Ann. appl. Biol. (1 996), 129:479489 Printed in Great Britain 479 Differentiation of citrus tristeza closterovirus (CTV) isolates by single-strand conformation polymorphism analysis of the coat protein gene By LUIS RUBIOl, Ma ANGELES AYLLONl, JOSE GUERRI', H A " U PAPPU', CHUCK NIBLElT2 and PEDRO MORENO1* 'Institute Valenciano de Investigaciones Agrarias, Apdo. Oficial, 46113- Moncada, Valencia, Spain 2Plant Pathology Department, University of Florida, Gainesville, FL 32611-0680, USA (Accepted 15 November 1996) Summary Citrus tristeza closterovirus (CTV) isolates of several geographical origins were compared for variations in their coat protein (CP) gene by analysis of single-strand conformation polymorphism (SSCP). The CP gene of 17 isolates was reverse transcribed, amplified by polymerase chain reaction (PCR), and 22 clones were inserted into a plasmid vector. These clones were sequenced and found to have between 91.7% and 99.8% sequence homology. Clones were amplified and the PCR products denatured and compared by SSCP analysis in 8% polyacrylamide gels. Using two different electrophoretic conditions, the patterns were different for 16 or 17 clones. Four pairs of clones (T36/T66, P1/Q2, 03/8Q, and E l E 2 ) differing by 10, 2, 1 and 1 nucleotides, respectively, could not be distinguished using either condition. When these clones were compared by SSCP after digestion with Eco91I (BstEII) three of the pairs (T36/T66, Pl/Q2, and 03/8Q) could be differentiated, whereas the clones E l and E2 (differing by 1 nucleotide) remained indistinguishable. Thus, SSCP analysis combining two electrophoretic conditions and restriction of eight clones with Eco91I allowed discrimination between 21 of the 22 CP gene clones selected. SSCP analysis may provide a procedure to identify and differentiate CTV isolates based on comparisons of several genes or gene regions. It is rapid and cheap and may drastically reduce the amount of sequencing necessary for accurate comparisons. Key words: SSCP, polymerase chain reaction, electrophoresis, nucleotide sequencing Introduction Citrus tristeza virus (CTV), the causal agent of one of the most economically important diseases of citrus, is a closterovirus with a single-stranded, positive-sense RNA genome of 19296 nucleotides (Karasev et al., 1995), encapsidated in flexuous particles about 2000 nm *To whom correspondence should be addressed 0 1996 Association of Applied Biologists

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ann. appl. Biol. (1 996), 129:479489 Printed in Great Britain 479
Differentiation of citrus tristeza closterovirus (CTV) isolates by single-strand conformation polymorphism
analysis of the coat protein gene
By LUIS RUBIOl, Ma ANGELES AYLLONl, JOSE GUERRI', H A " U PAPPU', CHUCK NIBLElT2 and PEDRO MORENO1*
'Institute Valenciano de Investigaciones Agrarias, Apdo. Oficial, 461 13- Moncada, Valencia, Spain
2Plant Pathology Department, University of Florida, Gainesville, FL 32611-0680, USA
(Accepted 15 November 1996)
Summary Citrus tristeza closterovirus (CTV) isolates of several geographical origins were
compared for variations in their coat protein (CP) gene by analysis of single-strand conformation polymorphism (SSCP). The CP gene of 17 isolates was reverse transcribed, amplified by polymerase chain reaction (PCR), and 22 clones were inserted into a plasmid vector. These clones were sequenced and found to have between 91.7% and 99.8% sequence homology. Clones were amplified and the PCR products denatured and compared by SSCP analysis in 8% polyacrylamide gels. Using two different electrophoretic conditions, the patterns were different for 16 or 17 clones. Four pairs of clones (T36/T66, P1/Q2, 03/8Q, and E l E 2 ) differing by 10, 2, 1 and 1 nucleotides, respectively, could not be distinguished using either condition. When these clones were compared by SSCP after digestion with Eco91I (BstEII) three of the pairs (T36/T66, Pl/Q2, and 03/8Q) could be differentiated, whereas the clones E l and E2 (differing by 1 nucleotide) remained indistinguishable. Thus, SSCP analysis combining two electrophoretic conditions and restriction of eight clones with Eco91I allowed discrimination between 21 of the 22 CP gene clones selected.
SSCP analysis may provide a procedure to identify and differentiate CTV isolates based on comparisons of several genes or gene regions. It is rapid and cheap and may drastically reduce the amount of sequencing necessary for accurate comparisons.
Key words: SSCP, polymerase chain reaction, electrophoresis, nucleotide sequencing
Introduction Citrus tristeza virus (CTV), the causal agent of one of the most economically important
diseases of citrus, is a closterovirus with a single-stranded, positive-sense RNA genome of 19296 nucleotides (Karasev et al., 1995), encapsidated in flexuous particles about 2000 nm
*To whom correspondence should be addressed 0 1996 Association of Applied Biologists

480 LUIS RUB10 ET AL.
long and 10-12 nm diameter (Bar-Joseph & Lee, 1989). A variety of CTV strains, differing by symptoms induced on different host species, aphid transmissibility or capacity to interfere with other strains, have been described (Raccah, Loebenstein & Singer, 1980; Ballester- Olmos et al., 1993; Van Vuuren, Collins & da GraGa, 1993). Economic losses caused by severe CTV strains can be avoided by early detection and eradication of these strains or by cross-protection using mild strains (Costa & Muller, 1980). Efficient application of these procedures requires reliable methods for strain identification and differentiation.
Several approaches have been tested for these purposes. Indexing on different indicator species is currently used for biological characterisation of CTV isolates, but it is time consuming, expensive and frequently imprecise for strain identification (Ballester-Olmos et al., 1993). A limitation of using monoclonal antibodies (Permar, Garnsey, Gumpf & Lee, 1990; Cambra et al., 1993), or peptide mapping (Guerri, Moreno & Lee, 1990), is that changes are only detected in the coat protein (CP) gene, which occupies just 3.5% of the genome. Additionally, peptide map analysis is too cumbersome and expensive for routine utilisation. Double-stranded RNA (dsRNA) analysis of infected plants (Dodds, Jarupat, Lee & Roistacher, 1987) is easy to perform and can detect variations in different portions of the genome, but dsRNA patterns and pathogenic characteristics are not necessarily correlated (Moreno, Guerri & Muiioz, 1990; Moreno et al., 1993). Hybridisation of the viral RNA with complementary DNA (cDNA) probes can potentially detect changes all along the genome (Rosner. Lee & Bar-Joseph, 1986; Albiach, 1995), but a wide panel of cDNA probes may be necessary for this approach.
Cillings, Broadbent, Indsto 8i Lee (1993) have used the polymerase chain reaction (PCR) (Saiki et al., 1988) to detect CTV in field trees, and restriction fragment length polymorphisms (RFLP) to compare isolates. The disadvantages of RFLP are that only nucleotide changes affecting restriction sites are detected and that PCR products may need to be tested with many different restriction enzymes.
Cloning and sequencing the CP gene has been used for comparisons of CTV isolates with different geographic origin and/or biological properties (Pappu et al., 1993~). Nucleotide
Table 1. Citrus tristeza virus isolates and clones Isolate'"' Clone Origin
T30 T30 Florida, USA T36 T36 Florida, USA T66 T66 Florida, USA T308 E l , E2 Spain T317 N2. N3 Spain T317L 03, 05 Spain T318 P1, P2 Spain T305 Q2 Spain T388 s3, s.5 Spain (originally from Japan) B53 B.53 USA (originally from Japan) IVIA QD 8Q 8Q Spain B7 B7 USA (originally from South Africa) B128 B128 USA (originally from Columbia) B272 B272 USA (originally from Columbia) B274 8214 USA (originally from Columbia) Cu 17b Cu 17b USA (originally from Cuba) B227 8227 USA (originally from India)
“%elates labelled with B and Cu 17b are from the international collection of exotic citrus pathogens maintained at the quarantine facilities of USDA-BARC, Beltsville, MD, USA.

Differentiation of CTV isolates by SSCP analysis 48 1
sequencing is the most accurate procedure for strain identification, but it is expensive and time consuming for routine purposes.
Single-strand conformation polymorphism (SSCP) analysis (Orita et al., 1989a,b) enables rapid differentiation of DNA fragments of the same size but with sequence variations. In SSCP analysis, double-stranded DNA is heat denatured in the presence of formamide and both DNA strands are separated by electrophoresis in a non-denaturing polyacrylamide gel. Migration of single-strand DNA (ssDNA) depends on its conformation under the electrophoretic conditions used, and this conformation depends on its nucleotide sequence. Small changes in the nucleotide sequence may alter conformation of ssDNA and, consequently, its electrophoretic mobility. This property can be exploited to detect mutations in selected DNA fragments by denaturing them and comparing their electrophoretic pattern. SSCP analysis has been used to evaluate sequence heterogeneity among isolates of hepatitis B (Yap et al., 1992) and C (Enomoto et al., 1994) viruses, and more recently to assign beet necrotic yellow vein virus isolates to a known strain group (Koenig, Liiddecke & Haeberle, 1995).
In this paper we describe the differentiation of CTV isolates by SSCP analysis of the DNA obtained after reverse transcription and PCR (RT-PCR) of the CP gene.
Materials and Methods
Virus isolates Twenty two clones of the CP gene obtained from 17 CTV isolates were used in this study.
The origin of the different clones is shown in Table 1.
Cloning and sequencing RNA was extracted from desiccated infected tissue by phenol-detergent treatment and
purified by column chromatography (Pappu et al., 1993b) using Sephadex G-50 (Pharmacia Biotech, Uppsala, Sweden). DsRNA was also purified from CTV-infected tissue following a protocol previously described (Moreno et al., 1990). Using purified total RNA or CTV dsRNA, cDNA of the CP gene was obtained by RT-PCR using the oligonucleotide primers CN150 (S'ATATATTTACTCTAGATCTACCA TGGACGACGAAACAAA) and CN 15 1 (5'GAATCGGAACGCGAA~CTCAAGTGTGTTAAA~CC?. These primers contain 17 and 20 bases of both ends of the CP gene (italics) (Pappu et al., 1994), and aXbaI and a EcoRI restriction site (underlined), respectively. RT-PCR was performed in 100 p1 reaction mixture containing 200 ng of each primer, 10 p1 PCR buffer 10 x (100 mM Tris-HC1, pH 9.0, 500 mM KCl, 15 mM MgC12, 10 ml litre-' Triton X-loo), 10 mM D?T, 400 p~ of each of the four nucleotides, 10 units RNasin, 15 units Ah4V reverse transcriptase, and 2.5 units Taq polymerase (Promega Corp., MD, USA). RNA was first denatured for 5 rnin at 95°C and chilled on ice. Then the following thermocycling conditions were used: 45 rnin at 42°C (1 cycle), 2 rnin at 94"C, 2 rnin at 42"C, and 2 min at 72°C (35 cycles), and 10 rnin at 72°C (1 cycle). The DNA ends were made cohesive by digestion with XbaI and EcoRI and then cloned in pUCl18 cut at these two sites. After ligation, DH a5 Escherichia coli cells were transformed with the plasmid, and colonies containing plasmid with insert were selected with X-gal (Sambrook, Fritsch & Maniatis, 1989).
The clones were sequenced as dsDNA using Sequenase Version 2.0 (US Biochemicals), according to the manufacturer's instructions.

482 LUIS RUBIO ET AL.
DNA amplification from clones Plasinids containing the CP gene clones were purified by the miniprep procedure
(Sambrook et ul., 1989) and 1-2 it1 aliquots were used for amplification with the primers CN150 and CN15 1. PCR was performed in 100 pl total volume containing 10 p1 PCR buffer lox, dNTPs at a final concentration of 200 p ~ , 100 ng of each primer, 2.5 units of Taq DNA polymerase (Promega Corporation) and 81.5 111 sterile double distilled H20. DNA amplification was for 35 cycles of denaturation for 2 min at 94"C, annealing for 2 min at 55 C. and DNA synthesis for 2 niin at 72 C, followed by a single chain extension period of 10 min at 72 C.
Occasionally PCR products were re-amplified in the above conditions. In these cases, template DNA was first purified using Wizard DNA Clean-up Purification Resin (Promega Corporation). following the manufacturer's instructions and eluting the DNA with 50 p1 sterile distilled water at 80 C. The concentration of DNA was estimated using an ethidum bromide stained agarose gel (8 g litre-'). An aliquot containing -10 ng of DNA was then used &:\ PCR template.
SSCP analy5ir SSCI' analysis was performed directly on the PCR products of the CP gene of different
CTV iwlates. or following restriction digests with EcoR91I (BsrEII) (MBI Fermentas, I *ithumla).
U~ually. 1 p1 of the PCR product was mixed with 9 1.11 of the denaturing solution (950 ml iitre formamide, 20 mM EDTA. 500 mg litre-' xylene-cyanol, and 500 mg litre-' bromophenol blue), heated for 10 min at 99 C, and chilled on ice. Denatured DNA was electrophoresed in a non-denaturing 80 g litre-' polyacrylamide minigel (80 mm x 70 mm x 0.75 mm) with or without 50 ml litre ' glycerol, using TBE (Tris-borate 89 mM, 2 mM EDTA, pFI 8 0) as electrophoresis buffer, and a constant voltage of 200 or 300 volts for 5 or 3 h, at 3 C or at rtoom temperature. The gels were stained with silver nitrate using the procedure of Beidler. Hilllard & Rill (1982).
1
Results
Nucleoticie differences between clones The 22 CP gene selected clones (Table 1) were sequenced, their nucleotide sequences
aligned using the program Clustal V (Higgins & Sharp, 1989), and the nucleotide differences obtained with the program ANDIST3 (kindly provided by Dr Dopazo, Departamento de GenCtica. Facultad de Biologia, Universidad de Valencia, Spain). Table 2 summarises these differences which were from 1 to 59 nucleotides (91.7% to over 99.8% sequence homology). The clones sharing over 98% homology were grouped in three clusters. One of them included clones El, E2. T36 and T66. the second included N2, N3,03 ,05 , SQ, T30, B272 and B274, and the third, P1, P2, Q2, S3 and S5.
Amplification of the clones and SSCP analysis Amplification from the plasmids containing the 22 clones yielded in all cases a single DNA
iragment of 710 bp. encompassing the 669 bp of the CP gene (Sekiya, Lawrence, McCaffery & Cline, 1991, Pappu et al.. 1993a) and the extra nucleotides of both primers.
In preliminary experiments, several electrophoretic conditions (voltage, acrylamide concentration. time, presence or absence of glycerol) were assayed. It was established that electrophoresis at a constant voltage of 200-300 volts, for 3-5 h, in 80 g litre acrylamide
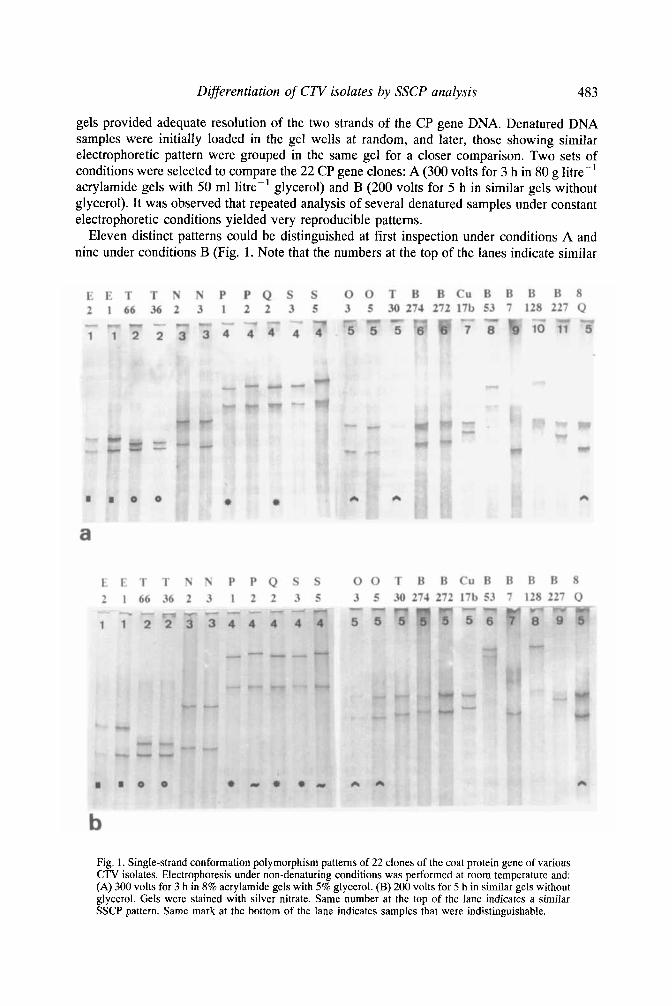
Differentiation of CTV isolates by SSCP analysis 483
gels provided adequate resolution of the two strands of the CP gene DNA. Denatured DNA samples were initially loaded in the gel wells at random, and later, those showing similar electrophoretic pattern were grouped in the same gel for a closer comparison. Two sets of conditions were selected to compare the 22 CP gene clones: A (300 volts for 3 h in 80 g litre-' acrylamide gels with 50 ml litre-' glycerol) and B (200 volts for 5 h in similar gels without glycerol). It was observed that repeated analysis of several denatured samples under constant electrophoretic conditions yielded very reproducible patterns.
Eleven distinct patterns could be distinguished at first inspection under conditions A and nine under conditions B (Fig. 1. Note that the numbers at the top of the lanes indicate similar
Fig. 1. Single-strand conformation polymorphism patterns of 22 clones of the coat protein gene of various CTV isolates. Electrophoresis under non-denaturing conditions was performed at room temperature and: (A) 300 volts for 3 h in 8% acrylamide gels with 5 % glycerol. (B) 200 volts for 5 h in similar gels without glycerol. Gels were stained with silver nitrate. Same number at the top of the lane indicates a similar SSCP pattern. Same mark at the bottom of the lane indicates samples that were indistinguishable.

484 LUIS RUB10 ET AL.
patterns). Some of them corresponded to individual clones (for instance, Cul7b, B53, B7, B128 and B227 in Fig. 1) whereas others included two or more clones (Fig. 1). Clones with a similar SSCP pattern always belonged to the same cluster in Table 2 and therefore shared over 98% sequence homology; however, some clones within the same cluster in Table 2 showed a distinct SSCP pattern (for example, the patterns of E l and E2 differed from those of T36 or T66; and the patterns of N2 and N3 also were different from those of 0 3 , 0 5 and T30, which are in the same cluster in Table 2).
After closer comparison, minor differences in electrophoretic mobility were also observed between some of the clones initially grouped within the same SSCP pattern. These clones were electrophoresed in the same gel in at least three different analyses, using the electrophoretic conditions (A or B) allowing better differentiation. When these differences were consistent in all gels they were considered to be sufficiently discriminating to identify different isolates. When these additional analyses were taken into account, 17 clones could be distinguished under conditions (A), whereas conditions (B) enabled discrimination of 16 (Fig. 1 . Samples marked with the same sign at the bottom of the lane were considered to be mdistinguishable).
Most of the clones differentiated under conditions (A) were also differentiated under conditions (B). Nevertheless, under conditions (A), clone T30 showed the same electrophoretic mobility as clones 03 and SQ, from which it differs in two and three nucleotides, respectively, whereas it could be discriminated under conditions (B). Conversely, under conditions (B) the clone S3 showed the same electrophoretic mobility as P1 and Q2 (it differs in 1 and 2 nucleotides, respectively), and it was difficult to distinguish S5 and P2 (two nucleotides difference). Both S3 and S5 could be discriminated from the other clones under conditions (A) (Fig. 1). Thus. using both sets of conditions up to 18 of the 22 clones analysed could be differential. The pairs T36F66 (10 nucleotides difference), P1/Q2 (2 nucleotides
Table 7. Summarv of nucleotide differences among 22 clones of the coat protein gene of various CTV isolates*
13128 31 13277 35 51 HS3 46 47 43 Cul7b 42 48 44 16 El 48 40 51 34 34 - - - - - E2 48 41 50 33 31 1 1 I
T36 50 45 54 37 36 I 2 4 I Th6 52 47 58 41 413 9 l a J@ Y2 53 57 49 52 5 1 44 43 47 5 1 - - - - - - - - - - - - ~
Y3 53 57 49 52 52. 46 45 49 53 2 I 0 3 49 53 44 48 4;3 42 41 45 49 1 5 5 I 0 5 50 54 45 49 4Y 43 42 46 50 I 6 6 1 HQ 50 54 45 49 49 43 42 46 50 I 6 6 1 2 r30 51 55 46 50 49 44 43 47 51 7 7 2 3 3 3272 55 59 49 54 54 48 47 51 55 113 13 8 9 9 10 I
13274 52 56 49 51 51 45 44 48 52 , 2- - -6 -6- -7- -7-1 PI 31 30 41 46 48 44 43 47 51 51 51 47 48 48 49 53 48 - - - - - - I
I
E i 32 31 42 47 49 45 44 48 52 52 52 48 49 49 50 54 49 I 1 2 3 I S5 37 31 42 47 4Y 45 44 48 52 52 52 48 49 49 50 54 49 I 1 2 3 2 1
R7 8128B227B53 Cul7b E l E2 T36T66 N2 N3 0 3 0 5 8Q T30B272B274 P1 P2 Q2 S3
I I I I
P2 32 31 42 47 49 45 44 48 52 52 52 48 49 49 SO 54 49 I 1 0 2 31 32 43 46 50 46 45 49 53 53 53 49 50 50 51 55 SO I 2 3 I
L - - - - - - J
I * ) The PCK product from each clone had 710 nucleotides.
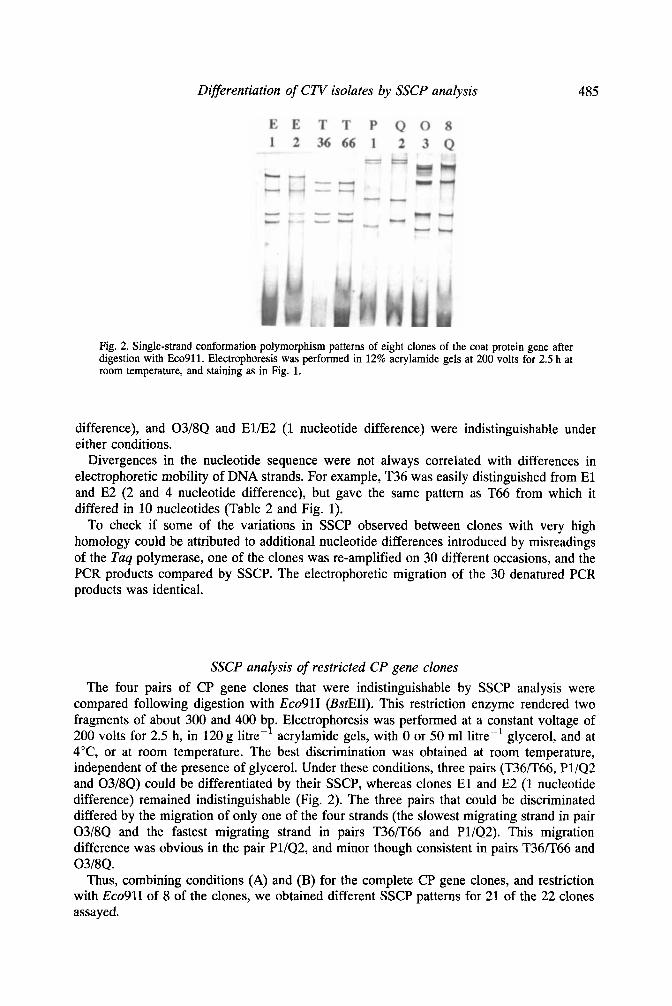
Differentiation of CTV isolates by SSCP analysis 485
Fig. 2. Single-strand conformation polymorphism patterns of eight clones of the coat protein gene after digestion with Eco911. Electrophoresis was performed in 12% acrylamide gels at 200 volts for 2.5 h at room temperature, and staining as in Fig. 1.
difference), and 03/8Q and EM32 (1 nucleotide difference) were indistinguishable under either conditions.
Divergences in the nucleotide sequence were not always correlated with differences in electrophoretic mobility of DNA strands. For example, T36 was easily distinguished from El and E2 (2 and 4 nucleotide difference), but gave the same pattern as T66 from which it differed in 10 nucleotides (Table 2 and Fig. 1).
To check if some of the variations in SSCP observed between clones with very high homology could be attributed to additional nucleotide differences introduced by misreadings of the Taq polymerase, one of the clones was re-amplified on 30 different occasions, and the PCR products compared by SSCP. The electrophoretic migration of the 30 denatured PCR products was identical.
SSCP analysis of restricted CP gene clones The four pairs of CP gene clones that were indistinguishable by SSCP analysis were
compared following digestion with Eco91I (BstEII). This restriction enzyme rendered two fragments of about 300 and 400 b . Electrophoresis was performed at a constant voltage of 200 volts for 2.5 h, in 120 g litre-' acrylamide gels, with 0 or 50 ml litre-' glycerol, and at 4"C, or at room temperature. The best discrimination was obtained at room temperature, independent of the presence of glycerol. Under these conditions, three pairs (T36n66, P1/Q2 and 03/8Q) could be differentiated by their SSCP, whereas clones E l and E2 (1 nucleotide difference) remained indistinguishable (Fig. 2). The three pairs that could be discriminated differed by the migration of only one of the four strands (the slowest migrating strand in pair 03/8Q and the fastest migrating strand in pairs T36F66 and P1/Q2). This migration difference was obvious in the pair Pl/Q2, and minor though consistent in pairs T36R66 and 03/8Q.
Thus, combining conditions (A) and (B) for the complete CP gene clones, and restriction with Eco91I of 8 of the clones, we obtained different SSCP patterns for 21 of the 22 clones assayed.

486 LUIS RUB10 E T M .
Discussion CTV isolates with 1 to 59 nucleotide differences in their CP gene could be distinguished by
SSCP analysis. Furthermore, some CP gene clones from the same CTV isolate, showing only 1 or 2 nucleotide variations in their sequence, could also be discriminated by this procedure. The sensitivity of SSCP analysis and the possibility of applying it to different virus genes makes it a very powerful procedure to discriminate CTV isolates.
Sometimes, the mobility differences between bands of two clones were small, but usually, repeated SSCP analysis of these clones in adjacent gel wells showed that the differences observed were consistent.
Since the DNA strand conformation can be altered by several factors (i.e. electrophoresis buffer, voltage, temperature, or the presence of glycerol or other additives) they also will alter the electrophoretic mobility. Different authors have recommended high acrylamide concentration (Savov et al. , 1992), low temperature (Hongyo, Buzard, Calvert & Weghorst, 1993), or glycerol addition for improved resolution of the DNA strands. Our experience with several DNA fragments suggests that, as indicated by other authors (Leren et al., 1993), there is no general rule, but rather conditions must be adjusted in each case.
Nucleotide incorporation errors in the initial cycles of the PCR could give rise to a significant population of molecules with nucleotide changes (mutations), and the resulting electrophoretic profile could be affected by these changes. However, we have found that the 30 PCR products prepared from a single clone showed identical electrophoretic profiles, indicating that under our experimental conditions this potential source of error was negligible. A5 indicated by Krawczak, Reiss, Schmidtke & Rosler (1989) with an initial population of more than lo4 DNA molecules it is very unlikely that nucleotide incorporation errors could ever give rise to a detectable sub-population of false mutants.
SSCP analysis has been used to detect single base mutations in DNA fragments from 135 to 400 bp (Orita et al.. 1 9 8 9 ~ ; Hayashi, 1991; Sheffield et al., 1993; Prosser, 1993) using long gels and electrophoresis runs of more than 8 h. Under the conditions used in this work we were able to detect single base mutations in DNA fragments of 710 bp using minigels and electrophoresing denatured samples for 3-5 h. Silver staining was also a quick and safe procedure providing sufficient sensitivity to detect the DNA strands without using hybridisation with radiolabelled probes.
Some electrophoretic profiles (i.e. B7 in Fig. 1) showed three instead of two bands. This might be due to the presence of two stable conformations for one of the strands (Hayashi, 1991) or. to an excess of primers that might bind to the DNA strands and form a detectable subspecies (Cai & Touitou, 1993). These types of artifact can be avoided by removing the oligonucleotides from the PCR product (Fujita & Silver, 1994) or just by diluting it. Nevertheless, when the PCR product appears as a single band in an agarose gel, this precaution is unnecessary (Hongyo el ai., 1993).
The genes or gene regions responsible for the pathogenic capabilities of CTV are unknown. The possibility of detecting single mutations in relatively large DNA fragments provides a new approach to compare the genome of CTV isolates and reduces the need for sequencing. Clones covering most of the genome could be obtained and only those clones differing in their SSCP patterns would need to be sequenced. A limitation of this approach is that obvious differences in the SSCP pattern do not necessarily mean great differences in the nucleotide sequence, and vice versa. For example, clones T36 and T66, that differ by 10 nucleotides could not be distinguished by their electrophoretic profile under the conditions used, whereas other pairs differing in one or two nucleotides were easily discriminated, thus confirming previous observations that the effect of base changes on the electrophoretic pattern is basically unpredictable (Orita et al., 1989~) . This problem can be partially avoided by using several

Differentiation of CTV isolates by SSCP analysis 487
conditions for SSCP of the selected DNA pieces and also testing restriction fragments obtained with one or more endonucleases.
Field isolates of CTV are presently being compared by SSCP analysis of RT-PCR products using primers for different genes. The procedure is also being applied to the study of cross protection between CTV strains.
Acknowledgements We are indebted to M E Martinez, M Boil, E Estela, and L Galipienso for technical
assistance. The first and second authors were recipients of fellowships of the Conselleria de Educaci6n y Ciencia de la Comunidad Valenciana and the CICYT, respectively. Dr. Niblett’s visit to the IVIA was supported by the Fulbright Commission and the Conselleria de Educaci6n y Ciencia de la Comunidad Valenciana. This work was supported by INIA project SC93-111.
References Albiach M R 1995. Diferenciacibn bioquimica de razas del virus de la tristeza de 10s citricos (CW) y
aplicacibn a la proteccibn cruzada. Tesis Doctoral. Universidad de Valencia, Facultad de Ciencias Quiniicas. 148 pp.
Ballester-Olmos J F, Pina J A, Carbonell E, Moreno P, Hermoso de Mendoza A, Cambra M, Navarro L. 1993. Biological diversity of citrus tristeza virus (CTV) isolates in Spain. Plant Pathology 42: 2 19-229.
Bar-Joseph M, Lee R F. 1989. Citrus tristeza virus. AAB Descriptions of Plant Viruses No. 353. 7 pp. Beidler L L, Hilliard P R, Rill R L. 1982. Ultrasensitive staining of nucleic acids with silver.
Analytical Biochemistry 126:374380. Cai Q Q, Touitou I. 1993. Excess PCR primers may dramatically affect SSCP efficiency. Nucleic Acid
Research 21:3909-3910. Cambra M, Camarasa E, Gorris M T, Garnsey S M, Gumpf D J, Tsai M C. 1993. Epitope diversity
of citrus tristeza virus (CTV) isolates in Spain. In Proceedings of the 12th Conference of the International Organization of Citrus Virologists, pp. 33-38. Eds P Moreno, J V da GraGa and LW Timmer. Riverside, CA, USA: IOCV.
Costa A S, Miiller G W. 1980. Tristeza control by cross protection: a US.-Brazil cooperative success. Plant Disease 64538-541.
Dodds J A, Jarupat T, Lee J G, Roistacher C N. 1987. Effects of strain, host, time of harvest, and virus concentration on double-stranded RNA analysis of citrus tristeza virus. Phytopathology 77:442- 447.
Enomoto N, Kurosaki M, Tanaka Y, Marumo F, Sat0 C. 1994. Fluctuation of hepatitis C virus quasispecies in persistent infection and interferon treatment revealed by single-strand conformation polymorphism analysis. Journal of General Virology 751361-1369.
Fujita K, Silver J. 1994. Single-strand conformational polymorphism. PCR Methods and Applications
Gillings M, Broadbent P, Indsto J, Lee R F. 1993. Characterisation of isolates and strains of citrus tristeza closterovirus using restriction analysis of the coat protein gene amplified by the polymerase chain reaction. Journal of Virological Methods 44:305-317.
Guerri J, Moreno P, Lee R F. 1990. Characterization of strains of citrus tristeza virus by peptide maps of virion coat protein. Phytopathology 80:692-698.
Hayashi K. 1991. PCR-SSCP: A simple and sensitive method for detection of mutations in the genomic DNA. PCR Methods and Applications 1:34-38.
4:137-140.

488 LUIS RUB10 ET AL.
Higgins P G, Sharp P M. 1989. Fast and sensitive multiple sequence alignment on a microcomputer.
Hongyo T, Buzard G S, Calvert R J, Weghorst C M. 1993. “Cold SSCP”: a simple, rapid and non- radioactive method for optimized single-strand conformation polymorphism analyses. Nucleic Acid Research 21:3637-3642.
Karasev A V, Boyko V P, Gowda S, Nikolaeva 0 V, Hilf M E, Koonin E V, Niblett C L, Cline K, Gumpf D J, Lee R F, Garnsey S M, Dawson W 0. 1995. Complete sequence of the citrus tristeza virus RNA genome. Virology 20851 1-520.
Koenig R, Luddecke P, Haeberle A M. 1995. Detection of beet necrotic yellow vein virus strains, variants and mixed infections by examining single-strand conformation polymorphisms of immunocapture RT-PCR products. Journal of General Virology 76:205 1-2055.
Krawczak M, Reiss J, Schmidtke J, Rosier U. 1989. Polymerase chain reaction: Replication errors and reliability of gene diagnosis. Nucleic Acid Research 17:2197-2201.
Leren T P, Solberg K, Rodningen 0 K, Ose L, Tonstad S, Berg K. 1993. Evaluation of running conditions for SSCP analysis: Application of SSCP for detection of point mutations in the LDL receptor gene. PCR Methods and Applications 3: 159-162.
Morenn P, Guerri J, Munoz N. 1990. Identification of Spanish strains of citrus tristeza virus (CTV) by analysis of double-stranded RNAs (dsRNA). Phytopathology 80:477482.
Moreno P, Guerri J, Ballester-Olmos J F, Albiach R, Martinez M E. 1993. Separation and interference of strains from a citrus tristeza virus isolate evidenced by biological activity and double- stranded RNA (dsRNA) analysis. Plant Pathology 42:3541.
Orita M, Suzuki Y, Sekiya T, Hayashi K. 1 9 8 9 ~ . Rapid and sensitive detection of point mutations and DNA polymorphisms using the polymerase chain reaction. Genomics 5:874-879.
Orita M, Iwahana H, Kanazawa H, Hayashi K, Sekiya T. 19896. Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polyrnorphisms. Proccedings of the Natiunal Academy of Science of the USA 86:2766-2770.
Pappu H R, Pappu S, Niblett C L, Lee R F, Civerolo E L. 1 9 9 3 ~ . Comparative sequence analysis of the coat protein of biologically distinct citrus tristeza closterovirus isolates. Virus Genes 7:255-264.
Pappu S S, Brand R, Pappu H R, Rybicki E P, Gough K H, Frenkel M J, Niblett C L. 19936. A polymerase chain reaction method adapted for selective amplification and cloning of 3‘ sequences of potyviral genomes: application to dasheen mosaic virus. Journal of Virological Methods 41:9-20.
Pappu H R, Karasev A V, Anderson E J, Pappu S S, Hilf M E, Febres V J, Eckloff R M G, McCaffery M, Boyko V, Gowda S, Dolja V V, Koonin E V, Gumpf D J, Cline K C, Garnsey S M, Dawson W 0, Lee R F. Niblett C L. 1994. Nucleotide sequence and organization of eight 3’ open reading frames of the citrus tristeza closterovirus genome. Virology 199:35-46.
Permar. T A, Garnsey S M, Gumpf D J, Lee R F. 1990. A monoclonal antibody which discriminates strains of citrus tristeza virus. Phytopatlzology 80:224-228.
Prosser J. 1993. Detecting single-base mutations. TIBTECH 11:238-245 Raccah B, Loebenstein G, Singer S. 1980. Aphid-transmissibility of citrus tristeza virus in infected
citrus trees. Phytopathology 70:89-93. Rosner A, Lee R F, Bar-Joseph M. 1986. Differential hybridization with cloned cDNA sequences for
detecting a specific isolate of citrus tristeza virus. Phytopathology 76:820-824. Saiki K K, Gelfand D H, Stoffel S, Scharf S J, Higuchi R, Horn G T, Mullis K B, Erlich H A. 1988.
Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239:48749 1.
Sambrook J, Fritsch E F, Maniatis T. 1989. Molecular Cloning. A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory.
Savov A, Angelicheva D, Jordanova A, Eigel A, Kalaydjieva L. 1992. High percentage acrylamide gels improve resolution in SSCP analysis. Nucleic Acid Research 20:67414742.
Sekiya M E, Lawrence S D, McCaffery M, Cline K. 1991. Molecular cloning and nucleotide
CABIOS 5:151-153.
- sequencing of the coat protein gene of citrus tristeza virus. Journal of General Virology 72:1013- 1020.

Differentiation of CTV isolates by SSCP analysis 489
Shemeld V C, Beck J S, Kwitek A E, Sandstrom D W, Stone E M. 1993. The sensitivity of single- strand conformation polymorphism analysis for the detection of single base substitutions. Genomics
Van Vuuren S P, Collins R P, da Graga J V. 1993. Evaluation of citrus iristeza virus isolates for cross
Yap E P H, Lo Y-M D, Cooper K, Fleming K A, McGee J 0 D. 1992. Exclusion of false-positive
(Received 5 August 1996)
16:325-332.
protection of grapefruit in South Africa. Plant Disease 77:24-28.
PCR viral diagnosis by single-strand conformation polymorphism. The Lancet 340:736.
Related Documents











