American Journal of Transplantation 2004; 5: 517–528 Blackwell Munksgaard Copyright C Blackwell Munksgaard 2005 doi: 10.1111/j.1600-6143.2005.00744.x Different Roles for Matrix Metalloproteinase-2 and Matrix Metalloproteinase-9 in the Pathogenesis of Cardiac Allograft Rejection Lacey G. Campbell a , Sabarinathan Ramachandran a , Wei Liu a , J. Michael Shipley b,c , Shigeyoshi Itohara d , Joseph G. Rogers b , Nader Moazami a , Robert M. Senior b,c and Andr ´ es Jaramillo a, ∗ Departments of a Surgery, b Internal Medicine and c Cell Biology & Physiology, Washington University School of Medicine, St. Louis, Missouri, USA and d Laboratory for Behavioral Genetics, Brain Science Institute, Riken, Saitama, Japan ∗ Corresponding author: Andr ´ es Jaramillo, [email protected] Recent studies have shown an increased expression of several matrix metalloproteinases (MMP) during car- diac, renal and pulmonary allograft rejection. To fur- ther define the roles of MMP-2 and MMP-9 in the pathogenesis of cardiac allograft rejection, BALB/c car- diac allografts were transplanted into MMP-2-deficient (−/−) and MMP-9−/− mice. Allografts rejected by wild-type mice revealed a significant increase in MMP- 2 and MMP-9 expression. MMP-2-deficiency signifi- cantly prolonged allograft survival time. Functioning allografts harvested from MMP-2−/− mice showed lower cellular infiltration and fibrosis than rejected al- lografts harvested from MMP-2+/+ mice at the same time. In contrast, MMP-9-deficiency significantly de- creased allograft survival time. Functioning allografts harvested from MMP-9+/+ mice showed lower cellu- lar infiltration and fibrosis than rejected allografts har- vested from MMP-9−/− mice at the same time. MMP- 2−/− recipients showed decreased T-cell alloreactivity mediated by a defect in dendritic cell stimulatory and T-cell responsive capacities. In contrast, MMP-9−/− recipients showed increased T-cell alloreactivity me- diated by a significant increased in dendritic cell stim- ulatory and T-cell responsive capacities. These results indicate that MMP2 and MMP-9 play significantly dif- ferent roles in the process of cardiac allograft rejection. Key words: Allograft rejection, dendritic cells, heart, matrix metalloproteinase-2, matrix metalloproteinase- 9, T-cells Received 19 August 2004, revised 21 October 2004 and accepted for publication 22 October 2004 Introduction Acute allograft rejection is mediated by the coordinated infiltration of T cells into the allograft followed by the ex- pression of several T-cell effector functions that induce the immune-mediated destruction of the allograft (1–3). This is associated with significant alterations in the com- ponents of the extracellular matrix compartment and this turnover of extracellular matrix proteins is mainly regulated by matrix metalloproteinases (MMP) (4–11). MMPs consti- tute a multigene family of over 25 secreted and cell sur- face zinc-dependent endopeptidases capable of degrading essentially all components of the extracellular matrix as well as other proteinases, clotting factors, chemokines, cytokines and adhesion molecules (4–11). MMPs partici- pate in numerous biological processes, such as embryo- genesis, cellular proliferation, tumor metastasis and angio- genesis, wound healing and inflammation (4–11). MMPs in general are secreted in a soluble pro-enzyme form and their activity is regulated through multiple events includ- ing pro-enzyme activation and interaction with tissue in- hibitors of metalloproteinases (4–11). Among the MMP family, MMP-2 and MMP-9 may be especially important in inflammation since both have been shown to play a critical role in the process of T-cell infiltration into tissues (12,13). In this regard, previous studies have shown that both CD4+ and CD8+ T cells produce significant levels of MMP-2 and MMP-9 upon stimulation (12–15). It has also been shown that T cells induce the production of MMP-2 and MMP-9 by macrophages, endothelial cells and vascu- lar smooth muscle cells through direct cell-cell interaction or through the production of inflammatory cytokines (16– 18). Related studies have shown that MMP-2 and MMP-9 are involved in immunoregulation by degrading biologically active molecules such as cytokines and chemokines (8– 10,19–22). On the other hand, MMP-2 and MMP-9 have been shown to be involved in the activation of several cy- tokines and chemokines (4–11). Recent studies in animal models (23–26) and humans (27,28) have shown an increased expression of both MMP- 2 and MMP-9 in association with acute rejection of car- diac and renal allografts. Other studies have also shown an association between increased expression levels of MMP-2 and MMP-9 with chronic rejection of pulmonary allografts in humans (29,30). Moreover, it has also been 517

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

American Journal of Transplantation 2004; 5: 517–528Blackwell Munksgaard
Copyright C© Blackwell Munksgaard 2005
doi: 10.1111/j.1600-6143.2005.00744.x
Different Roles for Matrix Metalloproteinase-2and Matrix Metalloproteinase-9 in the Pathogenesisof Cardiac Allograft Rejection
Lacey G. Campbella, Sabarinathan
Ramachandrana, Wei Liua, J. Michael Shipleyb,c,
Shigeyoshi Itoharad, Joseph G. Rogersb,
Nader Moazamia, Robert M. Seniorb,c
and Andres Jaramilloa,∗
Departments of aSurgery, bInternal Medicine andcCell Biology & Physiology, Washington University Schoolof Medicine, St. Louis, Missouri, USA anddLaboratory for Behavioral Genetics, Brain ScienceInstitute, Riken, Saitama, Japan∗
Corresponding author: Andres Jaramillo,[email protected]
Recent studies have shown an increased expression ofseveral matrix metalloproteinases (MMP) during car-diac, renal and pulmonary allograft rejection. To fur-ther define the roles of MMP-2 and MMP-9 in thepathogenesis of cardiac allograft rejection, BALB/c car-diac allografts were transplanted into MMP-2-deficient(−/−) and MMP-9−/− mice. Allografts rejected bywild-type mice revealed a significant increase in MMP-2 and MMP-9 expression. MMP-2-deficiency signifi-cantly prolonged allograft survival time. Functioningallografts harvested from MMP-2−/− mice showedlower cellular infiltration and fibrosis than rejected al-lografts harvested from MMP-2+/+ mice at the sametime. In contrast, MMP-9-deficiency significantly de-creased allograft survival time. Functioning allograftsharvested from MMP-9+/+ mice showed lower cellu-lar infiltration and fibrosis than rejected allografts har-vested from MMP-9−/− mice at the same time. MMP-2−/− recipients showed decreased T-cell alloreactivitymediated by a defect in dendritic cell stimulatory andT-cell responsive capacities. In contrast, MMP-9−/−recipients showed increased T-cell alloreactivity me-diated by a significant increased in dendritic cell stim-ulatory and T-cell responsive capacities. These resultsindicate that MMP2 and MMP-9 play significantly dif-ferent roles in the process of cardiac allograft rejection.
Key words: Allograft rejection, dendritic cells, heart,matrix metalloproteinase-2, matrix metalloproteinase-9, T-cells
Received 19 August 2004, revised 21 October 2004 andaccepted for publication 22 October 2004
Introduction
Acute allograft rejection is mediated by the coordinatedinfiltration of T cells into the allograft followed by the ex-pression of several T-cell effector functions that inducethe immune-mediated destruction of the allograft (1–3).This is associated with significant alterations in the com-ponents of the extracellular matrix compartment and thisturnover of extracellular matrix proteins is mainly regulatedby matrix metalloproteinases (MMP) (4–11). MMPs consti-tute a multigene family of over 25 secreted and cell sur-face zinc-dependent endopeptidases capable of degradingessentially all components of the extracellular matrix aswell as other proteinases, clotting factors, chemokines,cytokines and adhesion molecules (4–11). MMPs partici-pate in numerous biological processes, such as embryo-genesis, cellular proliferation, tumor metastasis and angio-genesis, wound healing and inflammation (4–11). MMPsin general are secreted in a soluble pro-enzyme form andtheir activity is regulated through multiple events includ-ing pro-enzyme activation and interaction with tissue in-hibitors of metalloproteinases (4–11). Among the MMPfamily, MMP-2 and MMP-9 may be especially importantin inflammation since both have been shown to play acritical role in the process of T-cell infiltration into tissues(12,13). In this regard, previous studies have shown thatboth CD4+ and CD8+ T cells produce significant levels ofMMP-2 and MMP-9 upon stimulation (12–15). It has alsobeen shown that T cells induce the production of MMP-2and MMP-9 by macrophages, endothelial cells and vascu-lar smooth muscle cells through direct cell-cell interactionor through the production of inflammatory cytokines (16–18). Related studies have shown that MMP-2 and MMP-9are involved in immunoregulation by degrading biologicallyactive molecules such as cytokines and chemokines (8–10,19–22). On the other hand, MMP-2 and MMP-9 havebeen shown to be involved in the activation of several cy-tokines and chemokines (4–11).
Recent studies in animal models (23–26) and humans(27,28) have shown an increased expression of both MMP-2 and MMP-9 in association with acute rejection of car-diac and renal allografts. Other studies have also shownan association between increased expression levels ofMMP-2 and MMP-9 with chronic rejection of pulmonaryallografts in humans (29,30). Moreover, it has also been
517

Campbell et al.
demonstrated that MMPs are involved in several car-diac pathologies including acute myocardial infarction,ischemic-dilated cardiomyopathy and congestive heart fail-ure (31–33). Overall, these observations indicate that theseenzymes may play a crucial role in the pathogenesis ofcardiac allograft rejection. However, as mentioned above,both these enzymes have the ability to suppress the in-flammatory response by means of their enzymatic activityon cytokines and chemokines (4–11,19–22). Therefore, thepurpose of this study was to determine the specific rolesof MMP-2 and MMP-9 in the process of cardiac allograftrejection.
Materials and Methods
Animals
MMP-2−/− and MMP-9−/− mice were originally developed by homolo-gous recombination on C57BL/6 (H-2b) and 129SvEv (H-2b) genetic back-grounds, respectively (34,35). Wild-type BALB/c (H-2d), C3H/He (H-2k) andC57BL/6 (MMP-2+/+) mice were obtained from The Jackson Laboratories(Bar Harbor, ME). Wild-type 129/SvEv (MMP-9+/+) mice were obtainedfrom Taconic Farms (Germantown, NY). All mice were housed in a pathogen-free facility and all experimental procedures were performed in adult males8–12 weeks of age. All animal procedures were performed according to theguidelines of the animal care committee of Washington University Schoolof Medicine.
Heterotopic cardiac transplantation
Cardiac grafts were transplanted heterotopically using the method of Corryet al. (36). Briefly, donor and recipient mice were anesthetized with phe-nobarbital. The donor hearts were harvested and placed in chilled, lactatedRinger’s solution while the recipient was prepared. Then, the donor heartwas anastomosed to the recipient abdominal aorta and inferior vena cavautilizing 10-0 sutures. Upon completion of the anastomosis and organ per-fusion, the heart grafts resumed spontaneous contractions. Cold ischemictime was always less than 30 min during recipient preparation. The strengthand quality of the cardiac graft impulses were examined each day by palpa-tion of the recipient abdomen. Rejection of cardiac grafts was defined bycessation of a palpable impulse and was confirmed histologically by H&Eand Masson’s trichrome stains.
Substrate zymography
Heart grafts were harvested, homogenized in PBS containing 0.1% BRIJ-35(Sigma, St. Louis, MO) and a protease inhibitor cocktail (Roche Diagnostics,Mannheim, Germany) and centrifuged at 15 000 rpm for 10 min at 4
◦C.
The protein concentration of the supernatants was determined using theBCA protein assay (Pierce, Rockford, IL), and 15 lg of supernatant proteinswere resolved by non-reducing 10% SDS-PAGE through Novex Tris-Glycinegels containing 0.1% gelatin (Invitrogen, Carlsbad, CA). The gels were thenwashed for 30 min in Novex zymogram denaturing buffer (Invitrogen) toremove the SDS, and then incubated in Novex zymogram developing buffer(Invitrogen) for 24 h at 37
◦C. Areas of protease activity were visualized after
staining the gels with 0.5% Coomassie blue R-250 (Invitrogen).
RT-PCR
Heart grafts were harvested, homogenized with TRIzol reagent (LifeTechnologies, Gaithersburg, MD) and total RNA was isolated accordingto the manufacturer’s instructions. cDNA was synthesized by extensionof primers using Powerscript reverse transcriptase (Clontech, Palo Alto,CA) according to manufacturer’s instructions. PCR was then performed
using Taq DNA polymerase and buffer mix (Invitrogen) in the presence ofoligonucleotide primers for MMP-2, MMP-9 and GAPDH (control) genes.MMP-2 primer sequences were 5′-CTTTGCAGGAGACAAGTTCTGG-3′
and 5′-TTAAGGTGGTGCAGGTATCTGG-3′ (701 bp product); MMP-9primer sequences were 5′-TGGCAGAGGCATACTTGTAC-3′ and 5′-AACTGGATGACAATGTCCGC-3′ (375-bp product); GAPDH primersequences were 5′-CCATGTTTGTGATGGGTGTGAACC-3′ and 5′-TGTGAGGGAGATGCTCAGTGTTGG-3′ (712 bp product). The PCR cyclingconditions for MMP-2 were as follows: initial denaturation at 94
◦C for
5 min, followed by 40 cycles of 94◦C for 20 s, 51
◦C for 30 s and 72
◦C
for 1 min and a final extension step at 72◦C for 10 min. The PCR cycling
conditions for MMP-9 were as follows: initial denaturation at 94◦C for
5 min, followed by 40 cycles of 94◦C for 20 s, 55
◦C for 30 s and 72
◦C
for 1 min and a final extension step at 72◦C for 10 min. The PCR cycling
conditions for GAPDH were as follows: initial denaturation at 94◦C for 1
min, followed by 30 cycles of 94◦C for 20 s, 58
◦C for 1 min and 72
◦C for
1 min and a final extension step at 72◦C for 10 min. Amplified cDNA was
electrophoresed on 2% agarose gels, stained with ethidium bromide andvisualized using an UV trans-illuminator.
Quantitative real-time PCR
Quantification of MMP-2 and MMP-9 gene expression was further an-alyzed by quantitative real-time PCR using the 2−��CT method (37).Briefly, heart grafts were harvested and total RNA was extracted usingan RNAqueous-4PCR kit (Ambion, Austin, TX) and reverse transcribedusing PowerScript reverse transcriptase (Clontech). cDNA from 20 ngof RNA was quantitatively analyzed using a SYBRO Green PCR Mas-ter Mix kit (Applied Biosystems, Foster City, CA) in the presence ofoligonucleotide primers for MMP-2, MMP-9 and COX-II (control) genes(gene bank accession numbers 008610, 013599 and 13838, respec-tively). MMP-2 primer sequences were 5′-TTTTGCTCGGGCCTTAAAAG-3′ (forward) and 5′-ATGGATTCGAGAAAAGCGCA-3′ (reverse); MMP-9 primer sequences were 5′-GTACACAGGCAAGACCGTGCT-3′ (for-ward) and 5′-CGCTGACGTGGGTTACCTCT-3′ (reverse); COX-II primersequences were 5′-CATCCCAGGCCGACTAAATC-3′ (forward) and 5′-TTTCAGAGCATTGGCCATAGAA-3′ (reverse). Gene-specific PCR productswere continuously measured with an ABI PRISM 5700 Sequence Detec-tion System (Applied Biosystems) during 40 cycles of PCR.
Collagen content determination
Heart grafts were harvested, homogenized in PBS containing 0.1% BRIJ-35 (Sigma) and a protease inhibitor cocktail (Roche Diagnostics) and cen-trifuged at 15 000 rpm for 10 min at 4
◦C. The protein concentration of the
supernatants was determined using the BCA protein assay (Pierce). The sol-uble collagen content included both denatured and un-denatured collagenwith the latter measured by means of hydroxyproline quantification usingthe Sircol collagen assay (Accurate Chemicals, Westbury, NY) following themanufacturer’s instructions.
Cytokine and chemokine production assay
Spleen cells from MMP-2+/+ and MMP-2−/− recipients were harvested at7 days after transplantation while spleen cells from MMP-9+/+ and MMP-9−/− recipients were harvested at 9 days after transplantation. The cellswere then treated with erythrocyte lysis buffer (0.1-mM EDTA, 155-mMammonium chloride, 10-mM potassium bicarbonate) for 3 min, washed(3×) and resuspended in RPMI-1640 medium (Gibco, Grand Island, NY)supplemented with FBS (10%, Hyclone, Logan, UT), L-glutamine (2 mM),non-essential amino acids (100 lM), HEPES buffer (25 mM), sodium pyru-vate (1 mM), penicillin (100 U/mL), streptomycin (100 lg/mL) and 2-ME (50lM). Then, the cells were plated in triplicate (3 × 105/well) in 96-well flat-bottom plates. BALB/c (allograft donor) and C57BL6 (syngeneic control for
518 American Journal of Transplantation 2004; 5: 517–528

MMPs in Cardiac Allograft Rejection
MMP-2+/+ and MMP-2−/− recipients) or 129/SvEv mice (syngeneic con-trol for MMP-9+/+ and MMP-9−/− recipients) spleen cells were irradiated(3000 rads) and plated (6 × 105/well) in the wells containing responder cells.After 72 h, the culture supernatants were collected and analyzed by meansof the TranSignal cytokine/chemokine antibody array (Panomics, RedwoodCity, CA) following the manufacturer’s instructions. The cytokine blots wereanalyzed using the Quantity One software (BioRad, Hercules, CA). Resultsare expressed as densitometry units.
ELISPOT assay
MultiScreen 96-well filtration plates (Millipore, Bedford, MA) were coatedfor 2 h at 37
◦C with an IFN-c -specific mAb (5.0 lg/mL, BD Pharmingen,
San Diego, CA) in 0.05-M carbonate–bicarbonate buffer (pH = 9.6). Next,the plates were blocked with 1% BSA for 1 h at 37
◦C and washed with
PBS (3×). Spleen cells from MMP-2+/+ and MMP-2−/− recipients wereharvested at 7 days after transplantation while spleen cells from MMP-9+/+and MMP-9−/− recipients were harvested at 9 days after transplantation.The cells were plated in quadruplicate (3 × 105/well) in the mAb-coatedplates. BALB/c (allograft donor), C3H/He (3rd party control) and C57BL/6(syngeneic control for MMP-2+/+ and MMP-2−/− recipients) or 129/SvEv(syngeneic control for MMP-9+/+ and MMP-9−/− recipients) spleen cellswere irradiated (3000 rads) and plated (6 × 105/well) in the wells containingresponder cells. After 48 h, the plates were washed with PBS (3×) andPBS supplemented with 0.5% Tween-20 (3×). Then, a biotinylated IFN-c -specific mAb (2.0 lg/mL, BD Pharmingen) diluted in PBS/BSA/Tween-20was added to the wells. After 2 h at room temperature, the plates werewashed with PBS/Tween-20 (3×) and HRP-labeled streptavidin (1:100, BDPharmingen) diluted in PBS/BSA/Tween-20 was added to the wells. After2 h at room temperature, the plates were washed with PBS/Tween-20 (3×)and PBS (3×). Then, AEC Substrate Reagent (BD Pharmingen) was addedto the wells for 2–5 min. The plates were washed with tap water to stopthe reaction and air-dried. Spots were analyzed in an ImmunoSpot Series Ianalyzer (Cellular Technology, Cleveland, OH) that was designed to detectELISA spots with pre-determined criteria for spot size, shape and colorimet-ric density.
Dendritic cell preparation
Mature dendritic cells (DC) were purified as previously described (38).Briefly, spleens were injected with collagenase D (100 U/mL, BoehringerMannheim, Indianapolis, IN), teased and passed through a cell strainer (poresize: 40 lm). The remaining tissue fragments were then treated with col-lagenase D (400 U/mL) at 37
◦C. After 45–60 min, the spleen fragments
were passed trough a cell strainer (pore size: 40 lm) and pooled with thesingle cell suspension collected before. Then, the cells were centrifuged, in-cubated with anti-mouse CD11c-coated magnetic beads (Miltenyi Biotech,Auburn, CA) and positively selected with a MACS separation column fol-lowing the manufacturer’s instructions. Isolated DCs were then allowed toundergo maturation by cultivation in culture medium for 24 h. Purified DCswere >98% CD11c+ and I-A+ as determined by FACS analysis (data notshown).
T-cell preparation
Spleens were teased and passed through a cell strainer (pore size: 40 lm).Then, the T cells were negatively selected using the pan-T-cell isolation kit(Miltenyi Biotech) following the manufacturer’s instructions. Purified T cellswere >98% CD3+ as determined by FACS analysis (data not shown).
APC stimulatory capacity assay
MMP-2+/+ and MMP-2−/− DCs or MMP-9+/+ and MMP-9−/− DCs wereirradiated (3000 rads) and plated in triplicate at varying concentrations(3–24 × 103/well) in anti-IFN-c -coated ELISPOT plates. Then, BALB/c Tcells were plated (3 × 105/well) in the wells containing DCs and used as
responder cells. After 48 h, the frequency of IFN-c -producing alloreactiveT cells was determined as described above. BALB/c T cells cultured in thepresence of syngeneic BALB/c DCs were used as negative controls.
T-cell responsive capacity assay
BALB/c DCs were irradiated (3000 rads) and plated in triplicate at vary-ing concentrations (3–24 × 103/well) in anti-IFN-c -coated ELISPOT plates.Then, MMP-2+/+ and MMP-2−/− T cells or MMP-9+/+ and MMP-9−/−T cells were plated (3 × 105/well) in the wells containing DCs and used asresponder cells. After 48 h, the frequency of IFN-c -producing alloreactiveT cells was determined as described above. MMP-2+/+ and MMP-2−/−T cells cultured in the presence of syngeneic C57BL/6 DCs or MMP-9+/+and MMP-9−/− T cells cultured in the presence of syngeneic 129/SvEv DCswere used as negative controls in the corresponding experiments.
Statistical analysis
Differences in graft survival curves were assessed by means of a 2-way log-rank (Mantel–Haenszel) test, differences in alloreactive T-cell frequencies incardiac allograft recipients and the collagen content of the allografts wereassessed by means of a 2-tailed Student’s t-test, and differences in T-cellresponses against allogeneic DCs in naıve mice were assessed by meansof a 2-way ANOVA test using the Prism 4.0 Statistical Program (GraphPadSofware, San Diego, CA). The a was set at p < 0.05 in all analyses.
Results
Increased MMP-2 and MMP-9 expression during
cardiac allograft rejection
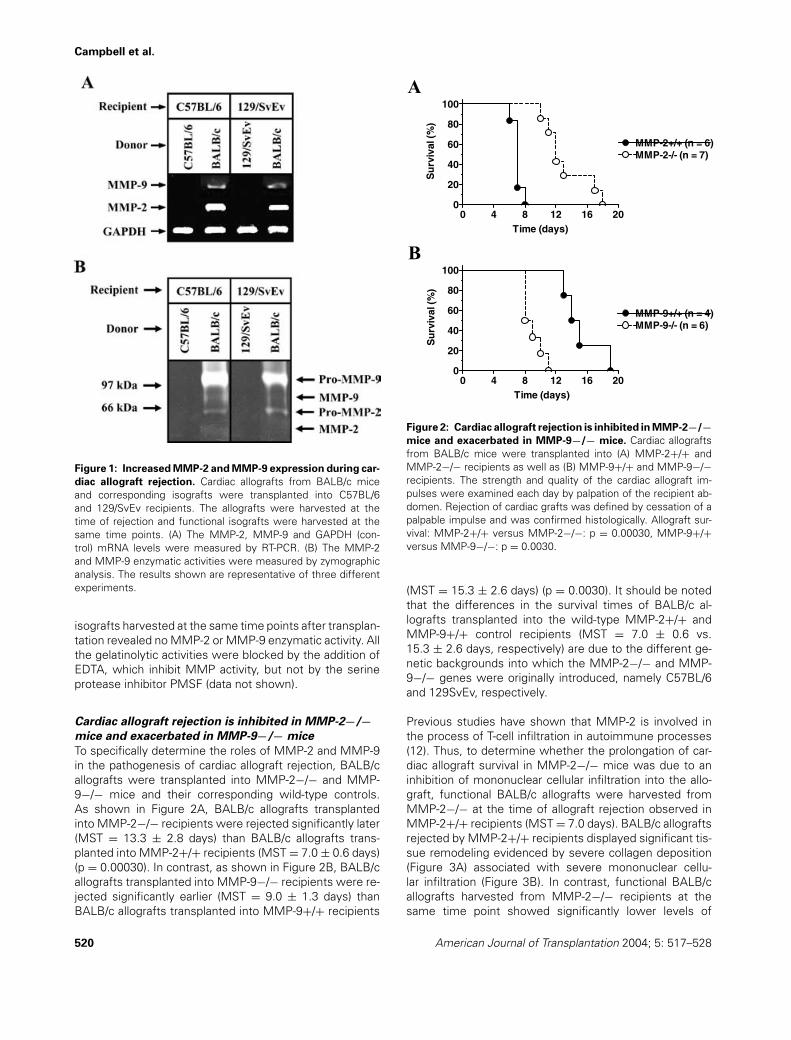
To determine whether MMP-2 and/or MMP-9 played a rolein the pathogenesis of cardiac allograft rejection, we trans-planted BALB/c allografts into C57BL/6 and 129/SvEv re-cipients. Then, the MMP-2 and MMP-9 mRNA levels inallografts harvested at the time of organ failure were de-termined by means of RT-PCR. As shown in Figure 1A, re-jected allografts revealed the presence of both MMP-2 andMMP-9 mRNAs. In contrast, the corresponding functioningisografts harvested at the same time points after transplan-tation revealed no MMP-2 or MMP-9 mRNA transcription.Further analysis of relative gene expression by real-timePCR of rejected allografts harvested from C57BL/6 recipi-ents showed that the levels of MMP-2 and MMP-9 mRNAwere 20.2-fold and 6.6-fold higher than the one observedin isografts, respectively (data not shown).
To determine the expression of MMP-2 and MMP-9 atthe protein level, we measured the enzymatic activity ofMMP-2 and MMP-9 in BALB/c allografts harvested fromC57BL/6 and 129/SvEv recipients at the time of organfailure by means of zymographic analysis. As shown inFigure 1B, rejected allografts revealed high levels of bothMMP-2 and MMP-9 enzymatic activity. Two prominentgelatinolytic bands at approximately 105 kDa and 64 kDawere present in the rejected allografts corresponding tothe pro-MMP-9 and pro-MMP-2 zymogens, respectively.Two less prominent gelatinolytic bands at approximately 94kDa and 60 kDa were also present in the rejected allograftscorresponding to the active forms of MMP-9 and MMP-2,respectively. In contrast, tissue extracts from functioning
American Journal of Transplantation 2004; 5: 517–528 519
Campbell et al.
Figure 1: Increased MMP-2 and MMP-9 expression during car-
diac allograft rejection. Cardiac allografts from BALB/c miceand corresponding isografts were transplanted into C57BL/6and 129/SvEv recipients. The allografts were harvested at thetime of rejection and functional isografts were harvested at thesame time points. (A) The MMP-2, MMP-9 and GAPDH (con-trol) mRNA levels were measured by RT-PCR. (B) The MMP-2and MMP-9 enzymatic activities were measured by zymographicanalysis. The results shown are representative of three differentexperiments.
isografts harvested at the same time points after transplan-tation revealed no MMP-2 or MMP-9 enzymatic activity. Allthe gelatinolytic activities were blocked by the addition ofEDTA, which inhibit MMP activity, but not by the serineprotease inhibitor PMSF (data not shown).
Cardiac allograft rejection is inhibited in MMP-2−/−mice and exacerbated in MMP-9−/− mice
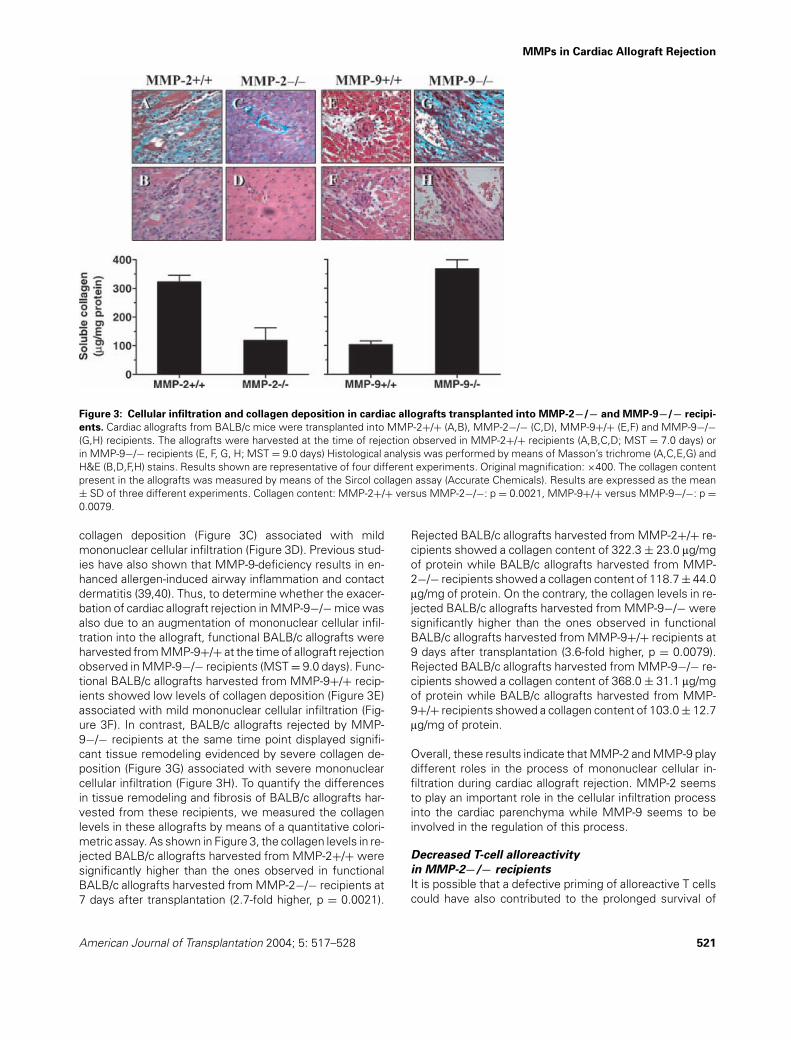
To specifically determine the roles of MMP-2 and MMP-9in the pathogenesis of cardiac allograft rejection, BALB/callografts were transplanted into MMP-2−/− and MMP-9−/− mice and their corresponding wild-type controls.As shown in Figure 2A, BALB/c allografts transplantedinto MMP-2−/− recipients were rejected significantly later(MST = 13.3 ± 2.8 days) than BALB/c allografts trans-planted into MMP-2+/+ recipients (MST = 7.0 ± 0.6 days)(p = 0.00030). In contrast, as shown in Figure 2B, BALB/callografts transplanted into MMP-9−/− recipients were re-jected significantly earlier (MST = 9.0 ± 1.3 days) thanBALB/c allografts transplanted into MMP-9+/+ recipients
0 4 8 12 16 200
20
40
60
80
100
MMP-2+/+ (n = 6)MMP-2-/- (n = 7)
Time (days)
Su
rviv
al (
%)
0 4 8 12 16 200
20
40
60
80
100
MMP-9+/+ (n = 4)MMP-9-/- (n = 6)
Time (days)
Su
rviv
al (
%)
A
B
Figure 2: Cardiac allograft rejection is inhibited in MMP-2−/−mice and exacerbated in MMP-9−/− mice. Cardiac allograftsfrom BALB/c mice were transplanted into (A) MMP-2+/+ andMMP-2−/− recipients as well as (B) MMP-9+/+ and MMP-9−/−recipients. The strength and quality of the cardiac allograft im-pulses were examined each day by palpation of the recipient ab-domen. Rejection of cardiac grafts was defined by cessation of apalpable impulse and was confirmed histologically. Allograft sur-vival: MMP-2+/+ versus MMP-2−/−: p = 0.00030, MMP-9+/+versus MMP-9−/−: p = 0.0030.
(MST = 15.3 ± 2.6 days) (p = 0.0030). It should be notedthat the differences in the survival times of BALB/c al-lografts transplanted into the wild-type MMP-2+/+ andMMP-9+/+ control recipients (MST = 7.0 ± 0.6 vs.15.3 ± 2.6 days, respectively) are due to the different ge-netic backgrounds into which the MMP-2−/− and MMP-9−/− genes were originally introduced, namely C57BL/6and 129SvEv, respectively.
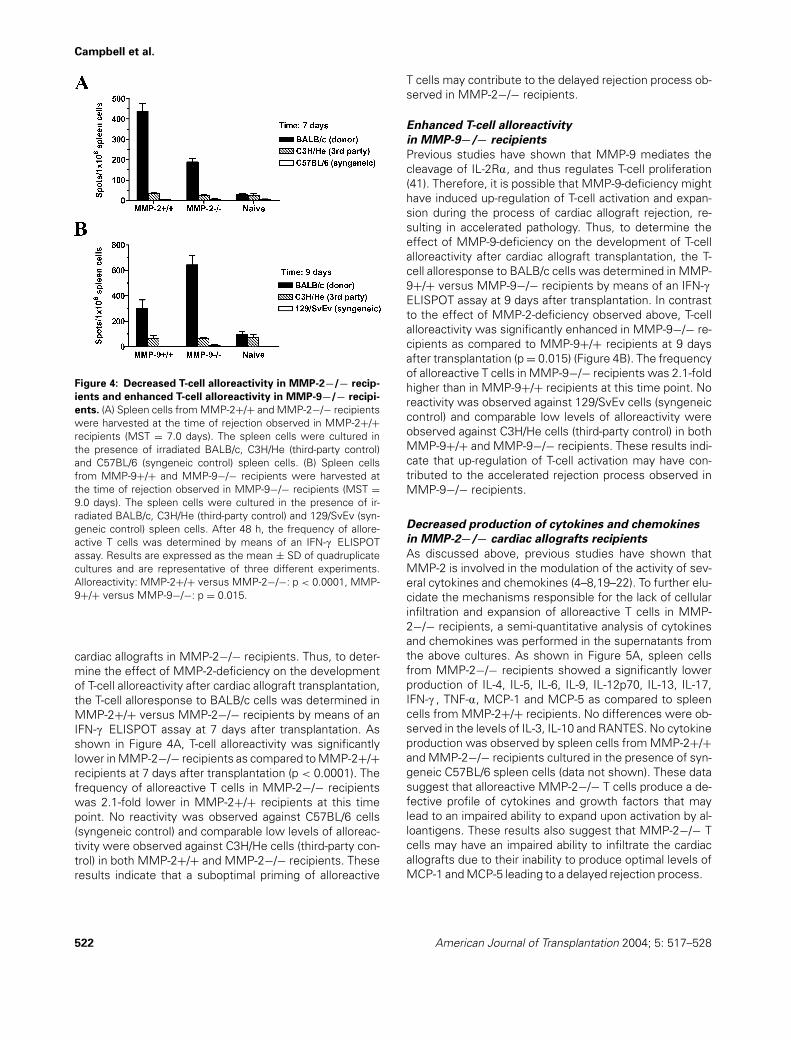
Previous studies have shown that MMP-2 is involved inthe process of T-cell infiltration in autoimmune processes(12). Thus, to determine whether the prolongation of car-diac allograft survival in MMP-2−/− mice was due to aninhibition of mononuclear cellular infiltration into the allo-graft, functional BALB/c allografts were harvested fromMMP-2−/− at the time of allograft rejection observed inMMP-2+/+ recipients (MST = 7.0 days). BALB/c allograftsrejected by MMP-2+/+ recipients displayed significant tis-sue remodeling evidenced by severe collagen deposition(Figure 3A) associated with severe mononuclear cellu-lar infiltration (Figure 3B). In contrast, functional BALB/callografts harvested from MMP-2−/− recipients at thesame time point showed significantly lower levels of
520 American Journal of Transplantation 2004; 5: 517–528
MMPs in Cardiac Allograft Rejection
Figure 3: Cellular infiltration and collagen deposition in cardiac allografts transplanted into MMP-2−/− and MMP-9−/− recipi-
ents. Cardiac allografts from BALB/c mice were transplanted into MMP-2+/+ (A,B), MMP-2−/− (C,D), MMP-9+/+ (E,F) and MMP-9−/−(G,H) recipients. The allografts were harvested at the time of rejection observed in MMP-2+/+ recipients (A,B,C,D; MST = 7.0 days) orin MMP-9−/− recipients (E, F, G, H; MST = 9.0 days) Histological analysis was performed by means of Masson’s trichrome (A,C,E,G) andH&E (B,D,F,H) stains. Results shown are representative of four different experiments. Original magnification: ×400. The collagen contentpresent in the allografts was measured by means of the Sircol collagen assay (Accurate Chemicals). Results are expressed as the mean± SD of three different experiments. Collagen content: MMP-2+/+ versus MMP-2−/−: p = 0.0021, MMP-9+/+ versus MMP-9−/−: p =0.0079.
collagen deposition (Figure 3C) associated with mildmononuclear cellular infiltration (Figure 3D). Previous stud-ies have also shown that MMP-9-deficiency results in en-hanced allergen-induced airway inflammation and contactdermatitis (39,40). Thus, to determine whether the exacer-bation of cardiac allograft rejection in MMP-9−/− mice wasalso due to an augmentation of mononuclear cellular infil-tration into the allograft, functional BALB/c allografts wereharvested from MMP-9+/+ at the time of allograft rejectionobserved in MMP-9−/− recipients (MST = 9.0 days). Func-tional BALB/c allografts harvested from MMP-9+/+ recip-ients showed low levels of collagen deposition (Figure 3E)associated with mild mononuclear cellular infiltration (Fig-ure 3F). In contrast, BALB/c allografts rejected by MMP-9−/− recipients at the same time point displayed signifi-cant tissue remodeling evidenced by severe collagen de-position (Figure 3G) associated with severe mononuclearcellular infiltration (Figure 3H). To quantify the differencesin tissue remodeling and fibrosis of BALB/c allografts har-vested from these recipients, we measured the collagenlevels in these allografts by means of a quantitative colori-metric assay. As shown in Figure 3, the collagen levels in re-jected BALB/c allografts harvested from MMP-2+/+ weresignificantly higher than the ones observed in functionalBALB/c allografts harvested from MMP-2−/− recipients at7 days after transplantation (2.7-fold higher, p = 0.0021).
Rejected BALB/c allografts harvested from MMP-2+/+ re-cipients showed a collagen content of 322.3 ± 23.0 lg/mgof protein while BALB/c allografts harvested from MMP-2−/− recipients showed a collagen content of 118.7 ± 44.0lg/mg of protein. On the contrary, the collagen levels in re-jected BALB/c allografts harvested from MMP-9−/− weresignificantly higher than the ones observed in functionalBALB/c allografts harvested from MMP-9+/+ recipients at9 days after transplantation (3.6-fold higher, p = 0.0079).Rejected BALB/c allografts harvested from MMP-9−/− re-cipients showed a collagen content of 368.0 ± 31.1 lg/mgof protein while BALB/c allografts harvested from MMP-9+/+ recipients showed a collagen content of 103.0 ± 12.7lg/mg of protein.
Overall, these results indicate that MMP-2 and MMP-9 playdifferent roles in the process of mononuclear cellular in-filtration during cardiac allograft rejection. MMP-2 seemsto play an important role in the cellular infiltration processinto the cardiac parenchyma while MMP-9 seems to beinvolved in the regulation of this process.
Decreased T-cell alloreactivity
in MMP-2−/− recipients
It is possible that a defective priming of alloreactive T cellscould have also contributed to the prolonged survival of
American Journal of Transplantation 2004; 5: 517–528 521
Campbell et al.
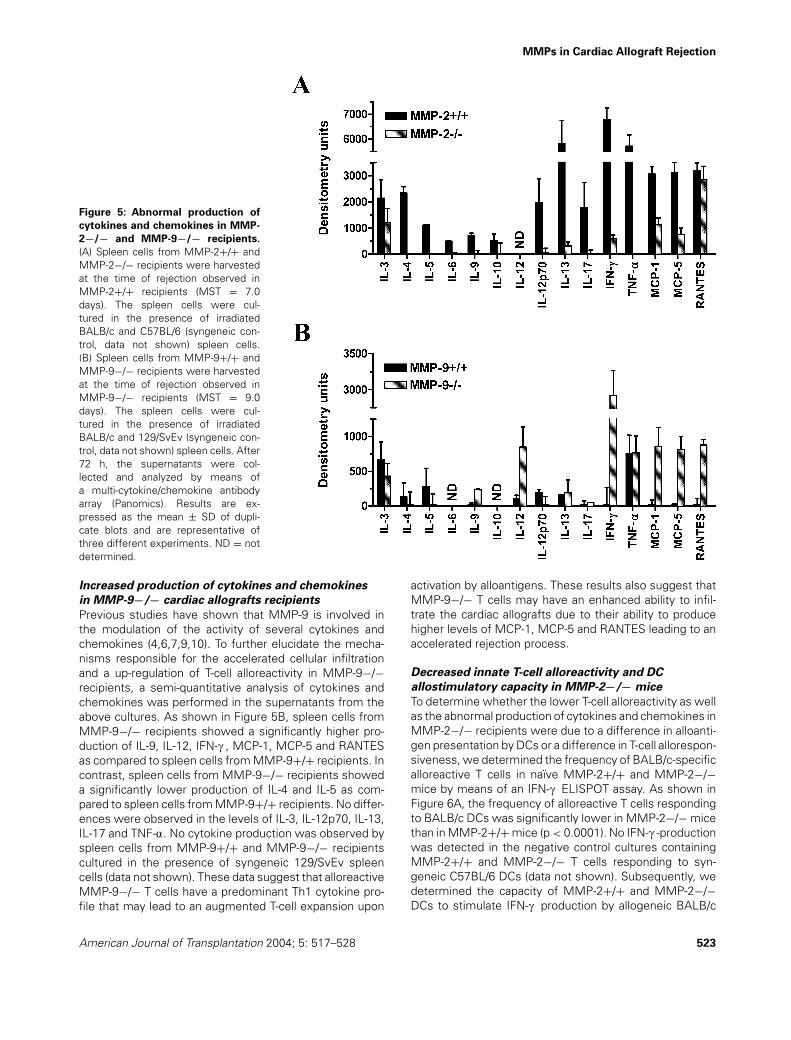
Figure 4: Decreased T-cell alloreactivity in MMP-2−/− recip-
ients and enhanced T-cell alloreactivity in MMP-9−/− recipi-
ents. (A) Spleen cells from MMP-2+/+ and MMP-2−/− recipientswere harvested at the time of rejection observed in MMP-2+/+recipients (MST = 7.0 days). The spleen cells were cultured inthe presence of irradiated BALB/c, C3H/He (third-party control)and C57BL/6 (syngeneic control) spleen cells. (B) Spleen cellsfrom MMP-9+/+ and MMP-9−/− recipients were harvested atthe time of rejection observed in MMP-9−/− recipients (MST =9.0 days). The spleen cells were cultured in the presence of ir-radiated BALB/c, C3H/He (third-party control) and 129/SvEv (syn-geneic control) spleen cells. After 48 h, the frequency of allore-active T cells was determined by means of an IFN-c ELISPOTassay. Results are expressed as the mean ± SD of quadruplicatecultures and are representative of three different experiments.Alloreactivity: MMP-2+/+ versus MMP-2−/−: p < 0.0001, MMP-9+/+ versus MMP-9−/−: p = 0.015.
cardiac allografts in MMP-2−/− recipients. Thus, to deter-mine the effect of MMP-2-deficiency on the developmentof T-cell alloreactivity after cardiac allograft transplantation,the T-cell alloresponse to BALB/c cells was determined inMMP-2+/+ versus MMP-2−/− recipients by means of anIFN-c ELISPOT assay at 7 days after transplantation. Asshown in Figure 4A, T-cell alloreactivity was significantlylower in MMP-2−/− recipients as compared to MMP-2+/+recipients at 7 days after transplantation (p < 0.0001). Thefrequency of alloreactive T cells in MMP-2−/− recipientswas 2.1-fold lower in MMP-2+/+ recipients at this timepoint. No reactivity was observed against C57BL/6 cells(syngeneic control) and comparable low levels of alloreac-tivity were observed against C3H/He cells (third-party con-trol) in both MMP-2+/+ and MMP-2−/− recipients. Theseresults indicate that a suboptimal priming of alloreactive
T cells may contribute to the delayed rejection process ob-served in MMP-2−/− recipients.
Enhanced T-cell alloreactivity
in MMP-9−/− recipients
Previous studies have shown that MMP-9 mediates thecleavage of IL-2Ra, and thus regulates T-cell proliferation(41). Therefore, it is possible that MMP-9-deficiency mighthave induced up-regulation of T-cell activation and expan-sion during the process of cardiac allograft rejection, re-sulting in accelerated pathology. Thus, to determine theeffect of MMP-9-deficiency on the development of T-cellalloreactivity after cardiac allograft transplantation, the T-cell alloresponse to BALB/c cells was determined in MMP-9+/+ versus MMP-9−/− recipients by means of an IFN-cELISPOT assay at 9 days after transplantation. In contrastto the effect of MMP-2-deficiency observed above, T-cellalloreactivity was significantly enhanced in MMP-9−/− re-cipients as compared to MMP-9+/+ recipients at 9 daysafter transplantation (p = 0.015) (Figure 4B). The frequencyof alloreactive T cells in MMP-9−/− recipients was 2.1-foldhigher than in MMP-9+/+ recipients at this time point. Noreactivity was observed against 129/SvEv cells (syngeneiccontrol) and comparable low levels of alloreactivity wereobserved against C3H/He cells (third-party control) in bothMMP-9+/+ and MMP-9−/− recipients. These results indi-cate that up-regulation of T-cell activation may have con-tributed to the accelerated rejection process observed inMMP-9−/− recipients.
Decreased production of cytokines and chemokines
in MMP-2−/− cardiac allografts recipients
As discussed above, previous studies have shown thatMMP-2 is involved in the modulation of the activity of sev-eral cytokines and chemokines (4–8,19–22). To further elu-cidate the mechanisms responsible for the lack of cellularinfiltration and expansion of alloreactive T cells in MMP-2−/− recipients, a semi-quantitative analysis of cytokinesand chemokines was performed in the supernatants fromthe above cultures. As shown in Figure 5A, spleen cellsfrom MMP-2−/− recipients showed a significantly lowerproduction of IL-4, IL-5, IL-6, IL-9, IL-12p70, IL-13, IL-17,IFN-c , TNF-a, MCP-1 and MCP-5 as compared to spleencells from MMP-2+/+ recipients. No differences were ob-served in the levels of IL-3, IL-10 and RANTES. No cytokineproduction was observed by spleen cells from MMP-2+/+and MMP-2−/− recipients cultured in the presence of syn-geneic C57BL/6 spleen cells (data not shown). These datasuggest that alloreactive MMP-2−/− T cells produce a de-fective profile of cytokines and growth factors that maylead to an impaired ability to expand upon activation by al-loantigens. These results also suggest that MMP-2−/− Tcells may have an impaired ability to infiltrate the cardiacallografts due to their inability to produce optimal levels ofMCP-1 and MCP-5 leading to a delayed rejection process.
522 American Journal of Transplantation 2004; 5: 517–528
MMPs in Cardiac Allograft Rejection
Figure 5: Abnormal production of
cytokines and chemokines in MMP-
2−/− and MMP-9−/− recipients.
(A) Spleen cells from MMP-2+/+ andMMP-2−/− recipients were harvestedat the time of rejection observed inMMP-2+/+ recipients (MST = 7.0days). The spleen cells were cul-tured in the presence of irradiatedBALB/c and C57BL/6 (syngeneic con-trol, data not shown) spleen cells.(B) Spleen cells from MMP-9+/+ andMMP-9−/− recipients were harvestedat the time of rejection observed inMMP-9−/− recipients (MST = 9.0days). The spleen cells were cul-tured in the presence of irradiatedBALB/c and 129/SvEv (syngeneic con-trol, data not shown) spleen cells. After72 h, the supernatants were col-lected and analyzed by means ofa multi-cytokine/chemokine antibodyarray (Panomics). Results are ex-pressed as the mean ± SD of dupli-cate blots and are representative ofthree different experiments. ND = notdetermined.
Increased production of cytokines and chemokines
in MMP-9−/− cardiac allografts recipients
Previous studies have shown that MMP-9 is involved inthe modulation of the activity of several cytokines andchemokines (4,6,7,9,10). To further elucidate the mecha-nisms responsible for the accelerated cellular infiltrationand a up-regulation of T-cell alloreactivity in MMP-9−/−recipients, a semi-quantitative analysis of cytokines andchemokines was performed in the supernatants from theabove cultures. As shown in Figure 5B, spleen cells fromMMP-9−/− recipients showed a significantly higher pro-duction of IL-9, IL-12, IFN-c , MCP-1, MCP-5 and RANTESas compared to spleen cells from MMP-9+/+ recipients. Incontrast, spleen cells from MMP-9−/− recipients showeda significantly lower production of IL-4 and IL-5 as com-pared to spleen cells from MMP-9+/+ recipients. No differ-ences were observed in the levels of IL-3, IL-12p70, IL-13,IL-17 and TNF-a. No cytokine production was observed byspleen cells from MMP-9+/+ and MMP-9−/− recipientscultured in the presence of syngeneic 129/SvEv spleencells (data not shown). These data suggest that alloreactiveMMP-9−/− T cells have a predominant Th1 cytokine pro-file that may lead to an augmented T-cell expansion upon
activation by alloantigens. These results also suggest thatMMP-9−/− T cells may have an enhanced ability to infil-trate the cardiac allografts due to their ability to producehigher levels of MCP-1, MCP-5 and RANTES leading to anaccelerated rejection process.
Decreased innate T-cell alloreactivity and DC
allostimulatory capacity in MMP-2−/− mice
To determine whether the lower T-cell alloreactivity as wellas the abnormal production of cytokines and chemokines inMMP-2−/− recipients were due to a difference in alloanti-gen presentation by DCs or a difference in T-cell allorespon-siveness, we determined the frequency of BALB/c-specificalloreactive T cells in naıve MMP-2+/+ and MMP-2−/−mice by means of an IFN-c ELISPOT assay. As shown inFigure 6A, the frequency of alloreactive T cells respondingto BALB/c DCs was significantly lower in MMP-2−/− micethan in MMP-2+/+ mice (p < 0.0001). No IFN-c -productionwas detected in the negative control cultures containingMMP-2+/+ and MMP-2−/− T cells responding to syn-geneic C57BL/6 DCs (data not shown). Subsequently, wedetermined the capacity of MMP-2+/+ and MMP-2−/−DCs to stimulate IFN-c production by allogeneic BALB/c
American Journal of Transplantation 2004; 5: 517–528 523
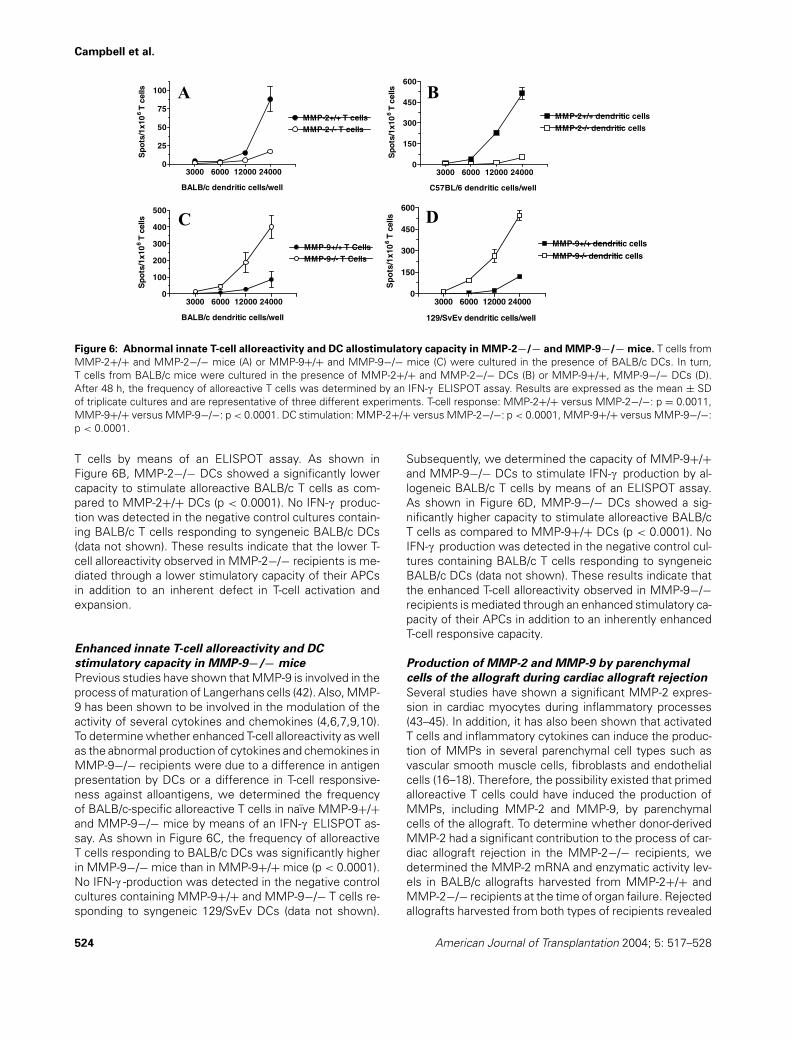
Campbell et al.
3000 6000 12000 240000
25
50
75
100
MMP-2+/+ T cellsMMP-2-/- T cells
BALB/c dendritic cells/well
Sp
ots
/1x1
06
T c
ells A
3000 6000 12000 240000
150
300
450
600
MMP-2+/+ dendritic cells
MMP-2-/- dendritic cells
C57BL/6 dendritic cells/well
Sp
ots
/1x1
06
T c
ells B
3000 6000 12000 240000
100
200
300
400
500
MMP-9+/+ T CellsMMP-9-/- T Cells
BALB/c dendritic cells/well
Sp
ots
/1x1
06
T c
ells C
3000 6000 12000 240000
150
300
450
600
MMP-9+/+ dendritic cells
MMP-9-/- dendritic cells
129/SvEv dendritic cells/well
Sp
ots
/1x1
06
T c
ells D
Figure 6: Abnormal innate T-cell alloreactivity and DC allostimulatory capacity in MMP-2−/− and MMP-9−/− mice. T cells fromMMP-2+/+ and MMP-2−/− mice (A) or MMP-9+/+ and MMP-9−/− mice (C) were cultured in the presence of BALB/c DCs. In turn,T cells from BALB/c mice were cultured in the presence of MMP-2+/+ and MMP-2−/− DCs (B) or MMP-9+/+, MMP-9−/− DCs (D).After 48 h, the frequency of alloreactive T cells was determined by an IFN-c ELISPOT assay. Results are expressed as the mean ± SDof triplicate cultures and are representative of three different experiments. T-cell response: MMP-2+/+ versus MMP-2−/−: p = 0.0011,MMP-9+/+ versus MMP-9−/−: p < 0.0001. DC stimulation: MMP-2+/+ versus MMP-2−/−: p < 0.0001, MMP-9+/+ versus MMP-9−/−:p < 0.0001.
T cells by means of an ELISPOT assay. As shown inFigure 6B, MMP-2−/− DCs showed a significantly lowercapacity to stimulate alloreactive BALB/c T cells as com-pared to MMP-2+/+ DCs (p < 0.0001). No IFN-c produc-tion was detected in the negative control cultures contain-ing BALB/c T cells responding to syngeneic BALB/c DCs(data not shown). These results indicate that the lower T-cell alloreactivity observed in MMP-2−/− recipients is me-diated through a lower stimulatory capacity of their APCsin addition to an inherent defect in T-cell activation andexpansion.
Enhanced innate T-cell alloreactivity and DC
stimulatory capacity in MMP-9−/− mice
Previous studies have shown that MMP-9 is involved in theprocess of maturation of Langerhans cells (42). Also, MMP-9 has been shown to be involved in the modulation of theactivity of several cytokines and chemokines (4,6,7,9,10).To determine whether enhanced T-cell alloreactivity as wellas the abnormal production of cytokines and chemokines inMMP-9−/− recipients were due to a difference in antigenpresentation by DCs or a difference in T-cell responsive-ness against alloantigens, we determined the frequencyof BALB/c-specific alloreactive T cells in naıve MMP-9+/+and MMP-9−/− mice by means of an IFN-c ELISPOT as-say. As shown in Figure 6C, the frequency of alloreactiveT cells responding to BALB/c DCs was significantly higherin MMP-9−/− mice than in MMP-9+/+ mice (p < 0.0001).No IFN-c -production was detected in the negative controlcultures containing MMP-9+/+ and MMP-9−/− T cells re-sponding to syngeneic 129/SvEv DCs (data not shown).
Subsequently, we determined the capacity of MMP-9+/+and MMP-9−/− DCs to stimulate IFN-c production by al-logeneic BALB/c T cells by means of an ELISPOT assay.As shown in Figure 6D, MMP-9−/− DCs showed a sig-nificantly higher capacity to stimulate alloreactive BALB/cT cells as compared to MMP-9+/+ DCs (p < 0.0001). NoIFN-c production was detected in the negative control cul-tures containing BALB/c T cells responding to syngeneicBALB/c DCs (data not shown). These results indicate thatthe enhanced T-cell alloreactivity observed in MMP-9−/−recipients is mediated through an enhanced stimulatory ca-pacity of their APCs in addition to an inherently enhancedT-cell responsive capacity.
Production of MMP-2 and MMP-9 by parenchymal
cells of the allograft during cardiac allograft rejection
Several studies have shown a significant MMP-2 expres-sion in cardiac myocytes during inflammatory processes(43–45). In addition, it has also been shown that activatedT cells and inflammatory cytokines can induce the produc-tion of MMPs in several parenchymal cell types such asvascular smooth muscle cells, fibroblasts and endothelialcells (16–18). Therefore, the possibility existed that primedalloreactive T cells could have induced the production ofMMPs, including MMP-2 and MMP-9, by parenchymalcells of the allograft. To determine whether donor-derivedMMP-2 had a significant contribution to the process of car-diac allograft rejection in the MMP-2−/− recipients, wedetermined the MMP-2 mRNA and enzymatic activity lev-els in BALB/c allografts harvested from MMP-2+/+ andMMP-2−/− recipients at the time of organ failure. Rejectedallografts harvested from both types of recipients revealed
524 American Journal of Transplantation 2004; 5: 517–528
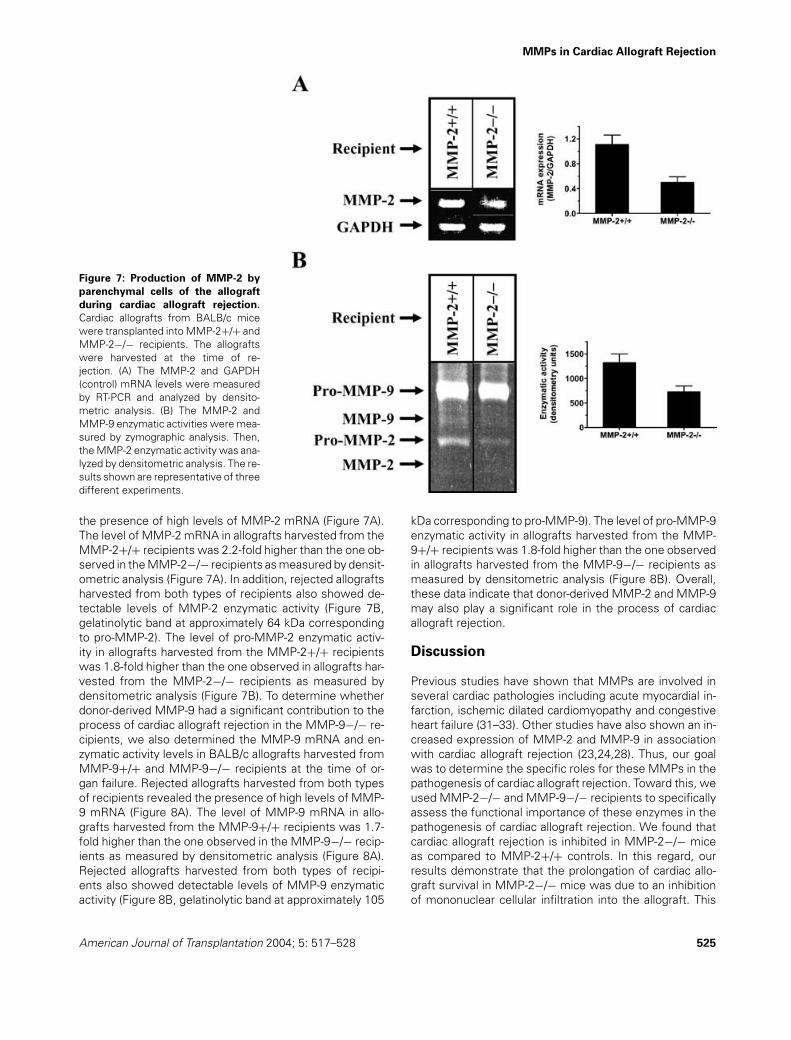
MMPs in Cardiac Allograft Rejection
Figure 7: Production of MMP-2 by
parenchymal cells of the allograft
during cardiac allograft rejection.
Cardiac allografts from BALB/c micewere transplanted into MMP-2+/+ andMMP-2−/− recipients. The allograftswere harvested at the time of re-jection. (A) The MMP-2 and GAPDH(control) mRNA levels were measuredby RT-PCR and analyzed by densito-metric analysis. (B) The MMP-2 andMMP-9 enzymatic activities were mea-sured by zymographic analysis. Then,the MMP-2 enzymatic activity was ana-lyzed by densitometric analysis. The re-sults shown are representative of threedifferent experiments.
the presence of high levels of MMP-2 mRNA (Figure 7A).The level of MMP-2 mRNA in allografts harvested from theMMP-2+/+ recipients was 2.2-fold higher than the one ob-served in the MMP-2−/− recipients as measured by densit-ometric analysis (Figure 7A). In addition, rejected allograftsharvested from both types of recipients also showed de-tectable levels of MMP-2 enzymatic activity (Figure 7B,gelatinolytic band at approximately 64 kDa correspondingto pro-MMP-2). The level of pro-MMP-2 enzymatic activ-ity in allografts harvested from the MMP-2+/+ recipientswas 1.8-fold higher than the one observed in allografts har-vested from the MMP-2−/− recipients as measured bydensitometric analysis (Figure 7B). To determine whetherdonor-derived MMP-9 had a significant contribution to theprocess of cardiac allograft rejection in the MMP-9−/− re-cipients, we also determined the MMP-9 mRNA and en-zymatic activity levels in BALB/c allografts harvested fromMMP-9+/+ and MMP-9−/− recipients at the time of or-gan failure. Rejected allografts harvested from both typesof recipients revealed the presence of high levels of MMP-9 mRNA (Figure 8A). The level of MMP-9 mRNA in allo-grafts harvested from the MMP-9+/+ recipients was 1.7-fold higher than the one observed in the MMP-9−/− recip-ients as measured by densitometric analysis (Figure 8A).Rejected allografts harvested from both types of recipi-ents also showed detectable levels of MMP-9 enzymaticactivity (Figure 8B, gelatinolytic band at approximately 105
kDa corresponding to pro-MMP-9). The level of pro-MMP-9enzymatic activity in allografts harvested from the MMP-9+/+ recipients was 1.8-fold higher than the one observedin allografts harvested from the MMP-9−/− recipients asmeasured by densitometric analysis (Figure 8B). Overall,these data indicate that donor-derived MMP-2 and MMP-9may also play a significant role in the process of cardiacallograft rejection.
Discussion
Previous studies have shown that MMPs are involved inseveral cardiac pathologies including acute myocardial in-farction, ischemic dilated cardiomyopathy and congestiveheart failure (31–33). Other studies have also shown an in-creased expression of MMP-2 and MMP-9 in associationwith cardiac allograft rejection (23,24,28). Thus, our goalwas to determine the specific roles for these MMPs in thepathogenesis of cardiac allograft rejection. Toward this, weused MMP-2−/− and MMP-9−/− recipients to specificallyassess the functional importance of these enzymes in thepathogenesis of cardiac allograft rejection. We found thatcardiac allograft rejection is inhibited in MMP-2−/− miceas compared to MMP-2+/+ controls. In this regard, ourresults demonstrate that the prolongation of cardiac allo-graft survival in MMP-2−/− mice was due to an inhibitionof mononuclear cellular infiltration into the allograft. This
American Journal of Transplantation 2004; 5: 517–528 525
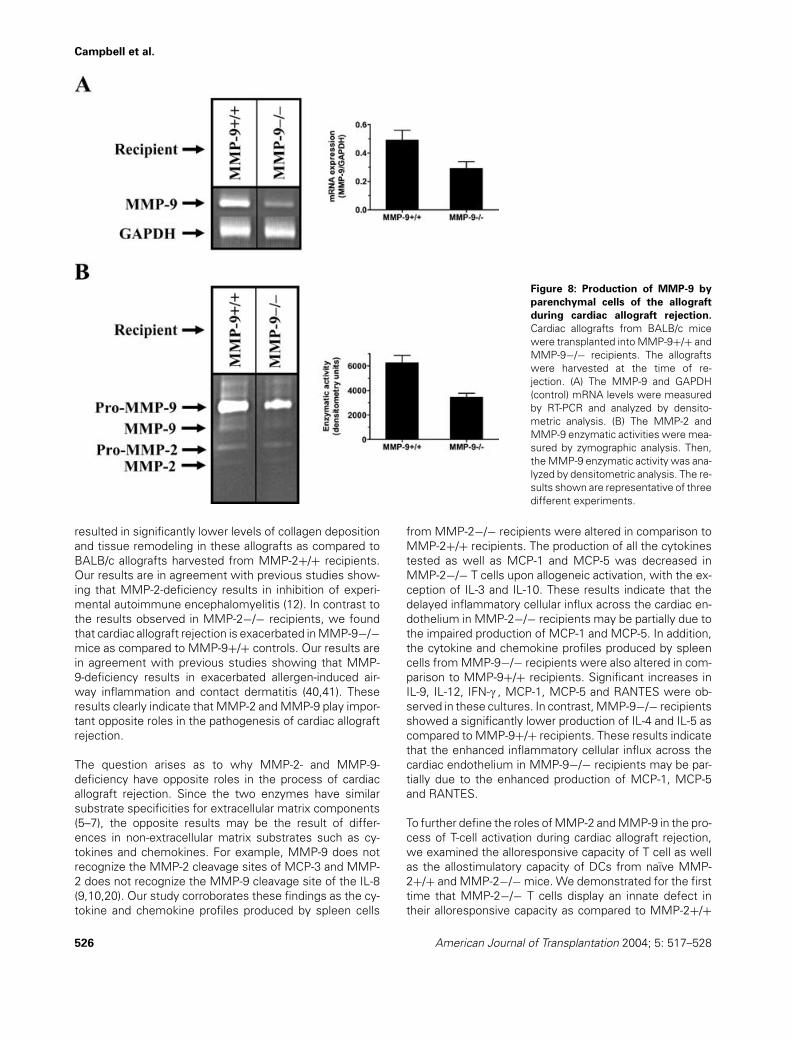
Campbell et al.
Figure 8: Production of MMP-9 by
parenchymal cells of the allograft
during cardiac allograft rejection.
Cardiac allografts from BALB/c micewere transplanted into MMP-9+/+ andMMP-9−/− recipients. The allograftswere harvested at the time of re-jection. (A) The MMP-9 and GAPDH(control) mRNA levels were measuredby RT-PCR and analyzed by densito-metric analysis. (B) The MMP-2 andMMP-9 enzymatic activities were mea-sured by zymographic analysis. Then,the MMP-9 enzymatic activity was ana-lyzed by densitometric analysis. The re-sults shown are representative of threedifferent experiments.
resulted in significantly lower levels of collagen depositionand tissue remodeling in these allografts as compared toBALB/c allografts harvested from MMP-2+/+ recipients.Our results are in agreement with previous studies show-ing that MMP-2-deficiency results in inhibition of experi-mental autoimmune encephalomyelitis (12). In contrast tothe results observed in MMP-2−/− recipients, we foundthat cardiac allograft rejection is exacerbated in MMP-9−/−mice as compared to MMP-9+/+ controls. Our results arein agreement with previous studies showing that MMP-9-deficiency results in exacerbated allergen-induced air-way inflammation and contact dermatitis (40,41). Theseresults clearly indicate that MMP-2 and MMP-9 play impor-tant opposite roles in the pathogenesis of cardiac allograftrejection.
The question arises as to why MMP-2- and MMP-9-deficiency have opposite roles in the process of cardiacallograft rejection. Since the two enzymes have similarsubstrate specificities for extracellular matrix components(5–7), the opposite results may be the result of differ-ences in non-extracellular matrix substrates such as cy-tokines and chemokines. For example, MMP-9 does notrecognize the MMP-2 cleavage sites of MCP-3 and MMP-2 does not recognize the MMP-9 cleavage site of the IL-8(9,10,20). Our study corroborates these findings as the cy-tokine and chemokine profiles produced by spleen cells
from MMP-2−/− recipients were altered in comparison toMMP-2+/+ recipients. The production of all the cytokinestested as well as MCP-1 and MCP-5 was decreased inMMP-2−/− T cells upon allogeneic activation, with the ex-ception of IL-3 and IL-10. These results indicate that thedelayed inflammatory cellular influx across the cardiac en-dothelium in MMP-2−/− recipients may be partially due tothe impaired production of MCP-1 and MCP-5. In addition,the cytokine and chemokine profiles produced by spleencells from MMP-9−/− recipients were also altered in com-parison to MMP-9+/+ recipients. Significant increases inIL-9, IL-12, IFN-c , MCP-1, MCP-5 and RANTES were ob-served in these cultures. In contrast, MMP-9−/− recipientsshowed a significantly lower production of IL-4 and IL-5 ascompared to MMP-9+/+ recipients. These results indicatethat the enhanced inflammatory cellular influx across thecardiac endothelium in MMP-9−/− recipients may be par-tially due to the enhanced production of MCP-1, MCP-5and RANTES.
To further define the roles of MMP-2 and MMP-9 in the pro-cess of T-cell activation during cardiac allograft rejection,we examined the alloresponsive capacity of T cell as wellas the allostimulatory capacity of DCs from naıve MMP-2+/+ and MMP-2−/− mice. We demonstrated for the firsttime that MMP-2−/− T cells display an innate defect intheir alloresponsive capacity as compared to MMP-2+/+
526 American Journal of Transplantation 2004; 5: 517–528

MMPs in Cardiac Allograft Rejection
T cells. Surprisingly, we also determined that MMP-2−/−DCs have an impaired stimulatory capacity as comparedto MMP-2+/+ DCs. Interestingly, we also determined thatMMP-9−/− DCs have an enhanced stimulatory capacityas compared to MMP-9+/+ DCs and that MMP-9−/− Tcells display an enhanced innate alloresponsive capacityas compared to MMP-9+/+ T cells. Our results are in di-rect contrast with previous studies showing that MMP-9−/− Langerhans cells display normal stimulatory capacityto alloreactive T cells (46). These differences in stimula-tory capacity may be explained by the different origin ofthese DCs or by a lower level of purity of Langerhans cellsprepared from epidermal cell cultures. Overall, our resultsindicate that both MMP-2 and MMP-9 play important rolesnot only in the process of T-cell alloactivation but also inthe process of alloantigen presentation by APCs.
Rejected allografts harvested from MMP-2−/− and MMP-9−/− recipients revealed the presence of detectable levelsof MMP-2 and MMP-9 production, respectively. These re-sults indicate that, upon activation, parenchymal cells in theallograft have a significant contribution to the rejection pro-cess through the production of MMP-2 and MMP-9 in situ.Experiments aimed to better define the effect of MMP-2-and MMP-9-deficiency on the pathogenesis of cardiac allo-graft rejection can only be performed with ‘clean’ BALB/callografts carrying the −/− genes. Thus, current studies inour laboratory are focused on the introduction of the MMP-2−/− and MMP-9−/− genes into the BALB/c backgroundto use these mice as donors in our cardiac transplantationprocedure.
In conclusion, our results indicate that MMP-2 and MMP-9 play different roles in the process of mononuclear cel-lular influx into the allograft as well as in the process ofactivation and expansion of alloreactive T cells during car-diac allograft rejection. Thus, MMPs may represent noveltherapeutic targets for the treatment and/or prevention ofcardiac allograft rejection.
Acknowledgments
The authors would like to thank Drs. T. Mohanakumar, Felix G. Fernandezand Jeff Atkinson for critical review of the manuscript.
This work was supported by an award from the American Heart Associ-ation (AJ), a Basic Scientist Faculty Grant from the American Society ofTransplantation (AJ), grants HL29594 and HL47328 from the National Insti-tutes of Health (RMS) and the Alan A. and Edith L. Wolff Charitable Trust(RMS).
References
1. Orosz CG. Local cellular immunology of experimental transplantvascular sclerosis. Clin Transplantation 1996; 10: 100–103.
2. Dengler TJ, Pober JS. Cellular and molecular biology of cardiactransplant rejection. J Nucl Cardiol 2000; 7: 669–685.
3. Joosten SA, van Koten C, Paul LC. Pathogenesis of chronic allo-graft rejection. Transpl Int 2003; 16: 137–145.
4. Goetzl EJ, Banda MJ, Leppert D. Matrix metalloproteinases inimmunity. J Immunol 1996; 156: 1–4.
5. Nagase H, Woessner JF, Jr. Matrix Metalloproteinases. J BiolChem 1999; 274: 21491–21494.
6. Shapiro SD, Senior RM. Matrix Metalloproteinases. Matrix degra-dation and more. Am J Respir Cell Mol Biol 1999; 20: 1100–1102.
7. Sternlicht MD, Werb Z. How matrix metalloproteinases regulatecell behavior. Annu Rev Cell Dev Biol 2001; 17: 463–516.
8. McCawley LJ, Matrisian LM. Matrix metalloproteinases: they’renot just for matrix anymore! Curr Opin Cell Biol 2001; 13: 534–540.
9. Opdenakker G, Van den Steen PE, Van Damme J. Gelatinase B: atuner and amplifier of immune functions. Trends Immunol 2001;22: 571–579.
10. Opdenakker G, Van den Steen PE, Dubois B et al. Gelatinase Bfunctions as regulator and effector in leukocyte biology. J LeukocBiol 2001; 69: 851–859.
11. Van den Steen PE, Dubois B, Nelissen I, Rudd PM, Dwek RA,Opdenakker G. Biochemistry and molecular biology of gelatinaseB or matrix metalloproteinase-9 (MMP-9). Crit Rev Biochem MolBiol 2002; 37: 375–536.
12. Graesser D, Mahooti S, Madri JA. Distinct roles for matrixmetalloproteinase-2 and a4 integrin in autoimmune T cell extrava-sation and residency in brain parenchyma during experimental au-toimmune encephalomyelitis. J Neuroimmunol 2000; 109: 121–131.
13. Dubois B, Masure S, Hurtenbach U et al. Resistance of younggelatinase B-deficient mice to experimental autoimmune en-cephalomyelitis and necrotizing tail lesions. J Clin Invest 1999;104: 1507–1515.
14. Biddison WE, Kubota R, Kawanishi T et al. Human T cell leukemiavirus type I (HTLV-I)-specific CD8+ CTL clones from patients withHTLV-I-associated neurologic disease secrete proinflammatorycytokines, chemokines, and matrix metalloproteinase. J Immunol1997; 159: 2018–2025.
15. Yakubenki VP, Lobb RR, Plow EF, Ugarova TP. Differential in-duction of gelatinase B (MMP-9) and gelatinase A (MMP-2) in Tlymphocytes upon a4b1-mediated adhesion to VCAM-1 and theCS-1 peptide of fibronectin. Exp Cell Res 2000; 260: 73–84.
16. Miltenburg AM, Lacraz S, Welgus HG, Dayer JM. Immobilizedanti-CD3 antibody activates T cell clones to induce the produc-tion of interstitial collagenase, but not tissue inhibitor of metal-loproteinases, in monocyte THP-1 cells and dermal fibroblasts.J Immunol 1995; 154: 2655–2667.
17. Mach F, Schonbeck U, Fabunmi RP et al. T lymphocytes induceendothelial cell matrix metalloproteinase expression by a CD40L-dependent mechanism: implications for tubule formation. Am JPathol 1999; 154: 229–238.
18. Schonbeck U, Mach F, Sukhova GK et al. Regulation of matrixmetalloproteinase expression in human vascular smooth musclecells by T lymphocytes: a role for CD40 signaling in plaque rupture.Circ Res 1997; 81: 448–454.
19. Ito A, Mukaiyama A, Itoh Y et al. Degradation of interleukin-1b bymatrix metalloproteinases. J Biol Chem 1996; 271: 14657–14660.
20. McQuibban GA, Gong JH, Tam EM, McCulloch CA, Clark-Lewis I,Overall CM. Inflammation dampened by gelatinase A cleavage ofmonocyte chemoattractant protein-3. Science 2000; 289: 1202–1206.
21. McQuibban GA, Butler GS, Gong JH et al. Matrix metallopro-teinase activity inactivates the CXC chemokine stromal cell-derived factor-1. J Biol Chem 2001; 276: 43503–43508.
American Journal of Transplantation 2004; 5: 517–528 527

Campbell et al.
22. Corry DB, Rishi K, Kanellis J et al. Decreased allergic lung in-flammatory cell egression and increased susceptibility to as-phyxiation in MMP-2-deficiency. Nat Immunol 2002; 3: 347–353.
23. Suzuki JI, Isobe M, Kawauchi M, Endoh M, Amano J,Takamoto S. Altered expression of matrix metalloproteinasesand tissue inhibitors of metalloproteinases in acutely re-jected myocardium and coronary arteriosclerosis in cardiac al-lografts of nonhuman primates. Transplant Int 2000; 13: 106–113.
24. Tsukioka K, Suzuki JI, Kawauchi M et al. Expression of membrane-type 1 matrix metalloproteinase in coronary vessels of allotrans-planted primate hearts. J Heart Lung Transplant 2000; 19: 1193–1198.
25. Ermolli M, Schumacher M, Lods N, Hammoud M, Marti HP. Dif-ferential expression of MMP-2/MMP-9 and potential benefit ofan MMP inhibitor in experimental acute kidney allograft rejection.Transpl Immunol 2003; 11: 137–145.
26. Laplante A, Liu D, Demeule M et al. Modulation of matrix gelati-nases and metalloproteinase-activating process in acute kidneyrejection. Transpl Int 2003; 16: 262–269.
27. Rodrigo E, Lopez-Hoyos M, Escallada R et al. Circulating levelsof matrix metalloproteinases MMP-3 and MMP-2 in renal trans-plant recipients with chronic transplant nephropathy. Nephrol DialTransplant 2000; 15: 2041–2045.
28. Yamani MH, Starling RC, Young JB et al. Acute vascular rejectionis associated with up-regulation of vitronectin receptor (avb3),increased expression of tissue factor, and activation of the extra-cellular matrix metalloproteinase induction system. J Heart LungTransplant 2002; 21: 983–989.
29. Trello CA, Williams DA, Keller CA, Crim C, Webster RO, Ohar JA.Increased gelatinolytic activity in bronchoalveolar lavage fluid instable lung transplant recipients. Am J Respir Crit Care Med 1997;156: 1978–1986.
30. Beeh KM, Beier J, Kornmann O, Micke P, Buhl R. Spu-tum levels of metalloproteinase-9 and tissue inhibitor ofmetalloproteinase-1 and their ratio correlate with airway obstruc-tion in lung transplant recipients: relation to tumor necrosis factor-a and interleukin-10. J Heart Lung Transplant 2001; 20: 1144–1151.
31. Coker ML, Thomas CV, Clair MJ et al. Myocardial matrix metallo-proteinase activity and abundance with congestive heart failure.Am J Physiol 1998; 274: H1516–H1523.
32. Coker ML, Zellner JL, Crumbley AJ, Spinale FG. Defects in matrixmetalloproteinase inhibitory stoichiometry and selective MMPinduction in patients with nonischemic or ischemic dilated car-diomyopathy. Ann N Y Acad Sci 1999; 878: 559–562.
33. Etoh T, Joffs C, Deschamps AM et al. Myocardial and interstitialmatrix metalloproteinase activity after acute myocardial infarctionin pigs. Am J Physiol Heart Circ Physiol 2001; 281: H987–H994.
34. Itoh T, Ikeda T, Gomi H, Nakao S, Suzuki T, Itohara S. Unalteredsecretion of beta-amyloid precursor protein in gelatinase A (ma-trix metalloproteinase 2)-deficient mice. J Biol Chem 1997; 272:22389–22392.
35. Vu TH, Shipley JM, Bergers G et al. MMP-9/gelatinase B is keyregulator of growth plate angiogenesis and apoptosis of hyper-trophic chondrocytes. Cell 1998; 90: 411–422.
36. Corry RJ, Winn HJ, Russell PS. Primarily vascularized allograftsof hearts in mice. Transplantation 1973; 16: 343–350.
37. Livak KJ, Schmittgen TD. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2−��CT method.Methods 2001; 25: 402–408.
38. Fukao T, Frucht DM, Yap G, Gadlins M, O’Shea JJ, Koyasu S.Inducible expression of stat4 in dendritic cells and macrophagesand its critical role in innate and adaptive immune responses. JImmunol 2001; 166: 4446–4455.
39. McMillan SJ, Kearley J, Campbell JD et al. Matrixmetalloproteinase-9 deficiency results in enhanced allergen-induced airway inflammation. J Immunol 2004; 172: 2586–2594.
40. Wang M, Qin X, Mudgett JS, Ferguson TA, Senior RM, WelgusHG. Matrix metalloproteinase deficiencies affect contact hyper-sensitivity: stromelysin-1 deficiency prevents the response andgelatinase B deficiency prolongs the response. Proc Natl AcadSci U S A 1999; 96: 6885–6889.
41. Sheu BC, Hsu SM, Ho HN, Lien HC, Huang SC, Lin RH. A novelrole of metalloproteinase in cancer-mediated immunosuppres-sion. Cancer Res 2001; 61: 237–242.
42. Kobayashi Y, Matsumoto M, Kotani M, Makino T. Possible in-volvement of matrix metalloproteinase-9 in Langerhans cell mi-gration and maturation. J Immunol 1999; 163: 5989–5993.
43. Tyagi SC, Kumar SG, Banks J, Fortson W. Co-expression of tissueinhibitor and matrix metalloproteinase in myocardium. J Mol CellCardiol 1995; 27: 2177–2189.
44. Coker ML, Doscher MA, Thomas CV, Galis ZS, Spinale FG. Matrixmetalloproteinase synthesis and expression in isolated LV my-ocyte preparations. Am J Physiol 1999; 277: H777–H787.
45. Tyagi SC, Campbell SE, Reddy HK, Tjahja E, Voelker DJ. Matrixmetalloproteinase activity expression in infarcted, noninfarctedand dilated cardiomyopathic human hearts. Mol Cell Biochem1996; 155: 13–21.
46. Ratzinger G, Stoitzner P, Ebner S et al. Matrix metalloproteinases9 and 2 are necessary for the migration of langerhans cells anddermal dendritic cells from human and murine skin. J Immunol2002; 168: 4361–4371.
528 American Journal of Transplantation 2004; 5: 517–528
Related Documents











