This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Diatom induction of reproductive failure in copepods: The effect of PUAs versus nonvolatile oxylipins
A. Barreiro a,⁎, Y. Carotenuto b, N. Lamari c, F. Esposito b, G. D'Ippolito c, A. Fontana c, G. Romano b, A. Ianora b,A. Miralto b, C. Guisande a
a Facultad de Ciencias, Universidad de Vigo, Lagoas-Marcosende, 36310 Vigo, Spainb Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italyc Instituto di Chimica Biomolecolare (ICB) del CNR, Via Campi Flegrei 34, 80078, Pozzuoli, Napoli, Italy
a b s t r a c ta r t i c l e i n f o
Article history:Received 30 November 2010Accepted 11 March 2011
Keywords:Chemical defenceCopepodsDiatomsEgg hatchingOxylipinsPUAs
Several species and strains of diatoms were tested for their effects on egg production, hatching success, faecalpellet production, ingestion rate and food selection in the copepod Temora stylifera. Two of the diatom speciestested were well known polyunsaturated aldehyde (PUA) producers: Skeletonema marinoi (SM) andThalassiosira rotula (TR1), while the other species tested, Skeletonema pseudocostatum, was a non-PUAproducing species. Another T. rotula strain (TR2) was also tested since previous studies had reportedcontradictory results, due either to absence of PUA production or production in small amounts. Our resultsshowed strong inhibitory effects on reproductive parameters of copepods with all diatom species tested.Although we confirm that S. pseudocostaum did not produce PUAs, this diatom produced large quantities ofother oxylipins such as (5Z,8Z,11Z,13E,15S,17Z)-15-hydroxy-5,8,11,13,17-eicosapentaenoic acid (15S-HEPE)and 13,14-13R-hydroxy-14S,15S-trans-epoxyeicosa-5Z,8Z,11Z,17Z-tetraenoic acid (13,14-HEpETE) and 15-oxo-5Z,9E,11E,13E-pentadecatetraenoic acid (15-oxoacid), all of which have already been found in S. marinoi(Fontana et al., 2007a) and Pseudonitzschia delicatissimia (d'Ippolito et al., 2009). Another unidentifiedoxylipin was also present in high quantities in LC–MS profiles. Some of these oxygenated fatty acid derivativeswere also found, together with PUAs, in TR1 and in TR2. Both PUA-producing and non-producing diatomscaused negative effects on hatching success with induction of apoptosis in newly hatched nauplii. Thissuggests that negative diatom effects can also depend on the production of non volatile oxylipins. Copepodfeeding behaviour was found to be non-selective when each diatom species was offered together with the nontoxic dinoflagellate Prorocentrum minimum suggesting that grazers are not aware of the toxicity of their food.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Several diatom species have been shown to induce reproductivefailure in copepods under laboratory conditions by reducing egg pro-duction, hatching success, and naupliar survival to adulthood (reviewedby Ianora and Miralto (2010)). Inhibition of copepod egg hatchingsuccess has been shown to be diatom density-dependent (Chaudronet al., 1996): with decreasing diatom concentrations, deleterious effectson hatching diminish and these effects take longer to induce (Ban et al.,1997). Mixed diets dilute but do not delete the negative effects of
diatoms on copepod reproduction (Kang and Poulet, 2000; Turner et al.,2001). Diatoms not only interfere with egg maturation but also inducestrong developmental aberrations in those nauplii that do develop tohatching. Teratogenic (abnormal) nauplii show a variety of birth defectssuch as asymmetrical bodies and malformed or reduced number ofswimming and feeding appendages (Ianora et al., 2004) and usually diesoon after birth because they are unable to swim or feed properly.
Pioneer studies (Miralto et al., 1999) led to the characterization ofshort-chained polyunsaturated aldehydes (hereafter PUAs) as themainmolecules responsible for the toxic effects observed in copepods. Thesemolecules are derived from lipoxygenase oxidation of polyunsaturatedfatty acids (PUFAs), mainly C16 and C20, liberated from chloroplastgalactolipids and membrane phospholipids after membrane damage ordisruption (Pohnert et al., 2002; d'Ippolito et al. 2004; Reviewed byPohner, (2005) and Fontana et al. (2007a)). Wichard et al. (2005)explored potential PUA production among 51 diatom isolates, reportingthat 30% are PUAproducers. Butwhether PUA production iswidespreadover different taxonomic groups of diatoms, from the genera to thestrain level, has yet to be clarified. Furthermore, some differences may
Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19
Abbreviations: PUA, polyunsaturated aldehyde; PUFA, polyunsaturated fatty acids;FAH, fatty acid hydroperoxide; LOX, lipoxygenase; TR1, Thalassiosira rotula strain CCMP1647; TR2, Thalassiosira rotula strain CCMP 1018; SM, Skeletonema marinoi; SPC,Skeletonema pseudocostatum; PRO, Prorocentrum minimum; FSW, filtrated sea water.⁎ Corresponding author at: Current address: Department of Ecology and Evolution-
ary Biology, Corson Hall, Cornell University, 14850 Ithaca, NY, USA.E-mail address: [email protected] (A. Barreiro).
0022-0981/$ – see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.jembe.2011.03.007
Contents lists available at ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r.com/ locate / jembe

Author's personal copy
not be due to genetic differentiation, but to environmental conditions orparticular physiological status of the cells (Ribalet et al., 2007).
Recently, an entirely new group of other oxygenated derivatives ofPUFAs (oxylipins) was identified in marine diatoms that were able(Thalassiosira rotula and Skeletonema marinoi) or unable (Chaetocerossocialis, Chaetoceros affinis, and Pseudonitzschia delicatissima) to pro-duce PUAs (d'Ippolito et al., 2005; Fontana et al., 2007b; d'Ippolitoet al. 2009). The presence of these compounds (here referred as non-volatile oxylipins) demonstrated that different oxygenase pathwaysare active in marine diatoms and that the production of PUAs iscomplemented by species-specific synthesis of other products derivedfrom the enzymatic oxidation of PUFAs. The process is mediated bymassive synthesis of fatty acid hydroperoxides (FAHs) and highlyreactive oxygen species (hROS). Both FAHs and non-volatile oxylipinsinduce reproductive failure in copepods even though the latter com-pounds seem to be less active (Fontana et al., 2007b).
PUA synthesis is therefore not sufficient to assess the potentialdeleterious effects of diatoms on copepod reproduction since com-pounds other than PUAs can also be involved in the defence againstpotential predators (Fontana et al., 2007b; d'Ippolito et al. 2009). Herewe continue to explore this possibility in the copepod Temora stylifera,a dominant copepod species in the Mediterranean Sea, by testing thepotential effect of a known PUA (Skeletonema marinoi) and non-PUA(Skeletonema pseudocostatum) producing species on egg production,egg hatching success, female survival and faecal pellet production. S.pseudocostatum is considered a non-PUA producing species accordingto Wichard et al. (2005). But it was unknown whether the strain usedin our experiment produced PUAs or not, since our strain was isolatedfrom the Gulf of Naples in 2002. We also tested the potential effect onthose reproductive parameters when T. stylifera was fed with twodifferent strains of the diatom T. rotula: Strain CCMP 1647, known as aPUA producer, and Strain CCMP 1018, which did not produce PUAs inPohnert et al. (2002), but showed minimal PUA production inWichard et al. (2005). Finally, we studied the feeding behaviour ofT. stylifera when offered with a mixture of each of these diatoms withthe non-oxylipin producing dinoflagellate, Prorocentrum minimum,to better understand the response of copepods under more naturalmixed diet conditions.
2. Methods
2.1. Phytoplankton cultures
The diatom species S. pseudocostatum (Strain SZN B77, hereafterSPC), S. marinoi (Strain SZN B118, hereafter SM), T. rotula Strain CCMP1647 (hereafter TR1), Strain CCMP 1018 (hereafter TR2) and thedinoflagellate P. minimum, (hereafter PRO) were cultured in 2 l jarswith 1 l of 0.22 μm filtered and autoclaved seawater enriched with f/2medium (Leftley et al., 1987) at 20 °C under 12 h:12 h light:dark cycle.Cultures were diluted daily by replacing ≈25% of the culture withfresh media. Cell carbon content of SM, TR1 and 2 and PRO wasconsidered to be the same as in Carotenuto et al. (2002). Cell carboncontent of SPCwas not determined but was assumed to be the same asSM due to its similar size. This assumption is supported by similarvalues measured in SPC cells isolated from field samples collected inthe Gulf of Naples (~20 pg C cell−1, D. Sarno, pers. comm.).
2.2. Zooplankton collection
Samples were collected weekly during 4 October to 30 November2005 at a fixed station of the Gulf of Naples (South Tyrrhenian, Italy)by towing a 250 μm mesh net with a non-filtering cod obliquely from≈50 m to the surface. Samples were poured in an insulated box andtransported to the laboratory within 1 h of collection.
2.3. Reproduction experiments
Healthy females and males of T. stylifera were sorted and trans-ferred to crystallising dishes containing 100 ml of 50 μm filtered sea-water (N=10–15, Table 1) and placed at≈20 °C. Each dish containedan individual couple to ensure re-mating and production of viableeggs (Ianora et al., 1989). Animals were maintained in this foodsuspension during 24 h. The next day, animals were transferred tonew food suspensions with each of the target algae offered as the onlyfood. The five microalgae tested were the diatoms SPC, SM, TR1, TR2and the control dinoflagellate PRO, (Table 1). Food suspensions wererenewed daily. Experiments were terminated after 12–15 days. PROwas used as a control diet since this alga was shown to induce highand constant egg hatching success in T. stylifera (Turner et al., 2001)and did not produce PUAs or other oxylipins (Fontana et al., 2007a).Cell concentrations of each alga were adjusted to keep the samecarbon concentration between treatments (0.98–1.24 μg C ml−1,Table 1) in 100 ml of food suspension. This carbon concentrationwas food saturating and cell abundances were close to natural bloomconditions. Faecal pellet production, egg production and crumpledegg membranes due to cannibalism were counted daily under abinocular inverted microscope. Female survival was estimated as the% of accumulated survival of the initial population. Hatching wasestimated daily by maintaining egg production containers for anadditional 48 h at ≈20 °C. Eggs were then fixed with 25 ml of 95%ethanol. Hatching success was calculated as the percentage of naupliiwith regards to egg production excluding eggs with crumpled mem-branes. Groups of treatments were performed on different datesduring the period from 4 October to 14 December.
2.4. Fluorescence labelling and confocal microscopy
Newly-hatched nauplius 1 and 2 embryos produced by females fedPRO, SKEor SPCwerewashed three times in FSWbeforefixation in 2–4%formaldehyde for 24 h. Fixed embryoswere then rinsed several times inPBS and incubated for 24 h in 250 μl of 0.5 U ml−1 chitinase enzyme(EC3.2.1.14; Sigma-Aldrich)dissolved in50 cmmol l−1 citrate buffer, pH6, at 25 °C, to permeabilize the chitinouswall. After rinsing several timesin PBS, embryoswere incubated for 10 min in 0.1% Triton X-100 at roomtemperature, rinsed in PBS containing 1% BSA, and further incubated for90 min in TUNEL (terminal-deoxynucleotidyl-transferase-mediateddUTP Nick End Labelling; Roche Diagnostics GmbH, Mannheim,
Table 1Protocol details of the experiments. SPC (Skeletonema pseudocostatum), SM (Skeleto-nema marinoi), TR1 (Thalassiosira rotula Strain CCMP 1647), TR2 (Thalassiosira rotulaStrain CCMP 1018), PRO (Prorocentrum minimum).
Reproductionexperiment
Cellabundance(cell ml−1)
Cell carboncontent(pg C cell−1)
Total carboncontent(μg C ml−1)
N°replicates
Days
SPC 60×103 20.7a 1.24 15 15SM 60×103 20.7b 1.24 15 14TR1 8×103 121.9b 0.98 15 15TR2 8×103 121.9c 0.98 15 15PRO 6×103 177.1b 1.06 10 12
Ingestionexperiment
Mean cellabundance(cell ml−1)
PRO–SPC 1095 PRO 0.19 PRO 3 feeding 48700 SPC 0.18 SPC 2 controls
PRO–TR1 874 PRO 0.15 PRO 3 feeding 4800 TR1 0.10 TR1 2 controls
PRO–TR2 1045 PRO 0.18 PRO 4 feeding 41212 TR2 0.15 TR2 2 controls
a Assumed to be the same as SM (see Methods section).b From Carotenuto et al., 2002.c Assumed to be the same as TR1.
14 A. Barreiro et al. / Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19
Author's personal copy
Germany) solution, at 37 °C. Whole-mount copepod embryos wereobserved with a Zeiss Axio-Imager M1microscope equipped with a 20´objective and GFP filter to detect TUNEL green fluorescence.
2.5. Grazing experiments
A mixture of PRO with each diatom was offered as food toT. stylifera in 25 ml bottles (0.10–0.18 μg C ml−1, Table 1). Foodconcentrations were much lower in the grazing experiments (belowsaturation levels) to better appreciate the removal of diatom cells byfemales. Three adult females were placed in each of three or four25 ml bottles whereas two 25 ml bottles without copepods were usedas a control for algal growth. Bottles were maintained on a rotatingwheel (1 rpm to ensure continuous algal suspension) at 20 °C under12 h:12 h light:dark cycle. Food suspensions were renewed daily and
experiments were terminated after 4 days. If any of the animals died,the sample was not considered for the calculation of ingestion rates.Two replicates of a sample of 1 ml for cell counting was taken fromeach bottle, fixed with 300 μl of Lugol iodide solution and placed into2 ml Eppendorf tubes. Triplicates of 1 ml samples for cell countingwere taken from initial cell suspensions, fixed with 300 μl of Lugoliodide solution and placed into 2 ml Eppendorf tubes. Samples werecounted under a direct microscope in 1 ml Sedgewick–Rafter cham-bers. Ingestion values were calculated following Frost's (1972)equations. Selection of food was calculated with Manly's α index(Manly, 1974).
2.6. Chemical analysis
10 l of cultures of each diatom species were centrifuged 10 min at1500 g at 4 °C. The pelletwas resuspended in distilledwater, sonicatedfor 1 min and, after 10 min, suspended in acetone (1 ml/g cell pellet).The hydro-acetone suspension was sonicated for 1 min and thencentrifuged at 2000 g for 5 min at 5 °C. The resulting pellet wasresuspended in H2O/acetone 1:1 (2 ml/g cell pellet), sonicated for1 min and centrifuged at 2000 g for 5 min. The supernatants werecombined prior to extraction with an equal volume of CH2Cl2. Theorganic layer was dried over dry Na2SO4, filtrated and evaporated atreduced pressure. A fraction (2/3 of the extract) of this material wassubmitted to PUAs analysis byGCMS in agreementwith d'Ippolito et al.(2002a). After addition of 16-hydroxy-hexadecanoic acid as aninternal standard, the remaining extractwasmethylatedwith etherealdiazomethane. The excess of organic solventwas removedat a reducedpressure and the residue dissolved inMeOH (1 μg/μl) was analysed byLCMS analysis on a Qtof-micromass spectrometer (Waters SpA, Milan,Italy) equippedwith an ESI source (positivemode) and coupledwith aWaters Alliance HPLC system (reverse phase column with a gradientfrom MeOH/H2O 70/30 to MeOH/H2O 80/20 in 15 min, followed byisocratic elution MeOH/H2O 80/20 at a flow of 1 ml/min). Content ofPUAs and non-volatile oxilipins was established by integration of areapeaks against internal standards, on an average of fourmeasurements.
3. Results
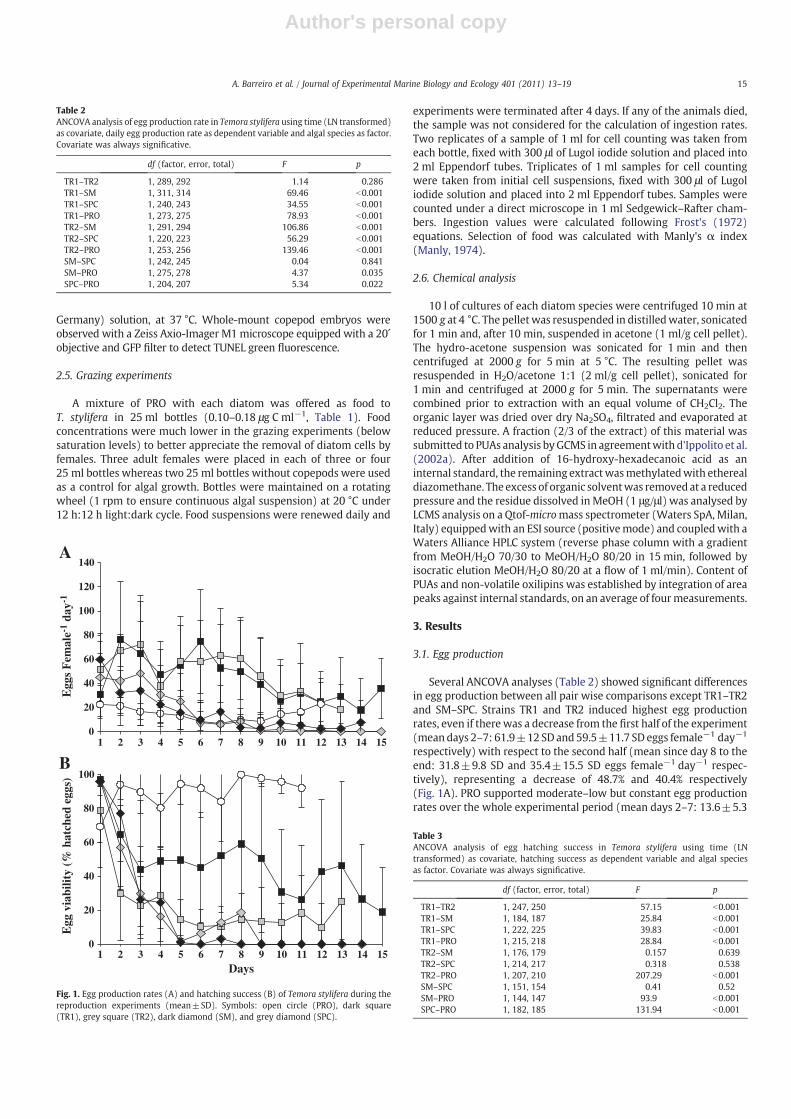
3.1. Egg production
Several ANCOVA analyses (Table 2) showed significant differencesin egg production between all pair wise comparisons except TR1–TR2and SM–SPC. Strains TR1 and TR2 induced highest egg productionrates, even if there was a decrease from the first half of the experiment(meandays 2–7: 61.9±12 SD and 59.5±11.7 SD eggs female−1 day−1
respectively) with respect to the second half (mean since day 8 to theend: 31.8±9.8 SD and 35.4±15.5 SD eggs female−1 day−1 respec-tively), representing a decrease of 48.7% and 40.4% respectively(Fig. 1A). PRO supported moderate–low but constant egg productionrates over the whole experimental period (mean days 2–7: 13.6±5.3
Table 2ANCOVA analysis of egg production rate in Temora stylifera using time (LN transformed)as covariate, daily egg production rate as dependent variable and algal species as factor.Covariate was always significative.
df (factor, error, total) F p
TR1–TR2 1, 289, 292 1.14 0.286TR1–SM 1, 311, 314 69.46 b0.001TR1–SPC 1, 240, 243 34.55 b0.001TR1–PRO 1, 273, 275 78.93 b0.001TR2–SM 1, 291, 294 106.86 b0.001TR2–SPC 1, 220, 223 56.29 b0.001TR2–PRO 1, 253, 256 139.46 b0.001SM–SPC 1, 242, 245 0.04 0.841SM–PRO 1, 275, 278 4.37 0.035SPC–PRO 1, 204, 207 5.34 0.022
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
1 2 3 4 5 6 7Days
8 9 10 11 12 13 14 15
0
20
40
60
Egg
via
bilit
y (%
hat
ched
egg
s)
80
100
120
140
0
20
40
60
80
100
A
B
Fig. 1. Egg production rates (A) and hatching success (B) of Temora stylifera during thereproduction experiments (mean±SD). Symbols: open circle (PRO), dark square(TR1), grey square (TR2), dark diamond (SM), and grey diamond (SPC).
Table 3ANCOVA analysis of egg hatching success in Temora stylifera using time (LNtransformed) as covariate, hatching success as dependent variable and algal speciesas factor. Covariate was always significative.
df (factor, error, total) F p
TR1–TR2 1, 247, 250 57.15 b0.001TR1–SM 1, 184, 187 25.84 b0.001TR1–SPC 1, 222, 225 39.83 b0.001TR1–PRO 1, 215, 218 28.84 b0.001TR2–SM 1, 176, 179 0.157 0.639TR2–SPC 1, 214, 217 0.318 0.538TR2–PRO 1, 207, 210 207.29 b0.001SM–SPC 1, 151, 154 0.41 0.52SM–PRO 1, 144, 147 93.9 b0.001SPC–PRO 1, 182, 185 131.94 b0.001
15A. Barreiro et al. / Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19

Author's personal copy
SD eggs female−1 day−1, mean as of day 8 to the end: 14.4±5.7 SD eggsfemale−1 day−1) (Fig. 1A). In contrast, SM and SPC resulted in higherinitial egg production rates that decreased sharply with time (meandays 2–7: 21.7±9.6 SD and 26.4±17.5 SD eggs female−1 day−1,respectively mean as of day 8 to the end: 4.5±2.2 SD and 1.5±2.7 SDeggs female−1 day−1, respectively), representing a decrease of 94.1%and 79.2% respectively, and the lowest egg production rates recorded inthis set of experiments (Fig. 1A).
3.2. Egg viability
Optimal hatching rates were observed on day 1 with all diets (mean87.3%±12.6 SD, Fig. 1B), but decreased thereafter with all testeddiatoms. Only PROsupportedoptimal egghatching rates throughout theexperiment (mean 89.9%±8.9 SD). Using an ANCOVA analysis, egg
viability with PRO was significantly different compared to all othertreatments (Table 3). With TR1 and TR2 diets egg hatching successdecreased slowly (mean day 2–day 7 50.7%±7.3 SD and 19.4%±8.7 SDrespectively;mean as of day 8 to the end: 37.6%±14 SD and 15.7%±5.3SD, respectively). There were statistically significant differences in eggviability between TR1 and TR2. SPC and SM induced strong egg hatchinginhibition after the first few days, with a drastic negative trend (meanday 2–day 7 20.5%±20.18 SD and 22%±29.4 SD respectively; mean asof day 8 to the end: 4.6%±9.2 SD and 0%±0 SD, respectively) reachingvalues of about 0% by day 5 (Fig. 1B). There were no differences in eggviability between TR2, SM and SPC (Table 3).
3.3. Apoptotic nauplii
88% of the nauplii that hatched after T. styliferawas fed SPC for 48 hwere TUNEL-positive, indicating apoptotic tissues and imminent death
Fig. 2. Apoptotic TUNEL-positive nauplii spawned by Temora stylifera females fedwith the oxylipin producing diatoms Skeletonema pseudocostatum (A,B) and Skeletonemamarinoi (C,D) in fluorescent and transmitted light.
Days1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Surv
ival
(%
fem
ale
surv
ival
)
0
20
40
60
80
100
Fig. 3. Female survival of Temora stylifera during reproduction experiments. Symbols:open circle (PRO), dark square (TR1), grey square (TR2), dark diamond (SM), and greydiamond (SPC).
AlgaeTR1 TR2 SM SPC
g m
g C
-1
0
2
4
6
8
10
12
14
16
18
20
µ
Fig. 4. Ingestion rates and Manly's α coefficient of food selection of Temora stylifera oneach alga during the whole feeding period (mean 4 days±SD). Symbols: black bars(PRO) and white bars (TR1, TR2 and SPC, respectively).
16 A. Barreiro et al. / Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19
Author's personal copy
(Fig. 2A,B). Similarly, females fed with the known apoptotic-inducerSM after 48 h spawned 100% TUNEL-positive nauplii (Fig. 2C,D). NoTUNEL-positive areas were observed in nauplii spawned by femalesfed the non-oxylipin producing control diet PRO (data not shown, seeIanora and Miralto (2010)).
3.4. Female survival
PRO supported 100% survival throughout the experimental period,and diatoms, in contrast, showed lower values at the end of theexperiment: 50%, 6.6%, 40.7%, and 30% respectively in TR1, TR2, SMand SPC (Fig. 3). An ANCOVA analysis with time (LN transformed) ascovariate, female survival as dependent variable and algal treatment
as factor was performed with the set of treatments without includingPRO. No significant differences were found (F3, 51=0.08, p=0.97).
3.5. Ingestion rates and food selection
Ingestion rateswith each algawere quite similar between PRO, TR1and TR2 (Fig. 4). The presence of SPC seemed to produce some kind ofinterference on feeding, since ingestion rates were simultaneouslyreduced with both PRO and SPC in that treatment. Food selectionbetween each algae showed feeding behaviours close to a nonselective pattern (α=0.5) for PRO–TR1 and PRO–TR2. A Wilcoxontest showed that there were no statistical differences between pairs ofManly'sα values, so theywere not different from0.5 (PRO–TR1 n=12,Z=0.94, p=0.34; PRO–TR2 n=14, Z=0.47, p=0.64; PRO–SPC n=9,Z=1.84, p=0.06). This indicates that our results do not show aselective feeding behaviour.
3.6. Metabolite analysis
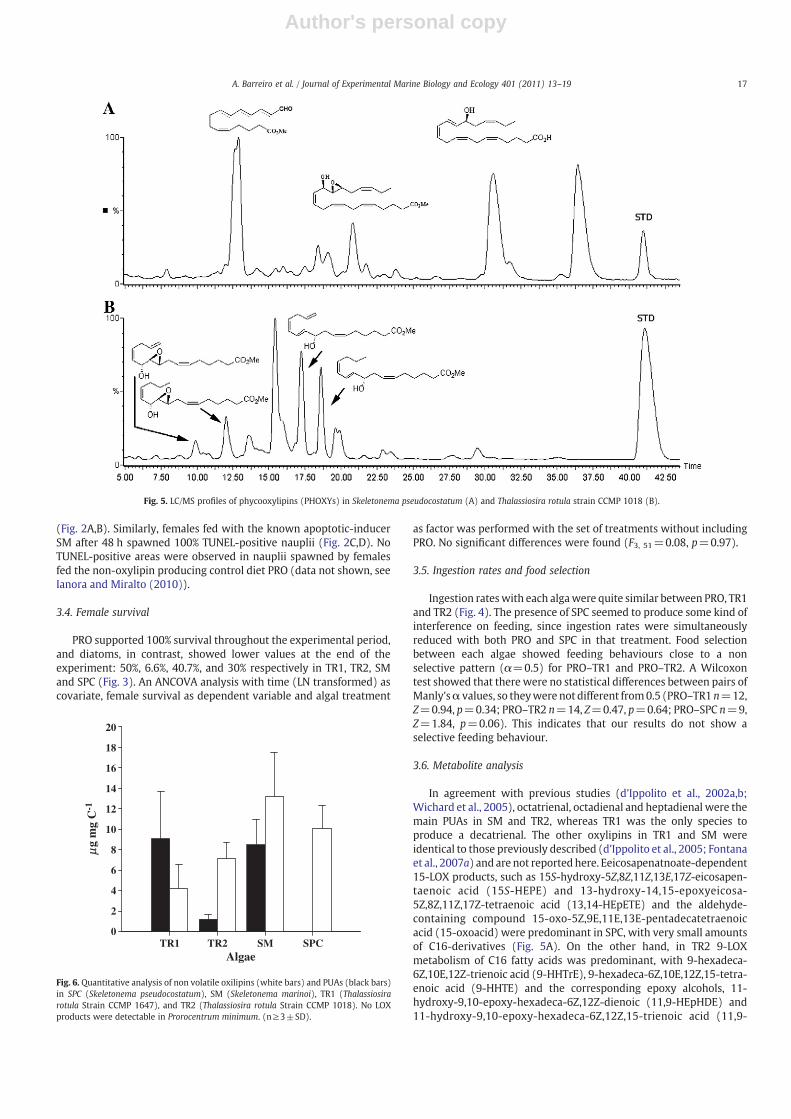
In agreement with previous studies (d'Ippolito et al., 2002a,b;Wichard et al., 2005), octatrienal, octadienal and heptadienal were themain PUAs in SM and TR2, whereas TR1 was the only species toproduce a decatrienal. The other oxylipins in TR1 and SM wereidentical to those previously described (d'Ippolito et al., 2005; Fontanaet al., 2007a) and are not reportedhere. Eeicosapenatnoate-dependent15-LOX products, such as 15S-hydroxy-5Z,8Z,11Z,13E,17Z-eicosapen-taenoic acid (15S-HEPE) and 13-hydroxy-14,15-epoxyeicosa-5Z,8Z,11Z,17Z-tetraenoic acid (13,14-HEpETE) and the aldehyde-containing compound 15-oxo-5Z,9E,11E,13E-pentadecatetraenoicacid (15-oxoacid) were predominant in SPC, with very small amountsof C16-derivatives (Fig. 5A). On the other hand, in TR2 9-LOXmetabolism of C16 fatty acids was predominant, with 9-hexadeca-6Z,10E,12Z-trienoic acid (9-HHTrE), 9-hexadeca-6Z,10E,12Z,15-tetra-enoic acid (9-HHTE) and the corresponding epoxy alcohols, 11-hydroxy-9,10-epoxy-hexadeca-6Z,12Z-dienoic (11,9-HEpHDE) and11-hydroxy-9,10-epoxy-hexadeca-6Z,12Z,15-trienoic acid (11,9-
Fig. 5. LC/MS profiles of phycooxylipins (PHOXYs) in Skeletonema pseudocostatum (A) and Thalassiosira rotula strain CCMP 1018 (B).
AlgaeTR1 TR2 SM SPC
0
2
4
6
8
10
12
14
16
18
20
g m
g C
-1µ
Fig. 6. Quantitative analysis of non volatile oxilipins (white bars) and PUAs (black bars)in SPC (Skeletonema pseudocostatum), SM (Skeletonema marinoi), TR1 (Thalassiosirarotula Strain CCMP 1647), and TR2 (Thalassiosira rotula Strain CCMP 1018). No LOXproducts were detectable in Prorocentrum minimum. (n≥3±SD).
17A. Barreiro et al. / Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19

Author's personal copy
HEpHTE), as the major products (Fig. 5B). Neither PUAs nor otheroxylipins were detectable in PRO (data not shown).
3.7. Dosage of LOX products
Non volatile oxylipins and PUA content normalised to total carbonare shown in Fig. 6. Production of oxylipins differed among the diatomspecies tested (one-way analysis of variance ANOVA, F3,11=7.8,pb0.01). This difference was due to the much lower amount of non-volatile oxylipins produced by TR1 (4.2±2.4 μg mg C−1) compared toSM (13.2±4.3 μg mg C−1) and SPC (10.0±2.3 μg mg C−1), since bothSkeletonema species and TR2 were not significantly different fromeach other (Tukey's post-hoc test, pN0.05). Interestingly, PUAs ac-counted for 39% of the total end-products in SM (8.5±2.5 μg mg C−1),and 68% of the total end-products in TR1 (9.0±3.5 μg mg C−1).However, the PUA content in both diatoms was not statisticallydifferent from non-volatile oxylipin content (Student's t-test:t4=1.64, pN0.05 for SM, and Student's t-test: t8=2.1, p=0.07 forTR1, respectively), though at the edge of the p value for TR1. Culturesof TR2 that have been reported as either able or unable to synthesisePUAs (Pohnert et al., 2002; Wichard et al., 2005) showed very lowlevels of PUA production (1.2±0.5 μg mg C−1) but much higher levelsof other oxylipins (7.1±1.6 μg mg C−1) (Student's t-test t4=6.2,pb0.01). No PUAs were detected in SPC.
4. Discussion
Results of ingestion experiments showed no clear evidence forselective feeding behaviourwith PRO, TR1 and TR2whichwere ingestedin similar amounts, as were PRO and SPC but at much lower con-centrations. Manly's coefficients were not significantly different thusindicating no selective behaviour even thoughwith PRO+SPC therewasa p value which was at the edge of statistical significance. Turner et al.(2001) also reported similar ingestion rates of TR1 andPROwhenofferedtogether, although food selection coefficients were not reported. Allthesefindings suggest that the chemicals produced by diatom species donot protect them from their grazers if the alternative prey is not of highfood quality, asmay be in the case of PRO. Of course, field conditionswilldiffer fromthis scenario because selective grazersmaybepresent, aswellas alternative good quality food. To date, little research has been con-ducted in thefield to study copepod feeding behaviour in the presenceofPUA-producing diatoms. The only study performed, by Leising et al.(2005a,b) showed a selective behaviour of Calanus pacificus, rejecting aPUA producing species of the genus Thalassiossira.
TR1 and TR2 induced higher egg production in T. stylifera com-pared to PRO, SM and SPC (Fig. 1, Table 2).
High egg production rates with diets of TR have already beenreported in the past for this copepod species (Turner et al., 2001;Pohnert et al., 2002; Ceballos and Ianora, 2003). On the other hand,both SM and SPC induced low fecundity similar to previous studies(Ban et al., 1997; Ceballos and Ianora, 2003). Intermediate eggproduction rates with PRO could be explained by the poor quality ofPRO as food (see review by Heil et al. (2005)) or by reduced“catchability” of the swimming PRO cells rather than the sinking TR2and TR1 cells. Female survival did not show differences among thediatoms tested compared to the control diet PRO. On the contrary, eggviability in Temora was affected by all diatom species tested (Fig. 1,Table 3). SM was already known to produce a strong reduction in egghatching (Ceballos and Ianora, 2003; Ianora et al., 2004; reviewed byIanora and Miralto (2010)).
SPC and TR2 had significantly stronger negative effects on hatchingviability than TR1 (Table 3) in Temora. In Pohnert et al. (2002), on thecontrary, TR2 (with no PUA production) and SPC did not induce asignificant decrease in egg viability in another copepod, Calanushelgolandicus. To date, such contrasting results have been explained asa result of a different sensitivity of different species or strains of
copepods to PUAs (Ianora et al., 2003) and/or to variations in PUAproduction (Ribalet et al., 2007) in response to environmental orphysiological conditions.
In agreement with previous reports (Wichard et al., 2005; Fontanaet al., 2007b; Dutz et al., 2008; Poulet et al. 2008), PUAs by themselvescannot explain reduced hatching success with all diatom diets. On theother hand, the negative effect of diatoms on copepod reproductionbecomes much clearer when other LOX products, namely oxygenatedlong chain fatty acid derivatives (hydroxyacids and epoxyalcohols), aretaken into consideration (Fontana et al., 2007b). Of the two species (SMand SPC)with strongest negative effects on copepod egg production andegg hatching success, only one, SM, was a PUA producer. In the absenceof PUAs, the strong activity of SPC on copepod egg viability can beexplained by the large quantities of other oxylipins, such as(5Z,8Z,11Z,13E,15S,17Z)-15-hydroxy-5,8,11,13,17-eicosapentaenoicacid (15S-HEPE) and 13,14-13R-hydroxy-14S,15S-trans-epoxyeicosa-5Z,8Z,11Z,17Z-tetraenoic acid (13,14-HEpETE) and 15-oxo-5Z,9E,11E,13E-pentadecatetraenoic acid (15-oxoacid), deriving fromthe oxidation of C20 fatty acids. Interestingly these metabolites wererecently reported in the pennate diatom P. delicatissima which doesnot produce PUAs but only these other oxylipins (d'Ippolito et al.2009). 15S-HEPE has already been shown to induce reproductivefailure in T. stylifera (Ianora et al., 2010) whereas similar aldehydic 9-oxo-nonadienoic and 12-oxo-dodecatienoic acids in Phaeodactylumtricornutum have been suggested to block cell divisions in sea urchinembryos (Pohnert et al., 2002). Herewe confirm that diatoms such asSPC (S. pseudocostatum) that produce these oxylipins can inducenegative effects comparable to those of SM (S. marinoi) even thoughthey do not produce PUAs. Such a response confirms previous resultswith SM and two species of Choetoceros that were not able tosynthesise PUAs (Fontana et al., 2007b).
Also in the case of the two T. rotula strains, TR2 was more toxiceven if it produced significantly lower quantities of PUAs compared toTR1. TR2 produced mainly 9-hexadecatrienoic acid (9-HHTrE), 9-hexadecatetraenoic acid (9-HHTE) and the corresponding epoxyalcohols which were present in lower quantities in TR1, possiblyexplaining the stronger activity of TR2 on hatching success.
Another interesting finding that has emerged from this studyregards with the chemical diversity of oxylipins, not only at thespecies but also at the strain level, as already noted by Wichard et al.(2005) for PUAs. The observed genetic differences between the ITSregions of TR1 (CCMP 1647) and TR2 (CCMP1018) support results byPohnert et al. (2002) that these two strains have been raised fromgenetically distinct individuals within T. rotula (Kooistra pers. comm.)This finding is not surprising because the strains originate from cellscollected ca. 25 years apart in different oceans (TR1, Gulf of Naples,Tyrrhenian Sea, Nov. 1993; TR2, La Jolla, California, 1968). What isinteresting is that different individuals exhibit different levels ofbioactive compounds and different cocktails of these, as a result ofdifferent enzymatic pathways committed to their production.
In termsof oxylipin uptake rates, Buttino et al. (2008) calculated thatT. stylifera females feeding on decadienal-loaded liposomes ingestedabout 9.7 ng of decadienal daily to reduce hatching success after10 days. Considering ingestion rates reported by Turner et al. (2001) inexperiments performed with the same copepod species under similarconditions as in our egg production experiments (with the speciesT. rotula, TR1), 13.2 μg mg C−1 of total oxylipins for TR1 (see ourFig. 5)×300 ng C ind−1 h−1 ingested (see Fig. 3 in Turner et al.(2001))×24 h corresponds to 95 ng of total oxylipins ingested ind−1
day−1, which is one order of magnitude higher than the concentrationsthat were active in Buttino et al. (2008). We do not have data for theother diatom species (genus Skeletonema), but the expected valuesshould be similarly high.
The copepods' inability to detect oxylipins may render diatom-grazer relationships unsuitable for co-evolution processes (Ianoraet al., 2006). Instead, evolution of diatom toxins could be driven by kin
18 A. Barreiro et al. / Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19

Author's personal copy
selection, as these compounds protect the entire population againstgrazers by reducing the number of grazer offspring. In this case, thebenefit would be on inclusive fitness (of genetically related in-dividuals or clonal members) relatively nearby rather than exclusivefitness of individual cells or chains of genetically identical cells. Inconclusion, the negative impact of diatoms is not due to a single classof molecules (i.e. PUAs), as previously believed, but rather to a cocktailof bioactive compounds, most of which have been discovered onlyrecently. The “rapid response” production process of these com-pounds probably evolved to deter grazers (Ianora et al., 2006).
Acknowledgements
We are grateful to Mario di Pinto, Massimo Perna and AntonioMaiello for their technical assistance. A.B. thanks the Stazione ZoologicaAnton Dohrn in Naples for providing general facilities. A.B. was fundedwith a FPU grant from the Spanish government. Funds were alsoobtained from the EU Network of Excellence Marine Biodiversity andEcosystem Functioning (contract number GOCE-CT-2003-505446). [SS]
References
Ban, S., Burns, Castel J., Chaudron, Y., Christou, E., Escribano, R., Fonda Umani, S.,Gasparini, S., Guerrero Ruiz, F., Hoffmeyer, M., Ianora, A., Kang, H.K., Laabir, M.,Lacoste, A., Miralto, A., Ning, X., Poulet, S., Rodríguez, V., Runge, J., Shi, J., Starr, M.,Uye, S.I., Wang, J., 1997. The paradox of diatom–copepod interactions. Mar. Ecol.Prog. Ser. 157, 287–293.
Buttino, I., De Rosa, G., Carotenuto, Y., Mazzella, M., Ianora, A., Esposito, F., Vitiello, V.,Quaglia, F., La Rotonda, M.I., Miralto, A., 2008. Aldehyde-encapsulating liposomesimpair marine grazer survivorship. J. Exp. Biol. 211, 1426–1433.
Carotenuto, Y., Ianora, A., Buttino, I., Romano, G., Miralto, A., 2002. is postembyonicdevelopment in the copepod Temora stylifera negatively affected by diatom diets?J. Exp. Mar. Biol. Ecol. 276, 49–66.
Ceballos, S., Ianora, A., 2003. Different diatoms induce contrasting effects on thereproductive success of the copepod Temora stylifera. J. Exp. Mar. Biol. Ecol. 294,189–202.
Chaudron, Y., Poulet, S.A., Laabir, M., Ianora, A., Miralto, A., 1996. Is hatching success ofcopepod eggs diatom density-dependent? Mar. Ecol. Prog. Ser. 144, 185–193.
D'Ippolito, G., Romano, G., Iadicicco, O., Miralto, A., Ianora, A., Cimino, G., Fontana, A.,2002a. New birth-control aldehydes from the marine diatom Skeletonemacostatum: characterization and biogenesis. Tetrahedron Lett. 43, 6133–6136.
D'Ippolito, G., Iadicicco, O., Romano, G., Fontana, A., 2002b. Detection of short-chainaldehydes in marine organisms: the diatom Thalassiossira rotula. Tetrahedron Lett.43, 6137–6140.
D'Ippolito, G., Cutignano, A., Briante, R., Febbraio, F., Cimino, G., Fontana, A., 2005. NewC16 fatty-acid-based oxylipin pathway in the marine diatom Thalassiosira rotula.Org. Biomol. Chem. 3, 4065–4070.
Dutz, J., Koski, M., Jónasdóttir, S.H., 2008. Copepod reproduction is unaffected by diatomaldehydes or lipid composition. Limnol. Oceanogr. 53, 225–235.
Fontana, A., D'Ippolito, G., Cutignano, A., Romano, G., Ianora, A., Miralto, A., Cimino, G.,2007a. Chemistry of oxylipins pathways in marine diatoms. Pure Appl. Chem. 79(4), 481–490.
Fontana, A., D'Ippolito, G., Cutignano, A., Romano, G., Lamari,N.,Massa-Gallucci, A., Cimino,G., Miralto, A., Ianora, A., 2007b. LOX-induced lipid peroxidation mechanism re-sponsible for the detrimental effect of marine diatoms on zooplankton grazers.Chembiochem 8 (15), 1810–1818.
Frost, B.W., 1972. Effects of size and concentration of food particles on the feedingbehaviour of themarine planktonic copepod Calanus pacificus. Limnol. Oceanog. 17,805–815.
Heil, C.A., Glibert, P.M., Fan, C., 2005. Prorocentrum minimum (Pavillard) Schiller. Areview of a harmful algal bloom species of growing worldwide importance.Harmful. Algae. 4, 449–470.
Ianora, A., Miralto, A., 2010. Toxigenic effects of diatoms on grazers, phytoplankton andother microbes: a review. Ecotoxicology 19, 493–511.
Ianora, A., Scotto di Carlo, B., Mascellaro, P., 1989. Reproductive biology of theplanktonic copepod Temora stylifera. Mar. Biol. 101, 187–194.
Ianora, A., Poulet, S.A., Miralto, A., 2003. The effects of diatoms on copepod reproduc-tion: a review. Phycologia 42 (4), 351–363.
Ianora, A., Miralto, A., Poulet, S., Carotenuto, Y., Buttino, I., Romano, G., Casotti, R.,Pohnert, G., Wichard, T., Colucci-D'Amato, L., Terrazzano, G., Smetacek, V., 2004.Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonicdiatom. Nature 429, 403–407.
Ianora, A., Boersma, M., Casotti, R., Esposito, F., Harder, J., Hoffmann, F., Pavia, H., Potin,P., Poulet, S.A., Toth, G., 2006. New trends in marine chemical ecology. EstuariesCoasts 29 (4), 531–551.
Ianora, A., Romano, G., Carotenuto, Y., Esposito, F., Roncalli, V., Buttino, I., Miralto, A.,2010. Impact of the diatom oxylipin 15S-HEPE on the reproductive success of thecopepod Temora stylifera. Hydrobiologia 666 (1), 265–275.
Kang, H.K., Poulet, S.A., 2000. Reproductive success in Calanus helgolandicus as afunction of diet and egg cannibalism. Mar. Ecol. Prog. Ser. 201, 241–250.
Leftley, J.W., Keller, D.K., Selvin, R.C., Claus, W., Guillard, R.R.L., 1987. Media for theculture of oceanic ultraphytoplankton. J. Phycol. 23, 633–638.
Leising, A.W., Pierson, J.J., Halsband-Lenk, C., Horner, R., Postel, J., 2005a. Copepodgrazing during spring blooms: does Calanus pacificus avoid harmful diatoms? Prog.Oceanog. 67, 384–405.
Leising, A.W., Pierson, J.J., Halsband-Lenk, C., Horner, R., Postel, J., 2005b. Copepodgrazing during spring blooms: can Pseudocalanus newmani induce trophiccascades? Prog. Oceanog. 67, 406–421.
Manly, B.F.J., 1974. A model for certain types of selection experiments. Biometrics 30,281–294.
Miralto, A., Barone, G., Romano, G., Poulet, S.A., Ianora, A., Russo, G.L., Buttino, I.,Mazzarella, G., Laabir, M., Cabrini, M., Giacobbe, M.G., 1999. The insidious effect ofdiatoms on copepod reproduction. Nature 402, 173–176.
Pohnert, G., 2005. Diatom/Copepod interactions in plankton: the indirect chemicaldefence of unicellular algae. Chembiochem 6, 1–14.
Pohnert, G., Lumineau, O., Cueff, A., Adolph, S., Cordevant, C., Lange, M., Poulet, S., 2002.Are volatile unsaturated aldehydes from diatoms the main line of chemical defenceagainst copepods? Mar. Ecol. Prog. Ser. 245, 33–45.
Ribalet, F., Wichard, T., Pohnert, G., Ianora, A., Miralto, A., Casotti, R., 2007. Age andnutrient limitation enhance polyunsaturated aldehyde production in marinediatoms. Phytochemistry 68 (15), 2059–2067.
Turner, J.T., Ianora, A., Miralto, A., Laabir, M., Esposito, F., 2001. Decoupling of copepodgrazing rates, fecundity and egg-hatching success on mixed and alternating diatomand dinoflagellate diets. Mar. Ecol. Prog. Ser. 220, 187–199.
Wichard, T., Poulet, S., Halsband-Lenk, Albaina A., Harris, R., Liu, D., Pohnert, G., 2005.Survey of the chemical defence potential of diatoms: screening of fifty one speciesfor α, β, γ, δ-unsaturated aldehydes. J. Chem. Ecol. 31, 949–958.
19A. Barreiro et al. / Journal of Experimental Marine Biology and Ecology 401 (2011) 13–19
Related Documents











