Lmnol. Oceanogr.. 36(7), 199 I, 1358-l 374 0 199 1, by the American Society of’ Limnology and Oceanography, Inc. Diagenesis of lbelowground biomass of Spartina alterniJlora irn salt-marsh sediments Ronald Benner Marine Science Institute, University of Texas at Austin, Port Aransas 78373 Marilyn L. Fogel Geophysical Laboratory, 525 I Broad Branch Rd., Carnegie Institution of Washington, Washington, D.C. 200 15 E. Kent Sprague MCrone Environmental Services, Norcross, Georgia 30093 Abstract Belowground biomass of Spartina alterniflora lost 55% of its organic matter during 18 months of decomposition in salt-marsh sediments. Significant losses of lignin were observed under highly reducing conditions confirming previous reports of lignin degradation in the absence of molecular oxygen. The submolecular components of lignin varied in susceptibility to degradation and were ranked in the following descending order of resistance to decomposition: V-P> S- C. The pref- erential degradation of 1 he cinnamyl and syringyl components constrains the quantitative usefulness of these compounds for estimating the contributions ofherbaceous and angiosperm tissues to pools of dissolved and particulate organic matter. The relative concentration of lignin increased about 1.6-fold during decomposition owing to the more rapid loss of polysaccharides. Two phases in nitrogen dynamics were evident during the decomposition study. During the first 4 months of decomposition there was a net loss of N from Spartina tissues at a rate of about 10 gg N g-l tissue d- I. After this initial phase of net N loss, a phase of N immobilization was evident and was tightly coupled with rates of microbial degradation of the tissues. Net rates of N immo- bilization were - 19 pg N g-’ tissue d -I; by the end of the 18-month study more N had accumulated in the tissues than was initially present. Stable N isotope composilions of decaying Spartina were also very dynamic and reflected the sources of N that were immobilized during decomposition and the N transformations that occurred on the detrital particles. The stable C isotope composition of the polysaccharide components of Spartina was -6%~ heavier than that of the lignin component. The isotopic compositions of these components retained the 6o/oo difference throughout the 18-month study. The stable C isotope composition of the bulk tissues, however, decreased gradually during decomposition as the isotopically heavier polysac- charides were preferentially lost. Residual material was relatively enriched in the isotopically lighter, lignin-deriveci C. ’ It has been recognized for some time that salt marshes are highly productive ecosys- tems. The smooth cordgrass, Spartina al- terniflora, is the dominant macrophyte in many salt marshes, and estimates of pri- mary production for Spartina in Georgia marshes range from I, 100 to 7,600 g dry mass m-2 yr-’ (Schubauer and Hopkinson 1984). By most estimates, belowground --- Acknowledgments We thank Steve Opsahl, John Hedges, Miguel Goni, and two anonymous reviewers for comments on the manuscript. This research was supported by NSF grant BSR 89- 10766 and grant NASYAA-D-SG 139 from the NOAA Ofice of Sea Grant. University of Texas Marine Science Institute Con- tribution 8 IO. production of Spartina roots and rhizomes exceeds production of aboveground com- ponents (Good et al. 1982; Schubauer and Hopkinson 1984). There have been numer- ous studies of the aerobic decomposition of aboveground portions of Spartina, but rel- atively little is known about the transfor- mations and fates of belowground biomass in the predominantly anoxic salt-marsh sediments. There are several possible fates for Spar- tina root and rhizome material including accretion in sediments, grazing by animals, and microbial decomposition. Salt marshes on the southeastern seaboard of the U.S. are accreting sediments at rates ranging from 1.4 to 11 .O mm yr-’ (Sharma et al. 1987), and the organic matter contents of salt- 1358

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lmnol. Oceanogr.. 36(7), 199 I, 1358-l 374 0 199 1, by the American Society of’ Limnology and Oceanography, Inc.
Diagenesis of lbelowground biomass of Spartina alterniJlora irn salt-marsh sediments
Ronald Benner Marine Science Institute, University of Texas at Austin, Port Aransas 78373
Marilyn L. Fogel Geophysical Laboratory, 525 I Broad Branch Rd., Carnegie Institution of Washington, Washington, D.C. 200 15
E. Kent Sprague MCrone Environmental Services, Norcross, Georgia 30093
Abstract
Belowground biomass of Spartina alterniflora lost 55% of its organic matter during 18 months of decomposition in salt-marsh sediments. Significant losses of lignin were observed under highly reducing conditions confirming previous reports of lignin degradation in the absence of molecular oxygen. The submolecular components of lignin varied in susceptibility to degradation and were ranked in the following descending order of resistance to decomposition: V-P> S- C. The pref- erential degradation of 1 he cinnamyl and syringyl components constrains the quantitative usefulness of these compounds for estimating the contributions ofherbaceous and angiosperm tissues to pools of dissolved and particulate organic matter. The relative concentration of lignin increased about 1.6-fold during decomposition owing to the more rapid loss of polysaccharides.
Two phases in nitrogen dynamics were evident during the decomposition study. During the first 4 months of decomposition there was a net loss of N from Spartina tissues at a rate of about 10 gg N g-l tissue d- I. After this initial phase of net N loss, a phase of N immobilization was evident and was tightly coupled with rates of microbial degradation of the tissues. Net rates of N immo- bilization were - 19 pg N g-’ tissue d -I; by the end of the 18-month study more N had accumulated in the tissues than was initially present. Stable N isotope composilions of decaying Spartina were also very dynamic and reflected the sources of N that were immobilized during decomposition and the N transformations that occurred on the detrital particles.
The stable C isotope composition of the polysaccharide components of Spartina was -6%~ heavier than that of the lignin component. The isotopic compositions of these components retained the 6o/oo difference throughout the 18-month study. The stable C isotope composition of the bulk tissues, however, decreased gradually during decomposition as the isotopically heavier polysac- charides were preferentially lost. Residual material was relatively enriched in the isotopically lighter, lignin-deriveci C. ’
It has been recognized for some time that salt marshes are highly productive ecosys- tems. The smooth cordgrass, Spartina al- terniflora, is the dominant macrophyte in many salt marshes, and estimates of pri- mary production for Spartina in Georgia marshes range from I, 100 to 7,600 g dry mass m-2 yr-’ (Schubauer and Hopkinson 19 84). By most estimates, belowground --- Acknowledgments
We thank Steve Opsahl, John Hedges, Miguel Goni, and two anonymous reviewers for comments on the manuscript.
This research was supported by NSF grant BSR 89- 10766 and grant NASYAA-D-SG 139 from the NOAA Ofice of Sea Grant.
University of Texas Marine Science Institute Con- tribution 8 IO.
production of Spartina roots and rhizomes exceeds production of aboveground com- ponents (Good et al. 1982; Schubauer and Hopkinson 1984). There have been numer- ous studies of the aerobic decomposition of aboveground portions of Spartina, but rel- atively little is known about the transfor- mations and fates of belowground biomass in the predominantly anoxic salt-marsh sediments.
There are several possible fates for Spar- tina root and rhizome material including accretion in sediments, grazing by animals, and microbial decomposition. Salt marshes on the southeastern seaboard of the U.S. are accreting sediments at rates ranging from 1.4 to 11 .O mm yr-’ (Sharma et al. 1987), and the organic matter contents of salt-
1358

Diagenesis of Spartina 1359
marsh sediments typically range from 5- 20% (Ember et al. 1987; Sharma et al. 1987). The accretion of organic matter in salt-marsh sediments ranges from 70 to 225 g m-2 yr-’ (assumes organic matter is 40 wt% C; Howes et al. 1985; Wiegert and Pomeroy 198 1) and may be an important fate for a significant fraction of belowground production. Esti- mates of belowground production of Spar- tina range from -2,000 to 4,500 g me2 yr-’ (Gallagher and Plumley 1979; Schubauer and Hopkinson 1984), so it is unlikely that accretion accounts for > 5-l 0% of be- lowground production. The direct utiliza- tion of roots and rhizomes by geese and other large vertebrates has been document- ed, and marsh invertebrates, such as fiddler crabs, also remove unknown quantities of root and rhizome material, but it is unlikely that grazing losses are significant (Good et al. 1982).
By far the major fate of Spartina be- lowground production appears to be micro- bial decomposition in undisturbed marsh sediments. During the decomposition of Spartina substantial quantities of dissolved organic matter are produced, but studies of water movement and dissolved organic C fluxes in pore waters indicate that there is very little export of dissolved organic mat- ter from salt-marsh sediments (Howes et al. 1985; Yelverton and Hackney 1986). Rates of C mineralization (to C02) in salt-marsh sediments are high enough to account for the decomposition of 80-100% of Spartina belowground production (Howes et al. 1985). It also appears from the high rates of sulfate reduction in salt-marsh sediments that most of the decomposition occurs un- der anoxic conditions (Howarth and Hob- bie 1982; Howes et al. 1984).
The extent to which Spartina detritus contributes to sedimentary organic matter in salt marshes is not clear. Stable C isotope compositions of sedimentary organic mat- ter in marshes dominated by Spartina are consistently depleted in 13C (613C of - 16 to -20%0) relative to Spartina plant tissues (613C of - 13%0) (Haines 1976; Ember et al. 1987). The relatively large difference bc- tween the bulk isotopic composition of Spartina tissues and the isotopic signature of sedimentary organic matter is suggestive
that other sources of organic matter with isotopic values similar to those of sedimen- tary organic matter, such as algae, are dom- inant contributors to the organic C in sed- iments or that alterations of isotopic compositions occur during decomposition of plant tissues or both.
Recent analyses of the stable C isotope compositions of the lignin and polysaccha- ride components of Spartina indicate that the lignin component is similarly depleted (4-7%0) in l 3C relative to polysaccharides which comprise the bulk of plant tissues (Benner et al. 1987). The polysaccharide components of Spartina and other plants are degraded by microorganisms at rates several-fold higher than the lignin compo- nent, leading to the gradual enrichment of lignin-derived C in remaining tissues (Ben- ner et al. 19843, 1986b, 1987). The obser- vation that the stable C isotope composition of Spartina shifted to more negative values during decomposition as the relative con- centration of lignin increased led to the hy- pothesis that Spartina may contribute sig- nificantly more to pools of sedimentary and particulate organic matter in salt marshes and adjacent coastal waters than would be predicted based on the C isotopic compo- sitions of undegraded tissues (Benner et al. 1987; Ember et al. 1987; Fogel et al. 1989).
In the present investigation, we charac- terize the overall rates and patterns of de- composition of Spartina root and rhizome material in salt-marsh sediments. Particular emphasis is placed on the fate of the lignin component of Spartina in predominantly anoxic sediments. Stable C isotope com- positions of bulk tissues and the lignin and polysaccharide components are followed throughout an 18-month study period. Sta- ble N isotope compositions of bulk tissues as well as total N dynamics are also char- acterized.
Materials and methods Litterbug study- Root and rhizome ma-
terial was collected in January 198 5 from a high-marsh area (Airport Marsh) on Sapelo Island, Georgia. Blocks of sediment with short-form S. alterniflora plants were re- moved from the marsh and returned to the laboratory where living root and rhizome

1360 Benner et al.
material was removed and thoroughly washed. Greater than 85% of the biomass collected was contained in rhizomes. The plant material was dried at 40°C for 1 week and - 1.8 g of material was placed in litter- bags made of polypropylene (Spectra/Mesh, Fisher Scientific) with mesh openings of 105 pm. The small mesh opening was used to prevent the loss of particles from bags due to physical fragmentation or the activity of larger invertebrates and to impede the pen- etration of growing root and rhizome ma- terial into the litterbags. The litterbags were buried with minimal disturbance of the sed- iments at depths of 8-l 0 cm in February 1985 in the same marsh a:rea where the plant material was collected.
Four replicate litterbags were collected every l-3 months over a period of 18 months. Sediment temperature at a depth of 9 cm was recorded at the time of collec- tion. After 15 months of decomposition, four replicate litterbags were removed from be- lowground and placed on the sediment sur- face under oxidizing conditions to compare aerobic decomposition characteristics with those of material that remained be- lowground under anoxic conditions. After collection from the field, litterbags were washed in deionized water to remove sed- iments and the contents of the bags were dried at 40°C weighed, and ground in a Wiley mill to pass a 40-mesh screen. Sub- samples of material from each bag were placed in a muffle furnace at 500°C for 2-3 h for determination of ash content. C, H, and N contents of the plant material were determined with a Perkin-Elmer 240C CHN analyzer. Weight losses a:nd elemental com- positions are reported on an ash-free dry weight (AFDW) basis. Oxygen content was determined by difference.
Lignin oxidation products-The lignin component of Spartina tissues was char- acterized by capillary gas chromatography of Cu0 oxidation products as described pre- viously (Hedges and Ertel I 982; Benner et al. 1990b). Recoveries of lignin-derived phenols were estimated from the recovery of an internal standard, ethylvanillin, which was added to the reaction mixture after ox- idation. Chemical alteration of lignin-de- rived phenols during oxidation in alkaline
Cu0 appears to be minimal as > 85% of these phenols is recovered typically (Hedg- es 1975). We found, however, that alkaline Cu0 oxidation of ferulic acid at 170°C yielded vanillyl phenols (of which vanillin was the major reaction product) as well as ferulic acid. It is important to note that ex- ternal rather than internal bomb tempera- tures were measured and maintained during Cu0 oxidation. Recent studies have shown that internal bomb temperatures can be sig- nificantly lower than external bomb tem- peratures and that yields of lignin-derived phenols are temperature-dependent (J. Hedges pers. comm.). In the present study 47.9 + 1.7% of the ferulic acid oxidized ( 10 mg) was recovered as ferulic acid. The other lignin-derived cinnamyl phenol, p-couma- ric acid, was recovered with fairly high ef- ficiency (77.8 + 1 .O%) after oxidation. The major reaction product recovered after ox- idation of p-coumaric acid was p-hydroxy- benzaldehyde. The relatively high conver- sion of ferulic acid to vanillyl phenols is suggestive that internal bomb temperatures were probably close to 170°C. No correc- tions for these alterations were made in the yields of lignin-derived phenols reported herein.
The weight percentages of lignin in de- caying Spartina tissues were estimated with lignin-derived phenol data for comparison to the weight percentages of lignin estimated by the acid-detergent extraction scheme (see below). Polymeric lignin in plant tissues is not stoichiometrically converted to simple phenols during oxidative degradation with alkaline CuO. Vanillyl phenols are pro- duced from the lignin polymer with -30% efficiency and syringyl phenols are produced with -90% efficiency (Sarkanen and Lud- wig 197 1; Manders 1987). Recovered yields of lignin-derived phenols were corrected for these losses as well as C losses from the side- chains of simple phenols to estimate weight percentages of lignin in Spartina. The meth- od of calculating lignin weight percentages from yields of lignin-derived phenols was given by Benner et al. (19906).
Isolation of specijk chemical fractions - A sequential detergent extraction procedure (Van Soest and Robertson 1980) was used to isolate specific chemical fractions from
Diagenesis of Spartina 1361
fresh and degraded Spartina. A neutral-de- tergent fiber (NDF) fraction consisting of hemicellulose, cellulose, and lignin, and an acid-detergent lignin (ADL) fraction con- sisting of acid-insoluble lignin were isolated for stable C isotope analysis. The stable C isotope composition of lignin was deter- mined directly, whereas the isotope com- position of the polysaccharides, hemicel- lulose and cellulose, was determined by mass balance with ash-free dry weight recoveries of the NDF and ADL fractions. In a pre- vious study, Benner et al. (1987) demon- strated that the stable C isotope composi- tions of hemicellulose and cellulose from vascular plant tissues, including Spartina, were similar. All isolated fractions were thoroughly washed with warm water to re- move detergents. The stable C isotope com- position of purified cellulose did not change (+ 0. lo/oo) after detergent extractions and washing, indicating that the detergents were completely removed during washing and did not interfere with determination of isotope compositions.
Stable C and N isotope analyses-The C and N isotope compositions of fresh and degraded Spartina were determined on - 10 mg dry wt samples. Samples were com- busted at 900°C in evacuated quartz tubes for 60 min before they were cooled at a controlled rate (0.6”C min-‘) to room tem- perature (modification of the procedure of Macko 198 1). In general, the precision of the analyses was +O. l?& for 613C and &0.2?&0 for 615N. The isotope composition of a sam- ple is defined as
aNE = [(Rsample/Rstd) - l] x lo3
where NE is the heavy isotope of an element (13Cor15N)andRistheratioof13C:12Cor 15N : 14N in the sample and the standard. The standard for 613C was PDB (O.O?&), and the standard for 615N was air (O.Oo/oo).
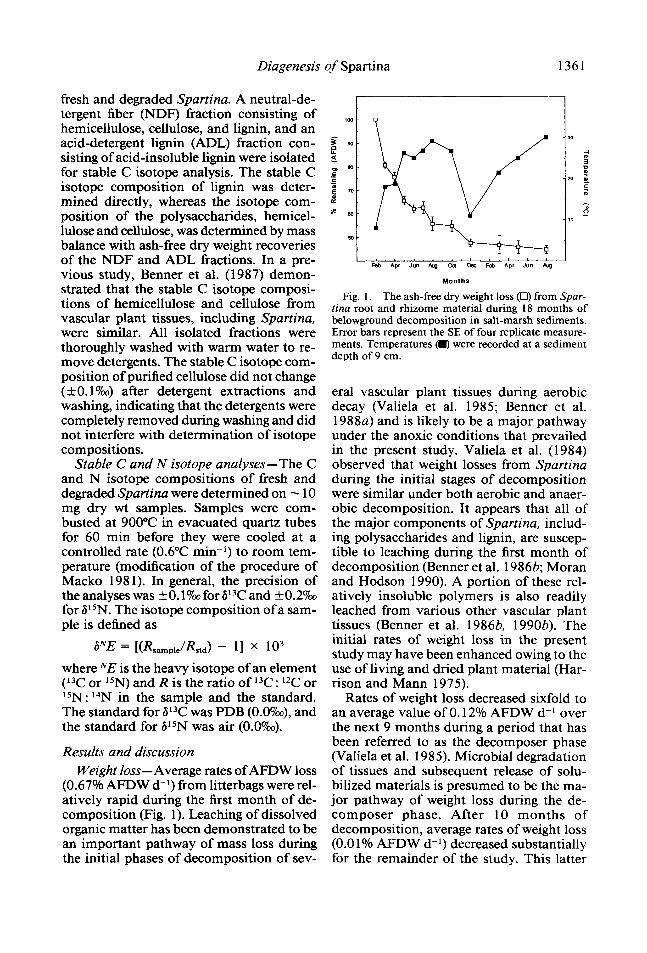
Results and discussion Weight loss-Average rates of AFDW loss
(0.67% AFDW d-l) from litterbags were rel- atively rapid during the first month of de- composition (Fig. 1). Leaching of dissolved organic matter has been demonstrated to be an important pathway of mass loss during the initial phases of decomposition of sev-
I. I. 1. I. 1. 1. I I. I .I. I .I Fob Apr Jun Pug Cd Dee Fob Apr Jun w
Months
Fig. 1. The ash-free dry weight loss (Cl) from Spar- tina root and rhizome material during 18 months of belownround decomDosition in salt-marsh sediments. Error bars represent ;he SE of four replicate measure- ments. Temperatures 0 were recorded at a sediment depth of 9 cm.
era1 vascular plant tissues during aerobic decay (Valiela et al. 1985; Benner et al. 1988a) and is likely to be a major pathway under the anoxic conditions that prevailed in the present study. Valiela et al. (1984) observed that weight losses from Spartina during the initial stages of decomposition - were similar under both aerobic and anaer- obic decomposition. It appears that all of the major components of Spartina, includ- ing polysaccharides and lignin, are suscep- tible to leaching during the first month of decomposition (Benner et al. 19863; Moran and Hodson 1990). A portion of these rel- atively insoluble polymers is also readily leached from various other vascular plant tissues (Benner et al. 19863, 1990b). The initial rates of weight loss in the present study may have been enhanced owing to the use of living and dried plant material (Har- rison and Mann 1975).
Rates of weight loss decreased sixfold to an average value of 0.12% AFDW d-l over the next 9 months during a period that has been referred to as the decomposer phase (Valiela et al. 1985). Microbial degradation of tissues and subsequent release of solu- bilized materials is presumed to be the ma- jor pathway of weight loss during the de- composer phase. After 10 months of decomposition, average rates of weight loss (0.0 1% AFDW d-l) decreased substantially for the remainder of the study. This latter

1362 Benner et al.
stage of decomposition is referred to as the refractory phase and is characterized by very low rates of weight loss (Valiela et al. 1985). Varying rates of decomposition primarily reflect changes in chemical composition, as the remaining material gradually becomes enriched in refractory components such as lignin (Benner et al. 19843; Valiela et al. 1985).
After 15 months of belowground decom- position, three replicate litterbags were re- moved and placed on the sediment surface to investigate differences in weight loss and chemical composition between material in- cubated under reducing and oxidizing con- ditions. Material incubated belowground continued to lose weight at an average rate of 0.0 1% AFDW d-l during the last 3 months of decomposition, whereas material incu- bated on the sediment surface lost weight at a sixfold higher rate. Similarly, Valiela et al. (1984) found that rates of aboveground decomposition were much higher than be- lowground rates after the initial leaching pe- riod. It is possible that the greater rates of weight loss for material incubated on the sediment surface result, in part, from a loss of fine particles (< 105 pm) from the litter- bags because of greater physical distur- bance.
Sediment temperatures at 9-cm depth were measured at the time of collection of the litterbags and ranged from 8°C in Feb- ruary to 30°C in August (Fig. 1). Minimum and maximum air temperatures, which are monitored daily at the Sapelo Island Marine Institute, ranged from - 9°C to 4 1 “C during the study period. Temperature has been shown to strongly influence rates of micro- bial degradation of Spartina in salt-marsh sediments (Benner et al. 1986a), but in the present study temporal changes in chemical composition and decomposability con- found the seasonal influence of temperature on rates of weight loss. Overall rates of be- lowground decomposition (55% AFDW loss in 1.5 yr) in the present study were relatively high compared to other studies of be- lowground decomposition of salt-marsh vegetation (Buth 1987; Hackney and de la Cruz 1980; Valiela et al. 1984).
Lignin parameter stability -Lignin is synthesized only by vascular plants and has
proven to be an effective geochemical tracer of terrigenous organic matter in aquatic en- vironments (Hedges et al. 1988). Upon ox- idation with alkaline CuO, herbaceous an- giosperm tissues, such as Spartina, produce a suite of 11 phenols which are derived pre- dominantly from the lignin polymer (Hedg- es and Mann 1979). In addition to being specific indicators of vascular plant mate- rial, lignin-derived phenols also provide in- formation about the major taxonomic groupings (angiosperm vs. gymnosperm) and types (woody vs. nonwoody) of plant tissues in environmental samples. Syringyl phenols (S) are indicative of angiosperm tissues and cinnamyl (C) phenols are indicative of non- woody tissues, whereas vanillyl phenols (I”) are found in all lignified tissues. The abun- dance ratios of these phenol types, S : Vand C : V, have been used to discriminate be- tween angiosperm and gymnosperm and be- tween woody and nonwoody plant tissues (Hedges and Mann 1979). The quantitative usefulness of lignin and the parameters gen- erated from yields of lignin-derived phenols in interpretating the amounts and types of vascular plant tissues in environmental samples, such as sediments, is dependent on the extent of lignin degradation and al- teration that occurs during diagensis. We investigated the early diagenesis of Spartina lignin in salt-marsh sediments to provide baseline information for future applications of lignin-derived phenols as indicators of salt-marsh vegetation in estuarine sedi- ments.
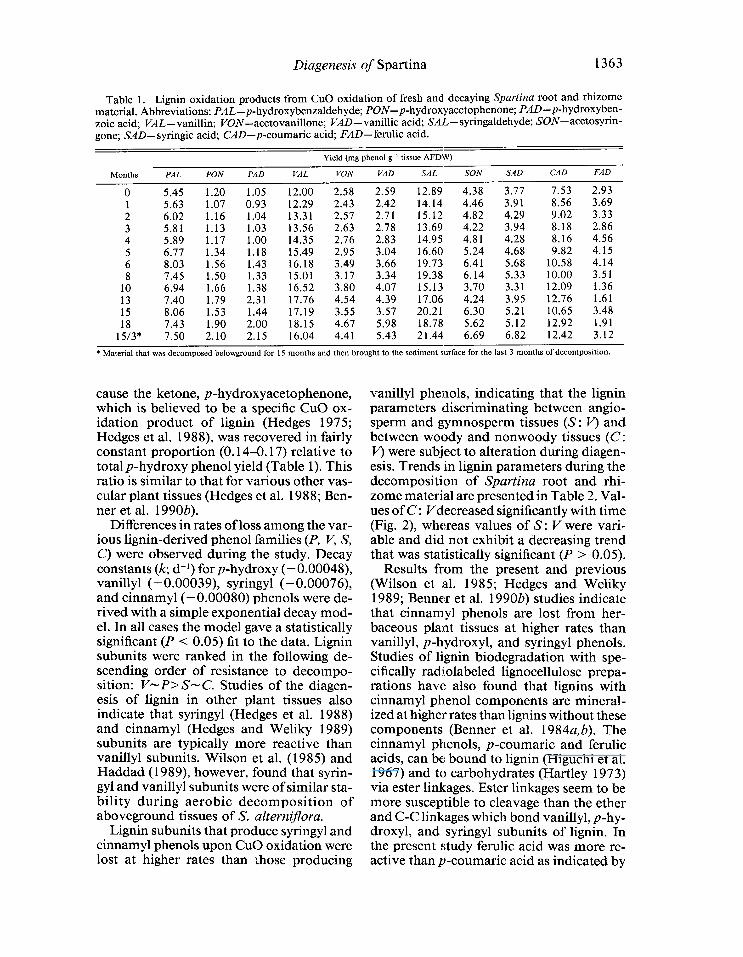
Lignin-derived phenol yields (mg phenol g-l tissue AFDW) for fresh and decomposed Spartina root and rhizome material are pre- sented in Table 1. With the exception of ferulic acid, yields increase with increasing decomposition as other components, such as polysaccharides, are degraded at higher rates than the lignin component (Benner et al. 1987). The p-hydroxy phenols are Cu0 oxidation products of lignin from most vas- cular plant tissues (Sarkanen and Ludwig 197 l), and they are included in the present discussion even though there are other non- lignin sources for p-hydroxybenzaldehyde and p-hydroxybenzoic acid (Hedges 1975). In the present study, the p-hydroxy phenols appear to be primarily lignin-derived be-
Diagenesis of Spartina 1363
Table 1. Lignin oxidation products from Cu0 oxidation of fresh and decaying Spartina root and rhizome material. Abbreviations: PAL-p-hydroxybenzaldehyde; PON-p-hydroxyacetophenone; PAD-p-hydroxyben- zoic acid; VAL- vanillin; VON- acetovanillone; VAD- vanillic acid; SAL- syringaldehyde; SON-acetosyrin- gone; SAD-syringic acid; CAD-p-coumaric acid; FAD-ferulic acid.
Yield (mg phenol g ’ tissue AFDW)
Months PAL PON PAD V/IL VON VAD SAL SON SAD CAD FAD
0 5.45 1.20 1.05 12.00 2.58 2.59 12.89 4.38 3.77 7.53 2.93 1 5.63 1.07 0.93 12.29 2.43 2.42 14.14 4.46 3.91 8.56 3.69 2 6.02 1.16 1.04 13.31 2.57 2.71 15.12 4.82 4.29 9.02 3.33 3 5.81 1.13 1.03 13.56 2.63 2.78 13.69 4.22 3.94 8.18 2.86 4 5.89 1.17 1 .oo 14.35 2.76 2.83 14.95 4.81 4.28 8.16 4.56 5 6.77 1.34 1.18 15.49 2.95 3.04 16.60 5.24 4.68 9.82 4.15 6 8.03 1.56 1.43 16.18 3.49 3.66 19.73 6.41 5.68 10.58 4.14 8 7.45 1.50 1.33 15.01 3.17 3.34 19.38 6.14 5.33 10.00 3.51
10 6.94 1.66 1.38 16.52 3.80 4.07 15.13 3.70 3.31 12.09 1.36 13 7.40 1.79 2.31 17.76 4.54 4.39 17.06 4.24 3.95 12.76 1.61 15 8.06 1.53 1.44 17.19 3.55 3.57 20.21 6.30 5.21 10.65 3.48 18 7.43 1.90 2.00 18.15 4.67 5.98 18.78 5.62 5.12 12.92 1.91
15/3* 7.50 2.10 2.15 16.04 4.41 5.43 21.44 6.69 6.82 12.42 3.12
* Material that was decomposed belowground for 15 months and then brought to the scdimcnt surface for the last 3 months of decomposition.
cause the ketone, p-hydroxyacetophenone, which is believed to be a specific Cu0 ox- idation product of lignin (Hedges 1975; Hedges et al. 1988), was recovered in fairly constant proportion (0.14-o. 17) relative to total p-hydroxy phenol yield (Table 1). This ratio is similar to that for various other vas- cular plant tissues (Hedges et al. 1988; Ben- ner et al. 1990b).
Differences in rates of loss among the var- ious lignin-derived phenol families (P, V, S, C) were observed during the study. Decay constants (k; d-l) for p-hydroxy (-0.00048), vanillyl (-0.00039), syringyl (-0.00076), and cinnamyl (- 0.00080) phenols were de- rived with a simple exponential decay mod- el. In all cases the model gave a statistically significant (P < 0.05) fit to the data. Lignin subunits were ranked in the following de- scending order of resistance to decompo- sition: I/-P>S- C. Studies of the diagen- esis of lignin in other plant tissues also indicate that syringyl (Hedges et al. 1988) and cinnamyl (Hedges and Weliky 1989) subunits are typically more reactive than vanillyl subunits. Wilson et al. (1985) and Haddad (1989), however, found that syrin- gyl and vanillyl subunits were of similar sta- bility during aerobic decomposition of aboveground tissues of S. alternijlora.
Lignin subunits that produce syringyl and cinnamyl phenols upon Cu0 oxidation were lost at higher rates than those producing
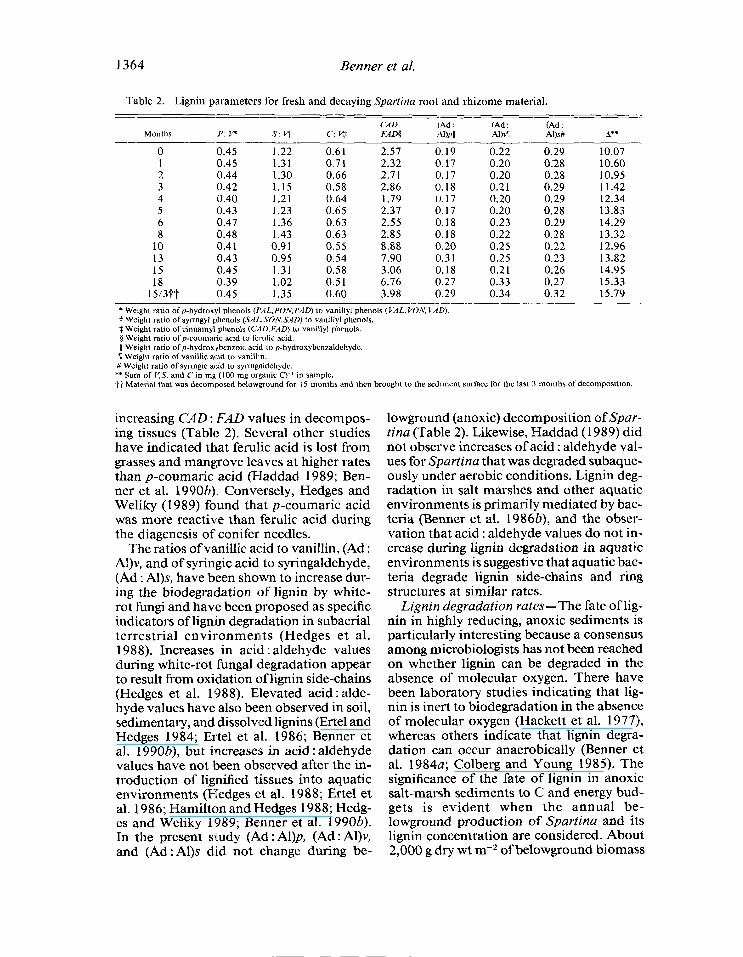
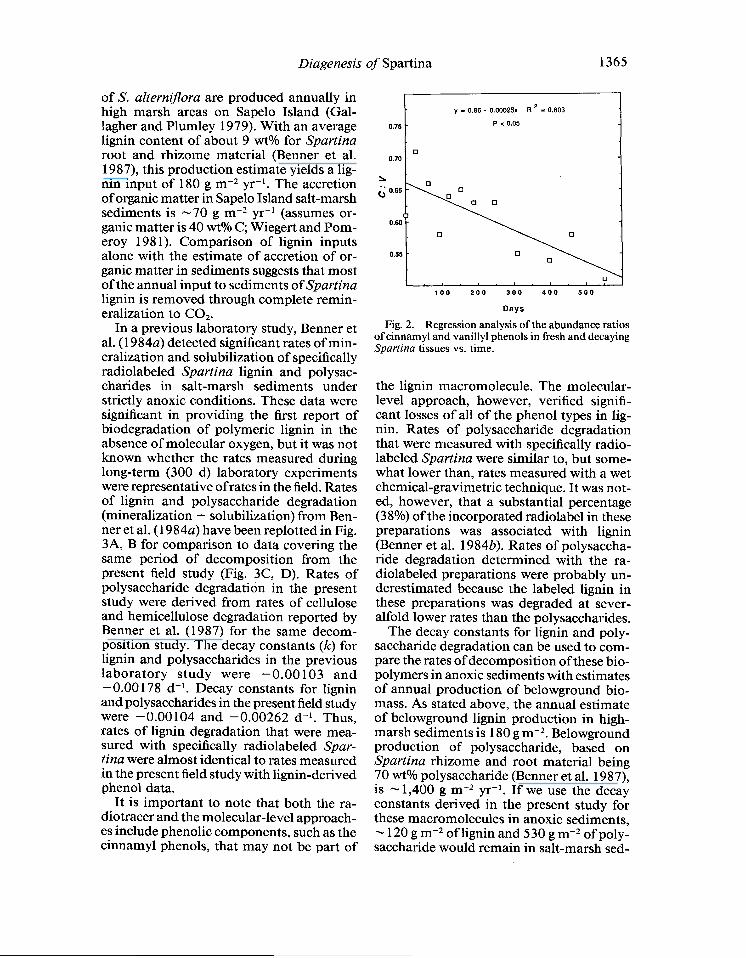
vanillyl phenols, indicating that the lignin parameters discriminating between angio- sperm and gymnosperm tissues (S: v) and between woody and nonwoody tissues (C: v) were subject to alteration during diagen- esis. Trends in lignin parameters during the decomposition of Spartina root and rhi- zome material are presented in Table 2. Val- ues of C : Vdecreased significantly with time (Fig. 2) whereas values of S: V were vari- able and did not exhibit a decreasing trend that was statistically significant (P > 0.05).
Results from the present and previous (Wilson et al. 1985; Hedges and Wcliky 1989; Benner et al. 1990b) studies indicate that cinnamyl phenols are lost from her- baceous plant tissues at higher rates than vanillyl, p-hydroxyl, and syringyl phenols. Studies of lignin biodegradation with spe- cifically radiolabeled lignocellulose prepa- rations have also found that lignins with cinnamyl phenol components are mineral- ized at higher rates than lignins without these components (Benner et al. 1984a,b). The cinnamyl phenols, p-coumaric and ferulic acids, can be bound to lignin (Higuchi et al. 1967) and to carbohydrates (Hartley 1973) via ester linkages. Ester linkages seem to be more susceptible to cleavage than the ether and C-C linkages which bond vanillyl, p-hy- droxyl, and syringyl subunits of lignin. In the present study ferulic acid was more re- active than p-coumaric acid as indicated by
1364 Benner et al.
‘Table 2. Lignin parameters for fresh and decaying Spartina root and rhizome material. = --- --_-- -~- ---- ~---.- ---_ --_-- --
CAD: (Ad : (Ad : (Ad : Months P: P s: vt c: vj: FADP mull Al)vll Al)s# A**
- ----
--- 0 0.45 1.22 0.61 2.57 0.19 0.22 0.29 10.07 1 0.45 1.31 0.71 2.32 0.17 0.20 0.28 10.60 2 0.44 1.30 0.66 2.7 1 0.17 0.20 0.28 10.95 3 0.42 1.15 0.58 2.86 0.18 0.21 0.29 11.42 4 0.40 1.21 0.64 1.79 0.17 0.20 0.29 12.34 5 0.43 1.23 0.65 2.37 0.17 0.20 0.28 13.83 6 0.47 1.36 0.63 2.55 0.18 0.23 0.29 14.29 8 0.48 1.43 0.63 2.85 0.18 0.22 0.28 13.32
10 0.41 0.91 0.55 8.88 0.20 0.25 0.22 12.96 13 0.43 0.95 0.54 7.90 0.31 0.25 0.23 13.82 15 0.45 1.31 0.58 3.06 0.18 0.21 0.26 14.95
1:;3+t 0.39 0.45 1.02 1.35 0.5 0.60 1 6.76 3.98 0.27 0.29 0.33 0.34 0.32 0.27 15.79 15.33
- -~ ._.-e-w-. ----- ---- * Weight ratio of p-hydroxyl phenols (PAL,PON,PAD) to vanillyl phenols (VAL, VON, MD). t Weight ratio of syringyl phenols (SrlL+K)N,S/iD) lo vanillyl phenols. $ Weight ratio of cinnamyl phenols (CAD. FAD) to vanillyl phenols. 9 Weight ratio of p-coumaric acid to ferulic acid. (1 Weight ratio ofp-hydroxybenzoic acid to p-hydroxybcnzaldchyde. B Weight ratio of vanillic acid to vanill n. # Weight ratio of syringic acid to syringaldehyde. ** Sum of V,S. and C in mg (100 mg organic C) ’ in sample. tt Material that was decomposed bclowground for 15 months and then brought to the sediment surface for the last 3 months of decomposition.
increasing CAD : FAD values in decompos- ing tissues (Table 2). Several other studies have indicated that ferulic acid is lost from grasses and mangrove leaves at higher rates than p-coumaric acid (Haddad 1989; Ben- ner et al. 1990b). Conversely, Hedges and Weliky (1989) found that p-coumaric acid was more reactive than ferulic acid during the diagenesis of conifer needles.
The ratios of vanillic acid to vanillin, (Ad : Al)v, and of syringic acid to syringaldehyde, (Ad : Al)s, have been shown to increase dur- ing the biodegradation of lignin by white- rot fungi and have been proposed as specific indicators of lignin degraldation in subaerial terrestrial environments (Hedges et al. 1988). Increases in acid : aldehyde values during white-rot fungal degradation appear to result from oxidation of lignin side-chains (Hedges et al. 1988). Elevated acid : alde- hyde values have also been observed in soil, sedimentary, and dissolved lignins (Ertel and Hedges 1984; Ertel et al. 1986; Benner et al. 1990b), but increase:s in acid : aldehyde values have not been observed after the in- troduction of lignified tissues into aquatic environments (Hedges et al. 1988; Ertel et al. 1986; Hamilton and Hedges 1988; Hedg- es and Weliky 1989; Benner et al. 1990b). In the present study (A,d : Al)p, (Ad : Al)v, and (Ad: Al)s did not change during be-
lowground (anoxic) decomposition of Spar- tina (Table 2). Likewise, Haddad (1989) did not observe increases of acid : aldehyde val- ues for Spartina that was degraded subaque- ously under aerobic conditions. Lignin deg- radation in salt marshes and other aquatic environments is primarily mediated by bac- teria (Benner et al. 19863), and the obser- vation that acid : aldehyde values do not in- crease during lignin degradation in aquatic environments is suggestive that aquatic bac- teria degrade lignin side-chains and ring structures at similar rates.
Lignin degradation rates-The fate of lig- nin in highly reducing, anoxic sediments is particularly interesting because a consensus among microbiologists has not been reached on whether lignin can be degraded in the absence of molecular oxygen. There have been laboratory studies indicating that lig- nin is inert to biodegradation in the absence of molecular oxygen (Hackett et al. 1977), whereas others indicate that lignin degra- dation can occur anaerobically (Benner et al. 1984a; Colberg and Young 1985). The significance of the fate of lignin in anoxic salt-marsh sediments to C and energy bud- gets is evident when the annual be- lowground production of Spartina and its lignin concentration are considered. About 2,000 g dry wt mm2 of belowground biomass
Diagenesis of Spartina 1365
of S. alterniflora are produced annually in high marsh areas on Sapelo Island (Gal- lagher and Plumley 1979). With an average lignin content of about 9 wt% for Spartina root and rhizome material (Benner et al. 1987), this production estimate yields a lig- nin input of 180 g mm2 yr- l. The accretion of organic matter in Sapelo Island salt-marsh sediments is -70 g m-2 yr-’ (assumes or- ganic matter is 40 wt% C; Wiegert and Pom- eroy 198 1). Comparison of lignin inputs alone with the estimate of accretion of or- ganic matter in sediments suggests that most of the annual input to sediments of Spartina lignin is removed through complete remin- eralization to CO,.
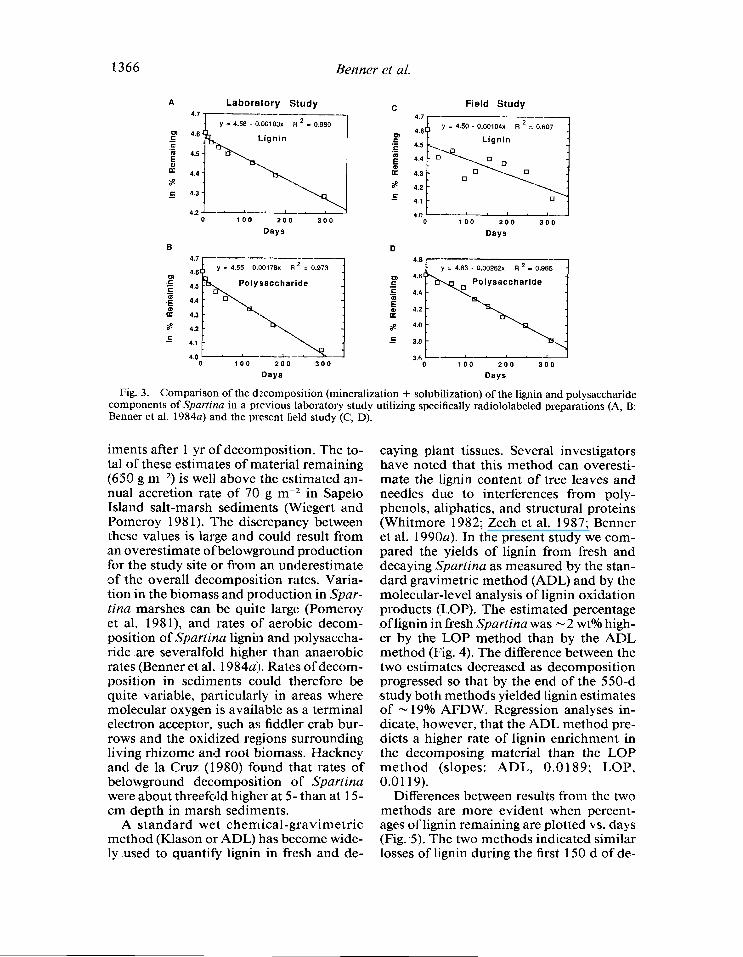
In a previous laboratory study, Benner et al. (1984a) detected significant rates of min- eralization and solubilization of specifically radiolabeled Spartina lignin and polysac- charides in salt-marsh sediments under strictly anoxic conditions. These data were significant in providing the first report of biodegradation of polymeric lignin in the absence of molecular oxygen, but it was not known whether the rates measured during long-term (300 d) laboratory experiments were representative of rates in the field. Rates of lignin and polysaccharide degradation (mineralization + solubilization) from Ben- ner et al. (1984a) have been replotted in Fig. 3A, B for comparison to data covering the same period of decomposition from the present field study (Fig. 3C, D). Rates of polysaccharide degradation in the present study were derived from rates of cellulose and hemicellulose degradation reported by ‘Benner et al. (1987) for the same decom- position study. The decay constants (k) for lignin and polysaccharides in the previous laboratory study were -0.00 103 and -0.00178 d-l. Decay constants for lignin and polysaccharides in the present field study were -0.00104 and -0.00262 d-l. Thus, rates of lignin degradation that were mea- sured with specifically radiolabeled Spar- tina were almost identical to rates measured in the present field study with lignin-derived phenol data.
It is important to note that both the ra- diotracer and the molecular-level approach- es include phenolic components, such as the cinnamyl phenols, that may not be part of
y = 0.66 - 0.00025x R ’ = 0.603
I . I * 1 . ’ . 1” 100 200 300 400 500
Days Fig. 2. Regression analysis of the abundance ratios
of cinnamyl and vanillyl phenols in fresh and decaying Spartina tissues vs. time.
the lignin macromolecule. The molecular- level approach, however, verified signifi- cant losses of all of the phenol types in lig- nin. Rates of polysaccharide degradation that were measured with specifically radio- labeled Spartina were similar to, but some- what lower than, rates measured with a wet chemical-gravimetric technique. It was not- ed, however, that a substantial percentage (38%) of the incorporated radiolabel in these preparations was associated with lignin (Benner et al. 19843). Rates of polysaccha- ride degradation determined with the ra- diolabeled preparations were probably un- derestimated because the labeled lignin in these preparations was degraded at sever- alfold lower rates than the polysaccharides.
The decay constants for lignin and poly- saccharide degradation can be used to com- pare the rates of decomposition of these bio- polymers in anoxic sediments with estimates of annual production of belowground bio- mass. As stated above, the annual estimate of belowground lignin production in high- marsh sediments is 180 g m-2. Belowground production of polysaccharide, based on Spartina rhizome and root material being 70 wt% polysaccharide (Benner et al. 1987), is - 1,400 g m-2 yr-‘. If we use the decay constants derived in the present study for these macromolecules in anoxic sediments, - 120 g m-2 of lignin and 530 g m-2 of poly- saccharide would remain in salt-marsh sed-
1366 Benner et al.
A Laboratory Study
100
Field Study
Days
Polysaccharide
Days
‘L-L.- 0 100 200 300 0 100 200 300
Days Days
Fig. 3. Comparison of the decomposition (mineralization + solubilization) of the lignin and polysaccharide components of Spartina in a previous laboratory study utilizing specifically radiololabeled preparations (A, B: Benner et al. 1984a) and the present field study (C, D).
iments after 1 yr of decomposition. The to- tal of these estimates of material remaining (650 g mp2) is well above the estimated an- nual accretion rate of 70 g m-* in Sapelo Island salt-marsh sediments (Wiegert and Pomeroy 198 1). The discrepancy between these values is large and could result from an overestimate of belowground production for the study site or from an underestimate of the overall decomposition rates. Varia- tion in the biomass and production in Spar- tina marshes can be quite large (Pomeroy et al. 198 I), and rates of aerobic decom- position of Spartina lignin and polysaccha- ride are severalfold higher than anaerobic rates (Benner et al. 1984a). Rates of decom- position in sediments could therefore be quite variable, particularly in areas where molecular oxygen is available as a terminal electron acceptor, such as fiddler crab bur- rows and the oxidized regions surrounding living rhizome and root biomass. Hackney and de la Cruz (1980) found that rates of belowground decomposition of Spartina were about threefold higher at 5- than at 15- cm depth in marsh sediments.
A standard wet chemical-gravimetric method (Klason or ADL) has become wide- ly used to quantify lignin in fresh and de-
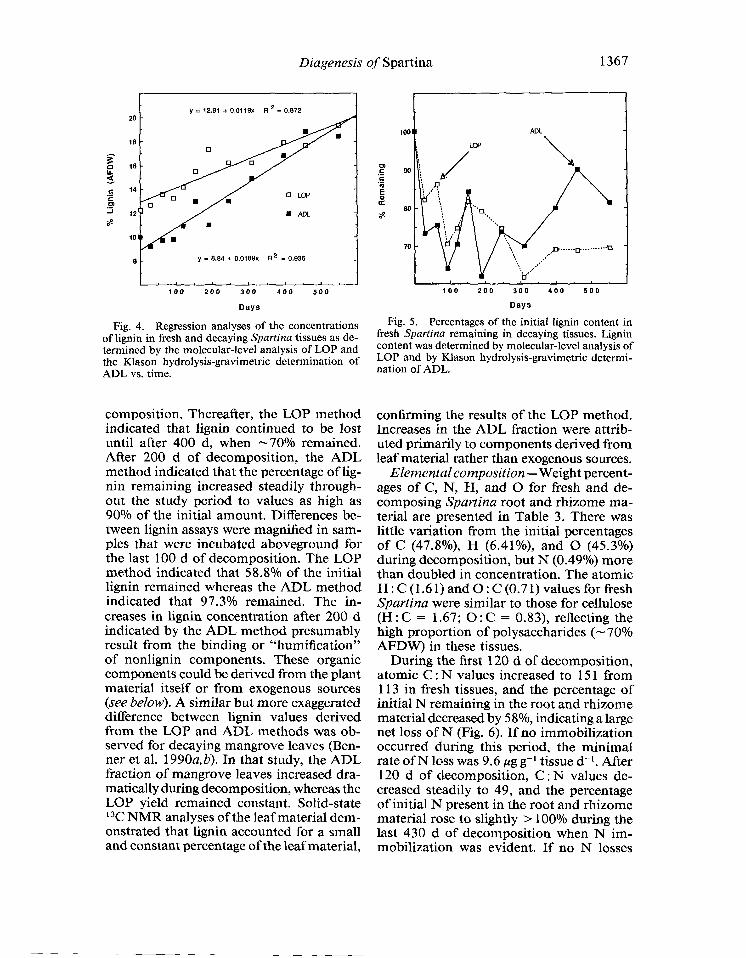
caying plant tissues. Several investigators have noted that this method can overesti- mate the lignin content of tree leaves and needles due to interferences from poly- phenols, aliphatics, and structural proteins (Whitmore 1982; Zech et al. 1987; Benner et al. 1990a). In the present study we com- pared the yields of lignin from fresh and decaying Spartina as measured by the stan- dard gravimetric method (ADL) and by the molecular-level analysis of lignin oxidation products (LOP). The estimated percentage of lignin in fresh Spartina was - 2 wt% high- er by the LOP method than by the ADL method (Fig. 4). The difference between the two estimates decreased as decomposition progressed so that by the end of the 550-d study both methods yielded lignin estimates of - 19% AFDW. Regression analyses in- dicate, however, that the ADL method pre- dicts a higher rate of lignin enrichment in the decomposing material than the LOP method (slopes: ADL, 0.0189; LOP, 0.0 119).
Differences between results from the two methods are more evident when percent- ages of lignin remaining are plotted vs. days (Fig. 5). The two methods indicated similar losses of lignin during the first 150 d of de-
Diagenesis of Spartina 1367
yz12.91 +0.0119x R2=0.672 20 -
8- y = 6.64 + 0.0189x R2 = 0.936
I. I . I . I .I . ‘.I
100 200 300 400 500
Days
Fig. 4. Regression analyses of the concentrations of lignin in fresh and decaying Spartina tissues as de- termined by the molecular-level analysis of LOP and the Mason hydrolysis-gravimetric determination of ADL vs. time.
composition. Thereafter, the LOP method indicated that lignin continued to be lost until after 400 d, when -70% remained. After 200 d of decomposition, the ADL method indicated that the percentage of lig- nin remaining increased steadily through- out the study period to values as high as 90% of the initial amount. Differences be- tween lignin assays were magnified in sam- ples that were incubated aboveground for the last 100 d of decomposition. The LOP method indicated that 58.8% of the initial lignin remained whereas the ADL method indicated that 97.3% remained. The in- creases in lignin concentration after 200 d indicated by the ADL method presumably result from the binding or “humification” of nonlignin components. These organic components could be derived from the plant material itself or from exogenous sources (see below). A similar but more exaggerated difference between lignin values derived from the LOP and ADL methods was ob- served for decaying mangrove leaves (Ben- ner et al. 1990a, b). In that study, the ADL fraction of mangrove leaves increased dra- matically during decomposition, whereas the LOP yield remained constant. Solid-state l 3C NMR analyses of the leaf material dem- onstrated that lignin accounted for a small and constant percentage of the leaf material,
Days
Fig. 5. Percentages of the initial lignin content in fresh Spartina remaining in decaying tissues. Lignin content was determined by molecular-level analysis of LOP and by Klason hydrolysis-gravimetric determi- nation of ADL.
confirming the results of the LOP method. Increases in the ADL fraction were attrib- uted primarily to components derived from leaf material rather than exogenous sources.
Elemental composition-weight percent- ages of C, N, H, and 0 for fresh and de- composing Spartina root and rhizome ma- terial are presented in Table 3. There was little variation from the initial percentages of C (47.8%), H (6.410/o), and 0 (45.3%) during decomposition, but N (0.49%) more than doubled in concentration. The atomic H:C(1.6l)andO:C(0.71)valuesforfresh Spartina were similar to those for cellulose (H : C = 1.67; 0 : C = 0.83), reflecting the high proportion of polysaccharides (- 70% AFDW) in these tissues.
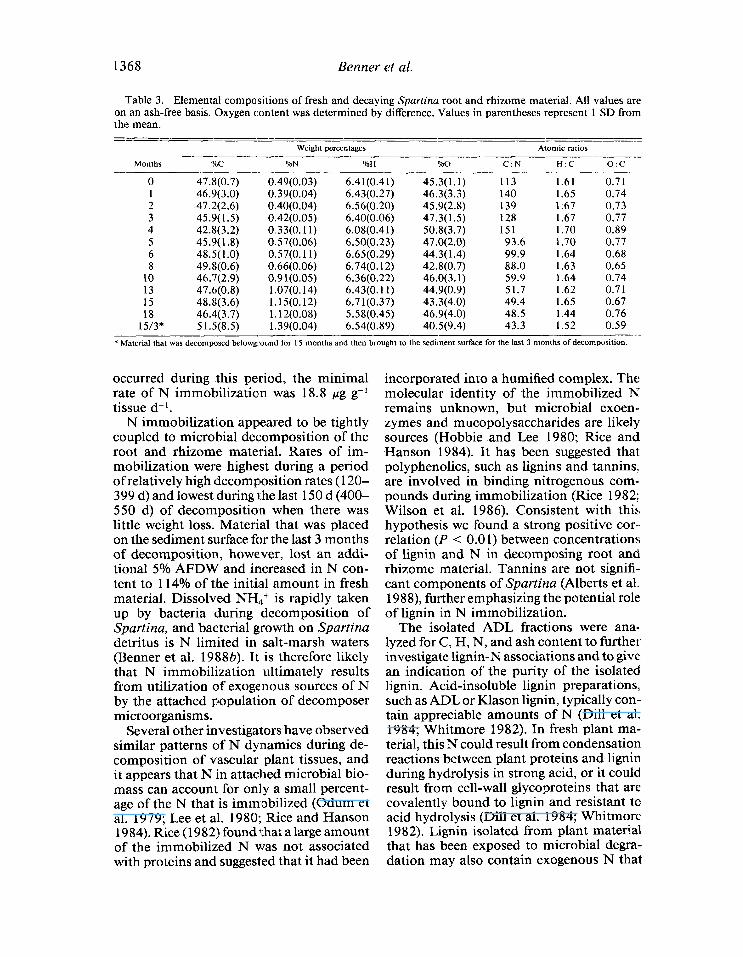
During the first 120 d of decomposition, atomic C : N values increased to 15 1 from 113 in fresh tissues, and the percentage of initial N remaining in the root and rhizome material decreased by 5 8%, indicating a large net loss of N (Fig. 6). If no immobilization occurred during this period, the minimal rate of N loss was 9.6 hg g-l tissue d-l. After 120 d of decomposition, C : N values de- creased steadily to 49, and the percentage of initial N present in the root and rhizome material rose to slightly > 100% during the last 430 d of decomposition when N im- mobilization was evident. If no N losses
1368 Benner et al.
Table 3. Elemental compositions of fresh and decaying Spartina root and rhizome material. All values are on an ash-free basis. Oxygen content was determined by difference. Values in parentheses represent 1 SD from the mean. = ---- -- -___---- -- --
Weight percentages Atomic ratios ----I_ -- ---
Months %C %N %H %O C:N - H:C 0:c
0 47.8(0.7) 1 46.9(3.0) 2 47.2(2.6) 3 45.9(1.5) 4 42.8(3.2) 5 45.9(1.8) 6 48.5(1.0) 8 49.8(0.6)
10 46.7(2.9) 13 47.6(0.8) 15 48.8(3.6) 18 46.4(3.7)
15/3* 5 1.5(8.5)
0.49(0.03) O-39(0.04) 0.40(0.04) 0.42(0.05) 0.33(0.11) O-57(0.06) 0.57(0.11) 0.66(0.06) 0.91(0.05) 1.07(0.14) 1.15(0.12) l-12(0.08) 1.39(0.04)
6.41(0.41) 6.43(0.27) 6.56(0.20) 6.40(0.06) 6.08(0.4 1) 6.50(0.23) 6.65(0.29) 6.74(0.12) 6.36(0.22) 6.43(0.1 1) 6.7 l(O.37) 5.58(0.45) 6.54(0.89)
45.3(1.1) 113 1.61 0.71 46.3(3.3) 140 1.65 0.74 45.9(2.8) 139 1.67 0.73 47.3(1.5) 128 1.67 0.77 50.8(3.7) 151 1.70 0.89 47.0(2.0) 93.6 1.70 0.77 44.3(1.4) 99.9 1.64 0.68 42.8(0.7) 88.0 1.63 0.65 46.0(3.1) 59.9 1.64 0.74 44.9(0.9) 51.7 1.62 0.71 43.3(4.0) 49.4 1.65 0.67 46.9(4.0) 48.5 1.44 0.76 40.5(9.4) 43.3 1.52 0.59
* Material that was decomposed belowg-ound for I5 months and then brought to the sediment surface for the last 3 months of decomposition.
occurred during this period, the minimal rate of N immobilization was 18.8 pg g- 1 tissue d-l.
N immobilization appeared to be tightly coupled to microbial decomposition of the root and rhizome material. Rates of im- mobilization were highest during a period of relatively high decomposition rates ( 120- 399 d) and lowest during the last 150 d (400- 550 d) of decomposition when there was little weight loss. Material that was placed on the sediment surface for the last 3 months of decomposition, however, lost an addi- tional 5% AFDW and increased in N con- tent to 114% of the initial amount in fresh material. Dissolved NH.,+ is rapidly taken up by bacteria during decomposition of Spartina, and bacterial growth on Spartina detritus is N limited in salt-marsh waters (Benner et al. 1988b). It is therefore likely that N immobilization ultimately results from utilization of exogenous sources of N by the attached population of decomposer microorganisms.
Several other investigators have observed similar patterns of N dy:namics during de- composition of vascular plant tissues, and it appears that N in attached microbial bio- mass can account for only a small percent- age of the N that is immobilized (Odum et al. 1979; Lee et al. 1980; Rice and Hanson 19 84). Rice ( 19 82) found that a large amount of the immobilized N was not associated with proteins and suggested that it had been
incorporated into a humified complex. The molecular identity of the immobilized N remains unknown, but microbial exoen- zymes and mucopolysaccharides are likely sources (Hobbie and Lee 1980; Rice and Hanson 1984). It has been suggested that polyphenolics, such as lignins and tannins., are involved in binding nitrogenous com- pounds during immobilization (Rice 19821, Wilson et al. 1986). Consistent with this hypothesis we found a strong positive car-a relation (B < 0.0 1) between concentrations of lignin and N in decomposing root and rhizome material. Tannins are not signifi-, cant components of Spartina (Alberts et al. 19 8 8) further emphasizing the potential role of lignin in N immobilization.
The isolated ADL fractions were ana-, lyzed for C, H, N, and ash content to further investigate lignin-N associations and to give an indication of the purity of the isolated lignin. Acid-insoluble lignin preparations, such as ADL or Klason lignin, typically con- tain appreciable amounts of N (Dill et al. 1984; Whitmore 1982). In fresh plant ma- terial, this N could result from condensation reactions between plant proteins and lignin during hydrolysis in strong acid, or it could result from cell-wall glycoproteins that are covalently bound to lignin and resistant to acid hydrolysis (Dill et al. 1984; Whitmore 1982). Lignin isolated from plant material that has been exposed to microbial degra- dation may also contain exogenous N that
Diagenesis of Spartina 1369
was covalently bound to lignin via highly reactive quinones formed during enzymatic degradation of phenolic compounds (Liu et al. 1985). The lignin fractions from fresh and degraded Spartina samples all had sig- nificant amounts of N (1.20-l .92% AFDW). N dynamics in the ADL fraction were sim- ilar to those in the total tissue. Accordingly, there was a significant (P < 0.05) correlation between the total tissue N and N in the ADL fraction. A total of 30.4+ 5.9% of the N in fresh and degraded tissues was recovered in the ADL fraction.
The elemental composition (CHNO) of the ADL fraction can be used to place some analytical constraints on its molecular com- position. The measured N concentrations in the isolated ADL fractions suggest that protein could account for as much as 7.5- 12.0% (N x 6.25) of the ADL fraction. The average C value for the ADL fractions, 59.0+_3.59% AFDW, is similar to the per- centage of C in angiosperm lignins (60.1% Sarkanen and Ludwig 197 l), and the av- erage atomic 0 : C (0.42) and H : C (1.36) values for the ADL fractions are also similar to values for angiosperm lignins (0.42 and 1.23; Sarkanen and Ludwig 197 1). Proteins have an average C content of 5 1.5% and average 0 : C and H : C values of 0.38 and 1.54 (Reuter and Perdue 1984). Given the above compositions for lignin and protein, a mixture of 90% lignin and 10% protein would be 5 9.3% C and would have 0 : C and H : C values of 0.42 and 1.38. These predicted 0 : C and H : C values (0.42, 1.38) are almost identical to measured values (0.42, 1.36) for the ADL fraction, indicat- ing, as does the N content of the ADL frac- tion, that protein accounts for - 10% of the ADL fraction.
Stable N isotopes -The stable N isotope compositions (615N) of fresh and decom- posed Spartina were measured to gain some insights about the source(s) of immobilized N in anoxic sediments. The initial 615N of root and rhizome material was 3.1%0 (Table 4), which is more negative than previously reported values for Spartina root and rhi- zome material from Sapelo Island (Peterson and Howarth 1987) and nearby St. Cath- erine’s Island (Fogel et al. 1989). Although there seems to be little variation in the sta-
100 200 300 400 500
Days
Fig. 6. Percentages of the initial N content in Spartina tissues remaining in decaying tissues.
fresh
ble C isotope composition of Spartina, spa- tial or temporal variations in stable N iso- tope composition are significant.
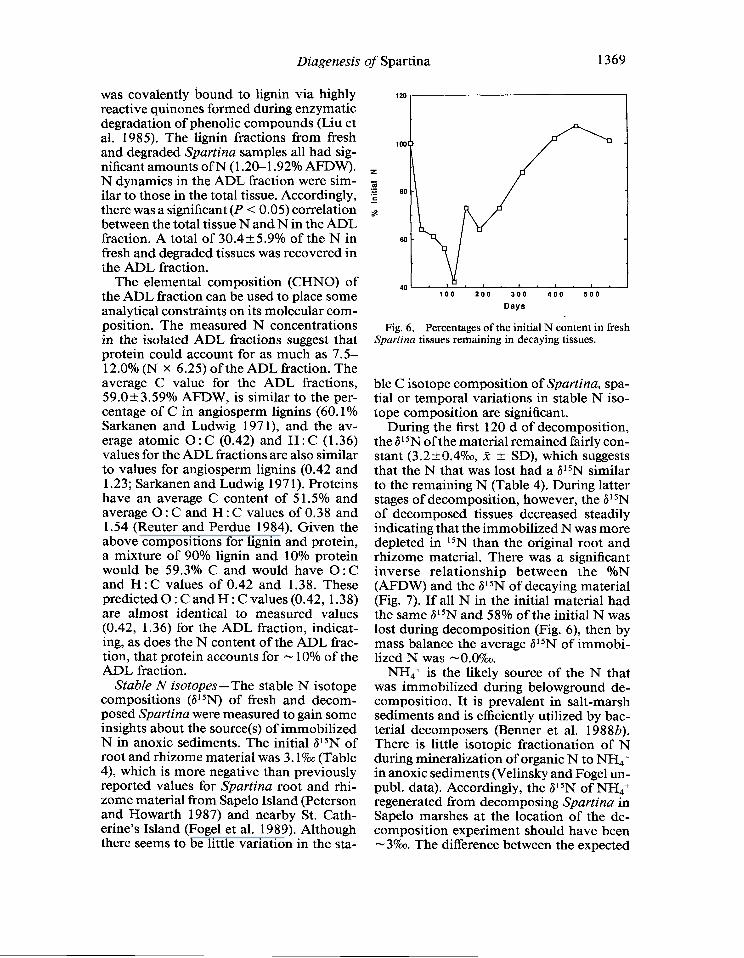
During the first 120 d of decomposition, the 6l 5N oft he material remained fairly con- stant (3.2+0.47~, X + SD), which suggests that the N that was lost had a 615N similar to the remaining N (Table 4). During latter stages of decomposition, however, the 615N of decomposed tissues decreased steadily indicating that the immobilized N was more depleted in 15N than the original root and rhizome material. There was a significant inverse relationship between the %N (AFDW) and the 615N of decaying material (Fig. 7). If all N in the initial material had the same 6’“N and 58% of the initial N was lost during decomposition (Fig. 6), then by mass balance the average a1 5N of immobi- lized N was - O.OY&
NH4+ is the likely source of the N that was immobilized during belowground de- composition. It is prevalent in salt-marsh sediments and is efficiently utilized by bac- terial decomposers (Benner et al. 1988b). There is little isotopic fractionation of N during mineralization of organic N to NH4 + in anoxic sediments (Velinsky and Fogel un- publ. data). Accordingly, the a1 5N of NH4+ regenerated from decomposing Spartina in Sapelo marshes at the location of the de- composition experiment should have been -3%0. The difference between the expected
1370 Benner et al.
Table 4. Stable C and N isotope compositions of whole tissues and major biochemical fractions from fresh and decaying Spartina rocIt and rhizome material. A sequential detergent extraction scheme was used to isolate two plant fractions, lignocellulose (lignin + hemicellulose + cellulose) and lignin. ‘The 6°C of the lignin fraction was measured directly, and the 613C of the polysaccharide fraction was calculated by mass bal- ance.
--I.- -- -__-- dl’C (%) -----
Months Whole Polysac- b”N (‘?‘a~) tissue charldes Lignin Whole tissue
-~ -_--.- ~--
0 -13.1 -11.9 -18.2 3.1 1 -13.2 -12.0 -19.0 3.6 2 - 13.5 -12.6 -19.4 2.8 3 -13.0 -13.7 -19.0 3.7 4 -14.1 - 13.2 - 18.9 3.0 5 -13.6 -12.0 -18.8 2.4 6 -13.5 - 12.7 -17.8 2.3 8 -13.7 -12.5 - 18.4 3.1
10 -13.8 -12.9 - 18.3 2.5 13 - 13.8 - 12.7 -18.2 1.8 15 - 14.0 -12.4 - 18.4 1.5 18 -14.1 -11.6 -17-6 1.1
15/3* -13.9 - 14.0 -18.9 4.5 -- ---~ * Material that was decomposed belowground for 15 months and then
brought to the sediment surface for the I,~st 3 months of decomposition.
615N of pore-water NHLI+ (-3o/oo) and the immobilized N (-OYm) could result from isotopic fractionation during uptake of NH,+ by heterotrophic bacteria (Hoch et al. un- publ.) or from the addition of N from an- other source, such as N,, fixation. Hanson ( 1977) estimated that rates of N2 fixation in short Spartina sediments at Sapelo Island are 13 g N m-* yr -l. Nz fixed by nitrogenase and incorporated into bacterial cells has a similar isotopic composition as dini trogen in air (615N = O.OY&).
An increase from 1.5 to 4.5?G~ was mea- sured in the 615N of material that was de- graded on the sediment surface for the last 3 months of decomposition (Table 4). Dur- ing this period the total N content of the material increased, indicating a net immo- bilization of N. If we assume that the N immobilized aboveground had an average 615N between 5 and 10% (Peterson and Howarth 1987; Fogel et al. 1989), then mass balance calculations indicate that between 30 and 85% of the N associated with the material from belowground was reminer- alized during the last 3 months of above- ground decomposition.
Stable C isotopes-Thle 613C of fresh and
1.5
% N (AFDW)
Fig. 7. Regression analysis of the stable N isotope composition in fresh and decaying Spartina tissues vs. the percentages of total N in tissues.
decomposed Spartina tissues and the iso- lated polysaccharide and lignin components are presented in Table 4. Polysaccharides were enriched in 13C relative to the whole tissue and lignin, and they remained 6.Ot-0.6% heavier than the lignin fraction throughout 1.5 yr of belowground decom- position (Table 4). There was a significant decrease of lo/oo, from - 13.1 to - 14.la/oo, in the a1 3C of whole tissues during decompo- sition Benner et al. (1987) attributed this 17~ decrease in 6l 3C to the relative increase in isotopically light, lignin-derived C in the remaining detritus. There is a significant in- verse relationship between the 613C ofwhole tissues and the lignin concentration in the detritus (Benner et al. 1987).
Temporal variability in the 613C values within the polysaccharide (- 12.5 +_ 0.6%, X + SD) and lignin (-18.5+0.5%, X + SD) fractions reflects the overall experimental error in extraction and isolation of these fractions. Analysis of the two replicate sam- ples collected at the end of the decompo- sition study yielded identical whole tissue values (-- 14.1%), similar polysaccharide values (- 11.6 and - 11.7%0), and identical lignin values (- 17.6o/oo), indicating that overall precision of the methods was very high. It appears, therefore, that the observed temporal variability results more from dif-. ferences in the chemical composition of tis- sues during decomposition and the accuracy
Diagenesis of Spartina
of the extraction methods than from low analytical precision. For instance, large temporal variations in the total N content of tissues were observed during decompo- sition (Fig. 6). These changes in total N con- tent are reflected in the isolated lignin frac- tion, which increased in the ratio of C to N during the initial phase of decomposition and decreased in the ratio of C to N during the latter phase of decomposition (Fig. 8). The isotopic composition of the C in the isolated lignin fraction varied along with the C : N ratio (Fig. 8). It appears that temporal variation in the 613C of the lignin fraction reflects the changing concentrations of pro- tein in this fraction. The most negative 613C values for the lignin fraction (- 19.0 to - 19.4”/,) were measured when the N con- tent in the ADL fraction was relatively low, and these values may most accurately reflect the isotopic composition of “pure” Spar- tina lignin. Consistent with this hypothesis, amino acids are typically slightly enriched in 13C relative to total organic C in cells (Abelson and Hoering 196 1).
Benner et al. (1987) presented a multiple exponential decay model to predict changes in the isotopic composition of Spartina dur- ing decomposition. In this model, the pro- cess controlling the isotopic composition of detrital tissues is the selective removal or degradation of components, such as poly- saccharides, that are less resistant to deg- radation than other components, such as lignin. An assumption of this model is that the C isotopic compositions of the major chemical components of Spartina remain constant during decomposition. Within the limitations of isolating pure chemical frac- tions from highly degraded plant material, the results presented herein indicate that isotopic compositions of the major bio- chemical components of Spartina, namely polysaccharides and lignin, remained con- stant during belowground decomposition.
Tracers of Spartina- The contributions of vascular plant detritus to pools of dis- solved and particulate organic matter in es- tuarine and coastal ecosystems have been difficult to quantify because the organic matter is chemically complex and there are typically many potential sources. The most useful approaches for determining the con-
100 200 300 400 500
Days
1371
-20
Fig. 8. Temporal variations in atomic C : N values and stable C isotope compositions in the ADL fraction of fresh and decaying Spartina tissues.
tributions of vascular plant detritus have been the use specific chemical compounds, or biomarkers (Hedges and Prahll99 l), and stable isotope compositions (Fry and Sherr 1984) as tracers of specific plant tissues. An important uncertainty with both of these approaches is the potential effects of micro- bial decomposition on biomarker concen- trations and stable isotope compositions. In the present study of the diagenesis of Spar- tina root and rhizome material in salt-marsh sediments, long-term trends in the signa- tures of lignin, a biomarker of vascular plant tissues, and of stable C and N isotopes were followed; the implications of alterations in biomarker and isotope signatures are dis- cussed below.
Lignin-derived phenols are very useful biomarkers for determining the contribu- tions of vascular plant tissues to complex mixtures of organic matter in waters and sediments (Hedges et al. 1988). However, two parameters calculated from lignin-de- rived phenol data, the C: V ratio and A, varied significantly during belowground de- composition of Spartina in salt-marsh sed- iments. The cinnamyl : vanillyl ratio (C : v) distinguishes herbaceous and woody tissues and A, the sum of the concentrations of vanillyl, syringyl, and cinnamyl phenols per 100 mg organic C, distinguishes vascular plant tissues and other sources of organic matter. The C: I/ ratio of Spartina de-

1372 Benner et al.
creased by -20% during the study period (Fig. 2) owing to preferential removal ofcin- namyl phenols. The observed shift in the C: Ratio indicates that the contribution of herbaceous tissues to pools of particulate organic matter will be underestimated and the contribution of woody tissues overes- timated if a simple mixi:ng model based on C: V values of fresh tissues is used to dis- tinguish sources.
The A parameter-a measure of the con- centration of lignin-derived phenols-is most useful for estimating the percentage of vascular plant material in environmental samples. The A of Spartzha increased - 1.5- fold during the study period and is likely to continue to increase as decomposition pro- gresses (Benner et al. 1987). These results for Spartina are probably typical for vas- cular plant tissues because the polysaccha- ride components of detritus are microbially degraded at higher rates than the lignin component, which results in gradual en- richment of lignin in the remaining detritus (Benner et al. 1986b). Thus, the contribu- tion of vascular plant material to pools of particulate organic matter will be overesti- mated if A values of fresh plant tissues are used to estimate the percentage of vascular plant detritus in sedimentary organic mat- ter.
Stable C and N isotope compositions of organic matter are extremely valuable trac- ers for determining the contributions of Spartina C to food webs and to pools of detrital materials in salt marshes (Haines 1976; Peterson and Howarth 1987). The stable C isotope composition of Spartina is, however, subject to alteration during de- composition as shown here and in previous studies (Benner et al. 1987; Ember et al. 1987; Alberts et al. 1988; Fogel et al. 1989). The C isotope composition of Spartina shifts to more negative values as the polysaccha- ride components are preferentially degrad- ed leaving a residual material that is en- riched in isotopically “light” lignin-derived C. Benner et al. (1987) measured the C iso- tope composition of the polysaccharide (cel- lulose and hemicellulose) and lignin com- ponents of a wide range of vascular plant tissues and found that the lignin component is consistently depleted in 1 3C by - 3-67~
relative to the polysaccharide components. Given that the polysaccharide components are typically degraded at higher rates than the lignin component and the fact that the lignin component is depleted in 13C relative to polysaccharides, it follows that C isotope compositions of plant tissues (> 5% Ilignin) will become increasingly depleted in 13C as decomposition progresses” In the case of Spartina, the shift in C isotopic composi- tion leads to d13C values similar to those of benthic algae and phytoplankton. End- member mixing models that do not account for this isotopic shift will underestimate the contribution of Spartina to estuarine organ- ic matter.
The N concentration in decomposing Spartina tissues was highly variable as was# reflected in the wide range (43-151) of C : N values. A major fraction (> 50%) of the initial N in the tissues was lost during the early stages of decomposition followed by the slow accumulation of exogenous N. The stable N isotope composition of Spar- tina also varied significantly ( l . l-4.5?&), re- flecting the dynamics of the detrital N pool. The mechanisms leading to shifts in N iso- tope compositions are variable and can re- sult in either enrichment or depletion of 15N in the remaining detritus. Therefore, stable N isotope compositions do not appear to be as applicable for tracing N flow and trophic relationships in detritus-based food webs as they are in grazing-based food webs.
References ABELSON, P. H., AND T. C. HOERING. 1961. Carbon
isotope fractionation in formation of amino acids by photosynthetic organisms. Proc. Natl. Acad. Sci. 47: 623-632.
ALBERTS, J. J., 2. FILIP, M. T. PRKE, D. J. WILLIAMS, AND M. C. WILLIAMS. 1988. Elemental compo- sition, stable carbon isotope ratios and spectro- photometric properties of humic substances OC- curring in a salt marsh estuary. Org. Geochem. 12: 455-467.
BENNER, R., M. L. FOGEL, E.K. SPRAGUE, AND R. E. HODSON. 1987. Depletion of 13C in lignin and its implications for stable carbon isotope studies. Nature 329: 708-7 10.
-, P. G. HATCHER, AND J. I. HEDGES. 1990a. Early diagenesis of mangrove leaves in a tropical estuary: Bulk chemical characterization using sol- id-state ‘“C NMR and elemental analyses. Geo- chim. Cosmochim. Acta 54: 2003-2013.
-, R. E. HODSON, AND D. KIRCHMAN. 1988a. Bacterial abundance and production on mangrove

Diagenesis of Spartina 1373
leaves during initial stages of leaching and biodeg- radation. Ergeb. Limnol. 31: 19-26.
-, J. LAY, E. K’NEES, AND R. E. HODSON. 19 88b. Carbon conversion efficiency for bacterial growth on lignocellulose: Implications for detri- tus-based food webs. Limnol. Oceanogr. 33: 15 14- 1526.
-, A. E. MACCUBBIN, AND R. E. HODSON. 1984~. Anaerobic biodegradation of the lignin and poly- saccharide components of lignocellulose and syn- thetic lignin by sediment microflora. Appl. En- viron. Microbial. 47: 998-1004.
.- - AND -. 19843. Preparation, chLracteriz;lion, and microbial degradation of specifically radiolabeled [ 14C]lignocellulose from marine and freshwater macrophytes. Appl. Envi- ron. Microbial. 47: 381-398.
- -, AND -. 1986~. Temporal rela- tic&ship between the deposition and microbial degradation of lignocellulosic detritus in a Georgia salt marsh and the Okefenokee Swamp. Microb. Ecol. 12: 291-298.
-, M. A. MORAN, AND R. E. HODSON. 19863. Biogeochemical cycling of lignocellulosic carbon in marine and freshwater ecosystems: Relative contributions of procaryotes and eucaryotes. Lim- nol. Oceanogr. 31: 89-100.
- K. WELIKY, AND J. I. HEDGES. 1990b. Early diigenesis of mangrove leaves in a tropical estu- ary: Molecular-level analyses ofneutral sugars and lignin-derived phenols. Gcochim. Cosmochim. Acta 54: 1991-2001.
BUTH, G. J. C. 1987. Decomposition of roots and rhizomes of three plant communities in a Dutch salt marsh. Aquat. Bot. 29: 123-138.
COLBERG, P. J., AND L. Y. YOUNG. 1985. Anaerobic degradation of soluble fractions of [14C-lig- ninllignocellulose. Appl. Environ. Microbial. 49: 345-349.
DILL, I., J. SALNIKOW, AND G. KRAEPELIN. 1984. Hy- droxyproline-rich protein material in wood and lignin of Fagus sylvadca. Appl. Environ. Micro- biol. 48: 1259-l 26 1.
EMBER, L. M., D. F. WILLIAMS, AND J. T. MORRIS. 1987. Processes that influence carbon isotopic variations in salt marsh sediments. Mar. Ecol. Prog. Ser. 36: 33-42.
ERTEL, J. R., AND J. I. HEDGES. 1984. The lignin component of humic substances: Distribution among soil and sedimentary humic, fulvic, and base-insoluble fractions. Geochim. Cosmochim. Acta 48: 2065-2074.
- - A. H. DEVOL, J. E. RICHEY, AND M. DE)N. G. I&BEIRO. 1986. Dissolved humic sub- stances of the Amazon River system. Limnol. Oceanogr. 31: 739-754.
FRY B., AND E. B. SHERR. 1984. 613C measurements as indicators of carbon flow in marine and fresh- water ecosystems. Contrib. Mar. Sci. 27: 13-47.
FOGEL, M. L., E. K. SPRAGUE, A. P. GIZE, AND R. W. FREY. 1989. Diagenesis of organic matter in salt marshes. Estuarine Coastal Shelf Sci. 28: 2 1 l-230.
GALLAGHER, J. L., AND F. G. PLUMLEY. 1979. Un- derground biomass profiles and productivity in Atlantic coastal marshes. Am. J. Bot. 66: 156-l 6 1.
GOOD, R. E., N. F. GOOD, AND B. R. FRASCO. 1982. A review of primary production and decomposi- tion dynamics of the belowground marsh com- ponent, p. 139-157. Zn V. S. Kennedy [ed.], Es- tuarine comparisons. Academic.
HACKETT, W. F., W. J. CONNORS, T. K. KIRK, AND J. G. ZEIKUS. 1977. Microbial decomposition of synthetic 14C-labeled lignins in nature: Lignin biodegradation in a variety of natural materials. Appl. Environ. Microbial. 33: 43-5 1.
HACKNEY, C. T., AND A. A. DE LA CRUZ. 1980. In situ decomposition of roots and rhizomes of two tidal marsh plants. Ecology 61: 226-23 1.
HADDAD, R. I. 1989. Sources and reactivity of organic matter accumulating in a rapidly depositing coast- al marine sediment. Ph.D. thesis, Univ. North Carolina. 403 p.
HAINES, E. B. 1976. Stable carbon isotope ratios in the biota, soils and tidal water of a Georgia salt marsh. Estuarine Coastal Mar. Sci. 4: 609-6 16.
HAMILTON, S. E., AND J. I. HEDGES. 1988. The com- parative geochemistries of lignins and carbohy- drates in an anoxic fjord. Geochim. Cosmochim. Acta 52: 129-142.
HANSON, R. B. 1977. Comparison of nitrogen fixation activity in tall and short Spartina alterniflora. Appl. Environ. Microbial. 33: 569-602.
HARRISON, P. G., AND K. H. MANN. 1975. Detritus formation from eelgrass (Zosferu marina L.): The relative effects of fragmentation, leaching, and de- cay. Limnol. Oceanogr. 20: 924-934.
HARTLEY, R. D. 1973. Carbohydrate esters of ferulic acid as components of cell-walls OC Lo&m mul- tiflorum. Phytochemistry 12: 661-665.
HEDGES, J. I. 1975. Lignin compounds as indicators of terrestrial organic matter in marine sediments. Ph.D. thesis, Univ. Texas, Austin. 137 p.
-, W. A. CLARK, AND G. L. COWIE. 1988. Or- ganic matter sources to the water column and sur- ficial sediments of a marine bay. Limnol. Ocean- ogr. 33: 1X16-1 136.
-, AND J. R. ERTEL. 1982. Characterization of lignin by gas capillary chromatography of cupric oxide oxidation products. Anal. Chem. 54: 174- 178.
-, AND D. C. MANN. 1979. The characterization of plant tissues by their lignin oxidation products. Geochim. Cosmochim. Acta 43: 1803-l 807.
AND F. G. PRAHL. 1991. Early diagenesis: Cdnsequences for applications of molecular bio- markers, in press. Zn M. Engel and S. A. Macko [eds.], Organic geochemistry. Plenum.
AND K. WELIKY. 1989. The diagenesis of co- niier needles in a coastal marine environment. Geochim. Cosmochim. Acta 53: 2659-2673.
HIGUCHI, T., Y. ITO, M. SHIMADA, AND I. KAWAMURA. 1967. Chemical properties of milled wood lignin of grasses. Phytochemistry 6: 155 l-l 556.
HOBBIE, J. E., AND C. LEE. 1980. Microbial produc- tion of extracellular material: Importance in ben- thic ecology, p. 341-346. Zn K. R. Tenore and B. C. Coull [eds.], Marine benthic dynamics. Univ. South Carolina.
HOWARTH, R. W., AND J. E. HOBBIE. 1982. The reg- ulation 0.f decomposition and heterotrophic mi-

1374 Benner et al.
crobial activity in salt marsh soils, p. 183-207. In V. S. Kennedy [ed.], Estuarine comparisons. Ac- ademic.
HOWES, B. L., J. W. DACEY, AND G. M. KING. 1984. Carbon flow through oxygen and sulfate reduction pathways in salt marsh sediments. Limnol. Ocean- ogr. 29: 1037-105 1.
--- -, AND .J. M. TEAL. 1985. Annual car- bon mineralization and belowground production of Spartina alterniflora in a New England salt marsh. Ecology 66: 595-605.
LEE, C., R. W. HOWARTH, AND B. I. Howss. 1980. Sterols in decomposing Spartina alterniflora and the use of ergosterol in esti mating the contribution of fungi to detrital nitrogen. Limnol. Oceanogr. 25: 290-303.
Lrrr, S.-Y., A. J. FREYER, R. 1~. MINARD, AND J.-M. BOLLAG. 1985. Enzyme-catalyzed complex-for- mation of amino acid esters and phenolic humus constituents. Soil Sci. Sot. Am. J. 49: 337-341.
MACKO, S. A. 198 1. Stable nitrogen isotope ratios as tracers of organic geoche mica1 processes. Ph.D. thesis, Univ. Texas, Austin. 181 p-
MANDERS, W. F. 1987. Solid-state 13C NMR detcr- mination of the syringyl/,guaiacyl ratio in hard- woods. Holforschung 41: 13-l 8.
MORAN, M. A., AND R. E. HODSON. 1990. Contri- butions of degrading Spadina alterniflora ligno- cellulose to the dissolved organic carbon pool of a salt marsh. Mar. Ecol. Prog. Ser. 62: 16 l-168.
ODUM, W. E., P. W. KIRK, AND J. C. ZIEMAN. 1979. Non-protein nitrogen compounds associated with particles of vascular plant detritus. Oikos 32: 363- 367.
PETERSON, B.J., AND R. W. HOWARTH. 1987. Sulfur, carbon, and nitrogen isotopes used to trace organic matter flow in the salt-marsh estuaries of Sapelo Island, Georgia. Limnol. Oceanogr. 32: 1195-I 2 13.
POMEROY, L. R., AND OTHERS. 1981. Primary pro- duction, p. 39-67. Zn L. 1R. Pomeroy and R. G. Wiegert [eds.], The ecology of a salt marsh. Springer.
REUTER, J. H., AND E. M. PERDUE. 1984. A chemical structural model of early diagenesis of sedimentary humus/proto-kerogens. h4itt. Geol.-Palaeontol. Inst. Univ. Hamburg 56: 249-262.
RICE, D. L. 1982. The detritus nitrogen problem: New observations and perspectives from organic geochemistry. Mar. Ecol. Prog. Ser. 9: 153-l 62.
--- , AND R. B. HANSON. 1984. A kinetic model for detritus nitrogen: Role: of the associated bac- teria in nitrogen accumulation. Bull. Mar. Sci. 35: 326-340.
SARKANEN,K.V.,ANDC.H. LIJDWIG. 1971. Lignins:
Occurrence, formation, structure, and reactions. Wiley.
SCHIJBAUER, J.P., ANDC.S.HOPKINSON. 1984. Above- and bclowground emergent macrophyte produc- tion and turnover in a coastal marsh ecosystem, Georgia. Limnol. Oceanogr. 29: 1052-l 065.
SHARMA, P., L. R. GARDNER, W. S. MOORE, AND M. S. BOLLINGER. 1987. Sedimentation and biotur- bation in a salt marsh as revealed by 210Pb, 137Cs, and ‘Be studies. Limnol. Oceanogr. 32: 3 13-326.
VALIELA, I. J., AND OTHERS. 1985. Decomposition in salt marsh ecosystems: The phases and major fac- tors affecting disappearance of above-ground or- ganic matter. J. Exp. Mar. Biol. 89: 29-54.
p,AND OTHERS. 1984. Importance of chemical composition of salt marsh litter on decay rates and feeding by detritivores. Bull. Mar. Sci. 35: 26 l- 269.
VAN SOEST, P. J., AND ROBERTSON. J. B. 1980. Sys- tems of analysis for evaluating fibrous feeds, p. 49-60. Zn W. J. Pigden et al. [eds.], Standardiza- tion of analytical methodology for feeds. Int. De- velop. Res. Center.
WHITMORE, F. W. 1982. Lignin-protein complex in cell walls of Pinus elliottii: Amino acid constitu- cnts. Phytochemistry 21: 3 15-3 18.
WIEGERT, R.G., AND L.R. POMEROY. 1981. The salt- marsh ecosystem: A synthesis, p. 219-230. Zn L.. R. Pomeroy and R. G. Wiegert [eds.], The ecology of a salt marsh. Springer.
WILSON, J. O., R. BUCHSBAUM, I. VALIELA, AND T. SWAIN. 1986. Decomposition in salt marsh cco- systems: Phenolic dynamics during decay of li ttcr of Spartina ufterniJlora. Mar. Ecol. Prog. Ser. 29: 177-187.
p--1 I. VALIELA,AND T. SWAIN. 1985. Sourcesand concentrations of vascular plant material in sed- iments of Buzzards Bay, Massachusetts, USA. Mar. Biol. 90: 129-137.
YELVERTON, F.G., AND C.T. HACKNEY. 1986. Flux ofdissolved organic carbon and pore water through the substrate of a Spartina alterniflora marsh in North Carolina. Estuarine Coastal Shelf Sci. 22: 255-267.
ZECH, W.,M.-B. JOHANSSON, L. HAUMAIER,AND R. L. MALCOLM. 1987. CPMAS 13C NMR and IR spectra of spruce and pine litter and of the Klason lignin fraction at different stages of decomposition. Z. Pflanzcnernaehr. Bodenkd. 150: 262-265.
Submitted: 23 November 1990 Accepted: 23 April 1994
Revised: 13 May 1991
Related Documents











