG). Revista Escola de Minas, Ouro Preto, 57: 157-164.
Wolf, F.G., 2002. Simulação de Processos de Deslocamento Miscível Utilizando
Modelos de Gás em Rede com Mediadores de Campo. Dissertação de Mestrado –
Universidade Federal de Santa Catarina.

58
CAPÍTULO 3
Alocação de recursos da hemiparasita Struthanthus
flexicaulis (Loranthaceae) em condições de estresse
hídrico do ambiente.
Fabiana Alves Mourão1, Claudia Maria Jacobi
1 & José Eugenio Cortes Figueira
1
1 Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade
Federal de Minas Gerais.

59
Resumo
Padrões de alocação de recursos em plantas podem variar em função dos
estresses ambientais aos quais estão submetidas. Entretanto, plantas parasitas poderiam
não sofrer consequências já que usufruem permanentemente dos nutrientes retirados de
suas hospedeiras. O objetivo desse trabalho foi verificar isto, comparando a alocação de
recursos da parasita Struthanthus flexicaulis em diferentes condições de estresse hídrico
do ambiente. Foram marcados 347 ramos principais que tiveram seu crescimento
acompanhado por 9 meses. Estes ramos parasitavam 32 indivíduos de Mimosa
calodendron, sua principal hospedeira em afloramentos ferruginosos. Mediu-se o
comprimento de cada ramo principal, sua localização (dentro ou fora da copa), o
número de ramos laterais, de folhas, de estruturas reprodutivas (botões, flores e frutos) e
de ramos danificados por herbivoria ou ressecamento. Além disso, foram comparados
ramos que cresciam em hospedeiras com três categorias de infestação. O crescimento de
ramos foi maior na estação seca do que na estação chuvosa, contrariamente ao número
de folhas. A mortalidade foi semelhante nos dois períodos, mas as causas foram
diferentes. A sobrevivência dos ramos exploratórios (fora das copas) foi maior do que a
dos internos (dentro das copas). Na estação seca não houve diferença significativa entre
o crescimento, número de estruturas reprodutivas e de folhas entre os ramos
exploratórios e internos. Já na estação chuvosa os ramos exploratórios cresceram mais e
produziram mais estruturas reprodutivas, sem, no entanto, diferir dos ramos internos
quanto ao número de folhas. Entre junho e agosto, os ramos em hospedeiras pouco
parasitadas apresentaram menor crescimento quando comparados aos de hospedeiras
muito parasitadas. O número de folhas seguiu o mesmo padrão e o de estruturas
reprodutivas foi semelhante entre as três categorias. Já entre os meses de novembro a
janeiro o crescimento não diferiu, mas os ramos que cresciam sobre as mimosas com
parasitismo intermediário tiveram maior número de estruturas reprodutivas e de folhas.
S. flexicaulis demonstrou sinais de estresse hídrico durante a estação seca, porque,
apesar de ter investido mais em crescimento vegetativo, não produziu estruturas
reprodutivas que são energicamente mais custosas. A parasita adotou diferentes
estratégias para garantir sua sobrevivência quando alocou recursos para realizar
diferentes funções durante as estações seca e chuvosa.
Palavras-chave: alocação de recursos, disponibilidade de água, seca.

60
Introdução
Os padrões de alocação de recursos em plantas influenciam sua habilidade de
aquisição de nutrientes (Poorter et al. 1990), interações competitivas (Bazzaz et al.
1987), crescimento vegetativo e reprodução (de Jong & Klinkhamer 2005). Durante o
crescimento, a fase vegetativa é o período de maior ganho de biomassa (Shaver &
Chapin 1991, Larcher 2006). É nesta etapa que as plantas alongam raízes e partes aéreas
e ainda investem em estruturas clonais como, por exemplo, os estolões, bulbos e
rametes (Grace 1993, Larcher 2006). O decréscimo no crescimento vegetativo é
observado quando se inicia a fase reprodutiva. Isso sugere que os recursos requeridos
para a reprodução são os mesmos utilizados para o crescimento (Bazzaz et al. 1987).
Essas mudanças na alocação de biomassa afetam a sobrevivência, o crescimento e a
reprodução das plantas (McConnaughay & Coleman 1999).
A alocação de recursos pode variar em função dos estresses ambientais aos quais
as plantas estão submetidas, tais como a falta de água, variações extremas de
temperatura e variações nas concentrações de nutrientes (Bloom et al. 1985). O estresse
hídrico, que ocorre quando a disponibilidade de água no solo é baixa e a temperatura é
capaz de causar perdas contínuas por transpiração ou evaporação (Jaleel et al. 2008),
por exemplo, pode provocar a redução das taxas fotossintéticas e de respiração,
distúrbios no metabolismo e até mesmo a morte do indivíduo (Jaleel et al. 2008, Farooq
et al. 2009). As respostas das plantas ao estresse podem ser rápidas e diretas visando
aumentar as chances de sobrevivência imediata (Mooney et al. 1991). O rápido
fechamento dos estômatos em resposta à seca pode evitar a perda de água, mas também
interfere na assimilação de carbono (Mooney et al. 1991). Em longo prazo, se as
condições desfavoráveis persistirem, as plantas passam a realocar os recursos
disponíveis para o desenvolvimento de mecanismos complexos envolvendo fenômenos
moleculares, bioquímicos e fisiológicos, afetando seu crescimento e reprodução
(Razmjoo et al. 2008).
Os efeitos do estresse hídrico sobre as plantas parasitas parecem não ser tão
acentuados quanto em outras plantas. Elas assumem o controle homeostático de água de
suas hospedeiras e intensificam a retirada de recursos (Glatzel & Geils 2009). Muitas
parasitas tem a capacidade de realizar fotossíntese e investem mais em crescimento
quando retiram grandes quantidades de água e nutrientes do xilema de suas hospedeiras

61
(Aukema 2003). Como o transporte de água e nutrientes ocorre devido a pressões
negativas no topo da planta pela evapotranspiração nas folhas (Glatzel & Geils 2009), as
hospedeiras sofrem duplamente com a falta de água e podem morrer principalmente
durante o período seco. Para que a parasita possa manter as taxas adequadas de seu
crescimento sem por em perigo a sobrevivência da hospedeira, é esperado um balanço
entre a quantidade de recurso disponível e o que é efetivamente retirado (Tennakoon &
Pate 1996). Mas quando a limitação de água e nutrientes é severa as parasitas buscam
novas fontes de recursos infectando mais de uma planta (Marvier 1996).
Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) é uma das hemiparasitas
brasileiras mais comuns, com ampla distribuição nas áreas de cerrado do Brasil Central
(Rizzini 1980). A espécie é generalista e pode parasitar diversas plantas vasculares
(Mourão et al. 2006). Seus ramos longos e flageliformes dispersam-se vegetativamente
e podem se fragmentar, formando novos rametes. Em fitofisionomias com elevadas
densidades de plantas a dispersão vegetativa é tão ou mais importante que a
ornitocórica, porque os ramos da parasita são capazes de infectar as copas de plantas
vizinhas espalhando-se no ambiente (Mourão et al. 2009), favorecendo a colonização de
novas hospedeiras e seu forrageamento. Esta espécie é dióica e possui folhas pequenas,
com flores e frutos pequenos (3-4 mm e 6-7 mm respectivamente).
O objetivo desse trabalho foi comparar a alocação de recursos de S. flexicaulis
em diferentes condições de estresse hídrico do ambiente. Devido à dificuldade de se
distinguir indivíduos, optou-se por adotar como unidade amostral os ramos da parasita.
As hipóteses que nortearam este trabalho foram: 1) A alocação de recursos dos ramos de
S. flexicaulis e sua mortalidade são influenciadas pela disponibilidade de água no
ambiente; 2) A alocação de recursos é afetada quando a infestação de parasitas for
elevada; 3) A sobrevivência dos ramos varia em função de sua posição com relação às
copas (dentro-internos ou fora-exploratórios) e 4) O crescimento de ramos é
influenciado por de sua posição com relação às copas (dentro-internos ou fora-
exploratórios). Acredita-se que na estação chuvosa os ramos de S. flexicaulis crescerão
mais, produzirão mais folhas e mais estruturas reprodutivas. A mortalidade deverá ser
menor. A alocação para os ramos que crescem sobre as copas de hospedeiras muito
parasitadas deverá ser menor, pois é esperada uma competição por recursos disponíveis
entre os ramos da parasita. Além disso, a sobrevivência deverá ser maior dentro das
copas já que seus ramos ficam menos expostos à predação e danos como quebra de suas

62
extremidades. Os ramos exploratórios (fora das copas) deverão investir mais em
crescimento linear porque se supõe que estes estejam à procura de novas hospedeiras.
Material e métodos
Local de estudo
O estudo foi desenvolvido numa área de afloramentos ferruginosos do Parque
Estadual da Serra do Rola Moça (20° 03’ 60”S, 44° 02’ 00” W, 1300 m), localizado na
porção sul da Cadeia do Espinhaço- Brasil, uma longa cadeia de montanhas paralela ao
oceano Atlântico. O clima da região é do tipo mesotérmico, de acordo com a
classificação de Köppen, é do tipo Cwa (Nimer & Brandão 1989). A precipitação anual
é de 1000 a 1500 mm, com uma pronunciada estação seca de abril a setembro, e
temperatura média anual de 25°C (Rizzini 1997). Nos afloramentos, localizados nos
topos da serra, os ventos constantes e altos índices de radiação ultravioleta contribuem
para a severidade do ambiente, agravada durante a estação seca. A vegetação é baixa,
composta por poucos indivíduos de porte arbóreo, e é dominada por eudicotiledôneas
como Asteraceae, Fabaceae e Myrtaceae, e monocotiledôneas como Poaceae,
Cyperaceae e Orchidaceae (Viana & Lombardi 2007, Jacobi et al. 2007). O solo é do
tipo neossolo litólico, ou seja, quando presente é raso (apresentando menos de 5 cm de
profundidade), pedregoso e cascalhento,é rico em ferro e apresenta baixos teores de
fósforo e micronutrientes (Benites et al. 2007). Na área de estudos Mimosa calodendron
(Fabaceae) é uma leguminosa abundante e frequentemente infectada pela hemiparasita
S. flexicaulis (Mourão et al. 2009). Acredita-se que M. calodendron seja endêmica dos
afloramentos rochosos ferrugíneos do Quadrilátero Ferrífero (Barneby, 1991) e
contribua para a estruturação da comunidade vegetal em que se encontra inserida
(Jacobi et al., 2007). Esta hospedeira é perene e de porte arbustivo sendo utilizada como
poleiro por diversas aves, inclusive as dispersoras de S. flexicaulis.
Desenho amostral
Para avaliar a alocação de recursos da parasita, foi delimitada uma área de
aproximadamente 2500 m2, contendo uma população expressiva de indivíduos de M.
calodendron parasitados. Foram marcados 347 ramos principais femininos que
parasitavam 32 indivíduos de M. calodendron, cujas alturas variaram de 80 e 120 cm.
Estes ramos foram marcados a 10 cm de suas extremidades para padronizar o início das
medidas (Fig. 1), que foram tomadas mensalmente durante nove meses, iniciando-se na

63
estação seca (maio) e finalizando na chuvosa (janeiro). Mediu-se o comprimento do
ramo principal, o número de ramos laterais, o número de folhas, a localização (dentro
ou fora da copa), o número de estruturas reprodutivas (botões, flores e frutos) e o
número de ramos danificados por herbivoria ou quebra. Os ramos poderiam apresentar
crescimento negativo quando: 1) cresciam sobre o substrato do afloramento ferruginoso
cujas temperaturas no verão chegam a 68° (F. F. Carmo com. pess.) e queimavam suas
extremidades ou 2) larvas de insetos no interior dos ramos que se rompiam após a
eclosão do adulto.
Fig. 1- Foto esquemática dos ramos da parasita onde A = ramo principal e B = ramos
laterais.
Para comparar o crescimento de ramos entre as estações seca e chuvosa, foram
considerados o incremento linear entre junho e agosto (estação seca) e entre novembro e
janeiro (estação chuvosa). Aplicou-se o teste U de Mann-Whitney, pois os dados não
apresentaram distribuição normal. O número de folhas de cada ramo, a partir da
marcação feita em maio, foi contado em agosto (estação seca) e janeiro (estação
chuvosa), e comparado por análise de covariância ANCOVA (covariável: número de
folhas), após a transformação usando log na base 10 para diminuir a variância. Em
seguida comparou-se cada reta através da inclinação das tangentes e os valores do
intercepto segundo metodologia de Zar (1999). As diferenças entre os valores
encontrados, tanto para as tangentes quanto para os interceptos foram comparados com
teste t de Student.
Os 347 ramos principais foram divididos em dois grupos: 142 ramos internos e
205 exploratórios. Os ramos internos são aqueles que permaneceram dentro das copas e
os exploratórios são aqueles que se estendem mais do que 10 cm além da borda da copa.
A
B
B

64
Estes grupos foram analisados quanto a: 1) sobrevivência (taxa de sobrevivência mensal
dos ramos principais); 2) porcentagem de ramos principais danificados nas estações
seca e chuvosa (morte por ressecamento, morte por herbivoria ou ressecamento, e
quebra das extremidades); 3) comprimento; 4) número de folhas/cm da parasita e 5)
número de estruturas reprodutivas/cm da parasita. Para calcular a taxa de sobrevivência
os ramos foram agrupados em cinco classes de tamanho de intervalos de 20 cm e foi
criada uma tabela de vida de todos os ramos marcados no início do experimento e a
proporção sobrevivente ao longo do tempo. Foi calculada a taxa de sobrevivência
mensal para cada grupo de ramos e comparada pelo teste de Wilcoxon. Para comparar o
número de ramos danificados exploratórios e internos foi utilizado um teste t pareado,
pois os dados apresentaram distribuição normal.
Como ao longo do experimento alguns ramos mudavam de posição para dentro ou
fora das copas, para comparar o crescimento, o número de folhas/cm da parasita e de
estruturas reprodutivas/cm da parasita em cada estação, considerou-se apenas os ramos
que permaneceram no mesmo grupo (somente dentro ou somente fora das copas) após
os nove meses de monitoramento. Assim, as análises de crescimento, número de folhas
e estruturas reprodutivas em cada estação foram realizadas com 71 ramos internos e 104
exploratórios. Os dados foram analisados pelo teste U de Mann-Whitney uma vez que
não apresentaram distribuição normal, mesmo após as transformações. Os danos
causados por morte por ressecamento, morte por herbivoria ou ressecamento e quebra
das extremidades na chuva e na seca foram expressos em porcentagens e comparados
pelo teste do Qui quadrado.
Para avaliar se os ramos que cresciam em hospedeiras com diferentes graus de
infestação apresentavam diferenças no crescimento linear, número de folhas e estruturas
reprodutivas nas estações seca (agosto) e chuvosa (janeiro), cada hospedeira foi
classificada em uma das seguintes categorias de parasitismo: 1) pouco parasitadas – até
10 ramos exploratórios; 2) parasitismo intermediário – 20 a 30 ramos exploratórios e; 3)
muito parasitadas – mais de 40 ramos exploratórios. Os ramos de cada categoria foram
comparados pelo teste não paramétrico Kruskal-Wallis, já que os dados não
apresentaram uma distribuição normal mesmo após as transformações.
Os dados de precipitação dos anos de 2009 e 2010, utilizados neste estudo, foram
extraídos da estação meteorológica do Parque Estadual da Serra do Rola Moça.

65
Resultados
Os ramos de S. flexicaulis alocaram os recursos para realizar diferentes funções
durante o seu desenvolvimento ao longo dos meses. O crescimento de ramos de S.
flexicaulis foi maior na estação seca do que na estação chuvosa (Z= - 9,513; n=175;
p<0,0001) (Fig. 2). Entretanto, o número de folhas foi maior durante a estação chuvosa
(Fig. 3).
Fig. 2 – Crescimento de ramos entre junho e agosto (período seco) e entre novembro e
janeiro (período chuvoso). A classe <0 representa os ramos que diminuiram seu
comprimento devido aos danos nas suas extremidades.
Para a estação seca a reta que descreveu a correlação entre o número de folhas e
comprimento dos ramos exploratórios e internos foi y = -0,098 + 0,858x (n = 175;
r2=0,860; p<0,001) e para a chuvosa, representada pelo mês de janeiro, foi y = -0,946 +
1,047x (n = 175; r2=0,846; p<0,001) (Fig. 3). Essas retas diferiram quanto a sua
inclinação (n=350; t= 4,157; p=0,001), mas não quanto ao intercepto (n=350; t=1,208;
p=0,228).
A menor quantidade de chuva durante a estação seca não impediu a produção
dos primeiros botões em julho, alcançando seu pico em outubro (Fig. 4). Já as primeiras
flores surgiram em agosto, e em setembro foi iniciada a produção de frutos.
Informações sobre os frutos foram coletados até o mês de janeiro e por esse motivo não

66
foi possível quantificar seu pico de produção, que acontece entre março e abril (Mourão
obs. pess.).
1 10 100
Log Comprimento (cm)
1
10
100
1 10 1001
10
100
Lo
g N
úm
ero
de f
olh
as
Fig. 3 – Correlação entre o número de folhas e o comprimento dos ramos. Para o
período seco (agosto, cinza), e para o período chuvoso (janeiro, pretos).
May Ju
n Jul
AugSep Oct
NovDec Ja
n0
10
20
30
40
50
60
70
80
Rep
roduct
ive
stru
cture
s/bra
nch
es
0
100
200
300
400
500
600
Rain
(mm
)
Bud
Flower
Fruit

67
Fig. 4 – Relação entre a quantidade de chuva (mm) e o número total de estruturas
reprodutivas por ramo (botões, flores e frutos) durante os meses de maio de 2009 a
janeiro de 2010.
Quando se considerou a produção de estruturas reprodutivas dos ramos notou-se
maior investimento nos meses de setembro a novembro, o que coincidiu com o período
de maior disponibilidade de água (Tab. 1). O início da produção de botões se deu no
mês de julho (4,26 / ramo) e teve seu pico no mês de outubro (77,36 / ramo). No mês de
novembro houve maior produção de flores (13,08 / ramo) e no mês de janeiro a
produção de frutos verdes e maduros se destacou com 28,17 / ramo (Tab. 1). Durante a
estação chuvosa, houve um maior investimento na produção de botões se comparado às
flores e frutos (Tab. 1). A produção de botões é energeticamente menos custosa para a
planta porque são menores e mais leves e, além disso, não precisam produzir
recompensas como as flores, e nem armazenar nutrientes como os frutos.
Tabela 1– Produção de estruturas reprodutivas e crescimento de ramos laterais nos
períodos de seca (maio a agosto) e chuva (setembro a janeiro).
Meses Botões Flores Frutos
Número de ramos
laterais
Número
de ramos
Produção
total
Número de
ramos
Produção
total
Número de
ramos
Produção
total
Maio 0 0 0 0 0 0 58
Junho 0 0 0 0 0 0 199
Julho 105 448 0 0 0 0 460
Agosto 157 2603 11 113 0 0 450
Setembro 218 11960 23 120 1 1 418
Outubro 202 15628 94 916 1 7 530
Novembro 165 6767 160 2093 25 499 452
Dezembro 87 1053 114 903 49 1145 443
Janeiro 16 121 17 118 41 1155 279
A mortalidade de ramos foi semelhante nos dois períodos, sendo de 3,7% na
seca e 4,2% na chuvosa. Entre as causas identificadas, a morte por ressecamento foi
mais comum durante a estação seca, enquanto a herbivoria ocorreu com maior
intensidade durante a estação chuvosa (Tab. 2). A porcentagem de quebra dos ramos
principais, sem, no entanto, leva-los à morte, foi maior durante a estação seca (Tab. 2).
Entre os meses de junho a agosto, o maior crescimento de ramos foi de 104 cm para um
que crescia fora da copa de uma hospedeira vigorosa, destoando fortemente do

68
crescimento médio para este período, que foi de 20,2 ± 17,6 cm, com mediana de 16
cm. Já durante a estação chuvosa, entre os meses de novembro a janeiro, o maior
crescimento foi de 68 cm e o crescimento médio de 4,5 ± 19,1 cm e a mediana foi de 1
cm.
Tabela 2. Porcentagem de danos e morte dos ramos principais nos períodos seco (de
junho a agosto) e chuvoso (novembro a janeiro).
SECA
N=205
CHUVA
N=176 χ2 gl p
Morte por ressecamento 19,5 6,1 6,53 1 0,01
Morte por herbivoria 0,5 27,8 27,35 1 0,00001
Quebras 35,5 19,6 4,02 1 0,05
Nestes últimos meses muitos ramos apresentaram crescimento negativo; por exemplo, o
que mais decresceu tinha 81 cm em novembro e em janeiro media 15 cm.
Quando a mortalidade foi analisada de acordo com posição dos ramos na copa, a
quebra foi mais comum nos ramos exploratórios enquanto a morte por ressecamento ou
herbivoria foi mais comum nos ramos internos (Tab. 3).
Tabela 3. Porcentagem de danos e morte dos ramos internos e exploratórios nos
períodos seco (de junho a agosto) e chuvoso (novembro a janeiro).
SECA CHUVA
Internos Exploratórios
N=109
Internos Exploratórios
N= 96 N=75 N=106
Morte por
ressecamento 12,6 6,9 3,2 2,9
Morte por
herbivoria 1 0 14,9 12,9
Quebras 12,5 23 7,8 11,8
Dos 347 ramos principais marcados, 142 se encontravam inicialmente dentro e
205 fora das copas. Os ramos que cresceram dentro das copas entrelaçaram-se com
outros ramos da parasita ou prenderam-se aos galhos da hospedeira. Estes ramos
internos tinham maior calibre e folhas maiores, escuras e menos maleáveis,
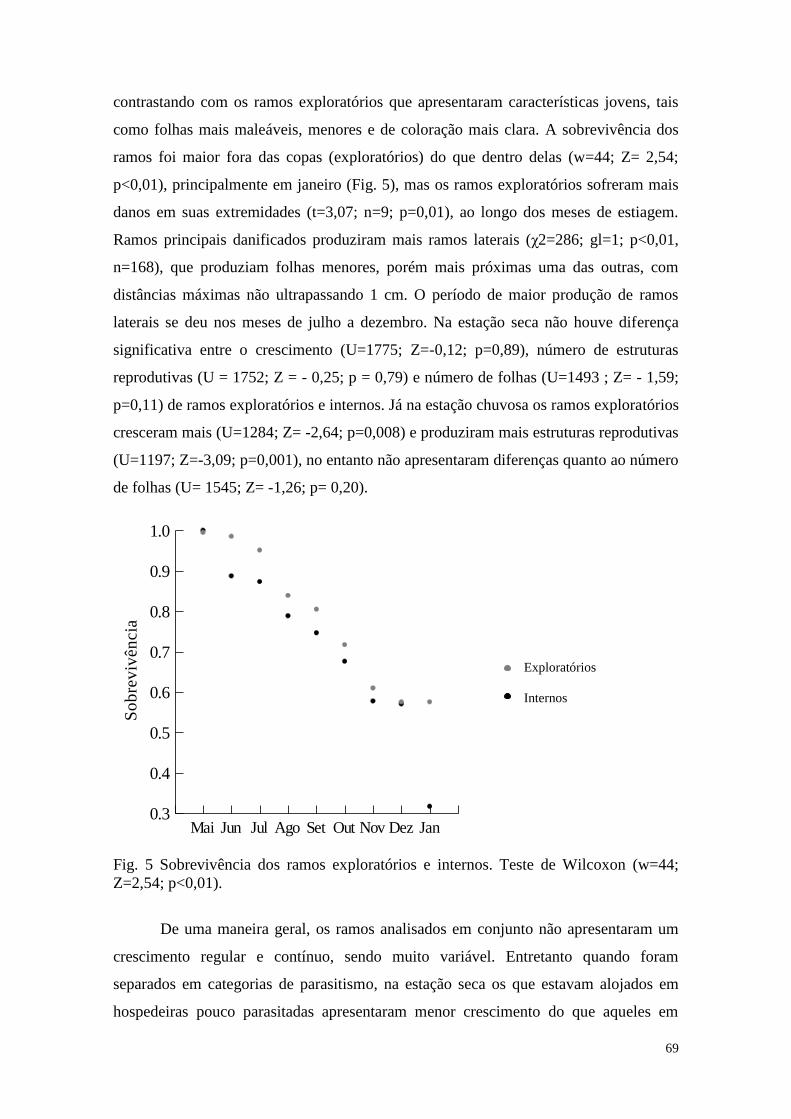
69
contrastando com os ramos exploratórios que apresentaram características jovens, tais
como folhas mais maleáveis, menores e de coloração mais clara. A sobrevivência dos
ramos foi maior fora das copas (exploratórios) do que dentro delas (w=44; Z= 2,54;
p<0,01), principalmente em janeiro (Fig. 5), mas os ramos exploratórios sofreram mais
danos em suas extremidades (t=3,07; n=9; p=0,01), ao longo dos meses de estiagem.
Ramos principais danificados produziram mais ramos laterais (χ2=286; gl=1; p<0,01,
n=168), que produziam folhas menores, porém mais próximas uma das outras, com
distâncias máximas não ultrapassando 1 cm. O período de maior produção de ramos
laterais se deu nos meses de julho a dezembro. Na estação seca não houve diferença
significativa entre o crescimento (U=1775; Z=-0,12; p=0,89), número de estruturas
reprodutivas (U = 1752; Z = - 0,25; p = 0,79) e número de folhas (U=1493 ; Z= - 1,59;
p=0,11) de ramos exploratórios e internos. Já na estação chuvosa os ramos exploratórios
cresceram mais (U=1284; Z= -2,64; p=0,008) e produziram mais estruturas reprodutivas
(U=1197; Z=-3,09; p=0,001), no entanto não apresentaram diferenças quanto ao número
de folhas (U= 1545; Z= -1,26; p= 0,20).
Mai Jun Jul Ago Set Out Nov Dez Jan
MES
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
So
bre
viv
ên
cia
Fig. 5 Sobrevivência dos ramos exploratórios e internos. Teste de Wilcoxon (w=44;
Z=2,54; p<0,01).
De uma maneira geral, os ramos analisados em conjunto não apresentaram um
crescimento regular e contínuo, sendo muito variável. Entretanto quando foram
separados em categorias de parasitismo, na estação seca os que estavam alojados em
hospedeiras pouco parasitadas apresentaram menor crescimento do que aqueles em
Exploratórios
Internos

70
hospedeiras com níveis intermediários de parasitismo e muito parasitadas (H = 14,97; gl
= 2; p < 0,0001) (Tab. 4). O número de folhas seguiu o mesmo padrão (H = 17,45; gl =
2; p < 0,001). A produção de estruturas reprodutivas foi semelhante entre as 3
categorias (H = 2,31; gl = 2; p = 0,31). Já na estação chuvosa o crescimento não diferiu
entre os grupos (H = 0,41; gl = 2; p = 0,81) (Tab. 4). O número de folhas (H = 3,90; gl =
2; p = 0,11) e de estruturas reprodutivas (H = 4,29; gl = 2; p = 0,14) seguiram o mesmo
padrão.
Tabela 4. Comparação da alocação de recursos dos ramos da parasita considerando-os
crescendo sobre diferentes classes de hospedeiras parasitadas nas estações seca e
chuvosa.
Seca Comprimento
Estruturas
Reprodutivas
Número de
folhas
Pouco Parasitada (N=43) 28,8 ± 15,4 A 9,5 ± 15,8
A 19,4 ± 13,4
A
Parasitismo Intermediário
(N=55) 40,5 ± 19,1 B 10,1 ± 12,8
A 23,8 ± 12,7
B
Muito Parasitada (N=68) 41,4 ± 17,1 B 13,1 ± 18,5
A 30,1 ± 14,3
B
Kruskal-Wallis Comprimento (H=14,97 gl=2 p<0,0001)
Kruskal-Wallis Estrutura Reprodutiva (H=2,31 gl=2 p=0,31)
Kruskal-Wallis Número de Folhas (H= 17,45 gl= 2 p<0,0001)
Chuva Comprimento
Estruturas
Reprodutivas
Número de
folhas
Pouco Parasitada (N=18) 30,7 ± 21,1 A
10,1 ± 31,8 A 20,8 ± 21,5
A
Parasitismo Intermediário
(N=42) 30 ± 19 A 8,2 ± 8,1
A 25,5 ± 18,4
A
Muito Parasitada (N=49) 34 ± 21 A 11,6 ± 23,4
A 29,5 ± 19,3
A
Kruskal-Wallis Comprimento (H=1,54 gl=2 p=0,46)
Kruskal-Wallis Estrutura Reprodutiva (H=4,29 gl=2 p=0,11)
Kruskal-Wallis Número de Folhas (H= 3,90 gl= 2 p=0,14)
Discussão
As condições de estresse hídrico do ambiente influenciaram a alocação de
recursos de S. flexicaulis. Durante a estação seca, seus ramos investiram mais no
crescimento sem, entretanto, produzir muitas folhas. Isso sugere uma estratégia para
buscar novas fontes de recursos (hospedeiras), já que neste período água e nutrientes são
fatores limitantes e críticos para a maioria das plantas. Embora as plantas parasitas

71
possam ser quase insensíveis à seca, pois dependem exclusivamente de suas
hospedeiras, estas últimas estão sujeitas à falta de água e podem não retirar recursos
suficientes nem mesmo para sua própria manutenção. O parasitismo reduz a biomassa e
altera a alocação de recursos das espécies parasitadas (Tennakoon & Pate 1996, Lei
2001, Press & Phoenix 2005, Shen et al. 2006), podendo leva-las à morte. Nos
afloramentos ferruginosos, cujas temperaturas podem ultrapassar 60° (Carmo, com.
pess.), a mortalidade de hospedeiras pode se intensificar durante o período seco devido
ao efeito combinado do parasitismo e do estresse hídrico a que estão sujeitas (Mourão et
al. 2009). A morte da atual hospedeira, sem que ocorra a dispersão para outra, significa
também a morte da parasita, e neste caso, investir em crescimento linear de ramos se
torna um ótimo investimento energético. Ao final da estação seca e início da chuvosa,
entre os meses de agosto e setembro, os ramos começaram a investir em novas folhas e
estruturas reprodutivas. O aumento da disponibilidade de água foi determinante para
que a parasita pudesse investir na produção de novas folhas. Com o acréscimo no
número de folhas, os ramos aumentaram a superfície fotossinteticamente ativa, o que
pode ter elevado a taxa de produção de energia (Larcher 2006). Isto permite o
investimento em estruturas reprodutivas, pois sua produção requer um elevado custo
energético (de Jong & Klinkhamer 2005).
Os padrões de alocação de recursos se tornaram mais claros quando os ramos
foram analisados quanto aos diferentes graus de parasitismo. Na estação seca, os ramos
que cresciam em hospedeiras mais parasitadas foram os que apresentaram maior
crescimento linear. Uma analogia pode ser feita entre o número de haustórios e a
ramificação de raízes de plantas durante a seca. Muitas plantas ramificam o sistema
radicular para suprir o déficit de água, que pode ser rapidamente perdida por
evapotranspiração (Jaleel et al. 2009). Comparativamente, a expansão das raízes aéreas
das plantas parasitas, que ocorre através do crescimento linear, teria a finalidade de
buscar novas fontes de recursos (colonização) e extrair mais nutrientes de suas
hospedeiras (estabelecimento e crescimento). Após o estabelecimento de novos
haustórios, os indivíduos captam mais recursos para investir em seu crescimento e
reprodução. Estudos afirmam que o número destes haustórios está correlacionado com o
crescimento da parasita e seus impactos sobre as hospedeiras (Tennakoon & Pate 1996,
Tennakoon et al. 1997, Aukema 2003). Isso sugere que hospedeiras muito parasitadas
devem abrigar poucas parasitas que estabelecem múltiplas conexões simultâneas ao
longo de seus galhos. Dessa forma, os poucos indivíduos parasitas poderiam desviar

72
maiores quantidades de recursos usados para o seu desenvolvimento. Em contrapartida
estas hospedeiras estariam sujeitas aos efeitos mais acentuados do parasitismo. Sabe-se
que indivíduos de Mimosa calodendron muito parasitados apresentam de 75 a 95% de
redução de sua cobertura foliar (Mourão et al. 2009). Assim, uma redução nas taxas
fotossintéticas é esperada e o estabelecimento de ramos de S.flexicaulis em outras
plantas seria vital para sua sobrevivência.
Na estação chuvosa o crescimento de ramos, número de folhas e estruturas
reprodutivas da parasita foi semelhante não importando a categoria de parasitismo.
Como a água deixa de ser um recurso limitante para as plantas, é possível que a retirada
de nutrientes das hospedeiras atinja seu máximo, permitindo um investimento similar
entre as categorias de parasitismo. Além disso, é esperado que o estresse hídrico e
nutricional das hospedeiras diminua com a chegada das chuvas, favorecendo a reposição
de folhas que foram perdidas durante o período seco, ampliando sua área
fotossiteticamente ativa. Como as hospedeiras mais parasitadas perdem mais folhas que
as pouco parasitadas (Mourão et al. 2009), o gasto de energia para a reposição desta
estrutura deverá ser maior, assim as hospedeiras muito parasitadas poderiam fornecer
tantos nutrientes quanto comparadas às pouco parasitadas. Sabe-se que no período seco
as plantas crescem pouco, reduzem sua área foliar disponível para fotossíntese (menor
tamanho das folhas ou abscisão), além de sofrerem uma série de modificações
fisiológicas e morfológicas para sobreviverem a essas condições (Crawley 2007).
Ao contrário do que se esperava a mortalidade de ramos não foi maior no período
seco. Nesta época a maioria dos ramos morreu devido ao esgotamento de hospedeiras
que muitas vezes (15%) não conseguiam sobreviver aos efeitos combinados do
parasitismo e escassez de agua. Além disso, a maior mortalidade de ramos internos
reforça a necessidade da parasita em investir em ramos que possam buscar por novas
fontes de recursos e sua dispersão. Já na estação chuvosa, a mortalidade de ramos
ocorreu principalmente pelo ataque de insetos galhadores, pulgões e larvas não
identificadas que cresciam no interior de muitos ramos. Os insetos podem causar
inúmeros prejuízos às plantas tais como redução no crescimento, aumentando a
vulnerabilidade ao ataque de outros insetos e doenças, reduzindo a habilidade
competitiva (Crawley 1989). Estes insetos podem ser uma importante fonte reguladora
do espalhamento de ramos das parasitas em comunidades naturais. Os ramos atacados
por insetos e que não morriam tinham seu crescimento comprometido, pois as larvas se

73
alimentavam de seu interior e o rompiam ao eclodir. Por esse motivo o crescimento
vegetativo na estação seca superou o da estação chuvosa.
Danos como as quebras da extremidade dos ramos principais, mais comuns nos
ramos exploratórios durante a estação seca, estimularam o desenvolvimento de ramos
laterais. As gemas danificadas perdem a dominância apical induzindo o crescimento de
novas partes (Cline 1997). É comum que plantas invistam no crescimento de novos
tecidos e estruturas para compensar as partes que foram perdidas (Harper 1985). Os
ramos exploratórios foram os que mais produziram ramos laterais e isso pode favorecer
a busca por novas hospedeiras da mesma ou de outras espécies.
O comportamento em relação ao crescimento linear, produção de estruturas
reprodutivas e folhas dos ramos principais internos e exploratórios foi semelhante nos
períodos de seca, sinalizando que a distribuição de recursos é proporcional, não
importando a posição de seus ramos na copa das hospedeiras. Já durante a estação
chuvosa os ramos exploratórios cresceram mais do que os internos e produziram mais
estruturas reprodutivas. Como os organismos clonais podem crescer assimetricamente,
priorizando partes do indivíduo que tem melhores condições desenvolvimento (Harper
1981, Harper 1985), é possível que os ramos mais expostos à luz tenham sido
beneficiados. Isso ocorre com raízes, por exemplo, que possuem uma diversidade de
respostas à heterogeneidade incluindo aumento de ramificações laterais, biomassa e
comprimento de raízes bem como a capacidade de retirada de nutrientes do solo (Hodge
2004).
Os principais efeitos do estresse hídrico nas plantas são as reduções nas taxas
fotossintéticas (Graves et al. 1992), redução na alocação de recursos afetando seu
crescimento (Crawley 2007) e diminuição no investimento reprodutivo (Mugabe 1983,
de Jong & Klinkhamer 2005). Além disso, os custos energéticos para a reprodução
esgotam os recursos necessários para manutenção e crescimento das plantas (Bazzaz et
al. 1987, de Jong & Klinkhamer 2005). S. flexicaulis alocou recursos de maneira
diferenciada durante as estações, adotando estratégias que favoreceram sua
sobrevivência. Como as parasitas assumem o controle homeostático de água das
hospedeiras, elas conseguem manter sua eficiência na captação de água e assimilação de
carbono (Glatzel & Geils 2009) mesmo durante o período de maior escassez de água.
Apesar disso, esta parasita demonstrou sinais de estresse hídrico durante a estação seca,
porque mesmo investindo em crescimento linear, não produziu estruturas reprodutivas,
que são energicamente mais custosas. Apenas na estação chuvosa, quando a agua não

74
era limitante, a parasita investiu em novas folhas e estruturas reprodutivas. O maior
investimento reprodutivo durante a estação chuvosa pode servir para garantir a
sobrevivência das hospedeiras.
Referências bibliográficas
Aukema, J. E. 2003. Vectors, viscin, and Viscaceae: mistletoes as parasites, mutualists
and resources. Frontiers in Ecology and the Environment, 1:212-219.
Barneby, R. C. 1991. Sensitivae censitae: a description of the genus Mimosa Linneaus
(Mimosaceae) in the New World. The New York Bootanical Gardens, New York,
835p.
Bazzaz, F. A.; Chiarielo, N. R.; Coley, P. D. & Pitelka, L. F. 1987. Allocation resources
to reproduction and defense. BioScience, 37:58-67.
Benites, V. M.; Shaefer, C. E. G. K.; Simas, F. N. B. & Santos, H. G. 2007. Soils
associated with rock outcrops in the Brazilian mountain ranges Mantiqueira and
Espinhaço. Revista Brasileira de Botânica, 4: 569-577.
Bloom,A. J.; Chapim III,F. S. & Mooney, H. A. 1985. Resource limitation in plants - an
economic analogy. Annual Review of Ecology and Systematics, 16:363-392.
Cline, M. G. 1997. Concepts and terminology of apical dominance. American Journal of
Botany, 84:1064-1069.
Crawley M. J. 1989. Insect herbivores and plant population dynamics. Annual Review
of Entomology, 34:531-64.
Crawley, M. J. 2007. Plant Ecology, 2 º Ed. 717 p. Ed. Blackwell Science, London.
de Jong, T. J., and P. G. L. Klinkhamer. 2005. Evolutionary Ecology of plants
Reproductive Strategies. University Press, Cambridge, UK.
Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D. & Basra, S. M. A. 2009. Plant drought
stress: effects, mechanisms and management. Agronomy for Sustainable
Development, 29:185-212.
Glatzel, G., & Geils, B. W. 2009. Mistletoe ecophysiology host-parasite interactions.
Botany, 87: 10-15.
Grace, J. B. 1993. The adaptive significance of clonal reproduction in angiosperms: an
aquatic perspective. Aquatic Botany. 44: 159-180.
Graves J. D., M. C. Press, S. Smith, and G. R. Stewart. 1992. The carbon economy of
the association between cowpea and the parasitic angiosperm Striga gesnerioides.
Plant, Cell Environmental, 15: 283–288.

75
Harper, J. L. 1981. The concept of population in modular organisms. In: Theoretical
ecology: principles and applications, 2 ed. P 53-77.
Harper, J. L. 1985. Modules, branches, and the capture of resources. In: Population
biology and evolution of clonal organisms (J.B.C. Jackson, L.W. Buss & R. Cook,
eds). Yale University Press, New Haven, p.1-34.
Hodge, A. 2004. The plastic plant: root responses to heterogeneous supplies of
nutrients. New Physiologist, 162:9-24.
Jacobi, C. M., F. F. Carmo, R. C. Vincent, and J. R. Stehmann. 2007. Plant communities
on ironstones outcrops: a diverse and endangered Brazilian ecosystem. Biodiversity
and Conservation, 16: 2185-2200.
Jaleel, C. A.; Gopi, R.; Sankar, B.; Gomathinayagam, M. & Panneerselvam, R. 2008.
Differential responses in water use efficiency in two varieties of Catharanthus roseus
under drought stress. Comptes Rendus Biologies, 1:42-47.
Jaleel, C. A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R. &
Panneerselvam, R. 2009. Drought stress in plants: A review on morphological
characteristics and pigments composition. International Journal of Agriculture &
Biology. 11:100-105.
Larcher, W. 2006. Ecofisiologia vegetal. São Carlos. 532p.
Lei, S. A. 2001. Survival and development of Phoradendron californicum and Acacia
greggii during a drought. Western North American Naturalist, 61: 78-84.
Marvier, M. A. 1996. Parasitic plant-host interactions: plant performance and indirect
effects on parasite-feeding herbivores. Ecology. 77:1398-1409.
McConnaughay, K. D. M & Coleman, J. S. 1999. Biomass allocation in plants:
ontogeny or optimality? A test along three resource gradients. Ecology, 80:2581-
2593.
Mooney, H. A.; Drake, B. G.; Luxmoore, R. J.; Oechel, W. C. & Pitelka, L. F. 1991.
Predicting ecosystem responses to elevated CO2 concentrations. BioScience, 41:96-
104.
Mourão, F. A.; Carmo, F. F; Ratton, P.; Jacobi, C. M. 2006. Hospedeiras da
hemiparasita Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) em campos
rupestres ferruginosos, Quadrilátero Ferrífero, MG. Lundiana, 7:103-110.
Mourão, F. A., Jacobi, C. M. Figueira, J. E. C & Batista, E. K. L. 2009. Effects of the
parasitism of Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) on the fitness of

76
Mimosa calodendron Mart. (Fabaceae), an endemic shrub from rupestrian fields over
ironstone outcrops, Minas Gerais State, Brazil. Acta Botânica Brasílica, 23:820-825.
Mugabe, N.R. 1983. Effects of Alectra vogelli Benth on cowpea (Vigna unguiculata (L.)
Walp). Some aspects on reproduction of cowpea. Zimbabwe. Journal of Agricultural
Research, 21:135-147.
Nimer, E. & Brandão, A. M.. 1989. Balanço Hídrico e clima da região dos Cerrados.
Instituto Brasileiro de Geografia e Estatística (IBGE), Rio de Janeiro, Brazil.
Poorter, H.; Remkes, C.& Lambers, H. 1990. Carbon and Nitrogen Economy of 24
Wild Species Differing in Relative Growth Rate. Plant Physiology, 94:621-627.
Press, M. C. & Phoenix, G. K. 2005. Impacts of parasitic plants on natural communities.
New Phytologist, 166:737-751.
Razmjoo, K.; Heydarizadeh, P. & Sabzalian, M. R. 2008. Effect of salinity and drought
stresses on growth parameters and essential oil contents of Matricaria chamomile.
International Journal of Agriculture e Biology 4:451-454.
Rizzini, C. T. 1980. Loranthaceae of the central Brazil. Arquivos do Jardim Botânico do
Rio de Janeiro,24: 19-50.
Rizzini, C. T. 1997. Tratado de Fitogeografia do Brasil: Aspectos Ecológicos,
Sociológicos e Florísticos 2 º ed. Âmbito Cultural Edições Ltda, Brazil.
Shaver, G. R. and Chapin, F. S., III, 1991: Production: biomass relationships and
element cycling in contrasting arctic vegetation types. Ecological Monographs, 61:
1–31
Shen, H.; Ye, W.; Hong, L.; Huang, H.; Wang, Z.; Deng, X.; Yang, Q.; Xu, Z. 2006.
Progress in parasitic plant biology: Host selection and nutrient transfer. Plant
Biology, 8: 175-185.
Silva, M. F. F.; Secco, R. S. & Lobo, M. G. A. 1996. Aspectos ecológicos da vegetação
rupestre da serra dos Carajás, estado do Pará, Brasil. Acta Amazonica, 26: 17-44.
Tennakoon, K. U. & Pate, J. S. 1996. Effects of parasitism by a mistletoe on the
structure and functioning of branches of its host. Plant, Cell and Environment,
19:517-528.
Tennakoon, K. U.; Pate, J. S. & Fineran, B. A. 1997. Growth and partitioning of C and
fixed N in the shrub legume Acacia litorea in the presence or absence of the root
hemiparasite Olux phyllanthus. Journal of Experimental Botany, 48:1047-1060.
Zar, J. H. 1999. Biostatistical Analysis. 4th Edition. Prentice-Hall, New York.

77
CAPÍTULO 4
Efeito top-down da hemiparasita Struthanthus
flexicaulis (Loranthaceae) e seu papel chave na
estruturação de comunidades vegetais em afloramentos
ferruginosos do Quadrilátero Ferrífero, Brasil.
Fabiana Alves Mourão1, Claudia Maria Jacobi
1, José Eugênio Côrtes Figueira
1 & Rafael
Barros1.
1 Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade
Federal de Minas Gerais.

78
Resumo
As plantas parasitas podem modificar a estrutura e dinâmica da comunidade onde estão
inseridas, reduzindo a biomassa e alterando a alocação de recursos das espécies
hospedeiras e as interações entre elas. O objetivo deste trabalho foi investigar a
importância da parasita Struthanthus flexicaulis (Loranthaceae) na estruturação de uma
comunidade vegetal sobre afloramento ferruginoso através do efeito top-down e
evidenciar se esta espécie é chave para este sistema. Foi realizado um levantamento de
espécies em duas áreas com diferentes densidades da parasita: A) alta (soma do
comprimento dos ramos da parasita superior a 180 m) e B) baixa (soma do
comprimento dos ramos da parasita inferior a 80 m), onde se mediu a cobertura vegetal
de cada indivíduo da comunidade durante a estação chuvosa, entre os meses de
fevereiro a abril de 2009. Foram amostradas cinco faixas de vegetação de 250m2 em
cada área, onde foi calculado o impacto da parasita (IP) sobre a perda de cobertura foliar
e mortalidade de hospedeiras. As densidades da parasita variaram de 3,62 a 6,4 m/m2
na
área A e 0,7 a 1,8 /m2
na área B. A composição florística da comunidade foi semelhante
entre as áreas, sendo formada por 20 famílias e 39 espécies na área A e 17 famílias e 38
espécies na B. O índice de diversidade de Shannon não apontou diferença entre a
comunidade com atuação da parasita e sem atuação da mesma. A leguminosa Mimosa
calodendron foi a mais representativa na comunidade, apresentando a maior cobertura
vegetal; também foi a espécie que mais perdeu (67%) sua cobertura com a atuação da
parasita. O efeito top-down foi negativo, pois o impacto da parasita sobre a cobertura
foliar variou de 39 a 66% na área A e 7 a 40% na B. S. flexicaulis foi considerada
espécie-chave na comunidade vegetal, apresentando valores de IP muito superiores a 1.
Os impactos que as parasitas provocam na comunidade vegetal poderiam ser
comparados aos de grandes herbívoros, já que ambos apresentaram elevado índice de
importância para as comunidades em que estão inseridos. S. flexicaulis reduziu
significativamente a cobertura vegetal de suas hospedeiras sem, entretanto, alterar a
diversidade e a composição de espécies. Sugere-se que seu papel chave e o efeito top-
down podem variar em função da densidade de plantas na paisagem.
Palavras-chave: comunidade vegetal, espécie-chave, interação planta-planta,
parasitismo.

79
Introdução
Compreender os fatores que determinam a manutenção das populações e a
estrutura de comunidades tem sido há várias décadas um dos grandes desafios da
ecologia (Chesson 2000). Após a obra clássica de Hairston et al. (1960), vários estudos
sobre cascata trófica que demonstraram o papel estruturador dos predadores (top-down)
e disponibilidade de recursos (bottom-up) foram publicados (Balciunas & Lawler 1995,
Hunter et al. 1997, Baum & Worm 2009). A maior parte desses estudos se concentrou
nos ambientes aquáticos porque são considerados habitats relativamente discretos e
homogêneos, estratificados e que geralmente mantêm fortes interações interespecíficas
(Polis et al. 2000). Em ambientes terrestres poucos estudos têm conseguido demonstrar
o efeito top-down (Schmitz et al. 2000, Costa et al. 2008), devido a sua maior
heterogeneidade, complexidade estrutural, e às interações entre as espécies serem
consideradas muitas vezes fracas e difusas (Strong 1992, Polis et al. 2000, Werner &
Peacor 2003). Nesses ambientes experimentos têm sido realizados através de cadeias
tróficas incompletas, dificilmente passando de três níveis tróficos e geralmente
envolvendo remoção ou adição de predadores (Chase 1998, Kneitel & Miller 2002,
Gruner 2004, Schmitz 2006, Nuttle et al. 2011). O efeito top-down em comunidades
terrestres têm sido atribuído principalmente à herbivoria, que pode alterar a biomassa
(Schmitz & Suttle 2001) e interferir na reprodução das plantas (Amsberry & Maron
2006), bem como modificar a diversidade local (Nuttle et al. 2011). Já outros grupos
que poderiam causar este efeito, como os parasitoides (Gomez & Zamora 1994), são
raramente mencionados nestes estudos.
Um grupo de organismos que poderia ser utilizado para investigar a atuação da
força top-down em comunidades vegetais é o das plantas parasitas. Elas podem
modificar a estrutura e dinâmica da comunidade onde estão inseridas, reduzindo a
biomassa e alterando a alocação de recursos das espécies hospedeiras (Tennakoon &
Pate 1996, Lei 2001, Press & Phoenix 2005, Shen et al. 2006). Como a produtividade e a
permanência destas parasitas na comunidade são dependentes da “qualidade” dos
recursos obtidos, elas são capazes de escolher entre suas potenciais hospedeiras aquelas
que ofereçam melhores condições de sobrevivência (Kelly 1992, Pennings & Callaway
2002, Press & Phoenix 2005). Esta escolha pode estar relacionada à abundância de
hospedeiras (Norton & Carpenter 1998) e seu tempo de permanência no ambiente
(perenes ou anuais) (Kelly et al. 1988), bem como suas concentrações de nitrogênio
(Radomiljac et al. 1999) ou por apresentarem sistema vascular facilmente acessível e/ou

80
com pouca capacidade de defesa (Press & Phoenix 2005). As parasitas podem afetar
negativamente processos fisiológicos, reprodutivos e ecológicos das plantas infectadas
(Press et al. 1999, Lei 1999, 2001, Mourão et al. 2009). Podem também interferir no
balanço de água e nutrientes de suas hospedeiras, reduzindo as taxas de fotossíntese e
respiração (Glatzel & Geils 2009) e, em algumas situações, podendo levá-las à morte
(Aukema 2003).
Apesar do efeito negativo nas espécies infectadas, as plantas parasitas podem
desempenhar um papel benéfico na comunidade controlando a expansão de algumas
espécies e beneficiando outras. O impacto na estrutura da comunidade pode ser grande,
principalmente se as espécies mais parasitadas forem as dominantes e o parasitismo
permitir a existência de espécies competitivamente subordinadas (Press 1998). As
plantas parasitas já foram consideradas espécies-chave em suas comunidades (Press &
Phoenix 2005) e até determinantes da diversidade local (Watson 2001). Além disso, as
plantas parasitas são utilizadas como fonte de recursos por uma variedade de
organismos, entre eles polinizadores e dispersores de sementes (Watson 2001, Aukema
2003, Kelly et al. 2004, Carlo & Aukema 2005, Azpeitia & Lara 2006). Foi sugerido que
a parasita Cuscuta salina poderia ser considerada chave na manutenção da diversidade
de espécies vegetais em marismas da Califórnia (Pennings & Callaway 1996).
Com exceção das referências acima, o papel ecológico das plantas parasitas têm
sido bastante ignorados em diversos estudos de comunidades vegetais (Norton & Reid
1997). Sabe-se que as plantas parasitas, com sua pequena biomassa de raízes sugadoras
de nutrientes, causam maior efeito em suas hospedeiras que herbívoros (Pennings &
Callaway 2002). As plantas parasitas podem alterar a o balanço competitivo entre
hospedeiras preferidas e outras plantas na comunidade (Callaway & Pennings 1998),
enquanto os herbívoros comumente alteram a composição de espécies (Bryant et al.
1991), podendo também alterar a composição de espécies caso o balanço competitivo
seja afetado.
O objetivo deste trabalho foi investigar a importância da parasita Struthanthus
flexicaulis (Loranthaceae) na estruturação das comunidades vegetais de afloramentos
ferruginosos através do efeito top-down e evidenciar se esta espécie pode ser considerada
chave nessas comunidades. Para isso foi formulada a seguinte hipótese: S. flexicaulis
influencia negativamente a permanência de algumas espécies nos afloramentos
ferruginosos, alterando a composição da comunidade. As predições que nortearam este
trabalho foram: 1) a parasita exerce efeito top-down significativo na comunidade vegetal

81
provocando a morte de indivíduos dominantes e favorecendo a diversidade; 2) a parasita
reduz a cobertura vegetal das hospedeiras, causando um grande impacto na cobertura
foliar das hospedeiras, na composição e diversidade de espécies, podendo ser
considerada uma espécie-chave. O termo top-down se refere originalmente ao controle
dos predadores sobre abundância de suas presas em uma cascata trófica (Hairston 1960).
Este controle pode afetar poucas espécies, onde os aumentos na abundância de um
predador provocam diminuições de suas presas sem que isso necessariamente afete a
comunidade como um todo. Já uma espécie-chave está entre as mais importantes e afeta
diretamente a comunidade seja através de mudanças na sua estrutura e diversidade,
processos ecológicos e interações (Power et al. 1996). De acordo com a definição
original de Paine (1969) espécie-chave refere-se a uma espécie que preferencialmente
consome e mantém sob controle outra que poderia dominar o sistema. A definição de
espécies-chave abordada neste trabalho está de acordo com Power et al. (1995) que as
descreve como sendo aquelas cujos efeitos em suas comunidades ou ecossistemas são
muito maiores do que o esperado por suas abundâncias ou biomassas.
Material e métodos
Local de estudo
Os estudos foram realizados em um dos afloramentos ferruginosos da porção sul
da Cadeia do Espinhaço localizado no Parque Estadual da Serra do Rola Moça, uma
unidade de conservação de proteção integral. Esta área está localizada dentro do
Quadrilátero Ferrífero (20o03’60”S, 44
o02’00”W, 1300m). Estes afloramentos são
caracterizados pela presença de metais pesados, baixa retenção de umidade, altas
temperaturas e escassez do solo (Benites et al. 2007). Seu substrato é muito compacto e
é composto por minério de ferro, de coloração marrom a negra, com baixíssima
erodibilidade, e porosidade e permeabilidade variáveis (Benites et al. 2007). Nessas
áreas os ventos são constantes e a radiação ultravioleta atinge altos índices, que agrava
durante a estação seca. Sob condições de elevadas temperaturas e baixa capacidade de
retenção de água, cresce uma vegetação baixa, composta por poucos indivíduos de porte
arbóreo, dominada por eudicotiledôneas como Asteraceae, Fabaceae e Myrtaceae, e
monocotiledôneas como Poaceae, Cyperaceae e Orchidaceae (Jacobi et al. 2007, Viana
& Lombardi 2007). O clima da região é do tipo mesotérmico, precipitação anual de
1000 a 1500 mm, correspondendo ao tipo Cwa na classificação de Köppen (Nimer &

82
Brandão 1989), com uma pronunciada estação seca de abril a setembro, e temperatura
média anual de 25°C (Rizzini 1997).
Espécie-alvo
Struthanthus flexicaulis pertence à família Loranthaceae e é considerada uma das
hemiparasitas brasileiras mais comuns, com ampla distribuição nas áreas de cerrado do
Brasil Central (Rizzini 1980). O hemiparasitismo é caracterizado pela capacidade da
planta em fabricar açúcares através da fotossíntese, mas depende da retirada de água e
nutrientes das hospedeiras por meio de raízes especializadas (haustórios) sendo
parcialmente dependentes destas. Esta espécie possui ramos longos, flageliformes,
folhas oblongo-abovadas, com flores e frutos pequenos (3-4 mm e 6-7 mm
respectivamente). Sua dispersão ocorre de duas maneiras: 1) ornitocoria, em que seus
frutos são consumidos pelas aves e as sementes depositadas nos galhos de suas
hospedeiras e 2) crescimento vegetativo, através do crescimento de ramos que podem
atingir a copa de novas hospedeiras. A parasita se fixa, através de raízes epicorticais
caulinares (Calvin & Wilson 2006), em diversas partes das hospedeiras tais como
caules, folhas e inflorescências (Mourão et al. 2006). Estes ramos emitidos podem
atingir a vizinha mais próxima e, através do contágio, dispersar vegetativamente.
Desenho amostral
Para avaliar a importância de cada espécie na comunidade e os impactos da
parasita sobre elas, foram selecionadas uma área com alta densidade de parasitas (Área
A) e uma com baixa densidade (Área B) do mesmo tamanho. Em cada área foram
traçados 5 faixas que mediram 1m x 50 m, totalizando 250 m2 e cada faixa foi dividida
em parcelas de 1m x 1m. Inicialmente a densidade da parasita foi estimada de modo
visual e posteriormente confirmada pela soma do comprimento dos seus ramos dividido
pela área da faixa (metros da parasita/250 m2). Para verificar se a densidade da parasita
diferia entre as áreas foi aplicado o teste t, uma vez que os dados apresentaram
distribuição normal. Em cada parcela das faixas foi registrada a riqueza, abundância de
indivíduos para cada espécie, e plantas parasitadas e/ou mortas. Plantas com
crescimento clonal como Orquidaceae, Cactaceae e Velloziaceae foram contadas como
único indivíduo quando os grupos da mesma espécie se encontravam isolados a uma
distância maior que cinco cm, não sendo possível identificar seu número exato. Além
disso, as Poaceae e Cyperaceae não foram identificadas, apenas tiveram suas coberturas

83
estimadas. O levantamento de espécies na comunidade foi realizado durante a estação
chuvosa, pois durante a seca há o desaparecimento de terófitas e muitas geófitas que
perdem suas partes aéreas, dificultando sua identificação. Este levantamento foi
realizado nos meses de fevereiro a abril de 2009.
Para avaliar a importância de cada espécie na comunidade foi calculada a
cobertura vegetal de cada indivíduo, através da projeção de suas copas sobre a
superfície do solo. Para as projeções que lembravam círculos foram tomadas medidas de
diâmetro máximo, as que lembravam triângulos mediam-se os lados e altura, quadrados
e retângulos mediam-se os lados e para formato elíptico mediam-se os diâmetros
máximos e mínimos. A determinação da cobertura vegetal de espécies permite
representar numericamente as áreas aproximadas ocupadas de cada espécie na
comunidade (Braun-Blanquet 1979). Estes valores foram somados para estimar o valor
total de área ocupada por cada espécie da comunidade. Foram relacionadas: 1) a
abundância das espécies que estiveram representadas por mais de 1% do total de
indivíduos na comunidade e o parasitismo; e 2) a cobertura sem a atuação da parasita
(CSEM) e cobertura com a atuação da parasita (CCOM) de todas as espécies que
tiveram a cobertura vegetal superior a 3% na comunidade.
Para estimar a cobertura da parasita foram coletados 10 ramos com o
comprimento igual a 1 metro, sendo 1 ramo por faixa e que se localizavam em Mimosa
calodendron, espécie mais parasitada em afloramentos ferruginosos. O ramo escolhido
possuía o maior diâmetro dentre os que parasitavam a hospedeira. Este procedimento
foi necessário para que se estimasse a maior área possível de cobertura da parasita.
Estes ramos tiveram suas folhas retiradas e contadas. A maior folha de cada ramo teve
sua área calculada, e este valor foi multiplicado pelo número total de folhas encontrado
em seu respectivo ramo para estimar a área média das folhas por metro da parasita. Em
campo, os ramos de S. flexicaulis foram medidos em seu diâmetro e comprimento.
Multiplicando o diâmetro por seu comprimento estimou-se a área ocupada pelo ramo e
a cada metro da parasita foi somado o correspondente ao valor médio da área ocupada
pelas folhas.
Para verificar a magnitude do efeito top-down por S. flexicaulis e se esta é uma
espécie-chave no sistema, foi calculado o impacto causado por sua presença sobre a
mortalidade de hospedeiras na comunidade fazendo uso da equação matemática proposta
por Power et al. (1996), que modelaram as relações predador-presa e os efeitos da
retirada da espécie (ou grupo) na estrutura da comunidade. A equação proposta por estes

84
autores é flexível e permite usar tanto a biomassa, a cobertura vegetal das espécies, a
abundância, produtividade ou riqueza de espécies, entre outras variáveis que consigam
exprimir a proporção da espécie-alvo (S. flexicaulis) em relação à comunidade que
estaria afetando. Fazendo uma adaptação da fórmula de Power et al. (1996), foi possível
estimar o efeito top-down que S. flexicaulis exerce sobre a comunidade vegetal. A
primeira expressão da fórmula multiplicada por 100 mede a porcentagem do impacto
causado pela parasita (top-down) (1). Para verificar se a parasita é uma espécie-chave,
fazendo uso da fórmula de Power et al. (1996) (2), foi verificado se a proporção de
cobertura vegetal de S. flexicaulis é muito menor em relação à cobertura vegetal da
comunidade.
IP= (CSEM-CCOM) / CSEM * 100 (1)
IP= │(CSEM-CCOM) / CSEM│ * [1/pi] (2)
Onde:
IP = Importância de S. flexicaulis na comunidade
CSEM = Comunidade sem a parasita. Em CSEM foi considerada a cobertura vegetal de
todos os indivíduos (vivos, parasitados e mortos) presentes na área.
CCOM = Comunidade com a parasita. Em CCOM foi considerada a cobertura vegetal
dos indivíduos vivos que estavam presentes nas faixas.
pi = proporção da cobertura vegetal da parasita na comunidade amostrada.
Os valores de cobertura de todas as espécies foram expressos em m2.
Se o valor de IP encontrado for igual a 0 (zero) ou tiver um valor próximo a 1
então a parasita não poderá ser considerada chave na comunidade vegetal. Para
espécie-chave, de acordo com Power et al. (1996), se a parasita tiver um efeito
desproporcional na comunidade então IP deverá ser muito maior que 1. Isto somente
ocorrerá se a cobertura vegetal de S. flexicaulis for desproporcionalmente inferior à da
comunidade e causar um grande impacto nas coberturas de suas hospedeiras. Neste caso
S. flexicaulis poderá ser considerada uma espécie-chave nesta comunidade.
Para verificar se a diversidade da comunidade se altera em função do parasitismo
foi calculado o índice de diversidade de Shannon H’ para: 1) comunidade com a
parasita (CCOM) e 2) comunidade sem a parasita (CSEM) nas áreas A e B. Os valores
de H’ foram expressos em nats/indivíduos. Estes valores foram comparados pelo teste t
de Hutcheson, que é o único teste recomendado para comparar os valores de H’ entre
áreas (Zar 2009).

85
Para comparar a composição de espécies foi calculado o índice de dissimilaridade
de Bray-Curtis para CCOM e CSEM nas áreas de maior e menor densidade da parasita,
conforme as considerações descritas acima. O índice de Bray-Curtis varia de 0 a 1 e as
áreas serão mais similares quanto maior for este valor. Este índice foi calculado para
duas variáveis: abundância de espécies e cobertura. Nas análises de abundância as
Poaceae e Cyperaceae foram excluídas, pois suas espécies não foram identificadas. Para
as famílias Orchidaceae, Cactaceae e Velloziaceae, agrupamentos de indivíduos da
mesma espécie isolados uma distância superior 5 cm, foram tratados como um único
indivíduo. Já para as análises de cobertura todas as espécies foram incluídas.
Resultados
Na área A as densidades da parasita variaram de 3,62 a 6,4 m/m2
(média= 4,96 ±
0,97) e a comunidade era composta por muitos arbustos (Tab. 1). Na área B, onde as
densidades da parasita variaram de 0,7 a 1,8 m/m2
(média=1,17 ± 0,31), houve
predominância de herbáceas e gramíneas, e os arbustos encontravam-se mais espaçados
na paisagem. As duas áreas diferiram quanto a densidade de parasitas (t = 7,39; n = 10;
p = 0,007). A cobertura total em ambas as áreas, entretanto, foi semelhante (veja adiante
Tab. 3) Houve uma correlação positiva entre a densidade de parasitas e arbustos
(rs=0,839; n=10; p<0,05).
Tabela 1. Densidade de parasitas e arbustos nas áreas A (alta densidade de parasitas) e B
(baixa densidade de parasitas) nas 5 faixas de 50 x 1 m2. (*) A densidade média de
espécies arbustivas foi calculada levando-se em consideração apenas indivíduos
lenhosos com diâmetro máximo de copas acima de 20 cm.
Faixas Área A S. flexicaulis (m)
Densidade (m de
S.flexicaulis/m2)
(*) Densidade média de
arbustivas/m2
1 181,00 3,62 2,4 ± 1,2
2 319,73 6,39 2,7 ± 1,5
3 266,06 5,32 3,1 ± 1,3
4 267,50 5,35 3,3 ± 2,1
5 207,39 4,15 2,7 ± 1,5
Faixas Área B
1 62,75 1,26 1,2 ± 1,1
2 78,20 1,56 1,9 ± 1,1
3 35,00 0,70 1,7 ± 0,9
4 46,74 0,93 1,5 ± 1,1
5 69,80 1,40 1,4 ± 1,1

86
Excluindo-se as gramíneas (Poaceae e Cyperaceae) a composição florística da
comunidade foi semelhante entre as áreas, sendo formada por 20 famílias e 39 espécies
na área A e 17 famílias e 38 espécies na área B (Anexo 2A e B). Tanto na área A quanto
na B, as famílias mais representativas foram Asteraceae com 7 e 8 espécies,
Orchidaceae com 4 e 6 espécies, Melastomataceae e Velloziaceae com 4 e 3 espécies
respectivamente. Foram encontrados 5.419 indivíduos na área A, sendo as espécies mais
abundantes a Orquidaceae Sophronitis cattleya (679), a Euphorbiaceae Microstachys
daphnoides (562), a Fabaceae Mimosa calodendron (500) e a Asteraceae Lychnophora
pinaster (415). Já na área B foram encontrados 4.871 indivíduos, sendo as espécies mais
abundantes S. cattleya (812), M. daphnoides (568), a Melastomataceae Tibouchina
heteromalla (400) e L. pinaster (391). Os valores de diversidade de Shannon não
apontaram diferenças entre as espécies da comunidade com a parasita (CCOM) e sem a
parasita (CSEM) (Tabela 2).
Tabela 2 - Valores de diversidade calculados para CSEM (comunidade sem a parasita) e
CCOM (com a parasita). S= número de espécies; H’= índice de diversidade de Shannon
e E’ = equitabilidade. Os números de 1 a 5 se referem às faixas. Foi usado o teste t de
Hutcheson para comparar os valores de diversidade da comunidade CSEM (H’CSEM)
com os da comunidade CCOM (H’CCOM).
Área A
(Alta densidade) S H'CCOM H'CSEM ECCOM E'CSEM teste t p
1 25 2,651 2,630 0,824 0,817 0,47 0,6
2 22 2,560 2,521 0,828 0,816 0,82 0,4
3 23 2,539 2,542 0,810 0,811 0,08 0,9
4 27 2,694 2,702 0,818 0,820 0,17 0,9
5 25 2,684 2,676 0,834 0,831 0,22 0,8
Área A 36 2,792 2,789 0,762 0,761 0,19 0,9
Área B (Baixa
densidade) S H'CCOM H'CSEM ECCOM E'CSEM teste t p
1 30 2,696 2,658 0,793 0,789 0,73 0,5
2 24 2,413 2,405 0,759 0,757 0,17 0,9
3 27 2,571 2,567 0,780 0,779 0,09 0,9
4 25 2,541 2,536 0,789 0,788 0,14 0,9
5 23 2,483 2,464 0,792 0,786 0,36 0,7
Área B 36 2,682 2,674 0,743 0,746 0,42 0,7
Considerando as áreas A e B, a espécie mais abundante foi M. daphnoides com
1.130 indivíduos, sendo apenas 39 parasitados. A espécie M. calodendron destacou-se,

87
representada por 858 indivíduos, dos quais mais de 57% estiveram parasitados (Fig. 1).
A composição das espécies tampouco variou. O índice de similaridade de Bray-Curtis
para CCOM e CSEM calculado com as abundâncias de espécies foi de 0,93 para a área
de alta densidade e de 0,97 para a de baixa densidade, e quando foi calculado com a
cobertura vegetal das espécies os valores foram 0,80 e 0,89 respectivamente.
0 5 10 15 20 25
Abundância (%)
0
10
20
30
40
50
60
Para
siti
smo
(%
)
Cm Cr
Bre
BorVel
Mdap
Mim
Lyc
TibStac
Sym
Fig. 1 – Relação entre o parasitismo e a abundância de 11 espécies na comunidade cuja
abundância relativa foi superior a 1%. Mim = Mimosa calodendron; Lyc =
Lychnophora pinaster; Mdap = Microstachys daphnoides; Bre = Baccharis reticularia;
Stac = Stachytarpheta glabra; Tib = Tibouchina heteromalla; Vel = Vellozia compacta;
Sym = Symphyopappus reticulatus; Cr = Croton serratoideus.; Cm = Cromolaena sp.;
Bor = Borreria sp.
De modo geral, em toda a comunidade (áreas A + B) todas as espécies arbustivas
sofreram reduções em suas coberturas vegetais devido à atuação do parasitismo (Fig. 2).
A espécie M. calodendron foi a que apresentou maior cobertura e também a que mais
sofreu com o parasitismo, pois perdeu 57% de sua cobertura total na comunidade. Esta
espécie é perene, fixadora de nitrogênio e muito abundante na área de estudo. Já L.
pinaster, representada por 806 e M. daphnoides com 562 indivíduos, tiveram uma
redução de 23% e 3,4% de suas coberturas vegetais, respectivamente.

88
Fig. 2 – Atuação do parasitismo sobre a cobertura vegetal das 14 espécies que estiveram
representadas por 3m2 ou mais na comunidade. Mim = Mimosa calodendron; Lyc =
Lychnophora pinaster; Poa = Poaceae; Mdap= Microstachys daphnoides; Bret =
Baccharis reticularia; Sop = Sophronitis cattleya; Stac = Stachytarpheta glabra; Tib =
Tibouchina heteromalla; Eri = Eriope macrostachya; vel= Vellozia compacta; Sym =
Symphyopappus reticulatus; Aci = Aciantera teres; velab = Vellozia albiflora; Micr =
Microlicia sp1.; Bser= Baccharis serrulata.
Representantes herbáceas e rasteiras como as Poaceae, Orchidaceae,
Velloziaceae e Cyperaceae que eventualmente foram parasitadas, entretanto, não
tiveram suas coberturas alteradas.
Quando as áreas A e B foram analisadas separadamente notou-se que a
identidade e a cobertura das espécies pouco se alteraram (Fig. 3). Entretanto, L. pinaster
foi a espécie mais representativa na área B e perdeu menos cobertura que M.
calodendron, que possuía a segunda maior cobertura. Na área de maior densidade (A),
todas as espécies sofreram reduções em suas coberturas em decorrência da mortalidade
pelo parasitismo (Fig. 3A). O mesmo não ocorreu na área de menor densidade (B) em
que espécies como T. heteromalla, Symphyopapus brasiliensis, Acianthera teres, S.
cattleya quase não sofreram perdas de suas coberturas (Fig. 3B).
Mim
Lyc
Poa
Mdap
Bret
Sop
Stac
Tibo
vel
Sym
Aci
velab
Cyp
Crot
Crom
Bor
Esp
écie
01020304050607080
Cobertura sem atuação da parasita (m²)
0 10 20 30 40 50 60 70 80
Cobertura com atuação da parasita (m²)

89
Fig. 3 – Atuação do parasitismo sobre a cobertura vegetal das espécies que estiveram
representadas por 3m2 ou mais na comunidade. A) Área de maior densidade da parasita
e B) Área de menor densidade da parasita. Aci = Aciantera teres; Bret = Baccharis
reticularia; Eri = Eriope macrostachya; Lyc = Lychnophora pinaster; Mdap=
Microstachys daphnoides; Mim = Mimosa calodendron; Poa = Poaceae; Sop =
Sophronitis cattleya; Stac = Stachytarpheta glabra; Sym = Symphyopappus reticulatus;
Tib = Tibouchina heteromalla; vel= Vellozia compacta.
O efeito top-down de S. flexicaulis sobre a comunidade vegetal esteve
relacionado à densidade de parasitas (Tab. 3). Nas faixas da área B este efeito foi
menor, pois a perda de cobertura foliar das espécies da comunidade variou de 7 a 40% e
seu valor médio foi de 22%. A faixa 3 foi a que menos sofreu este efeito, pois teve a
menor densidade de parasitas (Tab. 1). O contrário ocorreu nas faixas 1 e 2, de maiores
densidades de S. flexicaulis, que tiveram os maiores valores de perda de cobertura (Tab.
Lyc
Mim
Poa
Mdap
Bret
Tib
Stac
Sop
vel
Sym
Aci
Esp
écie
01020304050607080
Cobertura sem atuação da parasita (m²)
0 10 20 30 40 50 60 70 80
Cobertura com atuação da parasita (m²)
Mim
Lyc
Poa
Mdap
Sop
Bret
Stac
Eri
vel
SymE
spécie
01020304050607080
Cobertura sem atuação da parasita (m²)
0 10 20 30 40 50 60 70 80
Cobertura com atuação da parasita

90
1 e 3). Já na área A o efeito foi maior (Tab. 1 e 3). A perda de cobertura foliar foi 2,3
vezes maior que na área B e variou de 39 a 66%, com média de 50%.
Tabela 3- Magnitude do efeito top-down baseada em Power et al. (1996). CSEM
representa a cobertura vegetal da comunidade sem a parasita e CCOM representa a
cobertura vegetal da comunidade com a parasita. Os números de 1 a 5 se referem às
faixas.
Área A (Altas
densidades) CSEM (m
2) CCOM (m
2) (CSEM-CCOM)/CSEM top-down
1 13,69 19,65 -0,44 -43,54
2 25,43 42,18 -0,66 -65,87
3 22,27 34,44 -0,55 -54,65
4 29,56 41,20 -0,39 -39,38
5 22,24 32,65 -0,47 -46,81
Área A 113,19 170,12 -0,50 -50,30
Área B (Baixas
densidades) CSEM (m
2) CCOM (m
2) (CSEM-CCOM)/CSEM top-down
1 23,56 32,91 -0,40 -39,69
2 18,89 26,05 -0,38 -37,90
3 34,62 37,11 -0,07 -7,19
4 18,91 23,25 -0,23 -22,95
5 22,12 24,29 -0,10 -9,81
Área B 118,1 143,61 -0,22 -22,00
A parasita S. flexicaulis apresentou valores de IP indicativos de ser espécie-
chave na comunidade vegetal (Tabela 4). Os valores do IP variaram muito, de 10,85 a
51,94, nas faixas de baixas densidades (área B) de parasitas, sendo que a média para
esta área foi de 30. Já para as faixas de maiores densidades (área A) os valores do IP
variaram de 11,04 a 17,63 e a média para a área foi de 15,39. A cobertura vegetal da
comunidade sem a atuação parasita (CSEM) foi semelhante entre as áreas, mas a
cobertura da parasita foi mais de 4 vezes maior em A que em B, resultando em valores
de IP menores (Tabela 4).

91
Tabela 4- Resultados da aplicação da equação matemática proposta por Power et al.
(1996) para espécie-chave. CSEM = cobertura vegetal da comunidade sem a parasita;
CCOM = cobertura vegetal da comunidade com a parasita; Parasita = cobertura vegetal
da parasita; pi = proporção da cobertura vegetal da parasita. Os números de 1 a 5 se
referem às faixas.
Área A CSEM (m2) CCOM (m
2) Parasita (m
2) pi 1/pi Espécie-chave (IP)
1 13,69 19,65 0,54 0,039 25,35 11,04
2 25,43 42,18 0,95 0,037 26,77 17,63
3 22,27 34,44 0,79 0,035 28,19 15,41
4 29,56 41,20 0,80 0,027 36,95 14,55
5 22,24 32,65 0,62 0,028 35,87 16,79
Total Área A 113,19 170,12 3,7 0,033 30,59 15,39
Área B
1 23,56 32,91 0,18 0,008 130,89 51,94
2 18,89 26,05 0,23 0,012 82,13 31,13
3 34,62 37,11 0,10 0,003 346,20 24,90
4 18,91 23,25 0,14 0,007 135,07 31,00
5 22,12 24,29 0,20 0,009 110,60 10,85
Total Área B 118,1 143,61 0,85 0,007 138,94 30,01
Discussão
Os efeitos do parasitismo numa comunidade vegetal são dependentes da
densidade de plantas e a preferência da parasita por hospedeiras. Se por um lado a
produtividade e a permanência das plantas parasitas são dependentes da “qualidade” dos
recursos obtidos (Press & Phoenix 2005), hospedeiras muito parasitadas podem morrer
(Aukema 2003). Quando as plantas mais atacadas são competitivamente subordinadas, a
diversidade de espécies na comunidade poderá diminuir (Press & Phoenix 2005,
Grewell 2008). Por outro lado, se as mais atacadas forem as dominantes, é esperado um
aumento na diversidade local (Press 1998, Press & Phoenix 2005). A atuação da
parasita Cuscuta salina, por exemplo, modificou negativamente a riqueza e a
diversidade de plantas em marismas no norte da Califórnia (Pennings & Callaway
1996).
Em teoria S. flexicaulis poderia favorecer o aumento da diversidade local, pois
parasita preferencialmente a hospedeira M. calodendron, uma das espécies dominantes
dos afloramentos ferruginosos e possivelmente uma forte competidora. Entretanto, a
morte desta hospedeira poderia afetar negativamente a composição das espécies na

92
comunidade e até diminuir a diversidade, pois conforme sugerido por Jacobi et al.
(2007) M. calodendron seria uma espécie babá e auxiliaria no recrutamento de outras
espécies. Após considerar a atuação da parasita, verificou-se que a composição florística
da comunidade, tanto na área de maior quanto na de menor densidade da parasita, não
foi alterada. S. flexicaulis reduziu significativamente a cobertura vegetal de suas
hospedeiras sem que isso afetasse a diversidade de espécies. Embora a comunidade
vegetal não tenha sido acompanhada ao longo do tempo, estima-se que as espécies
parasitadas tenham morrido no período máximo de 5 anos. Isso por que os vestígios da
parasita estavam bastante visíveis e as espécies parasitadas demoram de 2 a 3 anos para
morrer (F. A. Mourão, obs. pess.). Dessa forma, seria possível que a comunidade
vegetal sofresse mudanças negativas em sua composição e diversidade de espécies no
decorrer de vários anos. Mas novos estudos precisam ser realizados para averiguar as
possíveis mudanças nesta comunidade.
Estudos que analisam o efeito top-down em comunidades terrestres têm
enfatizado as causas e consequências das variações de biomassa (Oksanen 1990, Dyer
& Letourneau 2003). Uma analogia pode ser feita com o estudo de Mourão e
colaboradores (2009) quando confirmaram que hospedeiras muito parasitadas
apresentavam de 75 a 95% de redução da cobertura vegetal. Para algumas espécies
como L. pinaster e M. calodendron foram nítidas as variações de cobertura vegetal após
a atuação da parasita. Esta última espécie foi que mais sofreu com o parasitismo na
comunidade. Por ser capaz de fixar nitrogênio, M. calodendron teria uma vantagem
competitiva sobre as demais plantas podendo se desenvolver e espalhar em locais com
sérias restrições nutricionais, já que as leguminosas são capazes de elevar o pH do solo
e favorecer a absorção de nutrientes necessários ao seu desenvolvimento (Nascimento et
al. 2003). Entretanto, as populações de M. calodendron são controladas por S.
flexicaulis, que reduzem o investimento reprodutivo desta hospedeira (Mourão et al.
2009), afetando o recrutamento de novos indivíduos na população e levando-as morte
por parasitismo. Isso porque M. calodendron é, dentre as potenciais hospedeiras, a que
mais poderia atender as necessidades nutricionais da parasita, já que o nitrogênio é um
recurso limitante para o desenvolvimento de plantas (Crawley 2007). Como há uma
relação direta e positiva entre a disponibilidade de nitrogênio e ganho de biomassa
(Crawford 1995, Ângre 1985, Cechim & Fumis 2004, Larcher 2006), ao parasitar as
leguminosas, S. flexicaulis poderia investir mais em crescimento e estruturas
reprodutivas. Outras espécies abundantes, mas pouco parasitadas como M. daphnoides e

93
T. heteromalla, por exemplo, não tiveram uma perda da cobertura vegetal tão
significativa. Estes resultados demonstram que a importância do efeito top-down pode
ser alterada em função da abundância e a composição de espécies na comunidade, em
função das “escolhas” das parasitas.
Apesar de este estudo tratar de interações de plantas em apenas dois níveis
tróficos, efeitos indiretos em outros níveis não mensurados neste trabalho podem
ocorrer. Assim como a adição de animais parasitas em teias tróficas pode aumentar a
riqueza de espécies, o número de ligações, os níveis tróficos e comprimento da teia
(Thompson et al. 2005), as plantas parasitas podem modificar as interações entre suas
hospedeiras e as demais espécies da cadeia trófica, podendo interferir na estabilidade,
nas forças de interação e no fluxo de energia (Lafferty et al. 2008). Como as espécies
parasitadas perdem cobertura vegetal, as aves que as utilizam como recursos tais como
alimento, poleiros e local para nidificação, poderiam se tornar mais vulneráveis à
predação. As aves da família Tyranidae Polystictus superciliaris, Elaenia cristata e
Embernagra longicauda, por exemplo, que se alimentam de insetos capturados nos
troncos de arbustos e constroem seus ninhos em galhos de M. calodendron (Hoffman et
al. 2009a, Hoffman et al. 2009b, Hoffmann & Rodrigues 2011) poderiam ser
prejudicadas. Por outro lado S. flexicaulis pode servir como fonte de recurso para aves
como E. cristata, que também se alimentam de frutos da parasita (Guerra 2005), insetos
como as formigas Camponotus crassus (Formicidae), que coletam o visco de sementes
da parasita recém-depositadas nos galhos das hospedeiras (Mourão obs. pessoal), e
polinizadores.
S. flexicaulis foi considerada espécie chave na comunidade estudada, segundo os
preceitos de Power e colaboradores (1996). Os valores de IP e de top-down foram
evidentes e variaram conforme diferentes densidades de arbustos. Na área A, que teve
quase o dobro de espécies arbustivas/m2 em relação à B, fazendo com que as distâncias
entre copas vizinhas fossem menores, o que possivelmente favoreceu o contágio de
outras plantas, elevando os valores de top-down. Entretanto, como a parasita também
apresentou maiores coberturas nestas áreas, os valores para espécie-chave foram
menores que em B. Já na área B, as distâncias entre as copas intuitivamente são maiores
e a dispersão vegetativa dos ramos da parasita pode ter sido dificultada, reduzindo a
importância do efeito top-down. As parasitas que se estabelecem em hospedeiras
isoladas, a mais de 20 cm de outras plantas (F. A. Mourão, dados não publicados),
dificilmente infectam outras plantas por dispersão vegetativa, pois quanto maior a

94
distância entre copas menor a chance de transmissão de ramos para outras hospedeiras.
O mesmo ocorre com a dispersão ornitocórica, pois a proximidade entre arbustos tende
a atrair mais aves, intensificando a deposição de sementes na área (Overton 1996). Se
este espalhamento for mais abrangente um efeito top-down maior é esperado, pois mais
plantas serão atacadas simultaneamente. O efeito top-down variou em função da
distribuição espacial de plantas na paisagem, pois a menor distância entre as
hospedeiras favorece o espalhamento de S. flexicaulis intensificando seus efeitos sobre a
comunidade vegetal. Além disso, a maior cobertura vegetal da parasita garantiu os
maiores valores deste efeito. Já a importância da parasita como espécie-chave variou
em função da perda de cobertura vegetal em relação à cobertura vegetal da parasita. Ao
contrário do efeito top-down, os valores de espécie-chave foram mais elevados onde a
pouca cobertura vegetal da parasita causou um grande efeito na cobertura das
hospedeiras. Os valores de espécie-chave refletiram bem esta relação uma vez que em
áreas de maior densidade de hospedeiras a parasita teve também maiores valores de
cobertura.
Neste trabalho os valores de IP que caracterizaram os impactos da parasita sobre
a vegetação foram até maiores que os valores encontrados por Knapp e colaboradores
(1999) para os grandes herbívoros das pradarias Norte Americanas (5 a 25). Eles
quantificaram os impactos diretos e indiretos causados por bisões que se alimentavam
seletivamente de espécies vegetais. Estes herbívoros alteraram as características físicas
e químicas do ambiente, aumentaram a heterogeneidade temporal e espacial da
vegetação e afetaram uma variedade de processos ecossistêmicos. Também,
diferentemente das parasitas, os bisões foram capazes de modificar a diversidade da
comunidade vegetal. Contudo, mesmo que a perda de espécies não tenha sido notada, a
parasita suprimiu consideravelmente a cobertura vegetal de plantas dominantes, que
pode ter ocorrido devido à constituição peculiar desta comunidade caracterizada por
elevada abundância e cobertura de M. calodendron. De maneira semelhante aos bisões,
que aumentam a quantidade de nutrientes no ecossistema, a queda acentuada de folhas
provocadas pelo parasitismo poderia contribuir para acelerar a entrada de nutrientes no
sistema. Isso também ocorreu em florestas temperadas de eucaliptos, cuja parasita
Amyema miquelli aumentou o retorno de nutrientes na serapilheira, duplicando as taxas
de nitrogênio e quadruplicando as taxas de fósforo (March & Watson 2010). S.
flexicaulis também poderia alterar a heterogeneidade espacial da paisagem e reduzir o
potencial competitivo de M. calodendron, já que hospedeiras muito infectadas morrem

95
com o passar do tempo.
Apesar de S. flexicaulis não ter alterado a diversidade e a composição de
espécies, foi capaz de modificar substancialmente a cobertura vegetal da hospedeira
dominante, o que sugere modificações espaciais na comunidade a longo prazo. A
variação do efeito top-down da parasita sobre a comunidade vegetal dos afloramentos
ferruginosos foi dependente da densidade de hospedeiras e contribuiu para sua atuação
como espécie-chave. Sua atuação pode contribuir para o aumento da heterogeneidade
ambiental, afetar a dinâmica das populações de suas hospedeiras e modificar as
interações entre as espécies, interferindo assim no funcionamento da comunidade.

96
Referências bibliográficas
Amsberry, L. K. & J. L. Maron. 2006. Effects of herbivore identity on plant fecundity.
Plant Ecology, 187:39–48.
Ågren, G. I. 1985. Theory for growth of plants derived from the nitrogen productivity
concept. Physiologia Plantarum, 64:17–28.
Aukema, J. E. 2003. Vectors, viscin, and Viscaceae: mistletoes as parasites, mutualists
and resources. Frontiers in Ecology and the Environment, 1:212-219.
Azpeitia, F. & Lara, C. 2006. Reproductive biology and pollination of the parasitic plant
Psittacanthus calyculatus (Loranthaceae) in central México. Journal of the Torrey
Botanical Society, 133:429–438.
Balciunas, D. & Lawler, S. P 1995. Effects of basal resources, predation, and alternative
prey in microcosm food chains. Ecology, 76:1322-1336.
Baum, J. K. & Worm, B. 2009. Cascading top-down of changing oceanic predator
abundances. Journal of Animal Ecology, 3:1-16.
Benites, V. M.; Shaefer, C. E. G. K.; Simas, F. N. B. & Santos, H. G. 2007. Soils
associated with rock outcrops in the Brazilian mountain ranges Mantiqueira and
Espinhaço. Revista Brasileira de Botânica, 4:569-577.
Braun-Blanquet, J. 1979. Fitosociologia, bases para el estudio de las comunidades
vegetales. H. Blume (ed.), Rosario, Madrid, 819 pp.
Bryant, J. P.; Provenza, F. D.; Pastor, J.; Reichardt, P. B.; Clausen, T. P. & du Toit, J. T.
1991. Interactions between woody plants and browsing mammals mediated by
secondary metabolites. Annual Review Ecological and Systemathics, 22:431-446.
Callaway, R. M. & Penning, S. C. 1998. Impact of a parasitic plant on the zonation of
two salt marsh perennials. Oecologia, 114:100-105.
Calvin, C. L. & Wilson, C. A. 2006. Comparative morphology of epicortical roots in
Old and New World Loranthaceae with reference to root types, origin, patterns of
longitudinal extension and potential for clonal growth. Flora, 201:345-353.
Carlo T. A, Aukema J. E. 2005. Female-directed dispersal and facilitation between a
tropical mistletoe and a dioecious host. Ecology 86: 3245–3251.
Cechim, I. & Fumis, T. F. 2004. Effect of nitrogen supply on growth and photosynthesis
of sunflower plants grown in the greenhouse. Plant Science, 166:1379-1385.
Chase, J. 1998. Central place forager effects on the food chain dynamics and special
pattern in Northern California meadows. Ecology, 79: 1236-1245.

97
Chesson, P. 2000. Mechanisms of maintenance of species diversity. Annual Review of
Ecology and Systematics, 31:343 -366.
Crawford, N. M. 1985. Nitrate: nutrient and signal for plant growth. The Plant Cell,
Rockville, 7:859-868.
Crawley, M. J. 2007. Plant Ecology, 2 º Ed. 717 p. Ed. Blackwell Science, London.
de Jong, T. J., & P. G. L. Klinkhamer. 2005. Evolutionary Ecology of Plant
Reproductive Strategies. University Press, Cambridge, UK.
Dyer, L.A. & Letourneau, D. 2003. Top-down and bottom-up diversity cascades in
detrital vs. living food webs. Ecology Letters, 6:60-68.
Glatzel, G. & Geils, B. W. 2009. Mistletoe ecophysiology host-parasite interactions.
Botany, 87:10-15.
Gómez, J. M. & Zamora, R. 1994. Top-down effects in a tritrophic system: Parasitoids
enhance plant fitness. Ecology, 75:1023-1030.
Grewell, B. J. 2008. Parasite facilitates plants species coexistence in costal wetland.
Ecology 89:1481-1488.
Gruner, D. S. 2004. Attenuation of top-down and bottom-up forces in a complex
terrestrial community. Ecology, 85:3010-3022.
Guerra, T. J. A. 2005 Componentes quantitativos e qualitativos da dispersão de
sementes de Struthanthus flexicaulis (Loranthaceae) em uma área de campo
rupestre do sudeste brasileiro. Dissertação de mestrado Universidade Estadual
Paulista, Rio Claro, 64 pp.
Hairston, N. G.; Smith, F. E. & Lawrence, B. S. 1960. Community structure, population
control and competition. The American Naturalist, 94:421-425.
Hoffman, D.; Lopes, L. E. & Vasconcelos, M. F. 2009a. Natural history notes on the
Pale-throated Serra-finch (Embernagra longicauda) in eastern Brazil. Ornitologia
Neotropical, 20:597-607.
Hoffman, D.; Gomes, H. B. & Guerra, T. 2009b. Biologia reprodutiva de Elaenia
cristata Pelzeln, 1868 (Passeriformes: Tyrannidae) em duas áreas de campo
rupestre de Minas Gerais, Brasil. Revista Brasileira de Ornitologia, 17:102-106.
Hoffmann, D.; Vasconcelos, M. F.; Lopes, L. E. & Rodrigues, M. 2007. Comportamento
de forrageamento e dieta de Polystictus superciliaris (Aves, Tyrannidae) no sudeste
do Brasil. Iheringia Série Zoologia, 97:296-300.
Hunter, M. D.; Varley, G. C. & Gradwell, G. R. 1997. Estimating the relative roles of
Top-down and Bottom-up forces on insect herbivore population: a classic study

98
revisited. Proceedings of the National Academy of Sciences of the United States of
America, 17:9176-9181.
Jacobi, C. M.; Carmo, F. F.; Vincent, R. C. & Stehmann, J. R. 2007. Plant communities
on ironstones outcrops: a diverse and endangered Brazilian ecosystem. Biodiversity
and Conservation, 16: 2185-2200.
Kelly, C. K.; Venable, D. L. & Zimmerer, K. 1988. Host specialization in Cuscuta
costaricensis. An assessment of use relative to host availability. Oikos, 53: 315-
320.
Kelly, K. C. 1992. Resource choice in Cuscuta europaea. Proceedings of the National
Academy of Sciences, 89:12194-12197.
Kelly, D.; Ladley, J. J. & Robertson, A. W. 2004. Is dispersal easier than pollination?
Two tests in New Zealand Loranthaceae. New Zealand Journal of Botany, 42:89-
103.
Knapp, A. K.; Blair, J. M.; Briggs, J. M.; Collins, S. L.; Hartnett, D. C.; Johnson, L. C.
& Towne, E. G. 1999. The keystone role of bison in North American Tallgrass
Prairie. Bioscience, 49:39-50.
Kneitel, J. M. & Miller, T. E. 2002. Resource and top-predator regulation in the pitcher
plant (Sarracenia purpurea) inquiline community. Ecology, 83:680-688.
Lafferty, K. D.; Allesina, S.; Arim, M.; Briggs , C. J.; De Leo, G. Dobson, P.; Dunne, J.
A.; Johnson,P. T. J.; Kuris, A. M.; Marcogliese, D. J.; Martinex, N. D.; Memmott,
J.; Marquet, P. A.; McLaughlin, J. P.; Mordecai, E. A.; Pascual, M.; Poulin, R. &
Thieltges, D. W. 2008. Parasites in food webs: the ultimate missing links. Ecology
Letters, 11:533-546.
Larcher, W. 2006. Ecofisiologia vegetal. São Carlos. 532p.
Lei, S. A. 1999. Age, size and water status of Acacia gregii influencing the infection
and reproductive success of Phoradendron californicum. American Midland
Naturalist, 141: 358-365.
Lei, S. A. 2001. Survival and development of Phoradendron californicum and Acacia
greggii during a drought. Western North American Naturalist, 61: 78-84.
March, W. A. & Watson, D. M. 2010. The contribution of mistletoe to nutrient returns:
Evidence for a critical role in nutrient cycling. Austral Ecology, 35:713-721.
Mourão, F. A.; Carmo, F. F; Ratton, P.; Jacobi, C. M. 2006. Hospedeiras da
hemiparasita Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) em campos
rupestres ferruginosos, Quadrilátero Ferrífero, MG. Lundiana, 7:103-110.

99
Mourão, F. A., Jacobi, C. M. Figueira, J. E. C & Batista, E. K. L. 2009. Effects of the
parasitism of Struthanthus flexicaulis (Mart.) Mart. (Loranthaceae) on the fitness of
Mimosa calodendron Mart. (Fabaceae), an endemic shrub from rupestrian fields over
ironstone outcrops, Minas Gerais State, Brazil. Acta Botânica Brasílica, 23:820-825.
Nascimento, J. T.; Silva, I. F.; Santiago, R. D. & Silva Neto, L. F. 2003. Efeito de
leguminosas nas características químicas e matéria orgânica de um solo degradado.
Revista Brasileira de Engenharia Agrícola Ambiental, 7:457-462.
Nimer, E. & Brandão, A. M. 1989. Balanço Hídrico e clima da região dos Cerrados.
Instituto Brasileiro de Geografia e Estatística (IBGE), Rio de Janeiro, Brasil.
Norton, D. A. & Carpenter, M. A. 1998. Mistletoes as parasites: host specificity and
speciation. Trends in Ecology and Evolution, 13:101-105.
Norton, D.A. & Reid, N. 1997. Lessons in ecosystem management from management of
threatened and pest loranthaceous mistletoes in New Zealand and Australia.
Conservation Biology, 11:759-769.
Nuttle, T; Yerger, E. H.; Stoleson, S. H & Ristau, T. 2011. Legacy of top-down
herbivore pressure ricochets back up multiple trophic levels in forest canopies over
30 years. Ecosphere, 2:1-11.
Oksanen L. 1990. Predation, herbivory, and plant strategies along gradients of primary
productivity. In: Perspectives on plant competition, pp. 445-474. Tilman, D. &
Grace, J. (Eds.). Academic Press, New York.
Overton, J. M. 1996. Spatial autocorrelation and dispersal in mistletoes: Field and
simulation results. Vegetatio, 125: 83-98.
Paine, R. T. 1969. A note on trophic complexity and community stability. American
Naturalist 103: 91-93.
Pennings, S. C. & Callaway, R. M. 1996. Impacts of parasitic plant on the structure and
dynamics of salt marsh vegetation. Ecology. 77:1410-1419.
Pennings, S. C.; Callaway, R. M. 2002. Parasitic plants: parallels and contrasts with
herbivores. Oecologia, 131:479-489.
Polis, G.A; Sears, A.L.W.; Huxel, G.R; Strong, D. R & Maron, J. 2000. When is a
trophic cascade a trophic cascade? Tree, 15:473-475.
Power, M. E. & Mills, L. S. 1995. The keystone cops meet in Hilo. TREE, 10: 182-184.
Power, M. E.; Tilman, D.; Estes, J. A.; Menge, B. A.; Bond, W. J.; Scott Mills, L.;
Daily, G.; Castilla, J. C.; Lubchenco, J. & Paine, R. T. 1996. Challenges in the
quest for keystones. Bioscience, 8: 609-620.

100
Press, M. C. & Phoenix, G. K. 2005. Impacts of parasitic plants on natural communities.
New Phytologist, 166: 737-751.
Press, M. C. 1998. Dracula or Robin Hood? A functional role for root hemiparasites in
nutrient poor ecosystems. Oikos. 82: 609-611.
Press M. C., Scholes J. D. & Barker, M. G. 1999. Plant physiological ecology. Oxford,
UK: Blackwell Science Ltd.
Radomiljac, A. M.; McComb, J. A. & Pate, J. S. 1999. Gas exchange and water
relations of the root hemiparasite Santalum album L. in association with legume
and non-legume hosts. Annals of Botany, 83: 215-224.
Rizzini, C. T. 1980. Loranthaceae of the central Brazil. Arquivos do Jardim Botânico do
Rio de Janeiro, 24: 19-50.
Rizzini, C. T. 1997. Tratado de Fitogeografia do Brasil: Aspectos Ecológicos,
Sociológicos e Florísticos 2 º ed. Âmbito Cultural Edições Ltda, Brazil.
Schmitz, O. J.; Hambäck, P. A. & Beckerman, A. P. 2000. Trophic cascade in terrestrial
systems: A review of the effects of carnivore removal plants. The American
Naturalist 155:141-153.
Schmitz, O. J. & Suttle, K. B. 2001. Effects of top predator species on direct and
indirect interactions in food web. Ecology 78: 1388-1399.
Schmitz, O.J. 2006. Predators have large effects on ecosystem properties by changing
plant diversity, not plant biomass. Ecology, 87:1432-1347.
Smith, D. 2000. The population dynamics and community ecology of root hemiparasitic
plants. American Naturalist 155:13-23.
Shen, H.; Ye, W.; Hong, L.; Huang, H.; Wang, Z.; Deng, X.; Yang, Q.; Xu, Z. 2006.
Progress in parasitic plant biology: Host selection and nutrient transfer. Plant
Biology, 8: 175-185.
Strong, D. R. 1992. Are trophic cascades all wet? Differentiation and donor-control in
speciose ecosystems. Ecology, 73: 747-754.
Tennakoon, K. U.; Pate, J. S. 1996. Effects of parasitism by a mistletoe on the structure
and functioning of branches of its host. Plant Cell and Environment, 19: 517-528.
Thompson, R. M, Mouritsen, K. N. & Poulin, R. 2005. Importance of parasites and
their life cycle characteristics in determining the structure of a large marine food
web. Journal Animal Ecology, 74: 77- 85.
Viana, P. L. & Lombardi, J. A. 2007. Florística e caracterização dos campos rupestres
sobre a canga na Serra da Calçada, Minas Gerais, Brasil. Rodriguésia, 58: 159-177.

101
Watson. D. M. 2001. Mistletoe - A keystone resource in forests and woodlands
worldwide. Annual Review of Ecology and Systematics, 32: 219-249.
Werner, E.E & Peacor, S.D. 2003. A review of trait-mediated indirect interactions in
ecological communities. Ecology, 84: 1083-1100.
Zar J. H. 2009. Biostatistical analysis, 5ª ed. Pearson Prentice-Hall, Upper Saddle River,
USA.

102
Anexo 2A – Lista das espécies vegetais - área A (Altas densidades da parasita).
Anemiacea Anemia sp.
Apocynaceae Ditassa mucronata Mart.
Asteraceae Ageratum fastigiatum (Gardn.) R.M. King & H. Rob.
Baccharis serrulata DC.
Baccharis reticulariaDC.
Cromolaena sp.
Lychnophora pinaster Mart.
Symphyopapus brasiliensis (Gardner) R. M. King & H. Rob.
Trixis vauthieri DC.
Bromeliaceae Dyckia cf. simulans L.B. Sm.
Cactaceae Arthrocereus glaziovii (K. Schum.) N.P.Taylor & D.C. Zappi
Celastraceae Maytenus gonoclada Mart.
Cyperaceae Cyperaceae
Euphorbiaceae Croton serratoideus Radcl.-Sm. & Govaerts
Microstachys daphnoides
Fabaceae Mimosa calodendron Mart.
Periandra mediterranea Taub
Lamiaceae Eriope macrostachya Mart. ex Benth.
Lauraceae Ocotea tristis (Nees) Mez
Loranthaceae Struthanthus flexicaulis Mart. Mart.
Tripodanthus acutifoliusThieg.
Melastomataceae Leandra australis (Cham.) Cogn.
Microlicia sp.
Tibouchina sp1
Tibouchina heteromalla
Myrtaceae Myrcia mutabilis (O.Berg) N.J.E. Silveira
Orquidaceae Acianthera teres (Lindl.) Borba
Epidendrum
Orchidaceae
Sophronitis cattleya
Phytolaccaceae Microtea paniculataMoq.
Piperaceae Peperomia decora Dahlst.
Poaceae Poaceae
Portulacaceae Portulaca hirsutissima Cambess.
Rubiaceae Borreria cf. capitata (Ruiz & Pav.) DC.
Velloziaceae Barbacenia
Vellozia albiflora
Vellozia graminea Pohl.
Vellozia compacta Mart.
Verbenaceae Lippia sp.
Stachytarpheta glabra Cham.

103
Anexo 2B – Lista das espécies vegetais - área B (Baixas densidades da parasita).
Alstroemeriaceae Alstroemeria plantaginea Mart.
Apocynaceae Ditassa mucronata Mart.
Asteraceae Ageratum fastigiatum (Gardn.) R.M. King & H. Rob.
Baccharis serrulata DC.
Baccharis reticularia DC.
Cromolaena sp.
Lessingianthus sp.
Lychnophora pinaster Mart.
Symphyopapus brasiliensis (Gardner) R. M. King & H. Rob.
Trixis vauthieri DC.
Bromeliaceae Dyckia cf. simulans L.B. Sm.
Vriesea minarum L.B. Sm.
Cactaceae Arthrocereus glaziovii (K. Schum.) N.P.Taylor & D.C. Zappi
Cyperaceae Cyperaceae
Euphorbiaceae Croton serratoideus Radcl.-Sm. & Govaerts
Microstachys daphnoides
Fabaceae Mimosa calodendron Mart.
Periandra mediterranea Taub
Lamiaceae Eriope macrostachya Mart. ex Benth.
Loranthaceae Struthanthus flexicaulis Mart. Mart.
Tripodanthus acutifoliusThieg.
Malpighiaceae Heteropteres sp.
Melastomataceae Leandra australis (Cham.) Cogn.
Microlicia sp.
Tibouchina heteromalla
Orquidaceae Acianthera teres (Lindl.) Borba
Epidendrum
Oncidium blanquet
Orchidaceae
Orquidea geófita
Sophronitis cattleya
Phytolaccaceae Microtea paniculataMoq.
Poaceae Poaceae
Portulacaceae Portulaca hirsutissima Cambess.
Rubiaceae Borreria cf. capitata (Ruiz & Pav.) DC.
Velloziaceae Vellozia albiflora
Vellozia graminea Pohl.
Vellozia compacta Mart.
Verbenaceae Lippia
Stachytarpheta glabra Cham.

104
Considerações finais
Durante a realização deste estudo surgiram novos questionamentos sobre o papel
de S. flexicaulis no funcionamento e estruturação da comunidade vegetal dos
afloramentos ferruginosos. Uma das perguntas a ser respondida é se a composição
vegetal e a riqueza de espécies se alterariam com o tempo de atuação da parasita. Como
este trabalho foi realizado dentro de uma unidade de conservação e as áreas foram
georeferenciadas, futuramente seria possível fazer um novo levantamento de espécies
vegetais. Outra solução seria buscar respostas mais imediatas através da incorporação de
novos parâmetros ao modelo matemático que, para esta tese, foram desconsiderados nas
simulações. A adição de novos parâmetros como densidade de hospedeiras (não
somente M. calodendron), configuração da paisagem considerando espécies herbáceas,
demografia das hospedeiras e o tempo que as espécies vegetais associadas ao
parasitismo levam para morrer, deverá dar respostas mais precisas sobre o destino desta
comunidade vegetal.
Futuros estudos deverão investigar como e quando a parasita forrageia. Através
de um experimento mais detalhado envolvendo as áreas de fisiologia e anatomia
vegetal, química e ecologia seria possível determinar os mecanismos de forrageamento
adotados pela parasita para otimizar as estratégias de busca por alimentos e
reconhecimento de hospedeiras. Como complemento a esta pergunta, recomenda-se
acompanhar a fenologia da parasita em diferentes hospedeiras. Estas respostas
ajudariam a compreender os diferentes efeitos do parasitismo por S. flexicaulis sobre
cada espécie hospedeira, indicando seu o sucesso e permanência em diferentes
comunidades vegetais.
Durante o desenvolvimento deste trabalho, alguns funcionários do Parque
Estadual da Serra do Rola Moça questionaram a necessidade do manejo de S. flexicaulis
na comunidade vegetal, uma vez que ela provoca a morte de indivíduos. Contudo não se
recomenda a retirada de S. flexicaulis da comunidade vegetal, uma vez que esta é uma
espécie-chave, nativa e não causou nenhuma alteração na composição e na diversidade
de espécies.
A
Related Documents











