FABPs in Mouse Adrenal Gland 77 77 Tohoku J. Exp. Med., 2004, 203, 77-86 Received March 29, 2004; revision accepted for publication April 9, 2004. Address for reprints: Yuji Owada, Division of Histology, Department of Cell Biology, Graduate School of Medicine, Tohoku University, 2-1 Seiryomachi, Aoba-ku, Sendai 980-8575, Japan. e-mail: [email protected] Differential Localization of Brain-Type and Epidermal- Type Fatty Acid Binding Proteins in the Adrenal Gland of Mice XIA Y UN, 1 MOHAMMAD REZA NOURANI, SOHA ABDELKAWI ABDELWAHAB, NORIKO KITANAKA, Y UJI OWADA, FRIEDRICH SPENER, 2 HIROO IWASA, AKIRA T AKAHASHI 1 and HISATAKE KONDO Division of Histology, Department of Cell Biology, 1 Division of Neuro-Intravas- cular Disease, Department of Neuroscience, Graduate School of Medicine, To- hoku University, Sendai 980-8575, and 2 Department of Biochemistry, Muenster University, Muenster, Germany Y UN, X., NOURANI, M.R., ABDELWAHAB, S.A., KITANAKA, N., OWADA, Y., SPENER, F., IWASA, H., T AKAHASHI, A. and KONDO, H. Differential Localization of Brain-Type and Epidermal-Type Fatty Acid Binding Proteins in the Adrenal Gland of Mice. Tohoku J. Exp. Med., 2004, 203 (2), 77-86 ── In immuno-light and -electron microscopy, brain-type fatty acid binding protein (B-FABP) is localized in the sus- tentacular cells enclosing the chromaffin cells in the adrenal medulla. This represents another new feature commonly shared by the sustentacular cells and ganglionic satel- lite cells, the latter of which has already been reported to localize this molecule, and suggests a common feature in lipid metabolism shared by the two cells enclosing pe- ripheral neurons and paraneurons. On the other hand, epidermal-type fatty acid bind- ing protein (E-FABP) is localized in two discrete cells in the adrenal gland: the one is a subpopulation of intra-adrenal macrophages which are intensely immunoreactive for F4/80, a marker of macrophages, and are rich in pleomorphic lysosomes. Because of their direct apposition to adjacent cortical endocrine cells and medullary chromaf- fin cells, the macrophages may be involved not only in phagocytosis of degenerating adrenal cells but also in exertion of some yet unknown effects on the endocrine func- tion of the cortical and medullary cells via humoral factors such as cytokines which have recently been known to be secreted by macrophages. The other is a population of cells having scanty perikaryal cytoplasm poor in organneles and several thinny ex- tended processes in the cortex and exhibiting weak immunoreactivity for E-FABP. The possible natures of these cells immunoreactive for E-FABP are discussed in view of a subpopulation of endothelial cells or the dendritic cells of antigen-presenting property. ──── B-FABP; E-FABP; sustentacular cells; adrenal gland; immuno- histochemistry © 2004 Tohoku University Medical Press

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

FABPs in Mouse Adrenal Gland 77
77
Tohoku J. Exp. Med., 2004, 203, 77-86
Received March 29, 2004; revision accepted for publication April 9, 2004.Address for reprints: Yuji Owada, Division of Histology, Department of Cell Biology, Graduate School of
Medicine, Tohoku University, 2-1 Seiryomachi, Aoba-ku, Sendai 980-8575, Japan.e-mail: [email protected]
Differential Localization of Brain-Type and Epidermal-Type Fatty Acid Binding Proteins in the Adrenal Gland of Mice
XIA YUN,1 MOHAMMAD REZA NOURANI, SOHA ABDELKAWI ABDELWAHAB, NORIKO KITANAKA, YUJI OWADA, FRIEDRICH SPENER,2 HIROO IWASA, AKIRA TAKAHASHI
1 and HISATAKE KONDO
Division of Histology, Department of Cell Biology, 1Division of Neuro-Intravas-cular Disease, Department of Neuroscience, Graduate School of Medicine, To-hoku University, Sendai 980-8575, and2Department of Biochemistry, Muenster University, Muenster, Germany
YUN, X., NOURANI, M.R., ABDELWAHAB, S.A., KITANAKA, N., OWADA, Y., SPENER, F., IWASA, H., TAKAHASHI, A. and KONDO, H. Differential Localization of Brain-Type and Epidermal-Type Fatty Acid Binding Proteins in the Adrenal Gland of Mice. Tohoku J. Exp. Med., 2004, 203 (2), 77-86 ── In immuno-light and -electron microscopy, brain-type fatty acid binding protein (B-FABP) is localized in the sus-tentacular cells enclosing the chromaffin cells in the adrenal medulla. This represents another new feature commonly shared by the sustentacular cells and ganglionic satel-lite cells, the latter of which has already been reported to localize this molecule, and suggests a common feature in lipid metabolism shared by the two cells enclosing pe-ripheral neurons and paraneurons. On the other hand, epidermal-type fatty acid bind-ing protein (E-FABP) is localized in two discrete cells in the adrenal gland: the one is a subpopulation of intra-adrenal macrophages which are intensely immunoreactive for F4/80, a marker of macrophages, and are rich in pleomorphic lysosomes. Because of their direct apposition to adjacent cortical endocrine cells and medullary chromaf-fin cells, the macrophages may be involved not only in phagocytosis of degenerating adrenal cells but also in exertion of some yet unknown effects on the endocrine func-tion of the cortical and medullary cells via humoral factors such as cytokines which have recently been known to be secreted by macrophages. The other is a population of cells having scanty perikaryal cytoplasm poor in organneles and several thinny ex-tended processes in the cortex and exhibiting weak immunoreactivity for E-FABP. The possible natures of these cells immunoreactive for E-FABP are discussed in view of a subpopulation of endothelial cells or the dendritic cells of antigen-presenting property. ──── B-FABP; E-FABP; sustentacular cells; adrenal gland; immuno-histochemistry© 2004 Tohoku University Medical Press

X. Yun et al.78 FABPs in Mouse Adrenal Gland 79
Because of their hydrophobic nature, fatty acids should be solubilized and transported by specific intracellular lipid binding proteins, the low molecular mass polypeptides of 14-15 kDa termed fatty acid binding proteins (FABPs). Multiple species of FABPs had been identified and named according to the tissues of their first isolation sites such as heart (H-), epidermal (E-), brain (B-) and adipocyte (A-) types, however, they showed much wider tissue distribution than first thought (Gordon et al. 1983; Alpers et al. 1984; Hunt et al. 1986; Siegenthaler et al. 1993; Feng et al. 1994; Kurtz et al. 1994). Several roles have been assigned to these FABPs such as con-trol of cellular uptake of fatty acids and their sub-sequent utilization and intracellular compartmen-tation, modulation of activity of enzymes involved in fatty acid metabolism, protection of cellular membranes and enzymes from detergent effects of fatty acids, and carriers of signaling fatty acids (Glatz et al. 1993; Coe and Bernlohr 1998).
Based on the idea that the exact localization of a given molecule is a first step and great help in understanding its functional significance, we have so far examined the localization of various types of FABPs in a series of studies (Iseki et al. 1988; Watanabe et al. 1991; Owada et al. 1996a, b, 1997, 2001, 2002a; Abdelwahab et al. 2003; Kitanaka et al. 2003). Here we show that B-FABP is localized in the sustentacular cells enclosing the chromaffin cells in the adrenal medulla, while E-FABP is localized in a small number of pre-sumptive dendritic cells distributed throughout the adrenal grand of postnatal mice.
MATERIALS AND METHODS
Male C57BL/6 mice at the stages of postna-tal day (P) 1 week (w), P2w and P7w were used in this study. All experimental protocols were re-viewed by the Committee on the Ethics of Animal Experiments of Tohoku University, and were car-ried out in accordance with the Guidelines for Animal Experiments issued by Graduate School of Medicine, Tohoku University. Mice were per-fused under Nembutal anesthesia through the
heart with 4% paraformaldehyde/0.1 M phosphate buffer (pH 7.4). The adrenal glands were extir-pated and immersed overnight in a phosphate buf-fer containing 30% sucrose. Antibodies used in this study include E-FABP antiserum at a concen-tration of 0.5 μg/ml, B-FABP antiserum at a con-centration of 2 μg/ml and a monoclonal rat anti-body against F4/80 (Serotec, Oxford, UK) at 1 :100 dilution. The characterization of anti-E-FABP and anti-B-FABP antibodies were described previously (Kurtz et al. 1994; Owada et al. 2002b). After incubation with the primary anti-body, the sections were incubated with biotinylat-ed anti-rabbit secondary antibody for single im-munostaining or with a combination of anti-rabbit IgG-Alexa488 and anti-rat IgG-Alexa594 (Molecular Probe, Eugene, OR, USA) for double immunostaining, respectively. The sections for single immunostaining were subsequently visual-ized using ABC (avidin-biotinylated peroxidase complex) system (Vector Laboratory, Burlingame, CA, USA) with DAB as a substrate. For immu-no-electron microscopy, some of the sections were postfixed with 1% OsO4 in 0.1% cacodylate buffer, pH 7.4, for 20 minutes after completion of the ABC procedure. They were subsequently em-bedded in Epon according to the conventional procedure and ultrathin sections were examined after brief staining with uranyl acetate. For con-trol of E-FABP-immunoreactivity, the adrenal gland of E-FABP-gene knockout mice were im-munostained which had been generated and char-acterized by us (Owada et al. 2002b), while the B-FABP-antibody was immunoabsorbed with its antigen at 10 μg/ml in antiserum for control of B-FABP-immunoreactivity.
RESULTS
Cells immunoreactive for B-FABP in the med-dula
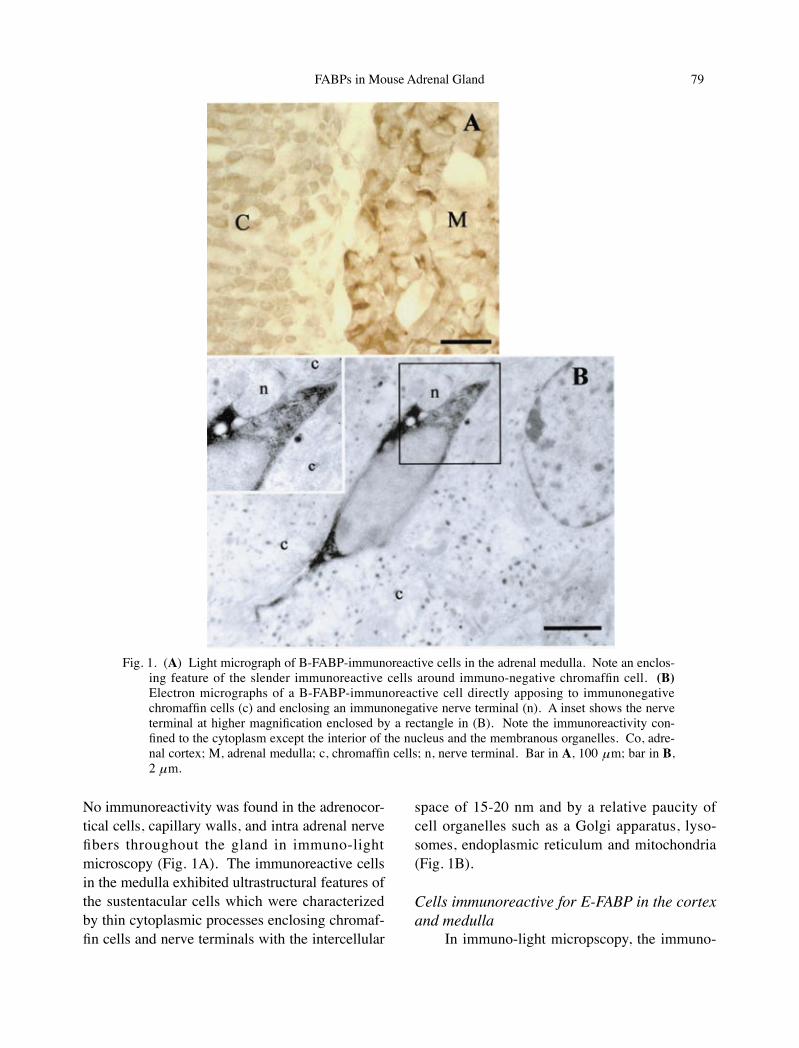
The immunoreactivity for B-FABP was de-tected in numerous small cells having thin pro-cesses in the medulla. They were distributed rather evenly in the medulla and enclosed multi-ple immunonegative chromaffin cells as groups.
X. Yun et al.78 FABPs in Mouse Adrenal Gland 79
No immunoreactivity was found in the adrenocor-tical cells, capillary walls, and intra adrenal nerve fibers throughout the gland in immuno-light microscopy (Fig. 1A). The immunoreactive cells in the medulla exhibited ultrastructural features of the sustentacular cells which were characterized by thin cytoplasmic processes enclosing chromaf-fin cells and nerve terminals with the intercellular
space of 15-20 nm and by a relative paucity of cell organelles such as a Golgi apparatus, lyso-somes, endoplasmic reticulum and mitochondria (Fig. 1B).
Cells immunoreactive for E-FABP in the cortex and medulla
In immuno-light micropscopy, the immuno-
Fig. 1. (A) Light micrograph of B-FABP-immunoreactive cells in the adrenal medulla. Note an enclos-ing feature of the slender immunoreactive cells around immuno-negative chromaffin cell. (B) Electron micrographs of a B-FABP-immunoreactive cell directly apposing to immunonegative chromaffin cells (c) and enclosing an immunonegative nerve terminal (n). A inset shows the nerve terminal at higher magnification enclosed by a rectangle in (B). Note the immunoreactivity con-fined to the cytoplasm except the interior of the nucleus and the membranous organelles. Co, adre-nal cortex; M, adrenal medulla; c, chromaffin cells; n, nerve terminal. Bar in A, 100 μm; bar in B, 2 μm.
X. Yun et al.80 FABPs in Mouse Adrenal Gland 81
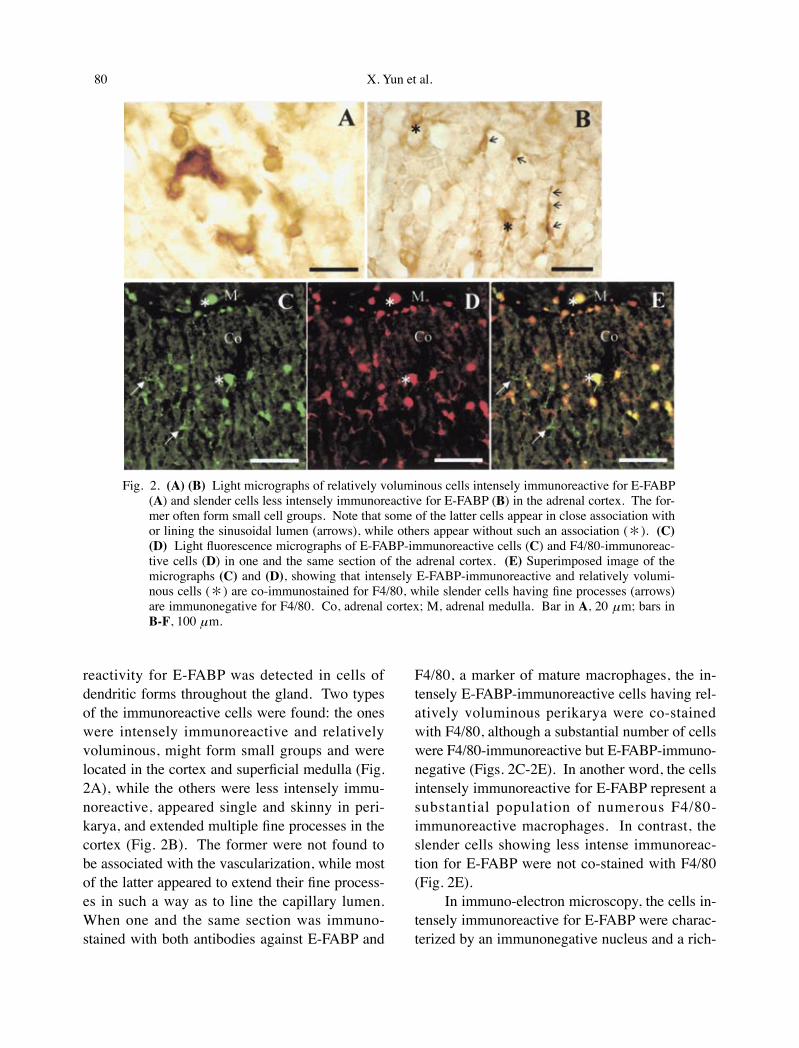
reactivity for E-FABP was detected in cells of dendritic forms throughout the gland. Two types of the immunoreactive cells were found: the ones were intensely immunoreactive and relatively voluminous, might form small groups and were located in the cortex and superficial medulla (Fig. 2A), while the others were less intensely immu-noreactive, appeared single and skinny in peri-karya, and extended multiple fine processes in the cortex (Fig. 2B). The former were not found to be associated with the vascularization, while most of the latter appeared to extend their fine process-es in such a way as to line the capillary lumen. When one and the same section was immuno-stained with both antibodies against E-FABP and
F4/80, a marker of mature macrophages, the in-tensely E-FABP-immunoreactive cells having rel-atively voluminous perikarya were co-stained with F4/80, although a substantial number of cells were F4/80-immunoreactive but E-FABP-immuno-negative (Figs. 2C-2E). In another word, the cells intensely immunoreactive for E-FABP represent a substantial population of numerous F4/80-immunoreactive macrophages. In contrast, the slender cells showing less intense immunoreac-tion for E-FABP were not co-stained with F4/80 (Fig. 2E).
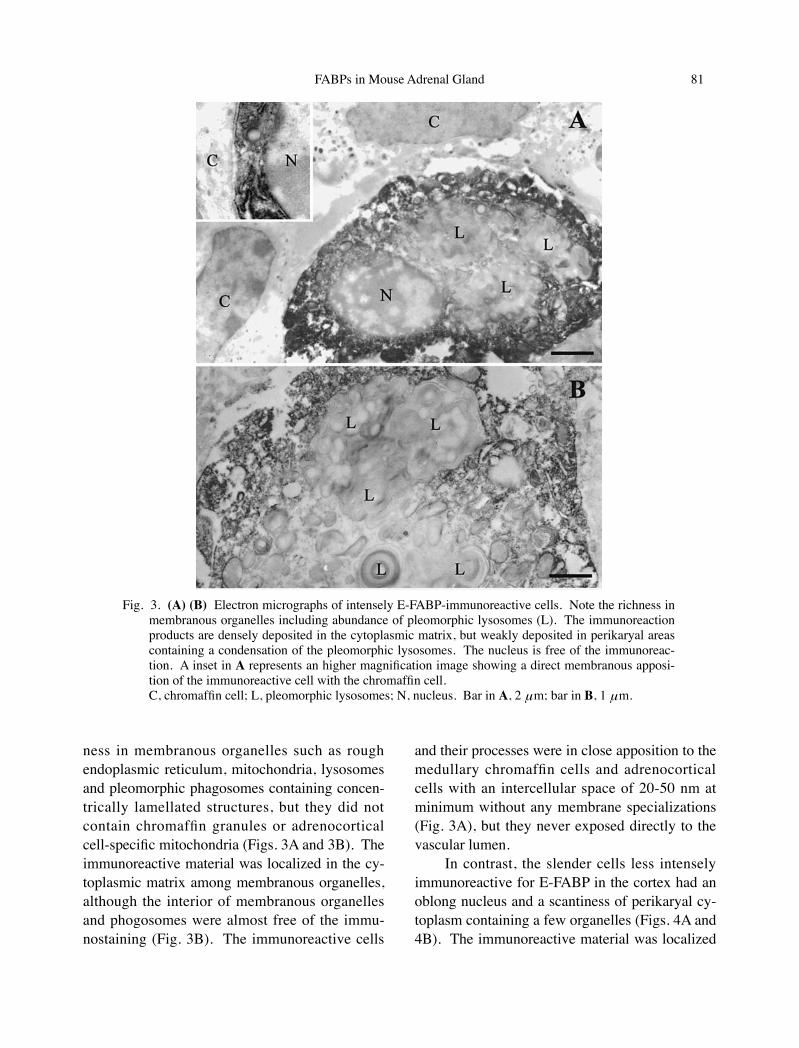
In immuno-electron microscopy, the cells in-tensely immunoreactive for E-FABP were charac-terized by an immunonegative nucleus and a rich-
Fig. 2. (A) (B) Light micrographs of relatively voluminous cells intensely immunoreactive for E-FABP (A) and slender cells less intensely immunoreactive for E-FABP (B) in the adrenal cortex. The for-mer often form small cell groups. Note that some of the latter cells appear in close association with or lining the sinusoidal lumen (arrows), while others appear without such an association (*). (C) (D) Light fluorescence micrographs of E-FABP-immunoreactive cells (C) and F4/80-immunoreac-tive cells (D) in one and the same section of the adrenal cortex. (E) Superimposed image of the micrographs (C) and (D), showing that intensely E-FABP-immunoreactive and relatively volumi-nous cells (*) are co-immunostained for F4/80, while slender cells having fine processes (arrows) are immunonegative for F4/80. Co, adrenal cortex; M, adrenal medulla. Bar in A, 20 μm; bars in B-F, 100 μm.
X. Yun et al.80 FABPs in Mouse Adrenal Gland 81
ness in membranous organelles such as rough endoplasmic reticulum, mitochondria, lysosomes and pleomorphic phagosomes containing concen-trically lamellated structures, but they did not contain chromaffin granules or adrenocortical cell-specific mitochondria (Figs. 3A and 3B). The immunoreactive material was localized in the cy-toplasmic matrix among membranous organelles, although the interior of membranous organelles and phogosomes were almost free of the immu-nostaining (Fig. 3B). The immunoreactive cells
and their processes were in close apposition to the medullary chromaffin cells and adrenocortical cells with an intercellular space of 20-50 nm at minimum without any membrane specializations (Fig. 3A), but they never exposed directly to the vascular lumen.
In contrast, the slender cells less intensely immunoreactive for E-FABP in the cortex had an oblong nucleus and a scantiness of perikaryal cy-toplasm containing a few organelles (Figs. 4A and 4B). The immunoreactive material was localized
Fig. 3. (A) (B) Electron micrographs of intensely E-FABP-immunoreactive cells. Note the richness in membranous organelles including abundance of pleomorphic lysosomes (L). The immunoreaction products are densely deposited in the cytoplasmic matrix, but weakly deposited in perikaryal areas containing a condensation of the pleomorphic lysosomes. The nucleus is free of the immunoreac-tion. A inset in A represents an higher magnification image showing a direct membranous apposi-tion of the immunoreactive cell with the chromaffin cell.
C, chromaffin cell; L, pleomorphic lysosomes; N, nucleus. Bar in A, 2 μm; bar in B, 1 μm.

X. Yun et al.82 FABPs in Mouse Adrenal Gland 83
in the cytoplasmic matrix but not within the nu-cleus and membranous organelles. Some of the slender immunoreactive cells appeared in single ultrathin sections as lining the sinusoidal capillary lumen: their perikarya were bulged into and ex-posed directly to the capillary lumen, and they had thin immunopositive cytoplasmic processes to line the capillary lumen together with immuno-
negative endothelial cells with simple membra-nous appositions (Fig. 4A). However, others ap-peared to locate their entire perikarya in the interstitium without direct exposure to the capil-lary lumen (Fig. 4B). They sent tapering process-es among adjacent adrenocortical cells or into subendothelial spaces (Fig. 4B).
Fig. 4. (A) Electron micrographs of a less intensely E-FABP-immunoreactive cell lining the sinusoidal lumen (S) together with immunonegative endothelial cells. (B) E-FABP-immunoreactive cell located in the interstitium among cortical cells (Co) without direct exposure to the sinusoidal lumen. Note the scanty in their cytoplasm and organelles and the extension of tapering processes. The immunoreactive products are present in the cytoplasmic matrix except for the nuclear interior (*). Co, cortical cells; E, endothelial cells; S, sinusoidal lumen; Bars in A and B, 1 μm.

X. Yun et al.82 FABPs in Mouse Adrenal Gland 83
Authenticity of the immunoreactivityWhen the adrenal gland from the gene
knockout mice for E-FABPs, the details of which had already been reported by us (Owada et al. 2002b), were immunostained with the E-FABP antibody, or the antigen-absorbed serum for B-FABP was applied to sections of the wild adre-nal gland, no immunoreaction was detected at a significant level in any portions of the gland (data not shown). These indicate that the immunoreac-tivity in the satellite cells and that in the cells of dendritic form are due to authentic E- and E-FABPs, respectively.
DISCUSSION
With regard to the present finding of B-FABP localization in the sustentacular cells enclosing the adrenal chromaffin cells, the immunohisto-chemical studies have so far clarified that this molecule is localized in the ganglionic satellite cells of the adult peripheral ganglia as well as the ventricular germinal zone cells and radial glia of the embryonic brain and astrocytes of the postna-tal brain (Kurtz et al. 1994). It is well known that both adrenal chromaffin cells and most peripheral ganglion neurons are of neural crest origin (Hamilton and Mossman 1972), that both the sus-tentacular cells and ganglionic satellite cells com-monly share the enclosing feature in relation to the chromaffin cells and ganglionic neurons re-spectively, and that the sustentacular cells and ganglionic satellite cells exhibit the histological contiguity to Schwann cells enclosing nerve fibers innervating the chromaffin cells and ganglionic neurons (Coupland 1965). Because of the com-mon features, it is generally expected to find con-stitutive molecules shared by both the sustentacu-lar cells, ganglionic satellite cells and Schwann cells, and a well-known example of the molecules shared by these cells is S-100β (Cocchia and Michetti 1981). Although B-FABP is shown in the present study as another new example of such common molecules in the sustentacular cells and ganglionic satellite cell, a peculiarity of B-FABP is that this molecule is not localized in Schwann
cells in normal mice (Kurtz et al. 1994). In this regard, it should be noted that B-FABP expression has been reported to be induced in the Schwann cells of sciatic nerve after nerve injury (Miller et al. 2003) and that the authors have suggested the possible mediation of its expression by the Ras-independent epidermal growth factor receptor sig-naling. It is possible to speculate that there is a yet unknown difference in the lipid metabolism and signaling between the sustentacular and satel-lite cells on one hand and the Schwann cells on the other hand, and that B-FABP expression is regulated by the fatty acid environment of given cells.
With regard to the nature of the cells intense-ly immunoreactive for E-FABP, because of their double-immunostaining with F4/80 antibody, they are regarded as a subpopulation of intra-adrenal macrophages whose abundant existence has al-ready been described by previous authors (Mausle 1974; Magalhaes and Magalhaes 1980, 1984; Hume et al. 1984; Gonzalez-Hernandez et al. 1994). The occurrence of E-FABP in such a sub-stantial, but not all, population of the macro-phages suggests that E-FABP may be involved in some specific roles, but not general to the intra-adrenal macrophages. Considering their direct ap-position to adjacent cortical endocrine cells and medullary chromaffin cells, the macrophages may be involved not only in phagocytosis of degener-ating adrenal cells but also in exertion of some yet unknown affects on the endocrine function of the cortical and medullary cells via humoral factors such as cytokines or eicosanoids which are secret-ed by macrophages (Currie et al. 2000).
The nature of the slender cells less intensely immunoreactive for E-FABP in the cortex, is enigma. Because of the absence of immunostain-ing for F4/80 and paucity of cell organelles in-cluding lysosomes in their cytoplasm, it is clear that the cells are discrete from the macrophages. Several interpretations are possible: the one is that the immunoreactive cells represent a subpopula-tion of fibroblasts because of their location. However, there have so far been no reports de-

X. Yun et al.84 FABPs in Mouse Adrenal Gland 85
scribing the occurrence of E-FABP-immuno-reactive fibroblasts in other tissues and organs. Another possible interpretation is that the cortical endothelial cells are heterogeneous in terms of fatty acid-related activities, and a type of the en-dothelial cells, which are highly involved in the activities, exhibits E-FABP-immunoreactivity, and that the immunoreactive cell profiles in the interstitium represent tangential section profiles of the immunoreactive endothelial cells. The occur-rence of E-FABP immunoreactivity in endothelial cells has been reported in some organs (Masouye et al. 1997). The other possibility to be considered is that the immunoreactive cells are the dendritic cells involved in the antigen presentation because of their dendritic morphology. There have been several studies indicating that the dendritic cells are present in the interstitium of most organs of so called non-immune nature excluding the brain, and that their occurrence is in general at a fre-quency so low that this has posed a major impedi-ment to their detection (Bell et al. 1999). The dendritic cells can usually be characterized by a high-level expression of class II MHC molecules. Because of the fact that the antigenicity of class II MHC molecules is in most cases preserved at suf-ficient levels for immunohistochemistry only by acetone fixation, there has been few information on the fine structural features of the dendritic cells. By partially overcoming these impedi-ments, the occurrence of the dendritic cells have recently been described in a light microscopic study of the adrenal gland (Sato 1998). The simi-larity of the present skinny cells showing less in-tense immunoreactivity for E-FABP to the MHC II-immunoreactive dendritic cells in the adrenal cortex in terms of the cell shape, size and distribu-tion, is in favor of the presumption that the pres-ent E-FABP-immunoreactive cells of dendritic forms are the dendritic cells although its confir-mation remains to be done by simultaneous ex-amination of expression of class II MHC antigen although it is actually difficult as discussed above. In this regard, we have recently disclosed that E-FABP-immunoreactivity is localized in the den-
dritic cells in the splenic white pulp, but not in the splenic marginal zone (Kitanaka et al. 2003). It is known that the dendritic cells in the splenic white pulp except for those in the follicles are positive for CD8, a surface antigen, while those in the marginal zone are CD8(−) and much more numer-ous than the former (Leenen et al. 1998). The rather frequent appearance of some of the immu-noreactive slender cells in direct contiguity to E-FABP-immunonegative endothelial cells and lining the capillary lumen suggests that the cells under discussion could move away constantly and frequently from the capillary lining into the lumi-nal blood flow or into the interstitium. This movement would be understandable if the cells under discussion be the dendritic cells which be-long to a hematopoietic lineage (Bell et al. 1999).
In order to further understand the functional significance of B-FABP in the sustentacular cells and that of E-FABP in adrenal macrophages and presumptive dendritic cells or endothelial cells in the adrenal cortex, the analyses of phenotypes of these respective cells from B- and E-FABP gene knockout mice recently generated by us (Owada et al. 2002b) is crucial, which is underway in our laboratory.
ReferencesAbdelwahab, S.A., Owada, Y., Kitanaka, N., Iwasa, H.,
Sakagami, H. & Kondo, H. (2003) Localization of brain-type fatty acid-binding protein in Kupffer cells of mice and its transient decrease in response to lipopolysaccharide. Histochem. Cell Biol., 119, 469-475.
Alpers, D.H., Strauss, A.W., Ockner, R.K., Bass, N.M. & Gordon, J.I. (1984) Cloning of a cDNA en-coding rat intestinal fatty acid binding protein. Proc. Natl. Acad. Sci. USA, 81, 313-317.
Bell, D., Young, J.W. & Banchereau, J. (1999) Den-dritic cells. Adv. Immunol., 72, 255-324.
Cocchia, D. & Michetti, F. (1981) S-100 antigen in satellite cells of the adrenal medulla and the su-perior cervical ganglion of the rat. An immuno-chemical and immunocytochemical study. Cell Tissue. Res., 215, 103-112.
Coe, N.R. & Bernlohr, D.A. (1998) Physiological properties and functions of intracellular fatty

X. Yun et al.84 FABPs in Mouse Adrenal Gland 85
acid-binding proteins. Biochim. Biophys. Acta, 1391, 287-306.
Coupland, R.E. (1965) Electron microscopic observa-tion on the structure of the rat adrenal medulla. J. Anat., 99, 231-272.
Currie, K.P., Zhou, Z. & Fox, A.P. (2000) Evidence for paracrine signaling between macrophages and bovine adrenal chromaffin cell Ca(2+) channels. J. Neurophysiol., 83, 280-287.
Feng, L., Hatten, M.E. & Heintz, N. (1994) Brain lip-id-binding protein (BLBP): a novel signaling system in the developing mammalian CNS. Neuron, 12, 895-908.
Glatz, J.F., Vork, M.M. & van der Vusse, G.J. (1993) Significance of cytoplasmic fatty acid-binding protein for the ischemic heart. Mol. Cell Bio-chem., 123, 167-173.
Gonzalez-Hernandez, J.A., Bornstein, S.R., Ehrhart-Bornstein, M., Geschwend, J.E., Adler, G. & Scherbaum, W.A. (1994) Macrophages within the human adrenal gland. Cell Tissue. Res., 278, 201-205.
Gordon, J.I., Alpers, D.H., Ockner, R.K. & Strauss, A.W. (1983) The nucleotide sequence of rat liver fatty acid binding protein mRNA. J. Biol. Chem., 258, 3356-3363.
Hamilton, W.J. & Mossman, H.W. (1972) Human Em-bryology, 4 ed., Heffer, Cambridge.
Hume, D.A., Halpin, D., Charlton, H. & Gordon, S. (1984) The mononuclear phagocyte system of the mouse defined by immunohistochemical localization of antigen F4/80: macrophages of endocrine organs. Proc. Natl. Acad. Sci. U S A, 81, 4174-4177.
Hunt, C.R., Ro, J.H., Dobson, D.E., Min, H.Y. & Spiegelman, B.M. (1986) Adipocyte P2 gene: developmental expression and homology of 5′-flanking sequences among fat cell-specific genes. Proc. Natl. Acad. Sci. U S A, 83, 3786-3790.
Iseki, S., Kondo, H., Hitomi, M. & Ono, T. (1988) Im-munocytochemical localization of hepatic fatty acid binding protein in the liver of fed and fast-ed rats. Histochemistry, 89, 317-322.
Kitanaka, N., Owada, Y., Abdelwahab, S.A., Iwasa, H., Sakagami, H., Watanabe, M., Spener, F. & Kondo, H. (2003) Specific localization of epi-dermal-type fatty acid binding protein in den-dritic cells of splenic white pulp. Histochem. Cell Biol., 120, 465-473.
Kurtz, A., Zimmer, A., Schnutgen, F., Bruning, G., Spener, F. & Muller, T. (1994) The expression
pattern of a novel gene encoding brain-fatty acid binding protein correlates with neuronal and glial cell development. Development, 120, 2637-2649.
Leenen, P.J., Radosevic, K., Voerman, J.S., Salomon, B., van Rooijen, N., Klatzmann, D. & van Ewijk, W. (1998) Heterogeneity of mouse spleen dendritic cells: in vivo phagocytic activi-ty, expression of macrophage markers, and sub-population turnover. J. Immunol . , 160 , 2166-2173.
Magalhaes, M.M. & Magalhaes, M.C. (1980) Effects of orchidectomy on the adrenal macrophage system. Anat. Rec., 198, 209-218.
Magalhaes, M.M. & Magalhaes, M.C. (1984) Effects of ovariectomy and estradiol administration on the adrenal macrophage system of the rat. Cell Tissue. Res., 238, 559-564.
Masouye, I., Hagens, G., Van Kuppevelt, T.H., Madsen, P., Saurat, J.H., Veerkamp, J.H., Pepper, M.S. & Siegenthaler, G. (1997) Endothelial cells of the human microvasculature express epidermal fat-ty acid-binding protein. Circ. Res., 81, 297-303.
Mausle, E. (1974) Ultrastructure and function of the mesenchyme of the rat adrenal cortex. Beitr. Pathol., 151, 344-360.
Miller, S.J., Li, H., Rizvi, T.A., Huang, Y., Johansson, G., Bowersock, J., Sidani, A., Vitullo, J., Vogel, K., Parysek, L.M., DeClue, J.E. & Ratner, N. (2003) Brain lipid binding protein in axon-Schwann cell interactions and peripheral nerve tumorigenesis. Mol. Cell Biol., 23, 2213-2224.
Owada, Y., Yoshimoto, T. & Kondo, H. (1996a) Spatio-temporally differential expression of genes for three members of fatty acid binding proteins in developing and mature rat brains. J. Chem. Neuroanat., 12, 113-122.
Owada, Y., Yoshimoto, T. & Kondo, H. (1996b) In-creased expression of the mRNA for brain- and skin-type but not heart-type fatty acid binding proteins following kainic acid systemic admin-istration in the hippocampal glia of adult rats. Brain Res. Mol. Brain Res., 42, 156-160.
Owada, Y., Utsunomiya, A., Yoshimoto, T. & Kondo, H. (1997) Changes in gene expression for skin-type fatty acid binding protein in hypoglossal motor neurons following nerve crush. Neuro-sci. Lett., 223, 25-28.
Owada, Y., Abdelwahab, S.A., Suzuki, R., Iwasa, H., Sakagami, H., Spener, F. & Kondo, H. (2001) Localization of epidermal-type fatty acid bind-

X. Yun et al.86
ing protein in alveolar macrophages and some alveolar type II epithelial cells in mouse lung. Histochem. J., 33, 453-457.
Owada, Y., Suzuki, R., Iwasa, H., Spener, F. & Kondo, H. (2002a) Localization of epidermal-type fatty acid binding protein in the thymic epithelial cells of mice. Histochem. Cell Biol., 117, 55-60.
Owada, Y., Takano, H., Yamanaka, H., Kobayashi, H., Sugitani, Y., Tomioka, Y., Suzuki, I., Suzuki, R., Terui, T., Mizugaki, M., Tagami, H., Noda, T. & Kondo, H. (2002b) Altered water barrier func-tion in epidermal-type fatty acid binding pro-tein-deficient mice. J. Invest. Dermatol., 118,
430-435.Sato, T. (1998) Class II MHC-expressing cells in the
rat adrenal gland defined by monoclonal anti-bodies. Histochem. Cell Biol., 109, 359-367.
Siegenthaler, G., Hotz, R., Chatellard-Gruaz, D., Jaconi, S. & Saurat, J.H. (1993) Characteriza-tion and expression of a novel human fatty acid-binding protein: the epidermal type (E-FABP). Biochem. Biophys. Res. Commun., 190, 482-487.
Watanabe, M., Ono, T. & Kondo, H. (1991) Immuno-histochemical studies on the localisation and ontogeny of heart fatty acid binding protein in the rat. J. Anat., 174, 81-95.
Related Documents











