ISSN 1672-9145 Acta Biochimica et Biophysica Sinica 2006, 38(9): 653–662 CN 31-1940/Q ©Institute of Biochemistry and Cell Biology, SIBS, CAS Differentially Expression of Tua1, a Tubulin-encoding Gene, during Flowering of Tea Plant Camellia sinensis (L.) O. Kuntze Using cDNA Amplified Fragment Length Polymorphism Technique Wan-Ping FANG 1,2 , Chang-Jun JIANG 1 *, Mei YU 1 , Ai-Hua YE 1 , and Zhao-Xia WANG 1,3 1 Key Laboratory of Tea Biochemistry and Biotechnology, Ministry of Agriculture, Anhui Agricultural University, Hefei 230036, China; 2 College Of Horticulture, Nanjing Agriculture University, Nanjing 210095, China; 3 Auhui Institute of Education, Hefei 230036, China Abstract The complementary DNA (cDNA) amplified fragment length polymorphism technique was used to isolate transcript-derived fragments corresponding to genes involved in the flowering of tea plant. Comparative sequence analysis of an approximately 300 bp differential fragment amplified by primer combi- nation E 11 M 11 revealed 80%−84% similarity to the corresponding part of an α-tubulin gene of other species. The complete cDNA sequence of this α-tubulin was cloned by the rapid amplification of cDNA ends technique; its full length is 1537 bp and contains an open reading frame of 450 amino acid residues with two N- glycosylation sites and four protein kinase C phosphorylation sites. The deduced amino acid sequences did show significant homology to the α-tubulin from other plants that has been reported to be a pollen-specific protein and could be correlated with plant cytoplasm-nucleus-interacted male sterility. We named this complete cDNA Tua1. The nucleotide and amino acid sequence data of Tua1 have been recorded in the GenBank sequence database with the accession No. DQ340766. This Tua1 gene was cloned into the pET-32a expression system and expressed in Escherichia coli BL21trxB(DE3). The molecular weight of expressed protein was deduced to be approximately 49 kDa. Western blot analysis was used to identify the temporal expression of Tua1 in tea plant. Further studies of the effect of Tua1 protein on pollen tube growth indicated the Tua1 solution obviously promoted the growth of tea pollen tube. Key words cDNA-AFLP; Camellia sinensis; flower bud; α-tubulin; Western blot; pollen Received: April 19, 2006 Accepted: May 30, 2006 This work was supported by a grant from the Natural Science Foun- dation of Anhui Province (No. 050410102) *Corresponding author: Tel, 86-551-5156265; Fax, 86-551- 5156265; E-mail, [email protected] DOI: 10.1111/j.1745-7270.2006.00202.x Tea plant (Camellia sinensis) is an important economic crop in China. It is evergreen, perennial and cross- pollinated. Under cultivated conditions, a bush height of 60−100 cm is maintained for harvesting the tender leaves, a process that can continue for more than 100 years. It bears flowers and fruit 2−3 years after being planted. Under natural pollination conditions, the flowering rate of most tea plant varieties is usually very high, but there is a big difference in the fruiting rate among different tea varieties. The fruiting rate of some varieties is very low (e.g., ap- proximately 1% in Wulong and Maoxie) or absolutely blank (e.g., 0% in Zhenghe big white tea and Foshou), but in other varieties the rate is very high (e.g., 10% in Longjing43 and Wuniuzao) [1]. The flower bud differentiation of tea plant usually begins in early summer and lasts until late fall. It takes approximately 4 months from bud differen- tiation to bloom (Fig. 1). The reproductive growth of tea plant, including flower bud differentiation, flowering, blos- soming and fruiting, requires large amounts of energy and nutrients, which restrains vegetative growth and severely influences immunity function. As a result, reproductive growth is not only reducing the production of tea leaves, but also influencing the quality and resistance character of tea, which directly relates to the income of tea farmers. Additionally, tea is propagated either through seed or cutting. The segregation character of tea plant filial generation is often unavoidable due to the limitation of its cross-fertility. at Pennsylvania State University on May 17, 2016 http://abbs.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN 1672-9145 Acta Biochimica et Biophysica Sinica 2006, 38(9): 653–662 CN 31-1940/Q
©Institute of Biochemistry and Cell Biology, SIBS, CAS
Differentially Expression of Tua1, a Tubulin-encoding Gene, during Floweringof Tea Plant Camellia sinensis (L.) O. Kuntze Using cDNA
Amplified Fragment Length Polymorphism Technique
Wan-Ping FANG1,2, Chang-Jun JIANG1*, Mei YU1, Ai-Hua YE1, and Zhao-Xia WANG1,3
1 Key Laboratory of Tea Biochemistry and Biotechnology, Ministry of Agriculture, Anhui Agricultural University, Hefei 230036, China;2 College Of Horticulture, Nanjing Agriculture University, Nanjing 210095, China;
3 Auhui Institute of Education, Hefei 230036, China
Abstract The complementary DNA (cDNA) amplified fragment length polymorphism technique wasused to isolate transcript-derived fragments corresponding to genes involved in the flowering of tea plant.Comparative sequence analysis of an approximately 300 bp differential fragment amplified by primer combi-nation E11M11 revealed 80%−84% similarity to the corresponding part of an α-tubulin gene of other species.The complete cDNA sequence of this α-tubulin was cloned by the rapid amplification of cDNA ends technique;its full length is 1537 bp and contains an open reading frame of 450 amino acid residues with two N-glycosylation sites and four protein kinase C phosphorylation sites. The deduced amino acid sequences didshow significant homology to the α-tubulin from other plants that has been reported to be a pollen-specificprotein and could be correlated with plant cytoplasm-nucleus-interacted male sterility. We named this completecDNA Tua1. The nucleotide and amino acid sequence data of Tua1 have been recorded in the GenBanksequence database with the accession No. DQ340766. This Tua1 gene was cloned into the pET-32aexpression system and expressed in Escherichia coli BL21trxB(DE3). The molecular weight of expressedprotein was deduced to be approximately 49 kDa. Western blot analysis was used to identify the temporalexpression of Tua1 in tea plant. Further studies of the effect of Tua1 protein on pollen tube growth indicatedthe Tua1 solution obviously promoted the growth of tea pollen tube.
Key words cDNA-AFLP; Camellia sinensis; flower bud; α-tubulin; Western blot; pollen
Received: April 19, 2006 Accepted: May 30, 2006This work was supported by a grant from the Natural Science Foun-
dation of Anhui Province (No. 050410102)*Corresponding author: Tel, 86-551-5156265; Fax, 86-551-
5156265; E-mail, [email protected] DOI: 10.1111/j.1745-7270.2006.00202.x
Tea plant (Camellia sinensis) is an important economiccrop in China. It is evergreen, perennial and cross-pollinated. Under cultivated conditions, a bush height of60−100 cm is maintained for harvesting the tender leaves,a process that can continue for more than 100 years. Itbears flowers and fruit 2−3 years after being planted. Undernatural pollination conditions, the flowering rate of mosttea plant varieties is usually very high, but there is a bigdifference in the fruiting rate among different tea varieties.The fruiting rate of some varieties is very low (e.g., ap-proximately 1% in Wulong and Maoxie) or absolutely blank(e.g., 0% in Zhenghe big white tea and Foshou), but in
other varieties the rate is very high (e.g., 10% in Longjing43and Wuniuzao) [1]. The flower bud differentiation of teaplant usually begins in early summer and lasts until latefall. It takes approximately 4 months from bud differen-tiation to bloom (Fig. 1). The reproductive growth of teaplant, including flower bud differentiation, flowering, blos-soming and fruiting, requires large amounts of energy andnutrients, which restrains vegetative growth and severelyinfluences immunity function. As a result, reproductivegrowth is not only reducing the production of tea leaves,but also influencing the quality and resistance character oftea, which directly relates to the income of tea farmers.Additionally, tea is propagated either through seed or cutting.The segregation character of tea plant filial generation isoften unavoidable due to the limitation of its cross-fertility.
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

654 Acta Biochim Biophys Sin Vol. 38, No. 9
©Institute of Biochemistry and Cell Biology, SIBS, CAS
At the beginning of the 20th century, some tea plantbreeding researchers attempted to obtain a pure-breedingvariety by self-fertilization, but failed. Tea vegetative propa-gation is an effective method for maintaining the excellentcharacter of the maternal plant. Until now, it has been akey issue to control the reproductive growth of tea plantdue to increasing product demand, the need for qualityimprovement, and for tea plant breeding.
Flowering of tea plant is a continuous process, meaningdifferent developmental stages, from flower bud differen-tiation to full flower period, can occur in one plantsimultaneously. All of these stages are asynchronousprocesses in nature and difficult to analyze individually. Atpresent, research on specific characteristic products relatedto the quality of tea is developing quickly [2−4], but thereis little research on the flowering period of tea plant. In1987, Jiang and Wang published their research on thegrowth of pollen, the formation of pollen, the growth ofovule, the formation of embryo and the process ofendosperm growth [5]. Dong classified tea plant into threetypes: high fruitage rate type; low fruitage rate type; andsterile type [6]. However, until now, no reports have beenpublished on the flower developmental genes of tea plant.
In this study, we exploited the complementary DNAamplified fragment length polymorphism (cDNA-AFLP)technique to isolate transcript-derived fragments (TDFs)corresponding to genes involved in the flowering periodof tea plant and aimed at finding some clues to the mecha-nism of the plant’s reproductive growth. Research on theflowering period of tea plant from a molecular biologyangle can provide insight into the theory of the molecularmechanism of flowering of tea. Thus, it might be possibleto control the reproductive growth of tea plant usingbiotechnology, according to product demand. It could alsogive us a chance to use crop hybridity, so as to lay a good
theoretical foundation for the development of tea’s output,quality and resistance characters. We will attempt toprovide leads for further research on the floweringmechanism, as well as the fertility mechanism, of tea plantthrough our study. To our knowledge, this is the first studyof the flowering character of tea plant from the point ofview of molecular biology.
Materials and Methods
Plant materials and RNA isolation
In this study, we selected two tea plant cultivars plantedin the tea variety orchard of Anhui Agriculture University(Hefei, China), Longjin43 and Wulong. The flowering ratesof both are as high as 60%, but the fruiting rate ofLongjing43 (10%) is considerably higher than that ofWulong (1%). Other characters of both are basicallysimilar. We harvested the small flower buds [Diameter (Dm)=3 mm)] of both lines and the big flower buds (Dm=6 mm)of both cultivars in the same stock plant 1 month later.Flower buds were collected and immediately frozen inliquid nitrogen (Fig. 2). For each sample, total RNA was
Fig. 1 Blossoming and fruiting of Camellia sinensis1, first flowering period; 2, flower developing; 3, blossoming; 4, period of endflowering; 5, bearing fruit.
Fig. 2 Tea plant materials used in complementary DNAamplified fragment length polymorphism technique(A) Small flower bud of Longjing 43 (Dm=3 mm), harvested on July 20, 2004.(B) Big flower bud of Longjing 43 (Dm=6 mm), harvested on August 20, 2004.(C) Small flower bud of Wulong (Dm=3 mm) harvested on July 20, 2004. (D) Bigflower bud of Wulong (Dm=6 mm), harvested on August 20, 2004.
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

Sep., 2006 Wan-Ping FANG et al.: Tua1, a Tubulin-encoding Gene Differentially Expressed 655
http://www.abbs.info; www.blackwellpublishing.com/abbs
extracted from approximately 100 mg of flower buds usingTrizol plant RNA purification reagent (Gibco BRL,Gaithersburg, USA). The total RNA quantity was measuredusing a spectrophotometer at a wavelength of 260 nm.The purity of the RNA was evaluated by the ratio ofabsorbency at 260 and 280 nm (A260/A280). Quality of theRNA was checked by formaldehyde denaturing gelelectrophoresis.
cDNA-AFLP procedure
Double-stranded cDNA was synthesized using theSMART polymerase chain reaction (PCR) cDNA synthesiskit (Clontech, Palo Alto, USA). The cDNA-AFLP proceduredescribed by Bachem et al. [7] was used with a fewmodifications. The EcoRI/MseI enzyme system was used.The cDNA was then digested using EcoRI and MseI. Thefollowing restriction digests were then mixed: 20 μl cDNA(approximately 100 ng), 1 μl EcoRI (10 U), 4 μl 10×PCRbuffer, and 15 μl H2O. The mixture was incubated at atemperature appropriate for the EcoRI (65 ºC) for 2 h. Tothe first digest mixture, the following agents were added:1 μl 10×PCR buffer, 1 μl MseI (10 U), and 8 μl H2O.This mixture was incubated at 37 ºC for 2 h. For ligationof the adaptors (Table 1) to the digest mixture, the following
Table 1 Sequences of the primers and adaptors used for amplified fragment length polymorphism
Adaptor or primer Nucleotide sequence (5'→3') Adaptor or primer Nucleotide sequence (5'→3')
EcoRI MseIEcoRI adaptor 1 CTCGTAGACTGCGTACC MseI adaptor 1 GACGATGAGTCCTGAGEcoRI adaptor 2 AATTGGTACGCAGTC MseI adaptor 2 TACTCAGGACTCATE00
† GACTGCGTACCAATTC M00 GATGAGTCCTGAGTAAE11
‡ GACTGCGTACCAATTCAA M11 GATGAGTCCTGAGTAAAAE12 GACTGCGTACCAATTCAC M12 GATGAGTCCTGAGTAAACE13 GACTGCGTACCAATTCAG M13 GATGAGTCCTGAGTAAAGE14 GACTGCGTACCAATTCAT M14 GATGAGTCCTGAGTAAATE15 GACTGCGTACCAATTCCA M15 GATGAGTCCTGAGTAACAE16 GACTGCGTACCAATTCCC M16 GATGAGTCCTGAGTAACCE17 GACTGCGTACCAATTCCG M17 GATGAGTCCTGAGTAACGE18 GACTGCGTACCAATTCCT M18 GATGAGTCCTGAGTAACTE19 GACTGCGTACCAATTCGA M19 GATGAGTCCTGAGTAAGAE20 GACTGCGTACCAATTCGC M20 GATGAGTCCTGAGTAAGCE21 GACTGCGTACCAATTCGG M21 GATGAGTCCTGAGTAAGGE22 GACTGCGTACCAATTCGT M22 GATGAGTCCTGAGTAAGTE23 GACTGCGTACCAATTCTA M23 GATGAGTCCTGAGTAATAE24 GACTGCGTACCAATTCTC M24 GATGAGTCCTGAGTAATCE25 GACTGCGTACCAATTCTG M25 GATGAGTCCTGAGTAATGE26 GACTGCGTACCAATTCTT M26 GATGAGTCCTGAGTAATT
† preamplification primer; ‡ selective amplification primer.
agents were added: 1 μl EcoRI adaptors (5 pM), 1 μl MseIadaptors (5 pM), 0.5 μl of 10 mM ATP, 0.5 μl 10×PCRbuffer, 0.2 μl T4 DNA ligase, and 2 μl double-distilledH2O. The mixture was incubated for 3 h at 37 ºC. Theligation product was termed the primary template and its20-fold dilution was used directly for pre-amplification.
For selective amplification, a total of 16 primer pairswere used (Table 1) with the following PCR system: 2 μlsecondary template, 2 μl each of primer E11−E18 and primerM11−M18, 2 μl of 10×PCR buffer, 2.5 μl Mg2+ (25 mM),1 μl dNTPs (25 mM), 0.3 μl Taq polymerase (10 U), and10.5 μl double-distilled H2O. Amplification was carried outfor 12 cycles with 30 s denaturation at 94 ºC, 30 s annealingat 65 ºC (the annealing temperature touching down to 1 ºCfor every cycle) and 1 min extension at 72 ºC; then 23cycles with 30 s denaturation at 94 ºC, 30 s annealing at56 ºC, and a 1 min extension at 72 ºC. After the last cycle,the amplification was extended for 10 min at 72 ºC.
The selective amplification products were separated ona 6% polyacrylamide gel containing 8 M urea at 110 Wuntil the bromophenol blue reached the bottom. The cDNAbands were stained with silver nitrate, following the protocoldescribed in the DNA sequencing system kit (Promega,Madison, USA).
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

656 Acta Biochim Biophys Sin Vol. 38, No. 9
©Institute of Biochemistry and Cell Biology, SIBS, CAS
Characterization of AFLP fragments
Selected fragments were excised from the polyacryla-mide gel, suspended in H2O and eluted DNA wasreamplified using the same PCR conditions and the sameprimer combination for selective amplification. Reamplifiedproducts were checked on a 1% agarose gel [8]. The clonedDNA fragments were sequenced by Shanghai SangonCompany (Shanghai, China). The sequences obtained werecompared to those in the GenBank database using BLASTsequence alignments.
Rapid amplification of cDNA ends (3'/5' RACE) ofTua1 gene
Comparison of cDNA-AFLP patterns revealed differentcDNA fragments among the four samples. These bandswere excised from gels, cloned into the plasmid, andsequenced. We picked out and further analyzed a 300 bpdifferential fragment, ChaH-1, amplified by primercombination E11M11, and it revealed 80%−84% to α-tubulinof other species. It was particular expressed in the bigflower buds of both lines. The RACE procedure wascarr ied out using the BD SMART RACE cDNAamplification kit (Clontech). We designed primers andnested primers for RACE on the basis of the sequence ofChaH-1. We named the complete cDNA sequence Tua1.
The secondary structure of tea Tua1 gene was predictedusing the Garnier method (GOR) biological tool inPROSITE (http://www.expasy.org/prosite/) and Swiss-Prot (http://www.expasy.org/sprot/). Sequence analysiswas carried out using BLAST-W and DNAStar softwarein GenBank (http://www.ncbi.nih.gov/).
Reverse transcription (RT)-PCR identification
RT-PCR was carried out using 1.0 μl of first-strandcDNA prepared as described above. Primers specific forthe 5' RACE of ChaH-1 were used to standardize the genetranscript levels in different samples (leaves, small flowerbuds and big flower buds). PCR reactions of 25.0 μl weresubjected to 30 cycles of 30 s denaturing at 94 ºC, 30 sannealing at 58 ºC and 1 min extension at 72 ºC. Fivemicroliters of the product was electrophoresed on anagarose gel alongside a DNA quantification ladder, and thelevels of template added to the reaction were alteredaccording to the amount of expected product.
Prokaryotic expression of tea Tua1 gene
According to the complete cloned cDNA sequence, twoprimers were designed using the Primer Select Programin DNAStar: 5'-CTCGGATCCATGAGAGAGTTCATTTC-
GATC-3' (forward) and 5'-CTCAAGCTTATACTCAT-CACCTTCATCATC-3' (reverse). PCR products were cutwith HindIII and BamHI, then ligated with the pET-32avector, and transformed into Escherichia coli BL21trx(DE3). Induced expression was carried out according tothe manufacturer’s instructions (Novagen, California,USA). Target protein analysis was also carried outaccording to the user manual of the pET-32a vector(Novagen).
Western blot analysis
Total soluble protein was extracted from leaves, smallflower buds and big flower buds of two tea plant cultivars(approximately 250 mg per sample) by homogenizingin 3 ml extraction buffer (50 mM Tris-HCl, pH 8.0, 1 mMEDTA, 500 mM NaCl, 100 μg/ml phenylmethylsulfonylfluoride) and centrifuging at 15,000 g for 15 min at 4 °C.Protein concentrations were determined using the Bradfordmethod with some modification [9]. The supernatant wasmixed with 0.1 volume of Tris saline azide buffer kept onice for 10 min and centrifuged at 5000 rpm for 5 min. Thesupernatant was boiled for 5 min in 5×sodium dodecyl-sulfate (SDS) sample buffer and separated using 10%SDS-polyacrylamide gel electrophoresis (1 h, 120 V), thentransferred to a nitrocellulose membrane using a semi-dryblotter (Mini Gel Transfer apparatus; Bio-Rad, California,USA). The membrane was incubated for 2 h in Tris-bufferedsaline with 5% non-fat milk power, in Tris-buffered saline/Tween-20 for 1 h at room temperature, then probed insuccession with anti-α-tubulin antibody made in rabbit ata dilution of 1:1000 in 0.5% bovine gelatin in phosphate-buffered saline. Bound antibodies were detected withalkalinephosphatase-conjugated goat anti-rabbit IgGantibodies (Pierce, Rockford, USA), diluted to 1:5000 to1:10,000 in blocking buffer (both antibodies were purchasedfrom Beijing Biosynthesis Biotechnology Company, Beijing,China). The membranes were developed with 0.33 mg/ml1-nitroblue tetrazolium and 0.165 mg/ml 5-bromo-4-chloro-3-indolyl phosphate in alkaline phosphatase buffer(100 mM Tris, pH 9.5, 100 mM NaCl, 5 mM MgCl2).
Functional study of tea Tua1
To study the effect of Tua1 protein on pollen tubegrowth, we first optimized the isolated culture mediumfor tea plant pollen germination, then collected the pollen,and cultivated it for 12 h in two kinds of pollen germina-tion medium (total volume 5 ml per medium): medium I,10.0% (W/V) sucrose, 0.01% (W/V) boric acid, 1.0% agar;medium II, 10.0% (W/V) sucrose, 0.01% (W/V) boric acid,1.0% agar, 0.1 μg/μl Tua1 protein expressed in E. coli
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

Sep., 2006 Wan-Ping FANG et al.: Tua1, a Tubulin-encoding Gene Differentially Expressed 657
http://www.abbs.info; www.blackwellpublishing.com/abbs
Fig. 3 Complementary DNA amplified fragment length polymorphism differential display profiles of four samples of tea plantduring flower bud development (A), and enlarged sections of profile created by primer combination E11M11 (B,C)1, small flower buds of tea plant variety Longjing43; 2, big flower buds of Longjing43; 3, small flower buds of tea plant variety Wulong; 4, big flower buds of Wulong.Arrow represents the band of transcript-derived fragment ChaH-1.
BL21trxB(DE3).
Results
cDNA-AFLP analysis of gene differentially expressedduring flower development
The gene differential expression of small flower budsand big flower buds was compared using cDNA-AFLPtechnology. In total, 256 primer set combinations wereused per cDNA sample for selective amplification. A typicalexample of the obtained cDNA-AFLP gel is shown in Fig.3(A). The cDNA-AFLP produced an average 50 amplifi-cation products per PCR and therefore a total estimatednumber of 12,800 fragments. Distinct transcriptionalchanges between the small flower buds and the big flowerbuds of both varieties were observed for approximately
90 cDNA fragments, of which 37 were only expressed insmall flower buds of both varieties, and the other 53appeared in big flower buds of both varieties. This indicatedthese fragments, which specially expressed at differenttimes, were potentially related with the flower developmentof tea plant.
Characterization of AFLP fragments
We selected a part of these special expression fragments,which appeared simultaneously at different developmentalstages of flower buds of both varieties, for sequencing.Twelve TDFs were successfully re-amplified by PCR usingthe selective primers that were used to obtain the frag-ments in the cDNA-AFLP analysis. Amplified fragmentswere sent for DNA sequencing. Those fragments werecompared with those in the GenBank database using theBLAST search tool (Table 2). One sequence, namedChaH-1 [Fig. 3(B)], revealed high (80%−84%) similarity
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from
658 Acta Biochim Biophys Sin Vol. 38, No. 9
©Institute of Biochemistry and Cell Biology, SIBS, CAS
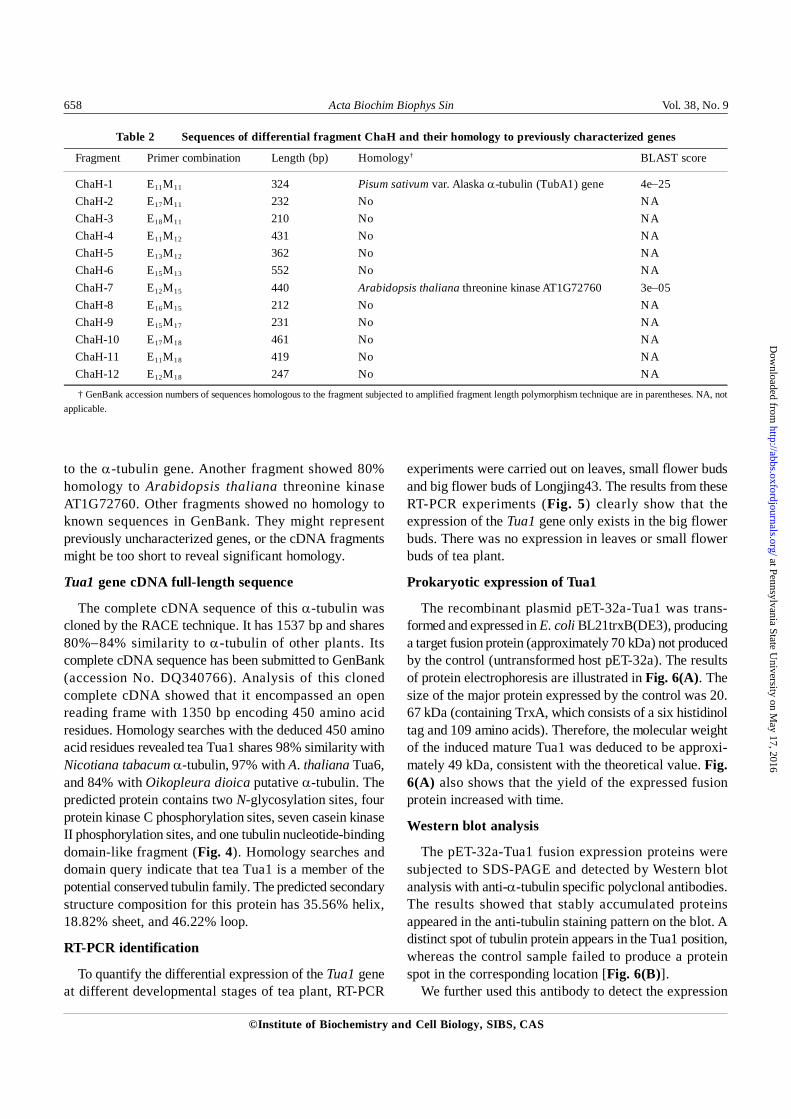
Table 2 Sequences of differential fragment ChaH and their homology to previously characterized genes
Fragment Primer combination Length (bp) Homology† BLAST score
ChaH-1 E11M11 324 Pisum sativum var. Alaska α-tubulin (TubA1) gene 4e−25ChaH-2 E17M11 232 No NAChaH-3 E18M11 210 No NAChaH-4 E11M12 431 No NAChaH-5 E13M12 362 No NAChaH-6 E15M13 552 No NAChaH-7 E12M15 440 Arabidopsis thaliana threonine kinase AT1G72760 3e−05ChaH-8 E16M15 212 No NAChaH-9 E15M17 231 No NAChaH-10 E17M18 461 No NAChaH-11 E11M18 419 No NAChaH-12 E12M18 247 No NA
† GenBank accession numbers of sequences homologous to the fragment subjected to amplified fragment length polymorphism technique are in parentheses. NA, notapplicable.
to the α-tubulin gene. Another fragment showed 80%homology to Arabidopsis thaliana threonine kinaseAT1G72760. Other fragments showed no homology toknown sequences in GenBank. They might representpreviously uncharacterized genes, or the cDNA fragmentsmight be too short to reveal significant homology.
Tua1 gene cDNA full-length sequence
The complete cDNA sequence of this α-tubulin wascloned by the RACE technique. It has 1537 bp and shares80%−84% similarity to α-tubulin of other plants. Itscomplete cDNA sequence has been submitted to GenBank(accession No. DQ340766). Analysis of this clonedcomplete cDNA showed that it encompassed an openreading frame with 1350 bp encoding 450 amino acidresidues. Homology searches with the deduced 450 aminoacid residues revealed tea Tua1 shares 98% similarity withNicotiana tabacum α-tubulin, 97% with A. thaliana Tua6,and 84% with Oikopleura dioica putative α-tubulin. Thepredicted protein contains two N-glycosylation sites, fourprotein kinase C phosphorylation sites, seven casein kinaseII phosphorylation sites, and one tubulin nucleotide-bindingdomain-like fragment (Fig. 4). Homology searches anddomain query indicate that tea Tua1 is a member of thepotential conserved tubulin family. The predicted secondarystructure composition for this protein has 35.56% helix,18.82% sheet, and 46.22% loop.
RT-PCR identification
To quantify the differential expression of the Tua1 geneat different developmental stages of tea plant, RT-PCR
experiments were carried out on leaves, small flower budsand big flower buds of Longjing43. The results from theseRT-PCR experiments (Fig. 5) clearly show that theexpression of the Tua1 gene only exists in the big flowerbuds. There was no expression in leaves or small flowerbuds of tea plant.
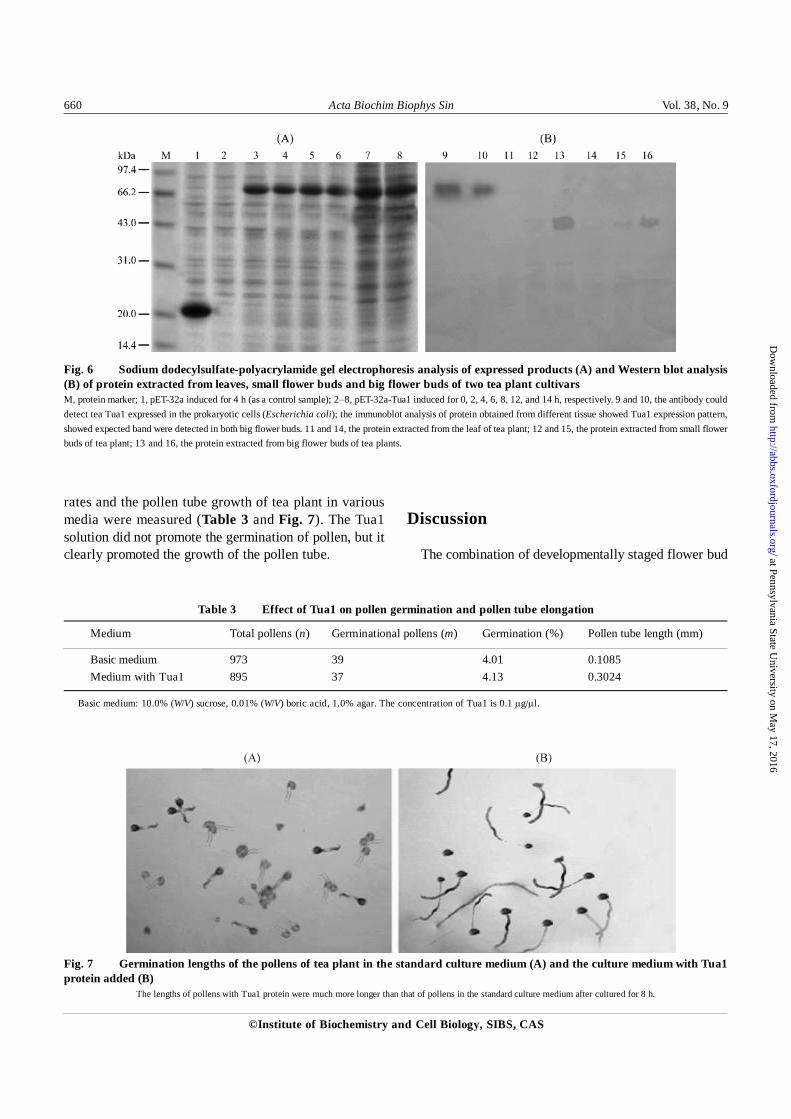
Prokaryotic expression of Tua1
The recombinant plasmid pET-32a-Tua1 was trans-formed and expressed in E. coli BL21trxB(DE3), producinga target fusion protein (approximately 70 kDa) not producedby the control (untransformed host pET-32a). The resultsof protein electrophoresis are illustrated in Fig. 6(A). Thesize of the major protein expressed by the control was 20.67 kDa (containing TrxA, which consists of a six histidinoltag and 109 amino acids). Therefore, the molecular weightof the induced mature Tua1 was deduced to be approxi-mately 49 kDa, consistent with the theoretical value. Fig.6(A) also shows that the yield of the expressed fusionprotein increased with time.
Western blot analysis
The pET-32a-Tua1 fusion expression proteins weresubjected to SDS-PAGE and detected by Western blotanalysis with anti-α-tubulin specific polyclonal antibodies.The results showed that stably accumulated proteinsappeared in the anti-tubulin staining pattern on the blot. Adistinct spot of tubulin protein appears in the Tua1 position,whereas the control sample failed to produce a proteinspot in the corresponding location [Fig. 6(B)].
We further used this antibody to detect the expression
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

Sep., 2006 Wan-Ping FANG et al.: Tua1, a Tubulin-encoding Gene Differentially Expressed 659
http://www.abbs.info; www.blackwellpublishing.com/abbs
Fig. 5 Electrophoresis analysis of reverse transcription-polymerase chain reaction (PCR) product of tissues from teaplant variety Longjing43M, marker; B, PCR product of big flower buds; S, PCR product of small flowerbuds; L, PCR product of leaves. Arrow represents the aimed PCR product.
pattern of Tua1 protein in tea plant. The immunoblot analy-sis of protein obtained from different tissue is shown inFig. 6(B). The expected band was detected in the big flowerbuds, but there was no protein corresponding to this bandin the leaves or small flower buds. Results demonstratedthat Tua1 protein was especially expressed in big flowerbuds of tea plant. This result is consistant with our cDNA-AFLP and RT-PCR analyses.
Effect of the Tua1 protein on the pollen tube growthof tea plant
The promotive experiments of pollen germination werecarried out on various mediums. The pollen germination
Fig. 4 Nucleotide sequence of the Tua1 gene and deduced amino acid sequenceBoxed amino acids are N-glycosylation sites; the amino acids underlined are protein kinase C phosphorylation sites; the amino acids underlined with dotted lines arecasein kinase II phosphorylation sites; the shading indicates tubulin subunits α, β, and γ signature. The asterisk indicates the termination codon.
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from
660 Acta Biochim Biophys Sin Vol. 38, No. 9
©Institute of Biochemistry and Cell Biology, SIBS, CAS
Fig. 6 Sodium dodecylsulfate-polyacrylamide gel electrophoresis analysis of expressed products (A) and Western blot analysis(B) of protein extracted from leaves, small flower buds and big flower buds of two tea plant cultivarsM, protein marker; 1, pET-32a induced for 4 h (as a control sample); 2−8, pET-32a-Tua1 induced for 0, 2, 4, 6, 8, 12, and 14 h, respectively. 9 and 10, the antibody coulddetect tea Tua1 expressed in the prokaryotic cells (Escherichia coli); the immunoblot analysis of protein obtained from different tissue showed Tua1 expression pattern,showed expected band were detected in both big flower buds. 11 and 14, the protein extracted from the leaf of tea plant; 12 and 15, the protein extracted from small flowerbuds of tea plant; 13 and 16, the protein extracted from big flower buds of tea plants.
Fig. 7 Germination lengths of the pollens of tea plant in the standard culture medium (A) and the culture medium with Tua1protein added (B)
The lengths of pollens with Tua1 protein were much more longer than that of pollens in the standard culture medium after cultured for 8 h.
rates and the pollen tube growth of tea plant in variousmedia were measured (Table 3 and Fig. 7). The Tua1solution did not promote the germination of pollen, but itclearly promoted the growth of the pollen tube.
Discussion
The combination of developmentally staged flower bud
Table 3 Effect of Tua1 on pollen germination and pollen tube elongation
Medium Total pollens (n) Germinational pollens (m) Germination (%) Pollen tube length (mm)
Basic medium 973 39 4.01 0.1085Medium with Tua1 895 37 4.13 0.3024
Basic medium: 10.0% (W/V) sucrose, 0.01% (W/V) boric acid, 1.0% agar. The concentration of Tua1 is 0.1 μg/μl.
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

Sep., 2006 Wan-Ping FANG et al.: Tua1, a Tubulin-encoding Gene Differentially Expressed 661
http://www.abbs.info; www.blackwellpublishing.com/abbs
samples with the cDNA-AFLP technique allowed us tocarry out a large screening for genes showing differentialexpression patterns during the flowering period.Mascarenhas distinguished two patterns of flower geneexpression. “Early” genes are transcribed soon aftermeiosis and are reduced or undetectable in mature pollen.Transcripts of the “late” genes are first detected aroundthe time of microspore mitosis and continue to accumulateas pollen matures [10]. In our study, we chose those TDFsthat appeared simultaneously in big flower buds of bothvarieties. Thus, we excluded the possibility of discrepancyin the two breeds themselves and confirmed that theseTDFs were exactly relevant to developmental stages offlower buds. According to the BLAST result of thesefragments, we selected a “late” gene fragment that wasdeduced to be an α-tubulin gene for further research andnamed it Tua1. The remaining TDFs obtained by cDNA-AFLP will be cloned and studied in more detail in the future.The α-tubulin gene of other species has been reported tobe a key protein in flower development [11−13]. Thephenomenon that the Tua1 gene was expressed preferen-tially in the big flower buds, and no expression was foundin the small flower buds or leaves of either line, indicatesthat the Tua1 gene might belong to the group of genesexpressed at the late stage of flower development.
Microtubules are components of the filamentous cyto-skeleton of eukaryotic cells and participate in many cellprocesses, including cell division, intracellular transport,cell motility, and cell morphogenesis [14−17]. In plants,microtubules have a number of specialized roles [18−20].The major structural component of microtubules is tubulin,a heterodimeric protein composed of two highly conservedsubunits, α and β. A less abundant form, γ-tubulin, is alsofound in higher plants [21,22]. Both α- and β-tubulins areencoded by multigene families in eukaryotes [23,24].Tissue-specific preferences in accumulation of tubulintranscripts have been reported in both Arabidopsis andmaize [11,25−27]. Up to the present, developmentallyregulated patterns of α-tubulin transcription in pollen havebeen mainly studied in Arabidopsis. The Arabidopsis α-tubulin gene, Tua1, is differentially expressed in flowerorgan of Arabidopsis, but no expression was detected inroot or leaf, and the peak transcription level of Tua1 wasnoted at the flowering period. It can be concluded that theTua1 gene plays a key role in the development of pollen ofArabidopsis [11,12]. The research on maize also indicatesthat the α-tubulin gene has a close relationship withcytoplasm-nucleus-interacted male sterility [13,28−30].
In our present study, the transcription and charac-terization analysis showed that Tua1 was expressed
References
1 Jiang CJ. Tea Plant Breeding. 1st ed. Beijing: China Agricultural Press2005
2 Jiang CJ. Tea Plant Breeding.1st ed. Beijing: China Agricultural Press2005: 11−13
3 Li YY, Jiang CJ, Wan XC, Zhang ZZ, Li DX. Purification and partialcharacterization of β-glucosidase from fresh leaves of tea plant [Camelliasinensis (L.) O. Kuntze]. Acta Biochim Biophys Sin 2005, 37: 363−370
4 Wang CX, Li YY, Jiang CJ, Yu YB. Molecular cloning and sequenceanalysis on cDNA of cystatin gene from tea leaves. J Tea Sci 2005, 25: 177−182
5 Jiang CJ, Wang ZH. Anatomical research on embryo development of teaplant. J Tea Sci 1987, 2: 23−28
6 Dong LJ. Fertility research of tea plant. J Tea Sci 1989, 1: 35−387 Bachem CWB, Oomen RJFJ, Visser RGF. Transcript imaging with cDNA-
AFLP: A step-by-step protocol. Plant Mol Biol Rep 1998, 16: 157−1738 Bachem CW, van der Hoeven RS, de Bruijn SM, Vreugdenhil D, Zabeau
M, Visser RG. Visualization of differential gene expression using a novelmethod of RNA fingerprinting based on AFLP: Analysis of gene expressionduring potato tuber development. Plant J 1996, 9: 745−753
9 Bradford MM. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding.Anal Biochem 1976, 72: 248−254
10 Mascarenhas JP. Gene activity during pollen development. Annu Rev Plant
differentially during the flowering period and might berelated to fertility of tea plant. The further functional studyindicated the Tua1 solution did not promote the germina-tion of pollen, but clearly promoted the growth of the pollentube. It will be useful to verify the gene function moredeeply and elucidate the biological roles and their relation-ship with the flower development of tea plant. Furtherwork will be focused on identifying the functionalmechanism of Tua1 in tea plant. A long-term goal of ourresearch is to identify expressed transcripts during flowerdevelopment, and to manipulate these genes or theirpromoter elements to regulate flower development andfertility mechanisms in tea plant. This work constitutesthe first report of genes activated during the floweringperiod of tea plant and has significance for tea plantbreeding.
Acknowledgements
We wish to thank Prof. Zheng-Zhu ZHANG (Key Labo-ratory of Tea Biochemistry and Biotechnology, Anhui Ag-riculture University) for helpful suggestions to our research,and Prof. Yu-Bao LI (Shanghai Institutes for BiologicalSciences, Chinese Academy Sciences) for his critical read-ing of this manuscript.
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from

662 Acta Biochim Biophys Sin Vol. 38, No. 9
©Institute of Biochemistry and Cell Biology, SIBS, CAS
Physiol Plant Mol Biol 1990, 41: 317−33811 Carpenter JL, Ploense SE, Snustad DP, Silflow CD. Preferential expres-
sion of an α-tubulin gene of Arabidopsis in pollen. Plant Cell 1992, 4:557−571
12 Carpenter JL, Kopczak SD, Snustad DP. Semi-constitutive expression ofan Arabidopsis thaliana α-tubulin gene. Plant Mol Biol 1993, 21: 937−942
13 Zhou HT, Xu L, Zhen WZ, Ling QT. Study on the relationship between α-tubulin gene and cytoplasm-nucleus-interacted male sterility (CMS) inMaize. Journal of Xiamen University (Natural Science) 2003, 42: 107−111
14 Goddard GH, Wick SM, Silflow CD, Snustad DP. Microtubule componentsof the plant cell cytoskeleton. Plant Physiol 1994, 104: 1−6
15 Cyr RJ, Palevitz BA. Organization of cortical microtubules in plant cells.Curr Opin Cell Biol 1995, 7: 65−71.
16 Kropf DL, Bisgrove SR, Hable WE. Cytoskeletal control of polar growthin plant cells. Curr Opin Cell Biol 1998, 10: 117−122
17 Bogre L, Calderini O, Merskiene I, Binarova P. Regulation of cell divisionand the cytoskeleton by mitogen-activated protein kinases in higher plants.Results Probl Cell Differ 2000, 27: 95−117
18 Giddings TH, Staehelin LA. Microtubule-mediated control of microfibrildeposition: A re-examination of the hypothesis. In: Lloyd CW ed. TheCytoskeletal Basis of Plant Growth and Form. London: Academic Press1991
19 Delmer DP, Amor Y. Cellulose biosynthesis. Plant Cell 1995, 7: 987−1000
20 Whittaker DJ, Triplett BA. Gene-specific changes in α-tubulin transcriptaccumulation in developing cotton fibers. Plant Physiol 1999, 121: 181−188
21 Oakley CE, Oakley BR. Identification of γ-tubulin, a new member of thetubulin superfamily encoded by mipA gene of Aspergillus nidulans. Nature1989, 338: 662−664
22 Liu B, Joshi HC, Wilson TJ, Silflow CD, Palevitz BA, Snustad DP. γ-Tubulin in Arabidopsis: Gene sequence, immunoblot, and immunofluores-cence studies. Plant Cell 1994, 6: 303−314
23 Cleveland DW, Sullivan KF. Molecular biology and genetics of tubulin.Annu Rev Biochem 1985, 54: 331−365
24 Silflow CD, Oppenheimer DG, Kopczak SD, Ploense SE, Ludwig SR,Haas NA, Snustad DP. Plant tubulin genes: Structure and differential ex-pression during development. Dev Genet 1987, 8: 435−460
25 Snustad DP, Haas NA, Kopczak SD, Silflow CD. The small genome ofArabidopsis thaliana contains at least nine expressed α-tubulin genes.Plant Cell 1992, 4: 549−556
26 Uribe X, Torres MA, Capellades M, Puigdomenech P, Rigau J. Maize α-tubulin genes are expressed according to specific patterns of cell differentiation.Plant Mol Biol 1998, 37: 1067−1078
27 Villemur R, Haas NA, Joyce CM, Snustad DP, Silflow CD. Characterizationof four new β-tubulin genes and their expression during male flowerdevelopment in maize (Zea mays L.). Plant Mol Biol 1994, 24: 295−315
28 Montoliu L, Puigdomenech P, Rigau J. The Tub α3 gene from Zea mays:Structure and expression in dividing plant tissues. Gene 1990, 94: 201−207
29 Montoliu L, Rigau J, Puigdomenech P. Multiple polyadenylation sites areactive in the α1-tubulin gene from Zea mays. FEBS Lett 1990, 277: 29−32
30 Montoliu L, Rigau J, Puigdomenech P. A tandem of α-tubulin genespreferentially expressed in radicular tissues from Zea mays. Plant Mol Biol1990, 14: 1−15
Edited byYu-Jun ZHANG
at Pennsylvania State University on M
ay 17, 2016http://abbs.oxfordjournals.org/
Dow
nloaded from
Related Documents











