Basic nutritional investigation Dietary supplementation with antioxidants improves functions and decreases oxidative stress of leukocytes from prematurely aging mice Carmen Alvarado, B.Sc. a , Pedro Álvarez, B.Sc. a , Marta Puerto, Ph.D. a , Nicolas Gausserès, Ph.D. b , Liliana Jiménez, Ph.D. b , and Mónica De la Fuente, Ph.D. a, * a Department of Physiology (Animal Physiology II), Faculty of Biological Sciences, Complutense University, Madrid, Spain b Danone Vitapole, RN 128, Palaiseau, France Manuscript received November 4, 2005; accepted May 10, 2006. Abstract Objectives: Aging is accompanied by chronic inflammation and oxidative stress, which lead to a marked impairment of immune function and therefore increased mortality. This study assessed the effect of dietary supplementation, for 15 wk, with 5% and 20% (w/w) of biscuits enriched with nutritional doses of vitamins C and E, zinc, selenium, and -carotenes on function and oxidative stress parameters of peritoneal leukocytes from middle-aged, prematurely aging mice (PAM) and non-prematurely aging mice (NPAM). Methods: After supplementation we measured leukocyte functions (adherence, chemotaxis, phago- cytosis, intracellular reactive oxygen species levels, lymphoproliferation, natural killer activity, and interleukin-2 release), antioxidant defenses (superoxide dismutase, glutathione peroxidase, and reduced glutathione), oxidant compounds (extracellular O 2 , glutathione disulfide, glutathione disulfide/reduced glutathione ratio, tumor necrosis factor-, nitric oxide, and prostaglandin E 2 ), and lipid and DNA oxidative damage, measured by malondialdehyde and 8-oxo,7,8-dihydro-2=-deox- yguanosine levels, respectively. Results: In general, leukocyte functions were improved and redox homeostasis was restored after intake of antioxidants. In consequence, malondialdehyde and 8-oxo,7,8-dihydro-2=-deoxyguanosine in PAM and NPAM were strikingly decreased after 5% and 20% supplementation (malondialde- hyde, P 0.001 in PAM; P 0.01 in NPAM after both treatments; 8-oxo,7,8-dihydro-2=- deoxyguanosine, P 0.01 after 5% supplementation and P 0.001 after 20% supplementation in PAM and NPAM). Moreover, the effect of the antioxidants was stronger in PAM than in NPAM, and 20% supplementation was more effective than 5%. Conclusion: Our data suggest that improvement of leukocyte function and restoration of redox balance after consumption of adequate levels of antioxidants from adulthood may be useful to attain healthy aging, especially in animals with premature aging. © 2006 Elsevier Inc. All rights reserved. Keywords: Aging; Antioxidants; Dietary supplementation; Immune system; Oxidative stress; Peritoneal leukocytes; Prematurely aging mice Introduction The immune system changes with age, developing a complex and progressive decline [1,2]. One of the most important consequences is the increased incidence of infec- tious and degenerative diseases that result in a high mortal- ity in elderly people [3]. Moreover, immunologic mecha- nisms have an influence on the process and manifestations of aging in many other systems [4], and the immune system appears to have a key role in the process of aging of all physiologic systems of the organism. Accordingly, cente- narians and very old mice show well-preserved leukocyte functions [5,6]. This work was supported by a grant from Danone Vitapole. E-mail address: (M. De la Fuente). * Corresponding author. Tel.: 34-913-944-986; fax: 34-913-944- 935. E-mail address: [email protected] Nutrition 22 (2006) 767–777 www.elsevier.com/locate/nut 0899-9007/06/$ – see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.nut.2006.05.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A
K
I
c
E
9
t
0d
Basic nutritional investigation
Dietary supplementation with antioxidants improves functions anddecreases oxidative stress of leukocytes from prematurely aging mice
Carmen Alvarado, B.Sc.a, Pedro Álvarez, B.Sc.a, Marta Puerto, Ph.D.a,Nicolas Gausserès, Ph.D.b, Liliana Jiménez, Ph.D.b, and Mónica De la Fuente, Ph.D.a,*
a Department of Physiology (Animal Physiology II), Faculty of Biological Sciences, Complutense University, Madrid, Spainb Danone Vitapole, RN 128, Palaiseau, France
Manuscript received November 4, 2005; accepted May 10, 2006.
bstract Objectives: Aging is accompanied by chronic inflammation and oxidative stress, which lead to amarked impairment of immune function and therefore increased mortality. This study assessed theeffect of dietary supplementation, for 15 wk, with 5% and 20% (w/w) of biscuits enriched withnutritional doses of vitamins C and E, zinc, selenium, and �-carotenes on function and oxidativestress parameters of peritoneal leukocytes from middle-aged, prematurely aging mice (PAM) andnon-prematurely aging mice (NPAM).Methods: After supplementation we measured leukocyte functions (adherence, chemotaxis, phago-cytosis, intracellular reactive oxygen species levels, lymphoproliferation, natural killer activity, andinterleukin-2 release), antioxidant defenses (superoxide dismutase, glutathione peroxidase, andreduced glutathione), oxidant compounds (extracellular O2
�, glutathione disulfide, glutathionedisulfide/reduced glutathione ratio, tumor necrosis factor-�, nitric oxide, and prostaglandin E2), andlipid and DNA oxidative damage, measured by malondialdehyde and 8-oxo,7,8-dihydro-2=-deox-yguanosine levels, respectively.Results: In general, leukocyte functions were improved and redox homeostasis was restored afterintake of antioxidants. In consequence, malondialdehyde and 8-oxo,7,8-dihydro-2=-deoxyguanosinein PAM and NPAM were strikingly decreased after 5% and 20% supplementation (malondialde-hyde, P � 0.001 in PAM; P � 0.01 in NPAM after both treatments; 8-oxo,7,8-dihydro-2=-deoxyguanosine, P � 0.01 after 5% supplementation and P � 0.001 after 20% supplementation inPAM and NPAM). Moreover, the effect of the antioxidants was stronger in PAM than in NPAM,and 20% supplementation was more effective than 5%.Conclusion: Our data suggest that improvement of leukocyte function and restoration of redoxbalance after consumption of adequate levels of antioxidants from adulthood may be useful to attainhealthy aging, especially in animals with premature aging. © 2006 Elsevier Inc. All rightsreserved.
eywords: Aging; Antioxidants; Dietary supplementation; Immune system; Oxidative stress; Peritoneal leukocytes;
Nutrition 22 (2006) 767–777www.elsevier.com/locate/nu
Prematurely aging mice
itinoapn
ntroduction
The immune system changes with age, developing aomplex and progressive decline [1,2]. One of the most
This work was supported by a grant from Danone Vitapole.-mail address: (M. De la Fuente).
* Corresponding author. Tel.: �34-913-944-986; fax: �34-913-944-35.
fE-mail address: [email protected]
899-9007/06/$ – see front matter © 2006 Elsevier Inc. All rights reserved.oi:10.1016/j.nut.2006.05.007
mportant consequences is the increased incidence of infec-ious and degenerative diseases that result in a high mortal-ty in elderly people [3]. Moreover, immunologic mecha-isms have an influence on the process and manifestationsf aging in many other systems [4], and the immune systemppears to have a key role in the process of aging of allhysiologic systems of the organism. Accordingly, cente-arians and very old mice show well-preserved leukocyte
unctions [5,6].
tdttaiadjcoi
iaadltiamfto
tdawsacocr
makweC
M
A
lwmaA
c�wdfbmooE
E
emothfomp
2ewoapitm
dgafsa8eCbedapftwiPmo
768 C. Alvarado et al. / Nutrition 22 (2006) 767–777
Moreover, leukocyte functions are strongly influenced byhe antioxidant/oxidant balance, because immune cells pro-uce reactive oxygen species (ROS) to support their func-ions [7]. From this point of view, the antioxidant levels ofhese cells are very important to maintain redox homeostasisnd, therefore, an adequate function, especially during ox-dative stress situations such as aging [7]. Thus, adequatemounts of neutralizing antioxidants are required to preventamage to the immune cells themselves. Therefore, it seemsustified to assess age-related function changes of diverseells that constitute the immune system and the redox statef these cells, factors that influence each other and that aremportant to determine the rate of aging [7–9].
Nutrition has great importance in the maintenance ofmmune function. Accumulating evidence suggests a strongssociation between diets rich in antioxidant compoundsnd a decreased incidence of cancer and other age-relatediseases, and it has been proposed that the immunomodu-atory effect of these diets might account, at least in part, forhis [3,10,11]. Further, most studies in the area of nutritionalmmunology have been carried out in aged experimentalnimals and elderly subjects using single-element supple-ents. In contrast, there are few data from studies per-
ormed during middle age in animals, when it is assumedhat the immune system is still functioning at a practicallyptimal level [10,12].
Although many dietary antioxidants have been suggestedo contribute to the improvement of immune function byifferent mechanisms, to our knowledge very limited datare available on the effect of supplementation of the dietith mixtures of antioxidants, which in general have been
hown to be more effective than administration of eachntioxidant separately [13,14]. This is an interesting con-ept, whereby we can obtain a greater effect of antioxidantsn leukocyte function by using lower concentrations of eachompound, because high doses of several antioxidants canesult in deleterious effects [15].
Thus, the objective of the present study was to deter-ine, in a model of premature senescence, several functions
nd oxidative stress levels (redox state) of peritoneal leu-ocytes from middle-age female mice (ICR/CD-1), after 15k of 5% and 20% supplementation of the diet with biscuits
nriched with nutritional doses of the antioxidants vitaminsand E, �-carotenes, selenium, and zinc.
aterials and methods
nimals
We used 270 adult female ICR/CD-1 mice (Mus muscu-us) purchased from Harlan Ibérica (Barcelona, Spain),hich were 28 � 2 wk old on arrival to our laboratory. Theice were specifically pathogen free, as tested by Harlan
ccording to Federation of European Laboratory Science
ssociations recommendations, and were housed five per dage and maintained in a temperature-controlled room (222°C), with a 12-h light/dark reversed cycle. All mice
ere initially fed tap water and standard Sander Mus (A04iet from Panlab L.S., Barcelona, Spain) pellets ad libitumor a 1-wk adaptation period and during performance ofehavioral probes. Diet was in accordance with the recom-endations of the American Institute of Nutrition for lab-
ratory animals. Mice were treated according to guidelinesf the European Community Council Directives (86/6091EC).
xperimental groups and animal diets
Previous work of our laboratory showed that the differ-nces in spontaneous exploratory behavior in a simple T-aze test of mouse populations may be related to their level
f senescence and longevity [16,17]. Thus, animals thatook longer to explore the T-shaped maze because of theirigher levels of anxiety [18] had a decline in immuneunction [16,17] and a neurochemistry similar to those ofld mice [19]. Thus, we designated these animals as pre-aturely aging mice (PAM) in contrast to non-
rematurely aging mice (NPAM).Animals were marked for their individual follow-ups. At
9 � 2 wk of age the spontaneous exploratory behavior ofach mouse was tested in a T-shaped maze [17]. This testas performed once a week for 4 consecutive weeks to sortut the PAM (which completed the exploration in �10 s atll four tests) from the NPAM (which completed that ex-loration in �10 s at all four tests). Animals showing anntermediate response were removed from the study. Thisest was always performed between 0900 and 1100 h, toinimize circadian variations, under red light.Non-prematurely aging mice and PAM were randomly
ivided into control and supplemented groups, with eachroup containing 30 mice. Control groups (control NPAMnd PAM, respectively) received 100% of a standard dietor maintenance (AIN-93M diet from Panlab L.S.) [20] andupplemented groups (5% and 20% supplemented NPAMnd PAM, respectively) received a diet made with 95% or0% of the standard diet plus 5% or 20% (w/w) of biscuitsnriched with nutritional doses of the antioxidants vitamins
and E, �-carotene, zinc, and selenium (Table 1); theseiscuits are readily and commercially accessible to consum-rs (Danone Vitapole, Palaiseau, France). The 5% and 20%iet supplementations were maintained for 15 wk and then,t the age of 50 � 2 wk, when mice were middle-aged,eritoneal leukocyte samples were obtained to evaluate theunctional and oxidative stress states of these cells. We usedhe same dose of these antioxidants (20% of biscuits,/w) as in a previous study in which we detected an
mprovement of several leukocyte functions in youngAM [21]. Moreover, we extended the period of supple-entation until 15 wk because the requirement of anti-
xidant ingestion increased with age [22]. Further, we
etermined whether supplementation of the diet with 5%
ot
nsdahupNaaeso
C
1soAcMfiaJ
wpHdrtii
A
wsRcsasRdit
pdcntiaomeIiltefSMfl(satl
TMV
t�E
it
wb
769C. Alvarado et al. / Nutrition 22 (2006) 767–777
f biscuits was sufficient to produce positive effects onhe parameters studied.
With regard to distribution of the antioxidant compo-ents of the biscuits in the physiologic systems, severaltudies made by other laboratories showed that the antioxi-ants present in those biscuits are bioavailable and wellbsorbed, because increased levels in plasma and tissuesave been observed after ingestion of doses similar to thosesed in the present work, without any adverse effect (datarovided by Danone Vitapole and presented at the Eighthordic Nutrition Conference, Norway, 2004). When the
ntioxidant-supplemented diet was administered, the aver-ge food intake and body weight of the mice of eachxperimental group during the supplementation period wereimilar to those found in control NPAM (4.79 g/d per mousef food intake and 42.25 g/mouse of body weight).
ollection of peritoneal leukocytes
Peritoneal suspensions were obtained between 0800 and000 h to minimize circadian variations of the parameterstudied without sacrificing the animals [17]. Briefly, 3 mLf sterile Hank’s solution was injected intraperitoneally.fter massaging the abdomen, peritoneal exudate cells were
ollected, allowing recovery of 80% of the injected volume.acrophages and lymphocytes were identified and quanti-
ed by their morphology with optical microscopy (40�)nd cytometric analysis (FACScan, Becton Dickinson, San
able 1ain nutrient composition of biscuits commercialized by Danoneitapole
Nutrient Biscuit
Energy (kcal/kg) 4581.3Proteins (g/kg) 82.8Lipids (g/kg) 155.03Carbohydrates (g/kg) 713.7�-3 (g/kg) 14.40�-6 (g/kg) 45.48Vitamin C* (g/kg) 0.693Vitamin E† (g/kg) 0.192�-Carotene‡ (g/kg) 0.034Zinc§ (g/kg) 0.077Selenium� (�g/kg) 730
* Ascorbic acid was added as a source of vitamin C.† Vegetables oil extracts that contained the natural form, RRR-d-�-
ocopherol, which exhibits more bioactivity than the synthetic one (all rac-tocopherol) and other natural forms, were added as a source of vitamin.
‡ Pure natural �-carotene included in a mixture of starch and vitamin E,n a minimum amount that was not significant in terms of supplementation,o avoid its oxidation, was added as a source of �-carotene.
§ Zinc oxide was added as a source of zinc.� Selenium-enriched yeast, containing an organic selenium form in
hich the main component is L(�)-selenomethionine, with the highestioavailability and biological activity, was added as a source of selenium.
ose, CA, USA) using Cell-Quest (Becton Dickinson) soft- s
are on the basis of the different sizes and structural com-lexities of the cells, and then adjusted by dilution withank’s solution depending on the parameter analyzed (in-icated in the corresponding method). Cellular viability,outinely measured before and after each experiment byrypan blue exclusion test, was �95% in all cases. Allncubations were performed at 37°C in a sterile and humid-fied atmosphere of 5% CO2.
ssays of phagocytic function in peritoneal macrophages
Macrophages were counted and then adjusted by dilutionith Hank’s solution to 5 � 105 macrophages/mL in the
amples in which we assayed these cell functions, except forOS levels in which macrophages were adjusted to 106
ells/mL. In the peritoneal suspension, we carried out thetudy of the main steps of the phagocytic process, namelydherence capacity, chemotaxis, phagocytosis, and the re-piratory burst (intracellular levels of superoxide anion andOS according to various standardized methods previouslyescribed by our group [8,17,21]). Thus, the adherencendex (AI) was calculated according to the following equa-ion:
AI � 1
� ��non-adherent cells ⁄ mL� ⁄ �initial cells ⁄ mL�� � 100(1)
The chemotaxis index (CI) was evaluated by the ca-acity of the macrophages to cross a filter, with a poreiameter of 3 �m included in a Boyden chamber. CI wasalculated by counting, in an optical microscope, the totalumber of macrophages in one-third of the lower face ofhe filters. Phagocytosis was determined as the number ofnert particles (latex beads) ingested by 100 macrophagesnd expressed as a phagocytic index and the percentagef macrophages with phagocytic capacity (with one orore latex beads ingested) was expressed as phagocytic
fficiency; these were counted in an optical microscope.ntracellular levels of superoxide anion were evaluated,n non-stimulated samples and samples stimulated withatex beads (diluted to 1% in Hank’s balanced salt solu-ion), at 525 nm in a spectrophotometer, and results werexpressed as nanomoles per 106 cells by extrapolatingrom a standard curve of Nitroblue tetrazolium (Sigma,t. Louis, MO, USA) reduced with 1,4-dithioerythritol.oreover, intracellular ROS levels were measured by
uorometry using dichlorodihydrofluorescein diacetateMolecular Probes, Eugene, OR, USA) in non-stimulatedamples and samples stimulated with phorbol myristatecetate (50 ng/mL; Sigma), and results were expressed ashe stimulation index, which is the percentage of stimu-ation in response to phorbol myristate acetate, with non-
timulated values being 100%.
A
L
tm
P
h51gpS(Sn3
hdCm
C
tctbroenre
I
maduSp
A
S
mtbsrm
G
btce1pawOtfi�dtma
G
Gc[aRcsv
O
Ta
gtd4csCPRwn
N
nmcM
770 C. Alvarado et al. / Nutrition 22 (2006) 767–777
ssays of peritoneal lymphocyte function
ymphocyte adherence and chemotaxis assayLymphocyte adherence and the chemotaxis capacity of
hese cells were evaluated as previously described for theacrophages.
roliferation assayA previously described method was used [21]. Two
undred microliters of peritoneal suspension, adjusted to� 105 leukocytes/mL in complete medium (RPMI-
640, PAA; plus 10% fetal bovine serum, Gibco; plus 1%entamicin, PAA, Pasching, Austria), was cultured inlates with concanavalin A (Con A; 1 �g/mL per well;igma, Paisley, Scotland, UK) and lipopolysaccharideLPS; Escherichia coli, 055:B5, 1 �g/mL per well;igma) as stimulated samples or in complete medium ason-stimulated samples. After 48 h of incubation, 5 �CiH-thymidine was added to each well and cells werearvested. Results were expressed as the stimulation in-ex, which is the percentage of stimulation in response toon A or LPS, with non-stimulated values (counts perinute) being 100%.
ytotoxicity assayMurine lymphoma YAC-1 cells were used as targets in
he natural killer (NK) activity assay [21]. An enzymaticolorimetric assay was used for cytolysis measurements ofarget cells (Cytotox 96 TM, Promega, Madison, WI, USA)ased on determination of lactate dehydrogenase using tet-azolium salts. Cells were cultured at an effector/target ratef 10/1 and incubated for 4 h. Then lactate dehydrogenasenzymatic activity was measured in 50 �L/well of super-atants by addition of the enzyme substrate and absorbanceecording spectrophotometrically at 490 nm. Results werexpressed as percentage of lysis of target cells.
nterleukin-2 release assayThe concentration of interleukin-2 (IL-2) was deter-
ined on culture supernatants of peritoneal leukocytes after48-h incubation with the mitogen Con A (previously
escribed in the proliferation assay method) and measuredsing an enzyme-linked immunosorbent assay kit (R&Dystems, Minneapolis, MN, USA) [21]. Results were ex-ressed as picograms per milliliter.
ntioxidant defences in peritoneal leukocytes
uperoxide dismutase assaySuperoxide dismutase (SOD) enzymatic activity was
easured using the pyrogallol (Sigma) compound to followhe reaction according to the technique previously describedy De la Fuente et al. [8]. Enzymatic activity was followedpectrophotometrically at 420 nm through oxidation of py-ogallol. Results were expressed as arbitrary units of enzy-
atic activity per 106 cells. llutathione peroxidase assayGlutathione peroxidase (GPx) enzyme was measured
y using the original method [23] with some modifica-ions. Peritoneal suspensions were adjusted to 106
ells/mL and aliquots of 500 �L were used to measurenzymatic activity. Cells were centrifuged at 1076g for0 min at 4°C, and pellets were resuspended in 50 mM ofhosphate buffer (pH 7.0). Then samples were sonicatednd centrifuged at 3200g for 20 min at 4°C. Total activityas determined with cumene hydroperoxide (cumene-OH; Sigma) that oxidized the reduced form of gluta-
hione (GSH) present in the samples to glutathione disul-de (GSSG), which is regenerated by the addition of-nicotinamide adenine dinucleotide phosphate in re-uced form (Sigma) in the presence of glutathione reduc-ase (Sigma). The reaction was followed spectrophoto-etrically at 340 nm. Enzymatic activity was expressed
s milliunits of GPx per 106 cells.
lutathione levelsTotal contents of glutathione and the oxidized form,
SSG, were determined by the Tietze enzymatic recy-ling assay, with some modifications made by our group8]. The reaction was monitored spectrophotometricallyt 412 nm for total glutathione and GSSG determinations.esults were expressed as nanomoles per 106cells. Toalculate the GSSG/GSH ratio, GSH was obtained byubtracting the GSSG value from the total glutathionealue.
xidant compound levels in peritoneal leukocytes
umor necrosis factor-� and prostaglandin E2 releasessay
Mouse tumor necrosis factor-� (TNF-�) and prosta-landin E2 (PGE2) releases were measured in superna-ants from cultures of peritoneal leukocytes (previouslyescribed in the proliferation assay method) [8]. After8 h of incubation, supernatants were collected and con-entrations of the proinflammatory mediators were mea-ured with enzyme-linked immunosorbent assay kits (Hy-ult Biotechnology for TNF-� and R&D Systems forGE2) (HyCult Biotechnology, Uden, The Netherlands;&D Systems, Minneapolis, MN, USA). The resultsere expressed as picograms per milliliter for TNF-� andanograms per milliliter for PGE2.
itric oxide release assayNitric oxide (NO) release was measured in the super-
atants from peritoneal leukocyte cultures according to aethod previously described by our group [8] using a
olorimetric assay kit (Cayman Chemical, Ann Arbor,I, USA). Results were expressed as micromoles per
iter.

Ll
L
acwwSm�mf(pnctuMn
Nd
nw8ycanoo
S
ttpgimEilbpoat
R
M
r5n0a2Pcvttpa0ro2umf0sbc�N
L
aaPNa0rtmts�2Pfctt
771C. Alvarado et al. / Nutrition 22 (2006) 767–777
ipid and nuclear DNA oxidative damage of peritonealeukocytes
ipid oxidative damage: levels of malondialdehydeLipid peroxidation was assessed as formation of malondi-
ldehyde (MDA) and measured by high-performance liquidhromatography (HPLC) as described by Chirico et al. [24],ith some modifications. Briefly, aliquots of 5 � 105 cellsere resuspended in 100 �L of 50 mM of KH2PO4 (pH 6.8;igma), and after sonication of the samples, the followingixture reaction was added: 150 �L of 0.44 M H3PO4 plus 50L of 2-thiobarbituric acid (6 mg/mL; Sigma) plus 10 �L of 3M butylated hydroxytoluene (Sigma) and incubated at 95°C
or 30 min. Reaction was stopped and 250 �L of n-butanolPanreac, Barcelona, Spain) were added to extract the organichase, where MDA is located. Afterward, 50 �L of the super-atant was injected in the HPLC column (Novapack C18, 15m � 3.9 mm; Waters, Rochester, MN, USA). The elute fromhe HPLC was monitored at � 532 nm in a Waters 486ltraviolet detector. Standard curves were obtained daily usingDA-bis (dimethyl acetal; Sigma). Results were expressed as
anomoles per 106 cells.
uclear DNA oxidative damage: levels of 8-oxo,7,8-yhidro-2=-deoxyguanosine
Nuclear DNA (nDNA) was isolated after sonication of perito-eal leukocyte samples containing 6 � 106 cells [8]. Then nDNAas digested to a deoxynucleoside level and concentrations of-oxo,7,8-dihydro-2=-deoxyguanosine (8-oxodG) and deox-guanosine (dG) were measured by HPLC with online electro-hemical and ultraviolet detection, respectively. Comparison ofreas of 8-oxodG standards injected with and without simulta-eous injection of dG standards ensured that no oxidation of dGccurred during HPLC analysis. Results were expressed as a ratiof 8-oxodG to dG multiplied by 10�5.
tatistical analysis
SPSS 11.5 (SPSS Inc., Chicago, IL, USA) was used for sta-istical analysis of results. All data are expressed as mean � SD ofhe number of values corresponding to the same number of ex-eriments and presented within parentheses for each experimentalroup and parameter measured in the different figures. Each values the mean of data from an assay performed in duplicate. Nor-ality of distribution was tested by the Kolmogorov-Smirnov test.quality of variance was checked by Levene’s test. Data from
mmune function parameters and oxidative stress state were ana-yzed by two-way analysis of variance, with the two factors beingehavior (PAM and NPAM) and supplementation (control, sup-lemented 5%, and supplemented 20%). Tukey’s test with a levelf significance set at P � 0.05 was used for post hoc analysis. Inddition, Student’s t test was used for comparisons between con-
rol NPAM and PAM. Minimum significance level was P � 0.05. sesults
acrophages function
The analysis of the results obtained on peritoneal mac-ophage functions is presented in Table 2. Regarding AI,% and 20% supplemented PAM showed statistically sig-ificant lower values than control PAM (P � 0.006 and P �.001, respectively), whereas no effect was observed afterntioxidant ingestion in NPAM. The CI increased only after0% supplementation in PAM and NPAM (P � 0.043 and� 0.042, respectively). With regard to phagocytic effi-
iency, 5% and 20% supplementation increased the controlalues obtained in PAM (P � 0.05 and P � 0.046, respec-ively), whereas no effect was observed in NPAM afterreatment. Antioxidant supplementation increased thehagocytic index in PAM (P � 0.002 and P � 0.001 for 5%nd 20% supplementation, respectively) and NPAM (P �.006 and P � 0.007 for 5% and 20% supplementation,espectively). Analysis of the stimulated intracellular super-xide anion showed statistically significant differences after0% supplementation in PAM (P � 0.001). However, val-es obtained in NPAM were unaffected after both treat-ents. In addition, the PAM group showed significant dif-
erences between 5% and 20% supplementation (P �.001), and the effect of 20% supplementation was moreignificant. With regard to ROS levels stimulated by phor-ol myristate acetate, 5% and 20% supplementation in-reased this leukocyte function in PAM (P � 0.007 and P
0.006, respectively), whereas no effect was observed inPAM.
ymphocyte functions
Different functions of peritoneal lymphocytes assessedre presented in Table 2. Adherence capacity (AI) decreasedfter 5% and 20% supplementation in PAM (P � 0.025 and� 0.013, respectively). There was no treatment effect inPAM. Statistical analysis showed significant differences
fter 20% supplementation with regard to CI (PAM, P �.05; NPAM, P � 0.001), showing an evident increase withespect to control values. Lymphoproliferation in responseo Con A improved in PAM (P � 0.001) after 20% supple-entation, whereas no differences were found in response
o LPS after treatment. The 5% and 20% supplementationtrikingly increased NK activity in PAM (P � 0.008 and P
0.003, respectively). With regard to IL-2 release, only0% supplementation was able to improve this parameter inAM (P � 0.001). Moreover, we obtained significant dif-erences in PAM between the two treatments in chemotaxisapacity (P � 0.006), Con A–stimulated lymphoprolifera-ion (P � 0.001), and IL-2 release (P � 0.001), showinghat 20% supplementation had a stronger effect than 5%
upplementation on these parameters.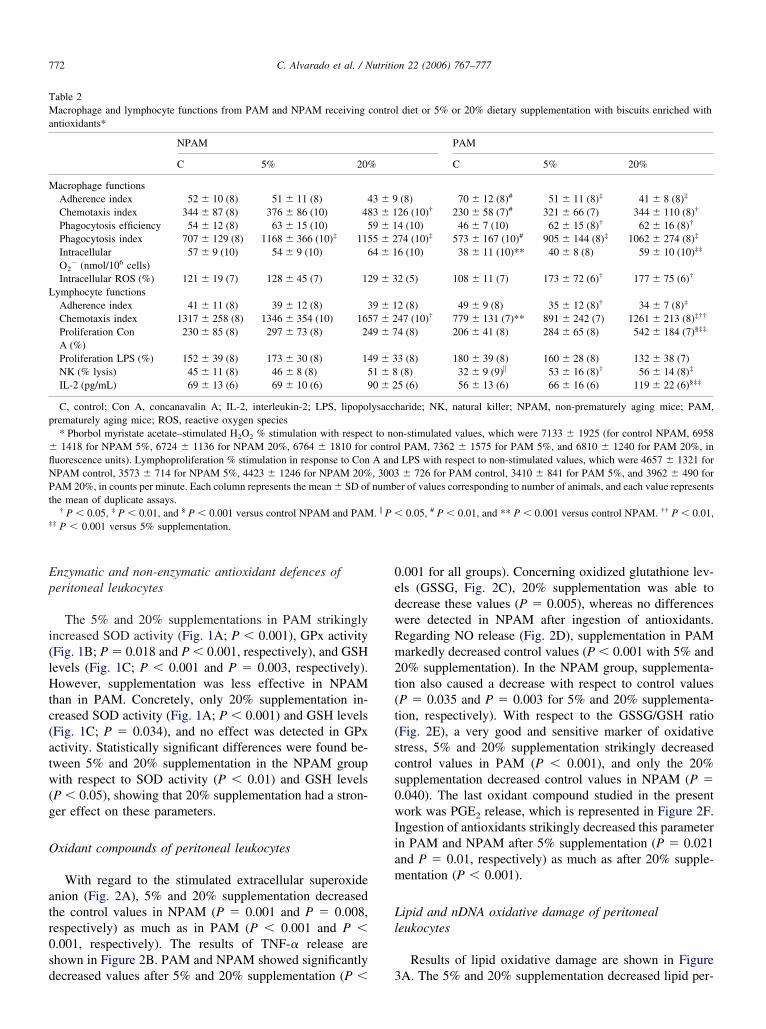
Ep
i(lHtc(atw(g
O
atr0sd
0edwRm2t(t(scs0wIiam
Ll
TMa
M
L
p
�flNPt
‡
772 C. Alvarado et al. / Nutrition 22 (2006) 767–777
nzymatic and non-enzymatic antioxidant defences oferitoneal leukocytes
The 5% and 20% supplementations in PAM strikinglyncreased SOD activity (Fig. 1A; P � 0.001), GPx activityFig. 1B; P � 0.018 and P � 0.001, respectively), and GSHevels (Fig. 1C; P � 0.001 and P � 0.003, respectively).owever, supplementation was less effective in NPAM
han in PAM. Concretely, only 20% supplementation in-reased SOD activity (Fig. 1A; P � 0.001) and GSH levelsFig. 1C; P � 0.034), and no effect was detected in GPxctivity. Statistically significant differences were found be-ween 5% and 20% supplementation in the NPAM groupith respect to SOD activity (P � 0.01) and GSH levels
P � 0.05), showing that 20% supplementation had a stron-er effect on these parameters.
xidant compounds of peritoneal leukocytes
With regard to the stimulated extracellular superoxidenion (Fig. 2A), 5% and 20% supplementation decreasedhe control values in NPAM (P � 0.001 and P � 0.008,espectively) as much as in PAM (P � 0.001 and P �.001, respectively). The results of TNF-� release arehown in Figure 2B. PAM and NPAM showed significantly
able 2acrophage and lymphocyte functions from PAM and NPAM receiving
ntioxidants*
NPAM
C 5% 20%
acrophage functionsAdherence index 52 � 10 (8) 51 � 11 (8) 4Chemotaxis index 344 � 87 (8) 376 � 86 (10) 48Phagocytosis efficiency 54 � 12 (8) 63 � 15 (10) 5Phagocytosis index 707 � 129 (8) 1168 � 366 (10)‡ 115IntracellularO2
� (nmol/106 cells)57 � 9 (10) 54 � 9 (10) 6
Intracellular ROS (%) 121 � 19 (7) 128 � 45 (7) 12ymphocyte functionsAdherence index 41 � 11 (8) 39 � 12 (8) 3Chemotaxis index 1317 � 258 (8) 1346 � 354 (10) 165Proliferation ConA (%)
230 � 85 (8) 297 � 73 (8) 24
Proliferation LPS (%) 152 � 39 (8) 173 � 30 (8) 14NK (% lysis) 45 � 11 (8) 46 � 8 (8) 5IL-2 (pg/mL) 69 � 13 (6) 69 � 10 (6) 9
C, control; Con A, concanavalin A; IL-2, interleukin-2; LPS, lipoporematurely aging mice; ROS, reactive oxygen species* Phorbol myristate acetate–stimulated H2O2 % stimulation with respec1418 for NPAM 5%, 6724 � 1136 for NPAM 20%, 6764 � 1810 for
uorescence units). Lymphoproliferation % stimulation in response to ConPAM control, 3573 � 714 for NPAM 5%, 4423 � 1246 for NPAM 20%AM 20%, in counts per minute. Each column represents the mean � SD of
he mean of duplicate assays.† P � 0.05, ‡ P � 0.01, and § P � 0.001 versus control NPAM and PAM
‡ P � 0.001 versus 5% supplementation.
ecreased values after 5% and 20% supplementation (P � 3
.001 for all groups). Concerning oxidized glutathione lev-ls (GSSG, Fig. 2C), 20% supplementation was able toecrease these values (P � 0.005), whereas no differencesere detected in NPAM after ingestion of antioxidants.egarding NO release (Fig. 2D), supplementation in PAMarkedly decreased control values (P � 0.001 with 5% and
0% supplementation). In the NPAM group, supplementa-ion also caused a decrease with respect to control valuesP � 0.035 and P � 0.003 for 5% and 20% supplementa-ion, respectively). With respect to the GSSG/GSH ratioFig. 2E), a very good and sensitive marker of oxidativetress, 5% and 20% supplementation strikingly decreasedontrol values in PAM (P � 0.001), and only the 20%upplementation decreased control values in NPAM (P �.040). The last oxidant compound studied in the presentork was PGE2 release, which is represented in Figure 2F.
ngestion of antioxidants strikingly decreased this parametern PAM and NPAM after 5% supplementation (P � 0.021nd P � 0.01, respectively) as much as after 20% supple-entation (P � 0.001).
ipid and nDNA oxidative damage of peritonealeukocytes
Results of lipid oxidative damage are shown in Figure
l diet or 5% or 20% dietary supplementation with biscuits enriched with
PAM
C 5% 20%
(8) 70 � 12 (8)# 51 � 11 (8)‡ 41 � 8 (8)‡
26 (10)† 230 � 58 (7)# 321 � 66 (7) 344 � 110 (8)†
4 (10) 46 � 7 (10) 62 � 15 (8)† 62 � 16 (8)†
74 (10)‡ 573 � 167 (10)# 905 � 144 (8)‡ 1062 � 274 (8)‡
6 (10) 38 � 11 (10)** 40 � 8 (8) 59 � 10 (10)‡‡
2 (5) 108 � 11 (7) 173 � 72 (6)† 177 � 75 (6)†
2 (8) 49 � 9 (8) 35 � 12 (8)† 34 � 7 (8)‡
47 (10)† 779 � 131 (7)** 891 � 242 (7) 1261 � 213 (8)‡††
4 (8) 206 � 41 (8) 284 � 65 (8) 542 � 184 (7)§‡‡
3 (8) 180 � 39 (8) 160 � 28 (8) 132 � 38 (7)(8) 32 � 9 (9)� 53 � 16 (8)† 56 � 14 (8)‡
5 (6) 56 � 13 (6) 66 � 16 (6) 119 � 22 (6)§‡‡
haride; NK, natural killer; NPAM, non-prematurely aging mice; PAM,
n-stimulated values, which were 7133 � 1925 (for control NPAM, 6958l PAM, 7362 � 1575 for PAM 5%, and 6810 � 1240 for PAM 20%, inLPS with respect to non-stimulated values, which were 4657 � 1321 for� 726 for PAM control, 3410 � 841 for PAM 5%, and 3962 � 490 for
er of values corresponding to number of animals, and each value represents
0.05, # P � 0.01, and ** P � 0.001 versus control NPAM. †† P � 0.01,
contro
3 � 93 � 19 � 15 � 24 � 1
9 � 3
9 � 17 � 29 � 7
9 � 31 � 80 � 2
lysacc
t to nocontroA and, 3003numb
. � P �
A. The 5% and 20% supplementation decreased lipid per-

oPnmCim
0r
D
otifctmwsilcllr
lrfiGNlsloapstaPlahpoapce
idcpta
Flm(EsaP�rt
773C. Alvarado et al. / Nutrition 22 (2006) 767–777
xidation in PAM (P � 0.001) and NPAM (P � 0.004 and� 0.005, respectively). Concerning oxidative damage to
DNA (Fig. 3B), in the PAM group, both kinds of supple-entation strikingly decreased control values (P � 0.001).oncerning NPAM, significant differences were found after
ngestion of antioxidants, with lower levels in the supple-
ig. 1. Enzymatic and non-enzymatic antioxidant defences of peritonealeukocytes from NPAM and PAM receiving the control diet, 5% supple-entation, or 20% supplementation: (A) SOD (arbitrary U per 106 cells),
B) GPx activity (mU per 106 cells), and (C) GSH levels (nmol/106 cells).ach column represents the mean � SD of the number of values corre-ponding to the number of animals, and each value is the mean of duplicatessays. *P � 0.05, **P � 0.01, and ***P � 0.001 versus control NPAM andAM; aP � 0.05 and cP � 0.001 versus control NPAM; #P � 0.05 and ##P
0.01 versus 5% supplementation. GPx, glutathione peroxidase; GSH,educed glutathione; NPAM, non-prematurely aging mice; PAM, prema-urely aging mice; SOD, superoxide dismutase activity; U, units.
ented group versus the non-supplemented group (P � o
.005 and P � 0.001 for 5% and 20% supplementation,espectively).
iscussion
Presently there is great interest in investigating the rolef nutrition in the prevention of age-associated disorders. Inhe present work, we observed that PAM show a worsemmune function than do NPAM, which agrees with datarom previous studies of our laboratory [17,16,21], and isharacteristic for chronologically older subjects [1]. Thus,he results obtained in the present study show an increasedacrophage adherence in PAM compared with NPAM,hich is a process of immune cells during aging [7]. This
uggests an impaired capacity of these cells to move to thenfectious focus. Chemotaxis capacity of macrophages andymphocytes, ingestion capacity of macrophages (phago-ytic index), their digestion activity (measured by intracel-ular levels of superoxide anion), and NK activity wereower in PAM than in NPAM, indicating a worse immuneesponsiveness of the former.
We also analyzed redox status and oxidative damage toipids and nDNA in leukocytes from the peritoneum. Ouresults have demonstrated for the first time that leukocytesrom middle-aged PAM show higher levels of oxidants andnflammatory compounds (extracellular superoxide, GSSG,SSG/GSH ratio, TNF-�, and PGE2) than those found inPAM, in addition to decreased antioxidant defences (GSH
evels and SOD and GPx activities). Therefore, PAMhowed an imbalance between oxidants and antioxidants,eading to an oxidative stress situation that may inducexidative damage through reaction with biomolecules suchs lipids and DNA, which is a characteristic of aging. Wereviously measured lipid and nDNA oxidative damage ashown by MDA and 8-oxodG levels, respectively, two ofhe most widely studied oxidative lesions that increase withge [8,25] and found higher levels of MDA and 8-oxodG inAM than in NPAM. In a recent study we observed that
evels of 8-oxodG in peritoneal leukocytes increase withging [8]. Because oxidative stress may lead to loss ofomeostasis in immune cells, our present results could ex-lain the immunosenescence exhibited by the PAM. More-ver, in previous work we demonstrated that the PAM haveshorter lifespan than do NPAM [16,17]; therefore, the
arameters of oxidative stress and damage studied hereould be used as biomarkers of aging and could estimate lifexpectancy.
Nutrition has great importance for the maintenance ofmmune functioning [26]. However, there is a scarcity ofata on the effect of supplementation of the diet with aombination of antioxidant compounds, even though it ap-ears to be more effective than administration of each an-ioxidant separately [13]. In the present study, ingestion ofdiet supplemented with two different doses, 5% and 20%,
f biscuits enriched with nutritional amounts of several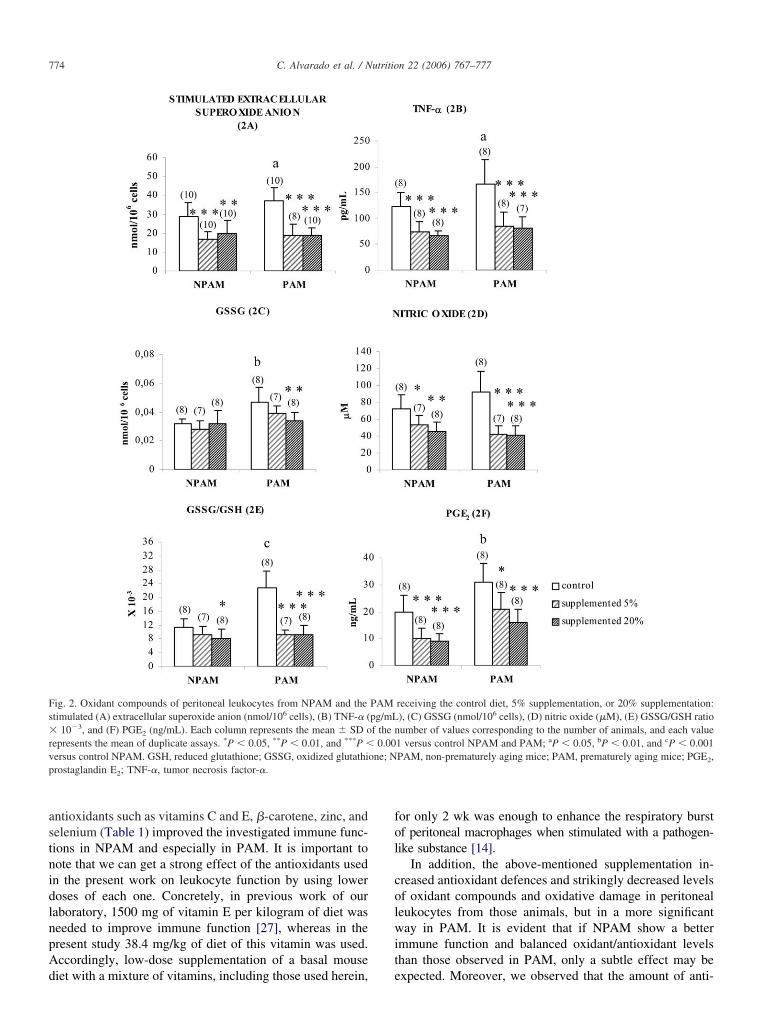
astnidlnpAd
fol
colwit
Fs�rvp
774 C. Alvarado et al. / Nutrition 22 (2006) 767–777
ntioxidants such as vitamins C and E, �-carotene, zinc, andelenium (Table 1) improved the investigated immune func-ions in NPAM and especially in PAM. It is important toote that we can get a strong effect of the antioxidants usedn the present work on leukocyte function by using loweroses of each one. Concretely, in previous work of ouraboratory, 1500 mg of vitamin E per kilogram of diet waseeded to improve immune function [27], whereas in theresent study 38.4 mg/kg of diet of this vitamin was used.ccordingly, low-dose supplementation of a basal mouse
ig. 2. Oxidant compounds of peritoneal leukocytes from NPAM and thetimulated (A) extracellular superoxide anion (nmol/106 cells), (B) TNF-� (
10�3, and (F) PGE2 (ng/mL). Each column represents the mean � SD oepresents the mean of duplicate assays. *P � 0.05, **P � 0.01, and ***P �ersus control NPAM. GSH, reduced glutathione; GSSG, oxidized glutathirostaglandin E2; TNF-�, tumor necrosis factor-�.
iet with a mixture of vitamins, including those used herein, e
or only 2 wk was enough to enhance the respiratory burstf peritoneal macrophages when stimulated with a pathogen-ike substance [14].
In addition, the above-mentioned supplementation in-reased antioxidant defences and strikingly decreased levelsf oxidant compounds and oxidative damage in peritonealeukocytes from those animals, but in a more significantay in PAM. It is evident that if NPAM show a better
mmune function and balanced oxidant/antioxidant levelshan those observed in PAM, only a subtle effect may be
receiving the control diet, 5% supplementation, or 20% supplementation:), (C) GSSG (nmol/106 cells), (D) nitric oxide (�M), (E) GSSG/GSH ratioumber of values corresponding to the number of animals, and each value
1 versus control NPAM and PAM; aP � 0.05, bP � 0.01, and cP � 0.001PAM, non-prematurely aging mice; PAM, prematurely aging mice; PGE2,
PAMpg/mLf the n
0.00one; N
xpected. Moreover, we observed that the amount of anti-

oltsmtf
pifpmvwSeaitimatsdaotcmfIT
arc
Hpooafapabppccatamrama
irmiAtdtmtc[rw
FscPN
775C. Alvarado et al. / Nutrition 22 (2006) 767–777
xidants needed to improve several immune functions isarger in aged than in adult or young mice [22]. In general,he functions studied in PAM after supplementation becomeimilar or better than those observed in NPAM. This im-unomodulator effect of the antioxidants is a general result
hat is more evident in subjects showing an initial worseunction [28].
All the antioxidants included in the biscuits used in theresent work have shown an enhancing effect on severalmmune functions. Thus, ascorbic acid improves immuneunctions such as those studied here [29,30]. Concretely,roduction of ROS, including O2; � and H2O2, by peritonealacrophages was increased after treatment of mice with
itamin C, vitamin E, vitamins C and E, or �-carotenes [30],hich is in agreement with our previous results [21,27].upplementation with vitamin E has also been shown tonhance cell-mediated and humoral immune responses inged experimental animals [27,31]. Similarly, �-carotenencreases NK activity, IL-2 receptors, and lymphoprolifera-ion [32,33]. Zinc and selenium also play an important rolen maintaining metabolic homeostasis and an adequate im-une function, and their deficiency is related to many age-
ssociated diseases [34–36]. Thus, zinc has a crucial role inhe maintenance of an adequate immune response and re-istance to infection during aging [36]. Klecha et al. [37]emonstrated in humans and rodents that zinc deficiencyffects T- and B-lymphocyte functions. Several studies havebserved that supranutritional amounts of selenium are ableo stimulate cytotoxic activity of NK cells [38]. An in-reased capacity to generate cytotoxic lymphocytes andacrophages that could eliminate tumor cells has been
ound to be related to increased expression of high-affinityL-2 receptor produced by selenium [39], thus modulating-cell–mediated responses generated by IL-2.
In relation to the effect of antioxidants on maintaining andequate redox balance, vitamin E can decrease the age-elated increase in PGE2 release from macrophages, which
ig. 3. Lipids and nuclear DNA oxidative damage of peritoneal leukocytesupplementation: (A) MDA levels (nmol/106 cells) and (B) 8-oxodG/dG raorresponding to the number of animals, and each value represents the meAM; aP � 0.05 and bP � 0.01 versus control NPAM. 8-oxodG, 8-oxo,PAM, non-prematurely aging mice; PAM, prematurely aging mice.
ontributes to decreased T-cell–mediated function [40]. �
ayek et al. [41] first showed that LPS-stimulated macro-hages from old mice produced more PGE2 than thosebserved in young mice because of the higher cyclo-xygenase-2 activity exhibited by the old. Moreover, NO,n important redox mediator that modulates several immuneunctions, such as lymphocyte chemotaxis [42], is decreasedfter the antioxidant diet supplementation used in theresent work, as is TNF-�, another proinflammatory medi-tor that increases during aging [21]. Vitamin E has alsoeen described as a very efficient compound to prevent lipideroxidation, as has vitamin C [43]. Levels of 8-oxodG ineritoneal leukocytes, an oxidative stress marker that in-reases during aging [8], showed a marked decrease in thoseells after diet supplementation with the combination ofntioxidants in PAM and NPAM with both doses adminis-ered. Similar results have been observed after vitamin Cnd E supplementation in relation to mitochondrial DNA inice [44]. With respect to �-carotene, Palozza et al. [45]
eported that its capacity to suppress cyclo-oxygenase-2nd, hence, PGE2 production may represent a molecularechanism by which this compound acts as an antitumor
gent in colon carcinogenesis.Zinc also prevents free radical formation [46], which
s very important for correct functioning of the immuneesponse [47]. Thus, supplementation with the recom-ended daily amount of zinc decreases the incidence of
nfections [46,47] and increases the rate of survival [46].nother study has suggested that selenium supplementa-
ion improves glutathione levels and decreases the oxi-ative stress measured by MDA levels in plasma andissue of experimental animals [48]. Selenium supple-entation also increases some antioxidant enzymatic ac-
ivities such as GPx and thioredoxin reductase, whichontributes to restore endogenous antioxidant systems49]. In addition, an increase in SOD and GPx has beeneported by Sánchez et al. [30] after treatment of miceith vitamin C, vitamin E, vitamins C and E, and
NPAM and PAM receiving the control diet, 5% supplementation, or 20%10�5). Each column represents the mean � SD of the number of values
duplicate assays. **P � 0.01 and ***P � 0.001 versus control NPAM andhidro-2=-deoxyguanosine; dG, deoxyguanosine; MDA, malondialdehyde;
fromtio (�
an of7,8-dy
-carotenes.

s[aisswrlchaiatbottcfhc
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
776 C. Alvarado et al. / Nutrition 22 (2006) 767–777
Despite the controversial results regarding antioxidantupplementation and lifespan presented in the literature15,50,51], we recently observed that ingestion of thiolicntioxidants, namely N-acetylcysteine and thioproline,ncreased the lifespan of mice in our model of prematureenescence (unpublished data). Moreover, we alsohowed that antioxidant supplementation of old miceith the same biscuits used herein improves function and
estores redox balance in leukocytes from peritoneum,eading to increased longevity (unpublished data). Inonclusion, because the immune function is a marker ofealth [52] and several of the immune parameters studiedre predictors of longevity [5,6,16,17], our data stronglyndicate, on the one hand, the importance of maintainingproper regulation of redox homeostasis in immune cells
o preserve their functions and, on the other hand, that theiscuits enriched with nutritional doses of several anti-xidants used in the present work appear to be a func-ional food that allows improvement of leukocyte func-ion through restoration of the redox balance of theseells. Therefore, consumption of antioxidant-enrichedood, included as a part of a regular diet, might improveealth, especially in animals with premature aging and, inonsequence, could increase their life expectancy.
eferences
[1] Pawelec G, Barnett Y, Forsey R, Frasca D, Globerson A, Mc Leod J,et al. T cells and aging. Front Biosci 2002;7:d1056–183.
[2] Stout RD, Suttles J. Immunosenescence and macrophage functionalplasticity: dysregulation of macrophage function by age-associatedmicroenvironmental changes. Immunol Rev 2005;205:60–71.
[3] Chandra RK. Impact of nutritional status and nutrient supplements onimmune responses and incidence of infection in older individuals.Ageing Res Rev 2004;3:91–104.
[4] Vasto S, Caruso C. Immunity and ageing: a new journal looking atageing from an immunological point of view. Immnun Ageing 2004;1:1–4.
[5] Franceschi C, Monti D, Sansoni P, Cossarizza A. The immunology ofexceptional individuals: the lesson of centenarians. Immunol Today1995;16:12–16.
[6] Puerto M, Guayerbas N, Álvarez P, De la Fuente M. Modulation ofneuropeptide Y and norepinephrine on several leucocyte functions inadult, old and very old mice. J Neuroimmunol 2005;165:33–40.
[7] De la Fuente M. Effects of antioxidants on immune system ageing.Eur J Clin Nutr 2002;56:5–8.
[8] De la Fuente M, Hernanz A, Guayerbas N, Álvarez P, Alvarado C.Changes with age in peritoneal macrophage functions. Implication ofleukocytes in the oxidative stress of senescence. Cell Mol Biol 2004;50(suppl):OL683–90.
[9] Albers R, Antoine JM, Bourdet-Sicard R, Calder PC, Gleeson M,Lesourd B, et al. Markers to measure immunomodulation in humannutrition intervention studies. Br J Nutr 2005;94:452–81.
10] Hughes DA. Effects of dietary antioxidants on the immune functionof middle-aged adults. Proc Nutr Soc 1999;58:79–84.
11] Watson RR, Zibadi S, Vazquez R, Larson D. Nutritional regulation ofimmunosenescence for heart health. J Nutr Biochem 2005;16:85–7.
12] Mc Donald RB. Some considerations for the development of diets formature rodents used in long term investigations. J Nutr 1997;127:
847S–50.13] Palozza P, Krinsky NI. �-Carotene and �-tocopherol are synergisticantioxidants. Arch Biochem Biophys 1992;297:184–7.
14] Jakus J, Kriska T, Vanyúr R. Effect of multivitamins in an efferves-cent preparation on the respiratory burst of peritoneal macrophages inmice. Br J Nutr 2002;87:501–8.
15] Hsieh CC, Lin BF. Opposite effects of low and high dose supple-mentation of vitamin E on survival of MRL/lpr mice. Nutrition2005;21:940–8.
16] Guayerbas N, Puerto M, Víctor VM, Miquel J, De la Fuente M.Leucocyte function and life span in a murine model of prematureimmunosenescence. Exp Gerontol 2002;37:249–56.
17] Guayerbas N, De la Fuente M. An impairment of phagocytic functionis linked to a shorter life span in two strains of prematurely agingmice. Dev Comp Immunol 2003;27:339–50.
18] Viveros MP, Fernández B, Guayerbas N, De la Fuente M. Behavioralcharacterization of a mouse model of premature inmunosenescence.J Neuroimmunol 2001;114:80–8.
19] De la Fuente M, Hernanz A, Medina S, Guayerbas N, Fernández B,Viveros MP. Characterization of monoaminergic systems in brainregions of prematurely aging mice. Neurochem Int 2003;43:165–72.
20] Reeves PG, Nielsen FH, Fahey GC Jr. AIN-93 Purified diets forlaboratory rodents: final report of the American Institute of Nutritionad hoc writing committee on the reformulation of the AIN-76Arodent diet. J Nutr 1993;123:1939–51.
21] Alvarado C, Álvarez P, Jiménez L, De la Fuente M. Improvement ofleukocyte functions in prematurely aging mice after a 5-week inges-tion of a diet supplemented with biscuits enriched in antioxidants.Antioxid Redox Signal 2005;7:1203–10.
22] De la Fuente M, Miquel J, Catalan MP, Víctor VM, Guayerbas N.The amount of thiolic antioxidant ingestion needed to improve sev-eral immune functions is higher in aged than in adult mice. Free RadicRes 2002;36:119–26.
23] Lawrence RA, Burck RF. Glutathione peroxidase activity in seleni-um-deficient rat liver. Biochem Biophys Res Commun1976;71:952–8.
24] Chirico S, Smith C, Marchant C, Mithison MJ, Halliwell B. Lipidperoxidation in hyperlipemic patients. A study of plasma using anHPLC-based thiobarbituric acid test. Free Radic Res Commun 1993;19:51–57.
25] Traverso N, Menini S, Maineri EP, Patriarca S, Odetti P, CottalassoD, et al. Malondialdehyde, a lipoperoxidation-derived aldehyde, canbring about secondary oxidative damage to proteins. J Gerontol ABiol Sci Med Sci 2004;59:B890–5.
26] Marcos A, Nova E, Montero A. Changes in the immune system areconditioned by nutrition. Eur J Clin Nutr 2003;57:S66–9.
27] De la Fuente M, Carazo M, Correa R, Del Río M. Changes inmacrophage and lymphocyte functions in guinea-pigs after differentamounts of vitamin E ingestion. Br J Nutr 2000;84:25–9.
28] De la Fuente M, Víctor VM. Anti-oxidants as modulators of immunefunction. Immunol Cell Biol 2000;8:49–54.
29] De la Fuente M, Víctor VM. Ascorbic acid and N-acetylcysteineimprove in vitro the function of lymphocytes from mice with endo-toxin-induced oxidative stress. Free Radic Res 2001;35:73–84.
30] Sánchez GM, Re L, Giuliani A, Núñez-Sellés AJ, Davison GP,León-Fernández OS. Protective effects of Mangifera indica L. extract,mangiferin and selected antioxidants against TPA-induced biomol-ecules oxidation and peritoneal macrophage activation in mice. Phar-macol Res 2000;42:565–70.
31] Adolfsson O, Huber BT, Meydani SN. Vitamin E-enhanced IL-2production in old mice: naive but not memory T cells show increasedcell division cycling and IL-2-producing capacity. J Immunol 2001;167:3809–17.
32] Chew BP, Park JS. Carotenoid action on the immune response. J Nutr2004;134:257S–61.
33] Stahl W, Sies H. Bioactivity and protective effects of natural carote-
noids. Biochim Biophys Acta 2005;1740:101–7.
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
777C. Alvarado et al. / Nutrition 22 (2006) 767–777
34] Navarro-Alarcon M, De LA Serrana HL, Perer Valero V, López-Martínez C. Serum selenium levels as indicators of body status incancer patients and their relationship with other nutritional and bio-chemical markers. Sci Total Environ 1998;212:195–202.
35] Shankar AH, Prasad AS. Zinc and immune function: the biologicalbasis of altered resistance to infection. Am J Clin Nutr 1998;68(suppl):447S–63.
36] Mocchegiani E, Muzzioli M, Giaconi R. Zinc and immunoresistanceto infection in aging: new biological tools. Trends Pharmacol Sci2000;21:205–8.
37] Klecha AJ, Salgueiro J, Wald M, Boccio J, Zubillaga M, LeonardiNM, et al. In vivo iron and zinc deficiency diminished T- andB-selective mitogen stimulation of murine lymphoid cells throughprotein kinase C-mediated mechanism. Biol Trace Elem Res 2005;104:173–84.
38] Kiremidjian-Schumacher L, Roy M, Wishe HI, Cohen MW, StotzkyG. Supplementation with selenium augments the functions of naturalkiller and lymphokine-activated killer cells. Biol Trace Elem Res1996;52:227–39.
39] Roy M, Kiremidjian-Schumacher L, Wishe HI, Cohen MW, StotzkyG. Supplementation with selenium and human immune cell functions.Effect of lymphocyte proliferation and interleukin 2 receptor expres-sion Biol Trace Elem Res 1994;41:103–13.
40] Beharka AA, Wu D, Serafini M, Meydani SN. Mechanism of vitaminE inhibition of cyclooxygenase activity in macrophages from oldmice: role of peroxynitrite. Free Radic Biol Med 2002;32:503–11.
41] Hayek MG, Mura C, Wu D, Beharka AA, Han SN, Paulson KE, et al.Enhanced expression of inducible cyclooxygenase with age in murinemacrophages. J Immunol 1997;159:2445–51.
42] Medina S, Del Río M, Hernanz A, Guaza C, De la Fuente M. Nitricoxide released by accessory cells mediates the gastrin-releasing pep-tide effect on murine lymphocyte chemotaxis. Regul Pept 2005;131:
46–53.43] Polidori MC, Mecocci P, Levine M, Frei B. Short-term and long-termvitamin C supplementation in humans dose-dependently increases theresistance of plasma to ex vivo lipid peroxidation. Arch BiochemBiophys 2004;423:109–15.
44] De la Asunción JG, Del Olmo LM, Gómez-Cambronero MG, SastreJ, Pallardó FV, Viña J. AZT induces oxidative damage to cardiacmitochondria: protective effect of vitamins C and E. Life Sci 2004;76:47–56.
45] Palozza P, Serini S, Maggiano N, Triangali G, Navarra P, RanelletiFO, Calviello G. �-Carotene downregulates the steady-state andheregulin-alpha-induced COX-2 pathways in colon cancer cells. JNutr 2005;135:129–36.
46] Mocchegiani E, Muzzioli M, Cipriano C, Giacconi R. Zinc, T-cellpathways, aging: role of metallothioneins. Mech Ageing Dev 1998;106:183–204.
47] Wellinghausen N, Rink L. The significance of zinc in leukocytebiology. J Leukocyte Biol 1998;64:571–7.
48] Sarada SKS, Sairam M, Dipti B, Anju B, Pauline T, Kain AK, et al.Role of selenium in reducing hypoxia-induced oxidative stress: an invivo study. Biomed Pharmacother 2002;56:173–8.
49] Tapiero H, Townsend DM, Tew KD. The antioxidant role of seleniumand seleno-compounds. Biomed Pharmacother 2003;57:134–44.
50] Bezlepkin VG, Sirota NP, Gaziev AI. The prolongation of survival inmice by dietary antioxidants depends on their age by the start offeeding this diet. Mech Ageing Dev 1996;92:227–34.
51] Navarro A, Gomez C, Sanchez-Pino MJ, Gonzalez H, Bandez MJ,Boveris AD, Boveris A. Vitamin E at high doses improves survival,neurological performance, and brain mitochondrial function in agingmale mice. Am J Physiol Regul Integr Comp Physiol 2005;289:R1392–9.
52] Wayne SJ, Rhyne RL, Garry PJ, Goodwin JS. Cell-mediated immu-nity as a predictor of morbidity and mortality in subjects over 60. J
Gerontol 1990;45:45–8.
Related Documents











