D型インフルエンザ 堀本泰介 東京大学大学院 農学生命科学研究科 獣医微生物学教室 ・D型インフルエンザウイルスの発見 ・ウイルス分子性状と病原性 ・わが国におけるD型インフルエンザ疫学調査 ・日本株の分離と性状解析 ・D型ウイルス粒子内ゲノム構造

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

D型インフルエンザ
堀本泰介
東京大学大学院農学生命科学研究科獣医微生物学教室
・D型インフルエンザウイルスの発見
・ウイルス分子性状と病原性
・わが国におけるD型インフルエンザ疫学調査
・日本株の分離と性状解析
・D型ウイルス粒子内ゲノム構造

型 A B C
宿主
亜型 HA (H1-H18)NA (N1-N11)
なし なし
ヒトブタウマ鳥
その他
ヒト(アザラシ)
ヒト(ブタ)
インフルエンザウイルス
ヒト流行
季節性パンデミック
季節性
ウシブタ・・
D
??
??
小規模

D型インフルエンザウイルス(Flu D)
呼吸器疾患のブタから分離(C/swine/Oklahoma/1334/2011)
AGID 抗原性がFlu Cとは異なる。ウシが主要な宿主動物

1. A, B, C型と免疫拡散法で抗原交差しない。2. 7本鎖。3. C型との遺伝学的距離が、A型とB型の差と
同じくらい離れている。4. C型と遺伝子交雑しない。5. ゲノム分節末端配列がC型と異なる。6. M1の転写の仕方がC型と異なる。7. ウシ, ヒツジ, ヤギに感染する。8. ヒトへの感染性が明らかでない。9. ヨーロッパ、アジアでも見つかる。
分離ウイルス
→ Influenza D virus(ICTV 2016)

Orthomyxoviridaeの分類(ICTV 2017)
Genus: AlphainfluenzavirusSpecies: Influenza A virus
human influenza A virus, avian influenza A virus, swine influenza A virus, equine influenza A virus,canine influenza A virus
Genus: BetainfluenzavirusSpecies: Influenza B virus
human influenza B virusGenus: Gammainfluenzavirus
Species: Influenza C virushuman influenza C virus
Genus: DeltainfluenzavirusSpecies: Influenza D virus
bovine influenza D virus, swine influenza D virusその他のGenus: Isavirus, Quaranjavirus, Thogotovirus

HEF: hemagglutinin-esterase-fusion
Wang and Veit, Protein Cell, 2016
インフルエンザウイルスの構造
Flu C/D Flu A/B7分節 8分節

Hause et al, mBio, 2014
Flu C
Flu A
Flu B
Flu D
Isa, Thogoto
Flu C
Flu D
NP
HEF
オルソミクソウイルス系統樹

タンパク質 PB2 PB1 HEF
相同性 (%)
Flu D vs. Flu C
NPP3 P42 NS1/2
53 70 53 4050 38 33/29
ORF pU3’-UCGUCUUCGUC-
3’-UCGUAUUCGUC-
3’-CCGUAUUCGUC-
-GAACGAUGACGA-5’
-GAACGAUGACGA-5’
-GAACGAUGACGA-5’
Flu C
Flu D
pU
Terminal sequence
pU
HEF, M, NS
3Ps & NP

Flu Dのゲノム構造
Su et al. Virulence, 2017

Flu A/B HA vs. Flu C/D HEF
Wang and Veit, Protein Cell, 2016
(C only)
(D only)
(D only)
HA HEF

Different acylation and lipid raft association of HA or HEF protein.
HA vs. HEF
Wang and Veit, Protein Cell, 2016
(Flu C) RTK(Flu D) KK
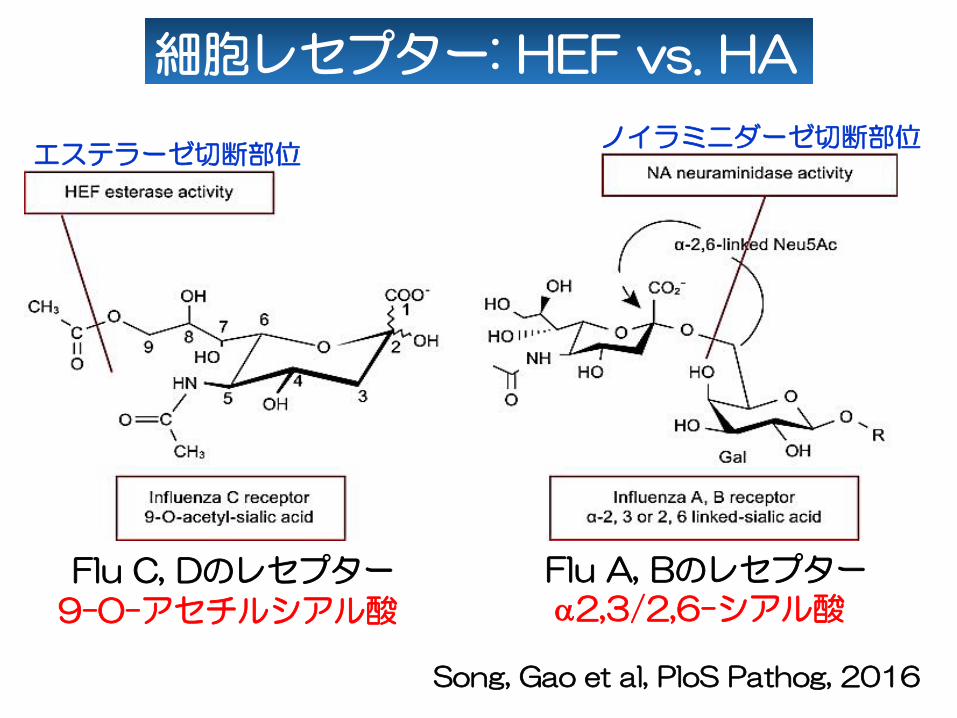
Flu C, Dのレセプター
9-O-アセチルシアル酸
細胞レセプター: HEF vs. HA
Song, Gao et al, PloS Pathog, 2016
エステラーゼ切断部位ノイラミニダーゼ切断部位
Flu A, Bのレセプターα2,3/2,6-シアル酸

牛呼吸器病症候群(BRDC)
ウイルス、細菌、ストレス等の複合的要因によって発生例:輸送熱、流行性肺炎
世界全体で3,600億円以上/年の経済的被害
ウイルス牛伝染性鼻気管炎ウイルス牛ウイルス性下痢ウイルス牛パラインフルウイルス3牛RS ウイルス牛アデノウイルス牛コロナウイルス牛鼻炎A/Bウイルス牛パルボウイルス
細菌マンヘミアマイコプラズマパスツレラヒストフィルス
環境因子ストレス、気温湿度、ガスほこり
臨床症状・急性期:食欲消失、高熱、呼吸促拍、沈鬱、咳、鼻汁・慢性期:衰弱、咳、呼吸困難

・メタゲノム解析で、これらよりもD型インフルエンザウイルスの方がBRDC牛から高頻度に検出(2016)
現在のBRDCの主要病原体はFlu Dである可能性
ウイルス牛伝染性鼻気管炎ウイルス牛ウイルス性下痢ウイルス牛パラインフルウイルス3牛RS ウイルス牛アデノウイルス牛コロナウイルス牛鼻炎A/Bウイルス牛パルボウイルス
・ワクチンを接種してもBRDCは発生する。
混合ワクチン
牛呼吸器病症候群(BRDC)

D型インフルエンザウイルスが検出された国(2015年まで)
日本の牛におけるD型ウイルスの疫学調査

血清(血漿)茨城県下の牧場飼育の乳用牛(ホルスタイン)28頭RDE・非働化処理
ウイルスD/swine/Oklahoma/1334/2011(カンザス大 Hause博士より分与)
0.6%七面鳥赤血球を用いてHI試験(≧1:40を陽性)
D型ウイルス特異抗体の検出
Murakami et al, EID, 2016

1/8採血採血時年齢
5y6m4y5m3y11m3y10m3y4m3y4m3y4m3y3m3y1m2y2m2y1m2y1m2y0m1y12m1y7m1y3m1y0m0y11m0y8m0y7m7y9m8y0m6y11m6y3m7y10m1y11m1y10m9y0m
<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<4080
16080
640160
8080
160
HI価
過去にFlu Dに感染わが国にもFlu Dは存在
No. 移動歴
123456789
10111213141516171819202122232425262728
茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城茨城北海道北海道茨城茨城北海道茨城・北海道茨城・北海道茨城

4080
160<40160
80160320160160160
8080
160160160
8040
32080
16080
160320
80160
4080
本牧場においてFlu Dの流行があった。
鼻スワブRT-PCR 陽性
ゲノム全塩基配列を決定(茨城株)ウイルス分離は陰性
No.1/8採血
123456789
10111213141516171819202122232425262728
<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<40<4080
16080
640160
8080
160
HI価
2/3採血

0%
~15%
~30%
~45%
45%~
Horimoto et al, PloS One, 2016
日本における抗体保有状況まとめ
陽性率
計30.4%
プレゼンター
プレゼンテーションのノート
まず鳥インフルエンザと研究の背景についてご説明いたします。 鳥インフルエンザウイルスはオルソミクソウイルス科のインフルエンザAに属する-鎖ssRNAウイルスです。 このウイルスの遺伝子は8本の分節状を呈し、それぞれウイルスのポリメラーゼや構造タンパクなどに関する情報をコードしています。

D型ウイルス抗体陽性率
0
20
60
40
80
陽性
率(%
)
ホルスタイン
黒毛和種 HI 価
0
20
60
40
80
100
40
80
16
0
32
0
64
0
120
140
ホルスタイン黒毛和種
陽性
数

0
20
60
40
80
陽性
率(%
)
採血年
20
10
20
12
20
13
20
14
20
15
20
16
0
20
60
40
80
陽性
率(%
)
年齢
1 2 3 4 5 6 7 8-0.5
D型ウイルス抗体陽性率

0
20
40
60
80
100
3 5 8 9 11 12 13 14 15 17 18 19 20 21 30 31 32 33 34 35 36 37 38 39 40 41
陽性
率(%
)
宮崎県内農場別抗体陽性率
農場番号
→ 高い水平伝播性を示唆

ケニア ラクダ99% 抗体陽性自然宿主??
Flu Dが検出された国(2017年まで)
Salem et al, EID, 2017

地域 動物種 サンプル数
Bati近郊
Jijiga近郊
Bati
Jijiga
ラクダヤギ
ヒツジウシロバ
21122082
ラクダウシヤギ
1778
合計 ラクダウシヤギ
ヒツジロバ
381520202
95
放牧地を共有。夜になって放牧地から連れて帰り家畜ごとに収容。
泉對博先生(日大獣医)
エチオピアのヒトコブラクダ血清

地域 動物種
Bati近郊 ラクダヤギ
ヒツジウシロバ
HI結果陽性数(陽性率%)
19/21 (90.5)1/12 (8.3)0/20 (0)0/8 (0)0/2 (0)
Jijiga近郊 ラクダウシヤギ
3/17 (17.6)0/7 (0)0/8 (0)
合計 ラクダヤギ
ヒツジウシロバ
22/38 (57.9)1/20 (5.0)0/20 (0)0/15 (0)0/2 (0)
エチオピアのヒトコブラクダ血清

15 samples
10 samples
15 samples
40検体全て陰性(<40)
HI結果
モンゴルのフタコブラクダ血清
ラクダは草原で放牧。所有者はゲルに居住し家畜とともに移動。夜になっても放牧したまま。

まとめ
・国内においてFluDによる牛の呼吸器疾患を初めて検出した。
・FluDはすでに全国的にウシ社会に侵淫している。
・FluDの自然宿主は東アフリカのヒトコブラクダである?

牧場 月齢 性別Flu D
(RT-PCR)
A 14 ♀ 陽性
A 14 ♀ 陽性
・呼吸器症状牛の鼻腔検体(山形)
ウイルス分離
HRT-18G(ヒト結腸癌)
ST(ブタ精巣)
trypsin存在下で継代
5日 5日
CPE観察HA価 8
D/bovine/Yamagata/2016(山形株)
ウシFlu Dの分離と性状解析

PB2 PB1 P3
HEF NP M NS
日本株の系統樹解析
D/bovine/Nebraska/9-5/2012D/bovine/Oklahoma/660/2013
D/bovine/Texas/3-13/2011D/bovine/Kansas/11-8/2012
D/bovine/Kansas/13-21/2012D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Italy/1/2014D/bovine/Italy/46484/2015
D/bovine/Mississippi/C00046N/2014D/bovine/Kansas/14-22/2012
D/swine/Oklahoma/1334/2011D/swine/Italy/199724-3/2015
D/bovine/Kansas/1-35/2010D/bovine/France/2986/2012
D/bovine/Shandong/Y125/2014D/bovine/Shandong/Y127/2014
D/bovine/Shandong/Y217/2014D/bovine/Miyazaki/B22/2016D/bovine/Ibaraki/7768/2016D/bovine/Ibaraki/7765/2016D/bovine/Yamagata/10710/2016
97100
10084
97
88
89
74
97
94
0.0050
D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Kansas/13-21/2012D/bovine/Kansas/1-35/2010D/bovine/Texas/3-13/2011
D/bovine/Nebraska/9-5/2012D/bovine/Kansas/11-8/2012
D/bovine/Oklahoma/660/2013D/bovine/Mississippi/C00046N/2014
D/swine/Oklahoma/1334/2011D/bovine/Kansas/14-22/2012
D/swine/Italy/199724-3/2015D/bovine/Italy/46484/2015
D/bovine/Italy/1/2014D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y127/2014D/bovine/Shandong/Y217/2014
D/bovine/France/2986/2012D/bovine/Miyazaki/B22/2016
D/bovine/Ibaraki/7768/2016D/bovine/Yamagata/10710/201695
100
100
99
8913
67
10095
82
98
7755
96
0.0050
D/bovine/Shandong/Y217/2014D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y127/2014D/bovine/France/2986/2012
D/bovine/Miyazaki/B22/2016D/bovine/Ibaraki/7768/2016
D/bovine/Ibaraki/7765/2016D/bovine/Yamagata/10710/2016
D/bovine/Mississippi/C00046N/2014D/swine/Oklahoma/1334/2011D/bovine/Kansas/14-22/2012
D/swine/Italy/199724-3/2015D/bovine/Italy/1/2014D/bovine/Italy/46484/2015
D/bovine/Texas/3-13/2011D/bovine/Kansas/1-35/2010
D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Kansas/13-21/2012D/bovine/Kansas/11-8/2012
D/bovine/Oklahoma/660/2013D/bovine/Nebraska/9-5/2012
62100
100
8692
5954
50
100
42
61
98
0.0050
D/bovine/Shandong/Y127/2014D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y217/2014D/bovine/Kansas/1-35/2010
D/bovine/Kansas/11-8/2012D/bovine/Nebraska/9-5/2012
D/bovine/Oklahoma/660/2013D/swine/Oklahoma/1334/2011
D/bovine/Mississippi/C00046N/2014D/bovine/Kansas/14-22/2012
D/bovine/Italy/46484/2015D/bovine/Italy/1/2014
D/swine/Italy/199724-3/2015D/bovine/France/2986/2012
D/bovine/Texas/3-13/2011D/bovine/Kansas/13-21/2012
D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013D/bovine/Miyazaki/B22/2016
D/bovine/Yamagata/10710/2016D/bovine/Ibaraki/7768/2016D/bovine/Ibaraki/7765/201692
94100
99100
10089
100
95
65
70
100
9474100
42
0.0050
D/bovine/Italy/46484/2015D/swine/Italy/199724-3/2015D/bovine/Italy/1/2014
D/bovine/Mississippi/C00030P/2014D/bovine/Mississippi/C00046N/2014
D/bovine/Kansas/14-22/2012D/swine/Oklahoma/1334/2011
D/bovine/Kansas/1-35/2010D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Shandong/Y127/2014D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y217/2014D/bovine/France/2986/2012
D/bovine/Kansas/13-21/2012D/bovine/Texas/3-13/2011D/bovine/Kansas/11-8/2012
D/bovine/Nebraska/9-5/2012D/bovine/Oklahoma/660/2013
D/bovine/Mississippi/C00014N/2014D/bovine/Mississippi/C00013N/2014
D/bovine/Miyazaki/B22/2016D/bovine/Yamagata/10710/2016D/bovine/Ibaraki/7768/2016D/Bovine/Ibaraki/7765/201676
87100
10099
99
100
42
40
100
100
100
98
7280
94
100
67
0.0050
D/swine/Italy/199724-3/2015D/bovine/Italy/1/2014
D/bovine/Italy/46484/2015D/bovine/Mississippi/C00046N/2014
D/swine/Oklahoma/1334/2011D/bovine/Texas/3-13/2011
D/bovine/Shandong/Y125/2014D/bovine/Shandong/Y127/2014
D/bovine/Shandong/Y217/2014D/bovine/France/2986/2012
D/bovine/Kansas/1-35/2010D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Kansas/13-21/2012D/bovine/Oklahoma/660/2013D/bovine/Nebraska/9-5/2012
D/bovine/Kansas/11-8/2012D/bovine/Kansas/14-22/2012D/bovine/Miyazaki/B22/2016
D/bovine/Yamagata/10710/2016D/bovine/Ibaraki/7768/2016D/bovine/Ibaraki/7765/201699
100
100
6366
95100
97
62
57
100100
99
98
97
100
0.0050
D/bovine/Italy/1/2014D/swine/Italy/199724-3/2015
D/bovine/Nebraska/9-5/2012D/bovine/Oklahoma/660/2013
D/bovine/Kansas/11-8/2012D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Texas/3-13/2011D/bovine/Kansas/1-35/2010
D/bovine/Kansas/13-21/2012D/swine/Oklahoma/1334/2011
D/bovine/Kansas/14-22/2012D/bovine/Mississippi/C00046N/2014
D/bovine/France/2986/2012D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y127/2014D/bovine/Shandong/Y217/2014
D/bovine/Miyazaki/B22/2016D/bovine/Ibaraki/7765/2016
D/bovine/Ibaraki/7768/2016D/bovine/Yamagata/10710/201699
100
10057
100
8999
100
100
75
58
55
59
0.0050

伊
仏
中
米2
米1
日
D/bovine/Italy/46484/2015D/swine/Italy/199724-3/2015D/bovine/Italy/1/2014
D/bovine/Mississippi/C00030P/2014D/bovine/Mississippi/C00046N/2014
D/bovine/Kansas/14-22/2012D/swine/Oklahoma/1334/2011
D/bovine/Kansas/1-35/2010D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Shandong/Y127/2014D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y217/2014D/bovine/France/2986/2012
D/bovine/Kansas/13-21/2012D/bovine/Texas/3-13/2011D/bovine/Kansas/11-8/2012
D/bovine/Nebraska/9-5/2012D/bovine/Oklahoma/660/2013
D/bovine/Mississippi/C00014N/2014D/bovine/Mississippi/C00013N/2014
D/bovine/Miyazaki/B22/2016D/bovine/Yamagata/10710/2016D/bovine/Ibaraki/7768/2016D/Bovine/Ibaraki/7765/2016
100
9999
100
40
100
100
9894
100
0.0050
OK系統
660系統
日本系統
HEF

D/bovine/Shandong/Y217/2014D/bovine/Shandong/Y125/2014
D/bovine/Shandong/Y127/2014D/bovine/France/2986/2012
D/bovine/Miyazaki/B22/2016D/bovine/Ibaraki/7768/2016
D/bovine/Ibaraki/7765/2016D/bovine/Yamagata/10710/2016
D/bovine/Mississippi/C00046N/2014D/swine/Oklahoma/1334/2011D/bovine/Kansas/14-22/2012
D/swine/Italy/199724-3/2015D/bovine/Italy/1/2014D/bovine/Italy/46484/2015
D/bovine/Texas/3-13/2011D/bovine/Kansas/1-35/2010
D/bovine/Minnesota/628/2013D/bovine/Minnesota/729/2013
D/bovine/Kansas/13-21/2012D/bovine/Kansas/11-8/2012
D/bovine/Oklahoma/660/2013D/bovine/Nebraska/9-5/2012
100
100
92
50
100
42
98
0.0050
伊
仏
中
米1
米2
日 OK系統
660系統
M

ホモロジー 山形株 vs. 米国OK株
分節 塩基 アミノ酸
PB2 96% 98%
PB1 96% 98%
P3 96% 96%
HEF 96% 95%
NP 95% 96%
M 97% 97%
NS 96% 96%
日本株間 99.5~99.8% 99.6~100%

OK系統vs.
日本系統
K212(OK) R212(660) S212
HEFの系統間で異なるアミノ酸部位
OK系統vs.
660系統
日本系統vs.
660系統

クローンウイルス
山形株 OK株 660株
B4 6,400 1,600 6,400
B13 <100 <100 <100
R36 12,800 25,600 6,400
G22 3,200 100 <100
G27 2,040,800 <100 <100
G74 400 <100 200
単クローン性抗体のHI反応性

まとめ
・日本株は外国株と系統的に異なり、国内で独自に進化した。
・FluD 日本株(山形株)を分離した。
・HEF抗原には共通エピトープと系統特異エピトープがある。
・日本株は外国株とHEF抗原性が異なる。

50nm
インフルエンザウイルス粒子内のRNP数
A型
B型
Nakatsu et al., mBio, 2016

8 vRNAs 7 vRNAs
HA分節欠損
人工的に作製した7分節A型ウイルスのRNP数
7本(17%)
8本(83%)
8本(100%)
N=11 N=12

Covera
ge D
ep
th(n
ucle
otid
e r
ead
s/to
tal n
ucle
otide r
ead
s)
PB2 HA MNANP NSPAPB1
28SrRNA
18SrRNA
Nucleotide position (cRNA for virus genes)
3′5′
0
0.005
0.010
0.015
0.020
0.025
0
0.005
0.010
0.015
0.020
0.025
8.8% 6.5%
12.1%野生型ウイルス
HA(-)ウイルス
HA(-)ウイルス粒子内のRNAの解析
Noda, Murakami et al. Nature Comm, 2018

C型ウイルス D型ウイルス
8本(70%)
8本(73%)
7本(27%)
7本(30%)
7分節のC型, D型ウイルスでは?
Nakatsu et al., JVI, 2018

まとめ
・8本のRNPが“1+7”に配置することが効率のよいゲノムパッケージングに重要と考えられる。
・ゲノム分節数にかかわらずインフルエンザウイルスは8本のRNPを取り込んでいる。
・ゲノムパッケージングの際に宿主RNAを利用する可能性がある。

カンザス大 Ben Hause北大 日尾野隆大、迫田義博宮大 目堅博久、乗峰潤三岩大 彦野弘一、村上賢二岐大 猪島康雄鹿大 安藤貴朗、乙丸孝之介、小澤真麻大 佐藤礼一郎、村上裕信、阪口雅弘岩医大 村木靖大阪府開業 石井一功日生研 大森崇司、田積晃浩、平修鳥取県 増田恒幸京大ウ再研 野田岳志東大医科研 河岡義裕東大農 小田切友葉、石田大歩、遠藤麻衣子、
上間亜希子、小林知也、村上晋
Acknowledgment
Related Documents











